THE FISHES 鱼类
• Fishes are the undisputed(无异议) masters of the
aquatic(水生) environment. Their numerous
structural adaptations have produced a great variety
of forms ranging from gracefully streamlined trout
(鲑鱼)to grotesque(奇异的) creatures that well in
the eternal blackness of the ocean's abyssal depths.
Considered either in numbers of species (more than
22 000 named species) or in numbers of individuals
(countless billions), they easily out number the four
terrestrial vertebrate classes combined.
• Characteristics
• 1. Skeleton with cartilage or bony, which represents the primitive
skeleton; vertebrae numerous; notocord may persist in part
• 2. Skin with mucous g1ands and with embedded dermal scales or
without scales
• 3. Fins both median and paired, with fin rays of cartilage or bone
• 4. Mouth terminal with many teeth (some tooth-less) jaws present;
olfactory (嗅觉) sacspaired and may or may not open into mouth
• 5. Respiration by gills
• 6. Swim bladder(鳔) often present with or without duct connected
to pharynx
• 7. Circulation consisting of a two-chambered heart, arterial and
venous systems, and four pairs of aortic arches; blood of nucleated red
cells
• 8. Nervous system of a brain with small olfactory lobes(嗅叶)
and cerebrum(大脑) and large optic lobes(视叶) and
cerebellum(小脑): ten pairs of cranial nerves
• 9. Sexes separate; gonads paired; fertilization usually external or
internal; larval forms may differ greatly from adults
• 在志留纪由原始有头类发展出一支具有上下颌的
脊椎动物,即鱼类的祖先,进而产生出现生的适
应水生的形形色色的鱼类。
• 主要特征
• 鱼类是最低等的有颌、变温脊椎动物,并适应水
生生活而发展出许多特有的结构。它具有比圆口
类更为进步的机能结构,主要表现在:
• 出现了能咬合的上下颌。
• 出现了成对的附肢(偶鳍)。
• 骨骼为软骨或硬骨。
• 脊柱代替了脊索,成为身体的主要支持结构。
• 头骨更加完整,脑和感觉器官更发达。
• 由于局限于水生环境,鱼类在进化上也有局限性,
适应水栖生活的特征主要表现为:
• 身体分为头、躯干和尾,缺少颈部,因此头不能
灵活转动。
• 体形多为流线型,体被骨质鳞片或盾鳞;体表富
黏液;具侧线。
• 以鳃为呼吸器官。
• 血液循环为单循环。
• 以鳔或脂肪调节身体比重获得水的浮力;靠躯干
分节的肌节的波浪式收缩传递和尾部的摆动获得
向前的推进力;有良好的调节体内渗透压的机制。
STRUCTURAL AND FUNCTIONAL
ADAPTATIONS OF FISH
• Locomotion in water
•
To the human eye, fish appear capable of swimming at
extremely high speeds. But our judgment(判断) is
unconsciously tempered by our own experience that water is
a highly resistant medium to move through. Most fishes,
such as a trout(鲑鱼) or a minnow(鲤科小鱼), can
swim maximally about l0 body lengths per second,
obviously an impressive performance by human standards.
Yet when these speeds are translated into miles per hour it
means that a foot long trout can swim only about 6.5 miles
per hour. The larger the fish, the faster it can swim. A 2-foot
long salmon can sprint l4 miles per hour and a 4-foot
barracuda(梭鱼), the fastest fish measured, is capable of
27 miles per hour.
• The propulsive mechanism of a
fish is its trunk and tail
musculature. When swimming,
the shark body assumes the form
of a sine wave. Waves of
contraction begin on one side of
the body at the front and proceed
to the tail. When this wave has
moved some distance, an other
wave is initiated at the front on
the opposite side of the body. The
process continues, with waves of
contraction moving posteriorly,
alternating from one side to the
other. In higher bony fishes, the
sweeping movement of the tail
assumes a greater role.
• Swimming is possible only because the density and
noncompressibility of water offers great purchase for forward thrust.
As a medium for locomotion, water offers another advantage: since
the density of water is only slightly less than that of protoplasm,
aquatic animals are almost perfectly supported and need expend no
energy overcoming the forcen of gravity. Consequently, swimming is
actually the most economic form of animal locomotion. However, the
low energy cost of fish swimming is by no means fully understood.
Relatively simple calculations show that a fish moves through water
with only about one tenth the drag of a rigid model of the fish's body.
The energy required to propel a submarine is many times greater than
that consumed by a whale(鲸) of similar size and moving at the same
speed. Aquatic mammals and fishes create virtually no turbulence, a
feat that man in his twentieth-century ingenuity is a long way from
matching. The secret lies in the way aquatic animals bend their bodies
and fins (or flukes) to swim, and in the textural properties of the body
surface. It has recently been shown, for example, that the slimy
surface of a fish reduces water friction by at least 66%.
• 鱼类的运动
• 鱼类的运动方式主要为波浪式运动,或称游泳。借助于
连续的肌节收缩与舒张,从头部开始的收缩在身体两侧
交替进行,形成波浪式的传递,使收缩波传向尾部,身
体则向收缩的一侧弯曲使成S型。收缩在尾部结束,尾
部将收缩的力传给水,这个力被水以同等大小、但方向
相反的反作用力作用于尾部。这个力向前的分力是鱼体
向前运动的主要推进力。金枪鱼(Thunnus thynnus)的游
泳速度可达75km/h。
• 跳跃或飞翔是鱼类另一种很突出的运动方式。如水温的
突变,受到某种刺激,追捕食物,越过障碍等,常能引
起跳跃运动。此外,鱼类还可靠鳍拨水、以鳃孔喷水作
为辅助运动方式。表皮内具有大量单细胞粘液腺,分泌
粘液使体表粘滑。这些特点均为减少水中游泳的阻力。
爬行也是鱼类运动方式之一,不过比较罕见。
鱼类结构和功能的适应
• 1.体形
• 绝大多数鱼类为流线型或纺锤型,以减少水中运动的
阻力,可快速而持久地游泳。由于水环境和生活方式
的不同,大致可分为四种体形:纺锤形、侧扁形、平
扁形、鳗形。此外,还有一些特殊体形如箱鲀、比目
鱼、海马、海龙等。
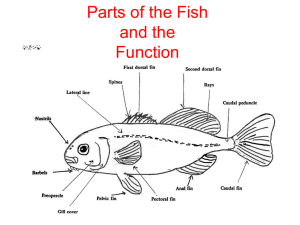
• 2.外形分部
• 身体分为头、躯干、尾3部分。
• 头:吻端至鳃盖后缘(硬骨鱼,不包括鳃盖膜)或最
后一对鳃裂(软骨鱼)。
• 躯干:最后1对鳃裂或鳃盖骨后方为躯干的开始,肛门
或泄腔孔为躯干和尾的分界。
• 尾:肛门到尾鳍基部长度或尾柄与尾鳍交界处。也可
说到最后一块脊椎骨。
• 3.水中运动器官—鳍
• 3.1分类
• 3.1.1按结构:可分为骨质鳍条也叫鳞质鳍条
(1epidotrichia)和角质鳍条两类。
• 软骨鱼的鳍,外面都覆盖有皮肤,内面由角质鳍条支持。
角质鳍条为一种纤维状的角质物,细长而不分节。
• 硬骨鱼的鳍由骨质鳍条支持,鳍条间以薄的鳍膜相联。
骨质鳍条也叫鳞质鳍条(1epidotrichia),系由鳞片衍生而
成。骨质鳍条分鳍棘和软条两种类型。
• 鳍棘:是一种由鳍条变形来的既不分支也不分节的硬棘,
为高等鱼类所具有。
• 软条:柔软而有节,其远端分支(叫分支鳍条)或不分支
(叫不分支鳍条),均由左右两半合并而成。不分支鳍条
常是最前面的几根。
• 在鲤形目中有些种类,例如鲤鱼,其背鳍与臀鳍前方的
硬棘,仍保留有分节和左右两列合并的痕迹,称为假棘,
系软条钙化后的变形物,不是真正的棘,因为真正的棘
始终为单一的结构而无法分开。
• 3.1.2按着生位置及功能
• 奇鳍(unparied fin),为不成对的鳍:包括背鳍(dorsal fin)、
臀鳍(anal fin)、尾鳍(caudal fin)。
• 偶鳍(paired fin),为成对的鳍: 包括胸鳍(pectoral fin)、
腹鳍(pelvic fin)。
• 硬骨鱼的偶鳍呈垂直位,而软骨鱼的偶鳍呈水平位。
• 3.2偶鳍的位置和作用
• 3.2.1胸鳍:
• 相当于陆生脊椎动物的前肢,着生于鳃盖后缘的胸部。
除有高低变化外,位置一般比较恒定。它的基本功能为
运动、平衡和掌握运动方向。当鱼类停止前进时,胸鳍
用于控制鱼体的平衡;缓慢地游动时,主要由胸鳍起着
船桨的拨水作用;高速行进时,胸鳍紧贴鱼体,胸鳍举
起则可减速和制动;当胸鳍一侧紧贴鱼体,一侧举起,
则鱼体朝举起的一侧拐弯,可见胸鳍的作用是多方面的。
• 3.2.2腹鳍:
• 相当于陆生脊椎动物的后肢,其作用不及胸鳍大,主要
协助背鳍、臀鳍维持鱼体的平衡,并有辅助鱼体升降和
拐弯功能。腹鳍着生的位置,各种鱼类并不相同,一般
软骨鱼类的腹鳍位于泄殖孔的两侧,形状与胸鳍相似而
稍小。硬骨鱼的腹鳍位置变化很大,有以下三种:
• 腹鳍腹位:位于躯干腹侧的腹部的,如鲤、鲇、鲱、鲑
等一些较原始的种类。
• 腹鳍胸位:位于胸鳍前方鳃盖之后的胸部的,如鲈、鲷、
鲽和黄鱼等。
• 腹鳍喉位:位于两鳃盖之间的喉部,如鲥科,腹鳍胸位
和喉位都是鱼类的高级特征,因此这种位置上的不同,
在鱼类分类和演化上都有很重要的意义。
• 3.3鳍式 鳍的组成和鳍条数目的记载叫鳍式,在鱼类分
类学上使用最为普遍。
• 一般记载方式是取用各鳍的拉丁名第一字母代表鳍的类
别,如“D”代表背鳍,“A”代表臀鳍,“V”代表腹鳍,
“P”代表胸鳍,“C”代表尾鳍。大写的罗马数字代表棘
的数目,阿拉伯数字代表软条的数目,棘或软条的数目
范围以“一”表示,棘与软条相连时用“一”表示,分
离时以“,”隔开。例如:
• 鲤鱼:D.I I I—IV—17—22 ;A.III—5—6;P.I—15
一16;V. I I—8—9
•
鲈鱼:D.X I I ,I—13;A.III—7—8;P.15一18;
V.I一5
• 以上两个鳍式表明:鲤鱼一个背鳍,3至4根硬棘和17至
22根软条;臀鳍3根硬棘和5至6根软条,胸鳍1根硬棘和
15至16根软条,腹鳍2根硬棘和8至9根软条。鲈鱼则有
两个背鳍,第一背鳍由12根硬棘组成,无软条,第二背
鳍包含1根硬棘和13根软条。
• 3.4尾鳍
• 3.4.1作用
• 有平衡、推进和转向的作用,尾的扭曲和伸直使
鱼体产生前进运动。
• 3.4.2分类
• 根据鱼类的尾鳍外形和尾椎骨末端的位置,一般
将尾部分为三种类型:
• 原形尾(protocercal tail):尾椎的末端平直伸展至
尾的末端,尾鳍的上、下叶大致相等,这是一种
原始的尾型。见于圆口纲,鱼纲仅见于胚胎期和
仔鱼期。
• 歪形尾(heterocercal tail):尾椎的末端曲而伸展
入较发达的尾鳍上叶内,下叶小而略为突出,这
种尾型内外均不对称。见于古代的甲胄类、盾皮
鱼类,现代的软骨鱼类和少数硬骨鱼类,如鲨、
鲟等种类。
• 正形尾(homocercal tail):在骨骼构造上仍
保留有歪形尾的痕迹,尾椎的末端仍翘向
尾鳍上叶,但仅达尾鳍上叶基部,其较发
达的下叶由增宽的尾下骨片(hypural bone)
支持着,因而尾鳍在外形上完全对称。
• 正形尾为高等鱼类的尾形。根据鳍形的变
化,正形尾还可归纳成多种鳍形(新月型、
叉型、内凹型、平截型、圆形、双凹型
等)。一般快速游泳的鱼类,尾鳍呈新月
形或叉形,尾柄也较细,如鲐、鲅等;游
泳速度慢的鱼类,尾鳍后端呈方形或圆形,
尾柄相应粗大,如纯类、虾虎鱼类。
• 软骨鱼为歪尾型,而硬骨鱼为正尾型。
Neutral buoyancy and the swim bladder
•
•
All fishes are slightly heavier than water because their
skeletons and other tissues contain heavy elements that are
present only in trace amounts in natural waters. To keep
from sinking, sharks(鲨鱼) must always keep moving
forward in the water. The asymmetric (heterocercal) tail (歪
尾)of a shark provides the necessary tail lift as it sweeps to
and fro in the water and the broad head and flat pectoral fins
act as planes to provide head lift. Sharks are also aided in
their buoyancy problem by having very large livers
containing a special fatty hydrocarbon called squalene(鲨烯)
that has a density of only 0.86. The liver thus acts like a
large sack of buoyant oil that helps to compensate for the
shark's heavy body.
