PPT - CCRG
advertisement

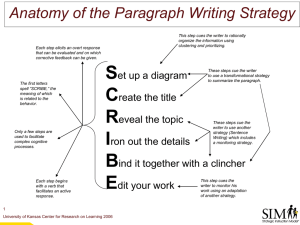
Neural Synchrony in Attention and Consciousness Lawrence M. Ward University of British Columbia Collaborators: Sam Doesburg, Kei Kitajo, Alexa Roggeveen, Tony Herdman, Jessica Green, John J. McDonald Funded by Themes Neural synchrony and its measurement Neural synchrony in attention: batonpassing in the cerebral cortex Neural synchrony in consciousness: binocular rivalry and the thalamic dynamic core Implications for LIDA Gray & Singer’s cats Neural synchrony occurs when neural activity, spiking or dendritic currents, in disparate locations rises and falls in a fixed relationship Ward et al’s humans Varela et al, 2001 Spectral power in a given frequency range reflects local synchrony Theta 6 Hz Spectral Power 10 20 30 40 50 Frequency Gamma 40 Hz Spectral Power 10 20 30 40 50 Frequency Alpha 10 Hz Spectral Power 10 20 30 Frequency 40 50 Roles of local neural synchrony High fidelity neural communication Perceptual/memorial/motor…. Binding: Increase 30-70 Hz: amplify post-synaptic effect by increasing spike cooccurrence (Fries et al, 2001) Decrease 6-15 Hz: increase post-synaptic impact by avoiding spikefrequency adaptation (Fries et al, 2001) Fries 2005 Roles of global neural synchrony Integration of neural activity through reciprocal (re-entrant) interactions between diverse brain regions (e.g., Varela et al, 2001) Exchange of data (upward) and hypotheses/templates (downward); sensory/perceptual processing Modulation of one region (e.g., hippocampus) by another (e.g., visual cortex) to store a memory (e.g., of a visual scene) Modulation of one region (e.g., visual cortex) by another (e.g., prefrontal cortex) to enhance processing of attended information (e.g., a sign for a sushi bar) Initiating an action in motor regions by computations from cognitive regions Consciousness (?) EEG/MEG synchronization analysis: calculation of phase locking value (PLV) i.e. surface Laplacian, or MEG field strength, Step.1 Obtain SCD^of filtered signals f(t) via bandpass filtering at chosen frequencies (µV) Fp1 F7 F3 T3 Fp2 ● F4 F8 30 Fz EEG (C3, O1, Fz) 10Hz 10 20Hz 0 ● C3 Cz -0.2 C4 T4 -10 0 0.2 0.4 0.6 0.8 1 (sec) 30Hz -20 T5 P3 Pz P4 T6 O1 ● SCD 20 C3 O1 Fz -30 stimulus O2 40Hz -0.2 0 stimulus 0.2 0.4 0.6 0.8 1 (sec) Step.2 instantaneous phase and amplitude 3 amplitude 30Hz (C3, O1, Fz) 2 3 30Hz (C3, O1, Fz) 2 1 1 -0.2 0 0 -1 0 0.2 0.4 0.6 0.8 1 (sec) -0.2 -2 -3 stimulus C3 O1 Fz -0.2 0 4 3 2 1 0 -1 0 -2 -3 -4 0.2 0.4 0.6 0.8 1 (sec) phase 30Hz (C3, O1, Fz) 0.2 0.4 0.6 0.8 1 (sec) ~ (t ) f (t ) i f t At expi t amplitude phase f ( ) ~ ~ 1 d where f (t ) is Hilbert transformof f (t ), f (t ) P.V . t Step.3 Calculation of phase locking value (PLV) for each time point 4 3 2 1 0 -1 0 -2 -3 -4 -0.2 PLV t 1, 2 1 N N e i (t ,n ) n 1 where (t , n) 1 (t , n) 2 (t , n) phase difference (30Hz) C3-O1 C3-Fz O1-Fz (sec) 0.2 0.4 ⇒ 0.6 0.8 1 complete synchronization:1 random phase difference:0 (phasedifference) N : the number of trials t : timepoints -0.2 1 : the phaseof thesignal fromelectrode1 2 : thephaseof thesignal fromelectrode2 4 3 2 1 0 -1 0 -2 -3 -4 phase difference (5 trials) C3-O1 (30Hz) (sec) 0.2 0.4 High PLV 1 C3-O1 (30Hz) 0.6 0.8 Low PLV 1 PLV (50 trials) 0.8 0.6 0.4 0.2 (sec) 0 -0.2 0 0.2 0.4 0.6 0.8 1 Step.4 standardization of PLV To reduce the effect of volume conduction of stable sources and compare between electrode pairs at different distances Standardized PLV 6 C3-O1 (30Hz) 4 2 -0.2 0 -2 0 0.2 0.4 0.6 0.8 1 (sec) -4 PLV PLVBmean PLVz(t ) -6 PLVBsd PLVBmean : themean of PLVin thebaseline period(400ms) PLVBsd : thestandarddeviationof PLVin thebaseline period(400ms) Standardized PLV and surrogate PLV 6 PLV (original) C3-O1 (30Hz) 4 Median PLVsurrogate 2 Step.5 statistical test using surrogate data -0.2 0 -2 0 0.2 0.4 0.6 0.8 -4 Note: Local and longrange PLVz must change together for spurious synchronization to be indicated (Doesburg,Ward, CC, 2007) significant PLV increase -6 (Hz) C3-O1 60 100 99 98 3-97 2 1 0 50 40 30 20 10 -0.2 1 ±95 percentle (sec) PLVsurrogate 0 0.2 0.4 0.6 