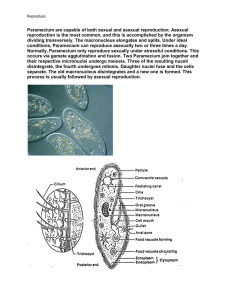
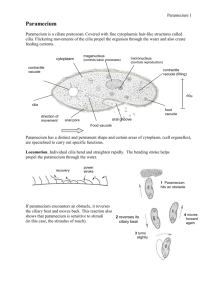
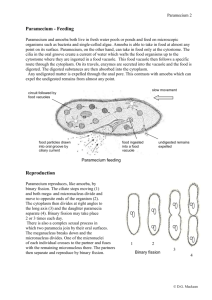
The V-ATPase in Paramecium: functional
advertisement

Pflugers Arch - Eur J Physiol (2009) 457:599–607 DOI 10.1007/s00424-007-0417-x CELL AND MOLECULAR PHYSIOLOGY The V-ATPase in Paramecium: functional specialization by multiple gene isoforms Thomas Wassmer & Ivonne M. Sehring & Roland Kissmehl & Helmut Plattner Received: 14 October 2007 / Revised: 23 November 2007 / Accepted: 29 November 2007 / Published online: 29 January 2008 # Springer-Verlag 2007 Abstract The vacuolar H+-ATPase (V-ATPase), a multisubunit, adenosine triphosphate (ATP)-driven proton pump, is essential for numerous cellular processes in all eukaryotes investigated so far. While structure and catalytic mechanism are similar to the evolutionarily related F-type ATPases, the V-ATPase’s main function is to establish an electrochemical proton potential across membranes using ATP hydrolysis. The holoenzyme is formed by two subcomplexes, the transmembraneous V0 and the cytoplasmic V1 complexes. Sequencing of the whole genome of the ciliate Paramecium tetraurelia enabled the identification of virtually all the genes encoding V-ATPase subunits in this organism and the studying of the localization of the enzyme and roles in membrane trafficking and osmoregulation. Surprisingly, the number of V-ATPase genes in this free-living protozoan is strikingly higher than in any other species previously studied. Especially abundant are V0-a-subunits with as many as 17 encoding genes. This abundance creates the possibility of forming a large number of different V-ATPase holoenzymes by combination and has functional consequences by differential targeting to various organelles. Keywords V-ATPase . Paramecium . Membrane traffic . Osmoregulation . Contractile vacuole complex . Trichocyst . Phagocytosis . Acidosomes T. Wassmer (*) Department of Biochemistry, School of Medical Sciences, University of Bristol, Bristol BS8 1TD, UK e-mail: twassmer@bristol.ac.uk I. M. Sehring : R. Kissmehl : H. Plattner Department of Biology, Universität Konstanz, Universitätsstraße 10, 78457 Konstanz, Germany Introduction The V-ATPase is a proton pump that drives many essential processes in organisms as distantly related as humans, plants, and protozoans (for reviews, see [1–3]). It hydrolyses adenosine triphosphate (ATP) and uses this energy to pump protons across membranes, building up a proton gradient as well as an electric potential. The electrochemical potential can be used for secondary active transport by antiporter systems for various processes. This makes the V-ATPase crucial as an acidifier of compartments and as an energizer of membranes. The structural organization of the V-ATPase resembles the evolutionarily related F-ATPase. They both have a globular cytoplasmic domain and a membrane embedded domain, the two being connected by a central stalk [4] (Fig. 1). While the F-ATPase has only one additional, peripheral stalk, it was shown that V-ATPase has more than one, connecting the cytoplasmic and transmembraneous subcomplexes [5, 6]. The cytoplasmic subcomplex is named V1 and the transmembrane subcomplex V0. The subunits forming the central stalk belong to V1 while subunits that compose the peripheral stalks belong to either V1 or V0. Early insights into the genes encoding V-ATPase subunits were gathered from the budding yeast Saccharomyces cerevisiae (for reviews, see [7, 8]). In yeast, the transmembraneous V0 complex is composed of six different subunits, named a, c, c′, c″, d, e, encoded by seven genes (Table 1) [3]. The soluble, cytoplasmic V1 complex is made of eight different subunits, termed A, B, C, D, E, F, G, H, encoded by eight genes (Table 1). Only the V0-a-subunit is encoded by more than one gene in S. cerevisiae (vph1 and stv1). The enzyme was shown to work in a way somewhat similar to a macroscopic turbine. The hydrolysis of ATP by DO00417; No of Pages 600 Pflugers Arch - Eur J Physiol (2009) 457:599–607 Table 1 Number of V-ATPase genes in Saccharomyces cerevisiae, Mus musculus, and Paramecium tetraurelia V1 A B C D E F G H V0 a c c′ c″ d e S. cerevisiae M. musculus P. tetraurelia 1 1 1 1 1 1 1 1 1 2 2 1 2 1 3 1 4 4 1 1 4 2 Not identified 4 2 1 1 1 1 1 4 1 0 1 2 1 17 6 0 0 3 2 Compiled from Forgac [3] and Wassmer et al. [33]. V1-subunit G could not be identified in P. tetraurelia probably because of limited sequence conservation but is expected to be existent. the cytoplasmic V1 A-subunits, assisted by the noncatalytic B-subunits, drives the rotation of the central stalk, composed of D- and F-subunits, which transmit the rotation to the membrane-embedded V0-c-subunit ring [3, 9, 10]. Each of the highly hydrophobic c-subunits has a glutamate residue critical for function that can pick up a proton delivered to this glutamate, supposedly by a hemichannel in the V0-a-subunit that allows proton entry from the cytoplasm [3]. The carboxy group of the glutamate is neutralized by the proton and then can swivel into the hydrophobic phase constituted by the membrane surrounding the V0-c-subunits. As there are five to six c-subunits organized in a ring that is driven by the central stalk, protons are continuously picked up from the cytoplasm. After a turn of the c-ring, protons are thought to be released from the c-subunits through a second hemichannel in the a-subunit leading to the luminal side of the membrane [3, 11]. One prerequisite is that