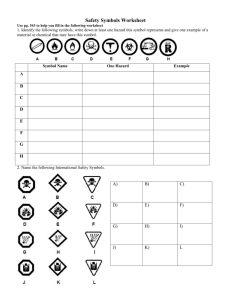
Serial Position Effects in the Identification of Letters, Digits, and
advertisement

Journal of Experimental Psychology: Human Perception and Performance 2009, Vol. 35, No. 2, 480 – 498 © 2009 American Psychological Association 0096-1523/09/$12.00 DOI: 10.1037/a0013027 Serial Position Effects in the Identification of Letters, Digits, and Symbols Ilse Tydgat Jonathan Grainger Ghent University Centre National de la Recherche Scientifique and Aix-Marseille University In 6 experiments, the authors investigated the form of serial position functions for identification of letters, digits, and symbols presented in strings. The results replicated findings obtained with the target search paradigm, showing an interaction between the effects of serial position and type of stimulus, with symbols generating a distinct serial position function compared with letters and digits. When the task was 2-alternative forced choice, this interaction was driven almost exclusively by performance at the first position in the string, with letters and digits showing much higher levels of accuracy than symbols at this position. A final-position advantage was reinstated in Experiment 6 by placing the two alternative responses below the target string. The end-position (first and last positions) advantage for letters and digits compared with symbol stimuli was further confirmed with the bar-probe technique (postcued partial report) in Experiments 5 and 6. Overall, the results further support the existence of a specialized mechanism designed to optimize processing of strings of letters and digits by modifying the size and shape of retinotopic character detectors’ receptive fields. Keywords: letter perception, serial position, alphanumeric character processing, crowding Second, from an empirical perspective, one well-established phenomenon in research investigating letter string perception is the W-shaped function relating identification accuracy to the position of target letters in the string. This is the so-called serial position function (for a recent investigation of letter visibility with varying fixation positions in the string, see Stevens & Grainger, 2003). For odd numbered strings with fixation on the central letter (e.g., five- or seven-letter arrays with central fixation), performance is optimal for the central letter and then drops as a function of eccentricity, except for the first and last letter, which recover a performance level similar to that of the central letter (Averbach & Coriell, 1961; Butler, 1975; Butler & Merikle, 1973; Haber & Standing, 1969; Merikle, Coltheart, & Lowe, 1971; Merikle, Lowe, & Coltheart, 1971; Mewhort & Campbell, 1978; Nazir, Ben-Boutayab, Decoppet, Deutsch, & Frost, 2004; Schwantes, 1978; Stevens & Grainger, 2003; Wolford & Hollingsworth, 1974). For letter detection speed, there is an M-shaped equivalent (Hammond & Green, 1982; Ktori & Pitchford, 2008; Mason, 1975, 1982; Mason & Katz, 1976; Pitchford, Ledgeway, & Masterson, 2008).1 However, this W/M-shaped function is unlikely to be due to a single process. One popular interpretation of the phenomenon is that it reflects the conjoint influence of two factors: (a) the drop in visual acuity as a function of distance from fixation and (b) the It is now a generally accepted fact that visual word recognition in languages with alphabetical orthographies involves processing the identities of the word’s component letters. Many researchers would further agree that a large part of this processing must be performed in parallel (e.g., Grainger, 2008; Grainger & Jacobs, 1996; McClelland & Rumelhart, 1981). Nevertheless, surprisingly little research has been aimed at determining precisely what factors are at play during the computation of letter identities in the earliest phases of printed word perception and whether this processing does, indeed, proceed in parallel. The present study builds on some classic work in this field in an attempt to highlight some of the key unresolved issues. First of all, from a methodological perspective, one particularly useful approach to addressing such issues is to examine the process of letter perception in random consonant strings. In this way, one can gather information about letter-level processing while minimizing the influence of higher level phonological and semantic processes. The assumption is that during visual word recognition, some form of letter-level processing must be performed before such higher order codes come into play (Grainger, Granier, Farioli, Van Assche, & van Heuven, 2006; Grainger & van Heuven, 2003). Ilse Tydgat, Department of Experimental Psychology, Ghent University, Ghent, Belgium; Jonathan Grainger, Centre National de la Recherche Scientifique and Laboratoire de Psychologie Cognitive, Aix-Marseille University, Marseille, France. Part of the research reported in this article was performed while Ilse Tydgat was on an Erasmus exchange between Ghent University and the University of Provence. Thanks to Marine Salery for running Experiments 5 and 6 and to Nikki Pitchford for discussion of this work. Correspondence concerning this article should be addressed to Jonathan Grainger, Laboratoire de Psychologie Cognitive, Université de Provence, 3 pl. Victor Hugo, 13331, Marseille, France. E-mail: jonathan.grainger@ univ-provence.fr 1 The W/M-shaped function is typically found in strings of five letters or greater with fixation on the central letter. Experiments testing four-letter strings and/or using parafoveal stimulus presentation have typically shown U-shaped functions. That is, for stimulus arrays where fixation does not fall on the middle letter of the array, a U-shaped serial position function for letter accuracy has been obtained, with optimal performance for the leftmost and rightmost letters (e.g., Bouma, 1973; Campbell & Mewhort, 1980; Estes, Allmeyer, & Reder, 1976; Jordan & Bevan, 1996; Jordan et al., 2000, 2003; Mewhort & Campbell, 1978; Tramer, Butler, & Mewhort, 1985). 480 SERIAL POSITION EFFECTS amount of lateral interference (crowding) determined by the number of flanking letters (Bouma, 1970, 1973; Estes, 1972; Estes, Allmeyer, & Reder, 1976; Haber & Standing, 1969; Van der Heijden, 1992). Because of the first factor, letter perception becomes worse for letters that are farther from fixation. Because of the second factor, perception of letters flanked by a letter on each side is worse than for letters with only one flanking letter (i.e., the leftmost and rightmost letter in the string). In summary, the reduced crowding as a result of having only one flanking letter compensates for the drop in visual acuity with the outermost letters. This interpretation of the W/M-shaped serial position function ignores, however, one fundamental finding that was first reported by Mason and Katz (1976) and was confirmed by Mason (1982) and Hammond and Green (1982). This finding is that, although letters and digits show an M-shaped search function, other keyboard characters (e.g., %, }, and §) do not. In these three studies, the researchers observed an inverse U-shaped serial position function for search times for symbol stimuli that can be interpreted uniquely in terms of variations in visual acuity. Clearly, if crowding (as defined by Pelli, Palomares, & Majaj, 2004) is a key ingredient of the M-shaped search function for letters, then symbol stimuli do not appear to be affected by crowding in the same manner as letter stimuli. Mason (1982) concluded that the absence of an advantage for outer positions (i.e., the first and last positions in the string) for symbol stimuli is strong evidence that the outer letter advantage observed in the processing of letter strings is not due to sensory mechanisms, such as lateral interference (crowding). She argued, instead, that these end-letter effects likely reflect top-down mechanisms specific to the processing of stimuli that typically occur in strings (i.e., letters and digits, but not symbols). Biases in the deployment of spatial attention could provide one such mechanism. Indeed, greater allocation of attention to the periphery might compensate for perceptual limitations (lower visual acuity) at these locations (Krumhansl & Thomas, 1976). A periphery-to-center scanning mechanism had already been proposed to account for the end-letter advantage (Bouma, 1973; White, 1976) and has been repeatedly proposed as the general mechanism behind a so-called ends-first hypothesis of letter string processing (Butler, 1975; Butler & Merikle, 1973; Krumhansl & Thomas, 1976; Merikle, 1974; Merikle & Coltheart, 1972; Merikle, Coltheart, & Lowe, 1971; White, 1976). In a simple version of this scanning explanation, outer letters would be processed first and would therefore be more rapidly and easily identified compared with inner letters. Combined with the effects of visual acuity, this scanning mechanism would then generate the W/M-shaped serial position function for letters and digits. Symbols, on the other hand, would show effects only of visual acuity. A parallel-processing version of this hypothesis might appeal to biases in attentional deployment that would improve processing of letters in the outer positions of strings (e.g., Rumelhart & McClelland, 1982). However, before speculating further on the origins of the endletter advantage and why it is not observed with strings of symbol stimuli, we decided to first provide further empirical support for the critical findings of Mason (1982) and Hammond and Green (1982). Perhaps these results are, for some unknown reason, specific to the target search task used in these experiments. Searching for a symbol in a string of symbols might invoke a different search 481 strategy compared with looking for letters or digits in strings of letters and digits, especially because stimulus category was blocked in these studies. The basic question guiding the present research is therefore whether distinct serial position functions would be found for symbol stimuli compared with letters and digits in an identification paradigm. To address this issue, the present study used data-limited identification accuracy as the dependent measure. Experiments 1– 4 used two-alternative forced choice (2AFC), whereas Experiment 5 used a postcued partial report bar-probe procedure (a probe indicated the position where item identity was to be reported). The 2AFC procedure was popularized by Reicher (1969) and Wheeler (1970) in their adaptation of the paradigm to study the word superiority effect while controlling for possible effects of differential memory loss during response read out and pure guessing strategies (McClelland & Rumelhart, 1981; Rumelhart & McClelland, 1982). For the present study, the 2AFC task allowed us to control for a possible influence of the number of elements in each stimulus category (not the actual number that are tested) that differed for letters, digits, and symbols. Category size is more likely to influence performance in an unconstrained partial report procedure than in 2AFC. Furthermore, 2AFC controls for effects of positional uncertainty that arise when a participant has identified the target character but incorrectly reports a character from the wrong position. The possible influence of positional uncertainty was examined in Experiment 5 with the partial report bar-probe procedure, and Experiment 6 provided a direct comparison of performance with the 2AFC and bar-probe techniques. Anticipating the results, in Experiments 1– 4, we found distinct serial position functions for letters and digits compared with symbol stimuli, and these functions were principally driven by a strong initial-position advantage for letters and digits. We did not observe the standard final-position advantage for letter stimuli when the 2AFC procedure involved presentation of the two alternatives above and below the target immediately following stimulus offset. A final-position advantage was found for letters and digits, but not for symbols, when a postcue stimulus replaced the two alternatives in Experiments 5 and 6. We explain how hypothesized differences in the size and shape of receptive fields of retinotopic character detectors can provide a unified account of the complete pattern of results. Experiment 1 Prior research examining serial position effects with the 2AFC procedure has exclusively used four-letter strings. These studies typically reported U-shaped functions with optimal performance on outer letters (e.g., Jordan & Bevan, 1996; Jordan, Patching, & Milner, 2000; Jordan, Patching, & Thomas, 2003). The aim of Experiment 1 was, therefore, to examine serial position functions for letter, digit, and symbol stimuli in a data-limited identification task with 2AFC. Figure 1 illustrates the 2AFC and bar-probe procedures used in the present study. Method Participants. Twenty-four students (3 men and 21 women) at the University of Provence, Marseille, France, were paid €5 for their participation in the experiment. Their mean age was 20 years, and they all reported having normal or corrected-to-normal vision. 