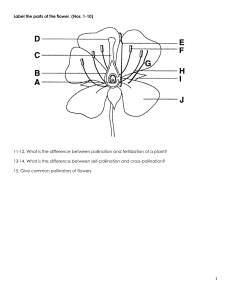
flowers")
Journal of ISSN 0972-1045 Vol. 18, No. 1, January-April, 2016 Applied Appl Hort Horticulture Journal of THE SOCIETY FOR ADVANCEMENT OF HORTICULTURE JOURNAL OF APPLIED HORTICULTURE Vol. 18, No. 1, Januar y-April, 2016 CONTENTS Further characterization of the action of pyridinedicarboxylic acids: multifunctional flower care agents for cut flowers of spray-type carnation —Shigeru Satoh, Yoshihiro Nomura, Shigeto Morita, and So Sugiyama (Japan) 3 Prohexadione-Ca provokes positive changes in the growth and development of habanero pepper —H. Ramírez, J. Mendoza-Castellanos, L.J. Ramírez-Pérez, J.H. Rancaño-Arrioja and M.G. Zavala-Ramírez (Mexico) 7 Assessing the influence of energy cost and other factors on profitability of greenhouse businesses in Tennessee —Hiren Bhavsar, Fisseha Tegegne and Krisden Ingram (USA) 12 Postharvest behaviour of minimally processed watercress —V.R. Logegaray, D. Frezza, A. Chiesa and A.P. León (Argentina) 16 Aroma profiling of jasmine (Jasminum sambac Ait.) flowers using electronic nose technology —X. Alex Isac, K.R. Rajadurai, M. Jawaharlal, K. Arul Mozhi Selvan, D. Uma, Hena Roy and Nabrun Bhattacharyya (India) 19 Genetic diversity assessment in Jasminum species using Amplified Fragment Length Polymorphism —K.S. Nirmala, B.V. Champa and A.P. Mallikarjuna Gowda (India) 25 Morphological characterization of herbaceous Phyllanthus spp. from Kerala —Shafna Kalarikkal, P.S. Udayan and M. Asha Sankar (India) 30 Optimizing the initial steps of immature endosperm culture of seeded banana (Musa sapientum L.) cultivar ‘Bhutia’ of Bangladesh —P.R. Paul, A.H.K. Robin and M.R. Hossain (Bangladesh) 34 A rapid screening method for salt stress tolerance of onion genotypes —Fatih Hanci, Esra Cebeci and Ayse Fidanci (Turkey) 39 A new and simple baiting technique for easy isolation of Phytophthora palmivora Butl. from bud rot affected tissue of coconut —K.M. Sharadraj and R. Chandra Mohanan (India) 44 Population distribution of Helicotylenchus species on Parkia biglobosa (Jacq) Benth and its association with some horticultural crops in Southern Guinea Savanna Ecological Zone of Nigeria —F.Y. Daramola (Nigeria) 48 Rose rootstocks position and auxins affect grafting take of ‘Inca’ —Millicent Adhiambo Otiende, Julius Omondi Nyabundi and Kamau Ngamau (Kenya) 54 Effect of plant density on mini-tuber production potential of potato varieties through micro-plants under net-house in North Eastern Himalayan region —A.K. Srivastava, S.K. Yadav, L.C. Diengdoh, R. Rai and T.K. Bag (India) 61 Assessment of genetic diversity in cucumber varieties using RAPD markers —Mahbube Someh, Ghaffar Kiani, Gholam Ali Ranjbar and Seyyed Mohamad Alavi (Iran) 64 Impact of seed priming on vigour in onion (Allium cepa L.) —C.N. Muruli, K. Bhanuprakash and B.C. Channakeshava (India) 68 Effect of chemicals and physical means on harvesting span, yield and quality of litchi (Litchi chinensis Sonn.) cv. Rose Scented —M. Pal, R.L Lal, P. Nautiyal and P. Joshi (India) 71 Media standardization for pre-hardening and hardening of in vitro regenerated plantlets of gerbera cultivars —S.K. Patra and S. Beura (India) 76 PCR mediated detection of sex and PaLCuV infection in papaya - A review —Sangeeta Saxena, Vijay K. Singh and Saurabh Verma (India) Journal of Applied Horticulture (http://horticultureresearch.net) 80 Journal Journal of Applied Horticulture, 18(1): 3-6, 2016 Appl Further characterization of the action of pyridinedicarboxylic acids: multifunctional flower care agents for cut flowers of spray-type carnation Shigeru Satoh1*, Yoshihiro Nomura1, Shigeto Morita2,3 and So Sugiyama2 Faculty of Agriculture, Ryukoku University, Otsu 520-2194, Japan. 2Graduate School of Life and Environmental Sciences, Kyoto Prefectural University, Kyoto 606-8522, Japan. 3Kyoto Prefectural Institute of Agricultural Biotechnology, Seika Town, Kyoto 619-0224, Japan. *E-mail: ssatoh@agr.ryukoku.ac.jp 1 Abstract Pyridinedicarboxylic acid (PDCA) analogs accelerate flower opening and retard senescence, which markedly extend the vase life of spray-type ‘Light Pink Barbara (LPB)’ carnation. In the present study, we characterized the activity of these chemicals to develop a novel flower care agent for a practical use. A representative PDCA analog 2,4-PDCA is effective in a wide range of spray-type carnation cultivars, ‘Barbara’, ‘Beam Cherry’, ‘Candle’, ‘Collin’, ‘Rascal Green’ and ‘Scarlet Ostara’, as well as ‘LPB’ and ‘Mule’. Treatment of ‘LPB’ flowers for the initial 24 h with 2,4-PDCA at 5 and 10 mM was almost as effective as the continuous treatment with the chemical at 2 mM. Key words: pyridinedicarboxylic acids, spray-type carnation, carnation cultivars, flower bud, time to flower opening, vase life, gross flower opening Abbreviations: ACC, 1-aminocyclopropane-1-carboxylate; FONS, fully-open and non-senescent; LPB, Light Pink Barbara; PDCA, pyridinedicarboxylic acid. Introduction Carnations are popular cut flowers and of highest economic importance in the floriculture industry in many countries. They are classified into standard and spray-type. The standard-type carnations have only one flower per stalk (stem), whereas the spray-type carnations have a main stalk with several offshoots, each having one or two flowers at the tip (inflorescence), making a plant with multiple flowers on a stalk, as a whole. During the senescence of carnation flowers, a climacteric increase in ethylene production occurs, and the evolved ethylene induces in-rolling of petals, resulting in wilting of whole flowers (Abeles et al., 1992; Satoh, 2011). The effect of ethylene on flower senescence can be diminished by treatment with inhibitors of ethylene biosynthesis or action, as discussed by Satoh et al. (2014). Treatment with these inhibitors prolongs the vase life of cut carnation flowers as well as other ethylene-sensitive ornamental flowers. Vlad et al. (2010) reported that 2,4-pyridinedicarboxylic acid (2,4-PDCA) inhibited ethylene production in detached flowers of ‘White Sim’, which is a standard-type carnation, and delayed senescence of the flowers. They hypothesized that 2,4-PDCA inhibited 1-aminocyclopropane-1-carboxylate (ACC) oxidase by competing with ascorbate, a co-substrate of the enzyme action. Then, Fragkostefanakis et al. (2013) showed that 2,4PDCA inhibited the in vitro activity of ACC oxidase prepared from tomato pericarp tissues. This observation supported the hypothesis that 2,4-PDCA inhibits ACC oxidase action by competing with ascorbate in carnation flowers. Satoh et al. (2014) confirmed that 2,4-PDCA inhibited ACC oxidase action using a recombinant enzyme produced in Escherichia coli cells from the carnation ACC oxidase gene (DcACO1 cDNA). They also showed that 2,4-PDCA treatment significantly prolonged the vase life of cut ‘Light Pink Barbara (LPB)’ and ‘Mule’ carnation flowers, both of which belong to the spray type, from the percentage of open flowers to the total number of initial flower buds (Satoh et al., 2005). Then, Sugiyama and Satoh (2015) evaluated the activity of 2,4-PDCA and its five analogs (2,3-, 2,5-, 2,6-, 3,4- and 3,5-PDCAs) to accelerate flower opening by determining the number of days to flower opening, in addition to their activity to extend the vase life of cut flowers of ‘LPB’ carnation. All six chemicals accelerated flower opening and extended vase life, although the effects varied with the chemical. They concluded that 2,3-PDCA and 2,4-PDCA were useful. Recently, Sugiyama et al. (2015) showed that PDCA analogs increased the number of open flowers according to their criterion ‘gross flower opening’. Satoh et al. (2014) suggested that gibberellin (GA) was involved in the promotive effect of 2,4PDCA on flower opening in cut flowers of spray-type carnation, because 2,4-PDCA is a structural analog of 2-oxoglutarate (2-OxoGA) (Vlad et al., 2010), and the latter acts as a co-substrate of enzymes responsible for GA biosynthesis and metabolism. The foregoing studies (Satoh et al., 2014; Sugiyama and Satoh, 2015; Sugiyama et al., 2015) demonstrated that PDCA analogs increased the number of open flowers, accelerated flower opening, and lengthened the vase life by retarding senescence, thereby markedly extending of the vase life of cut flowers of ‘LPB’ carnation. To the best of our knowledge, this is the first report of a chemical with multiple effects on cut carnation flowers. This novel agent, PDCA, may be applicable as a flower care agent in the near future. In the present study, we explored the applicability Journal of Applied Horticulture (http://horticultureresearch.net) 4 Pyridinedicarboxylic acids as flower care agents for cut flowers of spray-type carnation of 2,4-PDCA to cut flowers of spray-type carnation cultivars, including ‘LPB’ and ‘Mule’, and compared pulse treatment with continuous treatment. Materials and methods Carnation flowers: Flowers of seven cultivars of the spray-type carnation (Dianthus caryophyllus L.) were used, i.e., ‘Barbara’, ‘Beam Cherry’, ‘Candle’, ‘Collin’, ‘Light Pink Barbara (LPB)’, ‘Rascal Green’ and ‘Scarlet Ostara’. Flowers at the usual commercial stage of flowering, when the first flower out of six to eight flower buds on a stalk was partially open, were harvested with 65-cm-long stalks at the nursery of a commercial grower in Miyagi prefecture, Japan. The carnation flowers were harvested in the afternoon (in May and June 2015), immediately placed with their cut stalk ends in tap water, and sent in the next morning to the Faculty of Agriculture of Ryukoku University, Otsu city, Shiga prefecture, without supply of water during transportation. The flowers were not treated with any flower preservatives, including silverthiosulfate (STS), after harvest. Upon arrival the next day, the flowers were placed in plastic buckets with their cut stalk ends in tap water under continuous light from white fluorescent lamps (14 mmol m-2 s-1 PPFD) at 23 °C and 50-70% relative humidity, until experiments which were started several hours after they arrived. Determination of the days to flower opening, the vase life and the gross flower opening of carnation flowers treated with PDCA: Three samples (bunches) of 5 flower stalks (trimmed to 60-cm long), each having 5 flower buds (25 buds in total per sample), were put in 0.9 L glass jars with their stalk end in 300 mL of test solutions (one sample per glass jar). The flowers were kept under continuous light from white fluorescent lamps (14 mmol m-2 s-1 PPFD) at 23 °C and 50-70% relative humidity for 24 days or 30 days (‘Candle’). During this period the distilled water (control and the samples after pulse treatment) was replaced once a week, and test solutions were replenished as necessary in the continuous treatment. For pulse treatment (Fig. 2), flower samples were treated with 2,4-PDCA solutions at given concentrations for 24 h, and kept in distilled water thereafter. Fully-open and non-senescent (not wilted and turgid) flowers (FONS flowers), at flower opening stages Os 6 to Ss 2 (Harada et al., 2010; Morita et al., 2011), were counted daily and the percentage of these flowers to the total number (25) of initial flower buds per sample was calculated. Data are presented as changes of the percentages of FONS flowers during 24 days. Flower samples having 40% or more FONS flowers were regarded having display value. The vase life of flowers is expressed by the number of days during which the percentage of FONS flowers was 40% or more (Satoh et al., 2014). The time to flower opening was determined as the number of days from the start of the experiment to the time when the percentage of FONS flowers reached 40% (Sugiyama and Satoh, 2015). The gross flower opening was shown by the cumulative daily percentage at 40% or more of FONS flowers during incubation, which was shown by ‘scores’ as the unit (Sugiyama et al., 2015). The test solutions consisted of 2,4-PDCA at 0 (control), 2, 5, 10 and 30 mM dissolved in distilled water. The pH of these test solutions was not adjusted and 8-hydroxyquinoline sulfate at 100 mg L-1 was added to the solutions as a germicide. The stock solution of 100 mM 2,4-PDCA was made by dissolving 2,4-PDCA as Na-salt and adjusted at pH 7 with 1 M NaOH, and diluted with distilled water before use. Data are shown by the mean ±SE. 2,4-PDCA was purchased from Wako Pure Chemical Industries, Ltd., Osaka, Japan. Statistical analyses: Statistical analyses were carried out by Student’s t-test or Tukey’s multiple range test using an online statistical analysis program, MEPHAS (http://www.gen-info. osaka-u.ac.jp/testdocs/tomocom/, July 17, 2015). Values with P < 0.05 were considered significant. Results and discussion Effects of 2,4-PDCA on flower opening characteristics of different carnation cultivars: Figure 1 shows percentage of FONS flowers in cut ‘Barbara’ flowers treated continuously with 0 (control) or 2 mM 2,4-PDCA. Data are shown by those of three independent replicated samples. Each sample consisted of 5 flower stalks with 25 initial flower buds in total. Using this figure, we determined (1) the time to flower opening, (2) the vase life and (3) the gross flower opening which are shown by arrows with 1, 2 and 3. The treatment with 2 mM 2,4-PDCA tended to shorten the time to flower opening, lengthen the vase and increase the gross flower opening compared with the untreated control flowers. The same experiment as in Figure 1 was conducted using other cultivars, and the values for these three criteria for the flower opening profile are summarized