the V0-a-subunit and the V1subcomplex rest immobile with respect to the c-subunit ring and the central stalk. This suggests that the peripheral stalks, containing the N-terminal half of the V0-a-subunit part and V1-subunits C, E, H, G, serve as a “stator” [10, 12]. By the rotational coupling of ATP hydrolysis to a turbine-like ring, a molecular pump is created, which can build up huge proton potentials. Several mechanisms to regulate the activity of the VATPase have been suggested. One mechanism was shown to be reversible disulfide bond formation at the catalytic site on the A-subunit [13, 14]. Another possibility of regulation is reversible dissociation of the V1 and the V0 complex [3, 15, 16]. A third regulation mechanism was suggested to be the distribution of the enzyme to different cellular membranes [17]. Cell biology of the V-ATPase In eukaryotic cells ranging from simple, unicellular to complex, multicellular organisms, the V-ATPase fulfills a broad range of functions [1, 3, 18, 19]. In yeast, the V-ATPase is primarily localized in the late Golgi and in vacuoles [20]. The difference in localization is mediated by the different V0-a-subunits these two enzyme complexes contain [21]. They differ in coupling efficiency, state of dissociation/association of V1 and V0 and pump protons with different activities [3, 4]. This is probably one explanation for the late Golgi to be only slightly acidic while yeast vacuoles are strongly acidified. In yeast, knockout of any V-ATPase subunit except the two a-subunit genes, vph1 and stv1, results in the severe “vacuolar membrane ATPase” (vma) phenotype, which comprises growth deficiency in neutral medium and hypersensitivity to elevated Ca2 concentration in the medium [22]. Growth of yeast cells displaying a vma phenotype is only sustained at acidic pH and low Ca2+ concentration. Only the double knockout of both vph1 and stv1 lead to the vma-phenotype, suggesting that these two subunits can substitute for each other to some degree in yeast [20]. In mammals, the number of genes encoding V-ATPase subunits is strikingly higher than in yeast (compare Table 1), and therefore the subunit composition is more complex, with variations between cell types and tissues [3, 23]. The enzyme is targeted to numerous organelles, among them early endosomes, late endosomes/lysosomes, the transGolgi network, dense core secretory granules, synaptic vesicles, and in some specialized cells, to the plasma membrane. Those responsible for the differential targeting seem to be the V0-a-subunits, for example, subunits a3 and a4 targeting the enzyme to the plasma membrane of osteoclasts and renal intercalated cells [4, 11, 24]. In mammals the V-ATPase is of crucial importance for several membrane-trafficking events, e.g., endocytosis, endosomal sorting, endosomal maturation, and exocytotic capacity of dense core secretory granules and synaptic vesicles [1, 3]. Special attention has received the fact that the mammalian V-ATPase is important for acidification of extracellular spaces by an enzyme pool located in the plasma membrane [25, 26]. Extracellular acidification is necessary for bone resorption [27] and maturation of spermatozoa [28]. The V-ATPase was shown to play a major role in the invasiveness and metastatic potential of cancer cells [29]. It has also been Pflugers Arch - Eur J Physiol (2009) 457:599–607 suggested to be a pH sensor on the endocytic pathway where, depending on lumenal acidification, specific cytosolic proteins (small GTPases and regulator proteins) can be attached, thus governing interaction with downstream organelles [30]. Genes encoding subunits of the Paramecium V-ATPase The first V-ATPase gene to be described in Paramecium was the P. multimicronucleatum B-subunit [31]. Molecular data on P. tetraurelia has recently been boosted by the Paramecium genome project [32]. Sequence information from a draft assembly was used to identify most of the Paramecium V-ATPase subunits [33]. Table 1 gives an overview on gene numbers in P. tetraurelia in comparison with S. cerevisiae and M. musculus. One major outcome of the P. tetraurelia genome project is that the genome has globally and recently been duplicated [32]. Therefore, nearly all of the P. tetraurelia V-ATPase subunits are encoded by gene pairs that are greater than 80% identical in deoxyribonucleic acid sequence and nearly identical on the protein level. The Paramecium genome also shows traces of older genome duplications during which V-ATPase genes were duplicated and retained in the genome. This seems to be a general Fig. 1 Scheme of the V-ATPase. Subunits of the cytoplasmic V1 subcomplex are depicted with capital letters/gray, membrane embedded V0-subunits with small letters/white 601 phenomenon in this ciliate and could account for the large number of about 40,000 genes present in the genome. Because of these events, V-ATPase subunits in Paramecium are encoded by more genes than in any other organism investigated so far [33]. Among the most numerous genes are the V0-a-subunit genes that form a big multigene family with 17 paralogues. In this family, 16 genes can be grouped in pairs, while only one gene lacks a homologue from the most recent genome duplication [34]. As the V-ATPase genes forming a pair encode nearly identical proteins, it is difficult to imagine that there is any functional differentiation between them. In contrast, the presence of all the different gene pairs in the genome, which encode similar