Chapter 13
Genetic Variation
in Populations
Figure CO: An albino whitetail fawn
© Srcromer/Dreamstime.com
Overview
• Genetic variation arises from random
mutations
• Mutation rates vary
• Some loci are more likely to mutate than
others
• In the short term, when microevolution
operates, the frequency of different alleles in
the population from prior mutations is more
important than the creation of new mutations
Overview
• The ability of DNA to repair itself also limits the
impact of new, potentially harmful, mutations
• Quantitative trait loci (QTL) that contain groups
of genes often influence phenotypic characters
that show continuous variation
• Genetic Drift and Gene Flow also impact the
genome composition/gene pools of populations
• The genetic history of populations contributes to
our understanding of patterns of species
distributions
Mutation
• Mutation is occurs in all populations
• New mutations that arise, if not neutral in
effect, will rarely be better, and likely will be
worse, than the alleles already present
• Changes in environmental conditions can elicit
a genetic response based on the available
genetic variation in a population (evolution)
Resistance to DDT
in Common Houseflies
• Rapid genetic changes occur in many insect populations
exposed to pesticides such as dieldrin and DDT
Figure 01_INS: Resistance to DDT in
common houseflies
© Frank B. Yuwono/ShutterStock, Inc.
Adapted from Strickberger, M. W. Genetics, Third edition.
Macmillan, 1985; based on data from Decker, G. E., and W.
N. Bruce, Amer. J. Trop. Med. Hygiene 1 (1952): 395-403.
• Both dieldrin and DDT
are neurotoxins; DDT
opens sodium channels
and both DDT and
dieldrin stimulate Ach
synthesis and release
• Resistance develops
when mutant neuron
membrane proteins arise
or the numbers of
receptors change
Selection for Pesticide Resistance
Decreased Pesticide
Uptake
Decreased Pesticide
Targets
6
Resistance to DDT in Fruit Flies
• Experimental populations
were bred by hybridizing a
resistant and a susceptible
strain of D. melanogaster
• Resistant genes were found
on three chromosomes (2,
3, and X)
• Resistance was cumulative;
the more genes
contributing to resistance,
the greater the resistance
Figure 02: Percent survival of fruit flies
Adapted from Crow, J. F., Ann.
Rev. Entomol., 2 (1957): 227-246.
Peppered Moth Biston betularia
(Linnaeus, 1758)
• The peppered moth occurs in two color phases
(a) Both phases are displayed against an unpolluted, lichen-covered
tree
(b) Both phases are displayed against a dark tree, on which the lichen
were killed by pollution
Peppered Moth Biston betularia
• The typical wildtype white/speckled phenotype is
caused by a recessive allele that must be
homozygous to be expressed
• The recessive allele predominates in wild
population gene pools
• The melanic or black form is caused by a dominant
allele that occurs spontaneously in nature
• Peppered moths rest on trees and depend on
camouflage for protection
Peppered Moth Biston betularia
• In unpolluted areas, trees are
covered in lichens and the
light form of the moth is hard
to see
• In the mid 1800’s, air pollution
in British cities covered trees
with coal dust and soot
• In Victorian era cities, the dark
form became common and
the light form rare
Peppered Moth Biston betularia
• In 1848, 5% of the population were dark colored moths
while 95% were light colored
• In 1895, 98% were dark colored while 2% were light
colored
• In 1995, 19% were dark colored while 81% were light
colored
Peppered Moth Biston betularia
• The melanic phenotype is due to underlying homozygous (BB) and
heterozygous (Bb) dominant genotypes
• In the mid 1950’s, air pollution controls were introduced in Britain
• When smoke pollution decreased in Britain, natural selection acted
very quickly to favor survival of the wild type peppered morphs as
bird predation eliminated melanic forms in progressively less polluted
forests
• The frequency of the melanic form has declined ever since
Bernard Kettlewell (1907-1979) and
Industrial Melanism
Kettlewell performed extensive field
studies in Britain in the 1950s to test
the hypothesis that bird predators
were altering the frequencies of the
color morphs based on the moths’
contrast to their backgrounds, such
as tree bark, when they were at rest
Kettlewell’s Experiments
• Aviary studies demonstrated that native birds
did eat peppered moths
• Release and capture studies in two areas
indicated that peppered forms survived in
greater numbers in unpolluted forests while
melanic forms survived in greater numbers in
soot polluted forests
• Kettlewell also observed birds preying on the
two colors morphs in nature
Kettlewell’s Experiments
• Many other lepidopterists have confirmed
Kettlewell’s conclusions
• Kettlewell continued to work with melanism
throughout his career
• Kettlewell summarized his career’s work in The
Evolution of Melanism: a study of recurring
