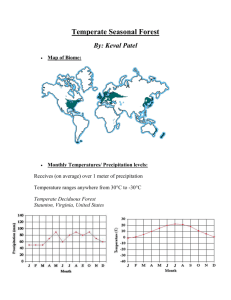
32 Global Ecology C h A p T e R
advertisement

Global Ecology 3 2 – 1 Chapter 32 Global Ecology Drunken forest As permafrost melts, a process now occurring at an increased rate because of climate change, trees formerly anchored in the icy soil shift and tilt, as seen in this “drunken forest” of black spruce near Fairbanks, Alaska. Organic material trapped in the ice for thousands of years decays, releasing enormous amounts of carbon dioxide and methane, gases that trap heat and warm the planet. chapter outline Life on the Land Rainforests Deciduous Tropical Forests Savannas Deserts Grasslands Temperate Deciduous Forests Temperate Mixed and Coniferous Forests Mediterranean Scrub The Northernmost Forests—Taiga and Boreal Forest Arctic Tundra A Final Word T o give a proper context for a broad view of world vegetation, we first zoom in to look at two small patches: one in the wet tropics and one in the north temperate zone. Consider first an area measuring 10 meters by 10 meters in the tropics. As we know, ecosystem function depends on the trapping of solar energy and its conversion into plant biomass, which then feeds not only the higher trophic levels but also the decomposers that digest dead biomass. The solar energy that drives photosynthesis is, however, only a tiny fraction of the energy picture. Most of the solar energy that reaches this tropical patch provides the heat that keeps it in the range of 20° to 35°C, driving the evaporation and transpiration (the “evapotranspiration”) that release water vapor into the air. Since we are in the wet tropics, the precipitation input exceeds the evapotranspiration 32–1 output of this small area, and the excess water runs off into rivers and streams. We also know that this patch will experience only moderate climate shifts from day to day, week to week, and month to month. Much of the climate variability in this ecosystem relates to the cycle of day and night, and much less to the seasons, which vary little in the wet tropics. Now let’s look at a patch of the same size in the moist temperate zone of North America. We find, as in the wet tropics, that precipitation exceeds evapotranspiration and that infiltration and runoff feed streams and rivers. But in contrast, there CHEC K POINT S After reading this chapter, you should be able to answer the following: 1. What is a biome, and what factors affect the distribution of biomes on Earth? 2. Given that tropical rainforests contain such a diversity of species, why are many kinds of tropical soils unsuitable for agriculture? 3. Which factors favor grasslands over wooded ecosystems? What role does fire play in grasslands? 4. Which best characterizes a desert: high temperature or low precipitation? How have plants adapted to desert living? 5. How does the appearance of the temperate deciduous forest biome change from one season to the next? What accounts for these changes? 6. What is the general global pattern for biodiversity, and what factors might explain it? 32–2 C ha p t e r 3 2 Global Ecology are large differences between seasons, a variation that is crucial to the functioning of this ecosystem. The seasons range from long, hot days in midsummer to short, often bitterly cold days in winter. During the winter, photosynthesis is either low (as for an evergreen spruce tree) or completely lacking (as for a deciduous maple tree). The growth that occurs between the last killing frost of the spring and the first killing frost of the autumn is substantial. It even equals, in some cases, that of the continuously productive tropics. Clearly, then, what happens locally depends on global patterns. Starting with energy, on any single day when plants are growing, the amount of solar radiation received that is in the restricted wavelengths active in photosynthesis (Chapter 7 of the textbook) sets an upper limit to productivity. The temperature of the air and of the soil must remain in a range that allows metabolic functioning. The soil moisture also sets a maximum for productivity. Without adequate water supplies, the photosynthetic machinery will slow or even shut down entirely. For rainfall to exceed evapotranspiration, the extra water must come from somewhere outside a local patch. And so we need to consider how moisture moves from places with more moisture—the oceans—to places with less. To understand any locality, we must understand the functioning of the global climate machinery by which heat and moisture are redistributed around the globe. But many other factors besides regional climate come into play in shaping a local ecosystem. If we were to shift our 10-square-meter patch by 100 meters in any direction, we would not find exactly the same collection of organisms. Usually, we would have some, perhaps most, of the species we had at the first location. But they might also be quite different. The changes we would see in this 100-meter shift would not be related to the regional solar radiation or rainfall. Rather, they would depend on many local factors such as the nature of the substrate, associated species, the slope of the land (which affects the solar radiation received), and the presence or absence of standing water. It would also be affected by the history of extreme events, such as fire, windthrow, or clearing of the land for agriculture. Suppose, for example, that our 100-meter shift in the temperate forest crosses a geologic boundary—from soil formed in glacial till, nutrientrich and high in the finer soil particles (silts and clays) that retain moisture, to a coarse, deep outwash sand, nutrient-poor and subject to drought. We would expect the plant communities to be different in these two places, even though the climate would be the same. Scientists who try to explain why vegetation in one place differs from that in another are very aware that some differences require attention to local factors, others to global factors. Despite the local variation, there are repeating patterns on the global scale. The similarity between two patches of vegetation in different parts of the world can arise in two ways. Species that are related by descent (they share a near ancestor) will, in general, look similar and function in a similar way. Species that are distantly related (they diverged from their common ancestor a long time ago) will be distinctly different genetically, but nevertheless may be similar in appearance and function. Consider deciduous forests in Europe and in the eastern United States. Many of the genera are the same—for example, oaks (Quercus), beeches (Fagus), and ashes (Fraxinus). Most of us, parachuted into the middle of such a forest without knowing which continent we were on, would not be able to deduce where we were from the general look of the vegetation. In this example, the abundant species are related by descent and are also morphologically and physiologically similar, because the climate and soils are very similar. We can repeat the experiment by dropping into a high-rainfall tropical area in South America or into an area with a similar climate in the Indonesian archipelago. The species and genera would mostly be different in these two areas, but unless you knew tropical rainforest taxonomy, the general appearance of the vegetation would give few clues to your geographic whereabouts. Similarities that arise despite genetic differences evolve because similar habitats favor similar morphologies and physiological adaptations (Figure 32–1). The process by which natural selection produces similarity in unrelated species is called convergent evolution (page 239). But there are changes over time as well as across space. If we revisit a 10-square-meter plot after leaving it for a day, we expect it to be much the same but not absolutely identical in every detail. If we revisited the site a year, 10 years, or 100 years later, we would not be surprised to find major changes. In a region subject to fire, we might find a forest recovering from a wildfire, seeing only young trees and shrubs. If we came back a million years later, a formerly dry site might now be under water, and even if the substrate were similar, new species would probably have evolved. These kinds of changes over time are another source of variation. In our modern world, changes caused by humans have become all-pervasive and often drastic. A forest can be replaced by a shopping center. Armed with these concepts, we begin our survey of world vegetation from what we might call the 3000-meter level—as viewed from an imaginary blimp flying over the globe and traversing the major gradients: tropics to temperate to Arctic, wet to dry, and low elevation to high elevation. Thanks to the Google Earth website, we can take such a virtual journey. It allows us to locate nearly any place on Earth and zoom in and out to view it on different scales (see the essay “Google Earth: A Tool for Discovering and Protecting Biodiversity” on page 241). If you do this, you will get a sense of the pervasiveness of human activity. There are places (such as Antarctica and the driest deserts) where human influence is not apparent, but overall, your virtual world tour will clearly show that humans have substantially modified the Earth. This means that a study of the Earth’s present ecosystems must include an understanding of the ecology of human land use and of the interactions between humans and nature. For more than 99.9 percent of the Earth’s history, though, there were no humans. And although other organisms have affected their environments—consider how photosynthesizers oxygenated the atmosphere—none have caused such drastic change so rapidly. It is now estimated that 40 percent of the Earth’s surface is devoted to agriculture, and much of the remaining 60 percent has been significantly altered by humans—just one example being the cutover forests of the United States. Understanding what the world’s landscapes looked like before their modification by humans is important if we are to understand our contemporary biosphere and, most importantly, if we are to grasp what we may lose if we fail to take steps to save this rich biological heritage. Life on the Land 3 2 – 3 (a) (b) 32–1 Convergent evolution at high elevations (a) Espeletia pycnophylla growing in the high-altitude ecosystem known as the páramo in the Andes, as seen here in Ecuador, South America. (b) Dendrosenecio adnivalis growing at a high elevation on Mount Gessi in Tanzania, Africa. The unique challenges of a habitat that freezes almost every night and warms in the day favor a compact tree form with robust leaves and well-protected growing points. These similar forms evolved independently on two distant continents, a striking example of convergent evolution. How can we determine what the Earth was like before the wholesale modifications brought about by our species? It is often possible to see the signature of the original vegetation. Imagine flying our blimp from coast to coast over the United States, starting in New York and heading west. You notice that until you cross the Mississippi River, uncultivated and minimally managed lands are usually forested with broad-leaved deciduous trees. If very wet, these areas are covered with marsh. Crossing central Illinois, you may not see clear evidence that in 1800, this area was mostly grassland, because most of the land is cultivated. As you head farther west, however, you see that uncultivated lands support fewer trees, except along rivers and streams, and it becomes obvious that grassland is the dominant natural vegetation. Over the Rocky Mountains, you cross from grasslands back into forests as the elevation increases, but now the dominant trees are evergreen conifers, though there will be grassy valleys and areas of treeless alpine tundra. On the western side of the Rockies in Colorado, with the land dropping in elevation and with dryness increasing, you see vast expanses of shrubby grasslands punctuated with forested hills. Crossing the Sierra Nevada in California, you encounter a complex landscape of coniferdominated mountain slopes, open oak woodlands, grasslands, and shrub communities of chaparral and coastal sage. Over the Central Valley you see one of the world’s most productive agricultural areas and little trace of the formerly extensive marsh areas drained to create it. Completing your trip on the beach in Santa Monica, California, you may wonder how to describe and explain the variation you have seen. To answer this question, we employ the concept of the biome, which we define as a type of vegetation that is reasonably homogeneous over large terrestrial areas with respect to the growth forms, sizes, and arrangement of the dominant plant species. Each biome also has an associated suite of animals. Like humans, they affect vegetation, but in general they are not defining characteristics of the biome. Life on the Land The scheme we present for classifying the world’s natural vegetation into a set of biomes is based on the work of the late A. W. Küchler, of the University of Kansas (Figure 32-2). This scheme is, of course, a broad-brush generalization. Making a map of this kind requires delineating areas. But a line on a map does not usually correspond to an easily discernible “line” on the ground. Similarly, the area inside the lines includes a lot of variation, and the larger the area of the unit, the greater the variation within. For example, biome 5 on the map—alpine tundra and mountain forests—encompasses continuous forest and treeless alpine tundra. Not mentioned, but also included in this biome, are mountain meadows covered with grasses, sedges, and other herbaceous plants, with no trees, or only widely scattered ones. Mapped biomes generally contain very different communities within their boundaries. What are the characteristics of the biomes, passing from the poles to the equator? By ignoring the often strong local effects of elevation (mountains) and moving only through regions with plenty of moisture during the growing season, we can attribute the dramatic changes to differences in temperature that, in turn, depend primarily on the amount of solar radiation received, which is largely explained by latitude. It is the geometry