in Table 1. The time to flower opening tended to be shortened by 2 mM 2,4-PDCA in all the cultivars tested, although it was significantly shortened in ‘Rascal Green’ cultivar. The vase life was significantly extended by the treatment with 2 mM 2,4-PDCA, in ‘Barbara’, ‘Candle’, ‘Collin’, and ‘Rascal Green’. The extension of the vase life was the greatest in ‘Collin’ cultivar (1.82 fold), followed by ‘Barbara’ (1.54 fold), ‘Candle’ Table 1. Effects of 2,4-PDCA on the time to flower opening, the vase life and the gross flower opening in cut flowers of various spray-type carnation cultivars Cultivars Barbara Candle Collin Beam Cherry Scarlet Ostara Rascal Green Time to flower opening (days) Control PDCA 3.7 ± 0.3 2.4 ± 0.3 2.4 ± 0.1 2.0 ± 0.2 2.8 ± 0.2 2.1 ± 0.2 6.3 ± 0.6 5.1 ± 0.5 3.0 ± 0.7 1.9 ± 0.2 9.5 ± 0.0 7.0** ± 0.5 Vase life (days) Control PDCA 11.2 ± 0.9 17.2** ± 0.5 16.5 ± 0.2 23.6** ± 0.2 10.0 ± 1.5 18.2** ± 0.8 10.8 ± 1.3 13.5 ± 0.9 13.1 ± 1.5 16.3 ± 0.9 9.8 ± 0.5 12.9** ± 0.1 Gross flower opening (scores) Control PDCA 126.7 ± 17.0 550.7** ± 26.6 429.3± 51.7 1138.7** ± 44.6 208.0 ± 31.7 713.3** ± 19.6 250.7 ± 40.7 548.0* ± 58.9 205.3 ± 19.2 534.7** ± 31.0 218.7 ± 33.7 480.0** ± 31.2 Data are shown by the mean ± SE of 3 replicated samples. * and ** show significant differences from the control by Student’s t-test at P < 0.05 and P < 0.01, respectively. Journal of Applied Horticulture (http://horticultureresearch.net) Fully-open and non--senescent flowers (%) Fully-open and non -senescent flowers (%) Pyridinedicarboxylic acids as flower care agents for cut flowers of spray-type carnation 100 80 60 1 3 40 2 20 0 0 5 15 20 10 Days after the start of treatment 25 5 100 80 60 40 20 0 0 5 10 15 20 25 Days after the start of treatment Fig. 1. Change in the percentage of fully-open and non-senescent flowers for cut flowers of ‘Barbara’ carnation treated with 2,4PDCA. Bunches of cut flowers, each with 5 main flower stalks with 5 flowers (buds) on each stalk (25 flower buds in total), were treated continuously with 0 (control) or 2 mM 2,4-PDCA. Three replicated data for the control (○, □, △) and 2,4-PDCA treatment (●, ■, ▲) are shown. The time to flower opening (1), the vase life (2) and the gross flower opening (3) are shown by arrows with numbers. Fig. 2. Effects of pulse and continuous treatments with 2,4-PDCA on flower opening characteristics of ‘Light Pink Barbara’ carnation. Bunches of cut flowers, similar to those described in the legend to Fig. 1, were treated with 2,4-PDCA at 0 (○), 5 (●), 10 (■) and 30 mM (◆) for 24 h, then kept in water (pulse treatment). The continuous treatment was conducted by leaving flower bunches continuously in 2 mM 2,4PDCA (□). Change in the percentage of fully-open and non-senescent flowers is shown by the mean of 4 replicated samples. (1.43 fold) and ‘Rascal Green’ (1.32 fold) cultivars. In ‘Beam Cherry’ and ‘Scarlet Ostara’, the vase life was extended by 2,4-PDCA treatment, 1.25 and 1.24 fold, respectively, though not significantly different with the control. The gross flower opening was increased by treatment with 2 mM 2,4-PDCA in all the carnation cultivars. The magnitude of increase in the gross flower opening was the largest with ‘Barbara’ (4.35 fold) followed by ‘Collin’ (3.43 fold) and 2.19 to 2.65 fold in other cultivars. being significantly different from the control (P < 0.05 by Tukey’s multiple range test). The continuous treatment with 2 mM 2,4PDCA shortened the time to flower opening to 4.3±0.3 days, though not significantly different with the control. Previous studies demonstrated the promotive effect of 2,4-PDCA treatment on flower opening in cut flowers of ‘LPB’ and ‘Mule’ (Satoh et al., 2014). The present findings further indicate that PDCA analogs, including 2,4-PDCA, promote flower opening and delay senescence in a wide range of spray-type carnation cultivars. Comparison of the effect of 2,4-PDCA between the continuous and pulse treatments: In previous studies (Satoh et al., 2014; Sugiyama and Satoh, 2015), PDCA was applied to cut carnation flowers continuously during experiments (the continuous treatment). This procedure sometimes caused detrimental side effects, resulting in browning of leaves or broken stalks (Sugiyama and Satoh, 2015), probably because of excess absorption of the chemicals. Also this procedure seems to be practically inadequate from application perspective of the chemicals, since it would need much labor work. Therefore, we tried to apply PDCA by a pulse treatment, in which the flowers were treated once after harvest for short period then kept in water. Fig. 2 shows the changes in the percentage of FONS flowers for cut ‘LPB’ flowers treated with 2,4-PDCA at 0, 5, 10 or 30 mM for 24 h, and thereafter left with their stalk end in water (pulse treatment). For comparison, the cut flowers were treated continuously with 2 mM 2,4-PDCA (continuous treatment). The mean of 4 replicated samples each for the control and the 2,4-PDCA-treated samples, each with 5 flower stalks having 25 initial flower buds in total, is shown. The time to flower opening was 6.0±0.5 days in the control. The pulse treatment with 5 and 10 mM 2,4-PDCA shortened it to 5.0±0.7 days and 3.1±0.4 days, respectively, the latter value The vase life in the control was 5.7±1.7 days. It was lengthened by the pulse treatment with 2,4-PDCA at 5 mM (11.9±0.7 days) and 10 mM (14.2±0.3 days), and by the continuous treatment with 2 mM 2,4-PDCA (11.2±0.5 days), with a significant difference from the control value. The gross flower opening score in the control was 94.0±35.7. The pulse treatment with 5 and 10 mM 2,4-PDCA increased the score to 335.0±28.8 and 495.0±8.5, respectively, and the continuous treatment with 2 mM 2,4-PDCA increased the score to 413.3±74.2. The pulse treatment with 30 mM 2,4-PDCA severely inhibited flower opening, resulting in browning and death of stems and leaves. The pulse treatment with 5 and 10 mM 2,4-PDCA did not have this adverse effect. The present results demonstrated that the effect of pulse treatment with 5 or 10 mM 2,4-PDCA on the flower opening characteristics was similar to or greater than that of the continuous treatment with 2 mM 2,4-PDCA in ‘LPB’ carnation. The present study revealed that 2,4-PDCA, probably other PDCA analogs as well, are effective on carnation cultivars, with spray-type flowers. These finding will help promote the practical use of PDCA analogs. Further studies are needed on the effect of PDCAs on cut flowers of other species with spraytype flowers, irrespective of the involvement of ethylene in the process of flower senescence, such as Eustoma, Gypsophila, and Alstroemeria flowers and spray-type Chrysanthemum. References Abeles, F.B., P.W. Morgan and M.E. Saltveit, Jr., 1992. Ethylene in Plant Biology. 2nd ed. Academic Press, San Diego, CA. Fragkostefanakis, S., P. Kalaitzis, A.S. Siomos and D. Gerasopoulos, 2013. Pyridine 2,4-dicarboxylate down regulated ethylene production in response to mechanical wounding in excised mature green tomato pericarp discs. J. Plant Growth Regul., 32: 140-147. Journal of Applied Horticulture (http://horticultureresearch.net) 6 Pyridinedicarboxylic acids as flower care agents for cut flowers of spray-type carnation Harada, T., Y. Torii, S. Morita, T. Masumura and S. Satoh, 2010. Differential expression of genes identified by suppression subtractive hybridization in petals of opening carnation flowers. J. Exp. Bot., 61: 2345-2354. Morita, S., Y. Torii, T. Harada, M. Kawarada, R. Onodera and S. Satoh, 2011. Cloning and characterization of a cDNA encoding sucrose synthase associated with flower opening through early senescence in carnation (Dianthus caryophyllus L.). J. Japan. Soc. Hort. Sci., 80: 358-364. Satoh, S. 2011. Ethylene production and petal wilting during senescence of cut carnation (Dianthus caryophyllus) flowers and prolonging their vase life by genetic transformation. J. Japan. Soc. Hort. Sci., 80: 127-135. Satoh, S., Y. Kosugi, S. Sugiyama and I. Ohira, 2014. 2,4-Pyridinedicarboxylic acid prolongs the vase life of cut flowers of spray carnations. J. Japan. Soc. Hort. Sci., 83: 72-80. Satoh, S., H. Nukui and T. Inokuma, 2005 A method for determining the vase life of cut spray carnation flowers. J. Appl. Hort., 7: 8-10. Sugiyama, S. and S. Satoh, 2015. Pyridinedicarboxylic acids prolong the vase life of cut flowers of spray-type ‘Light Pink Barbara’ carnation by accelerating flower opening in addition to an already-known action of retarding senescence. Hort. J., 84: 172-177. Sugiyama, S., S. Morita and S. Satoh, 2015. Three criteria for characterizing flower opening profiles and display values in cut spray-type carnation flowers. J. Appl. Hort.,17: 92-95. Vlad, F., P. Tiainen, C. Owen, T. Spano, F.B. Daher, F. Oualid, N.O. Senol, D. Vlad, J. Myllyharju and P. Kalaitzis, 2010. Characterization of two carnation petal prolyl 4 hydroxylases. Physiol. Plant., 140: 199-207. Received: October, 2015; Revised: November, 2015; Accepted: December, 2015 Journal of Applied Horticulture (http://horticultureresearch.net) Journal Journal of Applied Horticulture, 18(1): 7-11, 2016 Appl Prohexadione-Ca provokes positive changes in the growth and development of habanero pepper H. Ramírez1*, J. Mendoza-Castellanos1, L.J. Ramírez-Pérez1, J.H. Rancaño-Arrioja2 and M.G. Zavala-Ramírez1 Departamento de Horticultura, 2Dirección de Investigación, Universidad Autónoma Agraria Antonio Narro, Calz. Antonio Narro No. 1923, Saltillo, Coahuila, C.P. 25315. México. *E-mail: hrr_homero@hotmail.com 1 Abstract In recent years, the cultivated area of habanero pepper (Capsicum chinense Jacq.) has grown in México as a result of increasing the culinary diet among consumers and the knowledge on its high healthy components such as antioxidants, vitamins and nutrients. The actual worldwide demand of this vegetable requires the application of new production systems in order to increase yield per hectare as well as to improve the fruit quality of this commodity. The use of growth retardants is an alternative for this challenge, therefore, the effect of prohexadione-Ca (P-Ca) was evaluated on the vegetative growth, gibberellins in the stem apex, yield and antioxidants content in ripen fruits of habanero pepper cv. ‘Jaguar’. The dosages of P-Ca were: 0, 100, 175 and 250 mg L-1 sprayed to seedlings at one (10 days after transplanting) or two (10 and 31 days after transplanting) occasions. Results showed that P-Ca temporally reduced growth in height and diameter of main stem. This effect was related with a reduction in the synthesis of gibberellins A1, A4 and A7 in the apex. The fruit number and yield per plant increased with one application of P-Ca (at 175 mg L-1). The content of capsaicin and total carotenoids showed a remarkable increment in ripen fruits when plants have received one application of P-Ca at any concentration. Key words: Capsicum chinense Jacq., growth retardant, capsaicin, antioxidants, gibberellins. Introduction Vegetables such as habanero pepper (Capsicum chinense Jacq.) are a rich source of vitamins, minerals, fiber and antioxidants to humans. These elements in addition to cereals, grains and animal derived products, contribute to complete a dietary need for a healthy life. In recent years, medical research has established that fruits and vegetables given to patients through controlled diet provide protection to diseases such as cancer, arteriosclerosis, diabetes and liver injury (Charles, 2013). Under this expertise, the consumption of fruits and vegetables is highly advised in order to keep a good health and life quality (Mc Cormick, 2012). Habanero pepper is a crop with an increasing demand in the national and international markets. The fruit of this vegetable is gaining importance as a result of its high minerals, flavonoids and antioxidants content, in particular capsaicin. (Materska and Perucka, 2005; Ouzounidou et al., 2010). It is necessary to apply new alternative techniques which could contribute to increase both yield and fruit quality. On these bases, contemporaneous horticulture seeks technologies related to improve these referred components. Among them are: crop management systems, biotechnology and the use of bioregulators (Rademacher, 2000). Prohexadione calcium is a growth retardant used to control the excessive vegetative growth and to improve fruit quality in apple, pear and cherry trees (Costa et al., 2004). P-Ca inhibits the biosynthesis of the active gibberellins A1, A4, and A7 (Rademacher, 2000). Little is known upon the effects of P-Ca on vegetable crops. It has been suggested that P-Ca may participate in secondary metabolite pathways linked to antioxidant status in edible fruits (Mata et al., 2006; Rademacher and Kober, 2003; Roemmelt et al., 2003), as well as through modifying the enzyme system activity (Forkmann and Heller, 1999), reflected quite often in an increase in anthocyanin and color intensity in ripen grapes (Giudice et al., 2004). Therefore, the purpose of this work was to evaluate the effect of prohexadione-Ca on: the diameter and height of main stem; gibberellins at the apex; number of fruits and yield per plant and antioxidants content in ripen fruits of habanero pepper cv. ‘Jaguar’ under greenhouse conditions. Materials and methods Plant material and growing conditions: This research was conducted in a greenhouse facility at Universidad Autonoma Agraria Antonio Narro in Saltillo, Coahuila, Mexico. Seedlings of habanero pepper (C. chinense) cv. ‘Jaguar’ raised individually in peat moss and perlite (1:1) in black plastic boxes (Steiner, 1984), were used for this study. Treatment application: The growth retardant prohexadione-Ca at a concentrations of 0 (water-control), 100, 175 and 250 mg L-1 was applied with a back pack sprayer to seedlings at one (when the plants reached eight true leaves corresponding to 10 days after transplanting) or two (10 and 31 days after transplanting). All P-Ca solutions included 0.1% v/v Regulaid® as a surfactant. Horticultural evaluation: Height and diameter of main plant stem were evaluated every 14 days between time of P-Ca spray and the end of growing season. The number of fruits and yield per plant was recorded at each of the four harvested times. Endogenous gibberellins: Stem tips from control and P-Ca 175 mg L-1 plants were collected four days after the application of prohexadione-Ca. Removed samples were kept in liquid nitrogen, frozen, freeze-dried and ground. Later, tissue samples were Journal of Applied Horticulture (http://horticultureresearch.net) 8 Prohexadione-Ca provokes positive changes in the growth and development of habanero pepper analyzed for gibberellins using the gas chromatography-mass spectrometry (GCMS) technique (Ramirez et al., 2004). Purified extracts of tissue were dissolved in a few drops of methanol and methylated with diazomethane. A portion of a methylated extract was dissolved in pyridine and treated with trimethylchlorosilane and hexamethyl disilazane. Aliquots were examined using a pye 104 GLC coupled through a silicone membrane separator to an AEI MS30 dual beam mass spectrometer. Silanized glass columns (213 x 0.2 cm) were packed with 2% SE-33 on 80-100 Gas Chrom Q. The He-flow rate was 25 mL min-1, and the column temperature was programmed from 180 to 280 °C at 20/min at 280 °C . The MS were determined at 20 eV at a source temperature of 210 °C and a separator temperature of 190 °C with a scan speed of 6.5 sec. per mass decade. The spectra were recorded by a DEC Linc 8 computer. Capsaicin and carotenoids: The content of capsaicin in ripen fruits was determined at harvest time using the technique reported by Bennet and Kirby (1968), through which the antioxidant was extracted from fresh tissue utilizing a series of solvents and later measured in an spectrophotometer with 286 nm absorbance. Whilst total fruit carotenoids content was measured with the methodology of Tomas (1975). After organic solvent extraction and purification, the content of total carotenoids in the tissue sample was determined in the spectrophotometer at 454 nm and quantity of them established using the following formula: Fig. 1. Effect of prohexadione-Ca doses (mg L-1) on stem height growth in habanero pepper (C. chinense Jacq.) cv. ‘Jaguar’. Each point represents the mean of seven replicates. *Values statistically different at the Tukey´s P=0.05 level. A= Number of applications with P-Ca. µg carotenoids / 100 g fruit = % Abs × 3.857 × V ×100 / W Where: % Abs = percent of absorbance, V = measured volume in probet, and P = sample weight in grams. Experimental design and statistical analysis: A randomized factorial design with seven replicates plants per treatment was used. The results were analyzed with the PROC ANOVA (SAS 9.1, SAS Inst., Cary, NC). Significance was calculated using the Tukey´s method. Results and discussion Plant growth, gibberellins and yield: The application of P-Ca at all of the concentrations evaluated, resulted in a significant reduction (P=0.05) of stem growth on most measuring dates (Fig. 1). At the end of the growing season, plant height in P-Ca treated plants at 100 and 175 mg L-1 was similar to control; whereas those with the higher P-Ca concentration remained with lower growth. Fig. 2 shows that stem diameter increased between 65 and 100 days after transplanting in most P-Ca treatments. This increment was significantly noticeable (P=0.05) at the end of the stem growth in P-Ca sprayed plants with 175 and 250 mg L-1. The treatment with prohexadione-Ca at 175 mg L-1caused changes in the endogenous gibberellins status at the apex (Table 1). P-Ca samples showed GA9, GA20 and GA51; whilst in control tissue GA1, GA4 and GA7 were detected. The reduction in plant height and increment in stem diameter provoked by P-Ca (Figs. 1, 2) has also been observed in Mirador pepper (Ramirez et al., 2010) and apple trees (Costa et al., 2004). Prohexadione-Ca has been proven to be a strong retardant. This effect is explained in terms of its action as an inhibitor of the synthesis of gibberellins biologically active (Karhu and Hytönen, 2006; Rademacher, 2000). In this study, it was found that P-Ca inhibited at the apex the synthesis of gibberellins A1, A4 and A7 (Table 1), which are Fig. 2. Effect of prohexadione-Ca doses (mg L-1) on stem diameter growth in habanero pepper (C. chinense Jacq.) cv. ‘Jaguar’. Each point represents the mean of seven replicates.* Values statistically different at the Tukey´s P=0.05 level. A= Number of applications with P-Ca. required for shoot growth (Ramirez et al., 2003). This metabolic gibberellin inhibition remains for a few days and its synthesis is restored soon after (Rademacher, 2004); the behavior which may explain the recovery in stem growth of those P-Ca treatments seen at the end of the growing season (Fig. 1). The increment in stem diameter seen in P-Ca treated plants (Fig. 2) has been explained as a result of an increase in cell division followed by an increase of assimilate flux moving into that growing tissue (Ramirez et al., Journal of Applied Horticulture (http://horticultureresearch.net) Prohexadione-Ca provokes positive changes in the growth and development of habanero pepper 9 Table 1. Gibberellins in stem apices of habanero pepper (C. chinense Jacq.) cv. ‘Jaguar’ four days after being sprayed with P-Ca at 175 mg L-1 Gibberellins KRIa Principal ions and % relative intensity of base peak Control GA1 2651 [506(M+,100), 448(14), 377(15), 375(18)] GA4 2488 [418(M+,21), 403(2), 400(12), 386(25), 284(100)] GA7 2416 [416(M+,10), 193(12), 179(5), 155(13] Prohexadione-Ca 175 mg L-1 GA9 2295 [330(M+,5), 217(37), 183(19), 159(45)] GA20 2468 [418(M+,100), 403(17), 387(6), 375(82), 359(19)] GA51 2507 [418(M+,4), 403(3), 386(15),371(3), 358(1)] kovats retention index. M+ = Molecular ion. a 2003). The effect of prohexadione-Ca on fruits and yield per plant is shown in Table 2. The most remarkable effect on increase in yield per plant (P=0.05) was observed when the growth retardant was applied once at 175 mg L-1, where the fruit production per plant was 17% above control plants. Similar behavior was observed in the same treatment with respect to number of fruits per plant. The fact that P-Ca at 175 mg L-1 resulted in higher number of fruits and yield per plant, support the thesis that this given P-Ca concentration could be the optimum for habanero pepper plant under greenhouse conditions and also demonstrated for apple (Guak et al., 2004) and berries (Schildberger et al., 2011; Poledica et al., 2012). Other P-Ca concentrations used in this study resulted in low or too high doses for this habanero pepper commodity. The increment in fruit number and yield seen in P-Ca treated plants may reflect that the reduction in vegetative growth (Fig. 1) resulted in an increase in flower bud induction by the presence of more cytokinins in the meristematic tissue (Ramirez et al., 2010). This physiological condition also may promote more carbohydrates moving into developing fruitlets with a strong vascular connecting tissues which would avoid fruit drop (Costa et al., 2004; Jordan et al., 2001; Sridhar et al., 2009). Fig. 3. Effect of prohexadione-Ca doses (mg L-1) on capsaicin content in fruits of habanero pepper (C. chinense Jacq.) cv. ‘Jaguar’. Each column represents the mean of three replicates ± standard error. Values with the same letter are statistically similar according to Tukey test at P=0.05. Capsaicin and total carotenoids: The content of capsaicin increased significantly (P=0.05) in fruits collected from plants treated with any doses of prohexadione-Ca (Fig. 3). This effect was consistently higher when any concentration of P-Ca was sprayed only once. The highest increase in capsaicin occurred with P-Ca at 250 mg L-1; in which the amount of the antioxidant Table 2. Effect of prohexadione-Ca on fruit number and yield in habanero pepper (C. chinense Jacq.) cv. ‘Jaguar’ ProhexadioneYield per plant (g) Fruits per planty -1 Ca (mg L ) Applications 1 2 1 2 Control 764.13bz 760.03a 138.3b 143.75a 100 774.82b 698.88a 139.5b 175.95a 175 892.57a 541.01b 176.65a 109.85b 250 714.67b 401.54c 99.55b 84.4b VC 15.99 19.37 31.88 34.9 z VC: Variation coefficient. Within columns values with same letter are not statistically different at the 0.05% probability level using the Tukey´s test. yMean of seven plants. Fig. 4. Effect of prohexadione-Ca doses (mg L-1) on total carotenoids content in fruits of habanero pepper (C. chinense Jacq.) cv. ‘Jaguar’. Each column represents the mean of three replicates ± standard error. Values with the same letter are statistically similar according to Tukey test at P=0.05. was double when compared with control samples. Information on the effects of P-Ca on capsaicin in habanero pepper is scarce. It has been proposed that the increment in fruit antioxidants such as capsaicin after the application of P-Ca may be due to the ability of this growth retardant to inhibit the production of cellular free radicals (Díaz et al., 2004; Vázquez-Flota et al., 2007) which are normally produced during fruit ripening. This metabolic process requires the action of enzymes such as catalase and Journal of Applied Horticulture (http://horticultureresearch.net) 10 Prohexadione-Ca provokes positive changes in the growth and development of habanero pepper peroxidase as it has been demonstrated in apple (Rademacher, 2000). The content of total carotenoids in fruits also showed a significant increase when the P-Ca was applied only once at any dose and with 100 mg L-1 sprayed twice (Fig. 4). These values represent a two fold increase when compared with control fruits. This effect has also been observed in beans (Bekheta et al., 2009) and oranges (Graham and Smit, 2010). The promotion of carotenoids in the fruit by P-Ca could also be mediated through the hypothesis previously suggested for capsaicin. The increment in total carotenoids and capsaicin in fruits from P-Ca treated plants is an interesting contribution as an alternative to healthy food source since in recent years antioxidants consumption have been related to a high food quality (Da Silva et al., 2014; Mc Cormick, 2012), as well as for cancer, diabetes and heart diseases protection (Howard et al., 2000; Pramanik and Srivastava, 2013; Shaik et al., 2013). In conclusion, increased total carotenoids and capsaicin in fruits from P-Ca treated plants is an interesting contribution leading to high quality food production. More research is required to elucidate the mechanism of action by which higher carotenoid synthesis takes place as influenced by the growth retardant. Acknowledgements We thank Willbur Ellis Co. WA., USA for P-Ca donation. The authors Mendoza-Castellanos and Ramírez-Pérez thank CONACYT Mexico for grant. References Bekheta, M.A., M.T. Abdelhamid and A.A. El-Morsi, 2009. Physiological response of Vicia faba to prohexadione-calcium under saline conditions. Planta Daninha, 27(4): 769-779. Bennet, D.J. and G.W. Kirby, 1968. Constitution and biosynthesis of capsaicin. J. Chem. Soc. C., 442-446. Charles, D.J. 2013. Antioxidant Properties of Spices, Herbs, and other Sources. Springer Science Business Media, USA. 617 p. Costa, G., E. Sabatini, F. Spinelli, C. Andreotti, C. Bomben and G. Vizzotto, 2004. Two years of application of prohexadione-Ca on apple: effect on vegetative and cropping performance, fruit quality, return bloom and residual effect. Acta Hort., 653: 35-40. Da Silva Messias R., V. Galli, S.D. dos Anjos e Silva, and C.V. Rombaldi, 2014. Carotenoid biosynthetic and catabolic pathways: gene expression and carotenoid content in grains of maize landraces. Nutrients, 6(2): 546-563. Díaz, J., F. Pomar, A. Bernal and F. Merino, 2004. Peroxidases and the metabolism of capsaicin in Capsicum annuum L. Phytochem. Rev., 3: 141-157. Forkmann, G. and W. Heller, 1999. Biosynthesis of flavonoids. In: Comprehensive Natural Products Chemistry, Volume 1: Polyketides and Other Secondary Metabolites Including Fatty Acids and Their Derivatives. Barton D., K. Natkanishi and O. Meth-Cohn (eds.). Oxford, U.K.: Elsevier Science Ltd. p. 713-748. Giudice, D.L., T.K. Wolf and B.W. Zoecklein, 2004. Effects of prohexadione-calcium on grape yield components and fruit and wine composition. Am. J. Enol. Vitic., 55: 73-83. Graham, H.B. and L.R. Smit, 2010. Preharvest foliar sprays of prohexadione–calcium, a gibberellin-biosynthesis inhibitor, induce chlorophyll degradation and carotenoid synthesis in Citrus rinds. HortScience, 45: 242-247. Guak, S., M. Beulah and N.E. Looney, 2004. Controlling growth of super-spindle ‘Gala’/M.9 apple trees with prohexadione-calcium, and ethephon. Acta Hort., 653: 139-144. Howard, L.R., S.T. Talcott, C.H. Brenes and B. Villalon, 2000. Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum species) as influenced by maturity. J. Agric. Food Chem., 48: 1713-1720. Jordan, D.L., J.B. Beam, P.D. Johnson and J.F. Spears, 2001. Peanut response to prohexadione calcium in three seeding rate–row pattern planting systems. Agron. J., 93: 232-236. Karhu, S.T. and T.P.N. Hytönen, 2006. Nursery plant production controlled by prohexadione-calcium and mechanical treatments in strawberry cv. Honeoye. J. Hort. Sci. Biotech., 81: 937-942. Mata, A.P., J. Val and A. Blanco, 2006. Differential effects of prohexadione-calcium on red colour development in ‘Royal Gala’and ‘Fuji’apples. J. Hort. Sci. Biotech., 81: 84-88. Materska, M. and I. Perucka, 2005. Antioxidant activity of the main phenolic compounds isolated from hot pepper fruit (Capsicum annuum L.). J. Agric. Food Chem., 53: 1750-1756. McCormick, R. 2012. A Whole foods plant based health perspective, an opportunity for horticulture. Chron. Hort., 52(4): 5-9. Ouzounidou, G., I. Ilias, A. Giannakoula and P. Papadopoulou, 2010. Comparative study on the effects of various plant growth regulators on growth, quality and physiology of Capsicum annuum L. Pakistan J. Bot., 42(2): 805-814. Poledica-Milena, M., M. Milivojević-Jasminka, D. Radivojević-Dragan and J.J. Dragišić- Maksimović, 2012. Prohexadione-Ca and young cane removal treatments control growth, productivity, and fruit quality of the Willamette raspberry. Turk. J. Agric. For., 36: 680-687. Pramanik-Kartick, C. and S.K. Srivastava, 2013. Role of Capsaicin in Cancer Prevention. Springer Science+Business Media Dordrecht. Volume 3: 1-18. Rademacher, W. 2000. Growth retardants: effects on gibberellin biosynthesis and other metabolic pathways. Annu. Rev. Plant. Physiol. Plant Mol. Biol., 51: 501-531. Rademacher, W. 2004. Chemical regulation of shoot growth in fruit trees. Acta Hort., 653: 29-32. Rademacher, W. and L. Kober, 2003. Efficient use of prohexadione-Ca in pome fruits. Eur. J. Hort. Sci., 68: 107-107. Ramirez, H., A. Benavides and E. Rangel, 2004. Identification of gibberellins in seeds of a Golden Delicious apple mutant. Acta Hort., 653: 201-208. Ramírez, H., C. Amado, A. Benavides, V. Robledo and A. Osorio, 2010. Prohexadiona-Ca, AG3, anoxa y ba modifican indicadores fisiológicos en chile mirador (Capsicum annuum L.). Rev. Chapingo Ser. Hort., 16 (2): 83-89. Ramírez-Rodríguez, H., J.C. Gómez-Castañeda, A. Benavides-Mendoza, V. Robledo-Torres, L.I. Encina-Rodríguez and C.A. Coello-Cutiño, 2003. Influencia de prohexadiona-Ca sobre crecimiento vegetativo, producción y calidad de fruto en manzano. Rev. Chapingo Ser. Hort., 9: 279-284. Roemmelt, S., T.C. Fischer, H. Halbwirth, S. Peterek, K. Schlangen, J.B. Speakman, D. Treutter, G. Forkmann and K. Stich, 2003. Effect of dioxygenase inhibitors on the resistance-related flavonoid metabolism of apple and pears: Chemical, biochemical and molecular biological aspects. Eur. J. Hort. Sci., 68: 129-136. Schildberger, B., C. Faltis, M. Arnold and R. Eder, 2011. Effects of prohexadione-calcium on grape cluster structure and susceptibility to bunch rot (Botrytis cinerea) in cv. Grüner Veltliner. J. Plant Pathol., 93(1): S33-S37. Shaik-Dasthagirisaheb, Y.B., G. Varvara, G. Murmura, A. Saggini, A. Caraffa, P. Antinolfi, S. Tetè, M. Rosati, E. Cianchetti, E. Toniato, L. Speranza, A. Pantalone, R. Saggini, L.M. Di Tommaso, P. Conti, T.C. Theoharides and F. Pandolfi, 2013. Inhibitor effect of antioxidant flavonoids quercitin, and capsaicin in mast cell inflammation. Eur. J. Inflamm., 11(2): 353-35. Sridhar, G., R.V. Koti, M.B. Chetti and S.M. Hiremath, 2009. Effect of naphthaleneacetic acid and mepiquat chloride on physiological components of yield in bell pepper (Capsicum annuum L.). J. Agric. Res., 47: 53-62. Journal of Applied Horticulture (http://horticultureresearch.net) Prohexadione-Ca provokes positive changes in the growth and development of habanero pepper Steiner, A.A. 1984. The universal nutrient solution. Procceedings of the 6th International Congress Soilless Culture, International Society for Soilless Culture. PO Box 52, Wageningen, 1984, p. 633-649. Tomas, P. 1975. Effect of post-harvest temperature on quality carotenoids and ascorbic content of Alphonso mangoes on ripening. J. Food Sci., 40(4): 704-706. 11 Vázquez-Flota, F., M.L. Miranda-Ham, M. Monforte-González, G. Gutiérrez-Carbajal, C. Velázquez-García and Y. Nieto-Pelayo, 2007. La biosíntesis de capsaicinoides, el principio picante del chile. Rev. Fitotec. Mex., 30: 353-360. Received: May, 2015; Revised: July, 2015; Accepted: November, 2015 Journal of Applied Horticulture (http://horticultureresearch.net) Journal Journal of Applied Horticulture, 18(1): 12-15, 2016 Appl Assessing the influence of energy cost and other factors on profitability of greenhouse businesses in Tennessee Hiren Bhavsar*, Fisseha Tegegne and Krisden Ingram Department of Agricultural and Environmental Sciences, Tennessee State University, 3500 John A Merritt Blvd, Nashville, TN 37209, USA. *E-mail: hbhavsar@tnstate.edu Abstract The greenhouse industry is an important sub-sector of agriculture. On average, greenhouse and nursery farms in the U.S. have 57 percent more cash receipts than all farms (Muhammed, 2000). Tennessee cash receipts for greenhouse and nursery farms was 307 percent above than the average in the US. However, greenhouse operations in Tennessee have declined over the years. The goal of this study was to acquire a better understanding of energy use by greenhouse businesses in Tennessee. The latest database containing greenhouse businesses was provided by the Tennessee Department of Agriculture. A mailed survey was used to collect data that covered questions on energy use, marketing and skill needs of the businesses. The respondents were mixed in terms of size of their operations, education and income levels. Out of the 279 surveys, 56 were returned, resulting in a 20 percent response rate. To determine the different factors affecting profitability of greenhouse operations, this study utilized correlation and chi-square tests using different variables. Results indicate that profitability of greenhouse operations is influenced by the rising energy cost, economic downturn and size of operation. The survey also indicates that growers would consider adopting alternative energy saving methods depending on their income and age. The study shows the need to assist the growers in learning more about alternative energy saving methods and technologies. This study is beneficial not only for greenhouse businesses but also other stakeholders including policy makers and those working with growers. Other researchers can also undertake similar studies using the approach used here with appropriate modification. Key words: Greenhouse, energy use, middle Tennessee, profitability Introduction The greenhouse and nursery industries account for about 2.5 percent of all United States farms. According to the Census of Agriculture -by USDA, in 2012 - there were less than 100,000 greenhouse/nurseries in North America. Over the past few decades, there have been changes in the structure, conduct, and performance of the U.S. nursery and greenhouse industry. These industries can be very profitable for farmers and their communities. Yet, because of rise in energy costs, growers face the challenge of keeping their businesses profitable. One of the lessons learned over the years is that it is generally less expensive to consider energy conservation measures before implementing significant modifications to your heating system. Greenhouses in particular are harder to keep up, due to the many costs involved. The greenhouse structure is built to maintain proper temperature and climate for better gardening. This allows growers to extend their gardening season by growing plants inside when outside weather conditions are not ideal. Structures range from small to large buildings in an industry that has been around for more than a hundred years. Most greenhouses are seasonal businesses with maximum production in the spring. In Tennessee, it was reported that greenhouses must be heated from around October through April because Tennessee’s weather can be very unpredictable (Trainer, 2010). Surveys have examined the present business climate, but little has been done to understand what types of changes are taking place and whether or not these changes are regional in nature (Trainer, 2010). A large number of farmers depend on this important enterprise for their livelihood. But in recent years the business has been facing challenges due to the economic downturn, decline in demand for many of the products and high-energy costs. When oil prices are high, a typical commercial greenhouse operation would experience significant difficulty making a profit, and the industry would be in peril (Hall, 2006). Researchers have been searching for easier ways for greenhouse growers to save money on energy. Record low oil prices following the early energy crisis have shifted much of the attention away from increased self-reliance and the development of alternative energy sources. The results of the research efforts in the 1970s and 1980s led to substantial reductions in greenhouse energy use. It is important to identify not only the annual or seasonal expenses, but also the peak energy consumption amount (Stegelin, 2007). During the winter/ summer months, greenhouse growers spend a lot on energy costs. The greenhouse industry is one of the biggest sub-sectors in terms of economic output in the United States (Hall et al., 2006). The purpose of this study was to assess the benefit of considering alternative technologies for greenhouse businesses in Tennessee. Some of the specific objectives were to characterize current energy use, to examine consideration of alternative sources of energy and to assess the effect of energy costs on profitability by greenhouse businesses in Tennessee. Materials and methods For this study, primary data was collected from greenhouse operators. Certified growers from the Tennessee Department of Agriculture (2012) database were sent surveys in mail. There were 279 certified greenhouse growers on that list; in which Journal of Applied Horticulture (http://horticultureresearch.net) Influence of energy cost and other factors on profitability of greenhouse businesses in Tennessee we chose all to be surveyed. The survey focused on four major parts and asked general questions about the greenhouse such as the size, years in business, future plans, and other factors that might affect the business. Another section of the questionnaire was designed to assess energy use and costs over the years, plans to use alternative sources of energy, factors affecting increase in energy cost and their effect on profitability. Other questions focused on the demographics of the greenhouse owner such as income, age, and education. After getting approval from TSU’s Institutional Review Board (IRB), the surveys were distributed via mail to 279 greenhouse growers in Tennessee. Out of the 279 respondents, we received 56 completed mailed surveys, which gave us a 20 percent response rate. The coded surveys were then analyzed by Microsoft Excel and SPSS using descriptive analysis, correlations, chi-square tests and cross-tabulations. A series of statistical analyses were conducted using correlation models, contingency tables, and chi-square tests. Correlation models were used to investigate the strength of the relationship amongst variables. Contingency tables were formatted to display the frequency distribution of the variables. Chi-square tests were done in order to test our assumptions of what factors directly affected each other. Variables considered for contingency table are size, income, energy costs, and year in the business. Results Table 1 provides a summary of the section averaging the characteristics of the respondents. The table shows that 54 percent of the growers fell in the 60 + age group and only 3 percent of the respondents were under 30. The data also shows that a 32 percent of the respondents had an undergraduate degree and 64 percent had an annual income of less than $99,000 dollars. The survey asked about the size of their business in square foot. Fig. 1 shows the sizes of their operation. According to expert Table 1. Summary of the characteristics of the greenhouse operators Percentage Age Under 30 3 31-45 2 46-60 38 Over 60 54 Education Level High School/ GED 21 Some College 14 Undergraduate Degree 32 Graduate Degree 25 Other 4 13 opinion by Dr. Roger Sauve, a greenhouse operating in less than 10,000 sq-ft is considered small; 10,000- 30,000 sq-ft is considered medium and above 30,000 sq-ft is considered a large greenhouse. Results from the survey indicate that 61 percent of the respondents have small operations. Based on data collected from the survey, the respondents have been in the greenhouse industry for an average of 24 years. In some instances, some of the businesses have been around for more than 100 years. Being around for an extensive period of time is extraordinary because of the challenges they are facing such as building maintenance/ upkeep, the economic downturn, and the rise of energy costs. Table 2 displays the percentage of how each factor affects the growers’ businesses. A large number of respondents chose economic downturn and high-energy costs as the reason why their business is being affected. About 48 percent believed that economic downturn was the most important negative effect while 38 percent felt that high-energy cost was the most important negative factor affecting their business. A small number of respondents (18 percent) felt that competition and production problems were the reasons why their businesses were being affected. Table 2. Factors affecting the greenhouse business Category Economic High Energy Competition Production Downturn (%) Cost (%) in Market (%) Problem (%) Most important 48.2 37.5 16.1 1.8 More important Important 21.4 28.6 19.6 0 19.6 8.9 23.2 8.9 Less important 0 0 7.1 25 Least important 3.6 3.6 12.5 21.4 Energy costs are increasing yearly with an average cost of $18,791 in 2006, which has risen to $22,895 by 2011. Forty-six percent of the survey respondents indicated that there was decline in profitability in the business because of increase in energy costs. The following chart accurately displays a comparison between annual energy cost in 2006 and in 2011. Energy costs are the second largest cost for greenhouse owners behind labor costs, and heating totaling to about 70 percent of the total energy budget (Sanford, 2002). With the continuous rise in fuel cost, the survey focuses on key variables within the greenhouse industry that could potentially be altered to lower the Income 99,000 or less 64 100,000 - 299,000 23 300,000 - 399,000 2 400,000 - 499,000 4 500,00 or more 2 Fig. 1. Distribution of the respondents’ greenhouses by size Journal of Applied Horticulture (http://horticultureresearch.net) Fig. 2. Average energy cost of greenhouse businesses in 2006 and 2011. 