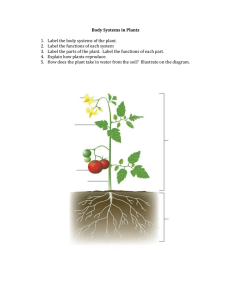
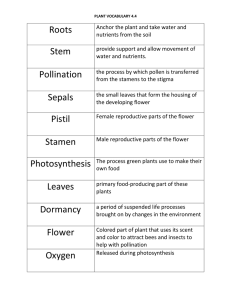
Lab 2 Flower Morphology and Plant Structure Overview In this lab you will have the opportunity to observe images of the structures (including flowers) for a reference strain of Arabidopsis thaliana plant and learn about the different functions for these structures. You will then compare these structures with those of a mutant Arabidopsis plant and make predictions about how these differences might impact how the mutant plant functions. Finally, you will receive published information on your mutant strain to determine if your predictions are correct. Objectives 1. Become familiar with plant growth and meristems, as well vegetative organs. 2. Become familiar with terms associated with plant structure, through lab exercises, as well as readings in textbook, lecture notes, and lab manual. Required Preparations 1. Read Lab 2 carefully before coming to your lab. 2. Read Chapter 27 and Chapter 29 in your textbook (Morris et al. 2019) and review relevant lecture notes. Lab 2 Exercises 1. Summary of plant organ function and observation of reference strain of Arabidopsis thaliana 2. Observation of mutant strain of Arabidopsis thaliana 3. Using internet resources for information on mutant strain of Arabidopsis thaliana 4. Complete Lab #2 Assignment on Quercus. This must be completed within 24 hours after the end of your scheduled lab period. Background Information: Plant Structure In this lab, you will have an opportunity to become familiar with some of the basic features of plant structure. Most of what you will learn relates primarily to vascular plants (primarily angiosperms), since these are the most prominent land plants currently found on the planet. The knowledge of plant structure is critical to understanding how plants function. To provide adequate context for this laboratory, the background information is divided into four subject areas: 1. 2. 3. 4. Growth and meristems Tissue systems Plant vegetative organs (leaf, stem, root) Plant reproductive organs (angiosperms only - flowers) 1. Growth and meristems For a plant to grow and develop, cell division, followed by cell enlargement, and finally cell differentiation are required. Unlike animals, plants may continue to grow throughout their lives due to the continued presence of regions of dividing cells. These regions are called meristems. There are two basic groups of meristems: (a) primary and (b) secondary meristems (Figure 2.1) (a) There are two primary meristems. Once is located at the tip or apex of the shoot and is called the shoot apical meristem. The second is at the tip or apex of the root and is called the root apical meristem. These two apical meristems give rise to all simple and complex tissues and tissue systems in the shoots and roots of a plant. (b) The secondary meristems have site-specific roles. See Figure 2.1 for names and locations of secondary meristems. Growth from the primary meristem is known as primary growth, generally resulting in a lengthening of root and shoot and in the addition of new plant parts. Figure 2.1. Primary and basic secondary meristems. Primary meristems consist of shoot apical meristem and root apical meristem, whereas secondary meristems include lateral shoot and root meristems, axial meristems, adventitious meristems, vascular cambium (e.g. in leaves; not indicated in the figure) or cork cambium (in the periderm; not indicated in the figure). From: http://vannocke.hrt.msu.edu/plb865/Meristem%20dynamics/meristems.html The leaves, stems, and roots of all monocotyledons and some eudicots have only primary growth. However, in all gymnosperms and some eudicots, a process known as secondary growth also occurs in stems and roots. This secondary growth is due to the appearance of two new lateral meristems, the vascular cambium and the cork cambium. Secondary growth from these regions results in an increase in the diameter of shoots and roots. Some plant parts can continue to grow throughout a plant’s lifetime. For example, the growth of roots and shoots over long periods of time shows such indeterminate growth. Determinate growth is growth that stops at a particular point. This is characteristic of most animals and of some plant parts such as leaves, flowers, and fruit. 