0.8 1(sec) sync desync Alpha, gamma and attention Local alpha power associated with active suppression Local gamma power associated with active processing Alpha suppression necessary for gamma binding? Roles of long-range alpha and gamma synchrony? Coordination of local and long-range synchronization? Theta-modulated gamma synchrony e.g., Klimesch, Jensen & Colgin, Palva & Palva, Ward Alpha/gamma local synchrony (power) indexes spatial attention orienting •Arrow cued box to attend to •Press button only if + in attended box, not if x nor if in unattended box •Cue-target SOA 1000-1200 ms •Cue onset at 0 ms in figures 15 + 20 P7 10 5 P8 10 0 0 Left Cue 11 Hz -5 Left Cue 11 Hz -10 Right Cue 11 Hz Right Cue 11 Hz Left Cue 39 Hz -10 Right Cue 39 Hz Left Cue 39 Hz -20 Right Cue 39 Hz -15 -30 -20 -40 -25 P7 -30 P8 -50 0 250 500 750 1000ms 0 250 500 Doesburg, Roggeveen, Kitajo & Ward, 2007, Cerebral Cortex 750 1000ms Long-distance gamma synchrony establishes focused attention network QuickTime™ and a decompressor are needed to see this picture. •At 250 ms (same as local power max gamma/min alpha) increase in global phase locking at diverse frequencies •Lateralized in gamma band: P7 (left) for right target, P8 (right) for left target (orienting?) •Increased synch in beta band also but not lateralized (readying?) •Desynch at 100 ms in gamma: erasing old network? MEG replication Increased lateralized synchronization in high alpha band from ~400 ms post cue onset until end of epoch Decreased synchronization side ipsilateral to cue Synchronization in alpha band associated with maintenance of attentional focus at cued location Left cue 14 Hz Doesburg, Ward, 2007, Proceedings BIOMAG2006 Right cue What is SAM beamformer? Synthetic Aperture Magnetometry (Vrba & Robinson 2001, Methods) Based on time/space covariance matrix of sensors Achieves estimate of source power for each voxel in brain region from weighted linear combination of all measurements (where weights are selected to attenuate signals from all other voxels): t ˆ S (t ) W m(t ) Optimal coefficients found by minimizing total power over time (computational tricks used in practice). MEG Replication MEG filtered at 14 Hz SAM beamformer sources in parietal (SPL?) and visual cortices (n=5) PLV analysis applied to broadband source activity filtered from 6 Hz to 60 Hz 14 Hz PLV shows lateralized increased synchrony similar to sensor analysis from 4001000 ms post cue onset (right; n=2) 40 Hz (gamma-band) PLV shows burst of lateralized increased synchrony at ~200-250 ms post cue onset Theta rate gamma synch bursts at least for right parietal-occipital when attending left Doesburg,Herdman, Ward, 2007, Cognitive Neuroscience Society MEG replication Beamformer sources projected to cortical surface (star=source in a sulcus so projection done by hand) Lateralized increases and decreases in synchrony between parietal and occipital sources in alpha band from 400-800 ms post cue onset (here 800 ms and 14 Hz) Occipital 14 Hz power replicates EEG data (blue bars, 800 ms) Doesburg, Herdman, Ward, 2007, Cognitive Neuroscience Society MEG Replication Endogenous orienting: dorsal fronto- parietal (FEF, IPS, V1/V2) We also found FEF sources but not consistent enough for PLV analysis Not enough to activate relevant areas - must be synchronized to be functionally effective? Corbetta and Shulman (2002) Nat Rev Neurosci; Wright & Ward, 2008, Orienting of Attention EEG Replication: BESA beamformer sources from theta band signals Green & McDonald, 2008, PLoS Biology EEG analyses BESA beamformer sources in theta band identified Broadband activity of dipoles at peak voxels computed based on EEG recordings Broadband signals filtered and analytic signal, PLV etc., computed between sources Lateralized increased synchronization in the alpha band Right brain Left-neutral Right-neutral Left brain 350 ms post cue onset until target onset: maintains attention at cued location Doesburg, Green, Ward & McDonald, in prep. Baton-passing in the cerebral cortex Auditory cues and targets Cue types: up or down glide for orient left or