necessity; with special reference to industrial
melanism in the Lepidoptera (1973)
Kettlewell’s Experiments
• In Bernard Kettlewell’s famous experiment, he
placed moths on dark and pale tree trunks and
showed that background colors strongly influenced
survival
• In the wild, however, moths take much more care
about where they settle and rarely settle on large
tree trunks
• Instead moths usually choose to rest in shady areas
where branches join the trunk
• If the moth’s choice of site is adaptive, then moths
in these positions should be taken less often by
predators than those on open space tree trunks
Peppered Moths
• In an experiment in which dead moths were
pinned to open tree trunks or the underside
of branches birds consumed fewer of those on
the undersides of branches
Peppered Moths
• Other moths also make
very specific choices
about where to rest
• The speckled moth
usually perches head up
with its forewings
covering its body
• When given a choice of
resting site these moths
prefer birch trees
Peppered Moths
• Pietrewicz and Kamil (1977) tested
whether these choices by moths
were selectively advantageous
• They trained blue jays to respond
to slides of moths by pecking a
button for a food reward whenever
they spotted a moth
• Results showed that blue jays
spotted moths less often on birch
trees and especially when a moth
was oriented with its head up
Try TechApps to hunt moths yourself!
Thus, moth’s choices
appear to reduce the
risk of detection by
visually oriented
predators
Kettlewell’s Critics: Scientists
• A minority of serious scientists have criticized
various aspects of Kettlewell’s methodology in
his industrial melanism studies
• But the scientific consensus remains that
Kettlewell‘s work was valid and that Kettlewell
had demonstrated microevolution in action
Kettlewell’s Critics: Others
• British journalist Judith Hooper
attacked Kettlewell in her book,
but her book has been
dismissed by scientists for lack
of scientific understanding,
prejudice and careless
journalism
• Creationists have seized on the
criticisms and claim that not
only was Kettlewell’s work
invalid but also that what he
claimed did not demonstrate
evolution since it did not
document the origin of a new
species
Mutation Rates
• Mutation rate is the probability that a gene will
mutate when the cell divides
• A spontaneous mutation rate for E. coli = 1 in 109
replicated base pairs or 1 in 106 replicated genes
• 1 in a million is easy to acheive in bacterial colonies
• Mutagens increase the mutation rate by 10 - 1000
times to as high as 1 in 103 replicated genes
Mutation Rates
• Though most mutations are harmful, rare superior
mutation rates are advantageous
• The mutation rates observed in nature allow
populations to retain common adaptive phenotypes
while accumulating new alleles which might contribute
to the origin of new features in a few individuals
• A remarkable way in which some organisms accumulate
mutations without experiencing their immediate
effects is to bind their mutant gene products with heat
shock proteins
Heat Shock Proteins (HSPs)
• Heat shock proteins (HSP) are expressed in response to
various biological stresses, including heat, high
pressures, and toxic compounds
• HSPs are among the most abundant cellular proteins
found under non-stress conditions
Heat Shock Proteins (HSPs)
• HSPs include a family of proteins
known as "chaperones," which
are solely dedicated to helping
other proteins fold and assume
their proper functions
• Cells are efficient about getting
the folding right because
misfolded proteins can change
the normal life of the cell
– In some cases change is good, in
others deadly
Heat Shock Proteins in Protein Folding
• As the ribosome moves along
the molecule of messenger
RNA, a chain of amino acids is
built up to form a new protein
molecule
• The chain is protected against
unwanted interactions with
other cytoplasmic molecules
by heat-shock proteins and a
chaperonin molecule until it
has successfully completed its
folding
Source:
(http://www.cs.stedwards.edu/chem/Chemistry/CHEM43/CHEM43/HSP/FUNCTION.HTML)
Heat Shock Proteins (HSPs)
• Heat shock proteins may be disabled in new stressful
environments
–
–
–
–
dramatic change in temperature
dramatic change in pH
dramatic change in salinity
etc.
• When the HSPs are disabled, mutant proteins may develop
into new conformations
• These new shapes may permit new functions, which may
allow natural selection to improve the mutant proteins over
time
Neo-Darwinism and Genetic Polymorphism
• New beneficial mutations seem to be very rare
• Neutral or deleterious but recessive mutations
provide a reservoir of potential genetic
variation
• Genetic variation in a population, where two or
more alleles exist at a locus, is termed a
genetic polymorphism
Genetic Variation in Nature
• Morphology, Physiology, Behavior
– Size, color, shape of cell or body parts,
etc.