of the Earth and its path through space with respect to the sun that creates the relationship between latitude and solar radiation. First consider the effects of the Earth’s nearly spherical shape. Imagine 32–4 C ha p t e r 3 2 Global Ecology 1 2 8 8 7 7 4 10 4 5 4 4 4 6 9 13 2 2 5 1 11 2 10 12 9 10 6 Mixed west-coast forests 7 Arctic tundra 8 9 Ice desert 10 11 3 16 5 Temperate mixed forests Subtropical mixed forests Taiga Northwestern coniferous forest Alpine tundra and mountain forests 3 4 4 8 7 12 13 10 5 16 Grasslands Savannas Mediterranean scrub Deserts and semideserts Juniper savanna Southern woodland and scrub 17 14 15 16 16 Tropical mixed forests Monsoon forests 17 Rainforests 17 14 Temperate deciduous forests 10 9 17 16 16 10 12 5 17 15 3 14 11 5 6 12 9 that the Earth has an axis of rotation perpendicular to the plane of its orbit. It does not, of course, because the Earth is tilted to approximately 23.45°, but if it were perpendicular, the solar radiation per unit surface area of the Earth would decrease with increasing north and south latitude, because the same amount of sun energy would be spread over an increasingly larger area. Moreover, since the Earth has an atmosphere that can reflect and absorb incoming energy, and the incoming solar radiation traverses a greater depth of atmosphere at higher latitudes before reaching the surface, the amount of solar energy hitting the ground is further diminished away from the equator. Tilting the Earth’s axis to its correct angle of 23.45° and setting the planet in motion around the sun, we can see that not only do the same equator-to-pole patterns still apply, but as the Earth travels around the sun, first the Northern Hemisphere and then the Southern Hemisphere is “tilted toward the sun.” At 23.45° north latitude, for example, at the June summer solstice, the sun at noon is directly overhead, and solar radiation per unit 17 32–2 Biomes of the world The information in these maps and the accompanying key was originally supplied by A. W. Küchler of the University of Kansas, one of the leading authorities on the distribution of biomes. Because of the global coverage of the maps, the scale is relatively small and the content is generalized. Any given biome is not always uniform, and all the biomes include considerable variations in vegetation. The boundaries between the biomes may be sharp, but more often they are blurred, consisting of broad zones of transition from one type of vegetation to another. area at the Earth’s surface is at its maximum—discounting the possible effect of clouds and dust. On this same day, poleward from 23.45° north, daylength increases, reaching 24 hours above the Arctic Circle. In temperate zones, this seasonal cycle reveals itself by the rising and falling of the path of the sun. Solar input is at a maximum at the summer solstice and a minimum at the winter solstice—a pattern well understood and carefully followed even in preliterate antiquity. Although temperature at any point is dependent on the solar radiation received, this is significantly modified by the air and water currents that redistribute heat. For the Earth as a whole, energy absorbed in the warmer tropical regions is transferred to higher latitudes, so that many parts of the globe are warmer than the solar radiation received at their latitude would indicate. Study of air and water movements has revealed large-scale global patterns (Figure 32–3) consisting of latitudinal circulation bands. Within the equatorial zone (23.45° south to 23.45° north), the heating promotes evaporation and transpiration, and warm, moist Life on the Land 3 2 – 5 8 8 7 7 7 4 2 1 10 10 1 4 10 1 4 5 9 9 10 2 5 11 5 9 4 5 12 12 2 11 5 9 2 1 3 5 3 12 12 17 16 16 5 10 17 16 16 5 10 17 12 17 17 5 5 17 10 15 16 16 9 10 9 17 12 10 12 12 9 10 11 11 17 11 3 6 air ascends. The rising air drops much of its moisture in the tropical zone, but it sets up a cyclical pattern, and the air, with much of its moisture removed, tends to descend at about 30° north and south. The descending air warms as pressure increases, causing the relative humidity to decline even further and to produce a stable atmosphere. The cyclical flow initiates winds that, because of the rotation of the Earth, trend away from the equator-to-pole flow to create the westerlies and trade winds (Figure 32–3).These global patterns explain why the most extreme deserts in the world are found around 30° north and south, and why the regions of highest rainfall in North and South America are found at the higher latitudes of their west coasts. Another factor that determines the aridity of a particular spot on land is how far it lies from the moisture-rich air of the oceans. The elevation of land—whether mountains or sea-level plains—has a significant influence on local climate, and the change in vegetation with increasing elevation can be dramatic over very short distances. To explain this, we need to remember 10 6 9 that it is primarily the absorption of short-wave energy at the Earth’s surface that captures the heat energy of the sun. The sunwarmed surface radiates long-wave radiation back into the atmosphere. Some of this is dissipated into space, but much is returned to Earth by the dust, water vapor, and carbon dioxide, as well as other greenhouse gases, that absorb the re-radiated heat, warming the air and the surface. In general, air temperature decreases with increasing elevation above the ground. There are reversals of this usual trend, called inversions, in which for some distance the air temperature actually increases with altitude, but these are local and generally temporary exceptions. On average, air temperature drops by about 6.4°C for each 1000 meters of elevation (3.5°F for each 1000 feet), with the observed rate of change depending on time of day, moisture content of the air, wind patterns, and other topographic and weather variables. Because of the decrease in temperature with altitude, locations on the Earth that are topographically higher have significantly cooler temperatures, on average, than 32–6 C ha p t e r 3 2 Global Ecology 32–3 Global air currents The Earth’s surface is covered by belts of air currents, which determine the major patterns of wind and rainfall. In this diagram, the blue arrows indicate the direction of movement of the air within the belts. The green arrows indicate regions of rising air, which are characterized by high precipitation, and the brown arrows indicate regions of descending air, characterized by low precipitation. The dry air descending at latitudes of 30° north and south is responsible for the great deserts of the world. The prevailing winds on the Earth’s surface, indicated by the black arrows, are produced by the twisting effect of the Earth’s rotation on the air currents within the individual belts. Green arrows: Moist air rises and cools— high precipitation 60° N Westerlies 30° N Northeast trade winds 0° Direction of Earth’s rotation Equatorial doldrums Southeast trade winds 30° S Brown arrows: Dry air descends and warms— low precipitation Westerlies 60° S locations at the same latitude at lower elevations. This effect is slightly counterbalanced by the fact that the air is thinner (because air pressure, and therefore density, also decreases with elevation) and the path of sunlight through the absorbing atmosphere is somewhat shorter. The cooling with increasing altitude has profound climatic consequences when air masses move across mountains. Because cool air cannot contain as much moisture as warm air, moisturerich air masses produce precipitation as they are carried upward. The highest rainfall on Earth occurs in places where this effect is particularly strong. For example, Mount Waialeale on Kauai, Hawaii, receives moisture-laden air driven by the northeast trade winds. The annual precipitation at the highest elevations (1554 meters) on the windward (north and northeast) slopes averages about 9.5 meters (31 feet). This is extreme, but we typically find wetter climates on mountains’ windward slopes than on the adjacent lowlands or on the leeward slopes. As the air descends, it warms, and its capacity to contain moisture correspondingly increases, making rainfall less likely. For example, the leeward side of Mount Waialeale receives only about 50 centimeters of annual precipitation. This downwind dryness is the “rain-shadow” effect (Figure 32–4). It is particularly strong along the Pacific coast of North America, with zones of aridity or semiaridity on the east side of mountain chains, from the northern United States south into Mexico. In South America the pattern is also strong, except that in northern South America the rain shadow is on the western side of the Andes, and in southern South America (Patagonia) the rain shadow is on the east—a pattern explained by the shift in prevailing wind direction (Figure 32–3). The climatic changes that you observe when climbing a mountain mimic the changes that occur as you travel toward the poles from the equator—mainly with respect to temperature and the occurrence of freezing conditions (Figure 32–5). An increase in elevation of 100 meters (328 feet) produces a change in mean temperature corresponding to an increase in latitude of approximately 1 degree. Climbing a tall mountain can take you through a sequence of ecosystem types that resembles the sequence you might encounter if you went north toward the Arctic. For example, in the southwestern United States, the highest mountains are topped by conifer-dominated forests, then, at higher elevations, by tundra-like meadows and scree fields, and eventually by a zone of perpetual snow cover. But this “replicate the latitudinal Descending, warming air Rising, cooling air Rain shadow Prevailing wind 32–4 Rain shadow The effect of coastal mountains on patterns of precipitation in the Northern Hemisphere. As winds come off the water, the air is forced upward by the contour of the land, cools, and releases its moisture in the form of rain or snow. As the air descends on the far side of the mountains and becomes warmer, its capacity to hold water increases, so the amount and frequency of precipitation decrease. Rainforests 3 2 – 7 Snowcapped peak lin Snow e Tundra l in Tree Altitude → e Taiga Temperate deciduous forest Polar ice Tropical rainforest Equator Latitude → Pole 32–5 Relationship between altitude and latitude In the Northern Hemisphere, the sequence of vegetation changes we would see when ascending a tall mountain is similar to the sequence we would see while traveling hundreds of kilometers to the north. Alexander von Humboldt was the first to point out the relationship between altitude and latitude. To experience a similar sequence of vegetation in the Southern Hemisphere, we could ascend a mountain. However, by simply traveling south, we would never encounter vegetation corresponding to the taiga and the tundra of the Northern Hemisphere. Can you explain why? change” pattern becomes less true as one moves into the tropics. On a tropical mountain, the climatic changes as you ascend are dramatic, but the vegetation of the colder upper elevations does not look much like what you would see in Alaska or Siberia. A key difference is that tropical mountains have the same limited seasonality as the lowland rainforests. As a result, the daily temperature variations are much greater than the seasonal average daily temperature extremes. As the ecologist O. Hedberg put it, high-elevation tropical systems have “summer every day and winter every night.” The demands of such extreme diurnal temperature variation have favored the evolution of bizarre plant types, and the high-altitude tropical mountain communities provide some of the most striking examples of convergent evolution (Figure 32–1). Soils and Fire Influence Regional Patterns of Distribution Variation in terrestrial vegetation is often explained by variation in the soil. Soil is best viewed as a sub-ecosystem, because it is not just ground-up rock and dead organic matter but a complex community of highly specialized organisms (see Figure 29–6). It is the functioning of this system that largely determines the abundance and chemical form of the elements made available to plants. It also affects the amount and distribution of soil moisture, which for most soils is strongly dependent on the sizes of the mineral particles, whether mostly microscopic clay particles or coarse sand and gravel. The kind of soil present within a region is partly determined by climate but also by the “parent material” in which the soil forms. Another factor is the length of time a region has been stable and therefore has had surfaces exposed to the processes of soil weathering. Like ecosystems generally, the soil is not a static system. Australia, for example, has been geologically stable over millions of years, and largely because of this, the continuous leaching by precipitation has produced nutrient-poor soils, with deficiencies of essential elements, especially phosphorus. Fire also strongly affects vegetation. Before humans became technologically sophisticated in their destruction of the natural world, fire was the major disturbance—where disturbance is defined as a factor that kills or significantly harms the organisms in an ecosystem. Fire is only possible with fuel (combustible plant material) that is dry enough to burn, and the presence of dry fuel is strongly influenced by climate (for example, whether or not there is a dry season) and by weather (such as wind, the pre-fire pattern of precipitation, and lightning). Thus fire, weather, and climate are intimately connected. Climate determines the plant types present, the overall conditions that control the death of plant parts, and weather conditions at the time of the burn, such as the moisture in the fuel, the humidity of the air, and, especially, the direction and strength of the wind. Few places are absolutely free of the possibility of fire, but those with the highest vulnerability combine a reliable moist growth period to build up biomass with annual episodes of drought to dry it out. These conditions exist over huge areas in the dry tropics and semiarid temperate zones. The boreal forest, though not thought of as an arid