2. Plant tissues In multicellular organisms, such as plants, cells often exist in communities. A biological tissue is a community of cells that work together to perform a specific function. In plants, ‘simple tissues’ are composed of just one cell type and include parenchyma, collenchyma, and sclerenchyma. Whereas ‘complex tissues’ are composed of more than one cell type and include the epidermis, periderm, xylem, and phloem. Plant tissues are organized into three basic tissue systems (Figure 2.2), based on their function: a. Dermal tissue system – This tissue system consists of the epidermis and periderm tissues. It provides an outer covering for the plant. b. Vascular tissue system – This tissue system is specialized for the transport of different substances from one part of the plant to another and consists of both xylem and phloem tissue. Among plants, only vascular plants (pteridophytes, gymnosperms, and angiosperms) contain the vascular tissue system. Non-vascular plants (bryophytes) lack this tissue system. c. Ground tissue system – This tissue system serves largely as a storage organ or as a filler of space between the dermal and vascular tissue systems. Reproductive organs Dermal tissue Shoot Leaf Stem Ground tissue Vascular tissue Root s Figure 2.2. The tissue systems and vegetative organs of plants. From Morris (2019). 3. Plant vegetative organs In vascular plants, the three tissue systems of a plant are organized into four major organ types: leaves, reproductive organs, stems, and roots (Figure 2.4). The leaf, stem, and reproductive organs form the shoot of the plant. Reproductive organs are covered in more detail later in this document. a. Leaves (Figure 2.3) The primary function of a leaf is to perform photosynthesis. There are two zones of photosynthetic cells, or cells that contain a large number of chloroplasts, within the ground tissue of a leaf: the palisade mesophyll and the spongy mesophyll. The palisade mesophyll consists of elongate cells found below the upper epidermis. The spongy mesophyll consists of irregular shaped cells found below the palisade mesophyll. Compared with the palisade mesophyll, spongy mesophyll cells are very loosely packed creating a large volume of intercellular airspace within this part of the leaf. Gas exchange for photosynthesis is dependent on this cell arrangement, as well as on the presence of pores called on the upper and more so on the lower surface of the leaf. These pores are called stomata (singular = stoma). The pair of cells that surround these openings, the guard cells, will change shape in response to environmental stimuli and cause the stomata to open and close. The dermal tissue system or epidermal cells are found on both the upper and lower surfaces of the leaf. To help mitigate water loss, the epidermal cells are covered by a waxy layer called the cuticle. Leaves may also possess tiny hairs called trichomes. These trichomes can be found on other parts of the shoot including the stem and sepals. The function of these tiny hairs to protect the plant from harmful UV light and herbivores. The vascular tissue system (xylem and phloem) is found in the veins of the leaf. The veins are enclosed by a layer of a cells known as the bundle sheath cells. The pattern of veins (leaf venation) facilitates close contact between the vascular tissue and the photosynthetic mesophyll cells. This arrangement facilitates efficient movement of water into the leaf from the roots and movement of sucrose out of the leaf to other parts of the plant. Palisade mesophyll cells near the upper surface of the leaf have a columnar arrangement that maximizes light interception. Cuticle Upper epidermis Mesophyll cells ~40,000 ppm Cuticle Guard cell Stoma Vein H2O CO2 gradient Lower epidermis Figure 2.3. Typical leaf structure. Adapted from Morris (2019). Water vapor gradient CO2 Spongy mesophyll cells close to the stomata have a honeycomb-like arrangement to allow CO2 to spread easily throughout the leaf. b. Stems (Figure 2.4) The function of the stem is to elevate and position both leaves and reproductive organs. Similar to the leaf, the dermal tissue system surrounds the stem and provides protection from pathogens and against water loss. The vascular tissue system transports substances between the leaves and roots of the plant. Unlike roots, the vascular tissue of the stem is usually arranged into discrete strands called vascular bundles, which are oriented such that the xylem is closest to the center of the stem and the phloem is closest to the epidermis. The arrangement of vascular bundles may differ in monocots and eudicots. For example, the sunflower, a eudicot, has vascular bundles that are arranged in a single ring, while the vascular bundles of a monocot are scattered throughout the stem. The ground tissue of stems is made up of undifferentiated parenchyma cells, or pith. Xylem Phloem Epidermis Thin-walled parenchyma cells Figure 2.4. Typical leaf structure. Adapted from Morris (2019). Sclerenchyma c. Roots (Figures 2.5 and 2.6) Roots enable vascular plants to take up water and nutrients from the soil, as well as provide an anchor for the plant, especially for those that can grow to great heights. The root apical meristem will form a root cap at the tip of the root through mitosis. Cells of the root cap mature into parenchyma cells toward the tip. The root cap protects the root apical meristem as it pushes its way through the soil and is unique to root apical meristems. The shoot apical meristem does not have an equivalent structure. There are three zones of root development behind the root apical meristem in this order: (1) the zone of cell division, (2) the zone of cell elongation, and (3) the zone of cell differentiation. (1) Zone of Cell Division: The root apical meristem and the cells immediately behind it are actively dividing through mitosis. This cell division will give rise to the three tissue systems in the roots. (2) Zone of Cell Elongation: As the name implies, following cell division the cells of a root undergo a period of rapid cell lengthening. (3) Zone of Cell Differentiation: In this zone, the now fully expanded cells differentiate into particular cell types. In the order of outer cells to inner cells: I. Epidermis: This is the outer layer of cells that constitutes the dermal tissue system of the root. Many epidermal cells grow long extensions called root hairs. Root hairs increase the absorptive surface of the root, allowing efficient uptake of nutrients and water from the soil. II. Cortex: This is a region usually made up of thin-walled parenchyma cells involved in the storage of food within the root. III. Endodermis: This is a single layer of cells. The cell walls of these cells contain a band of suberin called the Casparian strip. Suberin provides a barrier that prevents any further movement of water or solute through the cell walls (apoplastic water transport) and instead forces these substances to move through the cell membranes (symplastic water transport). Although water has no problem moving through osmosis across cell membranes, the Casparian strip allows plants to be selective about which solutes are taken up from the soil. IV. Stele: This central region of the root contains the vascular tissue system. The three tissue types that make up the stele are the pericycle, xylem, and phloem. The pericycle is the outermost layers of cells in the stele. Some of these cells may become meristematic and form root meristems, which give rise to branch roots. The arrangement of xylem (tracheids and vessel elements) and phloem (sieve tube elements and companion cells) in the stele differs in monocots and eudicots. In monocots, the central region of the stele contains additional ground tissue called the pith, which consists of thinwalled parenchyma cells. Figure 2.6. Typical root structure (part 1). From Morris (2019). Xylem Phloem Root hairs Zone of cell differentiation Epidermis Cortex Pericycle Endodermis Root apical meristem Root cap Root apical meristem Root cap Figure 2.5. Typical root structure (part 1). From Morris (2019) Zone of cell elongation Zone of cell division 3 At the endodermis, the Casparian strip prevents ions and water from moving in the walls, forcing them to pass through cell membranes. 1 Ions that enter the cytoplasm of a root cell can move to the xylem through plasmodesmata. Endodermis Casparian strip Phloem Epidermis Xylem Cortex Plasmodesmata Ion Root hair 2 Or they can move in the water-filled spaces of the cell walls. Figure 2.6. Typical root structure (part 2 – water movement through endodermal cells). From Morris (2019) 4. Plant/Angiosperm reproductive organs A flower is the reproductive structure found in flowering plants (angiosperms). The biological function of a flower is to accomplish reproduction through the union of pollen with eggs. Angiosperms show remarkable variation in flower form and gender function. Because a single flower can contain both male AND female organs or be unisexual (male OR female sexual organs), and because a single plant individual can produce one to many flowers (with varying genders), angiosperms are characterized by a very wide range of reproductive systems. Depending on the gender distribution within or among flowers, flowers may facilitate outcrossing (fusion of pollen and egg from different individual plants of the same species) or allow selfing (fusion of pollen and egg from the same individual plant). This varied gender distribution and function in flowering plants is in stark contrast to reproduction in animals, in which the majority of animals are unisexual. Following