Behavioural and population responses ... availability of Artemia prey by ...
advertisement

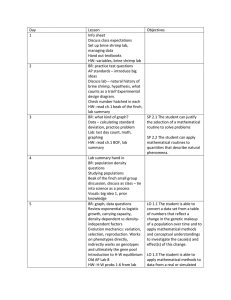
Hydrobiologia (2011) 664:163–171 DOI 10.1007/s10750-010-0596-x PRIMARY RESEARCH PAPER Behavioural and population responses to changing availability of Artemia prey by moulting black-necked grebes, Podiceps nigricollis Nico Varo • Andy J. Green • Marta I. Sánchez Cristina Ramo • Jesú s Gómez • Juan A. Amat • Received: 17 May 2010 / Revised: 9 December 2010 / Accepted: 30 December 2010 / Published online: 19 January 2011 Abstract We examined how availability of brine shrimps, Artemia parthenogenetica, influenced temporal aspects of foraging behaviour and population dynamics of moulting black-necked grebes, Podiceps nigricollis, from late August to early December in four salt ponds in the Odiel marshes, southern Spain, in 2008 and 2009. The moulting grebe population was higher in 2009, coinciding with an increase in shrimp biomass, with a peak of 2,500 birds in October. Grebes increased their time spent foraging as the season progressed, coinciding with decreases in shrimp biomass and water temperature. Foraging activity was lower in 2009, when shrimp biomass was greater. Diving was the most frequent feeding method, especially as the season progressed. Brine shrimps at the bottom of the water column were larger than those near the surface. Differences between years in grebe body mass suggest that changing shrimp availability and water temperature had an influence on body condition. The grebe population consumed an estimated 0.2–2.0% of the standing crop of Artemia per day, with this fraction Handing editor: Stuart Anthony Halse N. Varo (&) A. J. Green M. I. Sánchez C. Ramo J. Gómez J. A. Amat Department of Wetland Ecology, Estación Biológica de Doñana, C. S. I. C, Calle Américo Vespucio s/n, 41092 Sevilla, Spain e-mail: nico@ebd.csic.es increasing as the season progresses, thus contributing to the decline in the Artemia population. Our results suggest that moulting grebes are probably only able to adjust foraging effort within certain limits, especially at the end of moulting period when thermal stress is greatest and food supply is lowest. They may leave the study area when they can no longer meet their energy requirements. Keywords Brine shrimps Foraging behaviour Grebes Mediterranean wetlands Waterbirds Introduction Foraging behaviour is an important part of the daily routine in animals, forming an essential link between prey availability and ecological success (Martin, 1987; Varo & Amat, 2008). Before arriving at wintering areas, many bird species perform a moult migration from their breeding sites to moulting areas (Salomonsen, 1968; Jehl, 1990a). Such is the case of the black-necked grebe, Podiceps nigricollis, and flocks of 600,000–900,000 moulting grebes have been recorded in two saline lakes in North America (Jehl, 1988, 1990a). In Europe, smaller flocks of about 10,000 moulting black-necked grebes have been recorded in several moulting areas, the Odiel marshes in southern Spain being one of the most important moulting sites (Van Impe, 1969; Garcı́aJiménez & Calvo-Sendı́n, 1987; Iborra et al., 1991). 123 164 This moulting period is critical for grebes not only because they simultaneously lose all flight feathers (Jehl, 1988; Piersma, 1988) and are more vulnerable to predators, but also because they need to accumulate fat before moving to wintering areas (Winkler & Cooper, 1986; Jehl, 1988, 1990b). Therefore, grebes can be expected to make various foraging decisions, including prey and habitat selection and time budget allocation, in order to obtain the highest net energy gains. The Odiel marshes have low invertebrate species richness (Sánchez et al., 2006a), and the brine shrimp Artemia parthenogenetica is likely to be the most important food item for grebes, given its dominance amongst invertebrate biomass (Sánchez et al., 2006a). Brine shrimps A. franciscana are the main prey of this grebe species in hypersaline moulting areas in North America (Cooper et al., 1984; Caudell & Conover, 2006a). At the Odiel marshes, shrimps show a seasonal fluctuation in abundance across the moulting period (Sánchez et al., 