Trophic network
advertisement

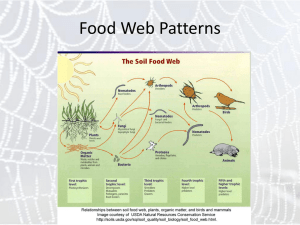
Trophic networks Food web Food chain Parasites Large herbivores Hyperparasitoids Parasitoids Large carnivores Large omnivores Larger carnivores Small carnivores Small herbivores Plant producers Parasites Larger decomposer Medium omnivores Bacteriophages Large predators Small predators Smaller decomposer Bacterial producers Typical terrestrial food with six trophic levels and 15 functional groups. Each functional group (guild) may contain many species. Herbivores Plant producers Web size The total number of elements S (species) in the web Connectance The proportion of realized links L in the web Linkage density 𝐿 𝐿 ≅ 2 𝑆(𝑆 − 1)/2 𝑆 𝐿 The proportion of realized connections per species 𝐷 = ≅ 𝐶𝑆 𝑆 𝐶= Food chain length The average length of single food chains Proportions of top, intermediate, and basal species. The proportion of omnivores. Omnivores are species that feed on more than one basic source of food (more than one trophic level) Loop, Web size: 5 intraspecific Links: 4 Predator Predator feeding Connectance: 4/(5*4/2) = 0.4 Linkage density: 4/5 = 0.8 Food chain length: 2 Omnivores: 0 Herbivore Herbivore Basal species: 1 Intermdeiate species: 2 Top species: 2 Plant Loops do not count Pitcher plant food (Nepenthes albomarginata) web Photo Nepenthes 1 2 4 5 6 12 Loop, Cannibalism 3 7 8 13 14 15 17 18 19 S = 19 Lmax = 19x18/2=171 L = 35 C = 35 / 171 = 0.2 Ch = 100 Li = 40 ChL = 100 / 40 = 2.5 L / S = 35 / 19 = 1.8 9 10 11 16 Trophical cascades Energy Abundance Biomass Large predators 2 2 Small predators 80 15 1 Herbivores 100 100 10 20 120 100 Plants 0.1 Trophical cascades vary from habitat to habitat. They are habitat specific In terrestrial habitats about 10% of energy is passed from each level to the next higher level (rule of Lawton). In marine food webs the biomass and abundance pyramids are sometime inverted. The paradox of the plankton Why is the world green? With predators of herbivores Venezuela Terborgh et al. 2006, J. Ecol. Without predators of herbivores Comparing the defoliation by herbivores on small (without predators), intermediate (some arthopod predators ) and large islands/mainlands (all types of predators) Terborgh et al. (2006) corroborated the hypothesis of Hairston, Smith, and Slobodkin (HSS) that herbivore predators control defoliation and keep the world green. Plant defense did not play a major role Mortality Recruitment Predaceous birds and snakes Parasitoid spider wasps Songbirds Predaceous insects Scorpions Lizards Spiders Rodensts Herbivorous insects Scavenging insects Detritovorous insects Seabird ectoparasites Fish and bird carcasses Seabirds Marince planctonic food web Land plands, seed detritus Seabird guano Algal detritus Polis 1998, Nature395: 744-745 Marine macroalgae An example how complex food webs might be. Each trophic level may contain several up to several hundreds of species. Islands in the Gulf of California. Terrestrial arthropod dominated food chains are often shorter than marine food chains Schoenly et al. 1991, Am. Nat 137: 597-638 Terrestrial food chains have rarely more than five levels. Torymus auratus Platygaster spec. Food chain involing insect parasitoids have often more than five levels. galls Mikiola fagi Do terrestrial and marine food webs differ in structure ? Schoenly et al. 1991, Am. Nat 137: 597-638 Haven, 1997, Oikos 78: 75-80 Terrestrial webs Marine webs Predator numbers increase linearly with the number of asvailable prey species The total richness of predators is often higher than the number of prey species Schoenly et al. 1991, Am. Nat 137: 597-638 Numbers of food chain in a web increase to the power of species richness. The upper boundary marks the limit of stability. Parasitoid – aphid relationship on oaks Lysiphlebus thelaxis Thelaxes dryophila Quercus robur Trioxys betulae Callaphidius elegans Mamamelistes betulinus Betulaphis brevipilosa Protaphidius wissmannii Trioxys pallidus Aphelinus chaonia Symydobius oblongus Stomaphis quercus Myzocallis castanicola Tuberculoides annulatus Praon flavinode Trioxys compressicornis