482 TYDGAT AND GRAINGER Figure 1. Illustration of the paradigms used in the present study. A pattern mask plus fixation bars were always followed by a stimulus presented briefly (for 100 ms), which was followed, in turn, by a pattern mask accompanied either by two alternatives (Experiments 1– 4) or by two hyphen marks (Experiments 5– 6) located above and below a specific location in the string. The two-alternative forced-choice paradigm of Experiments 1– 4 is illustrated in the left three schemas (Experiments 1 and 2 in the first schema, Experiment 3 in the second, and Experiment 4 in the third). The bar-probe procedure of Experiments 5 and 6 is shown in the rightmost schema. Stimuli and design. All stimuli consisted of horizontal arrays of five characters. Three different types of stimuli were used: consonant letters presented in uppercase (B, D, F, G, K, N, L, S, and T), digits (1 to 9), and symbols (%, /, ?, @, }, ⬍, £, §, and ). These three categories were assigned to three blocks, so that one block consisted of 90 letter-array stimuli, one of 90 digit-array stimuli, and one of 90 symbol-array stimuli. Although the presentation order of the blocks was counterbalanced among participants, all other factors were manipulated as within-subjects variables. These were (a) target type (letter, digit, or symbol) and (b) target position in the array (Positions 1–5). Each array consisted of a quasi-random sequence of characters, with each of the target characters being presented 2 times at each of the five target positions and 40 times at a nontarget position. Thus, accuracy for each type of target at each target position was based on 18 observations per participant. For the purposes of the forced-choice task, each target character was paired with an alternative character so that a target character was always presented with the same alternative at each of the five positions. Each target character served as the alternative for one other character. The incorrect alternative presented for forced choice was never present in the stimulus array. The correct alternative appeared once above the backward mask (with the incorrect alternative below) and once below the backward mask (with the incorrect alternative above) for each target position. Procedure. The experiment was run inside a dimly lit room and was controlled with E-Prime software (Psychology Software Tools, www.pstnet.com/eprime). Participants were seated in front of a computer screen at a viewing distance of approximately 60 cm. At that distance, target stimuli in the five different positions were located at ⫺1.2°, ⫺0.6°, 0°, 0.6°, and 1.2° of visual angle from the central fixation point, and each letter, digit, or symbol character subtended on average 0.44° of visual angle. Stimuli were displayed in white on a black background and were presented in 18-point Courier New font. Participants first saw instructions and practiced the task for 81 trials to become familiarized with the procedure and the stimuli. Each trial began with two vertical fixation bars, placed above and below the center of a forward mask. The forward mask consisted of five hash marks and stayed on the screen for 515 ms. Then the fixation bars and the mask disappeared, and the array of five characters immediately appeared for a duration of 100 ms. This was followed by a backward mask, which was identical to the forward mask and was accompanied by two characters, one above the mask and one below at one of the five possible positions. Participants had to decide which one of these two characters was present in the corresponding position of the preceding array. They were asked to respond as accurately as possible by pressing one of two keys on the keyboard. They had to choose either the upward arrow key (for the alternative above) or the downward arrow key (for the alternative below). After the response, the screen was cleared, and the two fixation bars appeared on the screen. There was a delay of 515 ms before the next trial started. The participants could take a short break between the three blocks of 90 trials each. The experiment lasted approximately 30 min. Results Mean accuracies for all target types and target positions are presented in Figure 2. An analysis of variance (ANOVA) was performed on the data with target type (letter, symbol, or digit) and target position (1–5) as within-subjects variables. In the item analysis, target type was a between-items variable, and target position was a within-items variable. In this and each of the following experiments, the Greenhouse and Geisser (1959) correction was applied to determine the significance values for variables with more than two levels. The main statistical results are presented in Table 1. Symbol targets were identified at slightly lower levels of accuracy than letter and digit targets (respectively, 57.6%, 59.9%, and 60.1%). Accuracy varied significantly as a function of serial position, and the serial position function for symbol stimuli differed from that found for letters and digits (Target Type ⫻ Target Position interaction). As can be seen from Figure 2, this interaction was mostly driven by differences in performance at the first position in strings. In line with this observation, the critical Target Type ⫻ Target Position interaction was significant when considering only the first and second positions in strings and was not significant when considering only the fourth and fifth positions in strings. Following Mason (1982) and Hammond and Green (1982), we also performed trends analyses to examine the best fitting serial position functions for the different types of stimuli. Both significant quartic and linear effects were found for letter stimuli, F1(1, 18) ⫽ 21.58, p ⬍ .001; F2(1, 8) ⫽ 13.34, p ⬍ .01, and F1(1, 18) ⫽ 8.71, p ⬍ .01; F2(1, 8) ⫽ 15.47, p ⬍ .01, respectively. These effects explained 95% of the observed variance, with 54% due to a quartic function alone (W shape) and 41% due to a linear function. For the symbol characters, there was a significant qua- SERIAL POSITION EFFECTS 483 Figure 2. Serial position functions for letters, symbols, and digits in Experiment 1 in which the different stimulus categories were presented in different blocks. Data are percentage correct for the two-alternative forced-choice task, and vertical bars represent standard errors. dratic effect (inverted U shaped), F1(1, 18) ⫽ 10.29, p ⬍ .01; F2(1, 8) ⫽ 18.68, p ⬍ .01, which explained 86% of the observed variance. For the digit characters, the effects were comparable to those of letters. There was a significant quartic effect, F1(1, 18) ⫽ 40.71, p ⬍ .001; F2(1, 8) ⫽ 25.93, p ⬍ .001, and a smaller linear effect, F1(1, 18) ⫽ 8.31, p ⬍ .01; F2(1, 8) ⫽ 13.40, p ⬍ .01. These two effects explained 85% of the variance, with 64% for the quartic function and 21% for the linear function. Discussion The results of Experiment 1 clearly demonstrate that the different serial position functions for letter, digit, and symbol stimuli reported by Mason (1982) and Hammond and Green (1982) are not specific to the target search task. Experiment 1 tested letter, digit, and symbol stimuli in a data-limited identification paradigm with 2AFC and found serial position functions that differed as a function of stimulus type. Letters and digits showed a distinct advantage for the first and central positions, with variance across position being best captured by a quartic function. Symbols, on the other hand, showed an inverted U-shaped function, with variance across position being best captured by a quadratic function. Experiment 2 In Experiment 1, the bulk of the difference between symbol stimuli, on the one hand, and letters and digits, on the other, was carried by performance for items at the beginning of strings. One possible account of this initial-position advantage is that the pres- ence of strings of letters or digits generates an endogenous bias toward the beginning of these strings. This would provide a mechanism for initiating a left-to-right attentional scanning of such stimuli (e.g., Mewhort & Popham, 1991). This attentional bias could be driven by the expectation that the stimuli to be processed are strings of letters or digits, and it would not occur when the expected stimuli are symbols. This type of strategy would be encouraged by the blocked presentation of stimulus type in prior studies that have used the target search paradigm and in Experiment 1 of the present study. Participants knew in advance what kind of stimulus they would receive and could therefore adjust endogenous attention as a function of the expected stimulus type. In Experiment 2, we tested this hypothesis by randomly intermixing the three types of stimuli during the experiment. This manipulation should cancel the initial-position advantage found for letters and digits and should produce similar serial position functions for all three types of stimuli. Method Participants. Twenty-four students (7 men and 17 women) at the University of Provence, Marseille, volunteered to participate in the experiment. Their mean age was 21 years, and all reported having normal or corrected-to-normal vision. Stimuli and design. Stimuli and design were exactly the same as in Experiment 1. Procedure. The procedure was also the same as in Experiment 1, except that the three blocks of 90 trials now contained a random Table 1 Main Statistical Results for Experiment 1 Effect F1 2 F2 2 Target type Target position Target Type ⫻ Target Position Only first and second positions: Target Type ⫻ Target Position Only fourth and fifth positions: Target Type ⫻ Target Position F1(2, 36) ⫽ 3.54ⴱ F1(4, 72) ⫽ 7.85ⴱⴱⴱ F1(8, 144) ⫽ 3.86ⴱⴱⴱ 0.01 0.12 0.05 F2 ⬍ 1 F2(4, 96) ⫽ 17.46ⴱⴱⴱ F2(8, 96) ⫽ 3.47ⴱⴱ 0.07 0.03 F1(2, 36) ⫽ 6.75ⴱⴱ 0.07 F2(2, 24) ⫽ 5.88ⴱⴱ 0.04 F1(2, 36) ⫽ 2.13 0.02 F2(2, 24) ⫽ 1.24 0.01 ⴱ p ⬍ .05. ⴱⴱ p ⬍ .01. ⴱⴱⴱ p ⬍ .001. 