region, dries sufficiently and has the right fuel characteristics to permit huge wildfires. There is consensus among ecologists that if, hypothetically, it were possible to prevent all fires, the vegetation of many parts of the world would look very different. We discuss some of these situations in the following sections. Rainforests We begin our exploration of biomes with the system where the fewest factors are limiting: the rainforests—where temperatures are relatively constant and never below freezing, where rainfall is 32–8 C ha p t e r 3 2 Global Ecology A l e x and e r v o n H u mb o ldt Alexander von Humboldt (1769–1859) was perhaps the greatest scientific traveler who ever lived and was certainly one of the greatest writers and scientists of his era. A native of Germany, Humboldt traveled widely across the interior of Latin America from 1799 to 1804 and climbed some of its highest mountains. Exploring the region between Ecuador and central Mexico, he was the first to recognize the incredible diversity of tropical life and, consequently, the first to realize just how vast a number of species of plants and animals there must be in the world. In his travels, Humboldt was impressed with the fact that plants tend to occur in repeatable groups, or communities, and that wherever there are similar conditions—relating to climate, soil, or biological interactions—similar groupings of plants appear. He also discovered a second major principle: the relationship between altitude and latitude. He found that climbing a mountain in the tropics is analogous to traveling farther north (or south) from the equator. Humboldt illustrated this point with his well-known diagram of the zones of vegetation on Mount Chimborazo in Ecuador, which he climbed. On this mountain, he reached the highest elevation on record for any human being up to that date. On leaving Latin America in 1804, Humboldt visited the United States for eight weeks. He spent three of these weeks as Thomas Jefferson’s guest at Monticello, talking over many matters of mutual interest. It is consistent and therefore soil moisture ample for growth throughout the year, where geology and topography allow deep soil formation and retention of at least an adequate supply of nutrients, and where the site is well drained (that is, not under water at any time). These conditions favor high and consistent photosynthesis and maximum plant growth. Plants can expand their canopies rapidly and, without the constraints of freezing and drought, can have large and/or thin leaves for maximum absorption of light. The result is strong competition for light. Plants that fall behind in capturing sunlight are quickly overtopped. Over evolutionary time, this has favored a proliferation of woody species capable of establishing their place in the sun. The wet lowland tropics are dominated by tropical rainforests—dense forests consisting of species that are active all year round. The plants are either evergreen (that is, they always have leaves that are actively photosynthesizing) or leafless for only brief periods. The dominance of trees influences all other aspects of the rainforest. The dense shade means that plant cover beneath the trees in undisturbed forests is sparse. But there are species that have adapted to these conditions and are capable of growing in low light. The seedlings and saplings of some trees are shade tolerant, meaning they can maintain a positive energy balance despite the limited light, and these are often the most abundant plants in the ground layer. thought that Humboldt’s enthusiasm for learning about new lands encouraged Jefferson’s own great scheme for the exploration of the western United States. Thus, it is fitting that Humboldt’s name is commemorated in the names of several counties, mountain ranges, and rivers in the American West. Another class of plants—the epiphytes (meaning “on plants”)—find light by growing on the trunks and branches of the trees. Algae, mosses, and lichens grow as epiphytes in all climatic regions. The wet tropics have these in abundance, but also a host of epiphytic species from the higher plant groups (Figure 32–6). Epiphytes survive without soil by having special adaptations to capture and retain moisture from the frequent rains and by acquiring nutrients from animal droppings, plant litter, dust, and dead insects. Epiphytes have evolved in many plant families, but two well-known groups are the orchids and bromeliads (pineapple is a bromeliad); there are also many epiphytic ferns. Lianas, or woody vines also known as climbers, represent another strategy for exploiting trees for support. Lianas are rooted in the soil but grow up along the trunks of trees into the canopy. Some lianas start as epiphytes and later drop stems to the soil, where they take root. The “strangler figs” (Ficus spp., see page 558) are a special case. They start as epiphytes, extend roots downward into the soil, and eventually envelope and kill their host trees, after which the strangler assumes an independent existence as a canopy tree. Because most edible leafy biomass in rainforests is in the treetops, large herbivores and the predators that feed on them are sparse on the forest floor. This, along with the favorable climate, Rainforests 3 2 – 9 32–6 Epiphytes By collecting and storing water and nutrients from the surrounding air, rain, and dust, epiphytes create little patches of soil from accumulated debris. Bromeliads, seen here as leafy tufts growing along the tree branches, are among the most common of the epiphytes. The leaves of many bromeliads merge at their bases to form watertight tanks that, in the larger species, can hold as much as 45 liters of rainwater. These pools of water are microcosms of bacteria, protozoa, larvae, insects, and insect-eaters. Many rainforest mosquitoes breed exclusively in bromeliad tanks. The bromeliads absorb water from their built-in reservoirs and are also supplied with nutrients from the debris. explains the abundance of arboreal (tree-dwelling) animals, including our primate relatives, such as the howler monkeys of the Americas and the gibbon species of Asia, as well as an abundance of birds (parrots in the Americas and hornbills in Africa). The mobility of primates and birds allows them to find the best patches of foods in the treetop environments—fruits, insects, and the more easily digestible new growth. It also makes them important dispersers of the seeds of rainforest plants. Ecologists are especially interested in the numbers of species that occupy given amounts of space. If the metric is simply the total number in an area, the term “species richness” is employed. Species diversity can be measured by species richness, but to ecologists, “diversity” is a more general term that includes consideration of the distribution of abundance among species. For this general survey, we equate diversity with richness. By any definition, though, tropical rainforests reach extraordinary levels of plant species diversity. A typical hectare (2.47 acres, or about 2.5 American football fields) of boreal coniferous forest has 1 to 4 tree species, and a rich temperate deciduous forest, 10 to 20 species, but tropical rainforests commonly have 100 species, and exceptionally rich forests have hundreds more. Epiphytes, lianas, and the herbaceous plants of the forest floor provide additional species. This diversity of plant species supports a commensurate diversity of arthropods. As with all generalizations in ecology, however, there are exceptions, and some forests of the wet tropics are dominated by one or a few species of trees over large areas. Ecologists are still searching for a full explanation of why tropical rainforests have such high diversity relative to other biomes. Both temperature and moisture conditions are probably important. The influence of warm temperatures is illustrated by the fact that diversity is relatively higher not only in the wet tropics but also in the dry tropics, whereas it is universally much lower in the polar regions. The situation with respect to moisture is not quite so simple. The moist tropics are species-rich, and the driest deserts are generally species-poor, but semiarid regions, such as the shrub-dominated fynbos region of South Africa, have quite high plant biodiversity. Species diversity ultimately is dependent on the process of evolution, and we expect diversity to be higher where there has been more time and more favorable conditions for speciation— so time for evolution may also be a controlling factor. Another idea focuses on trophic interactions, especially parasitism (in the general sense), as a source for selective pressure favoring the origin and persistence of species. The continuously warm and moist conditions of the tropics allow fungi, bacteria, and herbivorous insects to thrive and so to potentially act as selective agents in evolution. The need to fend off evolving parasites and predators may favor genetic diversity, which in turn may favor speciation. Like all natural systems, rainforests are dynamic and constantly changing. Because of the abundant rainfall, natural fires are rare. Rainforest trees have evolved to resist wind damage by a variety of mechanisms, including decay-resistant wood and buttressing (Figure 32–7). But eventually, what goes up must come down, and tree fall is a main natural disturbance in rainforests. Tropical rainforests that lie within the reach of hurricanes or typhoons can suffer widespread canopy loss, but such catastrophes are not essential for tree fall. Individual tree falls are also agents of change. A single falling tree often brings down other trees around it, by direct impact or because the trees are connected by lianas. Light can then penetrate to the forest floor, and a strong pulse of growth by herbs, shrubs, seedling trees, and resprouted trees follows. If the gap is large enough, sun-requiring species, dispersed to this spot, can establish themselves before the gap closes. Rainforests Are Rapidly Being Destroyed Tropical rainforests have already lost half of their original area, and the loss continues. It is estimated that at current rates, only about 5 percent of the Earth’s area originally covered by tropical rainforests will survive by the middle of this century. Since 32–10 C h a p t e r 3 2 Global Ecology (a) (c) 32–7 Tropical rainforest (a) The interior of a rainforest in Costa Rica. The broad-leaved plants with red flowers are Heliconia irrasa. A female green anole (Norops biporcatus) is seen in the foreground. (b) The diversity of the trees in the forest, which may reach several hundred species per hectare, is revealed when individual trees burst into bloom, such as these trees in the coastal rainforest of Brazil. (c) A buttressed tree in the rainforest of Ecuador. Note the woody vines known as lianas on the trunk. (b) the tropics contain half of the world’s species, the loss could have dire consequences—certainly for global biodiversity. The greatest threats are exploitative logging and the subsequent conversion of forest to cropland and open pastures. Agricultural clearing makes landscapes susceptible to fire to a degree that is historically unprecedented, as was tragically demonstrated by the destructive fires of 1997–1998 in the Kalimantan region of Indonesia, where over 5 million hectares of forest and former forest burned. The conversion of tropical ecosystems to cropland would be more defensible if it always resulted in highly productive agricultural lands. But experience shows otherwise (Figure 32–8). Tropical soils often prove to be fragile. They can lose fertility rapidly when cleared, because a high proportion of their nutrient capital is tied up in the vegetation and proportionately less in the strongly weathered mineral portion of the soil. Organic matter, important for soil structure and retention of nutrients, is confined mostly to a shallow layer easily depleted during the clearing of trees. Addition of fertilizer may not help much, because tropical soils often fix phosphorus into insoluble forms. Deciduous Tropical Forests Only some parts of the tropics receive reliable year-round rainfall. In regions with a prolonged dry season, tropical rainforest gives way to other, more drought-tolerant forest types. These forests can be grouped under the general term of tropical seasonal forests. For example, in central and southern Africa, the Indian subcontinent, and Southeast Asia, there are extensive Savannas 3 2 – 11 32–8 Erosion Of the native vegetation that once covered Madagascar, an island in the Indian Ocean, 420 kilometers east of the African mainland, only 10 percent remains, and that is fast disappearing. On Madagascar, as in other tropical areas, the soils exposed by clearing vegetation are often not suitable for sustainable agriculture. The abundant rains rapidly wash the topsoil off increasingly barren landscapes, choking the rivers with silt, as seen here, and sometimes causing floods and landslides. The sea around Madagascar is often rust-red from the loss of the island’s weathered red soil, and it is difficult to produce enough food to sustain the people of the world’s fourth-poorest country. areas of monsoon forest, and similar climates and forests are found in northern Yucatán Peninsula in Mexico, in northern Colombia and Venezuela, in Ecuador, and in eastern Brazil (Figure 32–2). These climates produce enough rain to permit vigorous tree growth resulting in dense and productive forests, but the dominant trees are deciduous, losing their leaves during the dry season. Elsewhere in areas of the tropics with seasonal rainfall, there are other types of dry forests, such as the short-tree forests of tropical Mexico and Central America and the biologically distinct dry forests of Madagascar. Tropical mixed forests, in which both evergreen trees and shrubs are present, occur locally in eastern and southern Brazil and northern Australia (Figure 32–2). Although tropical rainforests tend to dominate public attention, the seasonally dry forests of the tropics also contain very high levels of biodiversity. Like the rainforests, dry forests are also being rapidly lost to exploitative land use or abused by unsustainable forest harvest. Savannas