14 Influence of energy cost and other factors on profitability of greenhouse businesses in Tennessee grower’s energy cost. The survey asked respondents to provide the size of their greenhouse, fuel types they used within a five-year period, and energy saving technologies they used in a five-year period. The results from Pearson’s Correlations show that size of the greenhouse affected the respondent’s energy cost in 2006 (ρ=0.795, p-value=0.000) and 2011 (ρ=0.825, p-value=0.000). The respondents were also asked what fuel types they used in 2006 and what they used in 2011. Fig. 3 shows that many growers stopped using the same fuels. A percentage of growers stopped using kerosene (3.6 percent to 1.8 percent), propane (32.1 percent to 30.4 percent), and oil (12.5 percent to 8.9 percent) and switched to natural gas (26.8 percent to 30.4 percent), wood (1.8 percent to 5.4 percent), and electricity (5.4 percent to 7.1 percent). Fig. 4. Energy saving technology used within a five- year period percentage per greenhouse Fig. 3. Fuel type used within a five-year period percentages per greenhouse. The survey also asked the respondents to share what changes they made to their energy saving technologies. Fig. 4 shows the percentage of energy saving technologies and how they changed in the next 5 years. Many of the respondents that started off using plastic walls (19.6 percent to 14.3 percent) and new coverings (3.6 percent to 1.8 percent) in 2006, switched to using energy curtains (7.1 percent to 12.5 percent), lower temperatures (17.9 percent to 26.8 percent), efficient heaters (3.6 percent to 8.9 percent), and new heaters (0 to 3.6 percent). Some of the greenhouse growers found it so hard to maintain cost and decided to close down for a portion of the year (5.4 percent to 12.5 percent) and to use less space (1.8 percent to 3.6 percent). The survey asked the respondents to rank how optimistic they were about adopting a new energy source. In Fig. 5 about 25 percent of the respondents are optimistic about adopting a new source and a little over 30 percent are not sure. Only 5 percent were very optimistic and over 15 percent are not optimistic. There is no evidence to suggest that age of the greenhouse grower can affect his/her outlook on how they feel about future growth. Fig. 6 represents how different age groups felt about expecting to adopt new sources of energy. As indicated before, most of the respondents were from age 45 and above. Here we see that group Fig. 5. Optimism among respondents about adopting a new source of energy 46- 60 were more optimistic about adopting new sources than the respondents over 60. Another cross-tabulation was done for income vs. impacts rise in energy costs effect on profitability. A cross-tabulation was performed because rise in energy cost that affected profitability can also affect the greenhouse income range. Fig. 7 indicates that growers that fell in the $99,000 or less annual salary had either a significant decline or very little decline due to the increase in energy costs. Table 3 shows tests that shows the relationship among energy cost, income, adoption of new energy sources and age. The test values and p-values show that rise of energy cost was found to be affecting the income, as expected, of the greenhouse. However, adoption of new energy source wasn’t significantly related to income with a p-value of 0.091. The adoption of a new energy Journal of Applied Horticulture (http://horticultureresearch.net) Influence of energy cost and other factors on profitability of greenhouse businesses in Tennessee 15 Fig. 6. How different age groups feel about adopting new energy sources. sources was significantly related to age of the business owner. This result was particularly interesting since there are no previous studies that relate age with any significant factor of a business. The result shows that experience of the business owners play a role in decision making. Table 3. Chi-Square values and significance at P=0.05 level Variables χ2 Value P-value a Effect of energy cost related to 25.627 0.042 Income Adoption of new energy source 27.322a 0.038 related to age Adoption of new energy source 28.862a 0.091 related to income Results confirm the assumption that size affects the greenhouse energy cost. It also shows the relationship between age and adoption of a new energy source as well as the correlation between income and the effects that energy cost had in profitability. This study was focused on the certified growers because they highly depend on the operation. On this list there were 279 certified greenhouse operations in the state of Tennessee. The study analyzed the operations of greenhouse growers in the state of Tennessee and factors affecting their energy costs. The analysis showed that there is a high correlation between energy cost and size of the greenhouse. Results also indicate that the rise in energy cost directly affects profitability of the greenhouse business. Other important factors affecting profitability were economic downturn and cost of operation. This study also indicates that income level and the age of the grower have a significant positive effect in the interests of adopting new energy sources in the future. Understanding the fundamentals of what can greatly affect energy cost can help save greenhouse growers money in the future. Significant efforts are required to improve knowledge of greenhouse growers in Tennessee regarding energy cost. Findings of this study are consistent with other studies showing greenhouse growers have limited understanding of what factors affect their operations the most and what factors help lower costs. To enhance awareness there is a need to introduce different studies being done Fig. 7. Did a rise in energy cost affect profitability for Income – Groups? to analyze the effects of high-energy costs. It was found that a mix of factors affected their operations. The study relies on the people’s educated guess (survey questionsanswers) and just like any other primary data, opinions and guesses can change with time. From a policy stand point, onetime survey has its limitations in terms of sufficient information it can provide. Multiple surveys over time would serve as a more accurate measure for analysis and policy recommendations. The findings of this study can provide insights for other researchers on energy use by greenhouse operators. Cost efficient energy use is a key to maintaining profitability. It is also important that growers capture opportunities to acquaint themselves with alternative energy sources to enhance their knowledge and make choices that will give them positive economic returns. References Hall, C.R., A.W. Hodges and J.J. Haydu. 2006. The economic impacts of green industry in the United States. HortTechnology, 16: 345-353. Muhammed, S., E. Ekanem, S.P. Singh, F. Tegegne and A. AkuleyAmenyenu, 2000. Profile of the Nursery and Greenhouse Industry in the South. Southern Nursery Association Research Conference Proceedings, Atlanta, GA. Vol. 45. p. 496-499. Sanford, S. 2002. Reducing Natural Gas/ Propane use for Greenhouse Space Heating. Rural Energy Issues, p. 4. Stegelin, F. 2007. Economics of Renewable Energy Alternatives for the Green Industry. Economics and Marketing. SNA Research Conference. 52: 436-438. Tennessee Department of Agriculture 2012. Nursery and Greenhouse Database. TN Dept. of Agr., Nashville, TN Trainer, T. 2010. Can renewables etc. solve the greenhouse problem? The negative case. Energy Policy, 38(8): 4107-4114. United States Department of Agriculture, 2007. National Agricultural Statistics Services, Census of Agriculture, US. Dept. of Agr, Washington, D.C. Received: September, 2015; Revised: December, 2015; Accepted: December, 2015 Journal of Applied Horticulture (http://horticultureresearch.net) Journal Journal of Applied Horticulture, 18(1): 16-18, 2016 Appl Postharvest behaviour of minimally processed watercress V.R. Logegaray*, D. Frezza, A. Chiesa and A.P. León Departamento de Horticultura, Facultad de Agronomía, Universidad de Buenos Aires, Av. San Martín 4453 (C1417DSE) CABA, Argentina. *E-mail: logegara@agro.uba.ar Abstract Watercress (Nasturtium officinale R. Br.) is an aquatic plant of the Brassicaceae family and used as a leafy vegetable that grows in and around water. It is consumed raw or steamed and has a short shelf life of approximately seven days. The objective of this study was to evaluate the postharvest behaviour of watercress minimally processed and stored at optimal storage temperature vs. market temperature. Treatments were: shoots packed with plain film (PD961EZ, 31µm thickness) and stored in refrigerated chambers at 1 ± 0.5 ºC and 8 ± 2 ºC for 10 days. Overall visual quality, gas concentration inside the packages, chlorophyll, reducing sugar, ascorbic acid, oxalic acid and weight loss were evaluated. At the end of the storage period overall visual quality, gas concentration and reducing sugars were affected by storage time and temperature, whereas dehidro ascorbic, oxalic acid and weight loss were not. Key words: Nasturtium officinale, quality, gas concentration, chlorophyll, weight loss, ascorbic acid, reducing sugar, oxalic acid Introduction due to chlorophylls changes (Goncalves et al., 2009). Watercress (Nasturtium officinale R. Br.) is a leafy vegetable of the Brassicaceae family that grows in and around water. Raw watercress leaves are used as salad green or can be steamed and consumed as a normal processed vegetable. It is a good source of essential vitamins and minerals and beneficial phytonutrients and it has a short shelf life (approximately seven days) that can be extended through different techniques such as cold refrigeration and modified atmosphere packaging (Goncalves et al., 2009). Minimally processed products are prepared and handled to maintain their fresh nature while providing convenience to the consumer as ready to eat (Francis et al., 1999; Lanciotti et al., 2004). The attractiveness and convenience of fresh-cut vegetables are helping to bring about increased consumption of fresh produce, but these benefits are offset by the rapid deterioration and short shelf life of the products in the marketplace (Allende et al., 2004). The shelf life of a food can be defined as the time between the production and packaging of the product and the point at which it becomes unacceptable under defined environmental conditions. General appearance is the most important attribute that consumers use to evaluate the quality of fruits and vegetable, as people “buy with their eyes” (Piagentini et al., 2005). Exposure to higher temperatures and/or fluctuations of storage temperature produce cumulative adverse effects on the quality of stored foods, which is the primary cause of damage to food marketed through retail channels. Maintaining the shelf life of watercress represents a valuable advantage for distributors and retailers, and is also a convenient and healthy option to the final consumer (Piagentini et al., 2005). The loss of quality is caused by physical and chemical changes taking place in the product. Colour is one of the most important attributes which affects the consumer perception, and it is also an indicator of the vegetable pigment concentration. During postharvest, the colour of green vegetables suffers modifications Watercress is a major source of ascorbic acid that besides its vitamin action, is valuable for its antioxidant effect, stimulating the immune system and other health benefits. During processing, distribution and storage, the ascorbic acid oxidizes to dehydroascorbic acid which retains vitamin C activity (Cruz et al., 2008). The objective of this study was to evaluate the postharvest behaviour of watercress, minimally processed and stored at optimal storage temperature vs. market temperature (1 ºC and 8 ºC) for 10 days. Material and methods Watercress plants were grown in a floating system with a complete nutrient solution, in the greenhouse of the experimental field at the Horticultural Department of the Faculty of Agronomy, University of Buenos Aires, Argentina (35˚ 35’ S, 58˚ 31’ W). After transplant, plants were grown for forty five days and finally were harvested, selected, washed and sealed in polyolefin bags: multilayered polyolefin PD-961EZ non perforated (oxygen permeability: 6000-8000 cm³ m² 24 h, 1 atm at 23 ºC, carbon dioxide permeability: 19000-22000 cm³ m² 24 h, 1 atm at 23 ºC, and water vapor transmission: 0,90 - 1,10 g 100 square inch, 24 h, 23 ºC, 100 % RH). About 35 ± 5 g of shoots cut were packed in bags, sealed and stored in refrigerated chambers at 1 ± 0.5 ºC and 8 ± 2 ºC (optimal storage temperature versus market temperature) for 10 days. The samples were taken to evaluate overall visual quality, gas concentration (oxygen and carbon dioxide) inside the bags, total chlorophyll and weight loss at twenty four hours after sealed and every three days. Overall visual quality (OVQ) was evaluated using a scale of 9 to 1, where 9 = excellent and 1 = unusable, a score of 6 was considered as the limit of commercial acceptability (LópezGálvez et al., 1996). Journal of Applied Horticulture (http://horticultureresearch.net) Postharvest behaviour of minimally processed watercress 17 Oxygen (%) and carbon dioxide (%) concentration inside the packages were analyzed with a PBI-Dansensor Gas Analyzer Checkmate 9000 (Denmark). Table 2. Oxygen concentration (%) evolution inside the bags for watercress minimally processed packed in non perforated film (PD961EZ) and stored in refrigerated chambers at 1 ± 0.5 ºC and 8 ± 2 ºC for 10 days Total chlorophyll content (mg 100 g-1 fresh tissue) was estimated with a Minolta SPAD 502 according to León et al. (2010). Storage temperature 0 1 4 7 10 Reducing sugars (mg g fresh tissue) were measured according an adaptation of the technique developed by Somogyi-Nelson (Nelson, 1944). 