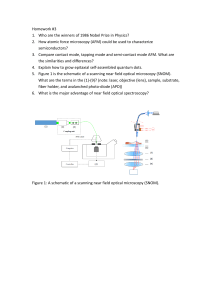
E BOOK Atomic Force Microscopy for Nanoscale Biophysics From Single Molecules to Living Cells 1st Edition by Mi Li
advertisement

Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Contents ix xi 1. Fundamentals and methods of atomic force microscopy for biophysics 1.1 Background of atomic force microscopy for biophysics 1.2 Atomic force microscopy topographical imaging modes 1.3 Atomic force microscopy force spectroscopy techniques 1.4 High-speed atomic force microscopy 1.5 Topography and recognition imaging mode atomic force microcopy References 2. Imaging and force detection of single deoxyribonucleic acid molecules by atomic force microscopy 2.1 Background 2.2 Sample preparation methods 2.3 Topographical imaging of single DNA molecules and events 2.4 Time-lapse imaging of individual DNA molecular dynamics 2.5 Extracting the persistence length of DNA molecules from atomic force microscopy images 2.6 Mechanically unzipping single DNA molecules by atomic force microscopy force spectroscopy 2.7 Probing individual DNA behaviors on DNA origami nanostructures 2.8 Summary References 3. High-resolution imaging and force spectroscopy of single membrane proteins by atomic force microscopy 3.1 Background 3.2 Topographical imaging of single native membrane proteins 3.3 Unfolding mechanics of individual native membrane proteins 3.4 Observing the dynamics of single membrane proteins by high-speed atomic force microscopy 3.5 Multiparametric atomic force microscopy imaging of single membrane proteins 1 1 5 19 31 33 35 43 43 44 47 52 55 58 61 66 68 75 75 77 84 88 91 Get all Chapters For Ebook vInstant Download by email at etutorsource@gmail.com We Don’t reply in this website, you need to contact by email for all chapters Instant download. Just send email and get all chapters download. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com You can also order by WhatsApp https://api.whatsapp.com/send/?phone=%2B447507735190&text&type=ph one_number&app_absent=0 Send email or WhatsApp with complete Book title, Edition Number and Author Name. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com vi Contents 3.6 Topography and recognition imaging of single membrane proteins 3.7 Summary References 4. Characterizing the nanostructures and mechanical properties of hydrogels by atomic force microscopy 94 97 99 105 4.1 Background 4.2 Nanostructures and nanomechanics of natural plant hydrogels 4.3 Characterizations of biopolymeric hydrogels inspired by carnivorous plant mucilage 4.4 Imaging and mechanical analysis of peptide-assembled nanofibrillar hydrogel 4.5 Probing the mechanical cues in cellhydrogel interactions 4.6 Summary References 105 108 116 122 127 130 131 5. Detecting the behaviors of single viruses by atomic force microscopy 135 5.1 Background 5.2 Imaging the fine structures of single viruses 5.3 Nanoindentation for mechanical measurements and manipulations of single viruses 5.4 Single-virus force spectroscopy for probing viral binding affinity 5.5 Multiparametric atomic force microscopy imaging of viruscell interactions 5.6 Visualizing individual viral dynamics by high-speed atomic force microscopy 5.7 Summary References 143 147 150 153 155 156 6. Imaging and mechanical analysis of single native exosomes by atomic force microscopy 161 6.1 Background 6.2 Exosome isolation and immobilization 6.3 Imaging single native exosomes in liquid 6.4 Measuring the mechanics of single native exosomes 6.5 Multiparametric imaging of single native exosomes 6.6 Single-molecule force spectroscopy on single exosomes 6.7 Summary References 161 164 168 172 174 177 179 182 7. Nanoscale imaging and force probing of single microbial cells by atomic force microscopy 187 7.1 Background 7.2 Immobilization methods of living microbial cells for atomic force microscopy imaging 135 139 187 191 Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Contents vii 7.3 Visualizing the nanostructures and their dynamics of living microbial cells by atomic force microscopy 7.4 Measuring the mechanical properties of single living microbial cells by atomic force microscopy 7.5 Single-molecule force spectroscopy and single-cell force spectroscopy of microbial adhesion 7.6 Multiparametric atomic force microscopy imaging of single living microbial cells 7.7 Atomic force microscopy cantilever as a nanomechanical sensor for monitoring microbial activities 7.8 Summary References 210 212 213 8. Investigating the structures and mechanics of single animal cells by atomic force microscopy 219 8.1 Background 8.2 Imaging the surface structures and their dynamics of single living adherent animal cells 8.3 Measuring the mechanical properties of single living adherent animal cells 8.4 Probing the molecular activities on the surface of single adherent cells 8.5 Visualizing the surface structures and their dynamics of single living suspended animal cells 8.6 Detecting the mechanical cues involved in the activities of lymphoma cells 8.7 Probing the molecular activities on the surface of primary lymphoma cells 8.8 Summary References 9. Characterizing the extracellular matrix for regulating cell behaviors by atomic force microscopy 9.1 Background 9.2 Detecting the mechanical properties of decellularized extracellular matrix 9.3 Investigating the structures and mechanics of basement membranes 9.4 In situ imaging of cell culture medium-forming nanogranular surface for cell growth 9.5 Hierarchical micro-/nanotopography of extracellular matrix for tuning cellular structures and mechanics 9.6 Summary References 193 200 203 207 219 222 229 241 246 248 258 261 262 269 269 272 275 279 281 284 285 10. Combining atomic force microscopy with complementary techniques for biophysics 289 10.1 Background 10.2 Scanning near-field ultrasound holography 10.3 Fluidic force microscopy 289 291 293 Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com viii Contents 10.4 Combining atomic force microscopy with micropipette 10.5 Combining atomic force microscopy with fluidic environment 10.6 Summary References 297 301 305 306 11. Future perspectives of atomic force microscopy for biophysics 309 References 313 Index 315 Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com C H A P T E R 1 Fundamentals and methods of atomic force microscopy for biophysics 1.1 Background of atomic force microscopy for biophysics Single-cell analysis is an emerging and promising area expected to remarkably benefit unveiling the underlying mechanisms guiding life activities. A cell is the fundamental structural and functional unit of living organisms. The study of the structure, function, and behavior of cells is critical for seeking the answers to the questions of what life is and how it works [1], which has been highlighted long ago by the pioneering cell biologist E. B. Wilson in his famous textbook The Cell in Development and Heredity: “The key to every biological problem must finally be sought in the cell, for every living organism is, or at some time has been, a cell” [2,3]. So far much of our understanding of cellular physiological and pathological activities has come from traditional ensemble-averaged assays [4], which reflect the dominant biological traits in a population and assume that an average response is representative of a typical cell within a population, whereas cell-to-cell differences are usually obscured [5,6]. Heterogeneity between individual cells is an intrinsic feature of dynamic cellular processes, including signaling, transcription, and cell fate [7]. Even for genetically identical cells that have been cultured under the same conditions, cellular heterogeneity can also be observed [8]. The basis of cellular heterogeneity is a field of active investigation, and multiple mechanisms are thought to contribute to this phenomenon, such as stochastic molecular interactions, phenotypic differences, stochastic transcriptional bursting, and so on [911]. Cellular heterogeneity represents one of the greatest challenges in tumor Get all Chapters For Ebook 1Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 2 1. Fundamentals and methods of atomic force microscopy for biophysics therapeutics [12,13]. For tumors, extensive genetic and phenotypic variations have been observed, including intertumoral heterogeneity (variations between tumors of different tissue and cell types), intratumoral heterogeneity (subclonal diversity within a tumor), and intercellular heterogeneity (cellular variations within a subclonal population of tumor cells) [14], which significantly add the complexity of tumors and have been shown to be closely associated with the therapeutic responses and drug resistance as well as disease relapse of tumors [15,16]. Single-cell analysis is therefore essential to understand the causes and consequences of cellular heterogeneity and how it changes during the physiological or pathological processes [17]. In fact, there have been significant advances in the development of single-cell biochemical analysis tools in the past years, which are able to detect diverse components of individual cells, including DNAs, RNAs, proteins and metabolites [1820]. The practical applications of single-cell biochemical analysis methods have significantly promoted our understanding of tumor heterogeneity, including the identification of metastasis-initiating cells [21], single-cell pathology landscape of breast cancer [22], revealing heterogeneity of intratumoral T cell states for cancer immunotherapy [23], discovering novel cell types of tumor microenvironment [24], and so on, showing the great potentials of single-cell analysis in addressing biological issues from the perspective of individual cells. It is increasingly evident in the communities of life sciences that mechanical cues play a fundamental role in regulating cellular activities. Cell life is not only a chemical process but also a mechanical one [25]. Cells generate, transmit and respond to forces through an intricate network of mechanical components, resulting in cell movement and shape change, as well as altered signaling, modulated expression and even genomic damage [26]. The mechanical interactions between the cell and extracellular matrix (ECM) often yield unique biochemical and mechanical properties of the cell [27]. For example, studies have shown that substrates with a stiffness similar to that of the brain (0.11 kPa), muscle (817 kPa), or bone tissues (2540 kPa) guide the stem cells to differentiate into neurons, myoblasts, and osteoblasts, respectively [28]. Particularly, changes in the biomechanical and biophysical properties of cells and subcellular structures influence, and are influenced by, the onset and progression of human diseases such as cancer [29]. Numerous studies have shown that cancerous cells are softer and more deformable than healthy cells due to a disorganized and less filamentous cytoskeletal network, which helps the malignant cells to squeeze through the narrow constrictions of the ECM and the tight endothelial cellcell junctions [3032]. In malaria disease, researchers have shown that progression through the parasite development stages (ring stage, trophozoite stage, schizont stage) leads to a considerable stiffening and decreased deformability of the host red blood Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com We Don’t reply in this website, you need to contact by email for all chapters Instant download. Just send email and get all chapters download. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com You can also order by WhatsApp https://api.whatsapp.com/send/?phone=%2B447507735190&text&type=ph one_number&app_absent=0 Send email or WhatsApp with complete Book title, Edition Number and Author Name. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.1 Background of atomic force microscopy for biophysics 3 cells compared to healthy ones [3335]. The knowledge of cell mechanics, combined with previously known biochemical cues, has greatly advanced our understanding of related diseases [36] and has inspired new therapeutic approaches for practical applications. In recent years researchers have been exploring targeting the mechanical cues involved in diverse human diseases (cancer, fibrosis, and cardiovascular disease) as a therapeutic intervention [37] and various agents that interfere with either the mechanical properties of the tissue or signaling responsive to changes in the tissue mechanics have been developed for preclinical and clinical trials [38], offering new opportunities for treating human diseases. Biointerfaces are surfaces at which tissues, microorganisms, cells, viruses or biomolecules make contact with other natural or synthetic materials, and observing and manipulating the interactions that occur at biointerfaces is critical for understanding the behaviors of biological systems nearly at all levels [39]. We know that in vivo cells are embedded within the fibrous ECM to form tissues and then many tissues are organized together to constitute organs [40]. The ECM provides not only essential physical scaffolding for the cellular constituents but also initiates crucial biochemical and biomechanical cues that are required for tissue morphogenesis, differentiation and homeostasis [41]. The resident cells of each tissue are responsible for and responsive to the ECM in a process referred to as dynamic reciprocity: cells modify their secreted ECM products in response to various stimuli (e.g., mechanical cues, oxygen and nutrient concentration), and in turn, the ECM sends mechanical and biochemical signals to resident cells through the engagement of cell surface receptors (e.g., integrins, ion channels [42]) and the subsequent activation of intracellular signaling cascades and ultimately the changes in gene expression and cell phenotype [43]. Abnormal interactions between cells and ECM are often accompanied by pathological changes in living organisms such as tumors [44] which leverages ECM remodeling to create a microenvironment that promotes tumorigenesis and metastasis [45] (e.g., thickening and linearization of ECM fibers are common in cancers, which facilitate the migration and progression of tumor cells [46]). At the level of individual cells, cells interact with their environments via their surfaces whose functions are mediated by the structurally complex and dynamic assembly of specific carbohydrates, proteins, lipids and other macromolecules distributed on cell surfaces [47]. Cell surface molecules regulate many essential cellular processes, including cell adhesion, tissue development, cellular communication, inflammation, tumor metastasis, and microbial infection [48], and therefore probing the molecular activities and behaviors on the cell surface is of remarkable significance for understanding the underlying mechanisms guiding cellular processes. At the level of individual viruses, in order to gain access to the cell interior to deliver the viral Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 4 1. Fundamentals and methods of atomic force microscopy for biophysics genome to the cell cytoplasm, viruses attach to the host cells via the specific binding interactions between molecules on the surface of the virus and receptors (proteins, carbohydrates, or lipids) on the cell surface [49]. Consequently, investigating the structures and properties as well as functions of biointerfaces is particularly meaningful for revealing the underpinnings of life processes. The advent of atomic force microscopy (AFM) provides a revolutionary, powerful, and multifunctional tool for investigating the structures and mechanics of living biological systems at biointerfaces under aqueous conditions with high resolution. AFM uses a sharp tip mounted at the end of a soft cantilever to raster scan the surface of specimens immobilized on stiff substrates, during which a laser beam is used to detect the deflections of the cantilever to sense the interaction forces between tip atoms and specimen surface atoms and a piezoelectric actuator drives the tip move vertically to maintain the interaction forces constant [50], yielding the three-dimensional topographical images of the specimen surface. In essence, the spatial resolution of AFM is determined by the contact area between the AFM tip and sample surface, which typically contains several factors, including probe geometry, tip sharpness, sticking of sample to AFM tip, mobility of the sample (or the substructures on the surface of the sample), mobility of substrate, and so on, and historically the spatial resolution of AFM on biological