2006a), so that foraging responses of grebes to changing shrimp abundance should be expected in order to meet energy demands. For example, increasing energy consumption by increasing foraging effort is an obvious way to deal with decline in food abundance (Guillemain et al., 2000a; Guillemain & Fritz, 2002; Varo & Amat, 2008). This study focused on the role of food availability in controlling and regulating the moulting black-necked grebe population at Odiel marshes over a 2 year period. We analysed the foraging behaviour of grebes, and how the shrimp abundance influenced temporal aspects of foraging behaviour and the population dynamics of grebes across the moulting period. Methods The Odiel marshes (37°140 N; 6°570 W) is an estuarine complex of 7,185 ha formed at the mouths of the rivers Odiel and Tinto and containing 1,174 ha of salt pans. In the main, intensively managed block of salt pans (1,118 ha), sea water is pumped from primary to secondary evaporation areas and finally to crystallizers. The salinity gradually increases until the final crystallization ponds are reached (see Sánchez et al., 2006a for more details). In this study, our field work was conducted from August to December 2008 and 123 Hydrobiologia (2011) 664:163–171 2009 in four ponds (76 h and 80 cm mean deep) within the secondary evaporation zone, where many grebes moult each year (L. Garcı́a, unpubl. data) and brine shrimp is the most abundant invertebrate (Sánchez et al., 2006a). Grebes are absent from crystallization ponds and less abundant in the primary than secondary evaporation ponds, and are much rarer outside our study period. For our analyses, we pooled data from all four ponds because they were adjacent, with hydrological connections and similar salinity, with regular interchange of both grebes and shrimps. During this study, we counted the number of grebes each week by telescope. Grebes displayed considerable avoidance and vigilance behaviours when they observed us, facilitating counts. Only when we were very close did they dive to avoid us. We also recorded behavioural observations in a tape recorder, i.e. foraging (diving or pecking at the water surface), swimming, preening, flapping or resting. Blacknecked grebes are not thought to feed at night (Cooper et al., 1984), and our behavioural observations were conducted between 09:00 and 12:00 h twice a month on days without wind. Each pond was scanned once on each date and at the same hour. We considered the percentage of all recorded grebes that were feeding as a measure of foraging effort. The moulting status (old plumage, moulting or new plumage) of the wing feathers of each grebe captured during weekly ringing operations by Estació n Bioló gica de Doñ ana in our study area was recorded, as well as body mass to the nearest gram on an electronic balance. Furthermore, we examined the gizzard contents of six grebes that died accidentally during ringing operations on different days, in order to obtain diet information. The oesophagi were empty. Gizzard contents were stored in 70% alcohol then later identified to the lowest possible taxonomic level using a binocular microscope. The percentage of total food content volume that was made up of each component was estimated by displacement (see Sánchez et al., 2005 for details). We estimated the brine shrimp abundance (BSA) by establishing 16 fixed study points in the study area. At each point, we regularly quantified shrimp biomass by filtering 100 l of the water column using a sieve with a 0.5 mm mesh. The shrimps collected were dried at 50° during 24 h, and then weighed to the nearest 0.1 mg with an electronic balance. Brine shrimp abundance was considered here as dry mass of Hydrobiologia (2011) 664:163–171 Results The moulting period of black-necked grebes extended from mid summer until early winter (Fig. 1). The moulting grebe population was larger in 2009 than moulting old plumage new plumage 2008 100 grebes (%) 80 60 40 20 0 2009 100 80 grebes (%) shrimp per 100 l. The size distribution of shrimps was also estimated in separate samples from the same study points. We sieved shrimps from 10 l of water from the bottom, and 10 l of water from the top of the water