1 ± 0.5 ºC 21 f 20.4 ef 19.2 cd 18.9 bc 19.2 cd 8 ± 2.0 ºC 21 f 19.9 de 18.7 bc 18 ab 17.2 a Dehidroascorbic acid (mg g-1 fresh tissue) was measured with a spectrophometric method according to Vicente et al. (2006). Table 3. Carbon dioxide concentration (%) evolution inside the bags for watercress minimally processed packed in non perforated film (PD961EZ) and stored in refrigerated chambers at 1 ± 0.5 ºC and 8 ± 2 ºC for 10 days -1 Weight of bags was recorded initially and after storage; the difference was used to calculate weight loss percentage (León et al., 2009). Days after packaging Different letters means significant difference (P<0.05) Storage temperature Days after packaging 0 1 4 7 10 Oxalic acid (mg g-1 fresh tissue) was analyzed according to the procedure descripted by Conesa et al. (2009). 1 ± 0.5 ºC 0.03 a 4.1 f 3.9 ef 2.9 bc 2.8 b 8 ± 2.0 ºC 0.03 a 4.1 f 3.5 de 3.2 bcd 3.2 cd An experimental design completely randomized was used; the experimental unit was each bag. The data obtained were subjected to an analysis of variance using SAS statistical program 8.5 and later for media comparisons of each treatment the Tukey test (P= 0.05) was used. For the analysis of the overall visual quality non parametric methods according to Friedman was used. Different letters means significant difference (P<0.05) Results and discussion Overall visual quality (OVQ): Overall visual quality was significantly affected by storage time and temperature. OVQ declined during storage period and quality losses were mainly attributed to wilting and yellowing (Table 1). Table 1. OVQ for watercress minimally processed packed in non perforated film (PD-961EZ) and stored in refrigerated chambers at 1 ± 0.5 ºC and 8 ± 2 ºC for 10 days Storage temperature 1 ± 0.5 ºC 8 ± 2.0 ºC 0 9a 9a Days after packaging 1 4 7 9a 8a 8a 9a 8a 7 ab 10 6b 3c Different letters means significant difference (P<0.05) At the end of postharvest, the best quality was obtained for watercress minimally processed and stored at 1 ± 0.5 ºC. Overall visual quality was below the commercial limit of acceptability when watercress plants were stored at 8 ± 2 ºC for 10 days. Gas concentration inside the bags: Gas concentration inside the non perforated bags was affected by storage time and temperature (Table 2 and 3). During storage period oxygen concentration decreased while carbon dioxide concentration increased as a consequence of the respiratory process. Gas concentration inside the bags is a consequence of a dynamic and interactive process among external environment, the permeability of the packaging material, the internal atmosphere inside the bags and the product itself (León et al., 2009). Chlorophyll content: Total chlorophyll content was affected by storage time and temperature (Table 4). At the end of the storage period, total chlorophyll was higher when watercress minimally processed was stored at 1 ± 0.5 ºC. This parameter is very important because these pigments are responsible for the Table 4. Total chlorophyll content ( mg 100 g -1 fresh tissue) for watercress minimally processed packed in non perforated film (PD961EZ) and stored in refrigerated chambers at 1 ± 0.5 ºC and 8 ± 2 ºC for 10 days Storage temperature 1 ± 0.5 ºC 8 ± 2.0 ºC 0 36.4 b 36.4 b Days after packaging 1 4 7 38.6 b 41.7 b 40.2 b 38.7 b 38.5 b 35.9 b 10 37 b 24.3 a green color and it is one of the major attributes which affects the consumer perception of quality (Francis, 1999). The loss of chlorophyll is responsible for the yellowing of fresh cut products and is the result of disruption of compartmentalization that occurs when cells are broken, allowing substrates and oxidases to come in contact (Tavarini et al., 2007). Reducing sugars: Significant differences were obtained for reducing sugars at the end of the storage period. Lower values obtained when plants were stored at 8 ± 2 ºC could be due to an increase in the respiration rate estimated through high oxygen depletion inside the bags. Table 5. Reducing sugar concentration ( mg g -1 fresh tissue) for watercress minimally processed packed in non perforated film (PD961EZ) and stored in refrigerated chambers at 1 ± 0.5 ºC and 8 ± 2 ºC for 10 days Storage temperature 1 ± 0.5 ºC 8 ± 2.0 ºC 0 10.6 cd 10.6 cd Days after packaging 1 4 7 11.5 cd 9.6 bc 9.5 bc 11.3 cd 7.0 ab 8.4 abc 10 12.9 d 5.5 a Different letters means significant difference (P<0.05) Dehidro ascorbic acid: At the end of postharvest, dehidro ascorbic acid content of watercress minimally processed and stored at 1 ± 0.5 ºC and 8 ± 2 ºC for 10 days did not differ significantly. Watercress minimally processed presents higher contents of dehidro ascorbic than most commonly consumed vegetables (peas 31-26 mg 100 g-1; green beans 25-100 mg 100 g-1; carrots 4 mg 100 g-1; spinach 31-22 mg 100 g-1) (Cruz et al., 2009). This result demonstrates that watercress is a good vitamin C source Journal of Applied Horticulture (http://horticultureresearch.net) 18 Postharvest behaviour of minimally processed watercress Table 6. Dehidro ascorbic acid concentration (mg g-1 fresh tissue) for watercress minimally processed packed in non perforated film (PD961EZ) and stored in refrigerated chambers at 1 ± 0.5 ºC and 8 ± 2 ºC for 10 days Storage Days after packaging temperature 0 1 4 7 10 1 ± 0.5 ºC 3.9 a 5.6 ab 8.5 b 6.4 ab 6.4 ab 8 ± 2.0 ºC 3.9 a 6.8 ab 6.2 ab 5.9 ab 6.4 ab Different letters means significant difference (P<0.05) and an important vegetable for the human diet. Weight loss: This parameter was significantly affected by storage time (Table 7). At the end of the storage period weight loss were lower than 1 % and these results could be due to the very high relative humidity inside the bags. These results agree with Hong and Kim (2004) for green onion who reported that nonperforated film acts like a water vapor barrier reducing weight loss of products. Table 7. Weight loss (%) for watercress minimally processed packed in non perforated film (PD-961EZ) and stored in refrigerated chambers at 1 ± 0.5 ºC and 8 ± 2 ºC for 10 days Storage temperature 1 ± 0.5 ºC 8 ± 2.0 ºC 0 0a 0a Days after packaging 1 4 7 0a 0a 0.3 ab 0a 0.2 ab 0.2 ab 10 0.3 ab 0.7 b Different letters means significant difference (P<0.05) Oxalic acid: At the end of the storage period oxalic acid content remained constant (Table 8) although at 1 ± 0.5 ºC it has a tendency to increase and this response is similar with those reported by Merry Evelyn et al. (2003) for spinach stored at dark who found increases of oxalic acid content after 24 days of storage. Table 8. Oxalic acid content (mg g-1 fresh tissue) for watercress minimally processed packed in non perforated film (PD-961EZ) and stored in refrigerated chambers at 1 ± 0.5 ºC and 8 ± 2 ºC for 10 days. Storage temperature 1 ± 0.5 ºC 8 ± 2.0 ºC 0 171 a 171 a Days after packaging 1 4 7 203 a 188 a 221 a 203 a 193 a 221 a 10 231 a 170 a Different letters means significant difference (P<0.05) PD 961EZ film was suitable for preserving watercress quality at 1 ± 0.5 ºC for ten days. The overall visual quality was maintained above the limit values of commercial acceptability. If the product was stored at higher temperature, the overall visual quality was below the limit of acceptability by the consumers, mainly due to significant yellowing that could be explained by the lower values of total chlorophyll recorded at the end of storage. Gas concentrations inside the bags were significantly affected by storage time and temperature. At the end of the storage, reducing sugar and total chlorophyll content were affected by temperature, while, oxalic, dehidro ascorbic acid and weight loss were not. References Allende, A., E. Aguayo and F. Artés, 2004. Quality of commercial fresh processed red lettuce throughout the production chain and shelf life. International Journal Food Microbiology, 91: 109-117. Conesa, E., D. Niñirola, M.J. Vicente, J. Ochoa, S. Bañon and J.A. Fernandez, 2009. The influence of nitrate/ammonium ratio on yield quality and nitrate, oxalate and vitamin C content on baby leaf spinach and bladder campion plants grown in a floating system. Acta Hortulturae, 843: 269-274. Cruz, R.M.S., M.C. Viera and C.L.M. Silva, 2008. Effect of heat and thermosonication treatments on watercress (Nasturtium officinale) vitamin C degradation kinetics. Innovative Food Science Emerging Technologies, 9: 483-488. Cruz, R.M.S., M.C. Viera and C.L.M. Silva, 2009. Effect of cold chain temperature abuses on the quality of frozen watercress (Nasturtium officinale R. Br.). Journal Food Engineering, 94: 90-97. Francis, F.J. 1999. Quality as influenced by colour. Food Quality Preference, 6: 149-155. Goncalves, E.M., R.M.S. Cruz, M. Abreu, T.R.S. Brandao and C.L.M. Silva, 2009. Biochemical and colour changes of watercress (Nasturtium officinale R. Br.) during freezing and frozen storage. Journal Food Engineering, 93: 32-39. Hong, S. and D. Kim, 2004. The effect of packaging treatment on the storage quality of minimally processed bunched onions. International Journal Food Science Technology, 39: 1033-1041. Lanciotti, R., A. Gianotti, F. Patrignani, N. Belletti, M.E. Guerzoni, and F. Gardini, 2004. Use of natural aroma compounds to improve shelf-life and processed fruits. Trends in Food Science Technology, 15(3): 201-208. León, A., G. De Santibañes, V. Logegaray, A. Chiesa and D. Frezza, 2010. Berro de agua (Nasturtium officinale R.Br.): Estimación no destructiva del contenido de clorofila. Resúmenes del XXXIII Congreso Argentino de Horticultura, 28 de setiembre al 1 de octubre, Santa Fe, Argentina, p.304. León, A., D. Frezza and A. Chiesa, 2009. Effect of nutrient solution composition and storage conditions on postharvest of minimally processed butterhead lettuce. Advances Horticultural Science, 23(1): 13-19. López-Gálvez, G., M. Saltveit and M. Cantwell, 1996. The visual quality of minimally processed lettuces stored in air or controlled atmosphere with emphasis on romaine and iceberg types. Postharvest Biology Technology, 8:179-190. Merry Evelin, A.T., U.Yoshinori, I. Yoshihiro and A. Mitsuko, 2003. L-ascorbic acid metabolism in spinach (Spinacia oleracea L.) during postharvest storage in light and dark. Postharvest Biology Technology, 28: 47-57. Nelson, N. 1944. A photometric adaptation of the somogyi method for the determination of glucose. Journal Biology Chemistry, 153: 375-380. Piagentini, A.M., J.C. Mendez, D.R. Guemes, and M.E. Pirovani, 2005. Modeling changes of sensory attributes for individual and mixed fresh-cut leafy vegetables. Postharvest Biology Technology, 38: 202-212. Tavarini, S., D. DeglInnoceti, A. Pardossi and L. Guidi, 2007. Biochemical aspects in two minimally processed lettuce upon storage. International Journal Food Science Technology, 42: 214219. Vicente, A., G.A. Martínez, A. Chaves and M. Civello, 2006. Effect of heat treatment on strawberry furit damage and oxidative metabolism during storage. Postharvest Biology Technology, 40: 116-122. Received: October, 2015; Revised: November, 2015; Accepted: December, 2015 Journal of Applied Horticulture (http://horticultureresearch.net) Journal Journal of Applied Horticulture, 18(1): 19-24, 2016 Appl Aroma profiling of jasmine (Jasminum sambac Ait.) flowers using electronic nose technology X. Alex Isac1*, K.R. Rajadurai1, M. Jawaharlal2, K. Arul Mozhi Selvan3, D. Uma4, Hena Roy5 and Nabrun Bhattacharyya5 Department of Floriculture and Landscaping, Tamil Nadu Agricultural University, India. 2Horticultural College & Research Institute for Women, Navalur Kutapattu, Trichy, India. 3Department of Soil Science & Agricultural Chemistry, Tamil Nadu Agricultural University, India. 4Department of Biochemistry, Centre for Plant Molecular Biology, Tamil Nadu Agricultural University, India. 