– Respiration, digestion, excretion, etc.
– Nutrient acquisition, reproduction,
migration, etc.
• Enzyme polymorphism
– Change in catalytic ability due to change
in temperature, osmotic environment,
pH, etc.
• DNA sequence polymorphism
– Changes in bases, codons, introns,
exons, etc.
29
Neo-Darwinism and Genetic Polymorphism
• In the fruit fly, Drosophila pseudoobscura,
populations in different localities are
polymorphic for a wide variety of gene
arrangements
• Many linked alleles are protected inside
inversion loops
Drosophila pseudoobscura Polymorphisms
• The different inversion
Figure 03B: Chromosomal inversions found at different months
during the year in one locality, Mount San Jacinto, California
Adapted from Dobzhansky, T., Heredity 1 (1947): 53-64.
loops contain different
linked alleles
• Those linked alleles
control different
metabolic phenotypes
• Since a fruit fly lives
only about a month,
different metabolic
phenotypes can be
selected over the course
of a single season
Drosophila pseudoobscura Polymorphisms
Figure 03A: Third chromosome gene
inversions in Drosophila pseudoobscura
Adapted from Dobzhansky, T., Carnegie
Inst. Wash. Publ., 554 (1944): 47-144.
• Those different phenotypes,
whose linked genes are
protected in the inversion
loops, are also adapted to
different climates in
different locations
• The different locations differ
in average temperatures,
average rainfall, altitude,
and other environmental
factors
Genetic Polymorphism
• Genetic polymorphism provides a much
greater source of genetic variation than do the
relatively few new mutations that arise each
generation
• Now let’s look again at the nature of genetic
variation
Polygenic Inheritance
• Polygenic inheritance, also known as quantitative
or multifactorial inheritance, refers to inheritance
of a phenotypic characteristic (trait) that is
attributable to two or more genes, or to the
interaction with the environment, or both
• Unlike monogenic traits, polygenic traits do not
follow patterns of simple Mendelian inheritance
(discontinuous traits)
• Instead, their phenotypes typically vary along a
continuous gradient depicted by a bell-shaped
curve (a normal distribution)
Continuous Variation
• Small heritable
changes provide most
of the variation on
which natural
selection acts
Figure 04: Heights of 1,000 Harvard
College students aged 18 to 25
Adapted from Castle, W. E. Genetics and Eugenics, Fourth
edition. Harvard University Press, 1932.
Continuous Variation
It is almost impossible for a single gene
locus to control continuous variation
Population Genetics
• The study of genes and
genotypes in a population
• We want to know the
extent of genetic
variation, why it exists
and how it changes over
time
• This knowledge helps us
to understand how
genetic variation is
related to phenotypic
variation
Gene Pool
• The gene pool is all of the
genes and different alleles
in a population
• We study genetic variation
within the gene pool and
how genetic variation
changes from one
generation to the next
Emphasis is often on variation in alleles between
members of a population at certain loci of interest
Populations
• Group of individuals of the same species that can
interbreed with one another
• Some species occupy a wide geographic range and
are divided into discrete populations (demes)
Genes in Natural Populations
Are Usually Polymorphic
• Polymorphism – many phenotypic traits display variation
within a population
– Due to 2 or more alleles at a locus that influence a phenotype
• Polymorphic gene/locus - 2 or more alleles
• Monomorphic gene/locus– predominantly a single allele
[“fixed” locus]
• Single nucleotide polymorphism (SNPs)
– Smallest type of genetic change in a gene; a point mutation
– Most common – 90% of the variation in human gene sequences
• Large, healthy populations exhibit a high level of genetic
diversity
• Polymorphisms are the raw material for evolution
Quantitative Trait Loci (QTL)
• The term quantitative trait
loci (QTL) is shorthand for
all of the loci or genes
(alleles) in a particular
region of a chromosome
that affect a quantitative
aspect of the phenotype
• In sticklebacks,
Gasterosteus aculeatus,
there is variation in the
pelvic girdle and in dermal
plates
Figure 06: Three spine sticklebacks
Reproduced from Trends in Ecology & Evolution, 19(9), Foster,
S. A, and Baker, J. A., Evolution in parallel…, pp. 456–459,
copyright 2004, with permission from Elsevier. Photographs
courtesy of Dr. W. A. Cresko, University of Oregon.