but distinct proteins, suggests that these differences are physiologically relevant in terms of function, as redundant gene copies tend to be eliminated from a genome in the course of evolution because of the lack of selection pressure. Localization and function of the V-ATPase in Paramecium Osmoregulation Fok et al. [35] were the first to establish the localization of the V-ATPase in Paramecium on the osmoregulatory system, also termed the “contractile vacuole complex.” To understand the role of the V-ATPase, its function was inhibited using concanamycin B. Thus, the VATPase has been found to be crucial for osmoregulation in this fresh water ciliate [35, 36]. Under normal conditions, Paramecium cells possess two contractile vacuole complexes, one being located close to the anterior and the other one close to the posterior pole of the cell. Each organelle is comprised of a central vacuole, the so called contractile vacuole that undergoes a rhythmic fusion and fission cycle with the plasma membrane. Upon fusion with the plasma membrane, its contents (mainly water and ions) are discharged into the medium. Closely attached to the contractile vacuole is an array of so called “radial arms.” Each radial arm is composed of an ampulla, which is adjacent to the contractile vacuole, and the radial canal, which is surrounded by two continuous complex membrane cisternae, called the “smooth” and the “decorated spongiome.” The current idea on the functioning of the contractile vacuole system is that ions are concentrated in the spongiome, which is osmotically followed by water influx [37, 38]. The water is collected via the radial canals in the ampullae, which fuse with and therefore pass their content on to the contractile vacuole. When this vacuole undergoes fusion with the plasma membrane and releases its content, one pump cycle is completed, and the cycle starts anew. For detailed information on the contractile vacuole complex, see the studies of Allen and Naitoh [36] and Allen [39]. 602 The main driving force for this pump cycle was shown to be the V-ATPase [35]. The enzyme pumps protons into the lumen of the decorated spongiome, where the V-ATPase is located in such abundance that in electron microscopy, it actually gives the decorated spongiome the electron dense, hence “decorated” appearance. Most likely, the proton potential is then used for secondary active transport of ions, among them K+ and Ca2+, into the lumen of the contractile vacuole complex [37, 38], probably by antiporter systems, which is followed by osmotic water influx. In fact, if the V-ATPase is inhibited by concanamycin B, the contractile vacuole pump cycle stops [35, 37, 38]. Localization and function of the V-ATPase in the contractile vacuole complex could be confirmed by studying green fluorescent protein (GFP)-tagged V-ATPase a-, c-, or F-subunits and by ribonucleic acid (RNA) interference with these subunits (RNAi) [33, 34]. GFPlabeled c-subunits 1, 4, and 5, representatives of all three c-subunit pairs (c1/2, c3/4, c5/6), all localized, among several other organelles, to the radial arms of the contractile vacuole system, as did a GFP-tagged F-subunit. Among the V0-a-subunits, only a2-1 specifically localized to the osmoregulatory system [34]. RNAi aiming at the expression of c- or F-subunits led to morphological defects in the structure of the contractile vacuole complex, both on the light and electron microscopical level, and a heavily impaired pumping cycle [33]. No specialization with respect to the c-subunit pairs involved in osmoregulation was detected, as RNAi with either of the three c-subunit pairs (c1/2, c3/4, c5/6) or with combinations of them led to similar results, suggesting that all three pairs are important for osmoregulation but leaving the question open, whether they occur mixed in V-ATPase complexes or whether they compose different V-ATPase complexes that are all necessary for the functioning of the contractile vacuole complex. An even more drastic phenotype was observed by interfering with a-subunit 2 expression. Osmoregulation by the contractile vacuole complex was disturbed to the point where Paramecium cells showed massive swelling and finally died by bursting [34]. Surprisingly, the contractile vacuole complex seems to be only mildly acidic with pH 6.4 [38] and is therefore not stained by the pH-sensitive dye acridine orange (Fig. 2a), despite the huge number of V-ATPase molecules it contains in its decorated spongiome [35]. The most likely explanation is that the proton potential is coupled so tightly to secondary active transport of ions into the lumen of the contractile vacuole complex that no bigger proton potential is being built up. To date, no comprehensive study has been carried out to dissect the role of the numerous putative antiporter systems encoded in the Paramecium genome [32], (http://paramecium. cgm.cnrs-gif.fr). As the major cations in the fluid expelled by Pflugers Arch - Eur J Physiol (2009) 457:599–607 the contractile vacuole are K+ and, to a lower degree, Ca2+, this suggests that the focus in the search for antiporter systems connected to the funtioning of the contractile vacuole complex should be concentrated on H+/K+ and H+/Ca2+ antiporters. Phagocytosis and endocytosis Another readily identified cellular function of the V-ATPase in Paramecium is phagocytosis. Paramecium feeds on bacteria and small eukaryotic microorganisms. In the process of phagocytosis, bacteria are concentrated by a