• By far the most efficient flotation device is a gas
filled space. The swim bladder(鳔) (or gas bladder
as it is often called) serves this purpose in the bony
fishes. It arose from the paired lungs of the primitive
Devonian(泥盆纪) bony fishes. Lungs were
probably a ubiquitous feature of the Devonian
freshwater bony fishes when, as we have seen, the
alternating wet and dry climate probably made such
an accessory respiratory structure essential for life.
Swim bladders are present in all pelagic bony fishes
but absent in most bottom dwellers such as
flounders(比目鱼) and sculpins(杜父鱼).
•
By adjusting the volume of gas in the swim bladder, a fish can
achieve neutral buoyancy and remain suspended indefinitely at any
depth with no muscular effort. There are severe technical problems,
however. If the fish descends to a greater depth, the swim bladder gas
is compressed so that the fish becomes heavier and tends to sink. Gas
must be added to the bladder to establish a new equilibrium buoyancy.
If the fish swims up, the gas in the bladder expands, making the fish
lighter. Unless gas is removed, the fish will rise with ever increasing
speed while the bladder continues to expand, until it pops helplessly
out of the water. There are two ways fish adjust gas volume in the
swim bladder. The less specialized fishes (trout, for example) have a
pneumatic duct that connects the swim bladder to the esophagus ;
these forms must come to the surface and gulpair to charge the
bladder and obviously are restricted to relatively shallow depths. More
specialized teleost shave lost the pneumatic duct. Gas exchange
depends on two highly specialized areas: a gas gland that secretes gas
into the bladder and a resorptive area, or "oval," that can remove gas
from the bladder. The gas gland contains a remarkable network of
blood vessels (rete mirabile 迷网) arranged so that a vast number of
arteries and veins in a tight bundle run in opposite directions to each
other. This is called countercurrent flow, and in some way, not yet
understood, this arrangement makes possible a tremendous
multiplication of gas concentration inside the swim bladder.
• The amazing effectiveness of this device is exemplified by a
fish living at a depth of 8,000 feet. To keep the bladder
inflated, the gas inside (mostly oxygen, but also variable
amounts of nitrogen, carbon dioxide, carbon monoxide, and
argon) must have a pressure exceeding 240 atmospheres.
Yet the oxygen pressure in the fish's blood cannot exceed
one-fifth atmosphere--equal to the oxygen pressure at the
sea surface. Physiologists suggest that oxygen and other
gases are actively transported within the gas gland's counter
current exchange system; this would involve a molecular
carrier system not yet identified.
• Other theories have been proposed to explain swim bladder
function, and this interesting organ is the object of
considerable current research.
• 鳔和浮力
• 由于水的密度大,受到水的浮力作用的鱼体如
果没有重力的影响,维持在一定深度只需最小限
度的能量。但所有的鱼类由于有密实的骨骼和肌
肉以及其他组织,比重稍重于水,为减少密度使
之不下沉,产生了一些结构以减少自身的比重。
一个有效的方法是增加脂肪在身体中的比例,如
软骨鱼类,具有巨大的肝脏,占体重的20%-25
%,其中含有大量的密度小于水(0.95g/mL)的
鲨烯。
• 另一个方法是以鳔的方式在体内增加气体,如大
部分硬骨鱼类具有鳔。淡水鱼类鳔的容量达鱼体
积的7%—10%,海鱼鳔容量达4%-6%。调节鳔
内气体的方法有2种,一是鳔有鳔管通入食管背
面,动物以吞咽或吐出空气来调节,这类动物称
开鳔类。另一种鳔不具鳔管,为闭鳔类,它们依
靠鳔的红腺和卵圆区来调节气体容量。
• 红腺位于鳔的前腹壁,卵圆区位于鳔的后背壁。
红腺处集中分布了大量毛细血管,形成奇异的网
(rete mirabile)。红腺分泌乳酸进入血液,可把血
液中的氧气分离出来进入鳔内,此时卵圆区的括
约肌收缩。当需要下沉而减少鳔内气体时,卵圆
区括约肌放松,空气进入卵圆区,渗入这里的血
管中。
皮肤及衍生物
• 5.1 皮肤
• 表皮和真皮均为多层细胞,且皮肤与肌肉紧密相
接,皮下组织极少,使整个身体成为坚实的实体。
表皮内具有大量单细胞粘液腺,分泌粘液使体表
粘滑。这些特点均为减少水中游泳的阻力。粘液
还可保护身体免遭病菌、寄生物的侵袭。
• 5. 2皮肤衍生物
• 5.2.1真皮鳞(dermal scale)
• 是鱼类特有的皮肤衍生物,是一种保护性的结构,
根据形状的不同,分为三种:
• 1 楯鳞(placoid scale) 这是软骨鱼类所特有的鳞片,构造比较原始,
分布全身,斜向排列,使身体表面显得很粗糙。楯鳞是由真皮
(derinis)和表皮(epidermis)联合形成。真皮演化为基板和板上的齿
质部分(dentine),齿质部分的尖峰指向后方。楯鳞执行着牙齿的
功能。楯鳞和牙齿应该是同源器官。
• 2 硬鳞(ganoid scale) 这是硬骨鱼中最原始的鳞片,单列排列,见于
鲟鱼和雀鳝等,由真皮演化 而成,典型鳞片呈斜方形,含有硬
鳞质(ganoin),发特殊亮光。
• 3骨鳞(bony scale) 这是绝大多数硬骨鱼类所具的鳞片,也由真皮
演化而成,略呈圆形,前端插入鳞囊内,后端游离,彼此作覆瓦
状排列,有利于增加躯体的灵活性。
• 游离的一端光滑的,称为圆鳞(cycloid scale),多见于鲤科鱼类
• 游离的一端生有许多细小锯状突起的,称为栉鳞(ctenoid scale),
在鲈科鱼类中常可见到。
• 骨鳞由许多同心圆的环片(sclerite)组成。环片因季节不同而表现
出生长速度的差异。鳞片上出现了年轮。根据年轮的情况,可以
推算鱼类的年龄、生长速度以及生殖季节等等,在养殖业和捕捞
业上都有极大的实践意义 。
• 侧线及侧线鳞
• 侧线:是低等水生脊椎动物(包括水生两栖类)特有的感
觉器官,由感受机械刺激的神经丘器官和感受电刺激的
壶腹器官组成。前者感觉水流的方向、速度和压力,后
者感觉水中动物因肌肉收缩而形成的微弱电场。侧线对
于鱼类的取食、避敌和求偶具有重要的生物学意义。
• 侧线鳞:有侧线器官穿孔的鳞片,叫做侧线鳞。
• 侧线鳞的数目以及侧线上鳞(由背鳍起点的基部至侧线
这一段距离上的鳞片)和侧线下鳞(由臀鳍起点的基部至
侧线这一段距离上的鳞片)的数目,通常都用鳞式来表
示,是鱼类分类方面需要记载的数据之一。
• 鳞式的写法是:侧线鳞数目(侧线上鳞数目/侧线下鳞
数目)
• 例如鲤鱼的鳞式为34~38(5/8)。这就是说:鲤鱼的
侧线鳞为34~38片,侧线上鳞为5片,侧线下鳞为8片。
• Coloration and concealment(隐蔽)
•
Tropical coral reef fishes bear some of the most
resplendent hues(色彩) and strikingly brilliant color
patterns in the animal kingdom. Viewed against the coral
reef background of invertebrate and plant life that create a
riot of color, the vivid(鲜明的) markings and coloration of
reef fishes attract relatively little attention. Their coloration
is not always concealing, however, since tropical bony
fishes tend toward vivid coloration even in areas of dull and
somber backgrounds. Although conspicuous in these
circumstances, they are protected by alertness and a gillty,
or by their poisonous flesh. In this instance the coloration is
an advertisement, warning would-be predators that they
should seek their meal elsewhere.
• Outside of coral reefs and other littoral habitats of the
tropical seas, fishes, like most other animals,
characteristically bear colors and patterns that serve to
conceal them from enemies. Freshwater fish as wear
subdued shades of green, brown, or blue above, grading to
silver and yellow-white below. This is obliterative shading.
Seen from above against its normal background of water
and stream bottom, the fish becomes almost invisible. Seen
from beneath as it might be viewed by an aquatic predator,
the palebelly of the fish blends with the water surface and
sky above. The obliterative coloration is frequently
enhanced by blotches, spots, and bars—conflicting patterns
that like the camouflaged ships of the Second World War
tend to break up the outline of the body. Some fishes, such
as many flatfish(比目鱼) species, can change their color
to harmonize with the patterns of their background.
• Fish colors are chiefly the result of pigment within
chromatophores(色素体) in the dermal layer of the skin. The
pigments are red, orange, yellow, and black and can be blended to
produce other shades. Pigment dispersion within the branched
chromatophores is controlled by the autonomic nervous system.
Fish also bear guanine in their skin, a purine (嘌呤)compound
that gives many fish their silvery appearance.
•
Another form of concealment is protective resemblance, or
mimicry. Pipe fishes, for example, are of the shape and color of
the seaweeds(海藻) among which they live . Pipe fishes even
sway slowly like seaweeds in a gentle current. There are many
other examples of protective body form that appear to turn their
owners into fragmented seaweed, or a leaf, or floating debris, or a
weed-covered rock on the bottom. The goosefish, or angler, with
its obliterative coloration and numerous fringes and branched
appendages of skin, not only becomes nearly invisible when
bedded down on the ocean floor, but sways a tempting bait above
its huge mouth to attract prey.