samples in liquids is about 1 nm at best [51]. Recently, a new technique called localization AFM (LAFM) is developed by Heath et al. [52], which increases the spatial resolution of AFM to 0.1 nm on biomolecules in a buffer solution. AFM is able to not only visualize the native structures of living specimens in near-physiological conditions without the need for chemical treatments (e.g., fixation, staining, labeling) [53], but also can measure the mechanical properties of specimens and quantify the molecular forces at force spectroscopy mode [54,55], making AFM particularly suited for applications in life sciences. Compared with other typical high-resolution imaging and force measurement methods used in molecular and cell biology, AFM has unique advantages. The spatial resolution of conventional optical microscopy is limited to about 200300 nm due to the effect of Abbe diffraction, making it impossible to visualize cellular structures and machineries as well as their assembly dynamics in nanoscopic details [56]. The development of superresolution optical microscopy [e.g., stimulated emission microscopy (STED), photoactivated localization microscopy (PALM), stochastic optical reconstruction microscopy (STORM)] shatters this limit and achieves the spatial resolution of 20 nm [57], enabling visualization of previously invisible molecular details in biological systems [58]. However, super-resolution optical microscopy requires the fluorescent labeling of target molecules, and strictly speaking the results only reflect the behaviors of the fluorescein, not Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.2 Atomic force microscopy topographical imaging modes 5 of the target molecules. Besides, fluorescent labeling inevitably reduces the fidelity of the target molecules and may affect the physiological activities of molecules and cells [59]. Electron microscopy [including scanning electron microscopy (SEM) and transmission electron microscopy (TEM)] is able to observe single molecules with high spatial resolution, but it requires the dehydration of the samples which can cause damage to samples [60,61]. Though cryo-electron microscopy (cryo-EM) is able to resolve the structures of proteins and macromolecular complexes at near-atomic resolution (,0.4 nm) without the need for crystals [62], it only provides static snapshots of molecules and is unable to capture the dynamics of molecules. Optical tweezers and magnetic tweezers are widely used methods for single-cell and single-molecule force assays [6368], but they cannot obtain the structural information of samples while AFM is able to simultaneously perform topographical imaging and force measurements on samples. The detailed comparisons between AFM and the other techniques described above are summarized in Table 1.1. The excellent merits of AFM have attracted the tremendous attention of researchers from various disciplines (e.g., biology, chemistry, physics, and engineering) to utilize AFM to probe cellular and molecular activities taking place at biointerfaces. In the past decades, AFM has achieved great success in characterizing the structures and mechanical cues of diverse biological systems with the unprecedented spatiotemporal resolution, yielding numerous insights into how mechanical forces regulate physiological and pathological behaviors at the single-cell and single-molecule levels. Particularly, the instrumental performances and functions of AFM have been being improved continuously since its invention, which has considerably promoted the wide applications of AFM, offering novel possibilities for unveiling the underlying mechanisms guiding life processes and human diseases from the perspective of biomechanics and biophysics. 1.2 Atomic force microscopy topographical imaging modes 1.2.1 Basic principles In order to understand the principles of AFM imaging, we need to discard the notions of conventional microscopy (optical microscopy, electron microscopy), since AFM does not have any lenses. Instead, AFM uses a tip mounted on a soft cantilever to feel the sample surface, much like a blind person feels a person’s face with their fingers and then forms a mental image of the person’s face [69]. As shown in Fig. 1.1A, an AFM is commonly composed of five parts, including a soft microcantilever with a sharp tip mounted at its end (the cantilever/tip assembly is often referred to as the probe), a piezoelectric tube actuator, an optical lever system, a Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com TABLE 1.1 biology. Comparisons of representative high-resolution imaging and force measurement tools for applications in molecular and cell Techniques/ features Atomic force microscopy General descriptions The deflection of a cantilever is measured by a laser beam and a position-sensing detector to obtain force and displacement for both topographical imaging and force spectroscopy. For force spectroscopy, ligand molecules are attached to the surface of AFM tip which is then controlled to touch the receptors on the substrate (or cell surface) to form receptorligand pairs and mechanically rupture the molecular pairs. Spatial resolution 0.11 nm Super-resolution optical microscopy (STED, PALM, STORM) Electron microscopy (SEM, TEM, Cryo-EM) Optical tweezers Magnetic tweezers Applying a focused excitation beam with a donut shape to create a region of fluorescent emission much smaller than a typical focal spot of conventional optical microscopy (STED) or stochastically turning on individual molecules within the diffraction-limited volume at different time points (PALM, STORM). Using electrons to penetrate a thin specimen and then image the transmitted electrons (TEM), or focusing electrons into a small-diameter electron probe to raster scan the specimen and then collect the secondary electrons released from each local region of the specimen (SEM). CryoEM is a type of TEM capable of observing samples at cryogenic temperatures. An optical trap is created by focusing a laser to a diffractionlimited spot with a high numerical aperture (NA) microscope objective to capture the dielectric particles (Bμm sized beads, bacteria, organelles). Polystyrene or silica beads used as “handles” attached to biological samples permit precise measurements of force and displacement. A pair of permanent magnets is placed above the sample holder of an inverted microscope outfitted with a charge-coupled device (CCD) camera, applying force and rotation on magnetic beads linked with biological samples. The three-dimensional position of the bead is obtained by video tracking. 2050 nm 0.210 nm B1 nm 110 nm Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Force range 10104 pN N/A N/A 0.1100 pN 0.01100 pN Sample preparation and work environment Samples immobilized on substrates in physiological conditions (cell growth medium or buffer solution, controllable temperature and CO2). Fluorescent labeling of target molecules in physiological conditions Dehydrated specimens on grid in vacuum (TEM); dehydrated and gilding specimens in vacuum (SEM); vitrified specimens at cryogenic temperatures (cryoEM). Beads on living cells or inside cells by phagocytosis, or beads coated with molecules in physiological conditions. Magnetic beads on living cells or inside cells by phagocytosis, or beads linked with molecules in physiological conditions. Advantages Imaging living samples and observing life dynamics under native conditions without the need of staining, labeling or fixation; simple sample preparation; high signal-to-noise ratio; assessment of multiple physical, chemical and biological parameters. Access to threedimensional cellular structures; high spatiotemporal resolution; monitoring biomolecular processes in living cells. Solving atomic structures of proteins; conformational snapshots of proteins and complexes; visualizing molecular structures within the cell; imaging the surfaces of specimens at nanometer resolution. High precision control of torque and force, parallel processing of multiple beads, and capable of altering the trap position at high frequencies (kHz). Insensitive to drift and low-frequency noise (,1 Hz), inherently providing ultrastable operation, no need of complex designs, passive force clamp. Limitations Restricted to the surface of specimens and unable to detect intracellular structures. Fluorescent labeling decreases specimen fidelity and could affect life processes. Providing only static snapshots and incapable of observing life dynamics. Photodamage and sample heating. Lower resolution and no 3D trapping. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com We Don’t reply in this website, you need to contact by email for all chapters Instant download. Just send email and get all chapters download. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com You can also order by WhatsApp https://api.whatsapp.com/send/?phone=%2B447507735190&text&type=ph one_number&app_absent=0 Send email or WhatsApp with complete Book title, Edition Number and Author Name. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 8 1. Fundamentals and methods of atomic force microscopy for biophysics FIGURE 1.1 Atomic force microscopy (AFM) configuration. (A) Schematic of AFM principle. (B and C) Actual photographs of commercial AFMs. (B) A commercial Bioscope Catalyst AFM (Bruker, Santa Barbara, CA, USA) is set on an inverted optical and fluorescence microscope. The inset shows the Petri dish. Cells are grown in dishes and the AFM probe is immersed in the medium in the dish to probe the cells under the guidance of optical (fluorescence) microscopy. (C) A commercial Dimension Icon AFM (Bruker, Santa Barbara, CA, USA) which has a lateral optical microscope for visual guidance of moving AFM probe to target areas on the substrates. The inset is the optical image of the working AFM probe in liquids in the dish. Source: (B) Reprinted with permission from M. Li, L. Liu, N. Xi, Y. Wang, X. Xiao, W. Zhang, Nanoscale imaging and mechanical analysis of Fc receptormediated macrophage phagocytosis against cancer cells, Langmuir 30(6) (2014) 16091621. Copyright 2014 American Chemical Society. (C) Reprinted with permission from M. Li, N. Xi, Y. Wang, L. Liu, In situ high-resolution AFM imaging and force probing of cell culture mediumforming nanogranular surfaces for cell growth, IEEE Trans. Nanobiosci. 19(3) (2020) 385393. Copyright 2020 IEEE. signal processing module and a feedback control electronics. The piezoelectric actuator acts as an electromechanical transducer that is able to convert electrical potentials into mechanical movements or vice versa. When a potential difference is applied on the opposite faces of a piezoelectric structure, the shape of the piezoelectric structure changes, and normally the Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.2 Atomic force microscopy topographical imaging modes 9 expansion coefficient for a single piezoelectric shape is on the order of 0.1 nm/V applied [70]. The piezoelectric materials used in AFM are commonly lead zirconate titanate (PZT). The piezoelectric scanner of AFM integrates independently operated electrodes for X, Y, and Z directions into a single tube, which allows the nanopositioning of the AFM probe in threedimensional directions (XYZ) on the surface of the sample being probed. Notably in the early days of scanning probe microscopy, the shape of the piezoelectric scanner is like a tripod, which has been totally superseded by tubes of piezoelectric ceramic materials now. Piezoelectric tube has many advantages over tripod arrangement and the principal one is that larger scan ranges are possible with the compact and symmetrical geometry of the tube scanner [71]. In essence, the piezoelectric material does not respond linearly to the applied voltage, and there are two prevailing methods in current commercial AFM to address this issue: one is adding XYZ position sensors to monitor the actual movements of the piezoelectric tube and then correcting the nonlinearity through a feedback loop (usually called close loop), and the other one is firstly modeling the nonlinearity and then using nonlinear voltage to drive the tube to obtain linear movements (usually called open loop) [72]. For AFM imaging, the AFM probe is driven by the piezoelectric tube to raster scan the surface of the sample in the horizontal plane (XY directions), during which the piezoelectric tube simultaneously drives the probe to move vertically (Z direction) to maintain the constant interaction forces between tip surface atoms and sample surface atoms. The interaction forces between the AFM probe and sample are sensed by the optical lever system. A laser beam is focused onto the backside of the cantilever and is reflected via a mirror to a fourquadrant position sensitive detector (PSD). Any cantilever deflections caused by the changes of tip-sample interaction forces will result in slight alterations in the direction of the reflected beam, which is tracked by the PSD. The PSD signals are analyzed by the signal processing module and then the feedback system will control the piezoelectric tube to move vertically according to the topography of the sample to maintain the tip-sample interaction forces constant. The displacements in the XYZ directions of the piezoelectric tube are recorded during scanning, which yields a threedimensional topographical image of the specimen being scanned. A computer is linked to the controller to set up AFM imaging parameters and collect data from the controller. In practice, an optical microscopy system is commonly integrated with the commercial AFM to facilitate the manipulator to visually move the AFM probe to the specimens [73,74], as shown in Fig. 1.1B and C. AFM imaging depends on monitoring the interaction forces between tip surface atoms and sample surface atoms. It is now well-known that there are four distinct forces in nature, two of which are the strong and weak interactions that act between neutrons, protons, electrons, and Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 10 1. Fundamentals and methods of atomic force microscopy for biophysics other elementary particles and have a very short range of action (less than 1025 nm) [75]. The other two types of forces are the electromagnetic and gravitational interactions that act between atoms and molecules. For the interaction forces between tip surface atoms and sample surface atoms involved in AFM imaging, the gravitational force is completely insignificant compared to electromagnetic force (e.g., the gravitational force between two ions 1 nm apart is 32 orders of magnitude weaker than the electrostatic force) [76]. Electromagnetic forces are the source of all intermolecular interactions which determine the properties of solids, liquids, and gases, the behaviors of particles in solution, chemical reactions, and the organization of biological structures [75]. We know that electrons within an atom are in continual motion and travel extremely fast. For a given atom, it may appear electrically neutral over conventional periods of time, but for a very short period of time (saying a snapshot), the distribution of electric charges due to the electron motions is not perfectly symmetrical and this gives rise to subtle charge imbalances referred to as dipoles. Each molecule, therefore, exhibits a slightly different distribution of charge within a given snapshot and the charge imbalance in one molecule can electrically induce a similar imbalance in a neighbor molecule, causing that the slightly positive end of one molecule will be attracted to the negative end of another neighboring molecule and this is the origin of the van der Waals force [71]. For simplicity, we can consider the interactions between an atom at the tip apex and another atom on the sample surface, which can be characterized by the LennardJones potential function [77]: EðrÞ 5 4ε σ 12 σ6 2 r r (1.1) where r is the separation between the two atoms, ε is the depth of the potential well (associated with interaction strength), and σ is the interatomic distance where the potential is zero. ε and σ are two constants that depend on the materials and need to be experimentally determined to describe the specific molecular interaction. Normally, σ is approximately equal to the diameter of the atom involved (typically a few Angstroms [77]), and is sometimes called the hard sphere diameter [71]. Plotting the LennardJones function gives the interaction forces between the AFM tip atom and sample surface atom, as shown in Fig. 1.2. When the tip atom is far away from the sample atom, there is no interaction force between the two atoms (stage ①). When the tip atom approaches the sample atom gradually, the van der Waals attractive force arises which pulls the tip atom toward the surface, causing the downward deflection of the AFM cantilever (stage ②). As the tip atom further contacts and compresses the sample atom, repulsive forces Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.2 Atomic force microscopy topographical imaging modes 11 FIGURE 1.2 Schematic showing the interactions between atomic force microscopy tip atom and sample surface atom modeled by LennardJones function. due to Pauli exclusion principle and electrostatic (Coulombic) interactions dominate the interaction forces between tip atom and sample atom, causing the upward deflection of the AFM cantilever (stage ③). Depending on the types of interaction forces (attractive force or repulsive force) between the AFM tip and sample sensed by the AFM cantilever, different AFM imaging modes are obtained (as denoted in Fig. 1.2), which will be described in the following paragraphs. It should be noted that the interaction forces between the AFM tip and sample surface are highly complicated and many other types of forces are involved in different imaging environments, for example, meniscus force arises from capillary condensation around the contact sites between the AFM tip and sample surface for AFM imaging under ambient conditions, whereas double-layer and hydrodynamic force emerge when imaging in liquids [78]. Readers are referred to the references [76,78] for more descriptions of the AFM tip-sample interaction forces. 