dense field of cilia at a region specialized for phagocytosis, termed the “cytostome.” There, the nascent food vacuole forms, enlarges, and pinches off the cytostome. Within 2 to 4 min, the food vacuole is acidified by fusing with a population of small vesicles called “acidosomes” that transport a high number of V-ATPase molecules [40, 41]. Acidosomes are thus constantly delivered to the cytostome and attach themselves to the nascent food vacuole. The pinching off process seems to trigger the fusion of acidosomes with the food vacuole. After the initial acidification burst, the pH rises to nearly neutral for the rest of the lifespan of the food vacuole. The acidosomal membrane is withdrawn from the phagosomes and cycles back to the cytostome. This process is paralleled by withdrawal of the V-ATPase being responsible for the strong acidification [41] and its recycling. In Fig. 2, acidosomes at the cytostome are stained by acridine orange and visible in the form of a horseshoe arranged around the cytostome. For a more detailed representation of the phagocytic cycle, see the study of Allen and Fok [42]. Recently, the observed cycle of V-ATPase delivery by acidosomes to phagosomes and the consecutive withdrawal from them could be correlated with molecular data on the V-ATPase. A subunit of V0, a4, was exclusively localized to acidosomes, the nascent food vacuole, and the freshly formed food vacuole from which it recycled back to the cytostome. In contrast, older food vacuoles contained the a-subunits a5, a6, and a9 [34]. This suggests that there are at least two cycles of V-ATPase delivery and withdrawal to and from food vacuoles, respectively. In the first cycle, a V-ATPase with a supposedly high activity containing subunit a4 is delivered to phagosomes to produce a rapid acidification burst to inactivate ingested microorganisms. This V-ATPase is recycled back to the cytostome, while a different V-ATPase enzyme set, containing a5, 6, and 9, is delivered to food vacuoles, possibly by lysosomes or lysosomal precursors. It is possible that a5, 6, and 9 are delivered in a sequential manner depending on the age of the food vacuole, but this has not been investigated yet. As the older food vacuoles do contain the V-ATPase, while they are not remarkably acidic, it is likely that the proton potential created by the pump is used for secondary active transport of nutrients out and to pump waste products into the vacuole. This is another aspect that remains to be elucidated. Pflugers Arch - Eur J Physiol (2009) 457:599–607 603 Fig. 2 Staining of acidic compartments of Paramecium using acridine orange. Fluorescent (a, c) and corresponding phase contrast images (b, d) show the distribution of various acidic vesicles in the cytoplasm (all median focus plane). Note the bright labeling of acidosomes around the cytostome (a, inset) and the punctate staining at the edge of the anterior and posterior end of the cells (arrows), indicating early endosomes. The surface view of a cell (e) shows the regular arrangement of endosomes below the cell surface. The corresponding phase contrast image (g) reveals that the osmoregulatory system is not stained (arrow). f Detail of e as indicated, showing cortical endosomes; h same detail on median focus. Bars=10 μm What happens to phagocytosis when the expression of the V-ATPase is inhibited? RNA interference with the expression of the c- or F-subunits of the V-ATPase not only strongly reduced acidification, it also inhibited formation of phagosomes [33]. The mechanism for this is not clear at the moment. A possible explanation might be that the initial acidification burst is necessary to signal further processing of the food vacuole in terms of transport and fusion with lysosomes. Lack of this signal might lead to a block in downstream events and, therefore, a failure in the retrieval of acidosomal membrane from the young food vacuole back to the cytostome. Ultimately, lack of acidosomes at the cytostome should cause the absence of phagocytosis. The role of V-ATPase in endocytosis and endosomal sorting in Paramecium has not been worked out yet. What is established is that this acidic compartment contains the V-ATPase with the V0-subunit a1 [34] (Figs. 2 and 3). Dense core secretory vesicle biogenesis The third process in Paramecium in which the V-ATPase was found to be involved in is the formation of dense core secretory granules, the so-called trichocysts [33]. In most organisms, dense core secretory granules are known to be acidified by the V-ATPase [43]. The finding that Paramecium trichocysts contain a V-ATPase was surprising, as trichocysts were shown by two independent reports using different methods not to be acidic compartments, neither the mature organelle nor trichocyst precursors [44, 45]. Interference with c-, F-, or a3-subunits of the Paramecium V-ATPase led to malformation of these organelles [33, 34]. They do not reach their usual carrot shape, fail to form proper tips, and are not attached to the plasma membrane. In their misshapen form, they resemble mutant trichocysts that are defective in the proteolytic maturation from trichocyst matrix precursor proteins to the correctly processed and assembly-competent trichocyst matrix proteins [46, 47]. 