• 色素细胞(chromatophore):来源于真皮,是许多
鱼类的体色得以丰富多采的物质基础。
• 毒腺:有些鱼类的皮肤中有由表皮转化而来的,
是它们攻击和防卫的武器,例见于玫瑰毒蚰、灰
刺虹等。
• 发光器官:有些鱼类、特别是生活在海洋深处的
鱼类,还具有另一种皮肤衍生物——发光器官。
鱼类发光可能是招引趋光的食物便于摄食,或作
同种间和雌雄间的联系信号。
• Migrations
• For centuries naturalists had been puzzled(迷) about the life
history of the freshwater eel(鳗鲡), Anguilla, a common and
commercially important species of coastal streams of the
North Atlantic(大西洋). Each fall(秋天), large numbers of
eels were seen swimming down the rivers toward the sea,
but no adults ever returned. And each spring countless
numbers of young eels, called “elvers,” each about the size
of a wooden match stick, appeared in the coastal rivers and
began swimming upstream(上游). Beyond the assumption
that eels must spawn somewhere at sea, the location of their
breeding grounds was totally unknown. When the mystery
was finally solved by the Danish scientist Johann Schmidt,
the story was so fantastic(幻想) that even today biologists
occasionally pause to wonder if it can be true.
• The first clue was provided by two Italian scientists, Grassi
and Calandruccio, who in l896 reported that elyers were not
in fact larval eels; rather they were relatively advanced
juveniles. The true larval eels, the Italians discovered, were
tiny, leaf-shaped, completely transparent creatures that bore
absolutely no resemblance to an eel. They had been called
leptocephali by early naturalists who never suspected their
true identity(丹麦). In l905 Johann Schmidt, supported by
the Danish government, began a systematic study of eel
biology that he continued until his death in l933. Enlisting
the cooperation of captains of commercial vessels plying the
Atlantic, thousands of the leptocephali were caught in
different areas of the Atlantic with the plankton(浮游生物)
nets Schmidt supplied them. By noting where in the ocean
larvae in different stages of development were captured,
Schmidt and his colleagues eventually reconstructed the
spawning migrations.
• When the adult eels leave the coastal rivers of Europe and
North America, they swim steadily and apparently at great
depth for l to 2 months until they reach the Sargasso Sea, a
vast area of warm oceanic water southeast of Bermuda.
Here, at depths of l,000 feet or more, the eels spawn and die.
The minute larvae then begin an incredible journey back to
the coastal rivers of Europe. Drifting with the Gulf Stream
and preyed upon constantly by numerous predators, they
reach the middle of the Atlantic after 2 years.
• By the end of the third year they reach the coastal waters of
Europe where the leptocephali metamorphose into elvers,
with an unmistakable eel-like body form. Here the males
and females part company; the males remain in the brackish
waters of coastal rivers and estuaries while the females
continue up the river soften penetrating hundreds of miles
up stream. After 8 to l5 years of growth, the females, now 3
to 4 feet long, return to the sea to join the smaller males;
both return to the ancestral breeding grounds thousands of
miles away to complete the life cycle.
• Schmidt found that the American eel (Anguilla rostrata)
could be distinguished from the European eel (A. vulgaris)
because it had fewer vertebrae—an average of l07 in the
American eel as compared to an average ll4 in the European
species. Since the American eel is much closer to the North
American coast line, it requires only about 8 months to
make the journey. Recently it has been suggested that both
American and European eels belong to the same species and
that the adult European forms all die before reaching the
spawning grounds. All eggs are laid by American eels
(according to the theory), but when hatched, the larvae in
the northern part of the spawning grounds will be carried by
currents to Europe; those in the southern part will be carried
toward the American coast.
• The larger number of vertebrae in the European eel
could be explained on the basis of Jordan's law,
which states that fish in colder waters have more
vertebrae than those in warmer temperatures.
• This theory remains to be confirmed. There are also
several species of eels in the western Pacific and
Indo-Pacific regions. They too undergo seaward
spawning migrations, but nothing is known of the
oceanic portion of their life histories. Many
biologists suspect that once these species are studied
with an investigation of the scale carried out by
Schmidt, and by someone having his patience, there
may well unfold an epic rivaling that of the common
Atlantic eel.
• 洄游:
• 一些鱼类在其生命过程中的一定时期会沿一定路线进行
集群的迁移活动,以寻求对某种生理活动的特殊要求,
并避开不利的环境。这种迁移活动称为洄游。洄的原因
和洄游路线的决定因素较为复杂,与遗传和环境因素关
系密切。洄游分为3类。
• 生殖洄游 从越冬或索饵场所向产卵地进行的迁移。如
成熟的海生大麻哈鱼(Oncorhynchus keta)在生殖期进行
溯河洄游,到达淡水的产卵地。而淡水的鳗鲡(Anguilla
japonica)则降河洄游,从淡水游向深海进行产卵,幼鱼
为透明柳叶状,经变态后再溯河而上,进入淡水生长。
鱼类识别洄游路线有自身的遗传因素,并靠识别沿途的
水流气味、温度梯度、食物构成、地球磁场等综合因素
来进行洄游。
• 索饵洄游 为寻找食物而进行的洄游。
•
越冬洄游 为寻找适宜的越冬场所而进行的洄游。
Osmotic regulation(渗透压调整)
•
Freshwater is an extremely dilute medium with a salt concentration
(0.00l to 0.005 gram moles per liter[M] much below that of the blood
of fresh water fishes (0.2 to 0.3 M). Water therefore tends to enter
their bodies osmotically and salt is lost by diffusion outward.
Although the scaled and mucus-covered body surface is almost totally
impermeable to water, water gain and salt loss do occur across the thin
membranes of the gills. Freshwater fish are hyperosmotic(高渗透压)
regulators that have several defenses against these problems . First,
the excess water. is pumped out by the mesonephric kidney , which is
capable of forming a very dilute urine. Second, special salt-absorbing
cells located in the gill epithelium are capable of actively moving
saltions, principally sodium (Na)and chloride(氯化物), from
the water to the blood. This, together with salt present in the fish's
food, replaces diffusive salt loss. These mechanisms are so efficient
that a freshwater fish devotes only a small part of its total energy
expenditure to keeping itself in osmotic balance.
• Marine bony fish are hypoosmotic regulators that encounter a
• completely different set of problems. Having a much lower blood salt
concentration (0.3to 0.4 M) than the seawater around them (about lM),
they tend to lose water and gain salt. The marine teleost fish quite
literally risks drying out, much like a desert mammal deprived of
water. Again, marine bony fishes, like their freshwater counterparts,
have evolved an appropriate set of defenses.To compensate for water
loss, the marine teleost drinks seawater. Although this behavior
obviously brings needed water into the body, it is unfortunately
accompanied by a great deal of unneeded salt. Unwanted salt is
disposed of in two ways: (l) the major sea salt ions (sodium, chloride,
and potassium K+) are carried by the blood to the gills where they are
secreted outward by special salt-secretory cells; (2)the remaining ions,
mostly the divalent ions (magneslum, sulfate(硫酸盐), and calcium
Ca), are left in the intestine and voided with the feces. However, a
small but significant fraction of these residual divalent salts in the
intestine, some l0% to 20% of the total, penetrates the intestinal
mucosa and enters the bloodstream. These ions are excreted by the
kidney. Unlike the freshwater fish kidney, which forms its urine by the
usual filtration-reabsorption sequence typical of most vertebrate
kidneys the marine fish's kidney excretes divalent ions by tubular
secretion. Since very little if any filtrate is formed, the glomeruli have
lost their importance and disappeared altogether in some marine
teleosts. The pipe fishes and the goosefish,are good examples of
排泄系统和渗透压的调节
• 成体鱼类的肾脏位于胸腹腔背面的中后部,以鲤鱼为例,
排泄系统包括肾脏、输尿管和膀胱。肾脏紧贴在胸腹腔
的背面,呈深红色,两个肾有一部分相连。每一肾的前
端为一头肾(hend kidney),头肾是拟淋巴腺,不是肾脏
本体。肾脏在头肾的后面,肾的宽度和厚度,由前到后
很不一致,它的最宽厚处是在与鳔的中部相接触的一段,
再往后变的很细。两肾各有一输尿管,沿胸腹腔背壁向
后走行,到将近末端处合而为一,稍为扩大,形成膀胱。
硬骨鱼的膀胱属于导管膀胱(tubal bladder),为中肾管后
端膨大形成。膀胱以后成为尿道,通至泄殖窦,以泄殖
孔开口于肛门后方。其中的肾小管仍保留少量从体腔中
收集废物的肾口,主要从血液中收集含氮废物排出体外,
由输尿管输尿。
•
• 鱼类的肾脏还对调节体内渗透压起着重要作用。
淡水和海水的含盐浓度相差很大,也与生活在其
间的鱼类的体液浓度不同,淡水鱼和海水鱼类具
有不同的体液调节机制来保证生命的正常运行。
海生硬骨鱼的血液盐浓度(0.3~0.4g/L)低于
周围海水(1g/L),体液大量渗出,机体面临失
水的威胁。为补偿体液的丧失,海生硬骨鱼大量
吞饮海水,而进人体内的过多的盐分通过位于鳃
上皮的泌氯腺(chloride—secreting gland)排出。为
保存水分,肾小体大部退化,肾脏排出的尿量极
少,如海生杜父鱼(Cottus gobio)每小时每公斤体
重排尿0.13-0.96mL。
• 海生软骨鱼类则在血液中积累大量尿素,浓度达
2%-2.5%,使血液渗透压高于周围海水,致使
海水不断渗入体内。进人体内的多余水分经肾脏
排出,多余的盐分经直肠背面的直肠腺(rectal
gland)排出。
• 淡水硬骨鱼的血液浓度(0.2-0.3g/L)高于周围
水环境(0.001-0.005 g/L),水分通过各种途
径进入体内,再以大量的稀尿液通过肾脏排出,
以取得平衡,因而肾小体数目极多。如鲤鱼
(Cyprinus carpio)每小时每公斤体重排尿5mL。同
时鳃上皮具有从水中吸收盐分的细胞,以补偿盐
的失去。
Respiration
• Fish gills are composed of thin filaments covered
with a thin epidermal membrane that is folded
repeatedly into plate-like lamella(薄片). These
are richly supplied with blood vessels. The gills are
located inside the pharyngeal cavity and covered
with a movable flap, the operculum(鳃盖骨).
This arrangement provides excellent protection to
the delicate gill filaments, streamlines the body, and
makes possible a pumping system for moving water
through the mouth, across the gills, and out the
operculum. Instead of opercular flaps as in bony
fishes, the elasmobranchs(软骨鱼) have a series
of gill slits out of which the water flows.
•
• In both elasmobranchs(软骨鱼) and bony fishes the
branchial mechanism is arranged to pump water
continuously and smoothly over the gills, even though to an
observer it appears that fish breathing is pulsatile(跳动).
The flow of water is opposite to the direction of blood flow
(countercurrent flow), the best arrangement for extracting
the greatest possible a mount of oxygen from the water.
Some bony fish can remove as much as 85% of the oxygen
from the water passing over their gills. Very active fish,
such as herring(青鱼) and mackerel(鲭), can obtain
sufficient(充足) water for their high oxygen demands only
by continually swimming forward to force water into the
open mouth and across the gills. Such fish will be
asphyxiated(窒息) if placed in an aquarium (养鱼池) that
restricts free swimming movements, even though the water
is saturated(饱和) with oxygen.
• A surprising number of fishes can live out of water for
varying lengths of time by breathing air. Several devices are
employed by different fishes. We have already described the
lungs of the lungfishes, polypterus. and the extinct
crossopterygians(总鳍鱼). Fresh water eels often make
overland excursions during rainy weather, using the skin as
a major respiratory surface. The bowfin, Amia. has both
gills and a lunglike swim bladder. At low temperatures it
uses only its gills, but as the temperature and the fish‘s
activity increases, it breathes mostly air with its swim
bladder. The electric eel has degenerate gills and must
supplement gill respiration by gulping air through its
vascular mouth cavity. One of the best air breathers of all is
the Indian climbing perch(河鲈) that spends most of its time
on land near the water's edge breathing air through special
air chambers above the much-reduced gills.
