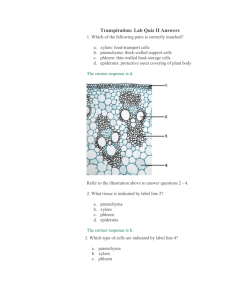
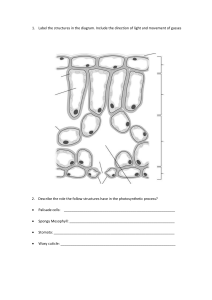
STUDY OF DIFFERENT TISSUE SYSTEMS The morphologically most easily and clearly recognizable units of the plant body are the cells. The united masses of cells are distinct from one another structurally as well as functionally. Such groupings of cells may be referred to as tissues which further may develop into a simpler or complex cellular organization. The arrangement of various tissues or tissue systems in the plant indicates its specialized nature. For example, vascular tissues are mainly concerned with the conduction of food and water, and for the efficient functioning; a complex network is developed with the places of water intake, sites of food synthesis and with areas of growth, development and storage. In the same way nonvascular tissues are also continually arranged which indicates the specific interrelationship of vascular tissues, storage tissues and supportive tissues. Plant tissues are generally categorized in to two categories. Ground Tissues Vascular Tissues Dermal tissues Differences between Meristematic and Permanent tissues In the plant body, the following three tissue systems can be distinguished. a) Dermal tissue system: It represents the outer most part of the plant which forms a protective covering line. It includes epidermis, periderm, etc. b) Vascular tissue system: It is concerned with trans-mission of material in the plant and represents stelar structures like xylem and phloem. c) Ground tissue system: It consists of simple cells which may be strengthened by addition of thickened cells. It represents ground tissue made up of parenchyma, collenchyma and sclerenchyma. Dermal Tissue System Epidermis The epidermal tissue system is derived from the dermatogen of the apical meristem and forms the epidermis (epi - upon, derma - skin) or outermost skin layer, which extends over the entire surface of the plant body. Epidermis is the outermost layer of the plant consisting normally of a single layer of flattened cells. The walls may be straight, wavy or beaded and often covered with a layer of cuticle made up of cutin. Functions 1. The primary function of the epidermis is protection of the internal tissues against mechanical injury, excessive heat or cold, fluctuations of temperature, attacks of parasitic fungi and bacteria, and against the leaching effect of rain. This is possible due to the presence of cuticle, hairs, tannin, gum, etc. 2. Prevention of excessive evaporation of water from the internal tissues by the development of thick cuticles, wax and other deposition, cutinized hairs, scales, multiple epidermis, etc., is another important function of the epidermis. 3. Strong cuticles and cutinized hairs, particularly a dense coating of hairs, protect the plant against intense illumination (i.e. strong sunlight) and excessive radiation of heat. 4. The epidermis also acts as a storehouse of water, as in desert plants. 5. The epidermis sometimes has some minor functions like photosynthesis, secretion, etc. Stomata Stomata are minute openings usually found in the epidermis of the leaves as in Digitalis, Senna, etc., or in young green stems as in Ephedra, in flower as in clove and in fruit as in fennel, orange peel. These openings are surrounded with a pair of kidney-shaped cells called guard cells. The term ‘stoma’ is often applied to the stomatal arrangement, which consists of slit like opening along with the guard cells. The epidermal cells surrounding the guard cells are called neighbouring cells or subsidiary cells. These, in many cases, as in Digitalis resemble the other epidermal cells, but in large number of plants they differ in size, arrangement and shape from the other epidermal cells. Types of stomatal arrangement: According to the arrangement of the epidermal cells surrounding the stomata, they have been grouped as follows: 1. Diacytic or Caryophyllaceous (cross celled): The stoma is accompanied by two subsidiary cells, the long axis of which is at right angles to that of the stoma. This type of stoma is also, called the Labiatae type as it is found in many plants of the family Labiatae such as vasaka, tulsi, spearmint and peppermint. 2. Anisocytic or Cruciferous (unequal celled): The stoma is surrounded by usually three subsidiary cells of which one is markedly smaller than the others. This type of stoma is also called the Solanaceous type as it is found in many plants of the family Solanaceae, such as Belladonna, Datura, Hyoscyamus, Stramonium, Tobacco; it is also found in many plants of the family Compositae. 