With further decreases in rainfall, a point is reached at which moisture is not sufficient to sustain a continuous tree canopy. Trees and shrubs are scattered individually or grow in groups in a grassland setting, and in places, woody plants nearly disappear. Such areas are called savannas, a term derived from the native languages of the Caribbean. The precise definition varies regionally. In South America, for example, savannas include tropical grasslands that are virtually free of trees, whereas in other places the term is restricted to grasslands with a significant woody component (Figure 32–2). Savannas of all kinds have annual rainfall in the range of 90 to 150 centimeters. In the tropics, savanna trees are broad-leaved deciduous and evergreen species, which may grow singly or in small groves. Other savanna types have a high proportion of shrubs (Figure 32–9). The cerrado vegetation of Brazil belongs to the savanna biome. It varies from open grassland to almost closed forest. In savannas, by definition, herbaceous plants, especially grasses, dominate. Bulbous plants can be abundant. Because of the dense cover of perennial herbs made possible by the relatively high seasonal rainfall, there are few annual herbs. Trees are well branched, but they are seldom more than 15 meters tall. Many trees, such as the acacias, are protected by stout spines, which discourage grazing (see Figure 31–9a). Leaves are generally smaller than those of the evergreen trees of the rainforest, and they are better able to regulate their water loss. The prolonged dry season, combined with the presence of highly flammable dry herbaceous growth, means that savannas are prone to burning. In general, fire is more destructive to woody plants than to herbaceous species. Unless protected by thick bark, a woody plant may lose all its investment in aboveground structures and will have to regrow years of accumulated biomass. If fires recur before recovery is complete, the tree may be killed or 32–12 C ha p t e r 3 2 Global Ecology 32–9 Savanna A savanna in East Africa, with giraffes surrounded by a herd of impala. The transitional nature of this biome, relative to the characteristic vegetation of the tropical rainforest biome and the desert biome, is evident in the grasses, shrubs, and short trees (acacias) seen here. reduced to a shrubby form, often arising from an enlarged base, or burl. But a herbaceous plant that is dormant at the time of a fire, and protected by having its live tissue below ground, will usually not suffer significant damage. Thus if fires occur regularly, they can gradually push back closed forests, opening the ground to full sun and encouraging the dominance of grasses and other herbaceous species. Current opinion is that natural fires must have been common enough to cause the evolution of fireresilient plants before burning by humans became so prevalent. This is not as certain in Africa, where fire-promoting hominoids have existed for periods long enough for evolution to act, but certainly applies in the Americas, where humans, in evolutionary terms, arrived very recently. There is little doubt that many savanna areas have been modified to some extent, possibly even created, by humans. At their poleward margins, tropical savannas and related plant communities grade into deserts, because of the low precipitation enforced by the global circulation (Figure 32–3). This transition is more or less continuous and occurs over a broad region in Africa, but it is more complex in the New World and in Asia. One such transitional community is the widespread juniper (Juniperus spp.) woodlands of western North America (Figure 32–10). These occur in places that are usually too dry to support grasslands but wetter and cooler than desert areas. In many places in the western United States, juniper, often found with pinyon pine, grows in areas adjacent to deserts but at higher elevations. Studies of pollen and macrofossils preserved in pack-rat middens show that vegetation types that seem to be permanent features of the landscape actually are dynamic over time. During the Pleistocene epoch, at the times of maximum expansion of the (a) (b) 32–10 Juniper woodlands (a) Juniper (Juniperus osteosperma) savanna in the Great Basin near Wellington, Utah. (b) Cold winters are characteristic of the pinyon-juniper savannas and woodlands of the Great Basin, as shown here south of Moab, Utah. Deserts 3 2 – 13 continental glaciers, juniper woodland communities moved down into the lowland areas that today support juniper-free deserts. Where temperatures are above freezing for much or all of the year, and where precipitation is so meager and unreliable that it can no longer support a continuous cover of perennial vegetation, we find the desert biome. All the great deserts of the world are located outside the equatorial zone, in the zones of atmospheric high pressure that flank the tropics at about 30° north and 30° south latitudes. They extend poleward in the interior of the large continents (Figure 32–2). Extensive deserts are located in North Africa and in the southern part of the African continent. Other deserts occur in the Near East, in southeast Mongolia and northern China, in western North America and western and southern South America, and in Australia. The Sahara, which extends all the way from the Atlantic coast of Africa to the Arabian peninsula, is the largest desert in the world. Seventy percent of the entire continent of Australia is covered in semiarid or arid areas. Less than 5 percent of North America is desert, and much of that is not extreme (Figure 32–11). (a) (b) (c) (d) Deserts 32–11 Desert Some representative plants of the principal deserts of North America. (a) The Sonoran Desert stretches from southern California to western Arizona and south into Mexico. A dominant plant is the giant saguaro cactus (Carnegiea gigantea), which is often as much as 15 meters high, with a wide-spreading network of shallow roots. Water is stored in a thickened stem, which expands, accordionlike, after a rainfall. (b) To the east of the Sonoran is the Chihuahuan Desert; one of its principal plants is the agave known as the “shin dagger” (Agave lechuguilla), a monocot. (c) North of the Sonoran is the Mojave Desert, with its characteristic plant, the Joshua tree (Yucca brevifolia). This plant was named by early Mormon colonists, who thought that its form resembled that of a bearded patriarch gesticulating in prayer. The Mojave contains Death Valley, the lowest point on the continent (90 meters below sea level), only 130 kilometers from Mount Whitney, with an elevation of more than 4000 meters. (d) The Mojave blends into the Great Basin Desert, a cold desert bounded by the Sierra Nevada to the west and the Rockies to the east. It is the largest and bleakest of the American deserts. The dominant plant is sagebrush (Artemisia tridentata), shown here with the snow-covered Sierra Nevada in the background. 32–14 C ha p t e r 3 2 Global Ecology The more severe deserts receive less than 20 centimeters of rainfall per year (see Figure 31–6). In the Atacama Desert of coastal Peru and northern Chile, the average rainfall is less than 2 centimeters per year. Averages, however, do not tell the whole story. As average rainfall decreases, the variability from year to year increases. In a humid region, a severe drought would be 50 percent less rain than average; in a desert, a drought is often no precipitation at all. The plants and animals of arid regions must be able to tolerate these periods of extreme dryness and to exploit the good years when they occur. This can favor some bizarre-appearing plant types, notable examples being Welwitschia (see Figure 18–41) found in the extreme coastal desert of Namibia in southern Africa and the cirio (Fouquieria columnaris) of Baja California (Figure 32–12). The desert regions of western North America illustrate the effect of larger-scale climatic variation. The hot deserts occur in the lower latitudes. The deserts of Arizona, which are of this type, are characterized by two peaks in rainfall, both variable with respect to exact timing and amounts. This pattern favors succulents that can store water from the infrequent and often scant rains. Temperatures can reach extremes, because the skies are 32–12 Surviving extreme dryness With leaves that emerge sporadically, depending on the presence of rain or coastal fog, the cirio (Fouquieria columnaris), also known as the boojum tree, is able to survive extensive periods of little or no moisture. “Boojum” comes from a nonsense poem by Lewis Carroll. usually clear, and little heat is lost to evaporation, because of dry soils, low plant cover, and limited transpiration. Summer temperatures of more than 36°C are common. The same clear-sky conditions that make deserts hot in the day cause them to cool rapidly at night. The heat stored during the day is lost through reradiation, and temperatures drop sharply as soon as the sun sets. As we move north from Arizona, the pattern in deserts shifts toward precipitation falling primarily in the winter, some of it in the form of snow. Such cold deserts are found, for example, in the Great Basin Desert of western North America, which lies between the Sierra Nevada–Cascade Mountain system and the Rocky Mountains (Figure 32–11d). In this region, winters can be bitterly cold. Succulent species are much less common, and small-leaved shrubs, such as the widespread sagebrush Artemisia tridentata, predominate. Moving further poleward, we reach areas of low precipitation in the Arctic and Antarctic regions, where the extreme cold combines with limited moisture to restrict plant life primarily to hardy lichens and mosses. Desert Plants Are Adapted to Low Precipitation and Extremes of Temperature High variability in moisture conditions means that desert plants must be able to either survive the dry season or dry periods in a dormant condition or have some way of averaging out variable moisture conditions. One solution is the annual-growth strategy, surviving the dry times as seeds that germinate when rains soak the surface, and flowering and setting a new crop of seeds before the window of moist conditions closes. Many species have been able to adopt this strategy, and there are proportionately more annual species in the deserts and semiarid regions of the world than in other biomes. For perennial plants, dormancy, water storage, access to deeper water, or a high level of resistance to drought stress are the only options. Grasses as a group have drought-tolerating features and are present at some level in all but the most extreme deserts. A typical perennial desert grass has a tussock growth form—a dense cluster of stems. With the onset of drought, the leaves die back, and the regenerative buds are protected in the mass of dead stems and leaves. Other herbaceous plants have deeply buried bulbs that break their dormancy when stimulated by rain. The amazing capacity of plants to adapt to desert conditions is exemplified by the grayish white wooly Arizona honeysweet (Tidestromia oblongifolia), a C4 (pages 140 to 145) perennial sub-shrub—that is, a plant with a shrubby growth form but with woody tissues only at the base of the plant—common in the Death Valley region of California and Nevada. The maximum photosynthetic rate for this species is achieved at temperatures between 45° and 50°C (around 120°F) in full sunlight in midsummer—temperatures that would be damaging or even fatal to plants of regions with higher moisture and cooler temperatures. Succulents are a distinctive class of water-storing desert perennials. The capacity to store water is found in many plant families, but two groups are notable: the cacti of the New World (family Cactaceae), essentially all of which are succulents, and the succulent euphorbs of the Old World (Euphorbiaceae). Members of these distantly related families provide some of the best examples of convergent evolution (see the essay “Convergent Grasslands 3 2 – 15 32–13 Creosote bushes One of the most characteristic plants of the Mojave, Sonoran, and Chihuahuan deserts of North America is the creosote bush (Larrea divaricata), which has small, leathery, water-conserving leaves. Creosote bushes in the Mojave Desert may form circular or elliptical clones, because of new stem production at the periphery of stem crowns and the segmentation and death of older stem segments, resulting in a ring of satellite bushes around a central bare area. The latter usually accumulates a mound of sand, which may reach a depth of about half a meter. Some clones may attain extreme ages: the King Clone, shown here, is estimated to be nearly 12,000 years old. Such ancient clones started from seeds that germinated near the end of the last glacial expansion. Evolution” on page 239). Succulents have shallow root systems that are designed to absorb water while it is available and then use it conservatively. To accommodate this water, succulents have a morphology that allows them to change volume without damage, a feature especially obvious in the accordionlike structure of many cacti. CAM (crassulacean acid metabolism—named for the Crassulaceae, a family with many succulent species; pages 145 to 147) photosynthesis provides another means of conserving the stored water. CAM plants reverse the usual pattern and open their stomata at night, then close them during the day, greatly reducing their loss of water (see the essay “How Does a Cactus Function?”). The success of the succulent life history is evidenced by the fact that succulents often are among the tallest and most robust plants of deserts—an example being the iconic saguaro cactus (Carnegiea gigantea) of the Sonoran Desert of the southwestern United States (Figure 32–11a). Another large group of perennial shrubs do not store water but have the ability to average out variation in rainfall by tapping into water reserves with their deep and extensive root systems. The success of this strategy is exemplified by the creosote bush (Larrea spp.), a genus of several species dominant over vast areas of New World deserts (Figure 32–13). Because deep water is accumulated by slow downward percolation, it varies much less