right (or vice versa), both for do not orient; each on 1/3 of trials White noise targets (respond to all targets) for gap discrimination presented left (1/3 of trials) or right (1/3 of trials) at random regardless of cue type; probes (respond only if at cued location) presented on 1/3 of trials at random BESA beamformer source RT analyses for theta-band signal Green, Doesburg, Ward & McDonald, submitted % Corr Valid Neutral Invalid 694 ms (25.5) 718 ms (26.6) 716 ms (26.7) 90.9 (.016) 90.7 (.014) 90.6 (.018) Theta-band BESA beamformer source activations Baton-passing: •1 Cue activates STG which in turn activates IPL and SPL •2 IPL/SPL activate IFG •3. IFG interprets cue •4. IFG tells IPL/SPL where to orient •5. IPL/SPL activate relevant STG and maintain activation until target Green, Doesburg, Ward & McDonald, submitted Interim Summary Brain configured to attend to a specific location by Confluence of burst of decreased local alpha power, increased local gamma power and increased long-range gamma synchrony at 250 ms post cue Attention maintained at specific location by increased long-range alpha-band synchrony (at least parietal-visual) that coincides with decreased local alpha power Information/control signals passed between brain regions by establishing and breaking synchronization in gamma band between those regions Still not understood: mechanism of synchrony used (thalamic control?); nature of signals exchanged (control/compliance signals?); how reactivity of sensory cortex is increased by control signal; …… Binocular rivalry: window to awareness Stimuli Apparent locus of fused object Constant stimulation, involuntarily alternating experience Prisms Eyes Corresponding retinal areas Neural synchrony and consciousness Cosmelli et al, (2004) Waves of consciousness: Ongoing cortical patterns during binocular rivalry. NeuroImage, 23, 128-140 Widespread 5 Hz synchrony associated with perception of the 5 Hz stimulus Face Rings Face Rings Face Rings Face Rings Face Synchrony of brain areas at 5 Hz for different subjects; note individual differences in both locations of active brain areas and in amount of synchrony between them. Binocular rivalry: window to awareness Constant stimulation, Stimuli involuntarily alternating experience; subjects press one of two buttons to indicate which pattern they see, neither to indicate parts of both Apparent locus of fused object Prisms Eyes Corresponding retinal areas Long-range theta-rate bursts of gamma-band synchrony in binocular rivalry of striped patterns Frequency (Hz) Button press at 0 Time (ms) 45Hz -450 7Hz -450 -400 -400 -350 -300 -250 -200 -150 -100 -50 0 50 100ms -350 -300 -250 -200 -150 -100 -50 0 50 100ms Doesburg, Kitajo & Ward, 2005, NeuroReport Frequency (Hz) BESA beamformer (280-220 ms prebutton-press, 36-46 Hz) applied to replication R L Time (ms – 0 indicates button press) Average PLVz over 8 active sources: R/L V1/V2, R sup occipital, R parietal, L temporal pole, L DLPF, R/L prefrontal Bursts of gamma-band synchronization occur at theta rate (arrows) around button press in presence of persisting synchronization in theta band; also bursts of synchronization in beta band Theta-modulated synchronization Left V1/V2 – Left DLPFC Right brain Left V1/V2 – Right parietal Left brain 40-50 Hz alt synch/desynch 40-50 Hz synch 30-40 Hz synch 20-30 Hz synch Right parietal – Right occipital 40-50 Hz desynch 30-40 Hz desynch surrogate level 0.025/0.025 Dynamic core hypothesis Neural correlate of conscious awareness is what Tononi & Edelman called the "dynamic core" Large-scale (brain-wide, 200-msec time scale) Coherent (statistically synchronous) activity Millions of neurons involved Dynamic core hypothesis DC simultaneously integrates activities of many brain areas (not all of them, a constantly changing subset) … And also differentiates current conscious state from many other, possible conscious states. My (radical) proposal: the thalamic dynamic core is the neural correlate of phenomenal awareness Cortex computes, thalamus experiences Human cortex, with more neurons and more corticocortical