3. Anomocytic or Ranunculaceous (irregular celled): The stoma is surrounded by a varying number of cells in no way differing from those of the epidermal cells as in Digitalis, eucalyptus, henna, lobelia, neem, etc. 4. Paracytic or Rubiaceous (parallel celled): The stoma is surrounded usually by two subsidiary cells, the long axis of which are parallel to that of stoma as in Senna and many Rubiaceous plants. 5. Actinocytic (radiate celled): The stoma is surrounded by circle of radiating cells, as in Uva ursi. Functions and distributions of stomata: Stomata perform the function of gaseous exchange and transpiration in the plant body. They are most abundant in the lower epidermis of a dorsiventral leaf and less abundant on the upper epidermis. In isobilateral leaves, stomata remain confined to the upper epidermis alone; in submerged leaves no stoma is present. In Buchu and Neem, stomata are present only on lower surface, while in case of Belladonna, Datura, Senna, etc.; stomata are present on the both surfaces. The distribution of stoma shows great variation between upper and lower epidermis. In desert plants and in those showing xerophytic adaptations, e.g. Ephedra, Agave, Oleander, etc., stomata are situated in grooves or pits in the stem or leaf. This is a special adaptation to reduce excessive evaporation, as the stomata sunken in pits are protected from gusts of wind. Trichomes Trichomes are more elongated outgrowths of one or more epidermal cells, and consist of two carts, a foot or root embedded in the epidermis and a free projecting portion termed as body. Trichomes usually occur in leaves but are also found to be present on some other parts of the plant as in Kurchi, Nux vomica and Strophanthus seeds, Andrographis and Belladonna stem, Cummin, and Lady’s finger fruits, etc. Trichomes are rarely present on the leaves of Bearberry, Buchu, Henna, etc., and are absent in glabrous leaves like Coca, Hemlock, Savin, etc. Functions of trichomes: Trichomes or hairs are adapted to many different purposes. A dense covering of trichomes prevents the damage by insects and the clogging of stomata due to accumulation of dust. Trichomes also aid the dispersion of seeds of Milkweed (Asclepias) and Madar (Calotropis), which are readily scattered by wind. In Peppermint, Rosemary, Tulsi, etc., trichomes perform the function of secreting volatile oil. Types of trichomes: Broadly, the trichomes are classified as: 1. Covering trichomes or clothing hairs or nonglandular trichomes and 2. Glandular trichomes Depending upon the structure, shape and number of cells, they are further classified as follows: [A] Covering trichomes a) Unicellular trichomes 1) Linear, strongly waved, thick walled trichomes— Yerba santa 2) Linear, thick walled and warty trichomes— Damiana 3) Short. conical trichomes—Tea 4) Short, conical, warty trichomes—Senna 5) Large, conical, longitudinally striated trichomes— Lobelia 6) Long, tubular, flattened and twisted trichomes— Cotton 7) Lignified trichomes—Nux vomica, strophanthus 8) Short, sharp, pointed, curved, conical trichomes— Cannabis 9) Unicellular, stellate trichomes—Deutezia scabra b) Multicellular unbranched trichornes 1) Uniseriate, bicellular, conical—Datura 2) Biseriate—Calendula officinalis 3) Multiseriate—Male fern c) Multicellular branched trichomes 1. Stellate (star shaped)—Hamamelis, Kamala 2. Peltate (shield-like structure)—cascarilla 3. Candelebra (branched)—Rosemary, Verbascum thapsus 4. T-shaped trichomes—Pyrethrum Covering trichomes [B] Glandular trichomes a) Unicellular glandular trichomes 1. Sessile trichomes—Without stalk - Piper betel, Vasaka b) Multicellular glandular trichomes 1) Unicellular stalk with single spherical secreting cell at the apex— Digitalis purpurea 2) Uniseriate, multicellular stalk with single spherical cell at the apex— Digitalis thapsi 3) Uniseriate stalk and bicellular head—Digitalis purpurea 4) Multicellular, uniseriate stalk and multicellular head—Hyoscyamus 5) Biseriate stalk and biseriate secreting head— Santonica 6) Short, unicellular stalk and head formed by a rosette of two to eight clubshaped cells—Mentha 7) Multiseriate, multicellular cylindrical stalk and a secreting head of about eight radiating club-shaped cells—Cannabis Glandular trichomes Periderm In the stem and root of mature plant, the layers immediately below the epidermis (phellogen) divide and redivide. On the