呼吸系统
• 鱼类必须从含氧量只及空气含氧量3%的水中获得足够
的氧气,并将其送入血液,其呼吸器官必须是高效的这
个器官即是外胚层形成的鳃。鱼类和圆口类都是用鳃呼
吸的水生脊椎动物,但圆口类的鳃丝(gill filament)起
源于内胚层而鱼类的鳃丝起源于外胚层。
• 软骨鱼类的鳃比较原始:鳃裂直接开口于体外,鳃隔
(gill septum)发达,其前后各有一个半鳃(hemibranch),
这两个半鳃总称全鳃(holobranch)。从图可以看出:软骨
鱼类咽部每侧具四个全鳃、一个半鳃,或者说,具九个
半鳃。
• 硬骨鱼类的鳃裂,在外侧另有鳃盖(operculum)保护,鳃
隔已退化,咽部每侧留下明显的四个全鳃。丝状或片状
的鳃瓣直接或间接地着生在鱼的5个鳃弓上。软骨鱼类
的鳃瓣是由上皮折叠形成栅板状附着 在鳃间隔上,硬
骨鱼类的鳃丝直接着生在鳃弓上。
血液循环
• 鱼类的血液循环与鳃呼吸密切相关,为单循环,即由心室压出的
缺氧血经人鳃动脉入鳃部进行气体交换,出鳃的多氧血经出鳃动
脉不再回心脏而是直接沿背大动脉流到全身,从各组织器官返回
的缺氧血经主静脉系统再流回心脏,形成一个大圈。血液在全身
循环一周只经过心脏一次,为单循环。
• 心脏
• 位于围心腔内,由静脉窦、一心房、一心室、动脉圆锥(或动脉
球)组成。动脉圆锥是心室的延伸,可主动收缩,为软骨鱼类所
具有;动脉球是腹大动脉基部的膨大,无收缩能力,为硬骨鱼类
所具有。
血管系统
• 动脉系统主要由腹大动脉、入鳃和出鳃动脉以及背大动脉组成,
静脉系统与文昌鱼的静脉系统相似(包括前主静脉、后主静脉和
总主静脉),为低等水栖脊椎动物所具有。门静脉包括肝门静脉
(收集消化道来的血液进入肝脏)和肾门静脉(收集尾部血液进入肾
脏)。
• 肺鱼除用鳃呼吸外,必要的时候,还可利用鳔代
行肺呼吸,它的循环系统因此也就发生了一系列
相应的改变。以非洲肺鱼为例,其心房隔为左右
两半,心室亦为一不完全的纵隔分为左右两半,
同时动脉圆锥(conus arteriosus)内又生出一个纵隔,
将圆锥腔分成两个通路。经心房右侧(右心房)来
的缺氧血,穿过心室进入后面两对入鳃动脉;从
肺(鳔)静脉来的多氧血,则经心房左侧(左心房)
和心室进入前面两对入鳃动脉,这样就在心脏内
初步划分了多氧血和缺氧血。最后一对出鳃动脉
再各分出一条到鳔去的肺动脉,把缺氧血运到鳔
去进行气体交换,完成气呼吸的全过程。
消化系统
•
绝大多数鱼类为主动捕食的肉食性动物。消化系统有
以下特点:
•
出现上、下颌。
•
出现真正的牙齿。在上下颌的有颌齿,不发达的种类,
其第5对鳃弓上着生咽喉齿,以不同的形状适应不同食
性,如食草的草鱼咽喉齿呈梳状,肉食性青鱼呈臼状。
• 出现食道与胃、肠的分化。这与出现上下颌、可摄取较
大型食物有关。胃与食道交界处为贲门,与肠交界处为
幽门。肠分为小肠和直肠。软骨鱼类的小肠包括极短的
十二指肠和螺旋瓣肠,肠内的螺旋瓣用于增加肠内消化
吸收的面积。硬骨鱼类的小肠分化不明显,有的种类在
幽门部生有幽门盲囊,以增加肠内吸收面积。
• 肠的长短与食性有关。软骨鱼类肠的末端是
泄殖腔,以泄殖腔孔通体外;硬骨鱼类则是
以肛门和泄殖孔与体外相通。
• 消化腺。软骨鱼类有独立的肝脏和胰脏,
尤以肝脏发达。除消化功能外,还有调节自
身比重的作用。大多数硬骨鱼类的肝脏和胰
脏组织混在一起称肝胰脏。
•
AXIAL SKELETON
• Apart from the limb supports, the somatic skeletal (膜质
骨)e1ements are classed as axial.
•
As an introduction to vertebral structure we may first
consider the relatively simple situation seen in typical
vertebrae of amniotes(脊椎动物) before considering the
wild assortment of variants seen in lower vertebrate classes.
Each vertebra, formed of bone in these forms, includes two
principal portions. a neural arch and a centrum(椎体).
Ventrally placed is the centrum which functionally replaces
the notochord. Typically this is a spool-shaped cylinder
articulated at either end with the adjacent centra. The cavity
thus present between successive centra is filled with soft
material which may be, in part at least, derived from the
embryonic notochord.
• In most living amniotes, however, the notochord has disappeared
except for, at the most, thin pads of tissuelying between successive
centra, and the centra have become solid structures. whose end faces
are closely apposed to those of their neighbors. Those vertebrae which
have flat terminal faces, as is common in mammals, are termed
acelous(凹形)). In many cases one end of the centrum expands in
spherical fashion(球形), to be received in a hollowed socket in the
neighboring centrum. lf the concave socket is at the anterior end of the
centrum (a condition found in most living reptiles ) the vertebra is
termed procelous . if posteriorly placed, opisthocelous. Since the
nature of the articular faces may change reglonally along the comlumn,
various irregularities:and combinations of structure may occur, and
there are numerous variants from the orthodox types. Notable is the
development in the neck region of birds of heterocelous vertebrae,
with saddle-shaped(马鞍形) articular faces between the centra, and
the development of complicited central connections in the retractable
neck of turtles.
• The base of each hemal arch consists of a pair of processes which
descend from the intercentrum to enclose a space in which lie the
major blood vessels of the tail; usually the two branches join below
the vessels to give V or Y shape as seen in end view. As will be seen
later. the hemal arches are represented in fishes not only in the tail, but
in the trunk as a type of rib--the ventral rib--not present in land
vertebrates.
•
In most cases a rib attachment is present in the centrally region.
Primitively this was with the intercentrum, but in some ammotes this
point of attachment shifts to the centrum, where there may develop a
special process, the parapophysis(横突), or it may come to lie. in
the trunk of mammals, at the point of junction between two successive
centra.
• The upper surface of the centrum froms the floor of the canal carrying
the spinal cord. At either side rises the broad base of a neural arch
(神经弓). firmly bound to the centrum in most adults. Above the
spinal cord the two arches fuse to form a neural spine. which runs
upward between the dorsal muscles of the two sides; successive spines
are usua11y bound together by an elastic ligament. At the base of the
arch on either side there is frequently a stout transverse process
(diapophysis) as a rib attachment.
骨骼系统
• 鱼类具有由典型的椎骨组成的脊柱、完整的头骨、带骨、附肢骨 。
软骨鱼类全部骨骼为软骨(cartilage),但有钙盐沉着而变得坚硬。
• 脊柱由双凹型的椎骨组成,分为尾椎和躯干椎,躯干椎具有肋骨。
• 硬骨鱼类的骨骼骨化程度加强,骨片增加,并更加复杂。肩带与头
骨愈合并加固,使头、肩带、躯干形成一个稳定支架,增加了运
动的力度。腰带不直接与脊柱相连。
• 6.1脊柱
• 鱼类脊柱的分化程度很低,脊椎只有两种,即躯干椎(trunk
vertebra)与尾椎(caudal vertebra)。
• 躯干椎附有肋骨(rib),尾椎特具血管弧,以此容易把二者区别开
来。每一脊椎的椎体(centrum),前后两面都是凹型,称为双凹型
椎体(amphicoelous centrum),为鱼类所特有。在相邻的两个椎体
之间的空隙以及贯穿椎体中心的小管内,还有脊索的残余存留。
• 6.2头骨 鱼类的头骨由比较容易区分的脑颅和咽颅两部分组成。
• 6.2.1脑颅
• 软骨鱼的脑颅,构造简单,只是一个软骨脑箱保护着脑
部。硬骨鱼的脑颅则由许多骨片所合成。这些骨片中,
一部分是软骨性硬骨(cartilage bone),如几块枕骨、
几块耳骨、几块蝶骨,几块筛骨等;另一部分是膜性硬
骨(membrane bone),如鼻骨、额骨、顶骨、犁骨等。
• 6.2.2咽颅(visceral skeleton)
• 鱼类的咽颅(visceral skeleton)最为发达,由一对颌弓
(mandibular arch)、一对舌弓(hyoid arch)、五对鳃弓
(branchialarch)共同组成。
• 6.3 附肢骨骼 可分为奇鳍骨骼与偶鳍骨骼两部分。
• 奇鳍中的背鳍和臀鳍,在基部有鳍担骨,在鳍中有鳍条
支持。尾鳍无鳍担骨。偶鳍骨骼包括带骨(肩带和腰带)
及支持鳍的骨骼(鳍担骨、鳍条)。鱼类成对的附肢骨骼
没有和脊柱发生联系,这也是它的特点之一。
肌肉系统
• 鱼类主要的肌肉是躯干部分节的肌节,由水平生
骨隔分为轴上肌和轴下肌,轴上肌发达而有力,
几乎是整个身体重量的一半。鱼的肌节呈锥形漏
斗状,彼此套叠,在鱼体的横切面上呈现一系列
同心圆状。这样一个肌节的收缩可以延伸几个骨
节,快速传递收缩的力量。作为附肢的偶鳍背腹
两面均有肌肉附着在鳍骨上,背面为伸肌,腹面
为屈肌,其收缩可使鳍依附躯体做整体运动,即
上提、外展和下降、内收。
• Electric fishes
•
The ability to produce strong electric shocks is confined to two
groups of vertebrates: elasmobranchs(for example, electric ray) and
teleosts (for example, electric eel, electric catfish). Best known and
studied is the famous electric eel, Electrophorus. of the Amazon and
Orinoco river systems of South America. This large, sluggish creature
contains powerful electric organs that in 4- to 8-foot adults can
produce paralyzing discharges exceeding 600 volts, quite ample to
stun or kill its prey, or discourage potential enemies including large
mammals. People who have accidentally met the electric eel under the
latter's own terms report it is about as pleasant as contacting an
uninsulated high-voltage wire. It is not necessary for the eel to
actually touch its victims since the shocks stretch out in an electric
field for many feet around the fish. Even small eels Iess than a foot
long can discharge pulses exceeding 200 volts. The source of this
impressive performance is the pair of electric organs lying on either
side of the vertebral column and extending almost the entire length of
the fish.
• Together they make up about 40% of the bodyweight. Each organ is
composed of longitudinal columns of 6,000 to l0,000 thin, wafer like
plates (electroplaxes) stacked one upon another like a long cylinder of
coins. There are about 60 such columns in each organ. Each plate, or
electroplax, is a modified muscle cell innervated by a nerve fiber and
has very special electrical properties. One side of the plate is a
nervous layer that will depolarize when stimulated; the opposite side
is a nutritive layer that remains inactive. All the nervous sides of the
plates face the tail of the animal. When the eel chooses to discharge its
organ, motor impulses are sent out to the electric organs from a
special neural center in the brain. These impulses travel to the
numerous plates in a highly synchronized way so that all of the
thousands of plates discharge simultaneously. Each plate develops a
potential charge of about l50 millivolts and because the plates are
arranged in series, the individual voltages summate like batteries
arranged in series. Thus a high-voltage current flows from the tail to
the head of the eel and completes the circuit in the water surrounding
the eel. Electric eels must develop high voltages to overcome the high
resistance of the freshwater in which they live.
