BIOC3800 Sensory Transducers
advertisement

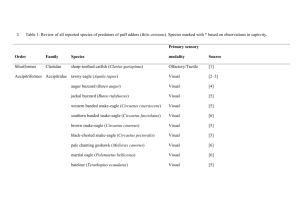
Taste circumvallate papilla labelled lines • There are five recognised modalities: sweet, sour, bitter, salt & umami. • All mature taste bud cells have prominent microvilli. • Taste bud cells are continuously renewed from a local population of stem cells and their half-life is about ten days. • Type I "dark" cells are glial (supporting) cells, but also contribute to salty (chloride) tastes. The full salt flavour may also require sodium channels. • Sub-populations of type II cells respond to sweet, bitter, umami (glutamate) and possibly fats. They release ATP which stimulates purinergic receptors on type III cells and also on sensory neurons. • Type III cells signal "sour" after intracellular acidification. Only type III cells make synaptic connections, where they release serotonin. • The sensory nerves are “labelled lines” but it is not known how they specifically communicate with the appropriate sensory cells. Taste bud Recognised tastes & receptors • • • • • Sour – PKD1L3 or PKD2L1 (disputed) Salty – ENaC or TRPV1 (disputed) Bitter – T2R family – about 30 members Sweet – T1R2/T1R3 heterodimer Umami – T1R1/T1R3 heterodimer – plus other glutamate receptors? • Fats? • In addition to the taste buds, there is a closely-related family of entero-endocrine cells throughout the GI tract, which use similar transduction mechanisms to control food intake, digestion and incretin release. Bitter, sweet & umami taste transduction 9 gustatory adaptation • It is difficult to study sensory adaptation to continuing taste stimulation and there are very few published reports. • It has recently become apparent that taste sensitivity is modulated by external factors, and this has become a prolific area for research. • Leptin, a cytokine produced by adipocytes (which reduces food intake and increases energy expenditure) inhibits the taste bud response to saccharin. • Glucagon (which is produced in response to hypoglycaemia) increases the taste bud response to sucrose, while the incretin hormone GLP-1 maintains and enhances the response to sweet stimuli. • Ghrelin increases the response to salty and sour tastants, and oxytocin also seems likely to modulate these responses. • Taste buds also respond to GABA, and it seems likely that complex networks of signalling molecules will be found to underpin taste bud physiology, just as they underpin other areas of the GI tract. Smell olfactory signal transduction – 1 • Vertebrate odorant receptors (ORs) all belong to the immunoglobulin superfamily. • Some mammals may have two thousand OR genes. • Species (e.g. humans) with a poor sense of smell, have fewer OR genes, and many of these are inactive pseudogenes. • Each OR can bind a range of molecules, but each gene has a unique pattern of binding affinities. • Human olfactory receptor neurons (ORNs) live for about two weeks and are replenished from a local population of stem cells. • Each ORN expresses one specific OR on about 20 non-motile cilia. • Olfactory cilia cannot be properly described as primary cilia because there is more than one per cell, but they are similar. • All the ORNs expressing a particular OR converge onto one or two matching olfactory glomeruli within the olfactory bulb. olfactory signal transduction – 2 • ORs are G protein-coupled-receptors (GPCRs) which activate adenyl cyclase type-III via the G protein, Golf • cAMP opens a nonselective, cyclic-nucleotide-gated (CNG) cation channel, depolarising the ciliary membrane. • ORNs express a Na/K/Cl cotransporter (NKCC10) which maintains a high intracellular [chloride] • Ca2+ influx through the CNG channels opens Ca2+-activated chloride channels on the ciliary membrane • Cl- leaves the ORN down its concentration gradient, which further depolarizes the cell and provides amplification. • Internal Ca2+ binds to calmodulin which initiates multiple negativefeedback pathways that facilitate adaptation. • Action-potential generation in ORNs involves voltage-gated sodium channels and low voltage-activated (T-type) calcium channels. olfactory signal transduction – 3 • Mammalian smell has great sensitivity and a very wide adaptation range, as might be expected for “outward facing” receptors. • Adaptation in mammals occurs by a variety of mechanisms with very different time scales, and their relative importance is disputed. • Olfactory adaptation in mammals may involve local