QTL Mapping
• Statistical analysis is required to demonstrate that different
genes interact with one another and to determine whether
they produce a significant effect on the phenotype
• QTL mapping identifies a particular region of the genome
containing a gene that is associated with the trait being
measured
• QTLs are shown as intervals across a chromosome where the
probability of association is plotted for each marker used in
the mapping experiment
• The QTL techniques were developed in the late 1980s and can
be performed on inbred strains of any species
• QTL analysis allows scientists to quantify the contributions of
both heredity and environment to polygenic traits
Population Genetics
• In our introductory course, we are not going
to explore the molecular details of QTL
analysis
• Instead, we will examine the foundation of
population genetics, the Hardy-Weinberg
Equilibrium
Population Genetics and Gene (Allele)
Frequencies in Populations
• The Neo-Darwinian theory
– Evolution is a population phenomenon
– Evolution is a change in gene (now allele) frequencies
in a population because of various natural forces such
as mutation, selection, migration, or genetic drift
– These changes in allele frequencies lead to differences
among populations, species, and higher clades
– This population genetics view of evolution became
known as Neo-Darwinian theory with its emphasis on
the frequency of genes and alleles in populations
Allele Frequencies
• A population’s gene pool includes all the alleles
for all the loci present in the population
• Diploid organisms have a maximum of two
different alleles at each genetic locus
• Typically, a single individual therefore has only a
small fraction of the alleles for a given locus that
are present in the population as a whole
Allele and Genotype Frequencies
• Related but distinct calculations
• Population 1 has 5 homozygous dominant and 5 heterozygous
individuals
• Population 2 has 7 homozygous dominant, 1 heterozygous,
and 2 homozygous recessive individuals
• Both populations have identical allele frequencies, 15 A : 5 a =
75% A and 25% a alleles
The Hardy-Weinberg Principle
Godfrey H. Hardy: English mathematician (1903)
Wilhelm Weinberg: German physician (1908)
W.E. Castle: American geneticist (1908) (left out)
Working independently just a few years after the
rediscovery of Mendelian genetics they concluded
that:
The original proportions of the allele frequencies in a
population remain constant from generation to
generation as long as five assumptions are met
47
The Hardy-Weinberg Principle
Five H-W Equilibrium assumptions: If:
1. The population size is very large
2. Random mating is occurring
3. No mutation occurs
4. No selection occurs
5. No alleles transfer in or out of the
population (no migration occurs)
Then allele frequencies in the population will
remain constant through future generations
48
Simplifying Assumptions for
The Hardy-Weinberg Principle
1) diploid organisms
2) sexual reproducing organisms
3) generations are non-overlapping
4) all genotypes are equally viable
• If these simplifying assumptions are not met, it
complicates the mathematics for the analyses
• Whether or not these assumptions are all met,
biologists can use Mendelian ratios and HardyWeinberg analysis to measure rates of evolution
The Hardy-Weinberg Principle
• p = frequency for first allele in the population’s
gene pool
• q = frequency for second allele in the population’s
gene pool
• Calculate allele frequencies with a binomial
equation:
p+q=1
• because there are only two alleles:
p + q must always equal 1 (100% of the alleles)
• [Note: more alleles can be handled, e.g., with three alleles: p + q + r = 1]
50
The Hardy-Weinberg Principle
Calculate genotype frequencies with a binomial
expansion
(p+q)2 = p2 + 2pq + q2 = 1
• p2 = individuals homozygous for first allele
• 2pq = individuals heterozygous for the alleles
• q2 = individuals homozygous for second allele
• because there are three phenotypic classes:
p2 + 2pq + q2 must always equal 1
51
The Hardy-Weinberg Principle
p = 0.6 and q = 0.4 and therefore p + q = 1.0
52
Product Law of Probability
Product Law of Probability: The probability of two
independent events occurring simultaneously is equal to
the product of their separate probabilities
Male gamete production is independent of female gamete production
53
The Hardy-Weinberg Principle
assume 100
viable offspring
36 BB and 48 Bb offspring have (36 + 36 + 24 + 24 = ) 120 B alleles; 48 Bb and 16 bb have
(24 + 24 + 16 + 16 = ) 80 b alleles. Freq of B = 124/200 = 0.6 and freq. of b = 80/200 = 0.4.
use the Hardy-Weinberg equation to predict frequencies in subsequent generations
54
The Hardy-Weinberg Equilibrium
If:
1.
2.
3.
4.
5.