outside they form cork or phellem and on the inner side they form phelloderm. Phellem + Phellogen + Phelloderm = Periderm Periderm The cork cells are rectangular brick shaped or polygonal; phelloderm cells are mostly parenchymatous in nature. Lenticels are present in the periderm, especially in the bark of old plants which are similar in function to stomata. These are open pores with absence of guard cells. The cork cells are impregnated with a layer of suberin. The various types of cork cells are shown bellow. Various types of cork cell Vascular Tissue System This system consists of a number of vascular bundles which are distributed in the stele. The stele is the central cylinder of the stem and the root surrounded by the endodermis. It consists of vascular bundles, pericycle, pith and medullary rays. Each bundle is made up of xylem and phloem, with a cambium in dicotyledonous stems, or without a cambium in monocotyledonous stems, or only one kind of tissue xylem or phloem, as in roots. Function The function of this system is to conduct water and raw food material from the roots to the leaves, and prepared food material from leaves to the storage organs and the growing regions. The vascular bundle of a dicotyledonous stem, when fully formed, consists of three well-defined tissues: 1. Xylem or wood 2. Phloem or bast, and 3. Cambium. [1] XYLEM Xylem or wood is a conducting tissue and is composed of elements of different kinds, viz. (a) tracheids, (b) vessels or tracheae, (c) wood fibres and (d) wood parenchyma. Xylem, as a whole, is meant to conduct water and mineral salts upwards from the root to the leaf to give mechanical strength to the plant body. (a) Tracheids: These are elongated, tube-like cells with hard, thick and lignified walls and large cell cavities. Their ends are tapering, either rounded or chisel-like and less frequently, pointed. They are dead, empty cells and their walls are provided with one or more rows of bordered pits. Tracheids may also be annular, spiral, scalariform or pitted (with simple pits). In transverse section, they are angular— either polygonal or rectangular. Tracheids (and not vessels) occur alone in the wood of ferns and gymnosperms, whereas in the wood of angiosperms, they are associated with the vessels. Their walls being lignified and hard, their function is conduction of water from the root to the leaf. (a) Tracheids with bordered pits (b) Scalariform tracheid (b) Vessels or tracheae: Vessels are cylindrical, tube-like structures. They are formed from a row of cells placed end to end, from which the transverse partition walls break down. A vessel or trachea is, thus, a tube-like series of cells, very much like a series of water pipes forming a pipeline. Their walls are thickened in various ways, and vessels can be annular, spiral, scalariform, reticulate, or pitted, according to the mode of thickening. Associated with the vessels are often some tracheids. Vessels and tracheids form the main elements of the wood or xylem of the vascular bundle. They serve to conduct water and mineral salts from the roots to the leaves. They are dead, thick-walled and lignified, and as such, they also serve the mechanical function of strengthening the plant body. Different kinds of vessels (c) Xylem (wood) fibres: Sclerenchymatous cells associated with wood or xylem are known as wood fibres. They occur abundantly in woody dicotyledons and add to the mechanical strength of the xylem and of the plant body as a whole. (d) Xylem (wood) parenchyma: Parenchymatous cells are of frequent occurrence in the xylem, and are known as wood parenchyma. The cells are alive and generally thin walled. The wood parenchyma assists, directly or indirectly, in the conduction of water, upwards, through the vessels and the tracheids. It also serves to store food. [2] PHLOEM The phloem or bast is another conducting tissue, and is composed of the following elements: (a) sieve tubes, (b) Companion cells, (c) phloem parenchyma and (d) bast fibres (rarely). Phloem, as a whole, is meant to conduct prepared food materials from the leaf to the storage organs and growing regions. (a) Sieve tubes: Sieve tubes are slender, tube-like structures, composed of elongated cells which are placed end to end. Their walls are thin and made of cellulose. The transverse partition walls are, however, perforated by a number of pores. The transverse wall then looks very much like a sieve, and is called the sieve plate. The sieve plate may sometimes be