Betula pendula Aphidius aquilus Trioxys tenuicaudus Aphidencyrtus aphidivorus Euceraphis betulae Calaphis betulicola Trioxys curvicaudus Aphidencyrtus quercicola Betulaphis quadrituberculata Rajmanek and Stary 1979, Nature 280: 311-313 Food web connection and stability The May equation predicts low linkage density at higher connection rate 𝐷 𝑆𝐶 < 1 Rajmanek and Stary 1979, Nature 280: 311-313 D: Linkage density S: species number C: connectivity The May equation predicts an upper limit of connectance for a stable food web. Schoenly et al. 1991, Am. Nat 137: 597-638 Food web complexity is limited by species richness 𝐷 𝑆𝐶 < 1 Schoenly et al. 1991, Am. Nat 137: 597-638 SC: measure of food web complexity Aquatic food webs Mechanisms that stabilize food webs: The May eqaution is based on simplified random food webs with density dependent regulation. • • • • Weak and variable links Low connectance Dietary switches Omnivory The temporal stability of food webs Temporal variability among species Omnivory stabilizes food webs High proportion of specialist species Intermediate proportion of specialist species High proportion of omnivorous species Undisturbed Disturbed Fagan 1997, Am. Nat. 150: 554-567 Mount Saint Helens blowdown zone Mount St. Helen’s recovery is a natural experiment on succession and community ecology. Food chain length and habitat properties Fresh water food chain length of North American lakes increase with lake size but not with productivity Post et al. 2000, Nature 405: 1047-1049 Compilation of well resolved food chains Hall and Raffaelli 1991, J. Anim. Ecol. 60: 823-841. Average food chain length asymptotically reaches a plateau independent of species richness. Food web complexity and ecosystem variability in ponds Linkage density of fresh water insect dominated small pond food webs increased with • Species richness • Habitat duration and decreases with • Pond environmental variability Schneider 1997, Oecologia 110: 567-575 Connectance was lowest at average species richness, variability, and pond duration. Empirical interaction matrices Pollination networks Plants Kratochwil et al. 2009, Apidologia 40: 634-650 Bees Plant Asclepias AsclepiasAspidonepsis Miraglossum Miraglossum PachycarpusSisyranthusXysmalobium Xysmalobium Pollinators cucullata woodii diploglossa verticillare pilosum natalensistrichostomus gerrardii involucratum Hemipepsis 0 0 0 18 9 20 2 41 1 Pompilidae sp. 2 0 0 0 0 0 0 0 1 0 Tiphia 0 1 0 0 0 0 0 0 0 Arge 0 0 0 0 0 0 1 0 0 Apis 0 0 1 0 0 0 1 3 0 Halictidae sp. 1 0 0 2 0 0 0 0 0 0 Halictidae sp. 2 1 0 0 0 0 0 0 0 0 Other wasps 0 1 1 0 0 0 0 1 3 Other bees 0 0 0 0 0 0 0 1 1 Other solitary bees 0 1 2 0 0 9 0 0 0 Atrichelaphinis 0 15 0 1 0 0 35 15 6 Cyrtothyrea 0 8 0 1 0 0 42 6 0 Lycidae sp. 0 0 0 0 0 0 0 2 0 Cantharidae sp. 0 0 0 0 0 0 0 2 0 Elateridae sp. 0 0 0 0 0 0 0 0 4 Chrysomelidae sp. 1 0 0 0 0 0 1 0 0 1 Chrysomelidae sp. 2 0 0 0 0 0 0 1 1 1 Scarabaeinae sp. 1 0 0 0 0 0 0 0 3 0 Scarabaeinae sp. 2 0 0 0 0 0 0 0 3 1 Scarabaeinae sp. 3 0 0 0 0 0 0 0 1 0 Curculionidae sp. 1 0 0 0 0 0 0 10 4 1 Curculionidae sp. 2 0 2 0 0 0 0 0 0 0 Coleoptera sp. 3 0 0 0 0 0 0 0 2 0 Coleoptera sp. 8 0 0 0 0 0 0 1 0 0 Other Coleoptera 0 0 0 0 0 0 0 4 4 Aspilocoryphus 1 0 0 1 0 4 1 139 1 Lygaeidae sp. 2 0 0 0 1 0 1 0 8 2 Coreidae sp. 0 0 0 0 0 0 0 1 0 Spilostethus 0 0 0 0 0 1 0 0 0 Homoecerus 0 0 0 0 0 1 0 0 0 Pentatomoidea sp. 0 0 0 0 0 0 0 1 0 Other Heteroptera 0 0 0 0 0 0 0 1 0 Calliphoridae genus 1 0 0 0 0 0 0 0 1 0 Calliphoridae genus 2 0 0 0 0 0 0 2 6 0 Calliphoridae genus 3 0 0 0 0 0 0 0 1 0 Sarcophaga sp. 0 1 0 6 0 11 0 53 1 Musca 0 0 2 0 0 0 0 3 0 Muscidae genus 2 0 0 1 0 0 0 0 0 0 Empididae sp. 1 2 0 0 0 0 1 0 0 0 Empididae sp. 2 0 0 0 0 0 0 0 1 0 Chloropidae 0 0 1 0 0 0 0 1 0 Microphthalma 0 0 0 0 0 1 0 0 0 Microphthalma 0 0 0 0 0 0 0 1 0 Tachinidae subfamily Goniinae 0 0 0 0 0 0 0 1 0 Tachinidae genus 2 0 0 0 0 0 0 0 1 0 Actea 0 0 0 0 0 0 0 1 0 Sepsidae sp. 1 0 0 0 0 0 0 0 3 1 Sepsidae sp. 2 0 0 0 0 0 0 0 0 1 Sepsidae sp. 3 0 0 0 1 0 0 0 0 0 Dacus 0 0 0 0 0 1 0 0 0 Bibionidae 0 0 0 0 0 0 0 1 0 Diptera sp. 3 0 0 0 0 0 0 1 0 0 