The population size is very large
Random mating is occurring
No mutation occurs
No selection occurs
No alleles transfer in or out of the
population (no migration)
Then allele frequencies in the population will
remain constant through generations
55
Another Example
• 49 red-flowered RR
• 42 pink-flowered Rr
• 9 white-flowered rr
• [100 diploid individuals
carry 200 alleles]
49 RR and 42 Rr offspring have
(49 + 49 + 42 = ) 140 R alleles
42 Rr and 9 rr have
(42 + 9 + 9 = ) 60 r alleles
Freq of R = 140/200 = 0.7 and freq.
of r = 60/200 = 0.3
No allele freq. change in the F1
The Hardy-Weinberg Equilibrium
• Relates allele and genotype frequencies
under certain limiting conditions
p2 + 2pq + q2 = 1 (the Hardy-Weinberg Equation)
If we apply the equation to the flower color gene, then:
p2 = the genotype frequency of RR
2pq = the genotype frequency of Rr
q2 = the genotype frequency of rr
If p = 0.7 and q = 0.3, then:
Frequency of RR individuals = p2 = (0.7) 2 = 0.49
Frequency of Rr individuals = 2pq = (2)(0.7)(0.3) = 0.42
Frequency of rr individuals = q2 = (0.3) 2 = 0.09
The Hardy-Weinberg Equilibrium
[In H-W equilibrium,
heterozygotes will
never be greater
than 50% of the
population]
• Hardy–Weinberg proportions for two alleles: the horizontal axis shows the two
allele frequencies p and q and the vertical axis shows the expected genotype
frequencies
• Each line shows one of the three possible genotypes and their relative
frequencies
The Hardy-Weinberg Principles Describes a
Population at Genetic Equilibrium
Genetic equilibrium requires:
1.
2.
3.
4.
5.
The population size is very large
Random mating is occurring
No mutation occurs
No selection occurs
No alleles transfer in or out of the population
(no migration occurs)
Five Agents of Evolutionary Change
A population not in Hardy-Weinberg equilibrium is one in
which allele frequencies are changing generation to
generation due to one or more of the five evolutionary agents
that are operating in the population
60
Agents of Evolutionary Change
• Small Population Size:
• When a population is large, then allele frequencies are very
unlikely to change due to random sampling error
• When a population is small, then, just by chance, some individuals
fail to mate at all, not because they are unfit
• When a population is small, then, just by chance, some offspring
fail to survive to reproduce, not because they are unfit
• When a population is small, gene frequencies may change due to
these sorts of random effects – this is called genetic drift
California condors
Genetic Drift
• Genetic drift: Random fluctuations in allele
frequencies over time due to chance events
• important in small populations
• founder effect – a few individuals
found a new population (with a
small allelic pool)
• bottleneck effect – a drastic
reduction in population, and
gene pool size and complexity
DNA studies indicate that polar bears have suffered repeated bottleneck
events when the arctic climate warmed and also repeated hybridization
62
causing gene introgression (HGT) from their sister group, the brown bears
Genetic Drift
Figure 08: Numbers of brown (bw75) alleles
in 107 populations of D. melanogaster
• In small populations, allele frequency
changes that have no predictable constancy
or direction from generation to generation
• Genetic drift is a consequence of random
fluctuations in gene frequencies that arise in
small populations
• In this experiment, 107 populations were
established with p = q = 0.5 for brown
(bw/recessive) and red (+/wildtype/
dominant) eye alleles, but at each
generation, only 16 parents were drawn, at
random, to produce the next generation
(+/+)
• By generation 19, brown eye had been
eliminated from 30 populations and fixed at
100% in another 28 populations.
(bw/bw)
Data from Buri, P., Evolution 10 (1956): 367-402.