formed in the side (longitudinal) wall. In some cases, the sieve plate is not transverse (horizontal), but inclined obliquely, and then different areas of it become perforated. A sieve plate of this nature is called a compound plate. At the close of the growing season, the sieve plate is covered by a deposit of colourless, shining substance in the form of a pad, called the callus or callus pad. This consists of carbohydrate, called callose. In winter, the callus completely clogs the pores, but in spring, when the active season begins, it gets dissolved. In old sieve tubes, the callus forms a permanent deposit. The sieve tube contains no nucleus, but has a lining layer of cytoplasm, which is continuous through the pores. Sieve tubes are used for the longitudinal transmission of prepared food materials—proteins and carbohydrates—downward from the leaves to the storage organs, and later upward from the storage organs to the growing regions. A heavy deposit of food material is found on either side of the sieve plate with a narrow median portion. A sieve tube in longitudinal section (b) Companion cells: Associated with each sieve lube and connected with it by pores is a thin-walled, elongated cell known as the companion cell. It is living and contains protoplasm and an elongated nucleus. The companion cell is present only in angiosperms (both dicotyledons and monocotyledons). It assists the sieve tube in the conduction of food. (c) Phloem parenchyma: There are always some parenchymatous cells forming a part of the phloem in all dicotyledons, gymnosperms and ferns. The cells are living, and often cylindrical. They store up food material and help to conduct it. Phloem parenchyma is, however, absent in most monocotyledons. (d) Bast fibres: Sclerenchymatous cells occurring in the phloem or bast are known as bast fibres. These are generally absent in the primary but occur frequently in the secondary phloem. [3] CAMBIUM This is a thin strip of primary meristem lying between the xylem and phloem. It consists of one or a few layers of thin-walled and roughly rectangular cells. Although cambial cells look rectangular in transverse section, they are very elongated, often with oblique ends. They become flattened tangentially, i.e. at right angles to the radius of the stem. Types of Vascular Bundles According to the arrangement of xylem and phloem, the vascular bundles are of the following types: (A) Radial vascular bundle: When the xylem and phloem form separate bundles which lie on different radii, alternat-ing with each other, as in roots. The radial vascular bundle is the most primitive type of vascular bundles. (B) Conjoint vascular bundle: When the xylem and phloem combine into one bundle, it is called as conjoint vascular bundle. There are different types of conjoint vascular bundles. (1) Collateral: When the xylem and phloem lie together on the same radius, the xylem being internal and the phloem external is called collateral. When cambium is present in collateral as in all dicotyledonous stems, the bundle is said to be open collateral, and when the cambium is absent, it is said to be closed collateral, as in monocotyledonous stems (2) Bicollateral: When the both phloem and cambium occur twice in a collateral bundle—once on the outer side of the xylem and again on the inner side of it, is called as bicollateral. The sequence is outer phloem, outer cambium, xylem, inner cambium and inner phloem. Bicollateral bundles are characteristics of Cucurbitaceae. They are also often found in Solanaceae, Apocynaceae, Convolvulaceae, Myrtaceae, etc. A bicollateral bundle is always open. (C) Concentric vascular bundle: When one kind of vascular tissue (xylem or phloem) is surrounded by the other is called as concentric vascular bundle. Evidently, there are two types, according to whether one is central or the other one is so. When the phloem lies in the centre and is surrounded by xylem, as in some monocotyledonous, the concentric bundle is said to be amphivasal (leptocentric). When, on the other hand, the xylem lies in the centre and is surrounded by phloem, the concentric bundle is said to be amphicribral (Hadrocentric). A concentric bundle is always closed. Ground Tissue System Ground tissue system is represented by the cortex, hypo-dermis, pith, mesophyll and portion of midrib of leaves and comprises of the following tissues. (a) Parenchyma The parenchyma consists of a collection of cells which are more or less isodiametric, that is, equally