column. These shrimps were photographed the same day in the lab with a digital camera, and then measured on a computer using Image J software. The total length (including tail) was measured for each individual. We aimed to sample all 16 points weekly, but stopped sampling in the presence of strong wind which has a strong effect on brine shrimp distribution (Sánchez et al., 2006a). The number of points sampled therefore ranged from 0 to 16 per week. Since our study area of 76 ha had a mean depth of 80 cm, we estimated the total water volume as 680,000 m3. Combined with data on shrimp abundance, this allowed us to estimate the standing crop of Artemia (CA). Thus, CAi = (680,000 m3 * BSAi)/ 0.1 m3, where CAi is the estimated crop of Artemia on day i and BSAi is the mean brine shrimp abundance measured on day i. In addition, based on the literature (Caudell & Conover, 2006a), we estimated the daily energetic requirements of the grebe population as 525.92 kJ per bird per day, and the energy content of dry Artemia as 21.88 kJ per g of dry mass. Thus, we could estimate how much Artemia the grebes ate to meet their requirements. Throughout the study period, water temperature was measured every hour using StowAway Tidbit underwater data loggers placed on the bottom. We used two-way ANOVAs to analyse the effects of week, year and their interaction on brine shrimp biomass (square root transformed). Student’s t tests were used to compare the size of shrimps at the top and bottom of the water column and annual variation in grebe body mass. A Wilcoxon Matched-Paired test was used to compare the foraging effort of grebes between years. The relationships between foraging effort and brine shrimp biomass was analysed using Spearman correlations. All statistical tests were performed using Statistica 6.0 (Statsoft Inc., Tulsa, OK, USA). 165 60 40 20 0 Fig. 1 Percentage of black-necked grebes in different stages of moult from the third week of August (a3) until the first week of December (d1), in 2008 and 2009. Sample sizes are shown above the bars. Moulting status refers to wings 2008, with the highest number of grebes recorded in the last week of October in both years (Fig. 2). Gizzard contents (n = 6) suggested that brine shrimps were the main prey, as Artemia represented 60–90% of the volume of food items in each gizzard. The other food items recorded were Ochthebius (Coleoptera), Hydrobia (Gastropoda) and Corixidae (Hemiptera). Habitat conditions for grebes varied as the moulting season progressed and also between years. Water temperature gradually declined between September 123 166 Hydrobiologia (2011) 664:163–171 3000 28 2008 2009 26 24 2000 1500 T (°C) number of grebes 2500 2008 2009 22 20 1000 18 500 16 0 14 and November, but with major variation between years for a given date (Fig. 3). For example, November was much cooler in 2008. Overall, temperature ranged from a minimum of 8.5°C on November 27th and a maximum of 30.8°C on September 10th in 2008 and from a minimum of 9.8°C on November 30th to a maximum of 32.8°C on September 7th in 2009. We found major seasonal and annual variation in shrimp biomass, with a strong week 9 year interaction (F7, 153 = 5.49, P \ 0.001; Fig. 4). In particular, shrimp biomass was greater in 2008 than 2009 in the second week of September (s2; Fig. 4), while the opposite occurred in the other weeks. When s2 was removed from the analysis, the week 9 year interaction was no longer significant (F6, 133 = 5.49, P = 0.190, Fig. 4). Brine shrimps were consistently larger at the bottom than at the top of the water column throughout the study (Table 1). At the start of the moulting period, grebes had a higher body mass in 2008 (Table 2). In contrast, from mid November onwards, the body mass of grebes was greater in 2009 (Table 2). Grebes increased their foraging effort as time passed in both 2008 (rs = 0.904, P \ 0.001) and 2009 (rs = 0.833, P = 0.01; Table 3). Overall, foraging effort was greater in 2008 123 Fig. 3 Variations in water temperature (mean ± s.e.) in 2008 and 2009 at salt ponds in Odiel marshes. s = September; o = October; n = November; 1 = first week; 2 = second week; 3 = third week; 4 = fourth week 1.4 4 1.2 brine shrimp