604 Pflugers Arch - Eur J Physiol (2009) 457:599–607 Fig. 3 Schematic distribution of V-ATPase subunits in the Paramecium cell. The red arrows indicate transport routes during phagosomal processing based on reviews [42, 48]. V-ATPase subunit distribution is based on data obtained with GFP localization in vivo and with antibody labeling [33, 34]. Blue: V0-a-subunits, black: V0-c-subunits, green: V1F-subunits; red filling: acidified vesicles/vacuoles; pink filling: mildly or not recognizably acidic compartments but with V-ATPase subunits detected in the membrane (presence in trichocyst precursors inferred from findings with mature trichocysts). The scheme contains elements of the osmoregulatory system (a ampula, cv contractile vacuole, ds decorated spongiome, ss smooth spongiome) and of the phagosomal apparatus (oc oral cavity, fv food vacuole, as acidosomes, cp cytoproct). Discoidal vesicles (dv) and other recycling vesicles (rv) are responsible for return transport of membranes. The endosomal system, composed of parasomal sacs (ps) and early endosomes (ee), is arrayed in a regular fashion. ci Cilia, er endoplasmatic reticulum, ga Golgi apparatus, trpc trichocyst precursor, tr trichocysts, gh ghosts (from released trichocysts), pm plasma membrane Correctly assembled trichocysts fuse with the plasma membrane upon a stimulus [48]. After membrane fusion, extracellular calcium enters the trichocyst body, and a major rearrangement of the paracrystaline trichocyst core takes place. It transforms into a long and insoluble needle, which is, thus, ejected into the medium. A possible role for the V-ATPase in the formation of trichocysts could be to provide an electrochemical proton potential that is exploited by an antiporter to pump out Ca2+ for making trichocysts competent for their exocytotic discharge, but this hypothesis remains to be tested. This would be in line with work on dense core secretory vesicles in mammalian cells where the V-ATPase has been shown to contribute to activation and specific sequestration of secretory contents [49, 50]. Remarkably, besides trichocysts, there are only few dense core secretory vesicles whose lumen is not remarkably acidic, e.g., parotis gland granules, although their membranes contain a H+-ATPase [51]. ments to which no specific function has been attributed or which had not been identified in any detail so far. Further functional aspects of Hþ -ATPase activity Unfortunately, it was not possible to assign any role to the VATPase V0 part of Paramecium in exocytotic membrane fusion [33] as had been suggested for other systems [52, 53, 54], based on experiments on homotypic yeast vacuole membrane fusion [55]. Apart from the three major functions of the V-ATPase in Paramecium presented here, it is likely to play many other yet unidentified roles in cellular processes as suggested by the presence of numerous a-subunits specific in compart- Functional overlap between the V-ATPase and other pumps Beyond the V-type ATPase described in this review, the Paramecium cell also possesses P-type ATPases that were reported to be involved in Ca2+ metabolism. This includes a plasmamembrane Ca2+-ATPase [56] and a SERCA-type (sarcoplasmic/endoplasmic reticulum Ca2+-ATPase) Ca2+ pump [57] whose biogenesis starts in the endoplasmic reticulum and finally is enriched in the cortical Ca2+ stores, the “alveoli” [58–60]. However, experimental analysis of the SERCA [61] and calculations from the kinetics of these pumps suggest that they are less powerful regulators in maintaining a low cytoplasmic Ca2+concentration than extrusion of Ca2+ ions via the contractile vacuole complex [62]. From studies with concanamycin B combined with Ca2+ imaging, we know that the H+-ATPase of the osmoregulatory system largely contributes to the re-establishment of Ca2+ homeostasis after signal transduction accompanying exocytosis stimulation (Sehring and Plattner, in preparation). This function complements the chemiosmotic, ΔH+-based water transport for osmoregulation, as described in the studies of Allen and Naitoh [36] and Allen [39]. In the Paramecium genome, a group of genes with homology to the vacuolar H+-translocating inorganic pyro- Pflugers Arch - Eur J Physiol (2009) 457:599–607 phosphatases of plants also exists (Wassmer, unpublished observation). Neither localization nor function of these genes have been studied, but the presence of a whole group of these genes suggests that not only V-type ATPases are used for H+ pumping in Paramecium but also pyrophosphatases. The specialists: functional diversification of V0-a-subunits Localization studies of the nine different a-subunit clusters (eight gene pairs and a single gene) showed them to be located in at least seven distinct membrane compartments [34]. Subunit a1-1 was found to be on the endocytic pathway, most likely on early endosomes. In many systems, it is known that early endosomes are acidified by the VATPase. The staining of Paramecium cells with acridine orange strongly labels early endosomes localized next to basal bodies directly below the plasma membrane (Fig. 2), suggesting acidification by V-ATPases containing a-subunit pair 1. Subunit a2-1 localized exclusively to the contractile vacuole complex, while a3-1 localized to trichocysts. a4-1 was found to shuttle between acidosomes and the nascent food vacuole, while a5-1, a6-1, and a9-1 localized to the membrane of older phagosomes. Subunit a7-1 is most likely localized in endoplasmic reticulum membranes and a8-1 in the Golgi apparatus. Whether these a-subunits can be rerouted and replace the function in the absence of another a-subunit was investigated by RNAi with a-subunits 2 and 3, located in the contractile vacuole complex