5Centre for Development of Advanced Computing (C-DAC), Kolkata, India. *E-mail: senthamil.alex@gmail.com 1 Abstract The Jasmine (Jasminum sambac Ait.) flowers are highly fragrant and used for extraction of essential oil, preparation of perfumes and scented water. Since there is a growing demand for the fresh flowers, there arises a need to develop a technique to identify the flower quality in non-destructive and quickest possible manner. A study was undertaken using hand held electronic nose technology (HEN) at the Department of Floriculture and Landscaping, Tamil Nadu Agricultural University, Coimbatore during the year 2013-2015. The result showed that, the HEN device generated Aroma Index (AI) score increased over the flower development stages and varied from 0.41 in immature bud (stage I) to 4.26 in matured bud (stage V). The comprehensive study on quantum of fragrance releasing pattern at different flower opening stages (physiologically matured bud to fully opened flower) over period of time interval showed that, minimum of 5.41 was recorded in an unopened closed bud stage which gradually increased upto 41.26 in the fully opened flowers. The biochemical constituents responsible for the unique jasmine flower fragrance were identified using Gas Chromatography –Mass Spectrometry (GC-MS). Key words: Jasminum sambac, electronic nose, MOS, volatile emission Introduction Jasmine occurrences are distributed in tropical and subtropical ecologies around the world. Jasmines are grown commercially in India, Thailand, China, Sri Lanka and the Philippines for its fresh flowers. The genus Jasminum contains more than 200 species and is mostly tropical in distribution (Abdul Khader and Kumar, 1995). Though there are a large number of species and varieties in jasmine, commercial cultivation is confined to only a very few, viz., Jasminum sambac, J. auriculatum and J. grandiflorum, which are largely cultivated and J. multiflorum (Syn: J. pubescence ) which is cultivated to a small extent. The flowers are highly fragrant and used for religious offerings in temples and highly preferred by ladies for adorning their hair. They are also used for extraction of essential oil which is used in the preparation of perfumes and scented water. The jasmine concrete is highly valuable for perfume, confectionaries, cosmetics and toiletry industries. Advancement of chemical and instrumental research in revealing the fragrance compounds in nature and also in synthesizing those components in laboratory has been achieved to some extent. The natural odor of the flowers are perfect and unchanged, cannot be completely extracted and analysed. There are various methods of analyzing these natural extract from flowers. Till date these fragrance estimation were mostly by subjective methods performed by panel of skilled persons. The analysis had been done through instruments like chromatography, spectrography etc., by trained operator. To simplify the fragrance estimation, portable and reliable tools are required. So far, a specially designed Electronic Nose has been successfully used to monitor volatile substances emission pattern over repeated measurements. Electronic Nose is a unique tool that is capable of sensing the volatile compounds of the given sample and reliably predicts scores with a high degree of accuracy. Neural Network based Soft Computing Techniques are used to tune near accurate correlation smell print of multisensor array. The software framework has been designed with adequate flexibility and openness, so that may train the system of scoring with reliable predictions of such smell print scores. Since there is a growing demand for the fresh flowers, there arises a need to develop a technique to identify the flower quality in non-destructive and quickest possible manner. These kind of technique would facilitate export of these flowers to both short and long distance overseas markets without much loss of the post harvest quality. Materials and methods The experiment was undertaken at the Department of Floriculture and Landscaping, Tamil Nadu Agricultural University, Coimbatore in collaboration with Centre for Development and Advanced Computing (C-DAC), Kolkata during the year 2013-2014. The experimental material consisted of flowers of major species of Jasmine i.e., J. sambac was used. The experiment was laid out in Completely Radomized Block Design (CRD) with three replications. Journal of Applied Horticulture (http://horticultureresearch.net) 20 Aroma profiling of jasmine flowers using electronic nose technology Fresh flowers (unopened fully matured flowers) from J. sambac were collected from randomly selected plants in the early morning hours around 5 am to 6 am during the entire period of study. The harvesting stages were classified based on the visual appearance of the flower bud. The different stages of harvesting are given in Table 1. Observations of E-nose generated Aroma Index (AI), ethylene emission rate and respiration rate was recorded for each stage of harvesting. Table 1. Different harvesting stages of J. sambac flowers Stage of harvest Age of flower bud (days) Stage I 5 Stage II 8 Stage III 10 Stage IV 12 Stage V 15 The matured unopened buds were harvested from the randomly selected plants. The flower buds were continuously observed using E-nose till it opened fully (i.e., from morning to late evening). Postharvest physiological parameters such as E-nose generated Aroma Index (AI), CO2 rate and ethylene emission rate were recorded at 10.00 am, 4.00 pm, 6.00 pm, 7.00 pm and 8.00 pm. The E-nose generated Aroma Index (AI) values for the above parameters were analysed statistically and least significant difference was applied to compare the differences among different time intervals at 5% as critical level of probability (α). The flower opening index was categorized based on the values in flower opening index chart (Table 2). Identification of different fragrances was performed by using electronic nose, equipped with a metal oxide semiconductor sensor (MOS). E-nose has a great potential to discriminate fragrances and would be a useful tool for the fragrance of ornamentals. Electronic Nose system for gradation of jasmine based on aroma characteristics comprised of two main components (i) The Sniffing Unit and (ii) Data Processing Unit. The sniffing unit consists of the sensory and sensing unit. The sniffing unit is the odor capture and delivery system to the sensor array and the data processing unit is responsible for data acquisition from the sensor array through a proper signal conditioning circuit and the acquired data is processed to generate and display the Fragrance Index. The experimental sniffing cycle consists of automated sequence of internal operations: (i) headspace generation, (ii) sampling, (iii) purging before the start of the next sniffing cycle. Initially these MOS sensors require heating for at least one hour to be stable. Heating is done by supplying 5 Volts to the heater coils of the sensors. This heating phase of sensors is referred to as Preheating. The MOS sensors react to volatile compounds on contact; the adsorption of volatile compounds on the sensor surface causes a physical change of the sensor. Table 2. Flower Opening Index chart of jasmine flowers (J. sambac) at different flower opening stages Flower opening index Flower opening pattern 0 Un-open bud 0.5 Nearly slight open 1.0 Slightly open 1.5 Nearly half open 2.0 Half open 2.5 Nearly full open 3.0 Fully open Principle of operation: The samples to be tested were placed in a sample holder (be specific) in the E-Nose set-up. Data was recorded separately for flowers and buds. It has been observed that the system was able to identify the bud and also the blossoming state of Jasmine. Air flow was blown into the sample container as pressure applied in time scale of second in order to ensure adequate concentration of volatiles in the air within the container. In the same time scale, output voltage was recorded with sensor when exposed to volatile substance influences. The time specified in seconds, for which the sensor array is exposed to fresh air in order to reestablish baseline values of the sensors. Due to the strong scent of jasmine, it is recommended that a purging time of at-least 15 minutes be used. The sensor array was exposed to a constant flow of volatiles emanated from Jasmine flower at time duration of 50 seconds. Data from all the sensors were stored all through this sampling operation, but the steady-state value for each sensor is considered for computation purposes. During the purging operation of 100 secs, sensor heads were cleared through the blow of air so that the sensors can go back to their baseline values. Data acquisition module: Sensor outputs were fed to this module. After signal conditioning, the channels were multiplexed and were fed to an data acquisition card. The DAQ output was fed to the processor for analysis and storage. Flower opening index (FOI): The fragrance emission was determined during fresh flowers opening period which is under influence of their development stage varying from Stage I to stage V. There were significant differences in opening of flower from tight bud stage to fully opened condition as in V stage. The various flower opening indices recorded at different time intervals were categorized (Table 3). Soxhlet extraction: The compounds responsible for fragrance emission in fully opened jasmine flowers were detected through the concrete extracted using Soxhlet extractor. The fresh samples of about 20 g were taken in the extraction chamber placed in a tube above the extraction solvent. The solvent used was food grade hexane (analytical reagent) to wash the sample using a reflux apparatus. When heated, the solvent evaporates into a gas, and then cools into a liquid in a condenser. It then refluxes back into the sample tube. This continuous cyclic process takes around 45 to 60 minutes per cycle until the concrete is separated from the sample. The solvent was evaporated off, by keeping it in the water bath and the amount of concrete was determined. Gas chromatography: Later the extracted concrete samples at different interval of time were subjected to Gas Chromatography –Mass Spectrometry analysis. About one micro litre of sample concentrate was injected into a Thermo GC - Trace Ultra Ver: 5.0,Thermo MS DSQ II gas chromatograph equipped with a flame ionization detector. The column used was DB 35 - MS Capillary Standard Non - Polar Column. The specifications of Gas Chromatography used for analysis; Column: 50m × 0.25mm internal diameter (i.d.) coated with PEG 20 M, film thickness: 0.15 pm, Carrier gas: N, with a flow rate at 1.2 mL/min, oven temperature: 60°C (4 min) + 220°C, injection and detector temperature: 200°C Split ratio: 10: l Journal of Applied Horticulture (http://horticultureresearch.net) Aroma profiling of jasmine flowers using electronic nose technology Results and discussion Fragrance parameters estimation at various development stages of flowers were recorded and given in the Table 3 and Table 4. Time taken for flower opening is an important character, which signifies the earliness or late flowering habit of the genotype. Both the habits are helpful in determining the availability of flowers for longer period (Khader and Kumar, 1995). It was observed that there was no ethylene evolution observed till stage III while it was triggered up to 1 ppm during the stage IV. Then after, ethylene was released up to 3.2 ppm after Stage V of harvest. Under ambient conditions, ethylene evolution rate with the range of 2.0 ppm and 22.8 ppm after harvest of fresh flowers at different time intervals were observed in fully matured opened flower (Table 4). The results indicated that the rate of ethylene evolution increased rapidly after harvest upto 16 hours, after which the senescence of flower starts (Fig. 2). Similar results were observed by Mayak and Halevy (1974), Suttle and Kende (1978, 1980) and Borochov et al. (1997). Also it was observed that as the flowers started showing symptoms of wilting, there was a rise in level of ethylene emission rate. The E-nose generated Aroma Index (AI) of jasmine flowers during different flower developmental stages were observed and it varied from 0.41 in I stage, 0.83 in II stage, 1.58 in III stage, 2.53 in IV stage to 4.26 in V stage. It clearly indicates that the Metal oxide Sensor (MoS) used in the E-nose instrument, sniffed the compounds responsible for fragrance and generated the Aroma Index (AI) as the stage of harvesting progressed. Fully matured harvested jasmine flowers were continuously observed from its closed unopened stage to fully opened condition. Based on our observation the E-nose generated Aroma Index (AI), gradually Table 3. E-nose generated aroma index (AI) of J. sambac at different flower developmental stages Harvesting stages E-nose Value (Aroma Index) 0.41 0.83 1.58 2.53 4.26 0.02 Stage I Stage II Stage III Stage IV Stage V SEm ± LSD (P=0.05) CO2 Rate (ppm) 0 0 1.2 2.3 8.4 0.1 0.1 0.07 Ethylene Emission Rate (ppm) 0 0 0 1.0 3.2 0.1 0.1 Table 4. Ethylene, CO2 and Aroma Index at different Flower Opening Index (FOI) of J. sambac Time *Flower opening index (FOP) 10.00 am 0 04.00 pm 06.00 pm E-nose value (Aroma Index) CO2 Rate (ppm) Ethylene emission rate (ppm) 5.41 8.4 2.0 0.5 16.83 27.8 9.5 1 24.58 36.3 14.9 07.00 pm 2 32.53 42.6 18.6 08.00 pm 3 41.26 56.8 22.8 SEm ± 0.01 0.1 0.1 LSD (P=0.05) 0.21 0.3 0.4 21 increased from 5.41 in 10.00 am (unopend closed bud stage) to 16.83 in 4.00 pm (nearly slight open stage), 24.58 in 6.00 pm (slightly opened stage), 32.53 in 6.00 pm (half opened) and 41.26 in 8.00 pm (fully opened). As the flower opens, the fragrance emission is higher and the senescence of flower takes place. Ethylene hormone has been known to play a crucial role in senescence of flowers, the