abundance Fig. 2 Number of black-necked grebes in the study area during the moulting period in 2008 and 2009. a = August; s = September; o = October; n = November; d = December; 1 = first week; 2 = second week; 3 = third week; 4 = fourth week 10 9 9 12 1.0 0.8 11 12 12 0.6 12 10 9 14 0.4 16 13 0.2 12 12 0.0 s2 s3 o1 o2 o3 o4 n1 n4 Fig. 4 Seasonal variations in brine shrimp abundance (dry mass per 100 l of filtered water, mean ± s.e.) in 2008 (solid line) and 2009 (dashed line). Each point denotes a different sampling day. Sample sizes (number of points sampled) are indicated beside points. s = September; o = October; n = November; 1 = first week; 2 = second week; 3 = third week; 4 = fourth week Hydrobiologia (2011) 664:163–171 167 Table 1 Size (mm, mean ± S.D.) of the brine shrimps recorded at the bottom and surface of the water column throughout the moulting seasons of grebes in 2008 and 2009 2008 n P Bottom Surface 6.17 ± 1.70 5.98 ± 1.57 4.76 ± 1.74 4.84 ± 1.33 3.85 4.04 4.61 3.27 s2 o1 865 841 o2 o3 o4 n1 1,973 990 651 1,454 5.08 5.33 5.49 3.67 n4 1,306 3.83 ± 1.37 ± ± ± ± 1.83 1.89 1.92 1.50 2009 P n Bottom Surface \0.001 \0.001 379 933 7.40 ± 1.17 6.01 ± 1.67 4.49 ± 2.32 4.55 ± 1.39 \0.001 \0.001 1.53 1.78 1.68 1.41 \0.001 \0.001 \0.001 \0.001 302 331 205 235 5.68 6.82 6.63 6.11 4.62 6.62 4.25 3.89 1.66 1.77 1.58 0.99 \0.001 0.269 \0.001 \0.001 3.23 ± 1.22 \0.001 159 6.41 ± 1.17 4.09 ± 1.26 \0.001 ± ± ± ± ± ± ± ± 1.88 1.39 1.83 1.68 ± ± ± ± Differences were analysed using Student t tests. a = August; s = September; o = October; n = November; d = December; 1 = first week; 2 = second week; 3 = third week; 4 = fourth week Table 2 Activities of grebes (% of individuals, n) recorded during the moulting seasons in 2008 and 2009 Foraging activities Date N 2008 a2 s1 s2 o1 Other activities Foraging effort (%) Diving (%) Pecking (%) div:pec Swimming (%) Preening (%) Flapping (%) Resting (%) 19.4 30.1 40 36.5 13.8 19.4 31.3 22.9 5.6 10.7 8.7 13.6 2:01 2:01 4:01 2:01 13.8 26.2 5.3 3.5 66.8 41.5 51.5 58.7 0 0 0.9 0.8 0 2.2 2.3 0.5 o2 1,452 51.2 n1 1,040 63.4 n2 857 74.2 d1 403 53.6 38.2 56.7 62.5 37.5 13.1 6.6 11.7 16.1 3:01 9:01 5:01 2:01 1.7 1.5 2.5 36.2 44.6 33.2 22.3 8.2 1.2 1.9 1.1 2 1.3 0 0 0 220 18.2 208 23.1 639 27.7 11.4 11.5 20.2 6.8 11.5 7.7 2:01 1:01 3:01 12.7 13 1.7 69.1 62.5 68.1 0 1.4 1.9 0 0 0.5 o1 861 32.3 o2 1,519 26.9 n1 640 62.4 21.6 15.9 47.2 10.7 10.9 15.2 2:01 1:01 3:01 1.6 0.4 0.3 63.5 71.1 36.6 2.6 1.7 0.8 0 0 0 n2 557 60 52.2 7.7 7:01 0.4 37.9 1.8 0 d1 421 59.2 51.3 7.8 7:01 0 37.9 2.9 0 2009 a2 s1 s2 232 366 470 882 a = August; s = September; o = October; n = November; d = December; 1 = first fortnight; 2 = second fortnight than 2009 (Wilcoxon Matched-Paired test; T = 4, P = 0.049; Table 3). Diving was always the dominant foraging activity, and tended to become relatively more important than pecking as time passed (Table 3). There was a significant negative correlation between the foraging effort of grebes and brine shrimp biomass in 2008 (rs = -0.785 P = 0.036), but not in 2009 (rs = 0.070, P = 0.879; Fig. 5). Discussion Changes in food abundance led to foraging responses by moulting grebes in the Odiel marshes, as previously recorded in other waterbirds (Guillemain et al., 2000a; Systad et al., 2000; Guillemain & Fritz, 2002; Varo & Amat, 2008). When brine shrimps were less abundant in 2008, grebes increased foraging effort, 123 168 Hydrobiologia (2011) 664:163–171 Table 3 Body mass (g) variations of black-necked grebes during the moulting periods of 2008 and 2009 80 2008 2009 N2 2008 n 2009 Mean s.d. n P Mean s.d. 70 N1 a3 a4 s1 82 48 73 386.6 405.1 376.5 43.8 46 41.1 84 72 35 379.9 357.7 382 38.3 35.3 45 0.288 \0.001 0.533 s2 s3 24 68 382 368.1 29.1 41.2 89 108 361.3 373.2 40.6 41.5 0.021 0.426 s4 o1 o2 o3 69 36 64 61 392 394.5 391.8 404.2 44.2 39.9 37.1 42.5 99 135 78 234 382.6 382.3 385.8 394.4 45 50.7 42.2 44.6 0.191 0.184 0.372 0.122 o4 n1 n2 56 152 68 391.6 389.5 387 40.6 48.3 41.9 229 85 198 383.4 386.7 398.9 43.8 46.3 42.1 0.206 0.663 0.045 n3 56 393.4 49 n4 d1 27 100 377.9 382 44.9 37.1 88 119 199 399 44.8 0.481 403.6 406.8 45.4 48.2 0.008 \0.001 foraging activity Date N1 N2 D1 60 D1 O2 50 S2 40 O1 S1 O1 30 S1 