expanded on all sides. Typical parenchymatous cells are oval, spherical or polygonal. Their walls are thin and made of cellulose. They are usually living. Parenchymatous tissue is of universal occurrence in all the soft parts of plants. Its main function is storage of food material. When parenchymatous tissue contains chloroplasts, it is called chlorenchyma. Its function is to manufacture food material. A special type of parenchyma develops in many aquatic plants and in the petiole of banana. The wall of each such cell grows out in several places, like rays radiating from a star and is, therefore, stellate or star-like in general appearance. These cells leave a lot of air cavities between them, where air is stored up. Such a tissue is often called aerenchyma. (a) Parenchyma, (b) Chlorenchyma and (c) Aerenchyma (b) Collenchyma This tissue consists of somewhat elongated, parenchymatous cells with oblique, slightly rounded or tapering ends. The cells are much thickened at the corners against the intercellular spaces. They look circular, oval or polygonal in a transverse section of the stem. The thickening is due to a deposit of cellulose, hemicellulose and protopectin. Although thickened, the cells are never lignified. Simple pits can be found here and there in their walls. Their thickened walls have a high refractive index and, therefore, this tissue in section is very conspicuous under the micro-scope. Collenchyma is found under the skin (epidermis) of herbaceous dicotyledons, e.g. sunflower, gourd, etc., occurring there in a few layers with special development at the ridges, as in gourd stem. It is absent from the root and the monocotyledon, except in special cases. The cells are living and often contain a few chloroplasts. Being flexible in nature, collenchyma gives tensile strength to the growing organs, and being extensible, it readily adapts itself to rapid elongation of the stem. Since it contains chloroplasts, it also manufactures sugar and starch. Its function is, therefore, both mechanical and vital. (a) Collenchyma in transaction and (b) Collenchyma in longitudinal section (c) Sclerenchyma Sclerenchyma (scleros means hard) consists of very long, narrow, thick and lignified cells, usually pointed at both ends. They are fibre-like in appearance and hence, they are also called sclerenchymatous fibres, or simply fibres. Their walls often become so greatly thickened that the cell cavity is nearly obliterated. They have simple, often oblique, pits in their walls. The middle lamella is conspicuous in sclerenchyma. They are dead cells and serve a purely mechanical function, i.e. they give the requisite strength, rigidity, flexibility and elasticity to the plant body and thus enable it to withstand various strains. Sclereids: Sometimes, special types of sclerenchyma develop in various parts of the plant body to meet local mechanical needs. They are known as Sclereids or Stone cells. They may occur in the cortex, pith, phloem, hard seeds, nuts, stony fruits, and in the leaves and stems of many dicotyledons and also gymnosperms. The cells, though very thick-walled, hard and strongly lignified (sometimes cutinized or suberized), are not long and pointed like sclerenchyma, but are mostly isodiametric, polyhedral, short-cylindrical, slightly elongated, or irregular in shape. Usually, they have no definite shape. They are dead cells, and have very narrow cell cavities, which may be almost obliterated, owing to excessive thickness of the cell wall. They may be somewhat loosely arranged or closely packed. They may also occur singly. They contribute to the firmness and hardness of the part concerned. (a) Sclerenchymatous fi bres and (b) Sclereids (Stone cells) Cell Differentiation and Development Cell differentiation is only part of the larger picture of plant development. As plant organs develop (the process of organogenesis), the precursors of the tissue systems form in response to positional signals. Then, within each tissue system precursor, cell types must be specified in the proper spatial pattern. For instance, the spacing of trichomes and stomates within the protoderm must be specified before their precursor cells begin differentiation. Exchange of signals among neighboring cells is an important aspect of the processes of spatial patterning and cell differentiation. In addition, long distance signals are required so that the strands of xylem and phloem cells within the leaf vascular bundles connect perfectly with those in the