Table 13.1: Definitions of Genetic Drift
and Comments on Them
Regardless of the definition, genetic drift tends to increase variation
between populations, but in no particular direction, including not
necessarily to increase the fitness of the population
Agents of Evolutionary Change
• Random Mating is required for the Hardy-Weinberg
Equilibrium
• The members of the population mate with each other without
regard to their phenotypes and genotypes
• The members of the population are (relatively) equally likely to
mate with any other member of the population of the opposite
sex, i.e., have relatively equal access to all members of the
population
• Humans mate without regard to ABO and Rh blood types
– This example shows that a particular gene can meet the H-W equilibrium
criteria, even though the species as a whole does not
• Wind-pollinated plant species and many aquatic species who
release their eggs and sperm into the water
• Animals who are members of large mobile schools, herds or flocks
65
Agents of Evolutionary Change
• Nonrandom Mating: mating between specific
genotypes shifts genotype frequencies
Non-Random mating could be either like
with like; also called Assortative Mating or
opposites attracting each other; also
called Non-Assortative Mating
66
Agents of Evolutionary Change
• Classification of mating systems
– Monogamy, polygamy, polyandry
(Darwin 1871)
– Monogamy, resource defense
polygyny, harem defense polygyny,
explosive mating assemblage, leks,
female access polyandry, etc. (Emlen
& Oring 1977)
– Promiscuous
– Self pollination/fertilization
– Asexual reproduction
• Apomixis: parthenogenesis in animals
and apogamy in plants
67
Agents of Evolutionary Change
• Nonrandom Mating: mating between specific
genotypes shifts genotype frequencies
• Ladies, would you prefer to mate and produce
offspring with one of these males over
another? If so, you are practicing non-random
mating based on phenotypic characteristics
68
Agents of Evolutionary Change
• Nonrandom Mating: mating between specific
genotypes shifts genotype and phenotype frequencies
– Assortative Mating: does not change frequency of individual
alleles; increases the proportion of homozygous individuals
– Disassortative Mating: phenotypically different individuals
mate; produces an excess of heterozygotes
Assortative Mating
Selfing Inbreeding
Random Mating
Disassortative Mating
Obligate Outcrossing
Less Genetic Diversity
Hardy-Weinberg More Genetic Diversity
More Homozygotes
Equilibrium
More Heterozygotes
Clones/Clonal Lineages
Many Genotypes
More Uniformly Fit Individuals
Individuals of Varying Fitness
Less Potential to Adapt to Change More Potential to Adapt to Change
69
Inbreeding
• Mating between relatives or selfing in plants
– Inevitable in smaller populations
– Occurs in nature because of proximity of relatives –
example: natural stands of tree whose relatives are
nearby because of limited seed dispersion
• If there is no natural selection, an inbreeding
population will acquire an increase in the
frequency of homozygotes without a change in
allele frequency in the population
Coefficient of Inbreeding
Isonymy: having the same surname from
both parents, an estimate of inbreeding in
population records, e.g., birth certificates
Coefficient of Inbreeding
10 children
Three of Darwin's 10 children died in childhood, while another three never had any
children of their own, despite being married for years
Agents of Evolutionary Change
• Mutation: Changes in a cell’s DNA
– Mutation is the ultimate source of
genetic variation
– Since a new mutation transforms an
allele into a different allele, it must
also change allele frequencies
– Mutation rates are generally so low
that they have little effect on
Hardy-Weinberg proportions of
common alleles in the short term,
over a few hundred generations
73
Agents of Evolutionary Change
Variation from
1/10000 to
1/1000000000
Agents of Evolutionary Change
• Mutation: Changes in a cell’s DNA
– Mutations are usually deleterious
Stumpy
Stumpy, born 2007, in UK,
on a duck farm, who
eventually lost the two
extra legs to accidental
traumas, and his
companion, Alice
75
Agents of Evolutionary Change
• Natural selection: environmental conditions
determine which individuals in a population
produce the most offspring
• Three conditions are required for natural
selection to occur:
– Variation must exist among individuals in a
population
– Variation among individuals must result in differences
in the number of offspring surviving
– Variation must be genetically inherited
• We will return to natural selection in Chapter 15
76
Agents of Evolutionary Change
• Gene Flow: A movement of alleles from one
population to another
– Migration of individuals or gametes between
populations
– Migration can be a powerful agent for evolutionary
change
– Migration tends to homogenize allele frequencies
between populations
– But migration is adding or removing alleles from
the gene pool, so migration is going to change gene
frequencies in the populations experiencing
immigration or emigration
77
Agents of Evolutionary Change
Migration – the movement of breeding individuals into or
out of isolated populations – results in evolutionary change
because alleles move with the individuals. We call this
movement gene flow.
• If enough migration occurs, the original isolates, with
their inherent limited genetic variability, may fuse and
form a new larger population with increased genetic
variability.