sensitivity of which varies depending on the flower species (Redman et al., 2002). Ethylene reduces the longevity of some flowers causing rapid wilting of petals (e.g., carnations), shedding or shattering of petals, or other changes to petal tissues, such as loss or change of colour. Earlier reports (Naidu and Reid, 1989) have indicated that the flowers are ethylene sensitive based on the fact that though the flowers produce moderate ethylene during opening and senescence, they do not respond to exogenously applied ethylene (Veen, 1983) indicating that this hormone is not involved in their senescence. No report is available on Jasminum spp. with respect to ethylene evolution, however some records on wilting of flowers other than jasmine caused by ethylene have been discussed. Involvement of ethylene in wilting of flowers (Borochov et al., 1997) has been observed in carnation (Ten Have and Woltering, 1997) and in Gypsophila paniculata (Vandoorn and Reid, 1992). It has also been noticed that the flower parts including petals, sepals, the ovary and labellum were the major site of ethylene production (Chao Chia et al., 1991) and that ethylene promoted the accumulation of sugars and inorganic materials in the ovary, with a simultaneous loss of fresh and dry weight of the petals. These are some evidences of ethylene sensitive species where in, ethylene is the major cause of wilting of flowers. Respiration rate: The minimum respiration rate of 8.4 ppm was observed at 10 am. It steadily increases and reached maximum of 56.8 ppm at 8.0 pm in the evening. The respiration of flowers started from III stage upto 1.2 ppm, followed by 2.3 ppm in IV stage and gradually increased in V stage upto 8.4 ppm (Table 3). All the flowers showed a climacteric rise in respiration rate after harvest. The lowest respiration rates were recorded immediately after harvest. This may be due to short supply of readily respirable substrates in the flowers due to onset of senescence. Similar results were reported by Coorts (1973) in cut flowers and Maxie et al. (1973) in carnation. Increased respiration leads to formation of free radicals with high oxidation potential. Free radicals promote senescence in tissues which in turn increases sensitivity to ethylene (Fig. 2). Respiration is the central process in living cells that mediates the release of energy through the oxidative breakdown of carbon compounds (starch, sugar and organic acids) and the formation of carbon skeletons necessary for maintenance and synthetic reactions after harvest (Wills et al., 1998). In the present study, a respiratory climacteric rise from the initial level and a decline thereafter was noticed with all the treatments. With regard to J. sambac under ambient conditions a similar trend was noticed, recording minimum respiration rate and sufficient amount of carbohydrate levels. These significant levels of carbohydrates might have served as the substrate for respiration for a longer duration. Evidences supporting this fact have been reported in case of flowers supplied with exogenous sugar, wherein pool of dry matter and respirable substrates were maintained at favourable levels thus promoting respiration (Coorts, 1973) and in turn extending the longevity (Rogers, 1973). The Journal of Applied Horticulture (http://horticultureresearch.net) 22 Aroma profiling of jasmine flowers using electronic nose technology observation of Maxie et al. (1973) that the respiratory activity in flowers and the production of carbon-di-oxide by flowers was similar to the pattern in climacteric fruits, characterized by a rise in level of respiration with senescence also supports the present study. Kaltaler and Steponkus (1976) have associated the decline in respiratory activity of aging rose petals with their inability to metabolise substrates consequent to decline in activity of mitochondria in the aging petals. Moreover, increased respiratory activity leads to the formation of free radicals with high oxidation potential and these free radicals have been found to promote senescence in the tissues, associated with an increased sensitivity to ethylene (Baker et al., 1977; Mishra et al., 1976). The typical climacteric respiratory rise reported in carnation cv. White Sim (Burger et al., 1986) and day-lily (Lukaszewski and Reid, 1989) is consistent with the present result. In contrary, Trippi and Paulin (1984) had reported a decrease in respiratory activity in carnation cv. White Sim. GC-MS analysis: The concrete obtained from soxhlet extractor was injected into Gas Chromatography–Mass Spectrometry instrument and constituents were identified by comparison of both mass spectra and retention indices, strictly measured on the same instrument with those of authentic jasmine samples. The identified constituents listed in Table 5 and Table 6 with their respective chromatogram obtained shown in Fig. 1 and 2 exhibited significant compounds identified at specific time intervals (10.00 am and 8.00 pm). The GC-MS chromatogram at 10 am shows, the preliminary indication about the composition of some major volatile components. However, the quantitative composition of these compounds differs considerably from the other samples. The jasmine flower possesses maximum composition and recorded peak during this time. They are Eicosanoic acid, phenylmethyl ester (Benzyl icosanoate) (26.47%), 9-Octadecenoic acid (Z), phenylmethyl ester (Benzyl oleate) (24.04%), Nonadecane (17.41%) and 2,6-Octadien-1-ol, 3,7 dimethyl-(Z)- (cis-Geraniol) (14.24%). Fig. 1. GC-MS Chromatogram of J. sambac extract (soxhlet extraction) at 10.00 am Fig. 2. GC-MS Chromatogram of J. sambac extract (soxhlet extraction) at 08.00 pm Journal of Applied Horticulture (http://horticultureresearch.net) Aroma profiling of jasmine flowers using electronic nose technology 23 Table 5. GC-MS analysis of J. sambac extract (soxhlet extraction) at 10.00 AM Table 6. GC-MS analysis of J. sambac extract (soxhlet extraction) at 08.00 PM RT Name of the compound RT 2.23 2,6-Octadien-1-ol, 3,7 dimethyl-(Z)- (cis-Geraniol) 2-Aminononadecane C19H41N Cyclooctyl alcohol C8H16O 1-Tetracosanol C24H50O 1-Methyldodecylamine C13H29N 2,4,6,8-Tetramethyl-1C15H30 undecene Octodrine C8H19N Heptadecane, 2-methylC18H38 1-Hexacosanol C26H54O Heptadecane, C21H44 2,6,10,15-tetramethylDidodecyl phthalate C32H54O4 Decane, 2,3,5,8-tetramethyl- C14H30 6H-Pyrazolo[1,2 a][1,2,4,5] C7H16N4 tetrazine, hexahydro-2,3dimethyl2-Nonen-1-ol C9H18O Octadecane, 6-methylC19H40 2,6,10-Dodecatrien-1-ol, C15H26O 3,7,11-trimethyl- (Farnesol) 1-Eicosanol C20H42O Nonadecane, 2-methylC20H42 Tetracontane, 3,5,24-trimethyl- C43H88 Nonadecane C19H40 Octadecane, 1-(ethenyloxy)- C20H40O 1-Octadecyne C18H34 Z,Z-2,5-Pentadecadien-1-ol C15H28O 9-Octadecenoic acid (Z)-, C25H40O2 phenylmethyl ester (Benzyl oleate) Eicosanoic acid, phenylmethyl C27H46O2 ester (Benzyl icosanoate) MW Peak Area (%) 154 14.24 283 128 354 199 210 1.23 0.57 1.51 0.38 1.49 129 254 382 296 0.26 0.35 1.06 0.31 2.58 3.51 10.80 14.76 15.80 16.10 17.44 18.79 502 198 156 1.07 0.76 0.67 142 268 222 0.17 0.76 0.37 22.48 22.85 23.03 298 282 604 268 296 250 224 372 0.09 1.54 0.86 17.41 0.94 2.53 0.92 24.04 23.66 24.16 25.46 26.75 28.05 28.63 28.99 402 26.47 31.42 10.44 10.82 12.45 13.92 14.76 15.80 16.10 17.39 18.79 19.49 20.09 21.48 22.44 22.79 23.01 23.79 24.13 25.40 26.67 27.95 28.67 29.47 32.51 33.60 Molecular Formula C10H18O Twenty eight constituents were identified as active principles in the jasmine samples taken at 8 pm. Major are, 9-Octadecenoic acid (Z)-, phenylmethyl ester (Benzyl oleate) (21.01%), 1-Octadecyne (15.35%) and Octadecanoic acid, phenylmethyl ester (Benzyl stearate) (14.04%). The jasmine volatile compounds responsible for its unique fragrance were released during this time. The study on ideal harvesting stage for Jasminum sp. fresh flowers revealed that the E-nose instrument aids in identifying the ideal harvesting stage for fresh flower stage and suitable time for concrete extraction. This ultimately helps in industrial utility to identify the perfect stage and time for higher concrete recovery. Considering the bulk of the components eluted under these chromatographic conditions, and assuming these compounds possesses a response for fragrance exclusive for jasmine species. The identified constituents from jasmine concrete were responsible for their unique fragrance. Acknowledgements The authors are thankful to Tamil Nadu Agricultural University (TNAU), Coimbatore for providing the necessary facilities and the Centre for Development and Advanced Computing (C-DAC), Kolkata and their staff for their funding to carry out the research and preparation of this paper. 2.21 19.48 20.16 21.14 21.51 29.51 30.47 32.76 33.63 Name of the compound Molecular Formula 2,6-Octadien-1-ol, 3,7 dimethyl- C10H18O (Z)- (cis-Geraniol) 1-Octanol, 2,7-dimethylC10H22O Cyclopropyl carbinol C4H8O Cyclooctyl alcohol C8H16O 2,4,6,8-Tetramethyl-1-undecene C15H30 Octodrine C8H19N Heptadecane, 2-methylC18H38 1-Hexacosanol C26H54O Heptadecane, C21H44 2,6,10,15-tetramethylDidodecyl phthalate C32H54O4 Decane, 2,3,5,8-tetramethylC14H30 Octadecanoic acid, C25H42O2 phenylmethyl ester (Benzyl stearate) 6H-Pyrazolo[1,2 a][1,2,4,5] C7H16N4 tetrazine, hexahydro-2,3dimethyl2-Nonen-1-ol C9H18O Octadecane, 6-methylC19H40 2,6,10-Dodecatrien-1-ol, C15H26O 3,7,11-trimethyl- (Farnesol) 1-Eicosanol C20H42O Nonadecane, 2-methylC20H42 Tetracontane, 3,5,24-trimethyl- C43H88 Nonadecane C19H40 Octadecane, 1-(ethenyloxy)C20H40O 1-Octadecyne C18H34 Dodeca-1,6-dien-12-ol, C14H26O 6,10-dimethylZ,Z-2,5-Pentadecadien-1-ol C15H28O Heptadecanoic acid, heptadecyl C34H68O2 ester 1,4-Dioxaspiro[4.5]decane, C9H16O2S 8-(methylthio)9-Octadecenoic acid (Z)-, C25H40O2 phenylmethyl ester (Benzyl oleate) Eicosanoic acid, phenylmethyl C27H46O2 ester (Benzyl icosanoate) MW Peak Area (%) 154 3.50 158 72 128 210 129 254 382 296 3.01 3.83 1.44 0.15 0.42 0.54 0.22 0.42 502 198 374 0.96 0.62 14.04 156 1.37 142 268 222 0.22 0.93 0.31 298 282 604 268 296 250 210 0.33 2.00 1.08 2.41 0.86 15.35 7.83 224 508 13.22 2.41 188 0.39 372 21.01 402 1.13 References Abdul Khader, JBM MD and N. Kumar, 1995. Genetic resources of jasmine. In: Advances in Horticulture., Vol.12. Ornamental Plants. Chadha, K.L. and S.K. Bhattacharjee (eds.), Malhotra Publishing house, New Delhi. pp. 121-132. Baker, J.E., C.Y. Wang, M. Lieberman and R. Hardenburg, 1977. Delay of senescence in carnation by a rhizobitoxine analog and Na- benzoate. HortScience, 12: 38-39. Burger, L., G.H. Swardt and A.H.P. Engelbrecht, 1986. Relationship between changes in membrane permeability, respiration rate, activities of lipase and phospholipase and ultra structure in senescing petal of Dianthus. S. Afr. J. Bot., 52(3): 195-200. Borochov, A., H. Spiegelstein and P. Hadas, 1997. Ethylene and flower petal senescence interrelation ship with membrane lipid metabolism. Physiol. Plant., 100(3): 606-612. Chao Chia, H., P. Meihwei, W. Tsutsuen, C.C. Huang, M.H. Pan, T.T. Wang.1991. Senescence, ethylene production and electrolyte leakage of different parts of chilled Phalaenopsis flower. J. Agric. Res. China., 46(2): 167-180. Coorts, G.D. 1973. Internal metabolic changes in cut flowers. HortScience, 8: 195-198. Journal of Applied Horticulture (http://horticultureresearch.net) 24 Aroma profiling of jasmine flowers using electronic nose technology Kaltaler, R.E.L. and P.L. Steponkus, 1976. Factors affecting respiration in cut roses. J. Amer. Soc. Hort. Sci., 101: 352-354. Lukaszewski, T.A. and M.S. Reid, 1989. Bulb type flower senescence, 1989. Acta Hort., 261: 59-62. Maxie, E.C., D.S. Farnhano, F.G. Mitchell and N.F. Sommer, 1973. Temperature and ethylene effects on cut flowers of carnation (Dianthus caryophyllus). J. Amer. Soc. Hort. Sci., 98(6): 568-572. Mayak, S. and A.H. Halevy, 1974. The action of kinetin in improving the water balance and delaying senescence processes of cut rose flowers. Physiol. Plant., 32: 330-336. Mishra, S.D., B.K. Gaur , V.W. Bedeker and B.B. Singh, 1976. Isolation, identification and significance of free radicals in senescing leaves. Acta Botanica, 4: 131-138. Naidu, S.N. and M.S. Reid, 1989. Postharvest handling of tuberose (Polianthes tuberosa L.). Acta Hort., 261: 313-317. Redman B. Paul, John M. Dole, Niels O. Maness and Jeffrey A. Anderson. 2002. Post harvest handling of nine speciality cut flower species. Scientia Hort., 92: 293-303. Rogers, M.N. 1973. An historical and critical review of post harvest physiology research on cut flowers. HortSci., 8: 189-194. Suttle, J.C. and H. Kende, 1978. Ethylene and senescence in petals of Tradescantia. Plant Physiol., 63: 267-271. Suttle, J.E. and H. Kende, 1980. Ethylene action and loss of membrane integrity during petal senescence in Tradescantia. Plant Physiol., 65: 1067-1072. Ten Have, A. and E.J. Woltering, 1997. Ethylene biosynthetic genes are differentially expressed during carnation (Dianthus caryophyllus L.) flower senescence. Plant Mol. Biol., 34: 89-97. Vandoorn, W.G. and M.S. Reid, 1992. Role of ethylene in flower senescence of Gypsophila paniculata L. Post Harvest Biol. Tech., 1(3): 265-272. Veen, H. 1983. Silver thiosulphate: an experimental tool in plant science. Scientia Hort., 20: 211-224. Wills, R., B. McGlasson, D. Graham and D. Joyce, 1998. Postharvest: An Introduction to the Physiology and Handling of Fruit, Vegetables and Ornamentals. 4th ed. UNSW Press, Sydney. Received: May, 2015; Revised: November, 2015; Accepted: December, 2015 Journal of Applied Horticulture (http://horticultureresearch.net)