1.2.2 Contact mode Contact mode is the first imaging mode developed for AFM, and it is the basis for AFM techniques. In the contact mode, the AFM probe tip is in continuous physical contact with the sample surface and the repulsive forces between the probe tip and sample surface are sensed by the AFM probe cantilever (Fig. 1.2). For contact mode AFM imaging, the piezoelectric tube drives the probe to raster scan the sample in the horizontal plane, during which the probe is dragged across the sample surface (Fig. 1.3A) and the topographical changes of the sample surface in the vertical direction will result in the alterations of the deflection of Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 12 1. Fundamentals and methods of atomic force microscopy for biophysics FIGURE 1.3 Typical types of atomic force microscopy (AFM) imaging modes. (A) Contact mode and tapping mode. (I) Schematic diagram of AFM imaging of biological specimens (an example of a cell membrane is shown) attached on the substrate. (II) Contact mode AFM imaging. In contact mode, AFM tip is scanned over the specimen surface, while the deflection of the cantilever is maintained constant. (III) Tapping mode AFM imaging. In tapping mode, commonly the amplitude of the oscillating cantilever is maintained constant. (B) PFT mode. (I) Schematic diagram of AFM imaging at PFT mode. The vibrating tip indents the specimen in the vertical direction (z) at each sampling point to record force curves during raster scanning (the movement trajectory of the AFM tip is denoted by the dashed yellow line) at the horizontal plane (xy). (II) Schematic diagram of the force curve. Each force curve is composed of two portions, including an approach curve and a retract curve. By analyzing the different parts of the force curve, various mechanical parameters of the specimen are obtained, including Young’s modulus, deformation, adhesion force, and energy dissipation. Young’s modulus and adhesion forces are obtained from the retract curve, deformation is obtained from the approach curve, and the energy dissipation represents the area (orange shaded area) between the approach curve and retract curve. AFM probe. The AFM signal processing and feedback module then compare the current cantilever deflection (called the feedback parameter) with a preset threshold (called the setpoint). The difference between the current cantilever deflection and the setpoint is called the error signal. If the current cantilever deflection is not equal to the setpoint, specific voltage is then applied to control the piezoelectric tube to move up or down to make Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com We Don’t reply in this website, you need to contact by email for all chapters Instant download. Just send email and get all chapters download. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com You can also order by WhatsApp https://api.whatsapp.com/send/?phone=%2B447507735190&text&type=ph one_number&app_absent=0 Send email or WhatsApp with complete Book title, Edition Number and Author Name. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.2 Atomic force microscopy topographical imaging modes 13 the error signal back to zero. By recording the movements of the piezoelectric tube in three-dimension directions, we get the three-dimensional topographical image of the sample being scanned. The setpoint of the cantilever deflection in contact mode is preset by the user before AFM scanning and this parameter determines how hard the AFM tip pushes against the surface of the sample (called the scan force), which is important for utilizing contact mode AFM imaging to observe fragile biological samples. Notably, the contact mode AFM imaging described here operates in a way of constant force mode. Constant force mode is by far the most widely used mode, and generally any references to contact mode AFM mean constant force mode [79]. Contact mode can also work in constant height mode, in which the probe maintains a fixed height above the sample and there is no feedback loop in this mode. In constant height mode, the image signal comes entirely from the cantilever deflection and the deflection force on the cantilever is directly used to calculate the separation between the tip and sample surface. Constant height mode is appropriate for quick scans of samples with small height differences (if height differences are large, the tip will very likely crash onto the surface, causing either the tip to be destroyed or the tip to damage the sample) [80] and this mode has specific biomedical applications (e.g., observing the behaviors of biomolecules firmly attached to the smooth substrate with high-speed AFM [81,82]). A notable point for contact mode AFM is the selection of the probe. Normally, there is a contradiction between the force sensitivity and the stability of the probe. A softer cantilever (with a lower spring constant) deforms more under the same load and thus has better force detection sensitivity. However, a softer cantilever is more susceptible to the thermal drift caused by the environmental temperature variation (e.g., the heat generated by the AFM laser or optical microscope illumination often results in significant drift in deflection of the cantilever with a spring constant lower than 0.01 N/m, in the range of a few volts in the PSD output signals [72]), which conversely influences the quality of AFM imaging. In practice, cantilevers with a spring constant between 0.01 and 0.1 N/m are usually good for living cell imaging by contact mode AFM [72]. Besides, commercial AFM probes are commonly fabricated with the use of silicon or silicon nitride. In order to increase the reflectivity, a thin layer of gold is often coated on the backside of the cantilever, or a ferromagnetic coating may be applied if magnetic sensitivity is required [71]. Obviously, due to the fact that the thermal expansion coefficient of the coated material is different from the cantilever material (silicon/silico nitride), coating only one side of the cantilever will lead to a substantial thermal drift of the cantilever in response to the changes of environmental temperature. In 2012, Churnside et al. [83] have shown that even coating AFM cantilevers on both sides has thermal drift effects on the probe, whereas removing the coating layers of Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 14 1. Fundamentals and methods of atomic force microscopy for biophysics the AFM probes significantly increases the force stability and precision. As an alternative way, we can perform gold coating only at the end of the probe cantilever for increasing laser reflectivity while the rest of the cantilever is uncoated (this type of probe has been commercially available [72]), which will produce the probe that is insensitive to environmental temperature alterations and at the same time has adequate force detection precision. Since contact mode AFM imaging senses the repulsive forces between AFM tip atoms and sample surface atoms to obtain the topographical image of the sample, and the repulsive forces are highly localized (the repulsive force between the tip atom and a target atom on the sample is not susceptible by the neighbor atoms of the target atom), contact mode has a high spatial resolution. In fact, contact mode has long been used to obtain the high-quality topographical image of single membrane proteins (which will be described in Chapter 3) immobilized on substrates in near-physiological buffer solutions in the past decades [84,85] and so far contact mode AFM is still the preferred method to obtain the high-resolution topography of single native membrane proteins under aqueous conditions [52]. Besides, contact mode is suited for imaging the fine structures of single living cells [86,87], which usually requires applying the adequate scan force on living cells depending on which cellular parts are to be visualized (e.g., cell surface structures, cytoskeletons beneath cell surface, and so on). Applications of AFM topographical imaging on single membrane proteins (Chapter 3) and living cells (Chapters 7 and 8) will be described in detail in the subsequent chapters of the book. Notably, the drawback of contact mode is that the lateral dragging of the probe tip across the sample surface can cause mechanical deformation or even damage to the sample, which is detrimental for observing soft and fragile biological samples which are loosely adsorbed onto the substrate. 1.2.3 Noncontact mode Noncontact AFM (NC-AFM) is the only one AFM mode which is able to detect the behaviors of single atoms (including true atomic resolution, control of atomic forces, measurement of atomic forces, measurement of atomic response, observing atomic defects, mechanical manipulation of individual atoms, and mechanical assembly atom by atom) [88]. Atomic imaging of molecules by NC-AFM is commonly performed in frequency-modulated NC-AFM, in which the cantilever is actively oscillated at constant amplitude at its resonance frequency by a feedback circuit and interaction of the tip with the sample surface causing the resonance frequency to shift by Δf which is the key measurement signal in NC-AFM [89]. In the limit of small Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.2 Atomic force microscopy topographical imaging modes 15 amplitudes, the frequency shift (Δf) is proportional to the gradient of the interaction force between tip and sample and therefore the tip-sample interaction forces can be measured by monitoring the frequency shift of the cantilever. NC-AFM usually works in constant height mode, in which the tip is scanned in a fixed plane above the sample surface while the frequency shift of the cantilever is recorded (typically, for atomic imaging, the height of the scanning tip above the sample is a few Angstroms [90]). A notable point is that atomic imaging by NC-AFM usually requires the use of a specific sensor called qPlus [91] which is based on the concept of piezoelectric quartz tuning forks similar to those used as timekeeping elements in wristwatches, allowing AFM operation at oscillation amplitudes down to a few Angstrom. A standard qPlus sensor is created by attaching one of the prongs of the tuning fork to a substrate and attaching a tip to the other prong which acts as a probe with the capability of self-sensing. For more descriptions of qPlus sensors, readers are referred to the recent review of literature by Giessibl [91]. An alternative to the self-sensing piezoelectric quartz sensor is the length extensional resonator or the so-called Kolibri sensor and the performance comparison between qPlus and Kolibri sensors is still under debate in the community [90]. Besides, utilizing NC-AFM to visualize the atomic structures of single molecules often requires tip functionalization [92]. For imaging single molecules, the AFM probe tip should be rather inert to prevent the target molecule from being picked up or moved by the tip, whereas the bare metal tips are often with high reactivity which could lead to the picking up of the target molecule before obtaining an atomic image. So far, the most popular functionalization for high-resolution NC-AFM is the CO tip [92], which is made by picking up a CO molecule with a metal tip (often copper [89]). Notably, NC-AFM is mainly applied in the fields of surface chemistry [93] for single molecular characterization with atomic resolution and requires substantial instrumentation as well as the harsh work environments (often in ultra-high vacuum (UHV) conditions [94]), which are uncommon for other applications, particularly for biomedical applications. The purpose of the brief description of NC-AFM here is to help readers understand AFM imaging techniques. 1.2.4 Tapping mode Tapping mode is also known as intermittent contact mode, dynamic force mode, oscillation mode, or AC mode. In tapping mode (Fig. 1.3A), the probe cantilever vibrates near its resonance frequency while raster scanning the sample surface and the probe tip intermittently touches the sample at its downward movement. When the probe is brought to the sample surface, the interactions between the tip and sample cause changes in both cantilever amplitude and resonance frequency. Either amplitude Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 16 1. Fundamentals and methods of atomic force microscopy for biophysics or resonance frequency of the cantilever can be used as the feedback parameter for tapping mode imaging, but using the amplitude as feedback is technically simpler because it requires only one feedback loop compared with using frequency as feedback which requires three such loops [61]. Hence, so far tapping mode AFM is commonly based on amplitude modulation. However, it should be noted that compared with amplitude modulation, the resonance frequency shift is more sensitive to the forces acting on the tip at the closest approach to the sample surface during the oscillation cycle [50], and frequency modulation is particularly suited for high-resolution imaging AFM such as in NC-AFM described above. During the raster scanning, the amplitudes of the cantilever are detected by the lock-in amplifier in the AFM controller and the piezoelectric tube drives the probe to move vertically to maintain the constant amplitude of the cantilever, yielding the three-dimensional topography of the sample by recording the XYZ displacements of the piezoelectric tube. The amplitude setpoint defines the amplitude of the cantilever oscillation signal to be maintained by the feedback loop and the setpoint reflects the tapping force exerted on the sample. In practice the amplitude setpoint is often chosen at about 80% of the free cantilever amplitude [72]. However, the adequate setpoint should be determined during experiments and it is associated with various factors, including specimen and probe as well as the imaging conditions. Generally, a larger tapping force often means better imaging quality, but also means more likely to cause tip damage (e.g., tip wear, tip break, and tip contamination) which could influence the life of the tip. A feasible way is starting the scanning with a lower tapping force and then increasing it slowly until the imaging quality does not improve anymore. Since tapping mode eliminates the lateral force between tip and sample, tapping mode is good at imaging loosely adsorbed and soft samples (e.g., biomolecules [50] and cellsurface fine structures [95]) which are often challenging for contact mode AFM. Incidentally, tapping mode AFM achieves great success in imaging DNA molecules, which will be described in Chapter 2. Nevertheless, one drawback of tapping mode is that it is unable to directly measure the interaction forces between tip and sample, as the lock-in amplifier only monitors the amplitude changes of the vibrating cantilever, making users unable to quantitatively characterize the interactions between tip and