484 TYDGAT AND GRAINGER mixture of target types (letters, symbols, and digits). This was also the case for the practice trials. attentional capture). Strings of letters and digits might in some way automatically draw attention to their beginnings. Results Experiment 3 Mean accuracies for each target type and target position are presented in Figure 3. The results of a 3 (target type) ⫻ 5 (target position) ANOVA performed on the accuracy data are shown in Table 2. Performance for symbol targets was slightly lower than for letters and digits, with 57.3%, 60.0%, and 61.7% mean accuracy, respectively. Accuracy varied significantly as a function of target position, and the resulting serial position functions differed as a function of target type. This Target Position ⫻ Target Type interaction reflects the superior performance on letter and digit stimuli that was mostly evident at the first position. For letters, there was a significant linear trend in the serial position function, F1(1, 23) ⫽ 17.95, p ⬍ .001; F2(1, 8) ⫽ 12.69, p ⬍ .01, which explained 83% of the observed variance, and there was a smaller quartic effect, F1(1, 23) ⫽ 4.21, p ⬍ .1; F2(1, 8) ⫽ 6.16, p ⬍ .05, which explained about 15% of additional variance. For the symbols, both a quadratic, F1(1, 23) ⫽ 6.95, p ⬍ .05; F2(1, 8) ⫽ 10.83, p ⬍ .05, and a quartic effect, F1(1, 23) ⫽ 6.36, p ⬍ .05; F2(1, 8) ⫽ 6.39, p ⬍ .05, were found, and they explained, respectively, 53% and 44% of the variance. For digits, there was a linear effect, F1(1, 23) ⫽ 17.38, p ⬍ .001; F2(1, 8) ⫽ 32.38, p ⬍ .001, and a quartic effect, F1(1, 23) ⫽ 8.47, p ⬍ .01, F2(1, 8) ⫽ 32.89, p ⬍ .001, which explained 60% and 30% of the variance, respectively. Discussion The results of Experiment 2 provide a replication of Experiment 1 and demonstrate that differences in the serial position function for symbols compared with letters and digits are not affected by randomly intermixing the three types of stimuli. On the contrary, the linear trend for letters and digits found in Experiment 1 was even more exaggerated in Experiment 2. This result allowed us to rule out an endogenous top-down attentional bias as the source of the initial-position advantage in the perception of letters and digits in strings. If some form of attentional mechanism is the basis of the first-position advantage found for letters and digits, then this mechanism must be automatically triggered on presentation of a string of letters or digits on each trial (i.e., bottom-up exogenous Experiment 3 provided a further test of the attentional bias hypothesis. If this bias is automatically triggered on presentation of a string of letters or digits, even when these trials are intermixed with symbol strings, then one ought to be able to remove the initial-position advantage by placing letter targets in a string of symbols. The assumption here is that it is the dominant stimulus type that should determine the nature of bottom-up processing of the whole string (i.e., four symbols with one embedded letter should not trigger an attentional bias). Therefore, single letters embedded in a string of symbols should not show an initialposition advantage, whereas symbol targets embedded in a string of letters should show the advantage. Experiment 3 also allowed us to test an alternative explanation of the first-position advantage found with letters and digits expressed in terms of a chunking mechanism that forms higher order combinations of items of the same category, with the higher order combinations being more resistant to masking than individual items. This chunking mechanism would be a basic component of the general process of word and number recognition and would therefore operate only on letter and digit strings and not on symbol strings (if we assume that there are no higher order symbol combinations stored in long-term memory). The first-position advantage found for letters and digits could then be due to the precise nature of this chunking process, with initial letters and digits playing a dominant role (e.g., Whitney, 2001). By embedding target letters in symbol strings in Experiment 3, we expected to block any influence of this hypothesized chunking mechanism, because letters and digits would not be able to combine with other letters and digits. Method Participants. Twenty-four students (2 men and 22 women) at the University of Provence, Marseille, volunteered to participate in the experiment. Their mean age was 20 years, and all reported having normal or corrected-to-normal vision. Stimuli and design. Target stimuli were the same as in the previous experiments, except that there were no digit stimuli; Figure 3. Serial position functions for letters, symbols, and digits in Experiment 2 in which the different stimulus categories were randomly intermixed during the experiment. Data are percentage correct for the two-alternative forced-choice task, and vertical bars represent standard errors. SERIAL POSITION EFFECTS 485 Table 2 Main Statistical Results for Experiment 2 Effect F1 2 F2 2 Target type Target position Target Type ⫻ Target Position Only first and second positions: Target Type ⫻ Target Position Only fourth and fifth positions: Target Type ⫻ Target Position F1(2, 46) ⫽ 6.26ⴱⴱ F1(4, 92) ⫽ 8.80ⴱⴱⴱ F1(8, 184) ⫽ 2.40ⴱ 0.02 0.11 0.03 F2 ⬍ 1 F2(4, 96) ⫽ 16.80ⴱⴱⴱ F2(8, 96) ⫽ 2.29ⴱ 0.19 0.05 F1(2, 46) ⫽ 2.68 0.03 F2(2, 24) ⫽ 4.05ⴱ 0.05 ⴱ p ⬍ .05. ⴱⴱ p ⬍ .01. ⴱⴱⴱ F1 ⬍ 1 F2 ⬍ 1 p ⬍ .001. therefore, there were only 180 trials. Also, the nine target letters were presented in a string of symbols, and the nine target symbol characters were now presented in a string of letters. Procedure. Participants practiced the task during 20 practice trials, randomly chosen out of the stimulus list of 180 trials. All other components of the procedure were exactly the same as in Experiment 2. The experiment lasted approximately 20 min. Results Mean accuracies for each target type and target position are plotted in Figure 4, and the results of a 2 (target type) ⫻ 5 (target position) ANOVA are shown in Table 3. Accuracy for symbol targets (63.1%) was somewhat higher than accuracy for letter targets (57.6%). The main effect of target position was significant, but this did not interact significantly with target type. However, there was a trend toward an interaction effect when only the first and second positions were analyzed. Accuracy for letter targets was higher in Position 1 than Position 2, whereas the opposite was true for symbol targets. For the letter stimuli, the sum of a linear, cubic, and quartic trend in the serial position function explained 97% of the observed variance, with 37%, 36%, and 24% for each trend, respectively. However, none of these trends were significant. For the symbol stimuli, there was a significant linear effect, F1(1, 23) ⫽ 4.74, p ⬍ .05; F2(1, 8) ⫽ 6.39, p ⬍ .05, a less robust quadratic effect, F1(1, 23) ⫽ 3.48, p ⬍ .1; F2(1, 8) ⫽ 6.26, p ⬍ .05, and a less robust quartic effect, F1(1, 23) ⫽ 4.74, p ⬍ .05; F2(1, 8) ⫽ 5.17, p ⬍ .1. These three effects explained, respectively, 27%, 38%, and 24% of the observed variance. Discussion The results of Experiment 3 allowed us to rule out an automatically triggered attentional bias account of the initial-position advantage found for letter-in-string perception compared with symbols. If this attentional bias was triggered only on presentation of a string of letters (or digits)— hence explaining why there is no initial-position advantage for symbols—then we expected to remove the advantage for letter targets by inserting them in strings of symbols, and we expected to create an advantage for symbol targets by inserting them in strings of letters. However, although the results were somewhat weaker in Experiment 3, an ANOVA that tested only the first and second positions did show a trend toward an interaction. This reflected the fact that performance tended to be better at the first position compared with the second position for letter targets, whereas symbol targets showed a difference in the opposite direction. An inspection of Figure 4 shows that the general pattern found in Experiments 1 and 2 was replicated in Experiment 3. Figure 4. Serial position functions in Experiment 3 for letter targets embedded in symbol arrays and symbol targets embedded in letter arrays. Data are percentage correct for the two-alternative forced-choice task, and vertical bars represent standard errors. TYDGAT AND GRAINGER 486 Table 3 Main Statistical Results for Experiment 3 Effect F1 2 F2 2 Target type Target position Target Type ⫻ Target Position Only first and second positions: Target Type ⫻ Target Position Only fourth and fifth positions: Target Type ⫻ Target Position F1(1, 23) ⫽ 14.83ⴱⴱⴱ F1(4, 92) ⫽ 3.56ⴱ F1(4, 92) ⫽ 1.50 0.05 0.05 0.02 F2(1, 16) ⫽ 2.24 F2(4, 64) ⫽ 3.07ⴱ F2(4, 64) ⫽ 1.25 0.06 0.07 0.03 F1(1, 23) ⫽ 3.74 0.03 F2(1, 16) ⫽ 3.03 0.03 ⴱ p ⬍ .05. ⴱⴱⴱ F1 ⬍ 1 F2 ⬍ 1 p ⬍ .001. This pattern was further confirmed by a combined analysis of Experiments 2 and 3, including only the data for letter and symbol targets from Experiment 2. The results of this joint ANOVA are shown in Table 4. There was no sign of a Target Position ⫻ Experiment interaction. Most important, the critical Target Type ⫻ Target Position interaction was significant in the combined analysis, and once again this interaction was shown to be carried completely by the difference in performance across Positions 1 and 2. There was an initial-position advantage (relative to the second position) for letter targets and a disadvantage for symbol targets; these were independent of the nature of the other items in the string. Experiment 3 also allows us to reject a chunking account of the different serial position functions for letters and symbols. According to this account, the critical difference between a string of letters and a string of symbols is that only the former are automatically subject to a chunking process whereby higher order letter combinations are formed. We predicted that embedding letter targets in strings of symbols would disrupt such a chunking process and remove any differences in performance for letter and symbol targets. The results of Experiment 3 do not support this prediction. It would appear that some other mechanism, also specific to the processing of stimuli that typically appear in strings, is the source of the distinct serial position functions obtained for letters and symbols. Experiment 4 Experiments 1–3 further confirmed the seminal findings of Mason (Mason, 1982; Mason & Katz, 1976) and Hammond and Green (1982) that symbol stimuli show distinct serial position functions compared with both letters and digits. The most parsimonious explanation for this systematic pattern of effects is that letters and digits benefit from a specialized processing system that endows an advantage for elements at the first position of a string. Therefore, a combination of visual acuity and an initial-position advantage for letters and digits provides a complete account of the pattern of results found in Experiments 1–3. According to this account of the results of Experiments 1–3, if we remove the central character in the stimuli tested in the previous experiments, then letter stimuli should show improved performance at Position 4 as opposed to Position 2, because Position 4 is now at the beginning of a string of two letters. On the other hand, symbol stimuli should not be affected by this manipulation, because visual acuity should not be affected by removing the central character. To test this, Experiment 4 examined identification of letters and symbols in conditions that were identical to those of Experiment 2, except that the central letter/symbol was removed. Therefore, on every trial, participants were presented with four letters or four symbols, two to the left of fixation and two to the right of fixation, in Positions 1, 2, 4, and 5 of the previous experiments. Method Participants. Twenty-four students (6 men and 18 women) at the University of Provence, Marseille, volunteered to participate in the experiment. Their mean age was 21 years, and all reported having normal or corrected-to-normal vision. Stimuli and design. Stimuli were the same as in Experiment 2, although there were no digit characters, and no character was Table 4 Main Statistical Results for the Combined Analysis of Experiments 2 and 3 F2 2 0.01 0.02 F2(1, 16) ⫽ 1.24 F2 ⬍ 1 F2(4, 64) ⫽ 12.55ⴱⴱⴱ F2 ⬍ 1 F2(4, 64) ⫽ 3.63ⴱⴱ F1(4, 184) ⫽ 1.79 0.01 F2(4, 64) ⫽ 1.22 0.02 F1(1, 46) ⫽ 7.19ⴱ 0.03 F2(1, 16) ⫽ 8.21ⴱ 0.04 Effect F1 Experiment Target type Target position Experiment ⫻ Target Position Target Type ⫻ Target Position Experiment ⫻ Target Type ⫻ Target Position Only first and second positions: Target Type ⫻ Target Position Only fourth and fifth positions: Target Type ⫻ Target Position F1 ⬍ 1 F1(1, 46) ⫽ 2.00 F1(4, 184) ⫽ 9.25ⴱⴱⴱ F1 ⬍ 1 F1(4, 184) ⫽ 3.53ⴱⴱ ⴱ p ⬍ .05. ⴱⴱ p ⬍ .01. ⴱⴱⴱ p ⬍ .001. F1 ⬍ 1 2 0.00 0.07 F2 ⬍ 1 0.10 0.03 SERIAL POSITION EFFECTS presented at the third position. Therefore, targets could appear at four possible positions, two to the left and two to the right of fixation, and on each trial, four letters or four symbols appeared at these four locations. In this design, target position is defined as the combination of two variables: visual field (left vs. right) and target position (first vs. second). Procedure. The procedure was the same as in Experiment 2, except for the removal of the central character in the stimulus strings and the pattern mask. Participants were first presented with a practice block of 20 randomly chosen trials, followed by the 144 experimental trials. The experiment lasted about 20 min. Results and Discussion The mean accuracies per experimental condition are given in Table 5. The results of