O2 S2 20 0.0 0.2 0.4 0.6 0.8 1.0 1.2 brine shrimp abundance Differences between years were tested with Student t tests. a = August; s = September; o = October; n = November; d = December; 1 = first week; 2 = second week; 3 = third week; 4 = fourth week Fig. 5 Relationship between brine shrimp abundance (dry mass of brine shrimp per 100 l of filtered water) and the percentage of grebes foraging in 2008 and 2009. Each point denotes a different sampling day. Sample dates are indicated beside points (s = September; o = October; n = November; d = December; 1 = first half of each month; 2 = second half of each month) probably to compensate for a reduced foraging intake rate. In both years, the foraging effort of grebes also increased as the moulting season progressed, coinciding with a decrease in the availability of brine shrimps. This seasonal increase in foraging effort may partly have been due to the reduction in the number of daylight hours available for foraging (Krams, 2000; Systad et al., 2000). Water temperature is likely to have also been important for two reasons: (1) as temperature decreased, heat loss by convection will have increased, raising the energy requirements of the grebes, and (2) the decline in shrimp abundance in early winter, towards the end of the moulting period, was related to cold winter temperatures (Sánchez et al., 2006a). Feeding behaviour might also be affected by other factors such as wind speed (Green et al., 1999; Heath et al., 2008), or the time of day when data were recorded (Heath et al., 2008), but in our study all foraging observations were conducted at the same time and on days without wind. In the Great Salt Lake, grebes also increased their time spent foraging when brine shrimp densities were low (Caudell & Conover, 2006b). Our analysis of gizzard contents confirmed the dominance of brine shrimps in the grebe diet at the Odiel marshes. The major alternative prey for waterbirds are benthic chironomid larvae (Sánchez et al., 2006c, d), whose hard head parts are resistant to digestion (Sánchez et al., 2005) but were absent from our samples. Shrimp availability determined the feeding method of moulting grebes at Odiel marshes. Grebes fed mainly by diving, despite a higher energetic cost than feeding at the surface (Bevan & Butler, 1992; de Leeuw, 1996; Quintana et al., 2007). Grebes forage only during daylight hours because they are visual predators (Cooper et al., 1984) and brine shrimps undergo vertical diurnal migrations, concentrating at the bottom during daytime so as to escape avian predation (Britton et al., 1986). The Odiel marshes are frequented by shorebirds and gulls unable to access Artemia on the bottom (Sánchez et al., 2006a, d). The largest Artemia were concentrated at the bottom, and these probably have a relatively higher energetic content than smaller ones and provide a higher intake rate for grebes (Caudell & Conover, 2006a; Sánchez et al., 2006c). Furthermore, 123 Hydrobiologia (2011) 664:163–171 169 Artemia occur at higher density at the bottom of the water column (C. Matesanz, M.I. Sánchez and N. Varo, unpublished data). Thus, grebes probably increase their energy intake by increasing diving effort to get to more and larger shrimps, thus meeting the high energy demands during moulting (i.e. Winkler & Cooper, 1986; Jehl, 1988, 1990b). As expected, shrimp abundance appeared to influence the population dynamics of grebes at the Odiel marshes. Food availability may regulate animal population dynamics (Martin, 1987; Odonoghue & Krebs, 1992; Abbott et al., 2008), and the greater availability of brine shrimps found in 2009 might explain the higher numbers of grebes that year. Similarly, it has previously been shown that the density of migrating shorebirds in our study area was related to the availability of aquatic invertebrates (Sánchez et al., 2006a, d). Likewise, the density of wintering shoveler Anas clypeata was related to zooplankton density in French sewage ponds (Guillemain et al., 2000b). Moreover, the combination of a change in shrimp supply and water temperature between years seems likely to explain the recorded differences in grebe body mass at the end of the two moulting periods. Grebes