• Electric fishes living in the sea, such as the electric
ray, Torpedo ,live in a lower sistance medium
where high voltages are not required. They generate
high amperages instead. In Torpedo each electric
organ contains some 2,000 columns made up of
about l,000 electroplaxes arranged in vertical stacks.
With fewer plates per column, the voltage produced
is relatively low (about 50 volts), but the power
output may be as much as 6 kilowatts! It is easy to
understand why, when seasoned commercial
fishermen happen to pick up one of these animals in
their trawl, they leave it strictly alone until it is dead
and safe to handle.
• 鱼类中有些种类具有发电器官,能放电以进行防
御或攻击。产于太平洋、大西洋和印度洋的电鳐
(torpedo marmorata),产于南美河流中的电鳗
(Eletrophorus electricus)等都能放电。它们的发电
器官都是由肌肉转变而来。电鳐的发电器在胸鳍
内侧、头的后方,放电量可达100伏特左右;电
鳗的发电器位于尾部,与体重之比为1:2.66,
放电时电压最大可达600伏特以上 。
• 产于非洲河流中的电鲇(Malapterurus electricus),
放电量为400—450伏特,但其发电器官并非肌肉
变成,乃是真皮腺的转化。
神经系统和感觉器官
• 鱼类的神经系统具有下列的几个特点
• 12.1神经系统
• 鱼类脑的结构较圆口类发达,明显分为5部分,
即大脑、间脑、中脑、小脑和延脑。其中软骨鱼
类的大脑较硬骨鱼类发达,脑顶部已出现神经物
质。大脑的主要部分是纹状体,以感嗅为主。脑
虽可分为明显的五部,但大脑所占的比例还是很
小,而且硬骨鱼类的大脑背面还只是上皮组织,
没有神经细胞。适应水生的鱼类在间脑底部有一
个突出的富含血管的血管囊,在深海鱼类尤为发
达,是一个水深度和压力的感受器。脑神经10对,
脊髓在每1体节发出1对脊神经。
感觉器官
• 12.2
• 12.2.1嗅觉
• 发达,有1对外鼻孔与1对嗅囊相连。鲨鱼的嗅觉尤为发
达,可在海水中嗅出稀释到百万分之一浓度的血液。
• 12.2.2听觉
• 听觉器官仅有内耳,由3个半规管、椭圆囊和球状囊组
成,主要是平衡觉。硬骨鱼类的鲤形目鱼类具有特有的
韦氏小骨,由前3块躯干椎的一部分演化而来,可将鳔
内气体的振动传给内耳的淋巴系统,从而产生听觉。
• 12.2.3视觉
• 鱼眼具有脊椎动物眼的基本模式,即3套膜(巩膜、脉络
膜和视网膜)和1套折光系统(角膜、房水、晶体和玻璃
体) 。
• 鱼眼的角膜平坦,无聚光作用。
• 晶体呈圆球形,没有弹性,其曲度又不能改变,
只能靠晶体后方的镰状突起(faliciform process)来
调节晶体和视网膜之间的距离,所以鱼类是近视
的。
• 软骨鱼类的晶体较靠后,适于远视;硬骨鱼类晶
体靠前,适于近视。
• 大多数鱼类由于水生而无眼睑。一些能短暂离水
爬行的鱼类如弹涂鱼等,适应了空气中的干燥环
境,也具有眼睑。鲨鱼的眼球外面有一片活动的
第三眼睑即瞬膜(nictitating membrane)遮盖着,鲻
鱼类还有所谓脂质眼睑,这些构造的作用,尚不
十分清楚。由于水生生活,鱼类没有泪腺。
Reproduction
•
The teleosts show many types of sexual reproduction
patterns. Although the hermaphroditic condition may
occasionally occur abnormally in many species, only one or
two families (for example, Serranidae)are truly
hermaphroditic. In hermaphroditic forms the gonads are
each divided into testicular and ovarian zones.
• The fish (Poecilia formosa) discovered in Texas illustrates
well the odd type of parthenogenesis called pseudogamy, or
gynogenesis. This process involves the entrance and
activation of an egg by a spermatozoon whose nucleus does
not make genetic fusion with the nucleus of the egg. Sperm
are furnished by males of related species, but the
descendants are all like that of the female because they are
genetically alike.
• The testes are usually elongated, whitish organs divided into lobules
that contain cysts of maturing germ cells. Within each cyst the
maturing cells are always of the same stage of development. The
lobules open into the spermatic duct (with secretory lining),which
runs into the urogenital sinus. Males of ten become sexually mature
before the females, and their testes may be active throughout the year;
in others the testes have seasonal rhythms in reproductive activity. The
ovaries may run the length of the abdominal cavity and are made up of
many ovarian follicles supported by connective tissue. The ovary, with
its membranous covering, may be continuous with the oviduct, or the
ovaries may be naked and discharge their eggs into the peritoneal
cavity, whence they are picked up by the oviducts (mullerian ducts).
The paired oviducts may open through a common urogenitall pore
behind the anus, or they may open through genital pores. Some fish
such as trout and salmon have no oviducts; others such as the
freshwater eel have neither sperm ducts nor oviducts. Usually eggs are
produced during a seasonal rhythm and the ovaries are quiescent at
other times. A few (for example, hake) are known to have active
ovaries at all times. Some fish such as the cod produce enormous
numbers of eggs (9 million have been found in the ovaries of a single
female).
• Most teleost fish are oviparous, laying eggs that are
fertilized in the water by the sperm discharged by
the male, usually in contact with the female.
Gametes in water have limited viability unless they
are united. Certain fish, however, are ovoviviparous
(young born alive but nourished by egg yolk in
mother) or viviparous (young with type of placental
attachment to wall of uterus). Ovoviviparity or
viviparity is found in the mosquito fish
(Cambusia)and certain sea perch (Embiotocidae) as
well as a few others.
•
Male fish have evolved many devices for transferring
sperm into the female. Copulatory organs include the
urogenital papillae, anal fins, and other specialized
structures. Fertilization usually takes place while the egg is
still in the ovarian follicle. In Rhodeus the female lays her
eggs in the gill spaces of afresh water mussel by means of a
very long urogenital papilla. In fish that are ovoviviparous,
the young develop within a cavity of the oviduct.
• Soon after eggs are laid in water, they take up water and
harden. Cleavage (meroblastic) occurs in the blastodisk of
the zygote and a blastoderm is formed. As cleavage
continues, the blastoderm spreads over and encloses the
yolk mass. The space between the blastoderm and yolk is
the segmentation cavity, or blastocoel. Development
proceeds, and eventually a larval form of fishlike
appearance with a large yolksac is hatched. Temperature has
a great effect on regulating the speed of hatching and
subsequent development.
生殖系统
• 鱼类的生殖系统一如陆生的脊椎动物,由生殖腺
(精巢和卵巢)及输导管(输精管和输卵管)共同组
成。一般的说,软骨鱼类雄鱼的输精管(vas
deferens)由中肾管转变而来,雌鱼的输卵管
(oviduct)则由一部分前肾管形成。
• 硬骨鱼类不利用肾管而是利用腹膜所围成的管道
来充作生殖腺的输导管。这样的输导管在雌体中
直接与卵巢连接,成熟的卵细胞不经过体腔就进
入输卵管。这种现象在其它脊椎动物是没有的。
有些鱼类,例如鲑鱼等甚至没有输导管,生殖细
胞成熟以后落入体腔,再由体腔经过特殊的生殖
孔排到体外。
• 大多数鱼类都是雌雄异体。两性在形态上的差别,
一般不显著
• 许多鱼在生殖季节,雄体常发生一些变化,容易
辨别,例如一些鲤科鱼(青、草、鲢、鳙、鲤等)
的雄体,胸鳍上会出现“追星”(由表皮细胞角
化而成的锥状突起);刺鱼、斗鱼等的雄体,颜
色变得很鲜艳,都属于雌雄异形。
• 在雌雄同体的鱼中,鲔属(Serranus)的许多种为永
久性的雌雄同体,而且能自体受精。
• 黄鳝的生殖腺,从胚胎到成体都是卵巢,只能产
卵。产卵过后,卵巢逐渐转化为精巢,育成精子
这种雌雄性的转变,称为性逆转(sex reversal)。
•
•
•
•
在生殖方式上,软骨鱼类较硬骨鱼类进化。
软骨鱼类为体内受精,受精卵的发育有3种方式。
一是卵生(体外发育,如虎头鲨),
二是卵胎生(卵滞留在子宫内靠自身卵黄发育完全后产
出,如棘鲨Echinorhinus brucus),
• 三是假胎生(卵在子宫内靠自身的卵黄发育,但发育后
期,卵黄囊与母体子宫壁相连形成卵黄囊胎盘,由母体
供给后期营养,如星鲨Mustelus mustelus)。
• 行体内受精的鱼,其雄体具有交接器官,借以把成熟的
精子注入雌体的生殖管道内,例如软骨鱼类的鳍脚
(clasper),(由腹鳍的一部分变来)和虹鳉的生殖足
(gonopodium)。
• 硬骨鱼类绝大部分为体外受精和发育,雌鱼产卵时雄鱼
在一旁同时排精。雌鱼产卵量大,如翻车鱼一次产卵达
3亿粒,但成活率低。多数硬骨鱼产卵受精后就各自分
离。但有少数鱼类交配前有筑巢习性,并对受精卵和幼
鱼加以保护,甚至将受精卵含在口中、鳃腔中或身体上
的特化囊袋中进行孵化。
• 鱼类受精和发育的关系,有如下的几种类型:
• 体外受精,体外发育。见于绝大多数鱼类。
• 体外受精,体内发育。卵子在体外受精后纳入亲
体内部去发育。例如非洲鲫鱼(Tilapia
mossambica )的雌体,把受精卵吞入口中孵化。
鲇科的一种鱼(Tachysurus barbus),其雄体在生殖
期间停食,把受精卵吞入胃中去孵化。
• 体内受精,体外发育。卵未产出前即在雌体的生
殖道受精,受精后不久,卵即产出体外完成发育
过程。例见于软骨鱼中的虎鲨及硬骨鱼中的霍鳉
(Horaichtny)等。
• 体内受精,体内发育。
• Class Elasmobranchii(板鳃亚纲).
• Six living orders, three extinct orders Subclass
Selachii (se-lay'kee-i) (Gr. selachos. fish having
cartilage instead of bones)--modern
elasmobranchs. Of the six orders, the two most
important are below:
• Order Squaliformes(角鲨目)--modern sharks
and rays.'
• Order Rajiformes(鳐形目)--skates.