a 2 (target type) ⫻ 2 (target position) ⫻ 2 (visual field) ANOVA are shown in Table 6. Mean accuracy for letters was 63.5%, and accuracy for symbols was 60.1%. There was a significant triple interaction reflecting the fact that the Target Type ⫻ Target Position interaction was significant in the left visual field (LVF) but not in the right visual field (RVF). In line with the results of Experiments 1–3, the results of Experiment 4 showed once again a strong interaction in the identification accuracy for letters and symbols at Serial Positions 1 and 2 in the LVF. Although overall performance was lower for symbols compared with letters, the pattern of results for targets presented to the right of fixation was almost identical for letters and symbols, with a 10% advantage for the innermost target. Figure 5 shows the results of Experiments 2 and 4 plotted together. To examine the influence of removing the central character on performance for letters and symbols, we performed a series of planned comparisons (Experiment 2 vs. Experiment 4) for each serial position. For letter stimuli, there was a significant effect at Position 1 in the RVF, F1(1, 46) ⫽ 10.97, p ⬍ .01, 2 ⫽ 0.19; F2(1, 16) ⫽ 6.99, p ⬍ .05, 2 ⫽ 0.30. For symbol stimuli, on the other hand, there was a significant difference at both Position 2 in the LVF, F1(1, 46) ⫽ 5.46, p ⬍ .05, 2 ⫽ 0.11; F2(1, 16) ⫽ 4.93, p ⬍ .05, 2 ⫽ 0.24, and Position 1 in the RVF, F1(1, 46) ⫽ 6.01, p ⬍ .05, 2 ⫽ 0.12; F2(1, 16) ⫽ 4.63, p ⬍ .05, 2 ⫽ 0.22. In line with the predictions of the initial-position advantage for letter stimuli, performance at Position 4 (innermost letter in the RVF) was significantly greater in Experiment 4 than in Experi- Table 5 Results (Percentage Correct Identification) in Experiment 4 for Letter and Symbol Targets Accuracy Letters Symbols Visual field/position % SE % SE Left/1 Left/2 Right/1 Right/2 69.9 63.4 65.3 56.5 2.9 2.7 2.0 2.3 57.4 66.7 63.2 53.2 2.7 2.4 2.6 2.5 Note. Targets were presented at the same positions as the first, second, fourth, and fifth positions in Experiments 1–3, thus giving two positions in the left visual field and two positions in the right visual field. 487 ment 2, whereas performance at all other positions remained statistically equivalent across experiments. However, there was also a significant increase in performance at Position 2 (innermost letter in the LVF) and Position 4 for symbol stimuli, which was not predicted by this account. Therefore, it appears from the combined results of Experiments 2 and 4 that a simple combination of visual acuity and an initial-position advantage for letters cannot provide a satisfactory account of performance for letters and symbols in these two experiments. This combined analysis clearly suggests that the presence of a symbol character at the central location in Experiment 2 exerted an inhibitory influence on the identification of symbols presented at Positions 2 and 4. This, therefore, reinstates crowding as one major factor determining the visibility of characters presented in strings and further raises the possibility that differential effects of crowding might underlie the different serial position functions for letters, digits, and symbols. The results of Experiment 4 further suggest that visual field differences in crowding effects might be part of the overall story. Experiment 5 One critical difference with respect to prior observations of outer-position effects for letters is that the bulk of the advantage was carried by the first letter in our experiments, with little evidence for a final-letter advantage.2 This result is all the more important given the nature of the paradigm we used, which, if anything, should have reduced sequential, beginning-to-end biases compared with the letter search task. Experiment 5 was designed to examine one possible reason for this divergence with respect to prior research on serial position functions for letter stimuli in data-limited identification tasks. The 2AFC task used in Experiments 1– 4 has the advantage of eliminating errors that occur when the target item has been identified but another item in the string is reported (location errors). These are eliminated in our 2AFC procedure because the alternative item provided for response is never present in the string. Thus, the 2AFC procedure provides a means of estimating how well a given item has been identified at a given position in the absence of contamination from such location errors. This implies that the first-position advantage that we have systematically found for letters and digits, but not for symbols, was not due to improved encoding of positional information for letters and digits that appear at the beginning of strings. On the other hand, the final-position advantage found for letter and digit stimuli in prior research, but not observed in Experiments 1– 4, could well have been due to a larger number of location errors when identifying interior letters/ digits than final letters/digits. There is evidence from experiments using the bar-probe technique that this is, indeed, the case. First, it has been shown that use of a backward mask (as was the case in the present experiments) has a greater influence on accuracy of report from inner positions compared with outer positions (Mewhort & Campbell, 1978). Second, it has been shown that the 2 It should nevertheless be noted that one recent study of serial position effects in the target search paradigm (Pitchford et al., 2008) did not find such a pronounced final-position advantage as in the earlier work of Mason (1982). The results of Pitchford et al. (2008) suggested that positional letter frequency might be one factor determining the size of outer-position effects. TYDGAT AND GRAINGER 488 Table 6 Main Statistical Results for Experiment 4 Effect Target type Target position Visual field Target Type ⫻ Target Position Target Type ⫻ Visual Field Target Position ⫻ Visual Field Target Type ⫻ Target Position ⫻ Visual Field Left visual field: Target Type ⫻ Target Position Right visual field: Target Type ⫻ Target Position ⴱ p ⬍ .05. ⴱⴱ p ⬍ .01. 2 F1 ⴱⴱⴱ F1(1, 23) F1(1, 92) F1(1, 23) F1(1, 23) F1 ⬍ 1 F1(1, 23) ⫽ ⫽ ⫽ ⫽ 4.99ⴱ 8.70ⴱⴱ 3.97 7.13ⴱ F1(1, 23) ⫽ 7.12ⴱ F1(1, 23) ⫽ 14.23 0.04 F2 ⬍ 1 F2(1, 16) F2(1, 16) F2(1, 16) F2 ⬍ 1 F2(1, 16) 0.03 F2(1, 16) ⫽ 12.13ⴱⴱ 0.02 0.02 0.03 0.02 ⫽ 11.73ⴱⴱ ⴱⴱⴱ F1 ⬍ 1 2 F2 0.09 ⫽ 6.51ⴱ ⫽ 6.57ⴱ ⫽ 5.43ⴱ 0.03 0.04 0.02 ⫽ 19.69ⴱⴱⴱ 0.05 F2(1, 16) ⫽ 20.44 ⴱⴱⴱ 0.03 0.12 F2 ⬍ 1 p ⬍ .001. use of a backward mask increases the number of location errors as opposed to intrusion (item) errors in the bar-probe identification task (Mewhort & Campbell, 1978; Mewhort, Campbell, Marchetti, & Campbell, 1981; Mewhort, Huntley, & Duff-Fraser, 1993). If the final-position advantage found in studies using a partial report bar-probe procedure was due to the larger number of positional errors for inner positions compared with the final position, then we would expect to observe this advantage with exactly the same stimuli and procedure as in Experiment 2, but with the bar-probe partial report procedure replacing the 2AFC procedure. Method Participants. Twenty-four students (4 men and 20 women) at the University of Provence, Marseille, were paid 5€ to participate in the experiment. Their mean age was 23 years, and all reported having normal or corrected-to-normal vision. Stimuli and design. Stimuli and design were exactly the same as in Experiment 2. Procedure. The procedure was identical to that of Experiment 2, except for a change in the report procedure. The 2AFC task was replaced by a bar-probe partial report task. The backward mask was accompanied by two hyphen marks positioned above and below one of the five positions in the string (hence mimicking the position of the two alternative letters in the 2AFC procedure). Participants were asked to decide as accurately as possible which character they had seen at this position, by choosing from nine letters, nine digits, and nine symbols indicated on the keyboard in front of the participant. They responded by pressing one of these 27 marked response keys on the keyboard. Results Two participants had to be excluded from the analyses because they had not followed the task instructions. For the remaining 22 participants, mean accuracy for each target type and target position is presented in Figure 6. A 3 (target type) ⫻ 5 (target position) ANOVA was performed on the accuracy data. The main statistical results are presented in Table 7. Performance for symbol targets was lower than for letter and digit targets, with 36.8%, 60.7%, and 61.0% mean accuracy, respectively.3 Target position significantly influenced performance and interacted significantly with target type, reflecting the different serial position function obtained for symbol stimuli compared with both letters and digits. The Target Type ⫻ Target Position interaction was significant in an analysis restricted to the first and second positions and in an analysis with only the fourth and fifth positions. Trends analyses were performed to examine the best fitting serial position functions for the different types of stimuli. For letters, there was a significant linear effect, F1(1, 21) ⫽ 4.61, p ⬍ .05; F2(1, 8) ⫽ 4.67, p ⬍ .1, quadratic effect, F1(1, 21) ⫽ 15.39, p ⬍ .001; F2(1, 8) ⫽ 21.18, p ⬍ .01, and quartic effect, F1(1, 21) ⫽ 47.59, p ⬍ .001; F2(1, 8) ⫽ 27.47, p ⬍ .001. These effects accounted for 12%, 40%, and 46% of the variance, respectively. For digits, such linear, quadratic, and quartic effects explained 5%, 16%, and 79% of the variance, respectively. Again, all of these functions were significant, with F1(1, 21) ⫽ 4.48, p ⬍ .05; F2(1, 8) ⫽ 12.62, p ⬍ .01, and F1(1, 21) ⫽ 8.17, p ⬍ .01; F2(1, 8) ⫽ 15.32, p ⬍ .01, and F1(1, 21) ⫽ 104.17, p ⬍ .001; F2(1, 8) ⫽ 76.65, p ⬍ .001, respectively. For symbols, we found a cubic effect, F1(1, 21) ⫽ 9.96, p ⬍ .01; F2(1, 8) ⫽ 3.60, p ⬍ .1, and a quartic effect, F1(1, 21) ⫽ 42.26, p ⬍ .001; F2(1, 8) ⫽ 29.68, p ⬍ .001. These functions explained 11% and 76% of the variance, respectively. In the partial report task used in Experiment 5, errors can be categorized as intrusion errors—when the erroneous response was not a member of the stimulus string— or location errors—when the erroneous response was present at another location in the string. 3 Because of the low overall level of accuracy for symbol stimuli, we analyzed the results of a group of 6 participants with the highest performance for symbols. In this group, accuracy for symbol targets at Positions 2, 3, and 4 was statistically equivalent to accuracy with digits and letters at these positions, F1(1, 5) ⫽ 2.72, p ⫽ .16; F2 ⬍ 1. For all positions, the main effect of target type was significant, F1(2, 10) ⫽ 31.90, p ⬍ .001, 2 ⫽ 0.12; F2(2, 24) ⫽ 5.26, p ⬍ .05, 2 ⫽ 0.11, as were the main effect of target position, F1(4, 20) ⫽ 3.22, p ⬍ .05, 2 ⫽ 0.23; F2(4, 96) ⫽ 13.58, p ⬍ .001, 2 ⫽ 0.21, and the interaction, F1(8, 40) ⫽ 4.82, p ⬍ .001, 2 ⫽ 0.10; F2(8, 96) ⫽ 2.86, p ⬍ .01, 2 ⫽ 0.09. The interaction was clearly driven by the lower levels of accuracy for symbol stimuli compared with letters and digits at Position 1, F1(1, 5) ⫽ 52.70, p ⬍ .001, 2 ⫽ 0.91; F2(1, 24) ⫽ 4.35, p ⬍ .05, 2 ⫽ 0.46, and at Position 5, F1(1, 5) ⫽ 11.75, p ⬍ .05, 2 ⫽ 0.70; F2(1, 24) ⫽ 3.66, p ⬍ .1, 2 ⫽ 0.40. SERIAL POSITION EFFECTS 489 Figure 5. Percentage correct for the two-alternative forced-choice task for letter and symbol targets in Experiment 4 (Positions 4 and 5 correspond to Positions 1 and 2 in the right visual field) and for the same positions in Experiment 2. In Experiment 2, there was also a character present at Position 3 in the array, whereas no character was present at this position in Experiment 4. Vertical bars represent standard errors. Expt. ⫽ Experiment. Performance for letter, symbol, and digit targets is broken down by error type in Figure 7. Separate ANOVAs were performed for location errors and intrusion errors (see Table 7). When only location errors were counted as errors, mean accuracies for letters, symbols, and digits were 72.1%, 60.3%, and 70.3%, respectively. When only intrusion errors were counted as errors, mean accuracies for letters, symbols, and digits were 88.5%, 76.5%, and 90.7%, respectively. The