fibers per thalamic fiber computes much more complex contents than do, e.g., rat, dog, or chimp cortices; pace Paul Nunez Cortical DC arises from synchronous activity in thalamus Thalamic dynamic core Dynamic core and supporting binocular rivalry data (Tononi & Edelman, Varela group); neural synchrony is key Lesions, neurosurgery, and anesthetic action point to thalamic “relay” nuclei as critical (Penfield, Alkire et al) Anatomy and function of thalamus and cortex (Mumford) We experience results (products) of computations, not the computations (processes) themselves (Lashley, Kinsbourne, Prinz, Rees, Koch, Baars) Lesions: Karen Ann Quinlan Karen Ann Quinlan’s Brain at Autopsy (see Kinney et al 1994) Drug/alcohol reaction; permanent vegetative state for 14 years Thalamus-massive loss Cortex-little loss Penfield’s neurosurgery and stimulation mapping M.M.’s cerebral cortex mapped via electrical stimulation by Penfield Patient M.M. treated for intractable epilepsy Neurosurgery and stimulation mapping Penfield (The Mystery of the Mind, 1975): The mechanisms of epilepsy and electrical stimulation mapping imply that “…there are two brain mechanisms that have strategically placed gray matter in the diencephalon …, viz. (a) the mind’s mechanism (or highest brain mechanism); and (b) the computer (or automatic sensory-motor mechanism).” (p. 40). Penfield’s “mind mechanism” Merker (2006, BBS): argued SC is locus of conscious analog simulation of world General anesthesia Alkire et al (2000) Consciousness & Cognition: Common brain loci and mode of action of different general anesthetics imply that the critical mechanism of general anesthesia is a hyperpolarization block of the thalamic relay nuclei neurons Common brain areas where halothane and isoflurane anesthesia significantly depresses activity; a. thalamus, b. midbrain reticular formation Alkire et al (2000) Consciousness & Cognition The thalamus Synchronizes cortical oscillations “Gateway” to cortex for major sensory systems (except smell) Evolved along with the cerebral cortex; a “seventh layer” of cortex (but with different neuron type) Each cortical area has an associated subnucleus of thalamus with massive cortico-thalamic projections and smaller thalamo-cortical projections; most thalamic subnuclei have no other inputs! Where is the thalamus? Thalamus Pineal body Gross Anatomy of some cortico-thalamic circuits Roles of the thalamus Relay station and gateway (attentional engagement) to cortex for sensory systems Synchronizes neural activity in remote cortical areas Active blackboard that echoes back to cortex results of latest computations (Mumford) Site of dynamic core of neural activity that gives rise to phenomenal experience(?): thalamic dynamic core We experience products… Crovitz: maximum rate of consciously following strobe light = 4 to 5 Hz (250 to 200 msec per cycle) Sternberg STM scanning: no awareness of process; 40 Hz (25 ms/item) scan? LTM search & Retrieval: no awareness of memory search codes, only memories Speech: not aware of composing utterences, phonemes, (co-)articulation, etc. Conclusions Synchronous neural oscillations occur at several scales and in particular frequency bands Local and global synchrony coordinate to establish an attentional network that enhances processing of attended stimuli Neural synchrony in the thalamo-cortical circuits, particularly in the thalamus, establishes a dynamic core of brain activity that supports (is?) conscious awareness Implications for GWT/LIDA Dynamic core is the GW GW (broadcasting) established by synchronization between various active processing modules mediated by thalamic dynamic core Transient networks of brain regions (processing coalitions) established by low frequency synchronization (carrier?), with high frequency synchronization establishing transient information exchange between processing areas in necessary temporal sequences (baton passing) Perception/action cycle governed by several temporal interactions based on processing speeds and communication requirements vis a vis available oscillation frequencies.