• Distinctive characteristics: The body of a shark such as a dogfish
Shark is fusiform or spindle shaped. In front of the ventral mouth is a
pointed rostrum; at the posterior end the vertebral column turns up to
form the heterocercal tail. The fins consist of the paired pectoral and
pelvic fins supported by appendicular skeletons, two median dorsal
fins (each with a spine in squalus), and a median caudal fin. A median
anal fin is present in the smooth dogfish opstelus). In the male the
medial part of the pelvic fin(腹鳍) is modified to form a clasper
(鳍脚), which is used in copulation(交配). The paired nostrils
(blind pouches) are ventral and anterior to the mouth. The lateral eyes
are lidless, and behind each eye is a spiracle(喷水孔) (remnant of
the first gill slit). Five gill slits are found anterior to each pectoral fin
(胸鳍). The leathery skin is covered with placoid scales(盾鳞)
(dermal denticles), each of which consist of a wide basal plate of
dermal dentin and a spine covered with vitrodentin, or a shiny enamellike dentin. These scales are modified to form teeth in the mouth and
are the remnants of the dermal plates of placoderms. A well-developed
lateral line system serves as a "distance touch" in water for detecting
and locating objects and moving animals (predators, prey, and social
partners). It is composed of a canal system extending along the side of
the body and over the head. The canal opens at intervals to the surface.
Inside are special receptor organs (neuromasts) that are extremely
sensitive to vibrations and currents in the water.
鱼纲分类
•
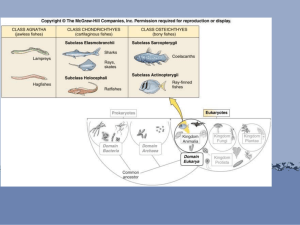
现存鱼类约22 000种,分为软骨鱼类(Chondrichthyes)
和硬骨鱼类(Osteichthyes)两大类群。我国产鱼类共2
830余种。
•
一、软骨鱼类
•
内骨骼全为软骨的海生鱼类;体被盾鳞;鼻孔腹位;
鳃间隔发达,鳃孔5-7对。鳍的末端附生皮质鳍条。歪
型尾。无鳔和“肺”。肠内具螺旋瓣。生殖腺与生殖导
管不直接相连;雄鱼有鳍脚,营体内受精。全世界约
800种,我国产190多种,绝大多数分布于热带及亚热带
海洋,包括两个亚纳。
•
(-)板鳃亚纲( Elasmobranchii) 体呈梭形或盘形。
鳃孔 5- 7对,各自开口于体外而无鳃盖;上颌不与颅
骨愈合。雄性仅有位于腹鳍内侧的鳍脚。共2总目。
• l.鲨形总目( Selachomorpha)体呈梭形,鳃孔侧位,
故又称侧孔总目( Pleurotremata)。胸鳍与头侧不愈合;
背鳍背位;歪型尾。
• ( l)六鳃鲨目( Hexanchiformes):我国产六鳃鲨科
( Hexanchidae)的3属3种,最常见的是分布于黄海、
渤海的扁头哈那鲨(Notorhynchus platycephalus),体
长4一5m,重200kg一300kg。鳃孔7个。底栖生活,游
泳缓慢.吞食小鱼和甲壳动物。卵胎生。肝含油量达65
%-70%,可制鱼肝油。东海和南海偶尔也见灰六鳃鲨
(Hexanchus griseus)。
• 灰六鳃鲨(Hexanchus griseus)
• (2)真鲨目(Carcharhiniformes):眼有瞬膜,是世界
上软骨鱼类中属、种最多的一个类群,共8科47属200余
种,我国产4科对属60多种,常见种类有体具斑点的梅
花鲨(Halaefurtus burgeri)、星鲨(Mlustelus)、颌齿
斜行排列的斜齿鲨(Scolidon sorrakowah)和锤头双告
鲨等。
• 斜齿鲨(Scolidon sorrakowah)
• (8)扁鲨目(Squatiniformes):是鲨形总目中惟一体
型平扁的类群。胸、腹鳍扩大,且彼此接近;背鳍2个,
形小而位于尾部上方。本目仅扁鲨科(Squatinidae)l属,
我国产2种,常见种为日本扁鲨( Squatina japonica),
俗称琵琶鲨。
• 日本扁鲨( Squatina japonica)
• 鳐形总目(Batomorpha)体形扁平,鳃孔腹位,又名下孔总目
(Hypotremata);胸鳍前部与头侧相连;背鳍常位于尾上;无臀鳍;尾鳍或
有或无。全世界有4目20科约430种,我国产约80种
• (l)锯鳐目(Pristiformes):吻狭长而平扁,似剑状突出,边缘具尖利的吻
齿。本目只有锯鳐科Pristidae),分布于热带和亚热带沿岸海区,有些种类也
可进入江河生活。我国南海和东海产尖齿锯鳐(Pritis cusridatus),鱼体最长
可达9 m,常用剑状的吻锯击毙或刺伤追食对象。
•
• 尖齿锯鳐(Pritis cusridatus)
•
(2)鳐形目(Rajiformes):吻圆钝或突出,侧缘无
吻齿。本目主要包括体盘呈犁状的、体盘呈团扇形的团
扇鳐(Platyrhinus sinensis)和体似斜方形的鳐鱼(Rala)
等。
• 犁头鳐(Rhinobatus hynnicephalus)
• 3 电鳐目
(Torpediniformes):体
盘椭圆形,皮肤光滑柔软;
头侧与胸鳍之间的皮下具
有特化而成的发电器官。
我国有2科4属 8种。常见
种类有产于南海的黑斑双
鳍电鳐(Narcine maculate)
和日本单鳍电鳐( Narke
japonica)等。
Class Holocephali(全头亚纲)
• Class Holocephali(全头亚纲)—chimaeras(银鲛) The
members of this small group, distinguished by such suggestive names
as ratfish, rabbitfish, spookfish and ghostfish, are remnants of an
aberrant line that diverged from the elasmobranches at least 300
million years ago (Carboniferous or Devonian). Fossil chimaeras
were first found in the Jurassic, reached their zenith in the Cretaceous
and early Tertiary (l20 million to 50 million years ago), and have
declined ever since. Anatomically they present an odd mixture of
sharklike and bony fish--like features. Instead of a toothed mouth,
their jaws bear large flat plates. The upper jaw is completely fused to
the cranium, a most unusual development in fishes. Their food is
seaweed, mollusks, echinoderms, crustaceans, and fishes—all in all a
surprisingly mixed diet for such a specialized grinding dentition.
Chimaeras are not commercial species and are seldom caught. Despite
their grotesque shape, they are beautifully colored with a pearly
iridescence and have vivid emerald-green eyes that must be seen to be
believed.
(二)全头亚纲(Holocephali)
• 体表光滑或偶有盾鳞。鳃腔外被一膜质
鳃盖,后缘具一总鳃孔。背鳍2个第一背
鳍前有一强大硬棘,能自由竖立或垂倒。
雄性除内侧的鳍脚外,尚有腹前鳍脚及
额鳍脚。全世界有1目3科近30种。我国
产5种,最常见的为银鲛科(Chimaeridae)
的黑线银鲛(Chimaera phantasma)_俗
名海兔子.
黑线银鲛(Chimaera phantasma)
Class Osteichthyes (Teleostomi 硬骨鱼)
• Class Osteichthyes (Teleostomi 硬骨鱼)-bony fish .
In no other major animal group do we see better
examples of adaptive radiation than among the bony
fishes. Their adaptations have fitted them for every
aquatic habitat except the most completely
inhospitable. Body form alone is indicative of this
diversity. Some have fusiform (streamlined) bodies
and other adaptations for reducing friction.
Predaceous, pelagic fish have trim, elongate bodies
and powerful tail fins and other mechanical
advantages for swift pursuit. Sluggish bottomfeeding forms have flattened bodies for movement
and concealment on the ocean floor.
• The elongate body of the eel is an adaptation for
wriggling through mud and reeds and into holes and
crevices. Some, such as pipefishes(尖嘴鱼), are
so whiplike that they are easily mistaken for
filaments of marine algae waving in the current .
Many other grotesque body forms are obviously
cryptic or mimetic adaptations for concealment from
predators or as predators. Such few examples cannot
begin to express the amazing array of physiologic
and anatomic specializations for defense and offense,
food gathering, navigation, and reproduction in the
diverse aquatic habitats to which bony fishes have
adapted them selves. Some of these adaptations are
described in the pages that follow.
• The Osteichthyes are divided into three clearly distinct
groups: Crossopterygii (lobe--finned fishes 总鳍鱼类),
Dipneusti (lungfishes肺鱼类), and Actinopterygii (rayfinned fishes 辐鳍鱼类).
• What are their origins? As pointed out earlier in this chapter,
it seems safest to steer a middle course through current
paleontologic debate and assign equal rank to all three
groups within the class Osteichthyes. In other words, it is
impossible to decide which one of these three groups, if any,
might have served as ancestral stock for the other two. It is
apparent from the fossil evidence that all three groups were
distinct in the Devonian, some 400 million years ago. They
are believed to have descended from an acanthodian of the
Silurian, perhaps a creature similar to the fossil Climatius.
• Classification of the Osteichthyes(Teleostomi)
•
Class Osteichthyes --bony fish. Three subclasses,
69 orders.
•
Subclass Crossopterygii --lobe-finned fish.
Three extinct orders, one living order
(Coelacanthoidei) containing one species,
Latimeria(矛尾鱼).
•
Subclass Dipneusti --lungfishes. Four extinct
orders, two living orders; these are Ceratodiformes
containing the genus Neoceratodus. and
Lepidosireniformes containing two genera,
Lepidosiren and Protopterus.
•
Subclass Actinopterygii --ray-finned fishes.
Three superorders and 59 orders.
二、硬骨鱼类
•
骨骼大多由硬骨组成;体被骨鳞或硬鳞,一
部分鱼类的鳞片有次生性退化现象;鼻孔位于
吻的背面;鳃间隔退化,鳃腔外有骨质鳃盖骨,
头的后缘每侧有一外鳃孔。鳍的末端附生骨质
鳍条,大多为正型尾。通常有鳔,肠内大多无
螺旋瓣;生殖腺外膜延伸成生殖导管,二者直
接相连。无泄殖腔和鳍脚,营体外受精。硬骨
鱼类包括2亚纲。
Subclass Crossopterygii
This group has had the least spectacular evolutionary
radiation of all the Osteichthyes. It is by far the most
important in an evolutionary sense because the tetrapods
arose from one or more of their ancient members. The
crossopterygians had nostrils that opened into the mouth
(choanae), lungs as well as gills, and paired lobed fins. They
first appeared in Devonian times, a capricious period of
alternating droughts and floods when their lungs would
have been a decided asset, if not absolutely essential, for
their survival. They used their strong lobed fins as four legs
to scuttle from one disappearing swamp to another that
offered more promise for a continuing aquatic existence.
• The crossopterygians are divided broadly into two groups.
The rhipidistians (骨鳞鱼目) appeared in the Devonian,
flourished in the late Paleozoic, and then disappeared. This
is the stock from which the amphibians have descended (Fig.
22-l). Among the primitive characteristics of the
rhipidistians were the fusiform shape, two dorsal fins, and a
heterocercal tail. The paired fins bore sharp resemblances to
a tetrapod limb, for they consisted of a basal arrangement of
median or axial bones, with other bones radiating out from
these median ones. Some of the proximal bones seem to
correspond with the three chief bones of the tetrapod limb.
The scales of these primitive fish were of the cosmoid type,
a thick complex scale of dentinlike cosmine, enamel, and
vascular pulp cavities. This type of scale is not found in
modern fish but has been replaced by the bony cycloid type.
• The other group of crossopterygians was the coelacanths. These also
arose in the Devonian, radiated somewhat, and reached their
evolutionary peak in the Mesozoic. At the end of the Mesozoic they
also disappeared but left one remarkable surviving species, the living
coeIacanth Latimeria chalumnae. Since the last coelacanths were
believed to have become extinct 60 million years ago, the
astonishment of the scientific world can be imagined when the
remains of a coelacanth were found on a dredge off the coast of South
Africa in l938. An intensive search was begun in the Comoro Islands
area near Madagascar where, it was learned, native Comoran
fisherman occasionally caught them with hand lines at great depths.