sample. Tapping mode AFM offers a powerful method for mapping the material and mechanical properties of the sample, which is called phase imaging [96]. Depending on the properties of the surface being probed (e.g., composition, friction, viscoelasticity, and adhesion), in addition to the amplitude changes, the interactions between tip and sample surface will also cause the phase changes of cantilever oscillation (called phase lag or phase shift), which can be recorded by the lock-in amplifier of AFM as well. Therefore, the cantilever’s vibration amplitude is used as Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.2 Atomic force microscopy topographical imaging modes 17 the feedback parameter to move the piezoelectric tube vertically to generate the topography of the sample while the cantilever’s vibration phase lag is used to simultaneously generate the material properties map of the sample. Phase lag is caused by the energy dissipation during each contact between the tip and sample, which includes several factors (the inelastic deformation of the sample caused by the tip during the approaching stage, the van der Waals attractive force (Fig. 1.2) and adhesion force needing to overcome by the tip during retracting stage) [72] and are associated with the properties of the sample. The dissipated energy due to the inelastic tipsample interactions can be mathematically described as follows [97]: E5 kω AA0 sinϕ 2 A2 2Q (1.2) where E is the dissipated energy, k is the spring constant of the cantilever, Q is the quality factor, A is the cantilever vibration amplitude during scanning, A0 is the free cantilever vibration amplitude without the tipsample contact, ω is the driving frequency, and ϕ is the phage lag compared to driver. We can see that, since the cantilever vibration amplitude is maintained constant during scanning, the energy dissipation is directly linked to the phase lag and therefore the phage images reflect the mechanical properties of the sample. Tapping mode phase imaging is undoubtedly useful for investigating the material properties and readers are referred to Ref. [96] for more descriptions of the phase imaging theory. Nevertheless, as described above, the force curves are not recorded during tapping mode imaging and thus information on the sample provided by tapping mode phase imaging is limited. Particularly, there has been a new AFM technique called peak force tapping (PFT) mode which provides powerful capabilities to quantitatively characterize the diverse material properties of samples and will be described in the following. 1.2.5 Peak force tapping mode PFT mode originates from the AFM-based force spectroscopy techniques [50,98]. Traditionally, for AFM-based force spectroscopy assays, the AFM tip is controlled to vertically indent the specimens, during which the force curves are recorded. The mechanical properties of the specimens can be extracted from the obtained force curves, which will be described in detail in the following contents. However, traditional force spectroscopy is time-consuming and inefficient with a poor spatiotemporal resolution, and recent developments in AFM techniques allow fast obtaining force curves on each sampling point on the specimen while simultaneously recording Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com We Don’t reply in this website, you need to contact by email for all chapters Instant download. Just send email and get all chapters download. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com You can also order by WhatsApp https://api.whatsapp.com/send/?phone=%2B447507735190&text&type=ph one_number&app_absent=0 Send email or WhatsApp with complete Book title, Edition Number and Author Name. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 18 1. Fundamentals and methods of atomic force microscopy for biophysics topographic images of the specimen [61,99,100], and this technique is called PFT. The principle of PFT is shown in Fig. 1.3B. During AFM’s raster scanning along the xy horizontal plane (the movement trajectory of the AFM tip is denoted by the dashed yellow line in Fig. 1.3BI), the vibrating tip performs approachretract cycle in the vertical direction (z) on the specimen at each sampling point (each tiny grid in Fig. 1.3BI is equal to a sampling point). During the approach-retract movement, the changes in the deflection of the cantilever and the vertical displacements of the piezoelectric tube are recorded, which yield the so-called forcedistance curves. The peak force (as shown in Fig. 1.3BII, the peak force is the force from the cantilever at its maximum deflection during approach stage) of the obtained force curve is then used as the feedback parameter for controlling the piezoelectric tube to move vertically to maintain the peak force constant, which subsequently constructs the topography image of the specimen as the tip horizontally raster scans the specimen. As described above, contact mode AFM and tapping mode AFM record the deflection or the oscillation amplitude (frequency) of the cantilever as the feedback parameter during the scanning, which cannot sense the contact processes between the tip and sample during imaging. On the contrary, PFT records the force curves and uses the peak force of the force curves as the feedback parameter for imaging, and thus the tipsample contact processes during imaging are available from the force curves (e.g., the contact point between the tip and sample is discernible from the force curve and can be used to determine when the tip contacts the sample), facilitating integrating AFM with other complementary techniques to realize the simultaneous multimodal probing (such as combining PFT imaging and infrared imaging [101]) with high controllability and precision, which is quite meaningful for comprehensively understanding life activities. PFT imaging is able to display the multiple properties of the sample as colored maps by analyzing the force curves, which are correlated with the topography of the sample. The force curve contains various physical properties of the sample, as shown in Fig. 1.3BII. For example, the adhesion force reflects the adhesive capabilities of the sample surface. When using a bare tip (without functionalization) to perform PFT imaging, the adhesion force map reflects the nonspecific adhesive features of the sample surface. When using the functionalized tip to perform PFT imaging [102], the adhesion force map reflects the specific adhesive interactions between the target molecules on the sample surface and the antibody molecules on the tip surface, which is meaningful for revealing the molecular behaviors of cells. Particularly, viruses can also be directly attached to the surface of the AFM tip via linker molecules [103], which is useful for probing viruscell interactions at the single-virus level. Detailed descriptions of AFM-based force spectroscopy with the use of functionalized tip will be presented in the Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.3 Atomic force microscopy force spectroscopy techniques 19 following texts. By fitting the force curve with mechanical models, Young’s modulus of the sample can be obtained. Notably, the commercially AFM produced by Bruker Company (Santa Barbara, CA, USA) uses DerjaguinMullerToporov (DMT) to extract Young’s modulus of samples. Some other theoretical models, such as HertzSneddon model (this mode will be described in detail in the following sections) and JohnsonKendallRoberts (JKR) [78], have also been widely used to analyze the force curves to obtain Young’s modulus of samples. The deformation of the sample can be obtained by analyzing the approach curve after the contact point, and the energy dissipation can be obtained by analyzing the area between the approach curve and the retraction curve. Since multiple properties maps of the specimen are generated simultaneously with the topographical image of the specimen during PFT imaging, PFT-based AFM imaging is also called multiparametric AFM [99,102]. The biomedical applications of PFT-based multiparametric imaging will be included in the subsequent chapters. Notably, for utilizing PFT-based AFM to investigate the mechanical properties of the specimens, we need to use appropriate AFM probes based on the characteristics of the specimens. Stiffer cantilevers can cause larger deformation on the specimen, while softer cantilevers have better detection sensitivity, and thus compromises are required for the specimens being probed. Generally, the cantilever spring constant should be comparable with the stiffness of the specimens being probed [104]. 1.3 Atomic force microscopy force spectroscopy techniques 1.3.1 Single-cell mechanical measurement AFM achieves great success in measuring the mechanical properties (e.g., elasticity, viscoelasticity) of single living cells in the past decades and now AFM indentation assay has become a standard method for characterizing the mechanics of cells. Both cellular elasticity and viscoelasticity can be simultaneously measured by applying an AFM probe to vertically perform approach-dwell-retract movements on the cell surface [50]. The process of the AFM approach-dwell-retract cycle is shown in Fig. 1.4A. Firstly, the AFM probe, which is far away from the cell, is controlled by the piezoelectric tube to gradually touch and indent the cell in the vertical direction (stage ①). The contact between the tip and cell will result in the deflection of the AFM cantilever (x) which is detected by the PSD of AFM (Fig. 1.1A). According to Hooke’s law: F 5 kx (1.3) Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 20 1. Fundamentals and methods of atomic force microscopy for biophysics FIGURE 1.4 Measuring cellular viscoelastic properties by atomic force microscopy (AFM) approach-dwell-retract experiments. (A) Schematic diagram of controlling AFM tip to vertically perform approach-dwell-retract movement on the cells. (B) A typical FD curve (I) and FT curve (II) obtained during the approach-dwell-retract process. (I) According to the contact point, the approach curve is converted into the indentation curve and the indentation curve is then fitted by the HertzSneddon model to extract cellular Young’s modulus (the inset). (II) The FT curve is fitted by the two-order Maxwell model to extract the cellular relaxation times (the inset). Source: Reprinted with permission from M. Li, N. Xi, Y. Wang, L. Liu, Nanotopographical surfaces for regulating cellular mechanical behaviors investigated by atomic force microscopy, ACS Biomater. Sci. Eng. 5(10) (2019) 50365050. Copyright 2019 American Chemical Society. where k is the spring constant of the cantilever and x is the deflection of the cantilever, the interaction force between probe and cell is obtained (also called the loading force of probe cantilever). Notably, in order to exactly obtain the loading force exerted by the cantilever, the spring constant of the cantilever needs to be accurately calibrated in advance. Diverse methods have been developed to calibrate the spring constant of cantilever [105], including the static mass hanging method, reference cantilever method, dynamic mass attachment method, resonant frequency method, and thermal noise method. In practice, commonly, force curves are firstly obtained on the stiff substrate to calibrate the deflection sensitivity of the cantilever, and then the spring constant of the cantilever is calibrated by the thermal noise [106] module of AFM. When the loading force of the cantilever achieves the preset value during the approach process, AFM probe stops approaching and dwells on the cell surface for a period of time (stage ②). The purpose of dwelling is to observe the relaxation process of the cell. Subsequently, the AFM Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.3 Atomic force microscopy force spectroscopy techniques 21 probe retracts from the cell surface to its original position (stage ③). During the approach-dwell-retract cycle, the vertical displacements of the AFM piezoelectric tube and the deflections of the AFM cantilever are obtained by AFM, which yields the forcedistance (FD) curve (also called force curve), as shown in Fig. 1.4BI. Each force curve is composed of two portions, an approach curve and a retract curve. Due to the dwelling of the AFM tip on the cell surface, the endpoint of the approach curve does not coincide with the start point of the retract curve. Generally, the approach curve is used for calculating the cellular elastic properties (such as Young’s modulus) [107], whereas the retract curve is used for probing adhesive interactions which usually requires using functionalized tips [47]. By linking an oscilloscope to the output signal of AFM, the PSD signal changes of AFM cantilever versus time are recorded [108,109], which yield the forcetime (FT) curves (also called relaxation curve). The FT curves obtained during the dwelling process of AFM tip on cells reflect the relaxation dynamics of cells, as shown in Fig. 1.4BII. By analyzing the FT curves, cellular viscoelastic properties (such as relaxation time) are obtained. The detailed processes of extracting cellular Young’s modulus from FD curve and relaxation time from FT curve are described in the following, respectively. Cellular Young’s modulus is obtained by fitting the FD curve with theoretical models. Several models have been presented for analyzing the indentation process of AFM tip, including HertzSneddon model, JKR model, DMT model, and OliverPharr model [40]. HertzSneddon model neglects the adhesion forces between the AFM tip and specimen, while JKR and DMT models consider the adhesion forces between the AFM tip and specimen (the JKR model considers the adhesion forces inside the contact area between tip and specimen, while the DMT model considers the adhesion forces outside the contact area) [110]. Thus, strictly speaking, HertzSneddon is applicable when the adhesion force is much smaller than the loading force, the JKR model is applicable in the case of large tips and soft samples with a large adhesion, and the DMT model is applicable in the case of small tips and stiff samples with a small adhesion [78]. The OliverPharr model is mainly used for determining the mechanical properties of thin films [111]. Nevertheless, for practical reasons, the HertzSneddon model is the most widely used one for calculating cellular Young’s modulus from FD curves [104,112114]. Hertz model is applicable to spherical tip and Sneddon extended the Hertz model to conical tip: pffiffiffiffi 4Eδ1:5 R Fspherical 5 (1.4) 3ð1 2 υ2 Þ Fconical 5 2Eδ2 tanθ πð1 2 υ2 Þ (1.5) Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 22 1. Fundamentals and methods of atomic force microscopy for biophysics where υ is the Poisson ratio of cell (cells are often considered as incompressible materials and thus υ 5 0.5), F is the loading force of the AFM probe, δ is the indentation depth, E is the cellular Young’s modulus, θ is the half-opening angle of the conical tip, and R is the radius of the spherical tip. For applying the HertzSneddon model to analyze the FD curves, the FD curve needs to be converted into the indentation curve according to the contact point in the FD curve (the indentation δ is equal to the difference between the vertical displacement change of AFM piezoelectric driver and the deflection of the cantilever). We can imagine that if the specimen being indented is extremely stiff and there is no deformation on the sample during the indentation process, the vertical displacement of the piezoelectric tube is therefore equal to the deflection of the cantilever and the indentation depth is zero. For the soft specimens such as cells, there is a deformation of the specimen during indentation, and therefore the vertical displacement of the piezoelectric tube is the sum of the indentation depth and cantilever deflection. The HertzSneddon model is then used to fit the indentation curve. The inset in Fig. 1.4BI is an example showing the comparison of the HertzSneddon fitting curve and experimental indentation curve. The fitting gives the cellular Young’s modulus E. Notably, the HertzSneddon model assumes that the sample is infinitely thick, but it works well when the indentation into the sample by the tip is less than 10% of the sample thickness [115]. In 2012, Gavara and Chadwick [116] improved the HertzSneddon model to make it applicable for thin samples by introducing a bottom effect correction factor, significantly facilitating probing of the mechanical properties of cellular lamellar structures such as pseudopodia. However, this method requires measuring the thickness of the cell and is more complex than the HertzSneddon model. It should be noted that cellular Young’s modulus measured by AFM is influenced by experimental conditions, such as tip shape, probe loading rate, cellular positions being indented, environmental temperature and measurement medium [117], and thus these conditions should be maintained identical during experiments to eliminate artificial errors and make the results comparable with each other. Besides, cells are highly heterogeneous, and the elastic properties of different intracellular structures are included in the different parts of the recorded force curves. With smaller indentations, the AFM tip senses the stiff cellular cortex which is composed of actin proteins. With larger indentations, the AFM tip senses the softer structures beneath the cellular cortex, such as cytoplasm. When further increasing the loading force to indent the cells, the cell nuclei may be probed by AFM. Another notable point regarding AFM-based cellular mechanical measurement is the geometry of the probe tip. Studies have shown that cellular Young’s modulus measured by the conical tip is significantly larger than that measured by the same cantilever modified with a spherical tip [118], and this is due to the fact that different intracellular Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com We Don’t reply in this website, you need to contact by email for all chapters Instant download. Just send email and get all chapters download. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com You can also order by WhatsApp https://api.whatsapp.com/send/?phone=%2B447507735190&text&type=ph one_number&app_absent=0 Send email or WhatsApp with complete Book title, Edition Number and Author Name. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.3 Atomic force microscopy force spectroscopy techniques 23 structures are probed by the different tips. The spherical tip has a much larger contact area than the conical tip. The conical tip primarily probes the stiff cell cortex, whereas the spherical tip probes both the cell cortex and the softer underlying cytoplasm. Cellular relaxation time is obtained by analyzing the FT curves recorded on cells. Young’s modulus only reflects the elastic properties of cells, while in fact living cells are intrinsically viscoelastic materials and the rheological behavior of cells is closely connected to the fundamental functions of cells. Hence, characterizing the viscoelastic properties of cells is quite meaningful for understanding cell behaviors from the perspective of biomechanics. By the way, AFM is a type of active method for characterizing cellular rheological properties, and passive methods (using the thermal fluctuations of embedded colloidal probes to measure cellular rheology) [119] have also been developed to characterize the rheology of cells. Generally, the FT curve can be fitted by a two-order Maxwell spring-dashpot model to obtain cellular relaxation time [108,109]: FðtÞ 5 A0 1 Ai 2 X e2t=τ i (1.6) i51 τi 5 ηi ; i 5 1; 2 Ei (1.7) where F is the loading force of the AFM probe, A0 is the instantaneous (purely elastic) response, Ai are the ith force amplitudes, τ i is the ith cellular relaxation time, ηi is the ith cellular viscosity, and Ei is the ith cellular Young’s modulus. The cellular relaxation time (τ) is the ratio of cellular viscosity (η) to cellular Young’s modulus (E), and thus cellular relaxation time indicates the viscoelastic properties of cells. In practice, the one-order Maxwell model often does not fit the FT curve well, while the two-order Maxwell model matches the FT curve well [108,109,120,121]. The inset in Fig. 1.4BII is an example showing the comparison of the two-order Maxwell fitting curve and experimental relaxation curve and we can see that the two curves are considerably consistent with each other. Fitting the relaxation curve with a two-order Maxwell model gives two cellular relaxation times (τ 1 and τ 2). During the AFM indentation process, the AFM tip successively probes the cell membrane and cytoskeleton. The first-order cellular relaxation time τ 1 (the fast relaxation time) originates from the rapid deformations of the cell membrane, while the second-order cellular relaxation time τ 2 (the slow relaxation time) originates from the relatively slow reorganizations of cytoskeletons. A notable point is that cellular relaxation times measured by AFM are dependent on the measurement parameters, such as surface dwell time and ramp rate [122], and thus experimental conditions should be identical to make the obtained results comparable. Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 24 1. Fundamentals and methods of atomic force microscopy for biophysics Modifying the AFM cantilever with an individual sphere to form a spherical tip is particularly useful for characterizing cell mechanics since the spherical tip has a defined geometry which facilitates theoretical modeling and the spherical tip could also better represent the mechanical properties of the whole cell compared with a conical tip which only detects the mechanical properties of local areas of cells. We have conveniently fabricated an AFM probe with spherical tips based on AFM micromanipulations [120,121] and the protocol is in the following as a reference to the readers (Fig. 1.5). First, a regular AFM probe is mounted onto the AFM head and the laser signal reflected on the cantilever is adjusted (for some commercial AFMs, such as the Dimension 3100 AFM used here, we need to adjust the laser signal of the cantilever to beyond a certain value and then the AFM head can be moved). Subsequently, a drop of the polystyrene sphere solution (the sphere diameter is B20 μm here and readers can use spheres with different sizes according to their experimental requirements) is placed on a fresh glass slide and a drop of mixed two-part epoxy adhesive (Araldite, USA) is placed on another position of the same glass slide by using a toothpick. The glass slide is then placed onto the specimen stage of AFM. Under the guidance of optical microscopy, the AFM cantilever is controlled to slightly touch the epoxy adhesive and then retract immediately. The AFM cantilever is then moved to contact a single sphere for 10 s and then retract from the glass slide. Fig. 1.5A shows the optical FIGURE 1.5 Fabricating spherical tip for single-cell mechanical analysis. (A) Under the guidance of optical microscopy, a sphere is glued to the end of the atomic force microscopy (AFM) cantilever. Optical images obtained before (I) and after (II) gluing a sphere to the AFM cantilever. (B and C) SEM images of the fabricated spherical tips. (B) Triangular cantilever glued with a sphere. (C) Rectangular cantilever glued with a sphere. Source: (A) Reprinted with permission from M. Li, L. Liu, X. Xiao, N. Xi, Y. Wang, Effects of methotrexate on the viscoelastic properties of single cells probed by atomic force microscopy, J. Biol. Phys. 42(4) (2016) 551569. Copyright 2016 Springer Nature. (B and C) Reprinted with permission from M. Li, L. Liu, N. Xi, Y. Wang, Atomic force microscopy studies on cellular elastic and viscoelastic properties, Sci. China Life Sci. 61(1) (2018) 5767. Copyright 2017 Science China Press and Springer-Verlag GmbH Germany. Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.3 Atomic force microscopy force spectroscopy techniques 25 images before (Fig. 1.5AI) and after (Fig. 1.5AII) gluing a sphere to the cantilever of the AFM probe. The prepared probes with spherical tips are placed in a probe box for 24 h at room temperature for the hardening of epoxy adhesive and then the probe can be used for single-cell mechanical measurement experiments. Experimental results have shown that either triangular or rectangular cantilevers can be easily modified with spherical tips by this method (Fig. 1.5B and C). 1.3.2 Single-cell force spectroscopy AFM-based single-cell force spectroscopy (SCFS) is able to quantify the adhesive interactions of individual cells. Cell adhesion, commonly defined as the binding of a cell to a substrate (the substrate can be another cell, a surface or an organic matrix), is broadly connected to physiological and pathological processes, including embryonic development, assembly of tissues, cellular communication, inflammation, wound healing, tumor metastasis, cell growth, and viral and bacterial infection [123]. AFM-based SCFS offers novel possibilities for seeking answers to the fundamental issues in cell adhesion. The prerequisite of SCFS is functionalizing the AFM cantilever to allow firmly attachment of individual living cells onto the cantilever to prepare single-cell probe. For single-cell probes, weak adsorption of the cell onto the cantilever may cause the sliding of cells during measurements, which results in the failure of the experiments. Commonly, AFM cantilevers are coated with concanavalin A (ConA) to adsorb single cells [124,125]. For doing so, AFM cantilever is firstly incubated with biotin-conjugated bovine serum albumin (BSA) at 37 C overnight, after which the cantilever is washed with phosphate buffered saline (PBS) to remove the unbound molecules. The cantilever is then incubated in streptavidin solution for 30 min at room temperature. After the incubation, the cantilever is washed with PBS again and then is incubated in biotin-conjugated ConA solution for 30 min at room temperature. After the incubation, the ConAfunctionalized cantilever can be stored in PBS for up to 1 week at 4 C. Individual living cells can be tightly attached to the ConA-functionalized cantilever by AFM micromanipulations for SCFS experiments, as shown in Fig. 1.6A. Under the guidance of optical microscopy, the ConAfunctionalized cantilever is moved to gradually approach individual living cells on the substrate (I in Fig. 1.6A). When the cantilever contacts the cell with adequate forces, the cantilever dwells on the cell for about 30 s to allow the formation of firm binding between the cell and cantilever (II in Fig. 1.6A), after which the cantilever retracts from the cell and usually a single-cell probe is prepared (III in Fig. 1.6A). A notable point is that after attaching the cell to the cantilever, the cell needs to recover for at least 10 min, during which the cell will form firm contacts with the cantilever Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 26 1. Fundamentals and methods of atomic force microscopy for biophysics FIGURE 1.6 Atomic force microscopy (AFM)-based single-cell force spectroscopy (SCFS). (A) Attaching single living cell to the ConA-coated cantilever by AFM micromanipulations. (I) The ConA-coated cantilever is approaching a cell on the substrate under the guidance of optical microscopy. (II) The cantilever contacts the cell and dwells on the cell for 30 s to allow the binding of the cell to the cantilever. (III) The cantilever retracts from the substrate and the cell is attached to the cantilever. (IV) Staining the cell probe with CFDA SE confirms the viability of the cell. (B) Schematic of SCFS for measuring the adhesive interactions between cell and surface. The single-cell cantilever is lowered toward the collagen substrate (I) until a preset force is reached (II). After a given contact time, the cantilever is retracted from the surface (III) until cell and substrate are completely separated (IV). (C) A typical FD curve showing steps (I, II, III, IV) corresponding to those outlined in (B). The maximum force required to separate the cell from the surface is referred to the detachment force (F). Source: (A) Reprinted with permission from D. Dang, R. Xiang, B. Liu, X. Liu, M. Li, Quantifying the adhesion forces of lymphoma cells by AFM single-cell force spectroscopy, Prog. Biochem. Biophys. 46(1) (2019) 8998. (B and C) are reprinted with permission from J. Friedrichs, J. Helenius, D. J. Muller, Quantifying cellular adhesion to extracellular matrix components by single-cell force spectroscopy, Nat. Protoc. 5(7) (2010) 13531361. Copyright 2010 Nature America, Inc. surface [125]. In order to examine the viability of the cell on the cantilever, the live/dead reagent CFSE DA is used to stain the cell on the cantilever, and we can see that cells exhibit bright fluorescence after the staining of CFSE DA (IV in Fig. 1.6A), indicating that the cell on the cantilever is alive. With the use of single-cell probe, the adhesive interactions between cell and substrate can then be probed by controlling the probe to vertically perform approach-retract cycle on the substrate, as shown in Fig. 1.6B. Incidentally, Fig. 1.6B only shows an example of adhesive interactions between cell and surface (collagen), and the surface can be replaced with a cell grown on a substrate and in that case the adhesive interactions between two cells are Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.3 Atomic force microscopy force spectroscopy techniques 27 measured. Firstly, the cell probe, which is originally far away from the surface, is controlled to vertically approach the surface. After the cell probe contacts the surface, the probe compresses the surface until the preset force is achieved (II in Fig. 1.6B) and the probe dwells on the surface for a period of time (called the contact time). Subsequently, the cell probe withdraws from the surface, during which the adhesion between the cell and surface could cause the downward deflection of the cantilever (III in Fig. 1.6B). When the pulling force is larger than the adhesion forces between the cell and surface, the cellsurface adhesive bonds rupture and the cell probe return to its original position (IV in Fig. 1.6B). By analyzing the force curves recorded during the approach-retract cycle (Fig. 1.6C) [123,125], the detachment force of the cell (denoted by F in Fig. 1.6C) is obtained. Particularly, studies have shown that two types of smaller unbinding events are often contained in the obtained force curves which are frequently named rupture (denoted by r in Fig. 1.6C) and tether (denoted by t in Fig. 1.6C) events, respectively [126]. In rupture events, the receptor remains anchored in the cell cortex and unbinds as the loading force increases, causing the shrinking of the contact area and subsequently the detaching of the cell body from the substrate. In tether events, receptor anchoring is lost and the membrane tethers are pulled out of the cell, causing the staircase-like manner of the force curve. 1.3.3 Single-molecule force spectroscopy AFM can probe the specific interaction force of single receptorligand pair by the technique called single-molecule force spectroscopy (SMFS). For doing so, ligands are attached to the surface of the AFM tip and then the functionalized tip is controlled to vertically perform approach-retract cycles on the cell surface. The ligands attached to the surface of the AFM tip can specifically bind to the particular receptors on the cell surface. As shown in Fig. 1.7A, the ligandconjugated tip, which is originally far away from the cell, gradually approaches the cell surface (I in Fig. 1.7A). The tip contacts and compresses the cell surface until the loading force achieves the preset maximum value (II in Fig. 1.7A). Subsequently, the tip retracts from the cell surface (III in Fig. 1.7A). If the receptorligand binding occurs during the contact between the AFM tip and cell, the receptorligand pair is stretched by the AFM tip (III in Fig. 1.7A) during the retract process. When the pulling force exerted by the AFM cantilever is larger than the bond strength of the receptorligand pair, the receptorligand pair ruptures and then the AFM tip returns back to its original position (IV in Fig. 1.7A). By analyzing the force curves (Fig. 1.7C) recorded during the approach-retract cycle, the rupture force of the individual Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com We Don’t reply in this website, you need to contact by email for all chapters Instant download. Just send email and get all chapters download. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com You can also order by WhatsApp https://api.whatsapp.com/send/?phone=%2B447507735190&text&type=ph one_number&app_absent=0 Send email or WhatsApp with complete Book title, Edition Number and Author Name. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 28 1. Fundamentals and methods of atomic force microscopy for biophysics FIGURE 1.7 Atomic force microscopy (AFM)-based single-molecule force spectroscopy (SMFS). (A) Schematic diagram of probing single receptors on cell surface by AFM-based SMFS. (I) AFM tip carrying ligands begins approaching cell surface. (II) AFM tip contacts the cell surface, allowing the binding of ligands and receptors. (III) AFM tip retracts from cell surface, which stretches the receptorligand pair. (IV) Receptorligand ruptures when the pulling force exerted by AFM cantilever is larger than the strength of receptorligand bond. (B) Schematic diagram of tip functionalization for SMFS. Not only various biomolecules (I), but also virus (II) can be attached to the AFM tip. (C) A typical force curve recorded during AFM-based SMFS experiments. The denoted steps (I, II, III, IV) correspond to those outlined in (A). Source: (B) Reprinted with permission from D. J. Muller, Y. F. Dufrene, Force nanoscopy of living cells, Curr. Biol. 21(6) (2011) R212R216. Copyright 2011 Elsevier Ltd. (C) is reprinted with permission from M. Li, X. Xiao, L. Liu, N. Xi, Y. Wang, Z. Dong, et al., Nanoscale mapping and organization analysis of target proteins on cancer cells from B-cell lymphoma patients, Exp. Cell Res. 319(18) (2013) 28122821. Copyright 2013 Elsevier Inc. receptorligand pair is obtained. Fig. 1.7C shows a practical force curve obtained during SMFS experiments [127]. The stretching of the receptorligand pair (III in Fig. 1.7C) yields an unbinding force peak in the retract portion of the curve (denoted by the black arrow in Fig. 1.7C) and the magnitude of this peak is equal to the rupture force of the receptorligand pair. When the density of ligands linked to the AFM tip is adjusted to an appropriate level that only one receptorligand pair is formed between the tip-cell contact area, the unbinding peak then corresponds to single receptorligand pair and thus the unbinding force of single molecular pair is measured. A critical point for AFM-based SMFS is linking biomolecules (which can specifically bind to the target receptors on the cell surface) onto the surface of AFM tips, and this process is commonly called tip functionalization (Fig. 1.7B) [128]. Originally, researchers attach biomolecules Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.3 Atomic force microscopy force spectroscopy techniques 29 onto the surface with the use of short rigid linker [129132] (typically avidinbiotin system), and the drawback of this method is that high coverage of ligands increases the risk of probing multiple binding events and increases steric hindrance, which can interfere with the binding process [55]. Besides, ligands linked to the AFM tip have weak conformational mobility. In order to overcome these defects, researchers have used long, flexible and distensible linkers such as polyethylene glycol (PEG) to attach ligands onto AFM tip [133]. For doing so, the PEG linker molecules typically carry two different functional ends. One end of the PEG linker molecule covalently reacts with the amine-functionalized tip surface, and the other end covalently reacts with the ligands. The use of heterobifunctional PEG crosslinker has several advantages. First, the binding strength of covalent bonds (12 nN) is about ten times stronger than typical receptorligand bonds [134], which ensures that the receptorligand pair ruptures during the SMFS process. Second, the ligands at the end of the PEG linker can freely orient in the solution to recognize the receptors on the cell surface, which facilitates the receptorligand binding interactions. Third, during the retract process, stretching the PEG linker will often cause a nonlinear shape of force peak in the force curve (denoted by the green arrow in Fig. 1.7C), which can be used to discriminate the specific receptorligand rupture events and nonspecific rupture events. Hence, so far PEG covalent coupling has been the predominant tip functionalization method for SMFS. Notably, in practice, one end of the PEG linker is commonly NHS, and the other end of the PEG linker is diverse depending on the types of ligands to be attached. In brief, amino-functionalization is firstly performed to coat the AFM probe with a layer of NH2. The NH2 group is able to form covalent bonds with the NHS group of the PEG linker, allowing attachment of the PEG linker to the AFM tip. The last step is attaching ligands to the reactive groups at the other end of the PEG linker, and various strategies (e.g., NHS-PEG-aldehyde, NSH-PEG-maleimide) have been presented to attach a wide range of ligands (e.g., protein, antibody, virus) [135]. Besides, methods based on the affinity between tetravalent nickel and histidine have also been developed for the attachment of His6-tagged ligands, which achieve tip functionalization in a controlled orientation of the ligands. There are several points which should be noted for practical SMFS experiments. Before applying AFM force spectroscopy to probe the target receptors on the cell surface, the expression levels of the receptor on the cell surface need to be confirmed, which can be performed by staining the target receptors with fluorescein and the subsequent recording of the fluorescent images [136]. For AFM force spectroscopy experiments, due to the fact that membrane proteins are often heterogeneously distributed on the cell surface, force curves are commonly recorded at different surface points of multiple cells (at least 10 cells) to Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 30 1. Fundamentals and methods of atomic force microscopy for biophysics obtain statistical results and the probability of finding specific molecular unbinding events usually ranges from 10% to 30% [137]. Besides, blocking experiments are often required to demonstrate the specific receptorligand binding interactions, during which free ligands are added to block the receptors on the cell surface and then the functionalized tip is applied to record force curves on the cell surface. For the force curves obtained on cells after blocking, there should be no, or few specific unbinding force peaks. Since receptorligand interactions are closely related to the cell states, one should carefully monitor cell morphology and cell viability during SMFS experiments and only force curves obtained from the same cell morphology/viability can be compared with each other [55]. In addition, the rupture force of receptorligand pair measured by AFM is dependent on the loading rate (retraction velocity multiplied by the spring constant of the cantilever) of the AFM cantilever, which is theoretically described by the BellEvans model [138]: F5 kB T rxβ ln xβ koff kB T (1.8) where F is the rupture force, r is the loading rate, koff is the kinetic dissociation rate at zero force, xβ is the distance to the transition state along the projection of the applied force, kB is the Boltzmann’s constant, and T is the temperature. The formula (1.8) indicates the linear relationship between the measured receptorligand rupture force and the logarithm of the loading rate. Hence, in practice, rupture forces are measured at different pulling velocities to investigate the dissociation dynamics of the molecular interactions and at least 1000 force curves [55] are required to record one pulling velocity. AFM-based SMFS also allows the revealing of the unfolding dynamics of single membrane proteins. Most proteins have to acquire an adequate three-dimensional structure to perform biological functions. The folding of proteins into their compact three-dimensional structures is one of the most fundamental examples of biological self-assembly. Protein misfolding is associated with many human diseases, including neurodegenerative diseases (e.g., Alzheimer’s disease, spongiform encephalopathies, Parkinson’s disease), nonneuropathic systemic amyloidosis (e.g., amyloid A (AA) amyloidosis), and nonneuropathic localized amyloidosis (e.g., type II diabetes) [139]. Hence, understanding the protein folding process significantly benefits understanding the underlying mechanisms guiding protein functions and behaviors. Utilizing AFM-based SMFS to mechanically unfold single proteins provides unique insights into the mechanics of proteins. For doing this, AFM tip does not require tip functionalization. After immobilizing membrane protein patches, either natural membranes containing highly Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.4 High-speed atomic force microscopy 31 ordered proteins (e.g., purple membrane) or artificial membranes containing reconstituted proteins [140], onto the substrate, the AFM tip is moved to above single membrane proteins based on high-resolution AFM imaging of single membrane proteins [141]. AFM tip is then controlled to approach and touch a single membrane protein with a contact force of B0.51 nN for B1 s, which results in firm adsorption of the protein to the AFM tip in about 15% of all cases [85]. Subsequently, the AFM tip retracts from the membrane, which mechanically unfolds the membrane protein. Force curves are recorded during the approach-retract cycle of the tip. Notably, if the polypeptide adsorbed to the AFM tip is the C- or N-terminal end of the protein, the force curves will exhibit a length corresponding to that of the entirely stretched protein. If the protein attaches with a polypeptide loop or slips off the tip before being completely unfolded, the force curves will be shorter and thus difficult to interpret. Therefore, only force curves representing the entirely unfolded and stretched protein should be used for analysis [142]. The force curve with a fully membrane protein unfolding process shows the sawtooth-like pattern of force peaks, each of which corresponds to the unfolding of a segment of membrane protein. The force curve is fitted by the worm-like chain (WLC) model to analyze the unfolding pathway of the protein [55,84]: " # kB T 1 x 22 x 1 12 1 2 (1.9) FðxÞ 5 lp 4 Lc L0 4 where F(x) is the force, Lc is the contour length of the peptide, lp is the persistence length of the peptide which describes the rigidity of the polymer, x is the extension of peptide, kB is the Boltzmann’s constant, and T is the temperature. Fitting each sawtooth-like peak with WLC model gives the number of amino acids of the stretched segment and thus the unfolding pathway of the protein is generated after fitting all the peaks. 1.4 High-speed atomic force microscopy The developments of high-speed AFM enable capturing the dynamics of biomolecular activities at work with unprecedented temporal resolution. Life is dynamic and the functions of biological systems depend on the dynamic processes that occur in biomolecules, organelles, and cells [143]. Observing the real-time dynamics of single biomolecules at work is considerably meaningful for uncovering the underpinnings of biological processes. AFM is the only technique which is able to visualize the structure of single native biomolecules with nanometer spatial resolution under aqueous conditions. However, the topographical imaging Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 32 1. Fundamentals and methods of atomic force microscopy for biophysics speed of AFM has long been slow. It often takes minutes to acquire a topographical image of biological specimens by conventional AFM, which is much greater than the time scale at which dynamic processes usually occur in biology [144]. Therefore, strong demand of AFM for biomedical applications is the ability to rapidly acquire successive high-resolution images of individual biomolecules in action. In 2001, Ando et al. [145] developed a high-speed AFM for studying biomolecules based on tapping mode, which is able to capture a 100 3 100 pixel2 image within 80 Ms and the experimental results exhibit the exciting capabilities of the high-speed AFM to capture the Brownian motions of single myosin V molecules in solution. Since then, highspeed AFM techniques has been continuously advanced and now high-speed AFM is coming of age [146] for biomedical applications. High-speed AFM is able to not only watch single biomolecules in action in real-time [147], but also can visualize the morphological dynamics of single living cells [148], contributing much to molecular and cell biology. High-speed AFM typically operates in tapping mode [149], in which the cantilever is excited to vibrate near its resonance frequency to intermittently contact the sample surface. The highest possible imaging rate of tapping mode AFM is determined by the bandwidth of feedback control to maintain the tapping amplitude constant during scanning as well as by scan parameters and the sample itself [146]. The details of the basic principles of high-speed AFM are beyond the scope of this book and readers are referred to the references [143,146,149] for more descriptions of the quantitative relationship between the feedback bandwidth and the various factors involved in AFM and the scanning conditions. Briefly, in the high-speed AFM system (Fig. 1.8A), various devices, including cantilevers (Fig. 1.8B), electronic circuits, the sample-stage scanner (Fig. 1.8C), and the cantilever deflection detection system, are optimized for achieving highspeed performance [150,151]. For example, cantilevers are miniaturized to achieve high resonant frequencies and small spring constants, while a new feedback control technique capable of maintaining weak tip-sample interactions and active camping techniques to suppress the scanner’s mechanical vibrations have been implemented. Notably, Fig. 1.8C shows a narrow-area scanner for high-speed AFM. Researchers have also developed a wide-area scanner for high-speed AFM [152] which has a maximum XY scan range of 46 3 46 μm2 and is able to capture the dynamic processes of single living bacterial and eukaryotic cells. High-speed AFM has now become an important method for observing the real-time dynamics of life processes and the practical applications of utilizing high-speed AFM to address biological issues will be presented in the subsequent chapters. Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com We Don’t reply in this website, you need to contact by email for all chapters Instant download. Just send email and get all chapters download. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com You can also order by WhatsApp https://api.whatsapp.com/send/?phone=%2B447507735190&text&type=ph one_number&app_absent=0 Send email or WhatsApp with complete Book title, Edition Number and Author Name. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 1.5 Topography and recognition imaging mode atomic force microcopy 33 FIGURE 1.8 High-speed atomic force microscopy (AFM). (A) Schematic diagram of the configuration of the high-speed system. (B) SEM image of a small cantilever for highspeed AFM. The inset shows an electron beam deposited amorphous carbon tip grown on the original tip. (C) Structure of a high-speed AFM scanner for narrow area imaging ( , 1 μm 3 4 μm). Source: (A) Reprinted with permission from T. Uchihashi, N. Kodera, T. Ando, Guide to video recording of structure dynamics and dynamic processes of proteins by high-speed atomic force microscopy, Nat. Protoc. 