had a higher body mass at the end of the moulting period in 2009, coinciding with greater brine shrimp biomass and higher water temperatures, suggesting that body mass, and especially fat stores, were influenced both by energetic gain when food supply is greater and energetic loss due to colder water. Diving birds typically decrease body temperature when submerged, so their foraging costs increase when thermal stress is greatest (Bevan & Butler, 1992; Heath et al., 2008). The sharp decline in brine shrimp abundance at the end of autumn observed in the Odiel marshes (see also Sánchez et al., 2006a) is also observed in North America (Cooper et al., 1984; Wurtsbaugh & Gliwicz, 2001). We estimated that grebes consumed between 0.18 and 2.04% of the standing crop of Artemia per day, with a gradual increase in this percentage as the season progressed (Table 4). Estimations of energetic Table 4 Estimated standing crop, energy content of standing crop, daily energetic requirements of the grebe population and daily percentage of standing crop consumed by grebes Standing crop (dry mass, g) Energy content of standing crop (kJ) 2008 s2 s3 5,836.800 5,289.600 127,709.184 115,736.448 445 451 234,034 237,190 0.18 0.20 o1 o2 o3 o4 2,128.000 4,256.000 3,708.800 4,560.000 46,560.640 93,121.280 81,148.544 99,772.800 848 823 987 1,391 445,980 432,832 519,083 731,555 0.95 0.46 0.64 0.73 n1 n4 2009 s2 s3 1,276.800 1,033.600 27,936.384 22,615.168 1,085 875 570,623 460,180 2.04 2.03 2,796.800 6,931.200 61,193.984 151,654.656 652 973 342,899 511,720 0.56 0.33 o1 o2 o3 4,195.200 5,350.400 6,080.000 91,790.976 117,066.752 133,562.521 1,743 1,920 1,914 916,679 1,009,766 1,006,611 1.00 0.86 0.75 o4 n1 n4 6,292.800 4,316.800 2,857.600 137,686.464 94,451.584 62,524.288 2,023 2,348 1,524 1,063,936 1,234,860 801,502 0.77 1.31 1.28 Date Grebe numbers Energetic requirements of grebe population (kJ) Standing crop consumed (%) Energy requirements of grebes were estimated as 525.92 kJ per bird per day, and the energy content of dry Artemia as 21.88 kJ per g of dry mass (after Caudell & Conover, 2006a). Pond volume was estimated as 680,000 m3 (76 h of surface * 0.80 m of mean depth). a = August; s = September; o = October; n = November; d = December; 1 = first week; 2 = second week; 3 = third week; 4 = fourth week 123 170 requirements of grebes did not account for the effects of changes in water temperature, so our estimates probably underestimated the strength of the seasonal increase in the daily % standing crop consumed by grebes. It is unclear what contribution grebe predation made to the seasonal decline in Artemia biomass. It is likely that grebe predation accelerates the decline in Artemia biomass, although it may not be the principal cause, which is more likely to be the declining temperatures (Cooper et al., 1984; Wurtsbaugh & Gliwicz, 2001). Similarly, in sewage ponds, shovelers were thought to accelerate a winter decline in zooplankton density (Guillemain et al., 2000a). Furthermore, predation by shorebirds on spring migration has been shown to reduce the density and size of benthic chironomid larvae at Odiel marshes (Sánchez et al., 2006c). Further studies analysing recruitment rates and the effect of cestodes (Sánchez et al., 2006b) on brine shrimp populations will help us to elucidate the relative importance of grebe predation on shrimp population dynamics. To our knowledge, this article presents the first detailed study of the foraging behaviour of moulting black-necked grebes in Europe. It is likely that moulting grebes are only able to adjust foraging effort within certain limits, given the number of daylight hours and the need to spend time in other activities (Caudell & Conover, 2006b). At the end of moulting period when thermal stress is greatest and food supply is lowest in the Odiel marshes, grebes may not be able to meet their energy requirements. Grebes probably responded by shifts in spatial distribution and left our study area when they were no longer in energy balance. This study highlights the value of studying foraging behaviour to infer the effects of changing habitat conditions on population dynamics and also has implications for conservation. The moulting grebe population in the Odiel marshes is highly dependent on the population of A. parthenogenetica, which is under threat of possible invasion by the invasive A. franciscana, which has eliminated native Artemia from many sites in the Iberian Peninsula (Amat et al., 2005) and appears to be a less suitable prey for waterbirds. In addition, the corixid Trichocorixa verticalis has a major predatory effect on Artemia in its native range in North America (Wurtsbaugh, 1992) and has recently invaded areas close to the Odiel marshes (Rodrı́guez-Pérez et al., 2009; Van de Meutter et al., 2010). 