main effects and interactions were significant in both analyses. As can be seen in Figure 7, for location errors, the interaction was mostly driven by the lower levels of accuracy for symbols as opposed to letters and digits at Positions 1 and 5. For intrusion errors, one can also note a sizeable difference at Position 2 for symbols relative to letters and digits. Discussion The results of Experiment 5 show that replacing 2AFC with a postcued partial report procedure reinstated the final-letter advantage for letters and digits. These stimuli then generated the classic W-shaped serial position function for letter and digit identification accuracy obtained with the bar-probe procedure (e.g., Haber & Standing, 1969; Merikle, Lowe, & Coltheart, 1971), the equivalent of the M-shaped function typically found for letter and digit detection latencies (e.g., Hammond & Green, 1982; Mason, 1982). Because the only difference between Experiment 2 and Experiment 5 was the switch from a 2AFC procedure to a bar-probe procedure, this suggests that the final-position advantage found in Experiment 5 was due to the larger number of location errors in Position 4 compared with Position 5. Such location errors would not arise in the 2AFC task because the alternative response was never a part of the target string. Furthermore, one would expect a greater number of location errors at inner positions compared with outer positions because the latter have only a single flanking character. An analysis of the different types of error (intrusion vs. location) provided some support for this analysis. Location errors did indeed show a more exaggerated W-shaped serial position function than intrusion errors. However, one aspect of the comparison of Experiments 2 and 5 does not fit an explanation of differences in performance in the 2AFC and bar-probe procedures being uniquely driven by the greater number of location errors in the latter. That aspect is the overall estimated superior performance in the bar-probe task over the 2AFC. To compare performance in the bar-probe task of Experiment 5 to performance in the 2AFC task of Experiment 2, a graph of transformed partial report accuracies was plotted by applying an equation for transforming partial report into approximate 2AFC Figure 6. Serial position functions for letters, symbols, and digits in Experiment 5. Data are percentages correct with a bar-probe partial report procedure, and vertical bars represent standard errors. TYDGAT AND GRAINGER 490 Table 7 Main Statistical Results for Experiment 5 Effect F1 2 F2 2 Target type Target position Target Type ⫻ Target Position Only first and second positions: Target Type ⫻ Target Position Only fourth and fifth positions: Target Type ⫻ Target Position Location errors Target type Target position Target Type ⫻ Target Position Intrusion errors Target type Target position Target Type ⫻ Target Position F1(2, 42) ⫽ 58.47ⴱⴱⴱ F1(4, 84) ⫽ 17.76ⴱⴱⴱ F1(8, 168) ⫽ 13.11ⴱⴱⴱ 0.16 0.17 0.05 F2(2, 24) ⫽ 21.26ⴱⴱⴱ F2(4, 96) ⫽ 30.78ⴱⴱⴱ F2(8, 96) ⫽ 4.89ⴱⴱⴱ 0.26 0.28 0.09 F1(2, 42) ⫽ 21.70ⴱⴱⴱ 0.04 F2(2, 24) ⫽ 8.21ⴱⴱ 0.06 F1(2, 42) ⫽ 24.35ⴱⴱⴱ 0.05 F2(2, 24) ⫽ 9.85ⴱⴱⴱ 0.10 F1(2, 42) ⫽ 19.59ⴱⴱⴱ F1(4, 84) ⫽ 16.90ⴱⴱⴱ F1(8, 168) ⫽ 5.70ⴱⴱⴱ 0.06 0.19 0.04 F2(2, 24) ⫽ 5.91ⴱⴱ F2(4, 96) ⫽ 23.67ⴱⴱⴱ F2(8, 96) ⫽ 2.47ⴱ 0.10 0.32 0.07 F1(2, 42) ⫽ 17.05ⴱⴱⴱ F1(4, 84) ⫽ 6.14ⴱⴱⴱ F1(8, 168) ⫽ 4.05ⴱⴱⴱ 0.17 0.03 0.03 F2(2, 24) ⫽ 19.34ⴱⴱⴱ F2(4, 96) ⫽ 4.42ⴱⴱ F2(8, 96) ⫽ 2.22ⴱ 0.33 0.06 0.06 ⴱ p ⬍ .05. ⴱⴱ p ⬍ .01. ⴱⴱⴱ p ⬍ .001. accuracy.4 Figure 8 compares these transformed data to the results of Experiment 2. If the different serial position functions found with the 2AFC and bar-probe procedures were due to a greater number of location errors in the bar-probe task, then a decrease in performance relative to the 2AFC data of Experiment 2 would be expected, yet the opposite pattern was found. This suggests that some other factor caused the shift in performance for final letters and digits when switching from a 2AFC to a bar-probe partial report procedure. One possibility is that the alternative items presented simultaneously with the backward mask in Experiments 1– 4 affected performance in this task. This could arise through meta-contrast masking (Breitmeyer & Ogmen, 2000) or through some other form of lateral interference or crowding (Huckauf & Heller, 2004), with the two alternative characters causing an additional interference effect adding to that of the backward mask. The interfering effect of two characters would be greater than that caused by the two bar-probe stimuli used in Experiment 5 because of the greater visual complexity of the characters and their closer spatial proximity to the target item (see Figure 1). This would therefore account for the overall lower levels of performance in Experiment 5 compared with Experiment 2. iment 6, and one within-subjects variable was added. This additional variable consisted of the two different report tasks. In one block of the experiment, a 2AFC task was used. In the other block of the experiment, a partial report task was used. The order of these blocks was counterbalanced across participants. Procedure. The procedure was almost identical to that of Experiment 5, except for the use of two different report tasks. The backward mask was always accompanied by two hyphen marks positioned above and below one of the five positions in the string. On the sixth line below the backward mask, the task was indicated on each trial. In the 2AFC block of the experiment, the task indication consisted of two response alternatives, with the word ou (French for or) in between (e.g., T ou B). Participants responded by pressing either the leftward arrow key (for the alternative on the left) or the rightward arrow key (for the alternative on the right) on the keyboard. In the partial report block of the experiment, participants saw the task indication quoi (French for what) and had to choose among the letters and symbols indicated on the keyboard. Participants responded by pressing 1 of 18 marked response keys on the keyboard. At the start of every task block, participants practiced the (2AFC or partial report) task during 20 trials. Both blocks consisted of 180 experimental trials. The experiment lasted approximately 30 min. Experiment 6 Experiment 6 was designed to test the interference hypothesis by presenting the two alternative letters in the 2AFC task at a different location at the same time as a bar probe indicating the location for report. Participants were tested with both procedures (2AFC and bar probe) in two different blocks, and the same bar-probe stimulus was used to indicate position of report in both procedures. Method Participants. Twenty-two students (5 men and 17 women) at the University of Provence, Marseille, were paid 5€ to participate in the experiment. Their mean age was 21 years, and all reported having normal or corrected-to-normal vision. Stimuli and design. Stimuli and design were the same as in Experiment 2, except that there were no digit characters in Exper- Results One participant had to be excluded from the analyses because of very low performance. For the remaining 21 participants, mean accuracy for each target type, target position, and report task is presented in Figure 9. The results of a 2 (target type) ⫻ 5 (target position) ⫻ 2 (report task) ANOVA on the accuracy data are shown in Table 8. Letter targets were identified more accurately 4 Performance on 2AFC can be estimated from partial report performance by assuming that in the 2AFC task, participants either identify the target and respond correctly or fail to identify the target and guess. Under these assumptions, 2AFC performance (FC) is calculated from percentage correct partial report performance (PR) with the following equation: FC ⫽ PR ⫹ [(100 ⫺ PR)/2]. SERIAL POSITION EFFECTS 491 Figure 7. Serial position functions for letters, symbols, and digits in Experiment 5 after categorizing errors into location errors (left) and intrusion errors (right). Vertical bars represent standard errors. than symbol targets, with 62.5% and 51.4% correct identifications, respectively. There was a main effect of target position, and performance was higher with the 2AFC procedure compared with (uncorrected) partial report. The three-way interaction and all two-way interactions were significant. The interactions with task simply reflect the fact that all effects were exaggerated in the partial report task given that the possible range of accuracy was twice as large with this procedure. In both tasks, there was a significantly more pronounced W-shaped serial position function for letter targets compared with symbol targets, and this pattern was more exaggerated with the partial report procedure. Trends analyses of the 2AFC data for letter targets revealed a significant linear effect, F1(1, 20) ⫽ 4.58, p ⬍ .05; F2(1, 8) ⫽ 6.48, p ⬍ .05, quadratic effect, F1(1, 20) ⫽ 4.80, p ⬍ .05; F2(1, 8) ⫽ 10.87, p ⬍ .05, and quartic effect, F1(1, 20) ⫽ 39.87, p ⬍ .001; F2(1, 8) ⫽ 41.10, p ⬍ .001. These accounted for 12%, 16%, and 72% of the variance, respectively. For symbols in the 2AFC task block, the quadratic and quartic functions explained the main part of the observed variance, with 49% and 48%, respectively, but these trends were not significant. For the partial report data with letter targets, there was a significant linear effect, F1(1, 20) ⫽ 7.77, p ⬍ .05; F2(1, 8) ⫽ 6.48, p ⬍ .05, a significant quadratic effect, F1(1, 20) ⫽ 50.24, p ⬍ .001; F2(1, 8) ⫽ 30.01, p ⬍ .001, and a significant quartic effect, F1(1, 20) ⫽ 77.42, p ⬍ .001; F2(1, 8) ⫽ 50.43, p ⬍ .001. These trends explained 10%, 49%, and 38% of the variance, respectively. For symbols, there was a significant quartic effect, F1(1, 20) ⫽ 21.66, p ⬍ .001; F2(1, 8) ⫽ 26.08, p ⬍ .001, which accounted for about 90% of the observed variance. The errors in the partial report task can again be categorized into intrusion and location errors. Performance for letter and symbol targets is broken down by error type in Figure 10, and the results of separate ANOVAs are shown in Table 8. When only location errors counted as errors, mean accuracies for letters and symbols were 69.4% and 62.1%, respectively. The pattern was the same as the overall analysis with the critical Target Type ⫻ Target Position interaction being driven largely by the lower levels of accuracy for symbols as opposed to letters at Positions 1 and 5. When only intrusion errors counted as errors, mean accuracy for letter targets was greater than for symbol targets, with 86.6% and 77.0% correct identifications, respectively. The effects of target position were less pronounced, but the critical interaction with target type was significant and was again due to the superior performance for letters compared with symbols at Positions 1 and 5. Discussion The results of Experiment 6 are clear-cut and provide a solution for integrating the apparently diverging results obtained in the previous experiments. Performance on the bar-probe procedure was an almost exact replication of performance in Experiment 5, Figure 8. Comparison of the transformed PR data of Experiment 5 with the 2AFC results of Experiment 2. PR data are transformed into 2AFC data by applying the equation described in footnote 4. PR ⫽ partial report; Expt. ⫽ Experiment; 2AFC ⫽ two-alternative forced choice. TYDGAT AND GRAINGER 492 Figure 9. Serial position functions for letters and symbols in Experiment 6 with a 2AFC procedure (left) and a PR procedure (right). In both cases, position of report was indicated by bar probes. Vertical bars represent standard errors. 