7(6) (2012) 11931206. Copyright 2012 Nature America, Inc. (B and C) Reprinted with permission from T. Ando, High-speed AFM imaging, Curr. Opin. Struct. Biol. 28 (2014) 6368. Copyright 2014 Elsevier Ltd. 1.5 Topography and recognition imaging mode atomic force microcopy The advent of simultaneous topography and recognition imaging (TREC) mode AFM offers novel opportunities for detecting molecular behaviors. Traditionally, in order to apply AFM force spectroscopy techniques to map the spatial distributions of particular receptors on single cells, the functionalized probes are used to obtain arrays of force curves at the small local areas on the cell surface [153]. For an array of force curves, the unbinding force of each force curve is calculated from the force peak in the retract curve (as denoted by the black arrow in Fig. 1.7C). If there are no unbinding force peaks, the unbinding force is set to zero. After normalizing the unbinding forces into gray colors (0255), a force map is constructed by using imaging processing software and the brightness of the pixel reflects the magnitude of unbinding forces. This method is called force volume mode which is time-consuming and inefficient. In 2004, Stroh et al. [154] developed a new molecular recognition mode based on AFM, which is called TREC. TREC is able to simultaneously obtain a topography image and a recognition image on the cell surface and the imaging time is comparable to Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 34 1. Fundamentals and methods of atomic force microscopy for biophysics conventional AFM imaging. TREC mode is based on tapping mode AFM imaging. In TREC mode (Fig. 1.9A), the oscillating tip which carries ligands raster scans the cell surface. Both the maxima and minima of the oscillation periods are recorded. During scanning, if the ligand FIGURE 1.9 Topography and recognition imaging (TREC) mode atomic force microscopy (AFM). (A) Schematic diagram of the principle of detecting cell surface receptors by TREC mode AFM. An oscillating AFM tip tethered with a ligand molecule scans across the cell surface to elucidate the nanoscale organization of receptor molecules. The cantilever oscillation amplitude is split into lower and upper parts, resulting in simultaneously acquired topography and recognition images. The topography image is generated from the lower part and the corresponding recognition map is constructed from the upper part of the oscillation amplitude. The recognition events originate from the PEG linker stretching which results in a damping of the upper part of the oscillation. (B) Amplitude signals of cantilever oscillations. The maxima signals of the cantilever oscillation amplitudes (denoted by red arrows) keep constant when scanning with a bare tip (I). Scanning with a functionalized tip results in significant changes in the maxima signals (denoted by blue arrows) while the topographical information is also contained in the minima signals (II). (C) Topography image (I) and corresponding recognition map (II) of thrombin protein molecules deposited on mica. The circles indicate those protein molecules that were recognized, whereas the square indicates the protein that was not recognized. (D) Topography image (I) and corresponding recognition map (II) of receptors on mica surface. In the recognition map, dark spots indicate the regions where the antibody on the tip is bound to the target receptors on surface. The white arrows indicate the interactions with large aggregates. Source: (A) Reprinted with permission from L.A. Chtcheglova, P. Hinterdorfer, Simultaneous AFM topography and recognition imaging at the plasma membrane of mammalian cells, Semin. Cell Dev. Biol. 73 (2018) 4556. Copyright 2017 Elsevier Ltd. (B) Reprinted with permission from C. M. Stroh, A. Ebner, M. Geretschlager, G. Freudenthaler, F. Kienberger, A. S. M. Kamruzzahan, et al., Simultaneous topography and recognition imaging using force microscopy, Biophys. J. 87(3) (2004) 19811990. Copyright 2004 The Biophysical Society. (C) Reprinted with permission from S. Senapati, S. Manna, S. Lindsay, P. Zhang, Application of catalyst-free click reactions in attaching affinity molecules to tips of atomic force microscopy for detection of protein biomarkers, Langmuir 29(47) (2013) 1462214630. Copyright 2013 American Chemical Society. (D) Reprinted with permission from P. Kaur, Q. Fu, A. Fuhrmann, R. Ros, L.O. Kutner, L.A. Schneeweis, et al., Antibody-unfolding and metastable-state binding in force spectroscopy and recognition imaging, Biophys. J. 100(1) (2011) 243250. Copyright 2011 the Biophysical Society. Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com References 35 molecules on the tip do not bind to the receptors on the cell surface, the tip vibrates like imaging with a nonfunctionalized tip. When the ligand molecules bind to the receptors on the cell surface, the subsequent unbinding events of receptorligand pairs will result in changes in the upper part of the vibration amplitude signal (maxima) but does not influence the lower part of the vibration amplitude signal (minima) (Fig. 1.9B) [155]. When using a bare tip (nonfunctionalized tip) to scan the surface of specimen, the maxima signals of cantilever oscillations keep constant while the topographical information is contained in the minima of the cantilever oscillations (I in Fig. 1.9B). When using a functionalized tip to perform scanning, not only the topographical information is contained in the minima of the cantilever oscillations, but also the specific molecular unbinding events are contained in the maxima of cantilever oscillations (II in Fig. 1.9B). By using a special electronic circuit called TREC box, the recognition image is generated simultaneously with the topographical image [156], therefore allowing correlating receptor distribution with cell surface structures. For TREC mode AFM, the cantilever oscillation amplitude should be slightly smaller than the extended PEG linker length, so that the ligand remains bound while passing a binding site and the reduction of the maxima of cantilever oscillations is of sufficient significance compared to the thermal noise. With TREC mode AFM, not only the purified biomolecules deposited on the substrate can be recognized [157159] (Fig. 1.9C and D), but also the proteins reconstituted in the artificial lipid bilayer membranes [160] and even the receptors on the cell surface [136,156,161,162] can be recognized, which provide novel insights into the molecular interactions and practical applications of TREC mode AFM will be presented in the subsequent chapters. Notably, TREC does not record force curves during imaging, and thus quantitative force information about molecular binding events is missing. References [1] B. Alberts, A. Johnson, J. Lewis, D. Morgan, M. Raff, K. Roberts, et al., Molecular Biology of the Cell, sixth ed., Garland Science, New York, 2014. [2] R.E. Dawes-Hoang, E.F. Wieschaus, Cell and developmental biology—a shared past, an intertwined future, Dev. Cell 1 (1) (2001) 2736. [3] E.B. Wilson, The Cell in Development and Heredity, third ed., Macmillan, New York, 1925. [4] X.S. Xie, J. Yu, W.Y. Yang, Living cells as test tubes, Science 312 (5771) (2006) 228230. [5] M. Labib, S.O. Kelley, Single-cell analysis targeting the proteome, Nat. Rev. Chem. 4 (3) (2020) 143158. [6] H. Yin, D. Marshall, Microfluidics for single cell analysis, Curr. Opin. Biotechnol. 23 (1) (2012) 110119. [7] P. Paszek, S. Ryan, L. Ashall, K. Sillitoe, C.V. Harper, D.G. Spiller, et al., Population robustness arising from cellular heterogeneity, Proc. Natl. Acad. Sci. USA 107 (25) (2010) 1164411649. Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 36 1. Fundamentals and methods of atomic force microscopy for biophysics [8] L. Pelkmans, Using cell-to-cell variability-a new era in molecular biology, Science 336 (6080) (2012) 425426. [9] N. Komin, A. Skupin, How to address cellular heterogeneity by distribution biology, Curr. Opin. Syst. Biol. 3 (2017) 154160. [10] Y. Li, Y. Shan, R.V. Desai, K.H. Cox, L.S. Weinberger, J.S. Takahashi, Noise-driven cellular heterogeneity in circadian periodicity, Proc. Nat. Acad. Sci. USA 117 (19) (2020) 1035010356. [11] B. Carter, K. Zhao, The epigenetic basis of cellular heterogeneity, Nat. Rev. Genet. 22 (4) (2021) 235250. [12] K. Hinohara, K. Polyak, Intratumoral heterogeneity: more than just mutations, Trends Cell Biol. 29 (7) (2019) 569579. [13] M.D. Brooks, M.L. Burness, M.S. Wicha, Therapeutic implications of cellular heterogeneity and plasticity in breast cancer, Cell Stem Cell 17 (3) (2015) 260271. [14] R.A. Burrell, N. McGranahan, J. Bartek, C. Swanton, The causes and consequences of genetic heterogeneity in cancer evolution, Nature 501 (7467) (2013) 338345. [15] A. Pribluda, C.C. de la Cruz, E.L. Jackson, Intratumoral heterogeneity: from diversity comes resistance, Clin. Cancer Res. 21 (13) (2015) 29162923. [16] I. Vitale, E. Shema, S. Loi, L. Galluzzi, Intratumoral heterogeneity in cancer progression and response to immunotherapy, Nat. Med. 27 (2) (2021) 212224. [17] X. Chen, J.C. Love, N.E. Navin, L. Pachter, M.J.T. Stubbington, V. Svensson, et al., Single-cell analysis at the threshold, Nat. Biotechnol. 34 (11) (2016) 11111118. [18] A.J. Hughes, D.P. Spelke, Z. Xu, C.C. Kang, D.V. Schaffer, A.E. Herr, Single-cell western blotting, Nat. Methods 11 (7) (2014) 749755. [19] J.R. Heath, A. Ribas, P.S. Mischel, Single-cell analysis tools for drug discovery and development, Nat. Rev. Drug. Discov. 15 (3) (2016) 204216. [20] V.Y. Kiselev, T.S. Andrews, M. Hemberg, Challenges in unsupervised clustering of single-cell RNA-seq data, Nat. Rev. Genet. 20 (5) (2019) 273282. [21] D.A. Lawson, N.R. Bhakta, K. Kessenbrock, K.D. Prummel, Y. Yu, K. Takai, et al., Single-cell analysis reveals a stem-cell program in human metastatic breast cancer cells, Nature 526 (7571) (2015) 131135. [22] H.W. Jackson, J.R. Fischer, V.R.T. Zanotelli, H.R. Ali, R. Mechera, S.D. Soysal, et al., The single-cell pathology landscape of breast cancer, Nature 578 (7796) (2020) 615620. [23] A.M. van der Leun, D.S. Thommen, T.N. Schumacher, CD81 T cells states in human cancer: insights from single-cell analysis, Nat. Rev. Cancer 20 (4) (2020) 218232. [24] M.A. Durante, D.A. Rodriguez, S. Kurtenbach, J.N. Kuznetsov, M.I. Sanchez, C.L. Decatur, et al., Single-cell analysis reveals new evolutionary complexity in uveal melanoma, Nat. Commun. 11 (2020) 496. [25] S. Dumont, M. Prakash, Emergent mechanics of biological structures, Mol. Biol. Cell 25 (22) (2014) 34613465. [26] A. Diz-Munoz, O.D. Weiner, D.A. Fletcher, In pursuit of the mechanics that shape cell surfaces, Nat. Phys. 14 (7) (2018) 648652. [27] D.E. Discher, P. Janmey, Y.L. Wang, Tissue cells feel and respond to the stiffness of their substrate, Science 310 (5751) (2005) 11391143. [28] A.J. Engler, S. Sen, H.L. Sweeney, D.E. Discher, Matrix elasticity directs stem cell lineage specification, Cell 126 (4) (2006) 677689. [29] S. Suresh, Biomechanics and biophysics of cancer cells, Acta Biomater. 3 (4) (2007) 413438. [30] D. Wirtz, K. Konstantopoulos, P.C. Searson, The physics of cancer: the role of physical interactions and mechanical forces in metastasis, Nat. Rev. Cancer 11 (7) (2011) 512522. [31] C. Alibert, B. Goud, J.B. Manneville, Are cancer cells really softer than normal cells? Biol. Cell 109 (5) (2017) 167189. Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com References 37 [32] P.K. Chaudhuri, B.C. Low, C.T. Lim, Mechanobiology of tumor growth, Chem. Rev. 118 (14) (2018) 64996515. [33] S. Suresh, J. Spatz, J.P. Mills, A. Micoulet, M. Dao, C.T. Lim, et al., Connections between single-cell biomechanics and human disease states: gastrointestinal cancer and malaria, Acta Biomater. 1 (1) (2005) 1530. [34] Y. Park, M. Diez-Silva, G. Popescu, G. Lykotrafitis, W. Choi, M.S. Feld, et al., Refractive index maps and membrane dynamics of human red blood cells parasitized by plasmodium falciparum, Proc. Natl. Acad. Sci. USA 105 (37) (2008) 1373013735. [35] D.A. Fedosov, B. Caswell, S. Suresh, G.E. Karniadakis, Quantifying the biophysical characteristics of plasmodium-falciparum-parasitized red blood cells in microcirculation, Proc. Natl. Acad. Sci. USA 108 (1) (2011) 3539. [36] C.F. Guimaraes, L. Gasperini, A.P. Marques, R.L. Reis, The stiffness of living tissues and its implications for tissue engineering, Nat. Rev. Mater. 5 (5) (2020) 351370. [37] M.C. Lampi, C.A. Reinhart-King, Targeting extracellular matrix stiffness to attenuate disease: from molecular mechanisms to clinical trials, Sci. Transl. Med. 10 (422) (2018). eaao0475. [38] H. Mohammadi, E. Sahai, Mechanisms and impact of altered tumor mechanics, Nat. Cell Biol. 20 (7) (2018) 766774. [39] D. Alsteens, H.E. Gaub, R. Newton, M. Pfreundschuh, C. Gerber, D.J. Muller, Atomic force microscopy-based characterization and design of biointerfaces, Nat. Rev. Mater. 2 (5) (2017) 17008. [40] M. Li, D. Dang, L. Liu, N. Xi, Y. Wang, Atomic force microscopy in characterizing cell mechanics for biomedical applications: a review, IEEE Trans. Nanobiosci. 16 (6) (2017) 523540. [41] C. Frantz, K.M. Stewart, V.M. Weaver, The extracellular matrix at a glance, J. Cell Sci. 123 (24) (2010) 41954200. [42] B.D. Hoffman, C. Grashoff, M.A. Schwartz, Dynamic molecular processes mediate cellular mechanotransduction, Nature 475 (7356) (2011) 316323. [43] G.S. Hussey, J.L. Dziki, S.F. Badylak, Extracellular matrix-based materials for regenerative medicine, Nat. Rev. Mater. 3 (7) (2018) 159173. [44] J.A. Eble, S. Niland, The extracellular matrix in tumor progression and metastasis, Clin. Exp. Metastasis 36 (3) (2019) 171198. [45] J. Winkler, A. Abisoye-Ogunniyan, K.J. Metcalf, Z. Werb, Concepts of extracellular matrix remodeling in tumor progression and metastasis, Nat. Commun. 11 (2020) 5120. [46] P. Lu, V.M. Weaver, Z. Werb, The extracellular matrix: a dynamic niche in cancer progression, J. Cell Biol. 196 (4) (2012) 395406. [47] D.J. Muller, J. Helenius, D. Alsteens, Y.F. Dufrene, Force probing surfaces of living cells to molecular resolution, Nat. Chem. Biol. 5 (6) (2009) 383390. [48] P. Hinterdorfer, M.F. Garcia-Parajo, Y.F. Dufrene, Single-molecule imaging of cell surfaces using near-field nanoscopy, Acc. Chem. Res. 45 (3) (2012) 327336. [49] D.S. Dimitrov, Virus entry: molecular mechanisms and biomedical applications, Nat. Rev. Microbiol. 2 (2) (2004) 109122. [50] M. Li, N. Xi, Y. Wang, L. Liu, Atomic force microscopy in probing tumor physics for nanomedicine, IEEE Trans. Nanotechnol. 18 (2019) 83113. [51] B.W. Hoogenboom, Stretching the resolution limit of atomic force microscopy, Nat. Struct. Mol. Biol. 28 (8) (2021) 629635. [52] G.R. Heath, E. Kots, J.L. Robertson, S. Lansky, G. Khelashvili, H. Weinstein, et al., Localization atomic force microscopy, Nature 594 (7863) (2021) 385390. [53] D.J. Muller, Y.F. Dufrene, Atomic force microscopy as a multifunctional molecular toolbox in nanobiotechnology, Nat. Nanotechnol. 3 (5) (2008) 261269. [54] M. Krieg, G. Flaschner, D. Alsteens, B.M. Gaub, W.H. Roos, G.J.L. Wuite, et al., Atomic force microscopy-based mechanobiology, Nat. Rev. Phys. 1 (1) (2019) 4157. Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com We Don’t reply in this website, you need to contact by email for all chapters Instant download. Just send email and get all chapters download. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com You can also order by WhatsApp https://api.whatsapp.com/send/?phone=%2B447507735190&text&type=ph one_number&app_absent=0 Send email or WhatsApp with complete Book title, Edition Number and Author Name. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 38 1. Fundamentals and methods of atomic force microscopy for biophysics [55] A. Viljoen, M. Mathelie-Guinlet, A. Ray, N. Strohmeyer, Y.J. Oh, P. Hinterdorfer, et al., Force spectroscopy of single cells using atomic force microscopy, Nat. Rev. Meth. Primers 1 (2021) 63. [56] J. Xiao, Y.F. Dufrene, Optical and force nanoscopy in microbiology, Nat. Microbiol. 1 (11) (2016) 16186. [57] B. Huang, Super-resolution optical microscopy: multiple choices, Curr. Opin. Chem. Biol. 14 (1) (2010) 1014. [58] Y.M. Sigal, R. Zhou, X. Zhuang, Visualizing and discovering cellular structures with super-resolution microscopy, Science 361 (6405) (2018) 880887. [59] M. Li, D. Dang, N. Xi, Y. Wang, L. Liu, Nanoscale imaging and force probing of biomolecular systems using atomic force microscopy: from single molecules to living cells, Nanoscale 9 (45) (2017) 1764317666. [60] R.F. Egerton, Physical Principles of Electron Microscopy: An Introduction to TEM, SEM, and AEM, second ed., Springer, Boston, 2016. [61] Y.F. Dufrene, T. Ando, R. Garcia, D. Alsteens, D. Martinez-Martin, A. Engel, et al., Imaging modes of atomic force microscopy for application in molecular and cell biology, Nat. Nanotechnol. 12 (4) (2017) 295307. [62] Y. Cheng, N. Grigorieff, P.A. Penczek, T. Walz, A primer to single-particle cryo-electron microscopy, Cell 161 (3) (2015) 438449. [63] K.C. Neuman, A. Nagy, Single-molecule force spectroscopy: optical tweezers, magnetic tweezers and atomic force microscopy, Nat. Meth. 5 (6) (2008) 491505. [64] Y. Seol, K.C. Neuman, Snapshot: force spectroscopy and single-molecule manipulation, Cell 153 (5) (2013) 1168. [65] F.M. Fazal, S.M. Block, Optical tweezers study life under tension, Nat. Photon. 5 (6) (2011) 318321. [66] I. De Vlaminck, C. Dekker, Recent advances in magnetic tweezers, Annu. Rev. Biophys. 