123 Hydrobiologia (2011) 664:163–171 Therefore, future management activities should attempt to minimize any impact these invasions may have on the A. parthenogenetica and blacknecked grebe populations. Acknowledgments Many volunteers participated in the capture of grebes, which was organized by Equipo de Seguimiento de Procesos Naturales (Estació n Bioló gica de Doñ ana, CSIC) and Sociedad Españ ola de Ornitologı́a. We also thank Enrique Martı́nez, Director of Paraje Natural Marismas del Odiel, for granting access to the study site and facilities to conduct the field work, as well as anonymous referees for their comments on previous versions. Our study was financially supported by the Consejerı́a de Innovació n, Ciencia y Empresa (Junta de Andalucı́a, project P07-CVI-02700). References Abbott, K. C., W. F. Morris & K. Gross, 2008. Simultaneous effects of food limitation and inducible resistance on herbivore population dynamics. Theoretical Population Biology 73: 63–78. Amat, F., F. Hontoria, O. Ruiz, A. J. Green, F. Hortas, J. Figuerola & F. Hortas, 2005. The American brine shrimp as an exotic invasive species in the western Mediterranean. Biological Invasions 7: 37–47. Bevan, R. M. & P. J. Butler, 1992. The effects of temperature on the oxygen consumption, heart rate and deep body temperature during diving in the Tufted duck, Aythya fuliga. Journal of Experimental Biology 163: 139–151. Britton, R. H., E. R. de Groot & A. R. Johnson, 1986. The daily cycle of feeding activity of the Greater Flamingo in relation to the dispersion of the prey Artemia. Wildfowl 37: 151–155. Caudell, J. N. & M. R. Conover, 2006a. Energy content and digestibility of brine shrimp (Artemia franciscana) and other prey items of eared grebes (Podiceps nigricollis) on the Great Salt Lake, Utah. Biological Conservation 130: 251–254. Caudell, J. N. & M. R. Conover, 2006b. Behavioral and physiological responses of Eared Grebes (Podiceps nigricollis) to variations in brine shrimp (Artemia franciscana) densities. Western North American Naturalist 66: 12–22. Charnov, E. L., 1976. Optimal foraging, the marginal value theorem. Theoretical Population Biology 9: 129–136. Cooper, S. D., D. W. Winkler & P. H. Lenz, 1984. The effect of grebe predation on a brine population. Journal of Animal Ecology 53: 51–64. de Leeuw, J. J., 1996. Diving costs as a component of daily energy budgets of aquatic birds and mammals: generalizing the inclusion of dive-recovery costs demonstrated in tufted ducks. Canadian Journal of Zoology 74: 2131–2142. Garcı́a-Jiménez, F. J. & J. F. Calvo-Sendı́n, 1987. El zampullı́n cuellinegro, Podiceps nigricollis, en la laguna de la Mata (Alicante). Ardeola 34: 102–105. Green, A. J., A. D. Fox, B. Hughes & G. M. Hilton, 1999. Time-activity budgets and site selection of white-headed Hydrobiologia (2011) 664:163–171 Ducks (Oxyura leucocephala) at Burdur Lake, Turkey in late winter. Bird Study 46: 62–73. Guillemain, M. & H. Fritz, 2002. Ecomorphology and coexistence in dabbling ducks: the role of lamellar density and body length in winter. Oikos 98: 547–551. Guillemain, M., H. Fritz & N. Guillon, 2000a. Foraging behavior and habitat choice of wintering Northern Shoveler in a major wintering quarter in France. Waterbirds 23: 353–363. Guillemain, M., H. Fritz & N. Guillon, 2000b. The use of an artificial wetland by Shoveler Anas clypeata in western France: the role of food resources. Revue D Ecologie-La Terre Et La Vie 55: 263–274. Heath, J. P., W. A. Montevecchi & G. J. Robertson, 2008. Allocating foraging effort across multiple time scales: behavioral responses to environmental conditions