than surface water over time and is available during dry seasons and even through several dry years. Tapping deep water would not work, however, if the plants were not able to use water efficiently. Creosote bushes produce small, leathery leaves with relatively few stomata and have stems resistant to the loss of water conductivity that is caused by embolisms (air pockets that block conducting elements). These characteristics enable close control of water loss and allow the plant to resist wilting when under extreme moisture stress. C4 photosynthesis is likewise more common among the plants of deserts and other seasonally dry, warm habitats than it is elsewhere. The plants also have the capacity to drop leaves and even entire branches in dry periods so that they can maintain their water balance. Then, when better conditions return, they can regenerate their canopies. Other woody plants, such as the various species of palo verde (e.g., Parkinsonia spp.), have green stems rich in chlorophyll and so are able to photosynthesize even when leafless. Grasslands Grasslands occur where the amount of rainfall is less than is needed to support vigorous tree growth but great enough to allow grasses and other herbaceous plants to produce a more or less uniform cover. They are found from the tropics to the edges of the boreal regions and from sea level to high mountain meadows. They grade into savanna (as noted above, grasslands in tropical areas are sometimes considered a type of savanna) and into desert. They occur as patches in woodland and forest when historical factors (such as fire) or soil factors (shallow dry soils) reduce tree growth. In the central and eastern United States, such grassy patches are often called glades, and they are typically dominated by prairie plants (Figure 32–14). Grasslands are most extensive, however, in the middle latitudes, where they coincide with the strongly seasonal climates of the temperate zones (Figure 32–2). Grasses regenerate at the beginning of each growing season from buds at or below ground level and have many fine roots designed to exploit the resources of the surface soil. Over long periods of time, grasslands, with the assistance of a complement of burrowing animals, tend to build soils with deep organic layers (see Figures 29–5, 29–6). Such soils are ideal for agriculture, and vast areas of grassland have been converted to cropland. In areas too dry for agriculture and without irrigation, grasslands have value as grazing lands. Although the growth of grassland plants is seasonal, there is little room for the development of annual herbs, and these are mostly limited to disturbed areas, such 32–16 C ha p t e r 3 2 Global Ecology 32–14 Glade Coreopsis dominating a glade in temperate deciduous forest near St. Louis, Missouri. Such openings in the forest usually occur in areas of shallow soil and are often dominated by prairie plants that form continuous stands beyond the borders of the forest. as prairie-dog towns or badger diggings or areas disturbed by humans, such as near buildings and along roadsides and railroads. The major variations in grasslands relate to precipitation. Grasslands receiving the highest precipitation are dominated by tall grasses in closely spaced clumps or dense sods. With decreasing precipitation, the stature of the plants decreases and the degree of coverage declines. In the driest grasslands the plants occur in widely spaced clumps, with sparse vegetation between them. The major gradients are exemplified in North America. Moving from west to east there is a transition from desertlike, western shortgrass prairie (the Great Plains), through intermediate midgrass prairie, to the moister and richer tallgrass prairie (now the Corn Belt) that gives way, in a complicated pattern, to the eastern temperate deciduous forest (Figure 32–15). Grasslands become progressively drier with increasing distance from the Atlantic Ocean and the Gulf of Mexico, which are the major sources of moisture-bearing winds in the eastern half of the North American continent. Farther north, grasslands become more moist again as evaporation decreases at relatively cool temperatures. Because most grasslands have more or less continuous cover and die back for part of the year, they carry fire readily. Where woody plants and grasses coexist, fires carried in grasses can “crown” (that is, spread fire within the tree canopies) or kill some or all of the meristematic tissues at the base of trees and shrubs. Though fires have always occurred naturally, it is thought that the purposeful burning of grasslands, for example, by Native Americans, may have increased the amount of grassland relative to forest. Those who manage natural grasslands today commonly use fire as an essential tool (see Figure 31–16b). Our emphasis here has been on plants, but all of the great grasslands of the world were once inhabited by herds of grazing mammals and associated large predators. Though there is no doubt that the grazers depend on an abundance of grass, there is debate about the extent to which grazing animals are necessary for maintaining the health of grassland vegetation. Some have argued that grasses have evolved to require pruning by grazers to (a) 32–15 Grasslands The grasslands of North America include large regions of shortgrass and tallgrass prairie. (a) A female bison nursing her calf in the shortgrass prairie of Custer State Park, South Dakota. (b) A June day on a tallgrass prairie in North Dakota. The cottonwood grove by the prairie creek is characteristic of this biome. (b) Grasslands 3 2 – 17 H o w D o e s a C act u s F u nct i o n ? The barrel cactus, Ferocactus acanthodes, which grows along the northwestern edge of the Sonoran Desert in southern California, gets its name from its barrel-like shape. It looks as if its stem has been folded like an accordion, and it is covered with spines like a porcupine. Why should this desert plant present such an unusual and formidable appearance? When the barrel cactus is full of water, the folds swell and are barely visible, but when the plant dries, the folds are deep and the stem can contract without crushing the cells. This ridge-and-valley folding of the stem has other advantages, too. Deep inside the valleys between the folds are the stomata. The spines help to break up the wind currents, and thus the valleys serve as protected retreats where dry desert winds cannot easily reach to carry moist air away from the vicinity of the stomatal chambers. These formidable spines also help to protect the cactus from the rodents and birds that are in constant search of water, even going so far as to steal it from a succulent stem. A study conducted by Park Nobel, of the University of California, Los Angeles, found that barrel cacti stored enough water in their succulent stems to permit them to open their stomata for about 40 days after the soil became too dry to furnish them with any additional water. Under such conditions, many of the fine roots are sloughed off, to prevent water loss to the soil. After seven months of drought, stomatal activity ceased, and the osmotic potential of the stem was more than double the value it had been during wet periods, despite the ability of the stem to fold and shrink. When rain finally returned, the shal- low root systems (with a mean depth of only about 8 centimeters) took in water so rapidly that the stomata were fully functional again within 24 hours of rainfall. But these are not the only mechanisms the barrel cactus uses for conserving water. Like many other desert plants, the barrel cactus exhibits crassulacean acid metabolism (CAM). It opens its stomata only at night and so undergoes gas exchange with cooler air, which can hold less water than warmer air. Consequently, the plant loses less water to the atmosphere through transpiration. Its ratio of mass of water transpired to mass of CO2 fixed is only about 70:1 for the entire year. This is considerably lower than for a typical C3 plant, which requires larger quantities of water to fix an equivalent amount of carbon but has a higher maximum rate of photosynthesis. Because seedlings of the barrel cactus, unlike their parents, cannot tolerate extremely high temperatures and prolonged drought, they survive only in certain years and in protected microhabitats. By the age of 26 years, these plants have usually grown to only about 34 centimeters tall, adding about 10 percent to the mass of their stems each year. Barrel cactus in a hydrated state (left), and in a dehydrated state (right). maintain a healthy photosynthesizing canopy. Whether or not this is correct, it is undeniable that grasses, grazers, and their predators have formed sustainable systems. Humans have disrupted natural systems, however. In the American West, the main natural grazer, the bison, was hunted to the brink of extinction and survives today only in refuges. The role of the bison in those grasslands that were not converted to row crops was taken over by domestic animals—mostly cattle and sheep. But domestic animals are not limited by wild predators, and since humans tend to maximize the size of their herds, overgrazing has been historically common. This is thought to account for the drastic depletion of the grasslands that formerly stretched from southern Arizona to western Texas. Over the course of the last century and half, these landscapes were converted to deserts or shrublands with sparse grass. The failure of farmers and ranchers to manage grasslands properly also contributed to the “dust bowl” disasters of the central United States in the 1930s (Figure 32–16). In extreme cases, the combination of grazers (mostly eating grass) and browsers (mostly eating woody vegetation) can turn lush areas into deserts of spiny and unpalatable plants. But responsible managers of grazing have learned from these past mistakes and have developed methods that make 32–18 C ha p t e r 3 2 Global Ecology 32–16 Dust bowl The prairie soils were once so bound together with the roots of grasses that they could not be cultivated until adequate plows were developed. But once the plants were removed by overgrazing or careless cultivation, prairie soils rapidly deteriorated and were blown away by the wind. This photograph of a badly eroded farmyard, taken in Oklahoma in 1937, vividly recalls the “dust bowl” conditions that led many thousands of people to migrate away from the central United States. John Steinbeck’s novel The Grapes of Wrath was based on the experiences of these migrants. sustainable grazing possible. With proper management, some lost grasslands, so it is hoped, can be restored. Temperate Deciduous Forests As we move poleward from the tropics, seasonality increases, and eventually, at about 35° latitude, we encounter climates experiencing long periods of temperatures below freezing. Even though the climates in these regions range between tropical heat and moisture in the summer and Arctic cold in the winter, it is traditional to describe them as “temperate” and the deciduous forests found there as temperate deciduous forests. These forests are best developed in areas with warm summers and cold winters (Figure 32–17). Annual precipitation generally ranges from about 75 to 250 centimeters and is either distributed evenly throughout the year or concentrated somewhat during the summer months. Temperate forests occur in both hemispheres, but because of the limited land area in the appropriate southern latitudinal bands, they occupy correspondingly small areas in the Southern Hemisphere (Figure 32–2). The tree form is favored in these regions, for the same reasons it is favored in the rainforests, though the kinds of trees and the structure and function of the forests are quite different. Leaf fall is strongly seasonal, with most of the trees remaining leafless through the winter. The deciduous trait can be explained by energetics. Relatively ample nutrients and moisture available (a) 32–17 Temperate deciduous forest Representative plants of the temperate deciduous forests of North America. (a) A beech and maple forest in Michigan, photographed in the spring. The forest floor is carpeted with large-flowered trillium (Trillium grandiflorum). (b) In the fall, as seen here in a forest in the southern Appalachian mountains, the leaves of maples and eastern sourwoods (Oxydendrum) turn a beautiful scarlet. (b) Temperate Deciduous Forests 3 2 – 19 32–18 Annual growth cycle The four seasons in a temperate deciduous forest in Illinois. The trees leaf out early in spring and begin to manufacture food; they lose their leaves in autumn and enter an essentially dormant state, in which they pass the unfavorable growing conditions of winter. Many herbs grow under the trees (see Figure 32–17), and some of them flower very early in spring, before the tree leaves reach full size and shade the forest floor. In spring, most of the trees produce abundant pollen, which is carried by the wind. throughout the summer assure high productivity, so that forming a new leaf produces larger gains and involves fewer risks than maintaining a more energetically expensive evergreen leaf through the rigors of the cold season. One of these risks is the lack of cold-season soil moisture because of frozen soil, a situation some describe as physiological drought. The annual cycle of growth is a defining feature of temperate deciduous forests (Figure 32–18). In winter, when the trees are leafless, their metabolic activity is greatly reduced. As temperatures warm, stored sugars are mobilized (the “sap rises”) and dormant buds begin to develop. It takes time for the new branches and leaves to expand fully, and this results in a short period when the forest floor receives substantial direct sunshine with temperatures above freezing. A variety of herbaceous plants, many with showy flowers, have evolved to exploit this situation (Figure 32–17a). Some (the spring ephemerals) have leaves that emerge seemingly overnight from bulbs or rhizomes. They complete most of their growth and reproduction before the trees are fully leafed out, then die back after the canopy closes. Others better adapted to reduced light (for example, early and late summer species and evergreen species) emerge