pollination, the flower transforms into a seed containing fruit. Fruit both protect immature seeds from being consumed by animals and enhance seed dispersal, once seeds are mature. Flower morphology: Introduction of the basic flower morphology The basic organs (Box 2-1) in a flower are arranged in four whorls (Fig. 2.7): (1) Sepals in the outer-most whorl. The typical role of sepals is the protection of a developing flower. They come in a variety of shapes and sizes. Some sepals are long and thin while others are short and thick. Some sepals are individualized or free while others are fused together to make a cup formation around all other whorls of the flower (Fig. 2.8). (2) Petals in the second whorl. The typical role of petals in animal-pollinated flowers is to attract pollinators, hence petals often are large and colorful. Similarly, as in sepals, petals can be either free or fused (Fig. 2.8). (3) Male sexual organs in the third whorl. Male sexual organs are called stamens and are made up of filaments and anthers (with two pollen sacks) (Fig. 2.7). (4) Female sexual organ in the fourth and inner-most whorl. The female sexual organ is called the pistil. It is composed of the ovary (containing the ovule and the egg), style, and the stigma (Fig. 2.7). Angiosperms, or flowering plants, include approximately 275,000 species from a large number of taxonomic groups, of which the eudicots and monocots are the largest and most important. Representative plant species of these two groups differ in their basic flower characteristics (Fig. 2.9). Flower organs of eudicots (approximately 200,000 species) often come in multiples of 4s or 5s, whereas flowers of monocots (approximately 60,000 species) come in multiples of 3s. In monocots, sepals and petals are often indistinguishable in size, shape, and color and, in such cases, are referred to as tepals (Fig. 2.10). Note that often the numbers of pistils, and, less so, stamens, do not follow these basic eudicot and monocot organ multiples. Both eudicot and monocot flower organs are arranged in two basic symmetries: in radial (actinomorphic) or bilateral (zygomorphic) symmetry (Fig. 2.11; Box 2-1). Floral symmetry describes whether, and how, a flower can be divided into two or more identical or mirror-image parts. Radially symmetric flowers can be divided into three or more identical sectors, whereas bilaterally symmetric flowers can be divided by only a single plane into two mirror-image halves, much like a person's face. TERMS OF FLOWER MOPHOLOGY; BOX 2-1 Sepal The outer parts of the flower inserted in the first whorl. Sepals are often green and leaf-like and protect the developing bud. Petal Flower parts inserted in the second whorl. In animal-pollinated flowers, petals are often conspicuously colored and used to attract the pollinators. Tepal This term is applied when the sepals and petals are of similar shape and color, or undifferentiated. This is often the case in monocots. Stamen The pollen-producing or male part of a flower, usually with a slender filament supporting the anther. Stamens are inserted in the third whorl. Anther The structure at the top of a stamen where pollen is produced. It typically consists of two pollen sacks. Filament The part of a stamen that supports the anther; the stalk of a stamen. Pistil The ovule-producing or female part of a flower, inserted in the fourth or inner-most whorl of the flower. The pistil consists of a stigma, style, the ovary, ovule and egg(s). Stigma The part of the pistil where pollen is supposed to land and germinate. The stigma is thus the landing platform for the pollen and located on top of the style. Style Pillar-like stalk through which pollen tubes grow to reach the ovary. Ovary The enlarged basal portion or chamber of the pistil in which ovules are produced. The ovary bears ovules (unfertilized and immature seeds) and ripens into a fruit. Ovule The plant part that contains the embryo sac and hence the female germ cell (egg), which after fertilization and maturation develops into a seed. Radial Flower of this symmetry can be divided into three or more identical (actinomorphic) sectors which are related to each other by rotation about the center of symmetry the flower. Typically, each sector might contain one petal and one sepal or one tepal. Bilateral (zygomorphic) symmetry Flower of this symmetry can be divided by only a single plane into two mirror-image halves, much like a person's face. Zygomorphic flowers generally have petals of two or more different shapes, sizes, and colors. Pedicel (the stalk that supports the flower) Fig. 2.7. Schematic diagram of a flower and an example of