Diptera sp. 22 0 0 0 0 0 1 0 0 0 Other Diptera 0 1 0 1 0 1 0 15 0 Unidentified butterfly 0 0 0 0 0 0 1 0 0 Unidentified micromoth 2 0 0 0 0 0 0 0 0 From Ollerton et al. 2003, Ann. Botany 92: 807-834 The matrix approach to mutualistic and food webs What are mutualistic webs: Plant – pollinator webs Plant seed disperser webs Plant herbivore webs Predator prey webs Host parasite webs Competition webs Generalists Plants • • • • • • Pollinators • Generalist pollinator visit most plant species • Specialist pollinator visit the most popular plant species • Mutualistic networks contain forbidden links Nestedness is defined as the ordered loss of links in a mutualistic matrix where rows and coloumns are sorted according to species richness. Specialists Generalists Specialists Unexpected link Linkages: number of filled cells in the matrix Linkage density: L/S1 Connectance: Matrix fill, L/(S1S2) The architecture of mutualistic networks Seed disperser Pollination networks Foods webs Jordi Bascompte 1967- Bascompte 2003, PNAS 100: 9383-9387 Weak Anthropic Principle (Carter 1973): We must be prepared to take account of the fact that our location in the universe is necessarily privileged to the extent of being compatible with our existence as observers. In ecology this means: Ecological systems do not have a random strucure. They have that non-random structure that enabled them to survive during evolution. Bastolla et al. 2009, Nature 458: 1018-1021 • Mutualistc networks are often nested. • The nested architecture promotes diversity and stability Nestedness as an emergent property of ecological systems Food webs are often compartmented A B C D E F G 1 1 1 1 1 0 0 0 2 1 1 1 1 0 0 0 3 1 1 1 1 0 0 0 4 1 1 1 1 0 0 0 5 0 0 0 0 1 1 1 6 0 0 0 0 1 1 1 7 0 0 0 0 1 1 1 8 0 0 0 0 1 1 1 S 4 4 4 4 4 4 4 4 4 4 4 3 3 3 3 28 Foods webs have a modular structure. Modularity tends to stabilize food webs. Modules itself have a nested structure. Nestedness tends to stabilize mutualistic networks. Mutualistic webs (comparirson of two trophic levels) are most often nested. A B C D E F G 1 1 1 1 1 1 1 1 2 1 0 1 1 1 0 0 3 1 1 1 1 0 0 0 4 1 1 1 1 0 0 0 5 1 1 1 0 0 0 0 6 1 1 0 0 1 0 0 7 1 0 0 0 0 0 0 8 1 0 0 0 0 0 0 S 8 5 5 4 3 1 1 S 7 4 4 4 3 3 1 1 27 Stability, resilience and tipping points Instable equilibrium Local stable Global stable state state Tipping point Low stability Local stability Global instanility Low resistence Ecologial systems (particularly networks) can be in various states: • Instable equilibria are at tipping points and can move towards different directions. • Local stable equilibria can easily be forced to achieve other stable states. • Global equilibria need much energy to leave their state. • Inequilibria can easily move between different states. • Resilience refers to the speed of a systen to return to a stable state. • Resistence is the ability of a system to avoid displacement. • Robustness is the ability of a system to exist witin a wide range of conditions. • Stability refers to the amplitude of variability • Sustainability i the capacity to endure Multiple states Probability Instable equilibrium Local stable Global stable state state A state reaches its tipping point Food webs and tipping points State Tipping points define states where a system irreversably changes the probability distribution of states. Indicators of critical tipping points: • Resilience slows down • Dominant eigenvectors of the food web matrices shorten • Increased variance • Variance / mean relationships increased • Multimodality of states • Increasing connectivity and decreasing diversity Robustness The importance of wild bees for pollination stability Meta-analysis of empirical food ebs Dunne 2002, Ecol. Lett 5: 558 Garibaldi et al. 2013. Science 339: 1608 In empirical foods webs robustness increases with connectance. In random foods webs robustness decreases with connectance. Therefore, empirical food webs have a special non-random structure that promotes stability. Wild insects increase fruit production more effectively than honey bees alone. Species richness increases ecological functioning