41 (2012) 453472. [67] D. Kilinc, G.U. Lee, Advances in magnetic tweezers for single molecule and cell biophysics, Integr. Biol. 6 (1) (2014) 2734. [68] C. Arbore, L. Perego, M. Sergides, M. Capitanio, Probing force in living cells with optical tweezers: from single-molecule mechanics to cell mechanotransduction, Biophys. Rev. 11 (5) (2019) 765782. [69] H.G. Hansma, Varieties of imaging with scanning probe microscopes, Proc. Natl. Acad. Sci. USA 96 (26) (1999) 1467814680. [70] H. Habibullah, 30 Years of atomic force microscopy: creep, hysteresis, cross-coupling, and vibration problems of piezoelectric tube scanners, Measurement 159 (2020) 107776. [71] V.J. Morris, A.R. Kirby, A.P. Gunning, Atomic Force Microscopy for Biologists, second ed., Imperial College Press, London, 2010. [72] W. Sun, Principles of atomic force microscopy, in: J. Cai (Ed.), Atomic Force Microscopy in Molecular and Cell Biology, Springer, Singapore, 2018, pp. 128. [73] M. Li, L. Liu, N. Xi, Y. Wang, X. Xiao, W. Zhang, Nanoscale imaging and mechanical analysis of Fc receptor-mediated macrophage phagocytosis against cancer cells, Langmuir 30 (6) (2014) 16091621. [74] M. Li, N. Xi, Y. Wang, L. Liu, In situ high-resolution AFM imaging and force probing of cell culture medium-forming nanogranular surfaces for cell growth, IEEE Trans. Nanobiosci. 19 (3) (2020) 385393. [75] J.N. Israelachvili, Intermolecular and Surface Forces, third ed., Elsevier, Amsterdam, 2011. [76] G. Haugstad, Atomic Force Microscopy: Understanding Basic Modes and Advanced Applications, John Wiley & Sons, New Jersey, 2012. [77] P. Klapetek, Quantitative Data Processing in Scanning Probe Microscopy, Elsevier, Amsterdam, 2013. Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com References 39 [78] H.J. Butt, B. Cappella, M. Kappl, Force measurements with the atomic force microscope: technique, interpretation and applications, Surf. Sci. Rep. 59 (2005) 1152. [79] P. Eaton, P. West, Atomic Force Microscopy, Oxford University Press, Oxford, 2010. [80] U. Maver, T. Velnar, M. Gaberscek, O. Planinsek, M. Finsgar, Recent progressive use of atomic force microscopy in biomedical applications, TrAC. Trends Anal. Chem. 80 (2016) 96111. [81] T. Ando, High-speed atomic force microscopy and its future prospects, Biophys. Rev. 10 (2) (2018) 285292. [82] G.R. Heath, S. Scheuring, High-speed AFM height spectroscopy reveals μs-dynamics of unlabeled biomolecules, Nat. Commun. 9 (2018) 4983. [83] A.B. Churnside, R.M.A. Sullan, D.M. Nguyen, S.O. Case, M.S. Bull, G.M. King, et al., Routine and timely sub-piconewton force stability and precision for biological applications of atomic force microscopy, Nano Lett. 12 (7) (2012) 35573561. [84] C.A. Bippes, D.J. Muller, High-resolution atomic force microscopy and spectroscopy of native membrane proteins, Rep. Prog. Phys. 74 (8) (2011) 086601. [85] M. Li, N. Xi, Y. Wang, L. Liu, Atomic force microscopy as a powerful multifunctional tool for probing the behaviors of single proteins, IEEE Trans. Nanobiosci. 19 (1) (2020) 7899. [86] M. Li, N. Xi, Y. Wang, L. Liu, Advances in atomic force microscopy for single-cell analysis, Nano Res. 12 (4) (2019) 703718. [87] C. Le Grimellec, E. Lesniewska, M.C. Giocondi, E. Finot, V. Vie, J.P. Goudonnet, Imaging of the surface of living cells by low-force contact-mode atomic force microscopy, Biophys. J. 75 (2) (1998) 695703. [88] S. Morita, R. Wiesendanger, E. Meyer, Noncontact Atomic Force Microscopy, Springer, New York, 2002. [89] N. Pavlicek, L. Gross, Generation, manipulation and characterization of molecules by atomic force microscopy, Nat. Rev. Chem. 1 (1) (2017) 0005. [90] S. Morita, R. Wiesendanger, E. Meyer, Noncontact Atomic Force Microscopy, vol. 3, Springer, Heidelberg, 2015. [91] F.J. Giessibl, The qPlus sensor, a powerful core for the atomic force microscope, Rev. Sci. Instrum. 90 (1) (2019) 011101. [92] L. Gross, B. Schuler, N. Pavlicek, S. Fatayer, Z. Majzik, N. Moll, et al., Atomic force microscopy for molecular structure elucidation, Angew. Chem. Int. Ed. 57 (15) (2018) 38883908. [93] F. Huber, J. Berwanger, S. Polesya, S. Mankovsky, H. Ebert, F.J. Giessibl, Chemical bond formation showing a transition from physisorption to chemisorption, Science 366 (6462) (2019) 235238. [94] J.V. Lauritsen, M. Reichling, Atomic resolution non-contact atomic force microscopy of clean metal oxide surfaces, J. Phys. Condens. Matter 22 (26) (2010) 263001. [95] M. Li, L. Liu, N. Xi, Y. Wang, Z. Dong, X. Xiao, et al., Progress of AFM single-cell and single-molecule morphology imaging, Chin. Sci. Bull. 58 (26) (2013) 31773182. [96] R. Garcia, R. Perez, Dynamic atomic force microscopy methods, Surf. Sci. Rep. 47 (2002) 197301. [97] S. Zhang, H. Aslan, F. Besenbacher, M. Dong, Quantitative biomolecular imaging by dynamic nanomechanical mapping, Chem. Soc. Rev. 43 (21) (2014) 74127429. [98] M. Li, N. Xi, L. Liu, Peak force tapping atomic force microscopy for advancing cell and molecular biology, Nanoscale 13 (18) (2021) 83588375. [99] Y.F. Dufrene, D. Martinez-Martin, I. Medalsy, D. Alsteens, D.J. Muller, Multiparametric imaging of biological systems by force-distance curve-based AFM, Nat. Meth. 10 (9) (2013) 847854. [100] M. Pfreundschuh, D. Martinez-Martin, E. Mulvihill, S. Wegmann, D.J. Muller, Multiparametric high-resolution imaging of native proteins by force-distance curvebased AFM, Nat. Protoc. 9 (5) (2014) 11131130. Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 40 1. Fundamentals and methods of atomic force microscopy for biophysics [101] L. Wang, H. Wang, M. Wagner, Y. Yan, D.S. Jakob, X.G. Xu, Nanoscale simultaneous chemical and mechanical imaging via peak force infrared microscopy, Sci. Adv. 3 (6) (2017) e1700255. [102] D.J. Muller, A.C. Dumitru, C. Lo Giudice, H.E. Gaub, P. Hinterdorfer, G. Hummer, et al., Atomic force microscopy-based force spectroscopy and multiparametric imaging of biomolecular and cellular systems, Chem. Rev. 121 (19) (2021) 1170111725. [103] D. Alsteens, R. Newton, R. Schubert, D. Martinez-Martin, M. Delguste, B. Roska, et al., Nanomechanical mapping of first binding steps of a virus to animal cells, Nat. Nanotechnol. 12 (2) (2017) 177183. [104] M. Lekka, Discrimination between normal and cancerous cells using AFM, Bionanoscience 6 (1) (2016) 6580. [105] R. Li, H. Ye, W. Zhang, G. Ma, Y. Su, An analytic model for accurate spring constant calibration of rectangular atomic force microscope cantilevers, Sci. Rep. 5 (2015) 15828. [106] J.L. Hutter, J. Bechhoefer, Calibration of atomic-force microscope tips, Rev. Sci. Instrum. 64 (7) (1993) 18681873. [107] D. Kirmizis, S. Logothetidis, Atomic force microscopy probing in the measurement of cell mechanics, Int. J. Nanomed. 5 (2010) 137145. [108] M. Li, N. Xi, Y. Wang, L. Liu, Nanotopographical surfaces for regulating cellular mechanical behaviors investigated by atomic force microscopy, ACS Biomater. Sci. Eng. 5 (10) (2019) 50365050. [109] M. Li, L. Liu, X. Xiao, N. Xi, Y. Wang, Viscoelastic properties measurement of human lymphocytes by atomic force microscopy based on magnetic beads cell isolation, IEEE Trans. Nanobiosci. 15 (5) (2016) 398411. [110] M. Li, L. Liu, N. Xi, Y. Wang, Nanoscale monitoring of drug actions on cell membrane using atomic force microscopy, Acta Pharmacol. Sin. 36 (7) (2015) 769782. [111] W.C. Oliver, G.M. Pharr, Measurement of hardness and elastic modulus by instrumented indentation: advances in understanding and refinements to methodology, J. Mater. Res. 19 (1) (2004) 320. [112] M. Radmacher, Measuring the elastic properties of living cells by the atomic force microscope, Methods Cell Biol. 68 (2002) 6790. [113] S. Kasas, G. Longo, G. Dietler, Mechanical properties of biological specimens explored by atomic force microscopy, J. Phys. D. Appl. Phys. 46 (13) (2013) 133001. [114] N. Gavara, A beginner’s guide to atomic force microscopy probing for cell mechanics, Microsc. Res. Tech. 80 (1) (2017) 7584. [115] C. Rotsch, K. Jacobson, M. Radmacher, Dimensional and mechanical dynamics of active and stable edges in motile fibroblasts investigated by using atomic force microscopy, Proc. Natl. Acad. Sci. USA 96 (3) (1999) 921926. [116] N. Gavara, R.S. Chadwick, Determination of the elastic moduli of thin samples and adherent cells using conical atomic force microscope tips, Nat. Nanotechnol. 7 (11) (2012) 733736. [117] M. Li, L. Liu, N. Xi, Y. Wang, X. Xiao, W. Zhang, Effects of temperature and cellular interactions on the mechanics and morphology of human cancer cells investigated by atomic force microscopy, Sci. China Life Sci 58 (9) (2015) 889901. [118] A.R. Harris, G.T. Charras, Experimental validation of atomic force microscopybased cell elasticity measurements, Nanotechnology 22 (34) (2011) 345102. [119] T.M. Squires, T.G. Mason, Fluid mechanics of microrheology, Ann. Rev. Fluid Mech. 42 (2010) 413438. [120] M. Li, L. Liu, X. Xiao, N. Xi, Y. Wang, Effects of methotrexate on the viscoelastic properties of single cells probed by atomic force microscopy, J. Biol. Phys. 42 (4) (2016) 551569. [121] M. Li, L. Liu, N. Xi, Y. Wang, Atomic force microscopy studies on cellular elastic and viscoelastic properties, Sci. China Life Sci 61 (1) (2018) 5767. Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com References 41 [122] M. Li, L. Liu, X. Xu, X. Xing, D. Dang, N. Xi, et al., Nanoscale characterization of dynamic cellular viscoelasticity by atomic force microscopy with varying measurement parameters, J. Mech. Behav. Biomed. Mater. 82 (2018) 193201. [123] J. Helenius, C.P. Heisenberg, H.E. Gaub, D.J. Muller, Single-cell force spectroscopy, J. Cell Sci. 121 (11) (2008) 17851791. [124] D. Dang, R. Xiang, B. Liu, X. Liu, M. Li, Quantifying the adhesion forces of lymphoma cells by AFM single-cell force spectroscopy, Prog. Biochem. Biophys. 46 (1) (2019) 8998. [125] J. Friedrichs, J. Helenius, D.J. Muller, Quantifying cellular adhesion to extracellular matrix components by single-cell force spectroscopy, Nat. Protoc. 5 (7) (2010) 13531361. [126] R. Schubert, N. Strohmeyer, M. Bharadwaj, S.P. Ramanathan, M. Krieg, J. Friedrichs, et al., Assay for characterizing the recovery of vertebrate cells for adhesion measurements by single-cell force spectroscopy, FEBS Lett. 588 (19) (2014) 36393648. [127] M. Li, X. Xiao, L. Liu, N. Xi, Y. Wang, Z. Dong, et al., Nanoscale mapping and organization analysis of target proteins on cancer cells from B-cell lymphoma patients, Exp. Cell Res. 319 (18) (2013) 28122821. [128] D.J. Muller, Y.F. Dufrene, Force nanoscopy of living cells, Curr. Biol. 21 (6) (2011) R212R216. [129] E.L. Florin, V.T. Moy, H.E. Gaub, Adhesion forces between individual ligandreceptor pairs, Science 264 (5157) (1994) 415417. [130] G.U. Lee, D.A. Kidwell, R.J. Colton, Sensing discrete streptavidin-biotin interactions with atomic force microscopy, Langmuir 10 (2) (1994) 354357. [131] S. Allen, X. Chen, J. Davies, M.C. Davies, A.C. Dawkes, J.C. Edwards, et al., Detection of antigen-antibody binding events with the atomic force microscope, Biochemistry 36 (24) (1997) 74577463. [132] X. Zhang, D.F. Bogorin, V.T. Moy, Molecular basis of the dynamic strength of the sialyl lewis X-selectin interaction, ChemPhysChem 5 (2) (2004) 175182. [133] P. Hinterdorfer, W. Baumgartner, H.J. Gruber, K. Schilcher, H. Schindler, Detection and localization of individual antibody-antigen recognition events by atomic force microscopy, Proc. Natl. Acad. Sci. USA 93 (8) (1996) 34773481. [134] P. Hinterdorfer, Y.F. Dufrene, Detection and localization of single molecular recognition events using atomic force microscopy, Nat. Meth. 3 (5) (2006) 347355. [135] L. Wildling, B. Unterauer, R. Zhu, A. Rupprecht, T. Haselgrubler, C. Rankl, et al., Linking of sensor molecules with amino groups to amino-functionalized AFM tips, Bioconjug. Chem. 22 (6) (2011) 12391248. [136] J. Zhang, L.A. Chtcheglova, R. Zhu, P. Hinterdorfer, B. Zhang, J. Tang, Nanoscale organization of human GnRH-R on human bladder cancer cells, Anal. Chem. 86 (5) (2014) 24582464. [137] T. Puntheeranurak, I. Neundlinger, R.K. Kinne, P. Hinterdorfer, Single-molecule recognition force spectroscopy of transmembrane transporters on living cells, Nat. Protoc. 6 (9) (2011) 14431452. [138] X. Zhang, X. Shi, L. Xu, J. Yuan, X. Fang, Atomic force microscopy study of the effect of HER 2 antibody on EGF mediated ErbB ligand-receptor interaction, Nanomedicine 9 (5) (2013) 627635. [139] T.P.J. Knowles, M. Vendruscolo, C.M. Dobson, The amyloid state and its association with protein misfolding diseases, Nat. Rev. Mol. Cell Biol. 15 (6) (2014) 385396. [140] D. Fotiadis, Atomic force microscopy for the study of membrane proteins, Curr. Opin. Biotechnol. 23 (4) (2012) 510515. [141] D.J. Muller, AFM: a nanotool in membrane biology, Biochemistry 47 (31) (2008) 79867998. [142] D.J. Muller, A. Engel, Atomic force microscopy and spectroscopy of native membrane protein, Nat. Protoc. 2 (9) (2007) 21912197. Atomic Force Microscopy for Nanoscale Biophysics Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com 42 1. Fundamentals and methods of atomic force microscopy for biophysics [143] T. Ando, T. Uchihashi, T. Fukuma, High-speed atomic force microscopy for nanovisualization of dynamic biomolecular processes, Prog. Surf. Sci. 83 (79) (2008) 337437. [144] D.J. Muller, Y.F. Dufrene, Atomic force microscopy: a nanoscopic window on the cell surface, Trends Cell Biol. 21 (8) (2011) 461469. [145] T. Ando, N. Kodera, E. Takai, D. Maruyama, K. Saito, A. Toda, A high-speed atomic force microscope for studying biological macromolecules, Proc. Natl. Acad. Sci. USA 98 (22) (2001) 1246812472. [146] T. Ando, High-speed atomic force microscopy coming of age, Nanotechnology 23 (6) (2012) 062001. [147] A.J. Katan, C. Dekker, High-speed AFM reveals the dynamics of single biomolecules at the nanometer scale, Cell 147 (5) (2011) 979982. [148] M. Shibata, H. Watanabe, T. Uchihashi, T. Ando, R. Yasuda, High-speed atomic force microscopy imaging of live mammalian cells, Biophys. Physicobiol. 14 (2017) 127135. [149] T. Ando, T. Uchihashi, S. Scheuring, Filming biomolecular processes by high-speed atomic force microscopy, Chem. Rev. 114 (6) (2014) 31203188. [150] T. Uchihashi, N. Kodera, T. Ando, Guide to video recording of structure dynamics and dynamic processes of proteins by high-speed atomic force microscopy, Nat. Protoc. 7 (6) (2012) 11931206. [151] T. Ando, High-speed AFM imaging, Curr. Opin. Struct. Biol. 28 (2014) 6368. [152] H. Watanabe, T. Uchihashi, T. Kobashi, M. Shibata, J. Nishiyama, R. Yasuda, et al., Wide-area scanner for high-speed atomic force microscopy, Rev. Sci. Instrum. 84 (5) (2013) 053702. [153] M. Li, L. Liu, N. Xi, Y. Wang, X. Xiao, W. Zhang, Imaging and measuring the biophysical properties of Fc gamma receptors on single macrophages using atomic force microscopy, Biochem. Biophys. Res. Commun. 438 (4) (2013) 709714. [154] C.M. Stroh, A. Ebner, M. Geretschlager, G. Freudenthaler, F. Kienberger, A.S.M. Kamruzzahan, et al., Simultaneous topography and recognition imaging using force microscopy, Biophys. J. 87 (3) (2004) 19811990. [155] L.A. Chtcheglova, P. Hinterdorfer, Simultaneous AFM topography and recognition imaging at the plasma membrane of mammalian cells, Semin. Cell Dev. Biol. 73 (2018) 4556. [156] S. Senapati, S. Lindsay, Recent progress in molecular recognition imaging using atomic force microscopy, Acc. Chem. Res. 49 (3) (2016) 503510. [157] S. Senapati, S. Manna, S. Lindsay, P. Zhang, Application of catalyst-free click reactions in attaching affinity molecules to tips of atomic force microscopy for detection of protein biomarkers, Langmuir 29 (47) (2013) 1462214630. [158] P. Kaur, Q. Fu, A. Fuhrmann, R. Ros, L.O. Kutner, L.A. Schneeweis, et al., Antibody-unfolding and metastable-state binding in force spectroscopy and recognition imaging, Biophys. J. 100 (1) (2011) 243250. [159] C. Stroh, H. Wang, R. Bash, B. Ashcroft, J. Nelson, H. Gruber, et al., Single-molecule recognition imaging microscopy, Proc. Natl. Acad. Sci. USA 101 (34) (2004) 1250312507. [160] R. Zhu, A. Rupprecht, A. Ebner, T. Haselgrubler, H.J. Gruber, P. Hinterdorfer, et al., Mapping the nucleotide binding site of uncoupling protein 1 using atomic force microscopy, J. Am. Chem. Soc. 135 (9) (2013) 36403646. [161] S. Lee, J. Mandic, K.J. Van Vliet, Chemomechanical mapping of ligand-receptor binding kinetics on cell, Proc. Natl. Acad. Sci. USA 104 (23) (2007) 96099614. [162] M. Duman, M. Pfleger, R. Zhu, C. Rankl, L.A. Chtcheglova, I. Neundlinger, et al., Improved localization of cellular membrane receptors using combined fluorescence microscopy and simultaneous topography and recognition imaging, Nanotechnology 21 (11) (2010) 115504. Microscopy for Nanoscale Biophysics Get all ChaptersAtomic ForForce Ebook Instant Download by email at etutorsource@gmail.com We Don’t reply in this website, you need to contact by email for all chapters Instant download. Just send email and get all chapters download. Get all Chapters For Ebook Instant Download by email at etutorsource@gmail.com You can also order by WhatsApp https://api.whatsapp.com/send/?phone=%2B447507735190&text&type=ph one_number&app_absent=0 Send email or WhatsApp with complete Book title, Edition Number and Author Name.