and trichocysts, respectively. Knockdown of a2-1/a2-2 subunits disrupted the function of the contractile vacuole complex and ultimately led to cell death, demonstrating that none of the other endogenous a-subunits could replace it. The same holds true for RNAi with a3-1/3-2. Absence of these V0-subunits caused malformation of trichocysts, showing that no other a-subunit can rescue this process [34]. The most likely explanation for the specificity is the differential localization of the a-subunits. The fact that none of the other a-subunits is rerouted in the absence of others suggests that they contain a targeting signal that specifies the localization of the holoenzyme, an observation previously made for a-subunits in other organisms [21, 24]. The presence of such a targeting signal was demonstrated by domain swap experiments and was found to lie within the C-terminal, transmembraneous half of the V0-a-subunits in Paramecium, although it could not be narrowed down to a short signal peptide sequence [34]. Any domain swapping within the C terminus of any a-subunit tested led to abnormal targeting of the chimeric subunit accompanied by cell division arrest and cell death. From this finding, it was concluded that the targeting signal is complex and 605 spreads over the whole C terminus of the V0-a-subunits in Paramecium [34]. The C-terminal localization signal in Paramecium contrasts with observations in yeast, in which the targeting signal was demonstrated to be in the Nterminal half of the a-subunits [21]. The difference can possibly be explained by the evolution of the a-subunits. A phylogenetic analysis reveals that the gene duplication in yeast and the a-subunit multiplication in Paramecium have occurred after these organisms have diverged from a common ancestor [34]. It is highly unlikely that similar signal recognition and targeting mechanisms should have evolved independently in the two organisms to correctly localize different a-subunits. Establishing and comparing the two different mechanisms should prove to be an exciting task for future work. Combinatorial enzyme design: a V-ATPase constructed from a building set? The puzzling number of V-ATPase genes raises several interesting possibilities. Are the V-ATPase molecules in the Paramecium cell a complex mosaic of the dozens of slightly different subunits that possibly changes over time? What are the physiological differences between them? Does a different subunit composition allow differential control, i.e., pumping kinetics, lumenal acidification, and binding of cytosolic regulators? The fact that the same c- and F-subunits associate with different V0-a-subunits in a compartment-specific way clearly demonstrates the presence of a variety of different V-ATPases in Paramecium. However, this phenomenon is most likely not only restricted to the V0 subcomplex. When an a-subunit chimera composed of a2-1 and a3-1 was introduced into the cell, it was correctly localized according to the targeting signal in the C terminus of the molecule. However, stunningly, the chimera introduced abnormalities specifically in the organelle, to which the V-ATPase would have been targeted, if its a-subunit had contained the correct C terminus [34]. The a-subunit chimera thus created a remote effect in an organelle in which it was not present. This scenario can most easily be explained under the assumption that the N terminus of the chimeric a-subunit binds to V1 complexes, which, in their unique subunit composition, “belong” to a different organelle. The a-subunit was shown to connect V0 with V1 as part of a peripheral stalk [63, 64]. Overexpression of a chimera would thus deplete the cytoplasmic V 1 subcomplex pool from which a different organelle recruits its specific V1 complexes. This model suggests that by the combination of slightly different V-ATPase subunits, one can create a multitude of 606 different isozymes. Paramecium thus offers an attractive experimental system to test how an enzyme is fine tuned by the use of different subunits in terms of function, structure, and localization. These possibilities will be exploited in future work. Acknowledgement We would like to thank Jean Cohen, Janine Beisson, and Linda Sperling (CNRS, Gif-sur-Yvette, France) for many helpful discussions and for access to the Paramecium Genome Database. We gratefully acknowledge the support from Genoscope, France, for sharing sequence information with us during sequencing and assembling the Paramecium genome. References 1. Sun-Wada GH, Wada Y, Futai M (2004) Diverse and essential roles of mammalian vacuolar-type proton pump ATPase: toward the physiological understanding of inside acidic compartments. Biochim Biophys Acta 1658:106–114 2. Schumacher K (2006) Endomembrane proton pumps: connecting membrane and vesicle transport. Curr Opin Plant Biol 9:595–600 3. Forgac M (2007) Vacuolar ATPases: rotary proton pumps in physiology and pathophysiology. Nat Rev Mol Cell Biol 8:917–929 4. Inoue T, Wang Y, Jefferies K, Qi J, Hinton A, Forgac M (2005) Structure and regulation of the V-ATPases. J Bioenerg Biomembr 37:393–398 5. Wilkens S, Inoue T, Forgac M (2004) Three-dimensional structure of the vacuolar ATPase. Localization of subunit H by difference imaging and chemical cross-linking. J Biol Chem 279:41942–41949 6. Venzke D, Domgall I, Kocher T, Fethiere J, Fischer S, Bottcher B (2005) Elucidation of the stator organization in the V-ATPase of Neurospora crassa. J Mol Biol 349:659–669 7. Arata