By the end of l97l, a total of 69 specimens had been caught, many in
excellent condition, although none have been kept alive after capture.
The "modern" marine coelacanth is a descendant of the Devonian
freshwater stock. The tail is . of the diphycercal type but possesses a
small lobe between the upper and lower caudal lobes, producing a
three-pronged structure. Coelacanths also show some degenerative
features, such as more cartilaginous parts and a swim bladder that was
either calcified or else persisted as a mere vestige. They also lack the
internal nostril so characteristic of crossopterygians, but this is
probably a secondary loss after the adoption of a deep-sea existence;
obviously neither nostrils nor functional lungs have any relevance for
such a life habit.
• Subclass Dipneusti The lungfishes are considered by many
paleontologists to be an offshoot of an early rhipidistian.
This view is contested by a minority of paleontologists,
among them the influential Swedish paleontologist E.
Jarvik, who prefers to leave their ancestry unassigned. The
lungfishes were a distinct group in the Devonian. The
earliest known lungfish, Uranolophus.shows a much closer
relationship to the crossop-terygians than do existing forms,
for it had a heterocercal tail, two dorsal fins, cosmoid scales,
and ossified shoulder girdle--structures that have undergone
considerable modification in modern lungfishes.
Evenmodern lungfishes resemble the crossoptery gians with
their lobe-shaped paired fins and lungs. At thesame time all
lungfishes, extinct or living, differ fromcrossopterygians in
several significant skeletal features, including the totally
different origin of theinternal nostrils.
• The three surviving genera of lungfishes have descended
from Devonian ancestors suchas Uranolophus or the
somewhat later fossil Dipterus.The least specialized is
Neoceratodus. the living Australian lungfish, which may
attain a length of s feet. This lungfish is able to survive in
stagnant, oxygen-poor water by coming to the surface and
gulping air into its single lung, but it cannot live out of
water. The south American lungfish Lepidosiren and the
African lungfish Protopterus are evolutionary sidebranches
of the Dipneusti, and they can live out of water for long
periods of time. Protopterus lives in African streams and
rivers that run completely dryduring the dry season, with
their mud beds bakedhard by the hot tropical sun. The fish
burrows downat the approach of the dry season and secretes
acopious slime that is mixed with mud to form a hard
cocoon in which it estivates until the rains return .
(一)内鼻孔亚纲(Choanichthyes)
• 本亚纲鱼类的口腔内具有内鼻孔(choana) ;有原鳍
型的偶鳍,即偶鳍有发达的肉质基部,鳍内有分节的
基鳍骨支持,外被鳞片,呈肉叶状或鞭状,故又称肉
鳍亚纲(Sarcopterygii)。肠内有螺旋瓣。共有2总目。
•
1.总鳍总目(Crossopterygiomorpha)这是一类出
现于泥盆纪的古鱼,也是当时数量最大的硬骨鱼类。
具有中轴骨骼是一条纵行的脊索而不存在椎体、颏下
有一块喉板(gular Plate)、肠内有螺旋瓣等一系列原
始特征。早期的总鳍鱼类都栖居于淡水中,有鳃、鳔
和内鼻孔,能在气候干燥和水域中周期性缺氧期间用
鳔呼吸空气。
• 总鳍鱼类一直被认为已于中生
代末期的白垩纪时完全绝灭,
但是1938年12月22日却在非洲
东南沿海哈隆河河口水深70m
首次捕获一尾体长1.8m、重
95kg的总鳍鱼,并依据其尾形
定名为矛尾鱼(Latimeria
chalumnae),该标本被保存
于东伦敦博物馆内(图16一
33)。以后又在科摩罗伊斯兰
联邦共和国科摩罗群岛附近
的海域中陆续捕得150-200尾
矛尾鱼。这是总鳍鱼类惟一现
存的残留种,隶属于腔棘鱼目
( Coelacanthiformes)、矛尾
鱼科( Latimeriidae)。这些
古鱼的孓遗已成为动物界最珍
贵的活化石之一。
• 2.肺鱼总目(Dipneustomorpha)本总目是与古总鳍
鱼类亲缘关系较近的同时代的鱼类,两者的主要区别是
肺鱼类口腔中有内鼻孔、偶鳍内具双列式排列(辐鳍骨
列于基鳍骨的两侧)的鳍骨和高度特化而适于压碎软体
动物硬壳的迷齿。肺鱼类一方面具有某些原始特征:另
一方面还具有与其生活环境相适应的特化性状或进化特
征。例如,肺鱼类的鳔不论在发生、构造或是呼吸机能
上,都与陆生脊椎动物的肺十分相似。鳔有鳔管与食道
相通,外界的空气可通过口腔和内鼻孔直接进人鳔内。
• 与鳔呼吸相联系的心脏及血液循环方式也发生了相应变
化:心房被不完全的房间隔分成左、右两半,在鳔内经
气体交换后的多氧血流回心房的左半边,从身体回归的
少氧血流进心房的右半边,是脊椎动物进化过程中首次
出现包括体循环和肺循环的双循环。本总目在世界各地
曾有过广泛的分布,但现生种类仅2目3科5种,并被隔
离分布于南美洲、非洲和大洋洲,我国四川省境内也出
土过肺鱼化石。
Subclass Actinopterygii
• This huge assemblage contains all our familiarbony fishes. The fossil
record reveals that the group had its beginnings in Devonian
freshwater lakes andstreams. The earliest actinopterygians were small
fish with large eyes and extended mouths. Their tails were
heterocercal. They had a single dorsal fin and a single anal fin: paired
fins were representedby the anterior pectoral fins and the posterior
pelvicfins. Their skeletons were largely bone. Their trunksand tails
were encased in an armor of heavy, rhombicscales (ganoid). Most of
these early fish had functional lungs, as did all the Devonian fishes,
but theylacked internal nostrils. All had gills (five pairs orless) and
spiracles. These early actinopterygians be-longed to the order
Palaeonisciformes. One common genus in the fossil record was
Cheirolepis. a generalized type that had some resemblance to certain
acanthodians. The palaeoniscids were distinctly different from their
freshwater contemporaries, the crossopterygians and the lung-fishes,
with whom they shared the Devonian swampsand rivers. The
palaeoniscids were rare in the Devonian, became a more secure group
in the Carbonif-erous and Permian, but nearly disappeared in the
triassic when their descendants returned to the sea.
• In their evolution, the actinopterygians have passed through
three stages. The most primitive group are the Chondrostei,
represented today by the freshwater and marine sturgeons
(Fig. 22-l4) and the bichir,Polypterus. of African rivers (Fig.
22-l). Polypterusis an interesting relic with a lunglike swim
bladderand many other primitive characteristics; it
resemblesan ancient palaeoniscid more than any other living
descendant. There is no satisfactory explanation for the
survival to the present of certain fish such as this one and
the coelacanth Latimeria when all of their kin perished
millions of years ago.
•
A second actinopterygian group is the Holostei. There
were several lines of descent within this group, which
flourished during the Triassic and Jurassic. They declined
toward the end of the Mesozoic as their successors, the
teleosts, crowded them out. But they left two surviving lines,
the bowfin, Amia (Fig. 22-l),of shallow, weedy waters of the
Great Lakes and Mississippi valley, and the garpikes (or
simply gars)of eastern North America (Fig. 22-l5).
• The third group is the Teleostei, the modern bony fishes (Fig. 22-l6).
No definite statements can be made about any criteria of success as
they apply to modern teleosts. Diversity appeared early in teleostei
evolution, foreshadowing the truly incredible variety of body forms
among teleosts today. The skeleton of primitive fish was largely
ossified, but this condition regressed to a partly cartllaginous state
among many of the Chondrostei and Holostei. Teleosts, however, have
an internal skeleton almost completely ossified like the primitive
members. The dermal investing bones of the skull (dermatocranium)
and the chondrocranium (endocranium) around the brain and sense
organs form a closer union among the teleosts than they did in the
primitive bony fish. Other evolutionary changes among the teleosts
were the movement of the pelvic fins forward to the head and thoracic
region, the transformation of the lungs of primitive forms into air
bladders (Fig. 22-l6) with hydrostatic functions and without ducts, the
changing of the heterocercal tail of primitive fish and of
theintermediate super orders into a homocercal form, and the
development of the thin cycloid and ctenoid scales from the thick
ganoid type of early fish. Among other changes were the loss of the
spiracles and the development of stout spines in the fins, especially in
the pectoral, dorsal, and anal fins.