2AFC ⫽ two-alternative forced choice; PR ⫽ partial report. thus demonstrating the robustness of the effects. Performance on the 2AFC procedure, on the other hand, differed from the pattern seen in Experiments 1–3, because there was a clear final-position advantage for letter stimuli. Because the only procedural difference in Experiment 6 was the way in which the two alternative letters were presented, this strongly suggests that interference from the alternative letters was the critical factor at play. Therefore, the absence of a final-position advantage for letter stimuli in Experiments 1–3 was likely due to the extra interference generated by the two alternative letters at this specific position. In the General Discussion, we describe in detail exactly how one simple mechanism can account for this surprising selective effect of interference from the two alternatives in the 2AFC task. Furthermore, in the absence of interference from the alternative letters, performance was very similar across the two procedures when applying the equation for transforming partial report into approximate 2AFC accuracy described in footnote 4. Figure 11 shows the transformed bar-probe data in comparison with the 2AFC data. There was still a small advantage for the partial report procedure, but this can be accounted for by assuming that partic- ipants also selected randomly from the nine alternative letters or symbols when they could not identify the target but knew which category it belonged to. This small advantage for performance with the partial report procedure appears to have been greatest for performance at the first position. An analysis of the different types of error (location vs. intrusion; see Figure 10) suggests that this might have been due to the reduced number of location errors at this position in the partial report bar-probe task. General Discussion In the present study, we examined serial position functions for letters, digits, and symbols, using data-limited identification paradigms. Our results confirm previous results obtained with a response-limited target search task (Hammond & Green, 1982; Mason, 1982; Mason & Katz, 1976), showing distinct serial position functions for symbol stimuli compared with both letters and digits. These results strongly suggest that the distinct serial position functions reflect fundamental differences in how strings of Table 8 Main Statistical Results for Experiment 6 Obtained With the Two-Alternative Forced-Choice (2AFC) Procedure and the Partial Report (PR) Bar-Probe Procedure Effect Target type Target position Task Target Type ⫻ Target Position Target Type ⫻ Task Target Position ⫻ Task Target Type ⫻ Target Position ⫻ Task 2AFC task: Target Type ⫻ Target Position PR task: Target Type ⫻ Target Position Location errors (PR task) Target type Target position Target Type ⫻ Target Position Intrusion errors (PR task) Target type Target position Target Type ⫻ Target Position ⴱ p ⬍ .05. ⴱⴱ p ⬍ .01. 2 F1 ⴱⴱⴱ p ⬍ .001. 2 F2 57.12ⴱⴱⴱ 12.85ⴱⴱⴱ 53.93ⴱⴱⴱ 20.82ⴱⴱⴱ 22.87ⴱⴱⴱ 9.85 ⴱⴱⴱ 3.48ⴱ 6.92ⴱⴱⴱ 16.72ⴱⴱⴱ 0.06 0.09 0.17 0.05 0.02 0.02 0.01 0.06 0.08 F2(1, F2(4, F2(1, F2(4, F2(1, F2(4, F2(4, F2(4, F2(4, F1(1, 20) ⫽ 12.59ⴱⴱ F1(4, 80) ⫽ 15.68ⴱⴱⴱ F1(4, 80) ⫽ 7.72ⴱⴱⴱ 0.04 0.23 0.06 F2(1, 16) ⫽ 2.06 F2(4, 64) ⫽ 14.29ⴱⴱⴱ F2(4, 64) ⫽ 3.84ⴱⴱ 0.05 0.25 0.07 F1(1, 20) ⫽ 17.82ⴱⴱⴱ F1(4, 80) ⫽ 1.50 F1(4, 80) ⫽ 4.60ⴱⴱ 0.10 0.01 0.04 F2(1, 16) ⫽ 10.03ⴱⴱ F2 ⬍ 1 F2(4, 64) ⫽ 2.52ⴱ 0.19 0.02 0.07 F1(1, F1(4, F1(1, F1(4, F1(1, F1(4, F1(4, F1(4, F1(4, 20) 80) 20) 80) 20) 80) 80) 80) 80) ⫽ ⫽ ⫽ ⫽ ⫽ ⫽ ⫽ ⫽ ⫽ 16) 64) 16) 64) 16) 64) 64) 64) 64) ⫽ ⫽ ⫽ ⫽ ⫽ ⫽ ⫽ ⫽ ⫽ 7.12ⴱ 16.03ⴱⴱⴱ 105.52ⴱⴱⴱ 8.97ⴱⴱⴱ 9.67ⴱⴱ 8.93ⴱⴱⴱ 2.83ⴱ 4.40ⴱⴱ 8.42ⴱⴱ 0.08 0.13 0.24 0.07 0.02 0.03 0.01 0.09 0.12 SERIAL POSITION EFFECTS 493 Figure 10. Serial position functions for letters and symbols in the bar-probe partial report task of Experiment 6 after categorizing the errors into location errors (left) and intrusion errors (right). Vertical bars represent standard errors. letters, digits, and symbols are processed and are not the consequence of how a particular laboratory task is performed. Experiment 1 found distinct serial position functions for symbol stimuli compared with both letters and digits in a 2AFC identification task with stimuli similar to those tested by Mason (1982). Experiments 2– 4 were designed to test several different interpretations of the different serial positions functions found in the 2AFC task of Experiment 1 and, in particular, examined the fact that most of this difference occurred at the first position. The same pattern was found when randomly mixing stimulus types (Experiment 2), thus suggesting that the different serial position functions were not caused by an endogenous attentional bias generated when expecting a particular type of stimulus (directing attention to the beginning of the string, e.g.). The same pattern was also found in Experiment 3 when target letters were embedded in strings of symbols and vice versa, thus suggesting that the different serial position functions were not caused by an exogenous attentional bias automatically triggered on presentation of a string of letters or digits (again orienting attention to the beginning of the string). The results of Experiment 3 also suggest that the different serial position functions were not caused by a chunking mechanism that attempts to create combinations of letters or digits. Any attempt to form higher order letter combinations should have been severely disrupted by having symbol stimuli as the surrounding context. Contrary to the results obtained with the target search task, the results obtained with the 2AFC task in Experiments 1–3 showed that the difference between symbols and letters/digits mostly concerned performance for targets presented at the initial position of strings. Experiments 5 and 6 were designed to examine why we did not find a final-position advantage for letters and digits in the 2AFC task. Indeed, prior research examining serial position functions for letter stimuli with a bar-probe identification procedure (postcued partial report) systematically found a W-shaped serial position function (Haber & Standing, 1969; Merikle, Lowe, & Coltheart, 1971; Mewhort & Campbell, 1978; Mewhort et al., 1981, 1993). Experiment 5 found exactly this pattern for letter and digit stimuli and further demonstrated that superior performance for such stimuli compared with symbols arose at both the first and the final positions in the string. Experiment 6 replicated this result for letter and symbol stimuli and further demonstrated that the same pattern can be obtained with a 2AFC procedure when the alternative letters are displaced relative to the target string and a bar-probe is used to indicate the position of report. On the basis of the results of Experiment 6, we concluded that the absence of a final-position advantage for letters and digits in Experiments 1–3 was due to additional interference from the alternative letters displayed with the postmask. This conclusion is consistent with prior findings showing that the outer-position Figure 11. Comparison of the transformed PR data (left) and 2AFC data (right) of Experiment 6. PR data were transformed into 2AFC data by applying the equation described in footnote 4. PR ⫽ partial report; Expt. ⫽ Experiment; 2AFC ⫽ two-alternative forced choice. 494 TYDGAT AND GRAINGER advantage for letter stimuli in the bar-probe task is diminished in the presence of a backward mask that extends beyond the outer positions (Mewhort & Campbell, 1978) and that adding parentheses before and after the letter string greatly reduces the outerposition advantage, particularly for the final position (Haber & Standing, 1969). Finally, Experiments 5 and 6 provided evidence that the presence of location errors in the bar-probe technique (when another item in the display was reported instead of the target) exaggerates the outer-position advantage compared with the 2AFC procedure. Next, we argue that the pattern of results found in the present study can be understood by considering the constraints imposed on a specialized system for processing strings of letters or digits. The key mechanism behind this account is crowding (see Levi, 2008, for a recent review), and the central hypothesis is that crowding affects letter and digit stimuli differently than symbol stimuli. There are two main differences, which we discuss in detail. One involves a reduction in crowding effects by means of a change in the size of the receptive fields of letter and digit detectors, and the other involves a change in the shape of these receptive fields (see Figure 12). Crowding, Stimulus Type, and Serial Position We hypothesize that when children learn to read words and numbers, they develop a specialized system that is custom built to handle the very specific nature of these stimuli (see McCandliss, Cohen, & Dehaene, 2003, for one version of this hypothesis). Most notably, this system would develop to optimize processing in the extremely crowded conditions that arise with printed words and numbers, compared with other visual objects that do not typically occur in such a cluttered environment. One good candidate for the specialized mechanism for processing strings of letters and digits is Grainger and van Heuven’s (2003) alphabetic array (see Figure 12). This is a bank of alphabetic character detectors that performs parallel independent character recognition. In this model, alphabetic character detectors simultaneously process all characters in a string of limited length. Each character detector signals the pres- Figure 12. Illustration of part of Grainger and van Heuven’s (2003) model focusing on the receptive field structure of retinotopic letter detectors in the alphabetic array. The size and shape of the receptive fields is hypothesized to change as a function of eccentricity and visual field, with receptive field size increasing with eccentricity and becoming asymmetrical toward the left in the left visual field. The example shows processing of a random string of consonants with fixation on the central letter. ence of a given character at a particular location along the horizontal meridian (i.e., a retinotopic map). These location-specific detectors then feed activation onto location-invariant orthographic representations. It should be noted that the alphabetic array in Grainger and van Heuven’s model is part of a long tradition of independent parallel-channels models of letter string processing (Estes, 1972; Gardner, 1973; McClelland & Rumelhart, 1981; Rumelhart, 1970; Shiffrin & Geisler, 1973; Wolford, 1975). Furthermore, the notion of overlapping receptive fields for letter detectors is also a central component in Dehaene, Cohen, Sigman, and Vinckier’s (2005) account of visual word recognition, and similar ideas can be found in the early models of Estes (1972), Rumelhart (1970), and Wolford (1975). An obvious extension of this account, in light of the present data, is that numbers are processed with the same mechanism as letter strings (in which case the term alphanumeric array would be more appropriate). The central hypothesis here is that receptive field size of retinotopic letter and digit detectors has adapted to the need to optimize processing of strings of letters and digits. The smaller the receptive field size of these detectors, the less interference from neighboring characters. This is the basic idea behind Bouma’s (1970) functional field and Pelli et al.’s (2004) integration field, as measured with a standard crowding manipulation.5 Thus, the hypothesized smaller receptive field sizes for letters and digits (Figure 12) suggest that it is a reduction in the spatial extent of crowding for letters and digits that is the major factor driving the different serial position functions for these stimuli compared with symbols. The larger receptive field size for symbol detectors would generate close to maximum crowding with only a single flanking symbol. In this way, the first and last symbols in an array of symbols are subject to practically the same amount of crowding as the inner letters. Letters and digits, on the other hand, would be subject to lower levels of crowding that would depend on the number of flanking characters (one or two) because of the hypothesized narrower receptive fields of the detectors in the alphabetic array. This, therefore, explains the classic W-shaped serial position function for letters and digits. The different spatial extent of crowding on letter, digit, and symbol stimuli also accounts for why the 2AFC task (Experiments 1–3) did not show the standard final-position advantage for letters and explains why this advantage was reinstated when the alternative letters were displaced relative to the target position (Experiment 6). Inner letters in a string are assumed to suffer close to maximal crowding, whereas