stem. Examples of Cell Differentiation Trichomes: The distinctive branched unicellular trichomes of plants such as Arabidopsis differentiate from undistinguished precursor cells in the protoderm. These precursor cells initiate the differentiation pathway by undergoing deoxyribonucleic acid (DNA) synthesis without accompanying cytokinesis, so that trichome precursors typically have eight or sixteen times the amount of DNA of adjacent pavement cells. Next, trichome precursors begin cell expansion in the plane perpendicular to the epidermis, forming a tubular extension. Once this stalk is formed, the nucleus migrates from the base of the stalk to its tip, using the cell's cytoskeleton to pull it to a new location. The trichome then undergoes an unusual pattern of cell wall growth, in which the cell wall balloons out at three locations, forming the Trichrome differentiation. three trichome branches. When the trichome cell has reached its full size and shape, it adds thickness to its cell wall and deposits sharp crystals of calcium oxalate on the surface of the trichome, adding to its effectiveness in defense against herbivores (see Figure above). Vessel Elements. Vessel elements differentiate from cells of the procambium. Vessel elements are first differentiated from other procambial cells because they expand more than their neighbors. Vessel element precursors next begin to deposit the thickened, lignified parts of their cell walls in the ringlike, helical, netlike, or pitted pattern. The pattern can be predicted by the location of elements of the cytoskeleton within the cytoplasm that help guide wall precursor to the proper location. When cell wall synthesis is complete, special wall-degrading enzymes attack the end walls of the cell, forming the perforation between adjacent elements in a vessel. Finally, the vessel elements undergo programmed cell death. The cell makes protease enzymes and nuclease enzymes that reduce proteins and nucleic acids to their simple building blocks. Surrounding parenchyma cells absorb these small molecules, leaving an empty vessel (see Figure below). Vessel element differentiation. Bundle Sheath Cells. In most plants, the cells of the photosynthetic ground tissue are uniform in size, shape, and chloroplast development. Two types of photosynthetic parenchyma cells are sharply differentiated in plants that have the C4 photosynthetic pathway, however. These two cell types, the mesophyll and bundle sheath cells, begin differentiation as similar appearing ground meristem cells. During leaf expansion, the bundle sheath cells begin to enlarge first. The cell wall becomes thickened and impermeable to the diffusion of gases. Their plastids replicate, grow, and become asymmetrically placed within the cell. In contrast, the mesophyll cells undergo a minimal amount of enlargement and have thin, permeable cell walls. The number of plastids is low and the plastids remain small. During cell differentiation the genes encoding the enzymes of the C4 biochemical pathway are expressed exclusively in the mesophyll cells, whereas the genes encoding the enzymes of the C3 pathway are expressed only in the bundle sheath cells (see Figure below). Bundle sheath cell differentiation Hormonal Influences Many aspects of differentiation are controlled by hormones. The hormone auxin, for example, plays an important role in the differentiation of vessel elements, both in intact and wounded plants. This role was first demonstrated in experiments where small incisions were made in stem internodes that cut though the phloem and xylem of a single vascular bundle. Auxin produced by the apical meristem and young leaves above the wound induces parenchyma cells to regenerate the damaged vascular tissue. Parenchyma cells undergo transdifferentiation. Although they already had differentiated as parenchyma cells from ground meristem precursors, they now repeat the steps that procambial cells take when they differentiate as vessel elements. Cells are induced to do this in a chainlike pattern, so that a new continuous strand of vascular tissue is formed as a detour around the original incision. Scientists know that auxin is involved, since transdifferentiation is blocked when the sources of natural auxin (young leaves and buds) are removed or when auxin transport inhibitors are applied. If natural sources of auxin are removed, and artificial sources added, transdifferentiation of parenchyma cells will occur, regenerating the vascular bundle.