more slowly and sustain photosynthetic activity during the shady conditions of summer. These species generally have leaves that are broader but thinner in cross section than those active only in spring, and they usually have smaller storage organs relative to their size. 32–20 C ha p t e r 3 2 Global Ecology Most of the species that ripen their seeds in spring are dispersed by ants (page 495), which are active when few other dispersers are present. In contrast, most of the fall-ripening species are dispersed by birds, during a season that coincides with the major bird migrations toward the equator. Very few annual plants occur in deciduous forests, and a smaller number of these behave as spring ephemerals. The cooler temperatures of these forests constrain the early growth that is crucial if an annual is to achieve reproductive size, a problem that spring ephemerals solve by having energy-producing stores (roots, bulbs, and rhizomes) below ground. A striking feature of the temperate deciduous forest is the sharing of genera—and, in some cases, even species—among three main regions in the Northern Hemisphere. Study of the fossil record shows that in the earlier Cenozoic era, these genera formed a band of deciduous forest across the Northern Hemisphere. Most of the deciduous trees and associated herbs were eliminated in western North America during the latter half of the Cenozoic, as the amount of summer rainfall was greatly reduced. This history explains why the herbaceous flora of the deciduous forests of China and Japan taxonomically resemble that of eastern North America more closely than either resembles that of western North America. But relics of the deciduous forests are still to be found in the western United States—for example, redbud (Cercis occidentalis) and big-leaf maple (Acer macrophyllum). Temperate Mixed and Coniferous Forests Although deciduous forests are the defining forest type in vast areas of the midlatitudes, there are also extensive areas with conifers, occasionally as the dominants, more often mixed with so-called broad-leaved species. We consider these as temperate mixed and coniferous forests. In keeping with the fact that, evolutionarily speaking, conifers predate the broad-leaved angiosperms, they tend to be most abundant where conditions are not the most favorable for tree growth—such as poorly drained sites, nutrient-deficient sites, and sites with shallow soil. Bordering the deciduous forests on the north (as noted above, the temperate forest is limited in the Southern Hemisphere) are forests in which conifers are prominent. Such temperate mixed forests (Figure 32–19) are characteristic of the Great Lakes–Saint Lawrence River region, eastern Europe, the northern and eastern border regions of Manchuria (in northeast China) and adjacent Siberia, eastern Korea, and northern Japan (Figure 32–2). They can be considered the intermediate condition between temperate deciduous forests to the south and the conifer-dominated taiga to the north. Temperate mixed forests occur in areas with colder winters and more reliable snow cover than are found in areas of temperate deciduous forest. The conifer element dominates on nutrient-deficient soils such as glacial outwash sands. There are also mixed forests on the southern edge of the temperate forest—the subtropical mixed forest (Figure 32–20). These forests occur in a complex mosaic of riparian (riverrelated) forest and swamp, with deciduous and broad-leaved evergreen forests on richer soils. Pines are especially prominent in these mixed forests, and in the southeastern United States, loblolly (Pinus taeda), slash (Pinus elliottii), and longleaf pines (Pinus palustris) are the common species. These pines 32–19 Temperate mixed forest In this forest in southern Ontario, the evergreen conifers appear dark green and the deciduous trees are showing their fall colors. are commercially important because of their fast growth and straight trunks. Although reduced soil fertility is a major explanation for the presence of conifer species, the cycle of disturbance also plays a role. 32–20 Subtropical mixed forest Slash pine (Pinus elliottii) is one of the widespread evergreen conifers growing in the subtropical mixed forests of the southeastern United States. Conifers tend to dominate on soils that are either nutrient-poor or seasonally flooded, or both. Longleaf pine has a distinctive life history related to fire. As a seedling, it exists in a grasslike form, with its long needles protecting its stem and growing point from fire. After some years, the seedling bolts, rapidly growing through the most vulnerable stages to a size that can survive fire. When fire is excluded from historical longleaf stands, they are invaded by broad-leaved species. Another fire-adapted species is the Florida sand pine (Pinus clausa), which grows in dense stands on nutrient-poor sandy soils. Unlike the longleaf pine, sand pine has serotinous (“late or delayed opening”) cones that are opened by the heat of fire. Because fire also kills most trees, the stands are reestablished by seed dispersed from the fire-opened cones. Cypress swamps are another conifer-dominated system characteristic of the subtropical mixed forest zone. Spanish moss– draped bald cypress (Taxodium distichum; “bald” because it is usually deciduous) is an iconic species of the swamps of the southeastern United States (Figure 32–21). It is most abundant in intermittently flooded sites, where it can reach ages in excess of 200 years and diameters of 2 meters or more. The current vegetation of western North America, the forested part of which has an abundance of conifer species, is an intricate mosaic complicated by the presence of mountain Mediterranean Scrub 3 2 – 21 32–22 Alpine tundra As seen here on the Olympic Peninsula in Washington State, alpine tundra is comparable in many respects to the Arctic tundra found hundreds of miles to the north. In this area, however, forested slopes are found within a hundred meters or so of the alpine meadows. ranges and their sequence of elevation-related changes in vegetation. Species that have expanded from the south formed, at lower elevations, grassland and shrubland communities along with savanna-like areas of evergreen and some deciduous oaks. Occurring at the highest elevations in these regions are the open herbaceous-dominated communities called alpine tundra (Figure 32–22), which in North America are intermixed with mountain forests. To the north, coniferous forests and mixed West-coast forests predominate, with such trees as the coast redwood (Sequoia sempervirens; Figure 32–23), the big tree or giant sequoia (Sequoiadendron giganteum), the Douglas fir (Pseudotsuga menziesii; Figure 32–24), and the sugar pine (Pinus lambertiana; Figure 31–15b), all of which were much more widely distributed in the past. Similar kinds of conifer-dominated vegetation are found in areas with similar climate characteristics in Scandinavia, central Europe, the Pyrenees, the Caucasus, the Urals, southern Tibet, and the Himalayas northward to eastern Siberia. Vegetation types that resemble these species are also found in areas in western South America, central New Guinea, southwestern New Zealand, the southern Arabian peninsula, Ethiopia, and the mountains of Central Africa (Figure 32–2). Mediterranean Scrub 32–21 Stabilized trees The bald cypress (Taxodium distichum) is a commercially valuable species that occurs in the low-lying, intermittently flooded swamps of the southeastern United States. The trees are often draped with Spanish moss (Tillandsia usneoides), which is not a moss but an epiphyte in the Bromeliaceae (pineapple family). The swollen bases and buttress roots stabilize the trees in the soft, usually highly organic substrate. Highly distinctive shrub communities have evolved from mixed deciduous-evergreen forests in areas with Mediterranean climates characterized by cool, moist winters and hot, dry summers. Such climates are found along the shores of the Mediterranean Sea, along the western edge of California and southern Oregon, in central Chile, on the southern coast of Africa, and along portions of the coast of southern and southwestern Australia (Figure 32–2). The plants in these areas have growing seasons concentrated in the cool, moist part of the year. The luxuriant growth of 32–22 C ha p t e r 3 2 Global Ecology 32–23 Mixed West-coast forest Redwoods (Sequoia sempervirens) 32–24 Pacific coast rainforest Douglas firs (Pseudotsuga are a prominent feature of the mixed west-coast forests of California. Watered by the frequent fogs of the region during the dry summers and protected from freezing temperatures by their proximity to the ocean, redwoods often form spectacular groves, many of which, like the one shown here in Muir Woods near San Francisco, are now protected in parks and reserves. menziesii) growing on the Olympic Peninsula in Washington State. Epiphytic mosses, liverworts, Selaginella, and lichens often grow luxuriantly on the trees. The forests of this area of North America experience heavy precipitation and are dominated by conifers. They are the basis for a profitable lumber industry. late winter and spring is followed by summer-fall drought, with some species becoming dormant or shedding most of their leaves, and others remaining evergreen and enduring the dry weather by drawing on deeper moisture. Though there are patches of woodland and grassland, vast areas are covered by shrublands with a continuous canopy, forming a kind of miniature forest. Shrublands of this kind are found in each of the widely dispersed Mediterranean climate regions and each has its own local name. In California and elsewhere in western North America, chaparral is the word for mostly evergreen and typically very dense shrublands. The equivalent vegetation around the Mediterranean Sea is called maquis (Figure 32–25), in Chile it is known as matorral, in South Africa fynbos, and in Australia mallee (kwongan in Western Australia). The quintessential growth form of the Mediterranean shrublands is the so-called sclerophyllous (“tough-leaved”) evergreen, deep-rooted species with stiff leaves that resist wilting. Another characteristic form is summer-drought deciduous, or semideciduous, with softer leaves, often with aromatic secondary compounds. Viewed from a distance, these different shrublands have a similar appearance. Even close up, species of different regions may appear superficially similar. But unlike the situation in the northern temperate forest, the similarities arise mostly because of convergent evolution. Each region has taxonomically unique species that have evolved toward morphological and physiological similarity. In all Mediterranean regions fire is a factor, though there is reason to believe that it is not uniformly so. Evidence of fire selection may be seen in the life histories. Most common—and not so clearly fire-selected—is vigorous resprouting after fire, often from an enlarged woody structure called a burl or lignotuber (“woody tuber”). Other species, the obligate seeders, are killed by fire but germinate from seeds that have accumulated in the soil between fires. Another strategy is to store seed in long-persisting, serotinous fruiting structures that release seeds in large numbers only after fire. Fire-stimulated seed release is hard to explain except for selection by fire, so the relative abundance of species with this strategy suggests that fire selection has been historically significant. Since Australia and South Africa have the largest number of such species, these regions also have probably experienced the most fire over evolutionary time. The California Mediterranean region, on the other hand, has many obligate seeding species that build up seed banks in the soil, with the serotinous trait found only in two genera of conifers (Cupressus and Pinus). The Northernmost Forests—Taiga and Boreal Forest 3 2 – 23 (a) (b) 32–25 Mediterranean-type vegetation Regions with Mediterranean-type climates are characterized by hot, dry summers and mild winters, with strongly seasonal rainfall that occurs primarily in the cooler part of the year. The vegetation is dominated by dense, low, mostly evergreen or summer-deciduous shrubs. There is a striking superficial similarity in the vegetation, whether in California, Chile, Australia, South Africa, or the Mediterranean region, as seen here for (a) chaparral in southern California and (b) similar vegetation, called maquis, on the Greek island of Corfu. Each region, however, has its unique set of species. The Mediterranean climate is one of the most pleasant for humans, especially near coasts, where cooling ocean breezes moderate the summer extremes. Before modern transportation and irrigation schemes, local sustainability had its limits, but today, largely because of the importation of water, huge numbers of people have migrated to the cities of these regions. This vast influx has had disastrous effects on the local plant life, mitigated only by the fact that steep topography, rocky soils, and a lack of access to imported water have put limits on agriculture and urban expansion. In less affluent areas, domestic grazing and browsing have reduced the native component of vegetation and increased the proportion of weedy species, many of them significantly less palatable because they are spiny or poisonous. Wildfire is a continuing problem in Mediterranean areas, the problem being proportional to the size of the human population, which increases the points of contact between people and fires. Also important is the state of the vegetation—that is, whether land use allows the shrub vegetation to approximate its naturally dense and continuous growth. Any wildfire is dangerous, but fires in dense shrublands are especially so. The heat generated by a wildfire can be fatal to humans, even if they are only near an area that is burning. In the windy conditions common in wildfires, houses can be ignited hundreds of meters from the flames when burning embers are lofted into suburban developments. Furthermore, wildfires that spread from a human ignition source (arson, campfires, fallen power lines, sparks thrown by mowing machines, and many more sources) can