(二)辐鳍亚纲(Actinopterygii)
• 本亚纲鱼类的各鳍均由真皮性的辐射状鳍条支持。
体被硬鳞、圆鳞或栉鳞,或裸露无鳞。无内鼻孔。
种类极多,占现生鱼类总数的90%以上,共包括
9总目、36目。产于我国的有8总目、26目。
• 1.硬鳞总目(Ganoidomorpha)本总目是鱼类中古老
类群的残余种,除了具有硬骨鱼类的主要特征外,仍留
有一些原始性状:体被外覆硬鳞质( ganoid)的菱形硬
鳞 ;心脏具动脉圆锥;肠内有螺旋瓣;尾鳍为歪型尾;
颏部常有喉板。包括4目,即鲟形目、多鳍鱼目、弓鳍
鱼目和省鳝目等(图16-35)。
•
(1) 鲟形目(Acipenseriformes):体形似鲨,吻长,
口腹位,有喷水孔。躯干部有5行纵列的骨板,或皮肤
裸露而仅在歪型尾的上叶列有少数硬鳞性质的叉状鳞。
我国产2科。
• 多鳍鱼目(Polypteriformes):背鳍分离为一列小鳍,故得名多
鳍鱼,每个小鳍前方各有一枚鳍棘。胸鳍有肉质的基叶,然其鳍
骨却与总鳍鱼明显不同。尾鳍为圆形的正型尾。鳔分2叶,内多
分隔,开口于食道腹面。幼鱼期有外鳃。主要种类为具有腹鳍的
多鳍鱼( Polypterus bichir)和体形似鳗而腹鳍退化的芦鳗
(Calamoichthys congicus)等。多鳍鱼类全产在非洲热带河流中,
栖于水下的泥底,性情凶猛,以小鱼、甲壳动物、昆虫为食;当
河湖接近干涸时,可用鳔直接呼吸空气。
• c 多鳍鱼( Polypterus bichir) d 芦鳗(Calamoichthys congicus)
• 弓鳍鱼目(Amiiformes):体被圆形硬鳞,颏下有一大形喉板。
鳔内分成许多小室,为辅助呼吸器官。中生代中期曾是弓鳍鱼类
全盛时期,而现存种类仅一种,即弓鳍鱼(Amia ciava)。弓鳍
鱼两颌具细齿,肠螺旋瓣和动脉圆锥极度退化,并出现了动脉球,
这些特点表明该鱼虽然还具有一些原始构造,但其演化地位已经
比较接近真骨鱼类。生活于北美的静水和缓流中,食物以鱼为主。
• e弓鳍鱼(Amia ciava) f雀鳝鱼(Lepidosteus Platystomus)
• (4)雀鳝目(Lepidosteiformes):体被不含闪
光质的菱形硬鳞。无喉板及喷水孔。鳔有呼吸作
用。现生种类仅雀鳝科(Lepidosteidae),代表动
物为扁口雀鳝鱼(Lepidosteus Platystomus),体
大,长达2m左右,两颌前伸成长吻,齿尖利,
是北美和中美洲淡水湖中的肉食性鱼类。
2.鲱形总目(Clupeomorpha)
• 腹鳍腹位,鳍条一般不少于6枚;胸鳍基部位置低,接
近腹缘;鳍无棘。圆鳞。
• (2)鲱形目(Clupeiformes):鳍无棘,背鳍单个。
体被圆鳞,无侧线。本目中有许多种类是很有经济价值
的海鱼,在世界渔业中占有重要的地位。鲥鱼
(Maccrura reevesi)
• 鲑形目(Salmoniformes):体形和特征与鲱形目相似,
但背鳍后方常具一脂鳍(adiposfin);有侧线。大多栖
居于北极和高纬度水域内,我国黑龙江、内蒙古、新疆
和西藏境内均有分布。本目包括在世界渔业中占重要地
位的鳟鲑鱼类。大麻哈鱼(Oncorhynchus beta) ,哲
罗鱼(Huchotaimen) ,大银鱼(Protosalanx
hyalocranius)。
3.鳗鲡总目(Anguillomorpha)
• 体呈鳗形。腹鳍腹位或缺失;背鳍及臀鳍的基底长,与
尾鳍相连。个体发育有变态。是栖息于热带和亚热带水
域中的海洋鱼类,只有少数种类(鳗鲡Anopuillo和头齿
鳗Pisodonorhis)可进入淡水中。成鱼在沿岸浅海内生活,
肉食性,生殖时到深海产卵。仔鳗变态后游向近岸。共
3目,我国产鳗鲡目(Anguilliformes)9科。
• 鳗鲡(Anguilla japonica) 海鳗( Muraenesox cinereus)
4 鲤形总目(Cyprinomorpha)
• 是比较低等的硬骨鱼类。腹鳍腹位,有些种类(鲇形目)有脂鳍;
鳔有管与食道相通;具韦伯氏器。广布于世界各州,大部分种类
生活于淡水,但以热带和亚热带水域中最多,可生活在高山、平
原、江河、山溪、激流中。包括许多重要的经济鱼类和养殖鱼种,
全世界约有5 000种,分鲤形目和鲇形目 2个目。
•
(1)鲤形目(Cypriniformes):体被圆鳞或裸露。许多种类口内
无齿,但下咽骨有发达的咽齿。全世界有3000余种,分布几乎遍
及各地。我国产6科。
• (2)鲇形目( Siluriformes):有韦伯氏器;口大齿利,口须 l-
4对;咽骨有细齿;体表裸露或局部被骨板;通常有脂鳍,胸鳍
和背鳍常有一强大的鳍棘。鲇形目中包括许多肉食性经济鱼类,
我国产10科。
• 鲤科(CyPrinidae)是鱼类中种类最多的一个科,约有2 000余种。
本科鱼类是我国淡水天然捕捞以及池塘和大水面养殖的主要对象,
有经济价值的不下400种,产量约占全国渔产总量的1/3。重要
的鲤科种类有:
• 草鱼( Ctenopharyngodon idellus),吻较宽钝,无口须;咽齿2
行,栉状。历来是我国的淡水养殖鱼种。
• 青鱼( Mylopharyngodon piceus)外形很像草鱼,但吻较尖,咽
齿一行,臼齿状;主要分布在长江以南的平原地区。
• 鲢(Hypophthal michthys molitris),头长约占全长的 1/4;眼下
侧位,鳃耙细密,呈海绵状;体银白色,故称白鲢;
• 鳙(Hyporhthalmichthxs nobilis),外形酷似鲢鱼,但因背侧的体
色较暗,并有不规则黑点而俗称花鲢;头长约为全长的 1/3,
故又名胖头鱼;主要分布在长江和珠江流域,与草鱼、青鱼、鲢
鱼合称为我国的“四大家鱼”。
• 鲤(Cyprinus),种类甚多,我国至少有20种,但以鲤鱼
(Cyprinus carpio)为最常见,其他种类大多局限于云南省境内。
鲤鱼是我国最早进行饲养的鱼种,现已成为世界性的重要养殖对
象。
• 鲫鱼( Carassins auratu),鳊鱼(Parabramis Pekinensis),体
侧扁,呈菱形,团头纺(Meglobrama amblycephala),主要分
布于长江中游地区,有‘武昌鱼”之称。
5.银汉鱼总目(Atherinomorpha)体被
圆鳞;腹鳍腹位,鳍条5-9枚,背鳍与臀
鳍对生。
• (1) 形目(Cyprinodontiformes):鳍无棘,背鳍一个,位于臀鳍
上方,无侧线,为热带及亚热带淡水中的小型鱼类。我国产2种,即青
科(Oryziatidae)的青(Oryzias latipes)和食蚊鱼科(Poeciliidae)的食
蚊鱼(Cambusia affinis)。
• (2)颌针鱼目(Beloniformes):鳍无棘,背鳍一个;测线低位,与腹
缘平行。大多是海洋鱼类,我国有飞鱼科(Exocoetida)和颌针鱼科
(Belonidae)。
• 飞鱼科胸鳍高位,长大如翼,背鳍与臀鳍同形相对,尾鳍深叉而下叶较
长;大多是热带和亚热带的浅海上层鱼类;喜集群,具趋光性,擅滑翔,
可伸展胸鳍在海面停留十余秒钟,最长滑行距离达百米以上,常见而有
经济价值的是尖头燕鳐鱼( Cypselurus oxycerhalus)。
• 颌针鱼科的两颌均延长如针状,外侧一列颌齿为稀疏的犬牙,我国沿海
常见的是扁颌针鱼(Ablennes anastomella),活泼擅泳,为近海上层肉
食性鱼类,肉味美。
燕鳐鱼( Cypselurus oxycerhalus)
颌针鱼(Ablennes anastomella)
6.鲑鲈总目(Parapercomorpha)
• 体被圆鳞或皮肤裸露;许多种类颐部有一小须;腹鳍胸位或喉位,
背鳍l- 3个,臀鳍l- 2个。
• 我国仅鳕形目
• 江鳕( Lota lota)有背鳍2个,臀鳍1个,最大个体可长达1m而重
25kg,日间隐伏在石缝洞穴中,夜间外出捕食鱼类,分布于黑龙
江水系及新疆额尔齐斯河。
• 大头鳕(Gadus macrocephalus)有背鳍3个,臀鳍2个,是黄渤海
经济鱼类之一。
7 鲈形总目(Percomorpha)
• 胸鳍大多为胸位或喉位,鳍通常有鳍棘;体常被栉鳞,但也偶有
骨板或皮肤裸露的情况。本总目绝大多数为海鱼,种类极其繁多,
共10目,常见有:
• (1)刺鱼目(Gasterosteiformes):吻大多呈管状,许多种类体被
骨板,背鳍l-2个,有时第一背鳍为游离的棘组成。我国产7科。
常见种有:
• 刺鱼科(Gasterosteidae)中华多刺鱼(Pungitis sinensis),体侧
扁,尾柄窄细;口大,两颌有锐齿;雄鱼具有善斗、以肾脏分泌
物筑巢及卫护鱼卵的习性,是北方淡水中的冷水性小鱼。
• 海龙科(Syngnatnidae),体长形,全身被环形骨板;吻呈长管
状,鳃孔小;无腹鳍,尾鳍小或消失;雄鱼常由尾部下方或腹部
皮肤形成育儿囊,容纳雌鱼产卵及受精卵发育,稚鱼孵化后,囊
即张开,稚鱼游出体外;本科包括海龙(Syngnathus)和海马
(Htopocampus)等近海小型鱼类,常栖于海藻丛中,吸食浮游
甲壳动物;于制品为传统中药材。
•
(2)鲻(zi)形目(Mugiliformes):背鳍2个,前后分离,第
一背鳍由鳍棘组成;腹鳍腹位或亚胸位。常见种为鲻鱼
(Mugilcephalus),为近海中下层鱼类,以下颌刮食泥底表面的
藻类和有机碎屑,是我国港养的主要对象。梭鱼(Liza soiuy),
脂眼睑不发达,眼稍带红色,4-6月为繁殖期。
•
(3)合鳃目(Synbranchiformes):体形似鳗,无胸鳍和腹鳍,
奇鳍彼此相连,无鳍棘。左、右鳃孔位于头的腹面合而为一,鳃
小而不发达,主要由咽腔内壁表皮代营呼吸作用。无鳔。黄鳝
(Monopterus albus),体光滑无鳞,全身黄褐色,满布不规则的
黑色斑点。栖于泥质上穴或堤岸的裂隙内。具有性逆转的特性。
除青藏高原外,分布遍及全国,是江南地区常见的食用鱼类。
•
(4)鲈形目(Perciformes):腹鳍胸位或喉位,鳍条l-5枚;
背鳍2个,分别由鳍棘和鳍条组成,无脂鳍;体大多被栉鳞。鳔
无鳔管。本目为鱼纲中种类最多的一个目,其中包括许多重要的
海产经济鱼种。我国产85科。
• 尤以鲈鱼( Lateolabrax japonicus)、鳜(gui)鱼( Siniperca
chuatsi)和石斑鱼等优质经济鱼类最负盛名。
• (5) 形目(ScorPaeniformes):第二眶下骨后延成一骨突,
并与前鳃盖骨连接。头部粗壮,常具棘棱或骨板。胸鳍基底比较
宽大。本目为一类广泛分布于热带、温带及寒带沿岸水域的海鱼
有些种类也进入河川、湖泊等淡水中。我国产11科,常见的有杜
父鱼科( Cottidae)的松江鲈鱼(Trichidermis fasciatas)、 科
(Scorpaenidae)的毒 等
• (6)鲽形目(Pleuronectiformes):即通常所说的比目鱼类。
体形侧扁,成鱼的眼、鼻、口、齿和偶鳍等均不对称,两侧的体
色也各不相同,无眼侧通常颜色浅淡。背鳍及臀鳍的基底长,腹
鳍胸位或喉位。肛门位置前移至胸鳍的后下方,且不在腹面正中。
无鳔。幼鱼身体侧扁而左右对称,泳姿正常;变态后随同头骨的
变化,一眼移位至另一侧,以无限侧平卧水底营底栖生活,比目
鱼即因此得名。本目是重要的海洋经济鱼类,我国产7科,种类
较多,资源十分丰富。
• (7)
形目(Tetrodontiformes):体短粗,皮肤裸露或被有
刺、骨板、粒鳞等。上颌骨常与前颌骨愈合,齿锥形或门齿状,
或愈合为喙状齿板。鳃孔小。腹鳍胸位或连同腰带骨一起消失。
有些种类具气囊,能使胸腹部充气和膨胀,用以自卫或漂浮水面。
大多为海洋鱼类,喜栖浅海海底或沿岸岩礁海区,只有少数种类
有定居淡水或在一定季节进入江河的习性。我国产11科,
鲻鱼(Mugilcephalus)
黄鳝(Monopterus albus)
鲈鱼( Lateolabrax japonicus)、鳜(gui)鱼( Siniperca chuatsi)
大黄鱼(Pseudosciaena crocea)
鱼(Echeneis naucrates)
小黄鱼(Pseudosciaena polyactis)
真鲷(Chrrsophrys major),又名“加级鱼”
带鱼(Trichiurus haumela)
鲐鱼( Pnematophorus japonicus)
银鲳(Stromateoldes argenteus )
金枪鱼 (Thunnus)
松江鲈鱼(Trichidermis fasciatas)
高眼鲽(Cleisthenes herzensteini)
条鳎(Zebria bra)
马面
(Navodon septentrionalis)虫斑东方 ( Fugu ocellatus)
角箱 (O. corntus)
翻车鱼(Mola mola)
8.蟾鱼总目(BatrachoidomorPha)
• 体短粗,平扁或侧扁,皮肤裸出,有小刺或小骨板。鳃
孔小,位于胸鳍外侧的腹面。腹鳍胸位或喉位。本总目
均为底栖的肉食性鱼类,以
目Lophiiformes)较常
见。