Y, Nishi T, Kawasaki-Nishi S, Shao E, Wilkens S, Forgac M (2002) Structure, subunit function and regulation of the coated vesicle and yeast vacuolar (H(+))-ATPases. Biochim Biophys Acta 1555:71–74 8. Kane PM, Smardon AM (2003) Assembly and regulation of the yeast vacuolar H+ATPase. J Bioenerg Biomembr 35:313–321 9. Yokoyama K, Nakano M, Imamura H, Yoshida M, Tamakoshi M (2003) Rotation of the proteolipid ring in the V-ATPase. J Biol Chem 278:24255–24258 10. Imamura H, Nakano M, Noji H, Muneyuki E, Ohkuma S, Yoshida M, Yokoyama K (2003) Evidence for rotation of V1-ATPase. Proc Natl Acad Sci USA 100:2312–2315 11. Nishi T, Forgac M (2002) The vacuolar (H)-ATPases—nature’s most versatile proton pumps. Nat Rev Mol Cell Biol 3:94–103 12. Hirata T, Iwamoto-Kihara A, Sun-Wada GH, Okajima T, Wada Y, Futai M (2003) Subunit rotation of vacuolar-type proton pumping ATPase: relative rotation of the G and C subunits. J Biol Chem 278:23714–23719 13. Feng Y, Forgac M (1992) A novel mechanism for regulation of vacuolar acidification. J Biol Chem 267:19769–19772 14. Feng Y, Forgac M (1994) Inhibition of vacuolar H(+)-ATPase by disulfide bond formation between cysteine 254 and cysteine 532 in subunit A. J Biol Chem 269:13224–13230 15. Kane PM (1995) Disassembly and reassembly of the yeast vacuolar H+ATPase in vivo. J Biol Chem 270:17025–17032 16. Qi J, Forgac M (2007) Cellular environment is important in controlling V-ATPase dissociation and its dependence on activity. J Biol Chem 282:24743–24751 17. Brown D, Breton S (2000) Sorting proteins to their target membranes. Kidney Int 57:816–824 Pflugers Arch - Eur J Physiol (2009) 457:599–607 18. Mellman I (1992) The importance of being acid: the role of acidification in intracellular membrane transport. J Exp Biol 172:39–45 19. Ijzendoorn SC (2006) Recycling endosomes. J Cell Sci 119:1679– 1681 20. Manolson MF, Wu B, Proteau D, Taillon BE, Roberts BT, Hoyt MA, Jones EW (1994) STV1 gene encodes functional homologue of 95-kDa yeast vacuolar H(+)-ATPase subunit Vph1p. J Biol Chem 269:14064–14074 21. Kawasaki-Nishi S, Bowers K, Nishi T, Forgac M, Stevens TH (2001) The amino-terminal domain of the vacuolar protontranslocating ATPase a subunit controls targeting and in vivo dissociation, and the carboxyl-terminal domain affects coupling of proton transport and ATP hydrolysis. J Biol Chem 276:47411– 47420 22. Graham LA, Flannery AR, Stevens TH (2003) Structure and assembly of the yeast V-ATPase. J Bioenerg Biomembr 35:301– 312 23. Sun-Wada GH, Yoshimizu T, Imai-Senga Y, Wada Y, Futai M (2003) Diversity of mouse proton-translocating ATPase: presence of multiple isoforms of the C, d and G subunits. Gene 302:147– 153 24. Toyomura T, Oka T, Yamaguchi C, Wada Y, Futai M (2000) Three subunit a isoforms of mouse vacuolar H(+)-ATPase. Preferential expression of the a3 isoform during osteoclast differentiation. J Biol Chem 275:8760–8765 25. Yao G, Feng H, Cai Y, Qi W, Kong K (2007) Characterization of vacuolar-ATPase and selective inhibition of vacuolar-H(+)-ATPase in osteoclasts. Biochem Biophys Res Commun 357:821–827 26. Breton S, Brown D (2007) New insights into the regulation of V-ATPase-dependent proton secretion. Am J Physiol Renal Physiol 292:F1–10 27. Ochotny N, Van Vliet A, Chan N, Yao Y, Morel M, Kartner N, von Schroeder HP, Heersche JN, Manolson MF (2006) Effects of human a3 and a4 mutations that result in osteopetrosis and distal renal tubular acidosis on yeast V-ATPase expression and activity. J Biol Chem 281:26102–26111 28. Pietrement C, Sun-Wada GH, Silva ND, McKee M, Marshansky V, Brown D, Futai M, Breton S (2006) Distinct expression patterns of different subunit isoforms of the V-ATPase in the rat epididymis. Biol Reprod 74:185–194 29. Sennoune SR, Luo D, Martinez-Zaguilan R (2004) Plasmalemmal vacuolar-type H+ATPase in cancer biology. Cell Biochem Biophys 40:185–206 30. Hurtado-Lorenzo A, Skinner M, El Annan J, Futai M, Sun-Wada GH, Bourgoin S, Casanova J, Wildeman A, Bechoua S, Ausiello DA, Brown D, Marshansky V (2006) V-ATPase interacts with ARNO and Arf6 in early endosomes and regulates the protein degradative pathway. Nat Cell Biol 8:124–136 31. Fok AK, Yamauchi K, Ishihara A, Aihara MS, Ishida M, Allen RD (2002) The vacuolar-atpase of Paramecium multimicronucleatum: gene structure of the B subunit and the dynamics of the V-ATPaserich osmoregulatory membranes. J Eukaryot Microbiol 49:185–196 32. Aury JM, Jaillon O, Duret L, Noel B, Jubin C, Porcel BM, Ségurens B, Daubin V, Anthouard V, Aiach N, Arnaiz O, Billaut A, Beisson J, Blanc I, Bouhouche K, Câmara F, Duharcourt S, Guigo R, Gogendeau D, Katinka M, Keller AM, Kissmehl R, Klotz C, Koll F, Le Mouël A, Lepère G, Malinsky S, Nowacki M, Nowak JK, Plattner H, Poulain J, Ruiz F, Serrano V, Zagulski M, Dessen P, Bétermier M, Weissenbach J, Scarpelli C, Schächter V, Sperling L, Meyer E, Cohen J, Wincker P (2006) Global trends of whole-genome duplications revealed by the ciliate Paramecium tetraurelia. Nature 444:171–178 33. Wassmer T, Froissard M, Plattner H, Kissmehl R, Cohen J (2005) The vacuolar proton-ATPase plays a major role in several membranebounded organelles in Paramecium. J Cell Sci 118:2813–2825 Pflugers Arch - Eur J Physiol (2009) 457:599–607 34. Wassmer T, Kissmehl R, Cohen J, Plattner H (2006) Seventeen asubunit isoforms of Paramecium V-ATPase provide high specialization in localization and function. Mol Biol Cell 17:917–930 35. Fok AK, Aihara MS, Ishida M, Nolta KV, Steck TL, Allen RD (1995) The pegs on