outer letters do not. Therefore, the masking effects of the alternative letters in 2AFC (when positioned next to the target) mostly affect performance at the end positions. Furthermore, because symbols are hypothesized to suffer maximal 5 Some particularly relevant data came to our attention during the preparation of a revision of this work. Kwon and Legge (2006) have shown that crowding effects are stronger in children than in skilled adult readers. This supports the notion that reduced crowding is a key ingredient in the optimization of printed word perception. Further support for this hypothesis was provided by Kwon, Legge, and Dubbels (2007), who demonstrated that development of reading speed is linked to development of the size of the visual span, which has in turn been shown to be determined by crowding (Pelli et al., 2007). Finally, recent research has shown that the spatial extent of crowding can be reduced by training observers to identify letters closely flanked by other letters (Chung, 2007). SERIAL POSITION EFFECTS crowding from a single flanking stimulus, the presence of the two alternative letters has little additional influence on the processing of these stimuli. This account therefore explains the observed variation in the final-position advantage for letter targets as a function of testing procedure but does not account for the robust advantage for the initial position that occurs over and above such procedural differences. We return to discuss this initial-position advantage later. Surprisingly, little research has used the standard flanker paradigm with eccentric target locations to directly investigate the effect of number of flanking stimuli (e.g., Bouma, 1970). We therefore set about running these experiments ourselves with the same set of letters and symbols as tested in the present study (Grainger & Tydgat, 2008). Targets were presented at eccentricities equal to the first and last positions of strings in the present study, either in isolation or accompanied by one or two flanking characters. The results revealed that performance for letter targets with a single flanker was closer to the no-flanker condition than to the two-flanker condition, whereas symbol targets showed practically as much interference from a single flanker as from two flankers and showed overall greater interference from flankers than did letter targets. This pattern nicely confirms the predictions derived from our crowding interpretation of the results of the present study. Symbols are more prone to crowding effects than are letters, and although symbols show close to maximal interference in the presence of a single flanker, letters, on the other hand, show very little interference from a single flanker. The Initial-Position Advantage: Sequential Processing or Specialized Detectors? In this section, we discuss some other mechanisms that might be at least partly responsible for the present data pattern. The initialposition advantage found in the present study might be the reflection of a serial processing mechanism specific to stimuli that typically appear in strings. According to such an account, processing moves continuously from the beginning of the string to the end; therefore, items at the beginning of the string are processed before items at the end of the string (e.g., the scanning model of Mewhort & Popham, 1991). Whitney’s (2001) SERIOL (sequential encoding regulated by inputs to oscillations within letter units) model provides a recent example of such an approach. In this model, a beginning-to-end activation gradient forms the starting point for orthographic processing (coding letter identities and letter positions). According to Whitney, the visual acuity function present in early visual areas (V1/V2) is transformed into a monotonically decreasing activation gradient (called the locational gradient) at the level of feature representations in V4. This locational gradient is then imposed on a set of position-independent letter representations, such that the first letter in the string has the highest activation, and activation diminishes across the string until the final letter. The activation levels of letter representations determine their order of firing, which then determines the formation of ordered letter sequences (contiguous and noncontiguous bigrams) at the bigram level of representation in the model. Whitney’s (2001) account therefore predicts a strong linear component in the identification of strings of letters, digits, and symbols, because the activation gradient is already present at the level of feature representations. The present results would appear 495 to rule out this possibility. Our results therefore suggest that, if there is indeed such a sequential processing bias, then it is most likely initiated at the level of representations in the alphabetic array. This could be achieved by a spatial attention mechanism that would impose a beginning-to-end activation gradient on letter representations. Grainger et al. (2006) suggested that such a mechanism would be particularly important for the correct assignment of grapheme–phoneme correspondences as in Perry, Ziegler, and Zorzi’s (2007) model of reading aloud. This would fit into a generic dual-route framework for visual word recognition (Grainger, Diependaele, Spinelli, Ferrand, & Farioli, 2003; Grainger & Ziegler, 2007) in which the alphabetic array feeds information into two different processing routes: a direct orthographic route for semantic access and an indirect route that implements sublexical spelling-to-sound conversion (e.g., through graphemes and phonemes). However, several aspects of the present results appear to rule out a beginning-to-end processing bias, whatever the precise nature of the bias. First, the results of Experiments 2 and 3 appear to rule out endogenous and exogenous attentional biases specific to strings of letters and digits (as discussed earlier). Second, the results of Experiments 5 and 6 reinstated a final-position advantage for letters and digits. This led to a diminution in the linear component and to an increase in the quartic component in the trends analyses. Third, performance for letter and digit stimuli at Positions 2 and 4 was always very similar across all experiments, suggesting that the linear trends were being driven almost entirely by performance at the first position. Overall, the results of the present experiments are more in favor of an advantage specifically for the first position of strings of letters or digits, rather than the combination of a beginning-to-end processing bias with an outer-position advantage (because of reduced crowding). Within an integrated account of the complete set of results reported here, we hypothesize that not only do the receptive fields of letter and digit detectors undergo a change in size because of experience with reading words and numbers, but they are also subject to a change in shape. This hypothesized change in shape of receptive fields (see Figure 12) would arise to optimize processing at the first position in strings of letters and digits. Letters in the first position tend to provide more constraint on lexical identity than letters in any other position in the string (e.g., Clark & O’Regan, 1999; Grainger & Jacobs, 1993). Furthermore, the first position is critical for translating an orthographic code into a phonological code, because the correct graphemes (letters and letter clusters corresponding to a phoneme) can be computed only with precise order information (Perry et al., 2007). Such optimization can be achieved by asymmetric receptive fields that are elongated in the direction of the initial position (i.e., to the left for languages read from left to right). Letter and digit detectors with receptive fields of this shape mostly suffer interference from characters immediately to their left, hence endowing an advantage for the initial position in strings. We further hypothesize that such optimization operates only for stimuli that are fixated and therefore only for detectors receiving input from the LVF. This visual-field specificity of the shape of receptive fields of letter and digit detectors accounts for why the initial-position advantage (LVF) is systematically larger than the final-position advantage (RVF). If one further assumes no change in the size (surface area) of receptive fields as a function of visual field, then 496 TYDGAT AND GRAINGER the elongated receptive fields in the LVF would have smaller vertical extent (because of the larger horizontal extent) than the more circular receptive fields of the RVF. This would explain why letters and digits at the initial position are less sensitive to the presence of the two alternative letters in 2AFC than are letters and digits at the final position. Bouma (1973) was among the first to report visual field differences in letter identification in unpronounceable nonwords (note that only the first and last positions were tested in Bouma’s study). For a fixed eccentricity, there was a strong advantage for initial over final position in the LVF, which diminished and even reversed (final position superior to initial position) in the RVF.6 This led Bouma to suggest that “the spatial extent of foveally oriented masking is smaller in the R field than the Left” (p. 775). This is in fact an alternative version of the present hypothesis. Because there is evidence for an inward– outward asymmetry in crowding effects from single flankers (Banks, Bachrach, & Larson, 1977; Banks, Larson, & Prinzmetal, 1979; Bouma, 1970; Chastain, 1982; Krumhansl & Thomas, 1977; White, 1976), this would suggest that receptive fields are elongated to the left in the LVF and to the right in the RVF (Bouma, 1978). Thus, our hypothesized change in shape of receptive fields for letters and digits could involve a general shift to the left operating over this initial asymmetry. This would generate an exaggerated elongation to the left in the LVF and a more symmetrical receptive field shape in the RVF. Our own recent crowding experiments (Grainger & Tydgat, 2008) have provided independent support for this hypothesized visual-field specificity of receptive fields. Letter targets presented in the LVF or RVF (at the same eccentricity as the outer letters in the present experiments), and flanked by a single letter either immediately to the right or to the left of the target, showed an asymmetric pattern of interference. Flanker position affected performance in the LVF but not in the RVF. That is, targets presented in the LVF flanked by another letter to the left were harder to identify than targets at the same location flanked by a letter to the right, whereas targets in the RVF showed equal amounts of interference from left and right flankers. This is exactly the pattern predicted by our account of the present set of results. Conclusions The present study examined the identification accuracy of letter, digit, and symbol targets presented in strings. In line with prior research with speeded target detection, the obtained serial position functions were similar for letters and digits and were different for symbols. In Experiments 1–3 we used a 2AFC procedure, and most of this difference occurred for performance at the first position in strings, with much higher levels of performance for letters and digits compared with symbols at this particular location. We showed that this first-position advantage for letters and digits compared with symbols was still present when the different types of target were randomly intermixed in the experiment, as opposed to being blocked by stimulus type. Furthermore, the initial-position advantage for letters compared with symbols was present even when letter targets were embedded in symbol strings and vice versa. Experiments 5 and 6 showed that a final-position advantage (and hence the classic W-shaped serial position function) was obtained for letters and digits when the alternative letters presented for 2AFC were replaced by a bar-probe stimulus. Our results suggest that the constraints imposed by reading words and numbers force an adaptive modification of basic visual processes to optimize identification of alphanumeric stimuli. This adaptive modification is hypothesized to involve a change in size and a change in shape of the receptive fields of retinotopic letter and digit detectors. 