be more frequent than those arising from natural ignitions—mostly lightning and blowups from lightning-caused smoldering fires. As long as there is a natural cover of shrubs or grasses, fire cannot be eliminated from the landscape, but it is commonly asserted that the problem is made much worse by past management policies. The argument is that in attempting to banish fire, we have favored biomass accumulation and therefore larger and more damaging fires. There is some truth to this, but the simple solution of suspending fire suppression is not feasible. Restoring fire to its “natural role” requires first reducing current fire hazards and then implementing a scheme of regular burning or something that simulates burning. Both of these solutions are costly and require careful planning. The giant wildfires of recent years suggest that we have not even begun to achieve this hypothetical benign fire landscape. The Northernmost Forests— Taiga and Boreal Forest Moving poleward from the mixed coniferous forest, the seasonal contrast becomes more pronounced, with very short and extremely cold days in winter and a short growing season of long days. In these conditions, trees still dominate, because long days during the brief summer permit them to accumulate enough energy to overtop competing life forms. These northern coniferous forests are often referred to by the Russian name taiga; in North America they are also called the boreal, or northern, forest. Taiga extends over much of Russia, Scandinavia, and northern North America (Figure 32–2). Throughout its range it is characterized by a persistent cover of snow in the winter. In the southern reaches of the taiga, the trees are taller and more luxuriant, often reaching 75 meters or more in height. In its main northern area, however, the trees are shorter, and thousands of square kilometers are covered by this uniform forest, with relatively few species of plants and animals (Figure 32–26). Taiga occurs in the interior of large continental masses. In such regions, extreme temperatures range from –50° to 35°C. 32–24 C ha p t e r 3 2 Global Ecology 32–26 Northern taiga The northern taiga, which covers hundreds of thousands of square kilometers in the cooler part of the north temperate zone, is dominated by white spruce (Picea glauca) and larch (Larix), a deciduous conifer. This photograph was taken in northern Manitoba, Canada. In the more northern part of their range, the trees are smaller than seen here. Taiga is flanked on the south by montane forests (as in western North America), deciduous forests, savannas, or grassland, depending on the amount of precipitation in the region. Because continental masses do not occur at the appropriate latitudes in the Southern Hemisphere, taiga is absent there. Due to the influence of the prevailing westerlies blowing over relatively warm ocean currents between 40° and 50° north latitude, the western portions of North America and Eurasia are characterized by milder climates than their eastern portions. Consequently, taiga is found somewhat farther north toward the Pacific coast than it is along the Atlantic coast in North America, and the same is true of the individual distributions of many kinds of plants and animals that inhabit the biome. The northern limit of taiga is determined by the severity of the Arctic climate, and it roughly corresponds to where the maximum monthly temperature is approximately 10°C. At its northern limits, taiga grades unevenly into tundra. In the extensive northern reaches of the taiga, most of the precipitation falls in the summer; the cold winter air in these regions has a low moisture content. The annual total precipitation usually amounts to less than 30 centimeters. Despite the low rainfall, it is a region of moisture surplus because of low evapotranspiration. Lakes, bogs, and marshes are common (Figure 32–26). A major shift occurs with the appearance of permafrost. In less severely cold climates, the zone of freezing moves downward from the surface, reaching a maximum depth, and then thaws from both the bottom and the top as the weather warms. But in more than three-quarters of the northern reaches of the taiga, this summer thawing does not extend through the frozen zone before the onset of winter, thus creating a permanently frozen zone, or permafrost. Permafrost has many ecological consequences. Since water cannot move through the permafrost, and water will be released in thawing, saturated conditions and the presence of ponds are favored. Permafrost provides a stable base for soils that are often high in organic matter. When thawing eliminates permafrost or descends to deeper levels, the surface soils can be destabilized, causing trees to tip and even fall over in “drunken forests” (page 32–1). Houses and other buildings can also be damaged or destroyed when the permafrost thaws. In general, taiga soils are highly acidic and very low in nutrients. Species of a few genera of trees are common in the northern taiga, including spruce (Picea), larch (Larix), fir (Abies), and poplar (Populus). Among the more common shrubs are dewberries (Rubus), Labrador tea (Rhododendron), willows (Salix), birches (Betula), and alders (Alnus). Pines (Pinus) can be common, usually in drier areas. The members of all these genera of trees and shrubs are ectomycorrhizal (pages 312 to 314, Figure 29–1), and they grow in dense stands consisting of only one or a very few species. Perennial herbs are common, but annual plants, except in areas disturbed by humans, are essentially absent. Nonvascular plants—mosses and lichens— play an important role in the taiga and other far poleward communities. It is common in these forests to have dense and continuous layers of mosses and lichens, often many centimeters thick (Figure 32–27). These plants have both positive and negative influences on their neighboring vascular plants. In extreme cases, mosses, especially Sphagnum species, can convert forest into bog by trapping rainfall and creating thick, moist mats that discourage establishment of trees. Because of the seasonally abundant light and favorable temperatures, cool-season cultivated plants, such as cabbages (Brassica oleracea var. capitata), may grow rapidly in cleared areas in the taiga, attaining large sizes in a remarkably short time. Yet the infertile, highly leached soils of the taiga do not allow most forms of agriculture. Arctic Tundra 3 2 – 25 local melting of the permafrost. But as proof that generalities have notable exceptions, there is also a genus of deciduous conifer, the larches (Larix spp.), that is widely distributed throughout the boreal regions and often dominant. Its success appears to be due to a combination of the superior cold-tolerance of conifers with an improved ability, attributable to its deciduousness, to exploit more favorable nutrient conditions. Coniferous forests extend far to the south of taiga, along the Pacific coast of North America. These magnificent evergreen coniferous forests (Figure 32–2) occur where there is a pronounced summer drought, but high and persistent rainfall during the cooler seasons. Because photosynthesis is limited by lack of moisture during the warm season, deciduous trees are at a disadvantage and are usually found only along stream banks. The evergreen conifers, however, can synthesize carbohydrates all year round and, because of their massive size, can store water and nutrients for use during the dry season. Arctic Tundra 32–27 Understory vegetation Taiga, such as this forest in Lac La Ronge Provincial Park in Saskatchewan, frequently has understory vegetation dominated by continuous mats of mosses and lichens. These nonvascular plants affect many processes, especially plant regeneration, nutrient cycling, and depth of thaw of the permafrost. The abundance of evergreen conifers raises a question: why is it advantageous to be evergreen in a climate that has such a long cold season? It is presumed (and partly proven) that the shortness of the growing season and the generally low fertility of boreal soils make the deciduous habit less efficient. During the short season with a less predictable start to a frost-free summer, unfolding a new leaf from a dormant bud is less efficient than retaining leaves that can immediately resume photosynthesis. Further, the cool temperatures tend to slow down the decomposition and recycling of nutrients. As nutrients become more limiting, a leaf that functions over more than one season is capable of more photosynthesis per unit of nutrient absorbed. Broad-leaved deciduous trees do occur in the most northern forests, but they often are restricted to situations such as the margins of streams, where frost does not penetrate so deeply into the ground, or in the early nutrient-rich stages of succession after fire. Fires are common in the taiga, and they result in generally warmer, more productive sites for at least 10 to 20 years afterward, due to the The Arctic tundra is a treeless biome that extends to the farthest northern limits of plant growth (Figure 32–28). It occupies an enormous area: fully one-fifth of the Earth’s land surface (Figure 32–2). Many species of plants that grow in the Arctic tundra have wide circumpolar ranges. As the name suggests, Arctic tundra lies mostly above the Arctic Circle, extending farther south along the eastern sides of the continents than along their western sides. The Arctic tundra essentially constitutes one huge band across Eurasia and North America, with alpine tundra, more closely related to the adjacent mountain forests, extending southward in the mountains (Figure 32–22; see also Figure 32–5). Tundra vegetation exists in the Southern Hemisphere, but it is not well developed, given the limited land area available at the appropriate latitudes. Tundra-like vegetation also occurs at the highest elevations of mountains. In the tundra biome, the seasonal variation in climate is extreme. Although daylength increases in summer—north of the Arctic Circle, the sun does not set for days—this is balanced by a dark winter. This portion of the Earth operates at a strong solar energy deficit, radiating more energy back to space than it receives in direct sunlight. Frosts can occur throughout the year, and winters are bitterly cold, with high winds that will desiccate any plant that rises above the snow level. At low temperatures, blowing snow acts as an abrasive that ruthlessly attacks any exposed tissues. During the summer, the limited supply of solar energy is insufficient to thaw more than a shallow layer of soil above the permafrost. The annual freezing and thawing of the surface has many effects unknown in milder climates, such as landscapes broken into polygonal blocks ranging from 3 to 30 meters across and small ice-cored hills (pingos) that rise up to 50 meters high. The soils are acidic to neutral and low in nutrients. Even though precipitation is usually less than 25 centimeters per year, much of it is held near the surface by underlying permafrost, and on more level areas the ground is usually wet, with many small pools. Drier, usually rocky upland areas are also present. Because there are only a few species of legumes and other plants with symbiotic bacteria that fix atmospheric nitrogen, available nitrogen is often in short supply in the soil. 32–26 C ha p t e r 3 2 Global Ecology (a) (b) 32–28 Arctic tundra (a) Arctic wet coastal tundra near Prudhoe Bay, Alaska, in late summer. The reddishbrown plants are the grass Arctophila fulva, the green ones are the sedge Carex aquatilis. Note that the water table is above the surface because of the presence of permafrost; such conditions are characteristic of the Arctic tundra. (b) Arctic tundra at Barrow, Alaska. A clone of the sedge Eriophorum angustifolium is growing out into a solid patch of another species of the same genus, Eriophorum scheuchzeri. Vegetative reproduction, as in E. angustifolium illustrated here, is characteristic of many tundra plants. The vegetation over such a vast area has many variants, with the moisture conditions of the substrate an important factor. There are often distinct plant assemblages alternating on a small scale from pond to dry rocky hilltops. Another variable is the length of time that snow remains. Areas that receive less solar radiation and accumulate deeper snow will thaw later, so the growing season can differ significantly in the space of only a few meters. More exposed rocky areas, which tend to be blown free of snow in the winter, have a low cover of sedges, grasses, and dwarf, mostly evergreen shrubs. Both lichens and mosses are present in abundance, with more moss in wetter places and more lichens in drier spots, especially on the surfaces of exposed rocks and boulders. Several genera of low shrubs, including birch (Betula), willow (Salix), blueberry (Vaccinium), and Labrador tea (Rhododendron), are common. Shrubs such as willows often occupy lower positions, with their canopies at roughly the same height as the surrounding, more herbaceous vegetation. This is due to the action of the freeze-drying winds of winter. Annual species are rare, with the widely distributed Koenigia islandica an exception (Figure 32–29a). The high proportion of evergreen species in the tundra can be explained by the same factors that prevail in the taiga. Nutrients are limited, and the brief growing season places a premium on photosynthesizing as soon as possible and intermittently between cold periods. For similar reasons, vegetative propagation is common and establishment by seed relatively rare. Much of the biomass of tundra plants—from about 50 percent to as much as 98 percent—is underground, consisting of roots and underground stems. A number of tundra plants have relatively large, showy flowers, the production of which costs the plants considerable quantities of energy (Figure 32–29b). Such flowers hold energyrich rewards for their pollinators—necessary rewards, given the low temperatures that prevail at high latitudes. North of the Arctic tundra is ice desert, where physical conditions are so extreme that vegetation is essentially absent. Ice desert is characteristic of the interior of Greenland; of Svalbard, a small group of islands off