an apple flower. Fig. 2.8. Free (individualized) or fused sepals and petals. Fig. 2.9. Typical eudicot and monocot flowers. Typically, the number of sepals and petals is indicative for whether a flower is a eudicot or a monocot, whereas the numbers of stamens and pistils are more likely to diverge from the basic numbers of 4s/5s and 3s. Fig. 2.10. If sepals and petals are indistinguishable (often the case in monocots), they are referred to as tepals. Left: schematic diagram of tepals. Mid, right: actual monocot flowers with indistinguishable tepals. Fig. 2.11. Radial or actinomorphic symmetry with more than one possible way to divide the flower in mirror-image parts (left) and bilateral or zygomorphic symmetry, in which a flower can only be divided by a single plane in two mirror-image halves (right). Exercise 1: Summary of plant organ function and observation of reference strain of Arabidopsis thaliana As you learned in Lab 1, Arabidopsis thaliana (Arabidopsis) was the first plant to have its genome completely sequenced. Although technically a weed, this plant has transformed into an important model system for plant research. Research experiments using Arabidopsis as a study species will often include a wild-type or reference strain. These strains have no or minimal mutations and therefore produce a wildtype or non-mutant phenotype. The images of the reference strain that you will be looking at today are of the Landsberg erecta (Ler-0) strain of Arabidopsis. This laboratory strain contains an X-ray induced mutation in the ERECTA gene, which causes the plants to have more upright growth. This mutation makes laboratory experiments easier for the researchers and does not affect any other part of the plant’s phenotype. For that reason Ler-0 is widely used to generate mutants, and serves as the reference strain for these mutants. 1. With your groups members in your breakout room, use the information in this lab, your textbook, and your lecture notes, fill in the column labelled ‘Function’ in the Lab 2 worksheet for all plant organs. Be as succinct as possible when filling out this table. 2. Carefully observe the images of the Landsberg erecta strain of Arabidopsis. With your groups members in your breakout room, fill in the column labelled ‘Observations of Reference Strain: Landsberg erecta’. Include as much detail as you can. Exercise 2: Observation of mutant strain of Arabidopsis thaliana Using various methods including X-ray, chemical mutagens, T-DNA inserts, as well as others, scientists have engineered a large number of mutant strains of Arabidopsis thaliana. These mutant strains often display a mutant phenotype due to a loss of function in one or more of the genes in their DNA. These mutant strains have allowed plant researchers to learn a great deal about plants and how they function. 1. Enter the name of the mutant strain in the heading of the column 2. Carefully observe the images of the mutant strain of Arabidopsis. With your groups members in your breakout room, fill in the column labelled ‘Observations of Mutant Strain’ in the Lab 2 worksheet. Include as much detail as you can. 3. After observing the mutant strain, predict how the phenotypic differences you observed would impact the function of the relevant organ in mutant. Complete the “Predictions as to how this would affect function of organ in mutant ” column in the worksheet. Exercise 3: Using internet resources for information on mutant strain of Arabidopsis thaliana 1. Follow the Arabidopsis Biological Resource Center link that is provided by your TA for the mutant strain you observed in Exercise 2. Paraphrase the phenotypic description given on this site for your mutant in complete sentences. Be sure to include an in-text citation and reference in CSE: Name Year format. For more information on this format please see this very comprehensive U of Guelph link: https://guides.lib.uoguelph.ca/CSENameYear/Basics The phenotypic information would be considered information from an ‘Organization/Group as Author’. 2. Does this phenotypic description match with your observations of your mutant strain? Describe how it does and does not as applicable. Exercise 4: Complete Lab #2 Assignment on Quercus After you have completed your online synchronous Lab 2, please complete Lab Assignment #2 on Quercus. This assignment will require you to interpret parts of the worksheet you completed. You will also be required to add to the paragraph you wrote in Exercise 3. You will have one three-hour attempt to complete this assignment. It must be completed 24 hours after the end of your scheduled Lab 2. No late assignments will be accepted