6 Other work has examined visual field effects in letter identification in four-letter strings (e.g., Jordan et al., 2003). In Jordan et al.’s (2003) study, eccentricity was matched for the whole string, such that initial and final letters had different eccentricities. In these conditions, initial and final letters were identified with equal accuracy in both visual fields. Under the modified receptive field hypothesis, this symmetrical effect is likely due to the trade-off between crowding and eccentricity. The initial position in the LVF suffers less crowding but is more eccentric than the initial position in the RVF, and the final position in the RVF suffers less crowding but is more eccentric than the final position in the LVF. References Averbach, E., & Coriell, A. S. (1961). Short-term memory in vision. Bell Telephone Technical Journal, 40, 19 –31. Banks, W. P., Bachrach, K., & Larson, D. W. (1977). Asymmetry of lateral interference in visual letter identification. Perception & Psychophysics, 22, 232–240. Banks, W. P., Larson, D. W., & Prinzmetal, W. (1979). Asymmetry of visual interference. Perception & Psychophysics, 25, 447– 456. Bouma, H. (1970). Interaction effects in parafoveal letter recognition. Nature, 226, 177–178. Bouma, H. (1973). Visual interference in the parafoveal recognition of initial and final letters of words. Visual Research, 13, 767–782. Bouma, H. (1978). Visual search and reading: Eye movements and functional visual field: A tutorial review. In J. Requin (Ed.), Attention and performance VII (pp. 115–147). Hillsdale, NJ: Erlbaum. Breitmeyer, B. G., & Ogmen, H. (2000). Recent models and findings in visual backward masking: A comparison, review, and update. Perception & Psychophysics, 62, 1572–1595. Butler, B. E. (1975). Selective attention and target search with brief visual displays. Quarterly Journal of Experimental Psychology, 27, 467– 477. Butler, B. E., & Merikle, P. M. (1973). Selective masking and processing strategy. Quarterly Journal of Experimental Psychology, 25, 542–548. Campbell, A. J., & Mewhort, D. J. (1980). On familiarity effects in visual information processing. Canadian Journal of Psychology, 34, 134 –154. Chastain, G. (1982). Confusability and interference between members of parafoveal letter pairs. Perception & Psychophysics, 32, 576 –580. Chung, S. T. L. (2007). Learning to identify crowded letters: Does it improve reading speed? Vision Research, 47, 3150 –3159. Clark, J. J., & O’Regan, J. K. (1999). Word ambiguity and the optimal viewing position in reading. Vision Research, 39, 843– 857. Dehaene, S., Cohen, L., Sigman, M., & Vinckier, F. (2005). The neural code for written words: A proposal. Trends in Cognitive Sciences, 9, 335–341. Estes, W. K. (1972). Interactions of signal and background variables in visual processing. Perception & Psychophysics, 12, 278 –286. Estes, W. K., Allmeyer, D. H., & Reder, S. M. (1976). Serial position functions for letter identification at brief and extended exposure durations. Perception & Psychophysics, 19, 1–15. Gardner, G. T. (1973). Evidence for independent parallel channels in tachistoscopic perception. Cognitive Psychology, 4, 130 –155. Grainger, J. (2008). Cracking the orthographic code: An introduction. Language and Cognitive Processes, 23, 1–35. Grainger, J., Diependaele, K., Spinelli, E., Ferrand, L., & Farioli, F. (2003). SERIAL POSITION EFFECTS Masked repetition and phonological priming within and across modalities. Journal of Experimental Psychology: Learning, Memory, and Cognition, 29, 1256 –1269. Grainger, J., Granier, J. P., Farioli, F., Van Assche, E., & van Heuven, W. J. (2006). Letter position information and printed word perception: The relative-position priming constraint. Journal of Experimental Psychology: Human Perception and Performance, 32, 865– 884. Grainger, J., & Jacobs, A. M. (1993). Masked partial-word priming in visual word recognition: Effects of positional letter frequency. Journal of Experimental Psychology: Human Perception and Performance, 19, 951–964. Grainger, J., & Jacobs, A. M. (1996). Orthographic processing in visual word recognition: A multiple read-out model. Psychological Review, 103, 518 –565. Grainger, J., & Tydgat, I. (2008). Crowding affects letters and symbols differently. Manuscript in preparation. Grainger, J., & van Heuven, W. (2003). Modeling letter position coding in printed word perception. In P. Bonin (Ed.), The mental lexicon (pp. 1–24). New York: Nova Science. Grainger, J., & Ziegler, J. (2007). Cross-code consistency effects in visual word recognition. In E. L. Grigorenko & A. Naples (Eds.), Single-word reading: Biological and behavioral perspectives (pp. 129 –157). Mahwah, NJ: Erlbaum. Greenhouse, S., & Geisser, S. (1959). On methods in the analysis of profile data. Psychometrika, 24, 95–112. Haber, R. N., & Standing, L. (1969). Location of errors with a poststimulus indicator. Psychonomic Science, 17, 345–346. Hammond, E. J., & Green, D. W. (1982). Detecting targets in letter and non-letter arrays. Canadian Journal of Psychology, 36, 67– 82. Huckauf, A., & Heller, D. (2004). On the relations between crowding and visual masking. Perception & Psychophysics, 66, 584 –595. Jordan, T. R., & Bevan, K. M. (1996). Position-specific masking and word–letter phenomenon: Reexamining the evidence from the Reicher– Wheeler paradigm. Journal of Experimental Psychology: Human Perception and Performance, 22, 1416 –1433. Jordan, T. R., Patching, G. R., & Milner, A. D. (2000). Lateralized word recognition: Assessing the role of hemispheric specialization, modes of lexical access, and perceptual asymmetry. Journal of Experimental Psychology: Human Perception and Performance, 26, 1192–1208. Jordan, T. R., Patching, G. R., & Thomas, S. M. (2003). Assessing the role of hemispheric specialization, serial-position processing, and retinal eccentricity in lateralised word recognition. Cognitive Neuropsychology, 20, 49 –71. Krumhansl, C. L., & Thomas, E. A. C. (1976). Extracting identity and location information from briefly presented letter arrays. Perception & Psychophysics, 20, 243–258. Krumhansl, C. L., & Thomas, E. A. C. (1977). Effect of level of confusability on reporting letters from briefly presented visual displays. Perception & Psychophysics, 21, 269 –279. Ktori, M., & Pitchford, N. J. (2008). Effect of orthographic transparency on letter position encoding: A comparison of Greek and English monoscriptal and biscriptal readers. Language and Cognitive Processes, 23, 258 –281. Kwon, M. Y., & Legge, G. E. (2006). Developmental changes in the size of the visual span for reading: Effects of crowding [Abstract]. Journal of Vision, 6, 1003a. Kwon, M. Y., Legge, G. E., & Dubbels, B. R. (2007). Developmental changes in the visual span for reading. Vision Research, 47, 2889 –2890. Levi, D. M. (2008). Crowding—An essential bottleneck for object recognition: A mini-review. Vision Research, 48, 635– 654. Mason, M. (1975). Reading ability and letter search time: Effects of orthographic structure defined by single-letter positional frequency. Journal of Experimental Psychology, 104, 146 –166. Mason, M. (1982). Recognition time for letters and nonletters: Effects of 497 serial position, array size, and processing order. Journal of Experimental Psychology: Human Perception and Performance, 8, 724 –738. Mason, M., & Katz, L. (1976). Visual processing of nonlinguistic strings: Redundancy effects and reading ability. Journal of Experimental Psychology, 105, 338 –348. McCandliss, B. D., Cohen, L., & Dehaene, S. (2003). The visual word form area: Expertise for reading in the fusiform gyrus. Trends in Cognitive Sciences, 13, 155–161. McClelland, J. L., & Rumelhart, D. E. (1981). An interactive activation model of context effects in letter perception: Part 1. An account of basic findings. Psychological Review, 88, 375– 407. Merikle, P. M. (1974). Selective backward masking with an unpredictable mask. Journal of Experimental Psychology, 103, 589 –591. Merikle, P. M., & Coltheart, M. (1972). Selective forward masking. Canadian Journal of Psychology, 26, 296 –302. Merikle, P. M., Coltheart, M., & Lowe, D. G. (1971). On the selective effects of a patterned masking stimulus. Canadian Journal of Psychology, 25, 264 –279. Merikle, P. M., Lowe, D. G., & Coltheart, M. (1971). Familiarity and method of report as determinants of tachistoscopic performance. Canadian Journal of Psychology, 25, 167–174. Mewhort, D. J. K., & Campbell, A. J. (1978). Processing spatial information and the selective-masking effect. Perception & Psychophysics, 24, 93–101. Mewhort, D. J. K., Campbell, A. J., Marchetti, F. M., & Campbell, J. I. D. (1981). Identification, localization, and “iconic memory”: An evaluation of the bar-probe task. Memory & Cognition, 9, 50 – 67. Mewhort, D. J. K., Huntley, M. F., & Duff-Fraser, H. (1993). Masking disrupts recovery of location information. Perception & Psychophysics, 54, 759 –762. Mewhort, D. J. K., & Popham, D. (1991). Serial recall of tachistoscopic letter strings. In W. E. Hockley & S. Lewandowsky (Eds.), Relating theory and data: Essays on human memory in Honor of Bennet B. Murdock (pp. 425– 443). Hillsdale, NJ: Erlbaum. Nazir, T. A., Ben-Boutayab, N., Decoppet, N., Deutsch, A., & Frost, R. (2004). Reading habits, perceptual learning, and recognition of printed words. Brain and Language, 88, 294 –311. Pelli, D. G., Palomares, M., & Majaj, N. J. (2004). Crowding is unlike ordinary masking: Distinguishing feature integration from detection. Journal of Vision, 4, 1136 –1169. Pelli, D. G., Tillman, K. A., Freeman, J., Su, M., Berger, T. D., & Majaj, N. J. (2007). Crowding and eccentricity determine reading rate. Journal of Vision, 7, 1–36. Perry, C., Ziegler, J. C., & Zorzi, M. (2007). Nested incremental modeling in the development of computational theories: The CDP⫹ model of reading aloud. Psychological Review, 114, 273–315. Pitchford, N. J., Ledgeway, T., & Masterson, J. (2008). Effect of orthographic processes on letter position encoding. Journal of Research in Reading, 31, 97–116. Reicher, G. M. (1969). Perceptual recognition as a function of meaningfulness of stimulus material. Journal of Experimental Psychology, 28, 115–121. Rumelhart, D. E. (1970). A multicomponent theory of the perception of briefly exposed visual displays. Journal of Mathematical Psychology, 7, 191–218. Rumelhart, D. E., & McClelland, J. L. (1982). An interactive activation model of context effects in letter perception: Part 2. The contextual enhancement effect and some tests and extensions of the model. Psychological Review, 89, 60 –94. Schwantes, F. M. (1978). Stimulus position functions in tachistoscopic identification tasks: Scanning, rehearsal, and order of report. Perception & Psychophysics, 23, 219 –226. Shiffrin, R. M., & Geisler, W. (1973). Visual recognition in a theory of information processing. In R. Solso (Ed.), Contemporary issues in 498 TYDGAT AND GRAINGER cognitive psychology: The Loyola Symposium (pp. 53–101). Washington, DC: Winston. Stevens, M., & Grainger, J. (2003). Letter visibility and the viewing position effect in visual word recognition. Perception & Psychophysics, 65, 133–151. Tramer, O., Butler, B. E., & Mewhort, D. J. (1985). Evidence for scanning with unilateral visual presentation of letters. Brain and Language, 25, 1–18. Van der Heijden, A. H. C. (1992). Selective attention in vision. London: Routledge. Wheeler, D. D. (1970). Processes in word recognition. Cognitive Psychology, 1, 59 – 85. White, M. J. (1976). Order of processing in visual perception. Canadian Journal of Psychology, 30, 140 –156. Whitney, C. (2001). How the brain codes the order of letters in a printed word: The SERIOL model and selective literature review. Psychonomic Bulletin and Review, 8, 221–243. Wolford, G. (1975). Perturbation model for letter identification. Psychological Review, 82, 184 –199. Wolford, G., & Hollingsworth, S. (1974). Retinal location and string position as important variables in visual information processing. Perception & Psychophysics, 16, 437– 442. Received February 15, 2007 Revision received April 9, 2008 Accepted April 28, 2008 䡲