Hardy-Weinberg Equation
• Relates allele and genotype frequencies under
certain conditions
• p2 + 2pq + q2 = 1
• p = frequency of the dominant allele
• q = frequency of the recessive allele
– The genotype frequencies of a population are
– p2 is frequency of homozygous dominant genotype
– 2pq is frequency of heterozygous genotype
– q2 is frequency of homozygous recessive genotype
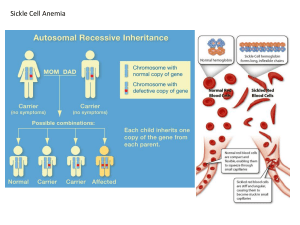
Sickle-Cell Anemia
• In sickle-cell anemia, hemoglobin (Hbs) has poor
oxygen affinity
• Sequencing of the hemoglobin gene revealed one
change from the normal amino acid sequence:
Sickle Cell Anemia
Normal globin gene
(H)
GAG (glutamate)
GTG (valine)
Sickle cell gene
(h)
Hh
Hh
HH
hh
Genotypes
Hh
Sickle-Cell Anemia
Evolution of populations is best understood
in terms of frequencies:
1. Phenotypes 2. Alleles 3. Genotypes
Phenotypes Gene
Alleles Genotypes
Normal
hemoglobin
hemoglobin
H
HH, Hh
Sickle Cell
hemoglobin
h
hh
Sickle-Cell Anemia
Actual Phenotype Frequencies: Sickle Cell
Anemia in the African American Population
Phenotype
Normal
Sickle Cell
Disease
Total # African
Americans
Homozygous
Dominants &
Carriers
Recessive
# people
29.94M
Phenotype
Freq.
0.998
0.06M
0.002
30M
1.000
Sickle-Cell Anemia
Actual Genotype Frequencies: Sickle Cell Anemia
in the African American Population
Phenotype
Genotype
# People
Genotype
freq
Dominant
HH
27.4M
0.915
Dominant
Hh
2.5M
0.083
Recessive
hh
0.06M
0.002
Total # African
Americans
30M
Sickle-Cell Anemia
H-W Expected Frequencies: Sickle Cell Anemia in
the African American Population
Genotype
H-W Expected
Phenotype
Frequency
Actual Phenotype
Frequency
Allele
Allele Freq
HH
0.996004
0.915 ↓
H
0.998
Hh
0.003992
0.083 ↑
hh
0.000004
0.002 ↑
h
0.002
If p (0.998) + q (0.002) = 1
Hardy–Weinberg Principle
• According to the Hardy–
Weinberg principle, in a
population of randomly
mating individuals, allele
frequencies are conserved
and in equilibrium unless
external forces act on them
• What is going on with Sickle
Cell Anemia in AfricanAmericans?
• [~400 years is only 16
human generations or so]
Figure 07: Hardy–
Weinberg equilibrium
Adapted from Falconer, D. S. and T. F. C Mackay. Introduction
to Quantitative Genetics, Fourth edition. Longman, 1996.
Sickle-Cell Anemia
1. Is the population size is very
large?
2. Is random mating is
occurring?
3. Is mutation significant (over
20-30 generations since
slave importation began)?
4. Is selection occurring?
5. Are alleles transferring in or
out of the population
(migration)?
1. Yes! 30M
2. Yes, as far as HgS is
concerned
3. No
[estimated at 5x10-8]
4. Yes; but against
homozygotes
5. Yes, from African
populations with a
higher incidence of HgS
Sickle-Cell Anemia’s
Heterozygote Advantage
• The recessive sickle-cell allele
produces hemoglobin with reduced
capacity to carry oxygen
• This mutation also confers malaria
resistance in heterozygotes
• This heterozygote advantage leads
to a larger proportion of the
recessive allele than usual in areas
where malaria is widespread
• These populations exhibit balanced
polymorphism between the
mutant and wild-type alleles
The Hardy-Weinberg Equilibrium
If a population meets all the assumptions of the HardyWeinberg Principle, then allele frequencies in the
population will remain constant through future generations
1.
2.
3.
4.
5.
The population size is very large
Random mating is occurring
No mutation occurs
No selection occurs
No alleles transfer in or out of the population (no migration)
A population in true Hardy-Weinberg equilibrium
is rarely seen in nature!
Additional Sources of
Variation in Populations
• The extent to which a population departs from
an optimal genetic constitution is called its
genetic load, and is marked by the loss of
some individuals through genetic death
• However, the presence of genetic variability in
a population can be considered a sort of
“insurance policy” for sexually reproducing
species (and HGT is similar for asexual species)
Additional Sources of
Variation in Populations
• On the other hand, there is, potentially, a major cost to
the less fit in the variable/polymorphic population
• In technologically advanced societies, we are actually
increasing the genetic load with modern medicine and
pharmacology:
– Diabetes mellitus
– Atherosclerosis
– Respiratory diseases
– Susceptibility to a variety of microbial pathogens
– Etc.