by Harlequin ducks wintering at Cape St. Mary’s, Newfoundland. Waterbirds 31: 71–80. Iborra, O., F. Dhermain & P. Vidal, 1991. L’hivernage du grèbe à cou noir sur l’Etang de Berre (Bouches-duRhône). Alauda 59: 195–205. Jehl Jr., J. R., 1988. Biology of the eared grebe and willson’s phalarope in the no-breeding season: a study of adaptations to saline lakes. Studies in Avian Biology 12: 1–74. Jehl Jr., J. R., 1990a. Aspects of the molt migration. In Winner, E. G. (ed.), Migration Physiology and Ecophysiology. Springer-Verlag, Berlin: 102–113. Jehl Jr., J. R., 1990b. Field estimates of energetics in migrating and downed black-necked grebes. Journal of Avian Biology 24: 63–68. Krams, I., 2000. Length of feeding day and body weight of great tits in a single-and a two-predator environment. Behavioral Ecology and Sociobiology 48: 147–153. Martin, T. E., 1987. Food as a limit on breeding birds: a lifehistory perspective. Annual Review of Ecology and Systematic 18: 453–487. Odonoghue, M. & C. J. Krebs, 1992. Effects of supplemental food on snowshoe hare reproduction and juvenile growth at a cyclic population peak. Journal of Animal Ecology 61: 631–641. Piersma, T., 1988. Breast muscle atrophy and constraints on foraging during the flightless period of wing moulting great crested grebes. Ardea 76: 96–106. Quintana, F., R. P. Wilson & P. Yorio, 2007. Dive depth and plumage air in wettable birds: the extraordinary case of the imperial cormorant. Marine Ecology Progress Series 334: 299–310. Rodrı́guez-Pérez, H., M. Florencio, C. Gómez-Rodrı́guez, A. J. Green, C. Diaz-Paniagua & L. Serrano, 2009. Monitoring the invasion of the aquatic bug Trichocorixa verticalis verticalis (Hemiptera: Corixidae) in the wetlands of Doana National Park (SW Spain). Hydrobiologia 634: 209–217. 171 Salomonsen, F., 1968. The moult migration. Wildfowl 19: 5–24. Sánchez, M. I., A. J. Green & E. M. Y. Castellanos, 2005. Seasonal variation in the diet of the Redshank Tringa totanus in the Odiel Marshes, southwest Spain: a comparison of faecal and pellet analysis. Bird Study 52: 210–216. Sánchez, M. I., A. J. Green & E. M. Castellanos, 2006a. Temporal and Spatial variation of an invertebrate community subjected to avian predation at the Odiel salt pans (SW Spain). Archiv für Hydrobiologie 166: 199–223. Sánchez, M. I., B. B. Georgiev, P. N. Nikolov, G. P. Vasilieva & A. J. Green, 2006b. Red and transparent brine shrimps (Artemia parthenogenetica): comparative study of their cestode infections. Parasitological Research 100: 111–114. Sánchez, M. I., A. J. Green & R. Alejandre, 2006c. Shorebird predation affects density, biomass, and size distribution of benthic chironomids in salt pans: an exclosure experiment. Journal of the North American Benthological Society 25: 9–18. Sánchez, M. I., A. J. Green & E. M. Castellanos, 2006d. Spatial and temporal fluctuations in presence and use of chironomid prey by shorebirds in the Odiel saltpans, south-west Spain. Hydrobiologia 567: 329–340. Schatz, G. S. & E. McCauley, 2007. Foraging behaviour of Daphnia in stoichiometric gradients of food quality. Oecologia 153: 1021–1030. Systad, G. H., J. O. Bustnes & K. E. Erikstad, 2000. Behavioural responses to decreasing day length in wintering sea ducks. Auk 117: 33–40. Van de Meutter, F., H. Trekels & A. J. Green, 2010. The impact of the North American waterbug Trichocorixa verticalis (Fieber) on aquatic macroinvertebrate communities in southern Europe. Fundamental and Applied Limnology 177: 283–292. Van Impe, J., 1969. Concentración enorme de Podiceps nigricollis, Brehm, en Dobroudja-Roumanie. Alauda 37: 77–79. Varo, N. & J. A. Amat, 2008. Differences in foraging behaviour of sympatric coots with different conservation status. Wildlife Research 35: 612–616. Winkler, D. W. & S. D. Cooper, 1986. Ecology of migrant black-necked grebes, Podiceps nigricollis, at Mono Lake, California. Ibis 128: 483–491. Wurtsbaugh, W. A., 1992. Food-web modification by an invertebrate predator in the Great-Salt-Lake (USA). Oecologia 89: 168–175. Wurtsbaugh, W. A. & Z. M. Gliwicz, 2001. Limnological control of brine shrimp population dynamics and cyst production in the Great Salt Lake, Utah. Hydrobiologia 466: 119–132. 123