negative feedback loops with calcium + calmodulin (see previous diagram). • Increased calcium export from ORNs may allow mammals to cope with sustained stimulation by high odorant concentrations. • Increased phosphodiesterase activity may help to remove excessive concentrations of cyclic AMP. • In addition, mammalian olfactory receptors may be phosphorylated, and / or internalised, and / or uncoupled from their G-proteins as part of the adaptation system. • Expression of mammalian OR genes may be downregulated as part of a long-term adaptation mechanism. olfactory signal transduction – 4 • Odorants produce a graded concentration-dependent depolarisation within mammalian olfactory cilia – an analogue response. • Spikes are superimposed on the receptor potential within the cell body of the ORN – analogue to digital conversion. • The axons from the ORNs to the corresponding glomerulus carry fully digital action potentials. • The glomeruli are arranged in a fixed, inherited, spatial map, but related odours are not associated within the olfactory bulb. • Olfactory processing involves contrast enhancement between different odours, similar in some respects to retinal processing. • Calculation of time derivatives d(smell)/dt and d2(smell)/dt2 is probably important for following odorant trails. • Sniffing and head movements provide mammals with additional information about the source of the smell. Olfactory and retinal processing Accessory olfactory areas • In addition to the main olfactory area, many species have various accessory areas, such as the vomeronasal organ, which are specialised for the reception of social and sexual smells. • Their function seems to be linked to the MHC system, with the result that many mammals prefer sexual partners that will optimise the immune capabilities of their offspring. It is disputed whether a similar system operates in humans. • MHC proteins and a subset of ORs are also expressed in sperm, where they may participate in sperm chemotaxis, thereby ensuring that eggs are more likely to be fertilised by an immunologically compatible sperm. more slides to be added here • There is one recent paper and three important recently published reviews, all of which are highly recommended. I am preparing slides from these, but this is taking me longer than intended. In any event, you ought to read them: • Kaupp (2010) Olfactory signalling in vertebrates and insects: differences and commonalities. Nature Reviews Neuroscience 11, 188-200. • Sibbering & Benton (2010) Ionotropic and metabotropic mechanisms in chemoreception: ‘chance or design’? EMBO reports 11, 173-179 • Kato & Touhara (2009) Mammalian olfactory receptors: pharmacology, G protein coupling and desensitization. Cell. Mol. Life Sci. 66, 3743-3753 • Pluznick et al (2009) Functional expression of the olfactory signaling system in the kidney. PNAS 106, 2059-2064. Sibbering & Benton (2010) Ionotropic and metabotropic mechanisms in chemoreception: ‘chance or design’? EMBO reports 11, 173-179 Mammals have metabotropic ligand binding receptors with second messenger signalling cascades, which indirectly activate ion channels, whereas insects use ionotropic receptors which are directly gated by the chemical stimuli. Kaupp (2010) Olfactory signalling in vertebrates and insects: differences and commonalities. Nature Reviews Neuroscience 11, 188-200. Key for next slide: The vertebrate nasal cavity contains several olfactory subsystems: the main olfactory epithelium (MOE), the vomeronasal organ (VNO), the Grüneberg ganglion (GG), the septal organ (SO) and guanylate cyclase D-containing cells (GCDs) in the MOE. Sensory cells of the MOE and the SO project axons to glomeruli of the main olfactory bulb (MOB). Sensory cells of the GG and GCDs in the MOE send their axons to the necklace glomeruli (NG). Sensory cells of the VNO send their axons into the accessory olfactory bulb (AOB). Olfactory receptor neurons (ORNs) in the MOE (right) have one dendrite, which ends in a dendritic knob. From each dendritic knob, approximately 15 cilia extend into the nasal mucus. ORNs are surrounded by supporting cells and are constantly generated from basal cells. Kaupp (2010) Nature Reviews Neuroscience 11, 188-200. from Kaupp (2010) Nature Reviews Neuroscience 11, 188-200. mammalian transduction mammalian adaptation Kaupp (2010) fig 3 Figure 4 from Kraup (2010) Competing models of insect OR signalling. Model “a” is considered more likely.