Populations, Allele Frequencies
and the Gene Pool
• A population defined as a group of
sexually interbreeding or potentially
interbreeding individuals
• A deme is a locally interbreeding
subset of the population
• The deme is most often the focal
point for evolutionary change
• Consider the California yarrow,
Achillea borealis and A. lanulosa
Figure B01B:
A. borealis
Figure B01C:
A. lanulosa
© Sergey Chushkin/
ShutterStock, Inc.
© Brzostowska/
ShutterStock, Inc.
Demes Can Adapt to Local Conditions
Average height, timing and length of growing
season, temperature and drought tolerance varied
among the populations on an east-west transect
Figure B01A: Locations of the populations and indication of genetic differences that have evolved
Adapted from Clausen, J. D., D. Keck, and W. M. Hiesey,
Carnegie Inst. Wash. Publ., No. 581, 1948: 1–219.
Demes Have Different Gene Pools
Figure B02: Responses of clones from
representative of the common yarrow
Adapted from Clausen, J. D., D. Keck, and W. M.
Hiesey, Carnegie Inst. Wash. Publ. No. 581, 1–219.
• Individual yarrow plants were
drawn from five different
locales, shoots were cut and
grown vegetatively into mature
individuals
• These clones of individuals
were then planted at different
locations
• They showed considerable
differences in fitness; some
even dying in a different locale
Gene Flow / Gene Migration
• At least three factors have an impact on the recipient
population:
1. the difference in gene frequencies between the two
populations;
2. the proportion of migrating genes incorporated into each
generation and its gene pool; and
3. the pattern of gene flow, whether occurring once or
continually over time
– Gene flow can hinder local evolutionary changes by
infusing genes from distant populations that are not
so well adapted to local conditions
– Example: global blood group gene frequencies
Global Blood Group Gene Frequencies
O
A
The original colonists of North
America were a small group of
apparently O+ founders; Caucasian
Type A individuals may have
originated in Scandinavia; Type B
individuals seem to have originated
in Central Asia
B
Different ABO blood group phenotypes may have little to do with variation in O2 transport, but
are correlated with other factors such as disease susceptibility, e.g., Type A individuals seem to
be less resistant to smallpox
Selection, Variation and Increased Fitness
• R. A. Fisher, one of the founders of population
genetics noted that the greater the genetic
variation upon which selection for fitness may
act, the greater the expected improvement in
fitness
• Variation itself is subject to selection, and so
the propensity to vary (variability) is an
important attribute of organisms
Phylogeography
• One tool for reconstructing the geographical
history of a lineage uses knowledge of
genetics to plot variation in allele frequencies
on the distribution map of the demes or
populations of a species
Distribution of
Mycobacterium
tuberculosis strains in
human populations
Phylogeography
Figure 09: Asian common wheat
Adapted from Ghimire, S. K., Y. Akashi, C. Maitani, M.
Nakanishi and K. Kato, Breeding Sci., 55 (2005): 75-185.
The pie charts record
frequencies of two isozyme
alleles, probably the tan
representing a more cold
and drought resilient form
than the brown
Figure 10: Asian common wheat
Adapted from Ghimire, S. K., Y. Akashi, C. Maitani, M.
Nakanishi and K. Kato, Breeding Sci., 55 (2005): 75-185.
• Another example: the spread of wheat
– Strains of the Asian common wheat, Triticum
aestivu, reflect adaptations of a single species to a
large number of environments
Why Doesn’t Natural Selection Eliminate
All Genetic Variation in Populations?
• Natural selection tends to reduce variability in
populations by eliminating less fit alleles
• Mechanisms which counteract that elimination to
preserve genetic variation include:
– The diploid condition preserves variation by “hiding”
recessive alleles (Bb)
– Balanced polymorphisms (2 or more phenotypes are
stable in the population) may result from:
1. heterozygote advantage: Aa superior to aa and AA
2. frequency-dependent selection
3. variation within the environment for a population
Frequency-Dependent Selection
• Frequency-dependent selection is the term given to an evolutionary
process where the fitness of a phenotype is dependent on its
frequency relative to other phenotypes in a given population
• In positive frequency-dependent selection, the fitness of a
phenotype increases as it becomes more common
• In negative frequency-dependent selection, the fitness of a
phenotype increases as it becomes rarer (this is an example of
balancing selection)
• Frequency-dependent selection is usually the result of interactions
between species (predation, parasitism, or competition) or between
genotypes within species (usually competitive or symbiotic), and has
been especially frequently discussed with relation to anti-predator
adaptations
negative frequency-dependent
selection in fruit flies
Chapter 13
End
Genetic Polymorphism