the decorated tubules of the contractile vacuole complex of Paramecium are proton pumps. J Cell Sci 108:3163–3170 36. Allen RD, Naitoh Y (2002) Osmoregulation and contractile vacuoles of protozoa. Int Rev Cytol 215:351–394 37. Stock C, Gronlien HK, Allen RD, Naitoh Y (2002) Osmoregulation in Paramecium: in situ ion gradients permit water to cascade through the cytosol to the contractile vacuole. J Cell Sci 115:2339–2348 38. Stock C, Gronlien HK, Allen RD (2002) The ionic composition of the contractile vacuole fluid of Paramecium mirrors ion transport across the plasma membrane. Eur J Cell Biol 81:505–515 39. Allen RD (2000) The contractile vacuole and its membrane dynamics. Bioessays 22:1035–1042 40. Allen RD, Fok AK (1983) Nonlysosomal vesicles (acidosomes) are involved in phagosome acidification in Paramecium. J Cell Biol 97:566–570 41. Allen RD (1988) Cytology. In: Goertz HD (ed) Paramecium. Springer, Berlin, pp 4–40 42. Allen RD, Fok AK (2000) Membrane trafficking and processing in Paramecium. Int Rev Cytol 198:277–317 43. Apps DK (1997) Membrane and soluble proteins of adrenal chromaffin granules. Semin Cell Dev Biol 8:121–131 44. Lumpert CJ, Glas-Albrecht R, Eisenmann E, Plattner H (1992) Secretory organelles of Paramecium cells (trichocysts) are not remarkably acidic compartments. J Histochem Cytochem 40: 153–160 45. Garreau de Loubresse N, Gautier MC, Sperling L (1994) Immature secretory granules are not acidic in Paramecium: Implications for sorting to the regulated pathway. Biol Cell 82:139–147 46. Gautier MC, Garreau de Loubresse N, Madeddu L, Sperling L (1994) Evidence for defects in membrane traffic in Paramecium secretory mutants unable to produce functional storage granules. J Cell Biol 124:893–902 47. Madeddu L, Gautier MC, Le Caer JP, Garreau de Loubresse N, Sperling L (1994) Protein processing and morphogenesis of secretory granules in Paramecium. Biochimie 76:329–335 48. Plattner, Kissmehl (2003) Molecular aspects of membrane trafficking in Paramecium. Int Rev Cytol 232:185–216 49. Taupenot L, Harper KL, O’Connor DT (2005) Role of H-ATPasemediated acidification in sorting and release of the regulated secretory protein chromogranin A. Evidence for a vesiculogenic function. J Biol Chem 280:3885–3897 50. Waterford SD, Kolodecik TR, Thrower EC, Gorelick FS (2005) Vacuolar ATPase regulates zymogen activation in pancreatic acini. J Biol Chem 280:5430–5434 51. Arvan P, Castle JD (1986) Isolated secretion granules from parotid glands of chronically stimulated rats possess an alkaline internal 607 52. 53. 54. 55. 56. 57. 58. 59. 60. 61. 62. 63. 64. pH and inward-directed H pump activity. J Cell Biol 103:1257– 1267 Hiesinger PR, Fayyazuddin A, Metha SQ, Rosenmund T, Schulze KL, Zhai RG, Verstreken P, Cao V, Zhou Y, Kunz J, Bellen HJ (2005) The v-ATPase V0 subunit a1 is required for a late step in synapic vesicle exocytosis in Drosophila. Cell 121:607–620 Liegeois S, Benedetto A, Garnier JM, Schwab Y, Labouesse M (2006) The V0-ATPase mediates apical secretion of exosomes containing Hedgehog-related proteins in Caenorhabditis elegans. J Cell Biol 173:949–961 Sun-Wada GH, Toyomura T, Murata Y, Yamamoto A, Futai M, Wada Y (2006) The a3 isoform of V-ATPase regulates insulin secretion from pancreatic β-cells. J. Cell Sci. 119:4531–4540 Peters C, Bayer MJ, Buhler S, Andersen JS, Mann M, Mayer A (2001) Trans-complex formation by proteolipid channels in the terminal phase of membrane fusion. Nature 409:581–588 Wright MV, Van Houten JL (1990) Characterization of a putative Ca2-transporting Ca2-ATPase in the pellicles of Paramecium tetraurelia. Biochim Biophys Acta 1029:241–251 Hauser K, Pavlovic N, Kissmehl R, Plattner H (1998) Molecular characterization of a sarco(endo)plasmic reticulum Ca2-ATPase gene from Paramecium tetraurelia and localization of its gene product to sub-plasmalemmal calcium stores. Biochem J 334:31–38 Kissmehl R, Huber S, Kottwitz B, Hauser K, Plattner H (1998) Subplasmalemmal Ca-stores in Paramecium tetraurelia. Identification and characterisation of a sarco(endo)plasmic reticulum-like Ca2-ATPase by phosphoenzyme intermediate formation and its inhibition by caffeine. Cell Calcium 24:193–203 Plattner H, Floetenmeyer M, Kissmehl R, Pavlovic N, Hauser K, Momayezi M, Braun N, Tack J, Bachmann L (1999) Microdomain arrangement of the SERCA-type Ca2 pump (Ca2-ATPase) in subplasmalemmal calcium stores of Paramecium cells. J Histochem Cytochem 47:841–854 Hauser K, Pavlovic N, Klauke N, Geissinger D, Plattner H (2000) Green fluorescent protein-tagged sarco(endo)plasmic reticulum Ca2-ATPase overexpression in Paramecium cells: isoforms, subcellular localization, biogenesis of cortical calcium stores and functional aspects. Mol Microbiol 37:773–787 Mohamed I, Husser M, Sehring I, Hentschel J, Hentschel C, Plattner H (2003) Refilling of cortical calcium stores in Paramecium cells: in situ analysis in correlation with storeoperated calcium influx. Cell Calcium 34:87–96 Ladenburger EM, Korn I, Kasielke N, Wassmer T, Plattner H (2006) An Ins(1,4,5)P3 receptor in Paramecium is associated with the osmoregulatory system. J Cell Sci 119:3705–3717 Xu T, Vasilyeva E, Forgac M (1999) Subunit interactions in the clathrin-coated vesicle vacuolar (H(+))-ATPase complex. J Biol Chem 274:28909–28915 Inoue T, Forgac M (2005) Cysteine-mediated cross-linking indicates that subunit C of the V-ATPase is in close proximity to subunits E and G of the V1 domain and subunit a of the V0 domain. J Biol Chem 280:27896–27903