Norway, one of which is the site of the Global Seed Vault (page 677); and of Novaya Zemlya, two islands off the north coast of Siberia. Much of Antarctica, not mapped in Figure 32–2, is also covered by ice or by Arctic deserts with patches of mosses and lichens. If projections of climatologists prove true, we can expect a considerable diminution of the ice cover of polar, and especially Arctic, regions. A Final Word What will be left of the natural biosphere in 2100? According to the United Nations, the population of the world will peak at 9 to 10 billion people in 2100. We are going to find out which bumper-sticker sentiment is closer to the truth: “More people means more hands to work and brains to think” or “More people means more mouths to feed, more pollution and land degradation.” For human welfare, it will matter a great deal which of these best describes the future. To a conservationist, it is not clear that there is a lot of difference between them. History tells us that when pressure from the human population increases, there is further exploitation of the natural world. If we are optimistic conservationists, we can hope for the first outcome. Perhaps one of the “extra” 3 billion people will be an agronomical genius who devises sustainable schemes to double the world’s agricultural output. Then it would be possible to hold the line on further destruction of natural habitats. But even if this worldwide transformation of agriculture could be achieved, it would still be true that 42 percent more people would be encroaching on wild habitats, building more houses, roads, and trails, and demanding Summary 3 2 – 27 (a) (b) 32–29 Tundra flowers (a) Koenigia islandica, also known as Iceland, or island, purslane, grows on damp rocky areas, especially near persistent patches of snow, in the tundra and alpine meadows of the Northern and Southern Hemispheres. (b) Mountain avens (Dryas octopetala), a large-flowered Arctic plant of wide distribution, is seen here growing in Siberia. Its pollen has been used as a marker, or indicator, of colder climates, and its name has been adopted to describe periods of cooling during glacial times, as in “the younger and older Dryas periods.” the control of dangerous or annoying wild animals. If the pessimists are correct, and more people means more threats to the biosphere, the future for conservation is even bleaker. If we increase agricultural productivity 10 percent instead of 100 percent, and simultaneously need to produce biomass energy, much more land will be needed. And this additional land will come primarily from places where human populations are low—that is, from whatever remains of our natural landscapes. If the more dire predictions of climate scientists prove true, there will be further disruptions and instabilities. What is the appropriate response of a conservationist to this possibility of a bleak future? Given the intense pressure from the disdainers of wild nature, it is clear that to save as much of the natural part of the biosphere as possible, we will need continuing counter-efforts across a broad front. There are utilitarian as well as aesthetic and moral reasons for not destroying nature. We need all of these arguments—and articulate persons to make them. But as believers in science (which we expect you are, too), we recognize the critical importance of a scientific understanding of how the world ecosystems work. We hope this brief overview of Earth’s natural ecosystems will give you a foundation on which to build and will motivate you to do your part in saving the natural world. SUmmary Vegetation Differs from Place to Place and from Time to Time, but Large-Scale Patterns Are Discernible On a small scale, vegetation is highly variable because of patterns of past disturbance and differences in the substrate. But when viewed on a larger scale, vegetation separated by many thousands of kilometers can appear very similar. This similarity can arise because the species are closely related or because similar climates and substrates in different areas favor convergent evolution, or both. Biomes Are Terrestrial Ecosystems Characterized by Distinctive Vegetation Biomes are more or less homogeneous regional or global ecosystems characterized by plant species that share physical and physiological traits. The similarity in species traits arises primarily because of the similarity in climate. Because they cover broad areas, biomes also contain local variation, areas that differ from the defining communities. So, for example, the temperate deciduous forest biome includes extensive herbaceous marshes. Tropical Rainforests Have a Great Diversity of Species Tropical rainforests, with their consistently warm temperatures and ample moisture, are the richest biomes in terms of number of species. The trees are predominantly evergreen and broad-leaved. A poorly developed layer of herbaceous plants grows on the forest floor, but there are many vines and epiphytes at higher levels. Tropical soils are often acidic and very poor in nutrients; such soils lose their fertility rapidly when the forest is cleared. Savannas and Deciduous Tropical Forests Occur Where Rainfall Is Seasonal Mostly tropical and subtropical communities that are characterized by a seasonal drought are termed savannas, subtropical mixed forests, monsoon forests, tropical mixed forests, and southern woodland and scrub. The trees and shrubs of these communities are wholly or partly deciduous, shedding their leaves during times of drought. Herbaceous perennials are common. Savannas also occur between the prairies and temperate deciduous forests and between the prairies and the taiga in North America. Subtropical mixed forests cover most of Florida and the coastal plain of the southeastern United States. In these forests, pines and other evergreen trees grow intermixed with deciduous trees. 32–28 C ha p t e r 3 2 Global Ecology S u mmar y T abl e Some Characteristics of the Earth’s Principal Biomes B IO M E T E M PE R AT U R E A N D P R E C IPI TAT IO N Rainforests High temperature and high rainfall year round CHARACTERISTIC PLANTS M I S C E L L A N EOU S F E AT U R E S Broad-leaved evergreen trees, The biome with the greatest diversity of species; epiphytes, and lianas infertile soils Savannas and High temperature and seasonal drought Grasslands with scattered broad-leavedPeriodic burning is an important aspect deciduous tropical deciduous or evergreen trees or shrubs forests DesertsPrecipitation generally very low except for occasional peaks; maximum temperature varies with the type of desert In warmer, slightly wetter deserts, succulents such as cacti; in all deserts, deep-rooted drought-resisting shrubs and annual plants that flourish after rare rains Grasslands Moderately low precipitation, cold winters, and Perennial bunchgrasses and sod- warm summers forming grasses Temperate deciduous forests Moderate precipitation evenly distributed, cool winters, and warm summers Adaptations include small leaves, thick cuticles, and photosynthetic rates with high midsummer temperatures; spacing of dominant perennial plants reflects competition for underground water more than for light Heavily exploited for agriculture or for grazing of domestic animals Deciduous trees and many perennialThe dominant herbaceous plants vary with the herbs seasons Temperate mixed Moderately low precipitation and moderately Mixtures of deciduous trees and and coniferous cold winters conifers forests Occur as a transition zone north of the deciduous forest; also found in areas with nutrient-poor soils or with less seasonal environments Mediterranean Cool, moist winters and hot, dry summers scrub Evergreen or summer-deciduous drought-resistant trees and shrubs in dense thickets Called chaparral in California and maquis around the Mediterranean Sea Taiga Moderately low precipitation and cold winters Forests mostly of conifers, mostly evergreen but with a significant component of deciduous Larix species in some places Soils are highly acidic and very low in nutrients; permafrost may be present Arctic tundra Low shrubs, grasses, sedges, andPermafrost present throughout; much of the lichens biomass is underground Very low precipitation in both summer and winter, very cold winters Desert Plants Are Adapted to Low Precipitation and Extremes of Temperature Away from the equator, tropical and subtropical communities grade into deserts and semideserts, which are characterized by low precipitation and often by high daytime temperatures during at least part of the year. Succulent plants are common in the warmer and less extreme deserts, and drought-tolerant smallleaved shrubs are found in most deserts. Grasslands Occur Where Precipitation and Fire Interact to Support Grasses but Not Trees Grasslands, which intergrade with savannas, deserts, and temperate forests, are characterized by a general lack of trees, except along streams. The height and density of the grass-dominated cover varies from scattered tussocks in desert grasslands to shortgrass turfs, to mid- and tallgrass prairies, where rainfall is greatest. Grasslands favor the accumulation of organic matter, often to considerable depths. The most productive soils for temperate agriculture are grassland soils. Temperate Deciduous Forests Are Made up of LeafShedding Trees and Many Types of Perennial Herbs In the temperate deciduous forests, most of the trees lose their leaves during the cold (usually snowy) winters, when moisture may be unavailable for growth. Many genera are common to the temperate deciduous forests of eastern North America, northern Europe, and eastern Asia. Temperate deciduous forests are bordered by temperate mixed and coniferous forests to the north, in which conifers play an important role. Mediterranean Climate Scrub Is Characterized by Evergreen, Drought-Resistant Shrubs or Trees That Form Thickets Distinctive shrub communities, called chaparral in North America and maquis in the Mediterranean region, have evolved in the five widely separated areas of the world with a Mediterranean climate—a dry summer and a cool, rainy winter growing season. Such communities occur in western North and South America, around the Mediterranean, in the Cape Region of South Africa, and in southwestern Australia. In all regions they are subject to more or less frequent intense wildfires. The Taiga Is Characterized by Forests of Coniferous Trees The taiga is a vast northern coniferous forest that extends in unbroken bands across Eurasia and North America. In its southern reaches, the taiga is dominated by tall trees with a lush growth of mosses and lichens; northward, it consists of vast monotonous stretches of forest with very few tree species. The Arctic Tundra Has Low-Lying Shrubs and Grasses but No Trees Tundra occurs poleward of the evergreen taiga-type forests, where the intense winter cold and short growing season preclude tree growth. It extends around the Northern Hemisphere, mostly PASS 1 Photo Credits 3 2 – 29 above the Arctic Circle, in a band that is broken only by bodies of water. Both the northern reaches of the taiga and all of the tundra are underlain by permafrost. Because of this permafrost, and especially because of the low rates of evapotranspiration, tundra and taiga soils are relatively moist and highly leached of nutrients. The Earth’s Biodiversity Is under Siege, and Efforts Must Be Made to Protect It The growing human population continues to diminish our global stock of natural areas. If we are to save enough of it to prevent massive extinctions, we must redouble our efforts at conservation and restoration. QUESTIONS 1.Describe the influence of latitude and altitude on the distribution of organisms on Earth. 2.Describe the effect of mountains on local precipitation. 3.Compare tropical and temperate forests in terms of the numbers of species found in each, and in the size and appearance of the trees. 4.Explain why annual plants are better represented, both in number and in kind, in the deserts and semiarid regions of the world than anywhere else. 5.Compare the relative amounts of nutrients found in forest and grassland soils. 6.How are the evergreen conifers of the Pacific Northwest of the United States and Canada adapted to the winter-wet, summer-dry environment of that region? 7. What are the principal differences between taiga and tundra? What role does permafrost play in these biomes? 8. The concept of “ecological services”—benefits that humans derive from the existence of natural systems—has become a guiding idea in recent decades. What are these benefits, and how are they likely to change as the effects of human interference continue to increase? 9.Using Google Earth, find the latitude of your current location. Follow this latitude around the globe, zooming in at enough points to get an impression of how the vegetation changes. What causes the changes you observe? What proportion of the surface shows clear evidence of human activity? How much looks more or less natural? PHOTO CREDITS Chapter Opener Ashley Cooper, Visuals Unlimited/Science Photo Library; 32.1a Wikipedia Commons; 32.1b Martin Zeick, Age Fotostock; 32.2 After A. W. Küchler; page 32-8 Institut für Auslandbeziehungen, Stuttgart, Germany; 32.7a Michael Fogden/ DRK Photo; 32.7b Martin Wendler/NHPA; 32.7c E. S. Ross; 32.8 Frans Lanting/Minden Pictures; 32.9 Peter Ward/Bruce Coleman, Inc.; 32.10a, b J. Reveal; 32.11a Martin Wendler/NHPA; 32.11b Creative Commons, photo by Stan Shebs; 32.11c Ric Ergenbright; 32.11d Greg Vaughn/Tom Stack and Associates; 32.12 Corbis RF/ Alamy; 32.13 F. C. Vasek; 32.14 Ray Evert; page 32-17 (both) Ronald F. Thomas/Bruce Coleman, Inc.; 32.15a Jeff Foott; 32.15b Pat Caulfield; 32.16 Soil Conservation Service; 32.17a Rod Planck/ Tom Stack and Associates; 32.17b P. White; 32.18 (all) J. H. Gerard; 32.19 R. Burda/Taurus Photos, Inc.; 32.20 J. Dermid; 32.21 Tom Salyer/Alamy; 32.22 E. Beals; 32.23 E. S. Ross; 32.24 David Woodfall/WWI/Peter Arnold, Inc.; 32.25a Jack Wilburn/Earth Scenes; 32.25b Ardea Photographics; 32.26 J. Bartlett and D. Bartlett/Bruce Coleman, Inc.; 32.27 Ron Erwin Photography; 32.28a, b W. D. Bellings; 32.29a Bob Gibbons/Science Photo Library; 32.29b Serg Zastavkin, Shutterstock Images