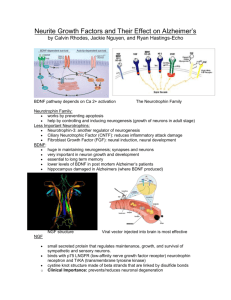
REGULATION AND FUNCTION OF ADULT NEUROGENESIS: FROM GENES TO COGNITION
advertisement

Physiol Rev 94: 991–1026, 2014 doi:10.1152/physrev.00004.2014 REGULATION AND FUNCTION OF ADULT NEUROGENESIS: FROM GENES TO COGNITION James B. Aimone, Yan Li, Star W. Lee, Gregory D. Clemenson, Wei Deng, and Fred H. Gage Cognitive Modeling Group, Sandia National Laboratories, Albuquerque, New Mexico; and Laboratory of Genetics, Salk Institute for Biological Studies, La Jolla, California Aimone JB, Li Y, Lee SW, Clemenson GD, Deng W, Gage FH. Regulation and Function of Adult Neurogenesis: From Genes to Cognition. Physiol Rev 94: 991–1026, 2014; doi:10.1152/physrev.00004.2014.—Adult neurogenesis in the hippocampus is a notable process due not only to its uniqueness and potential impact on cognition but also to its localized vertical integration of different scales of neuroscience, ranging from molecular and cellular biology to behavior. This review summarizes the recent research regarding the process of adult neurogenesis from these different perspectives, with particular emphasis on the differentiation and development of new neurons, the regulation of the process by extrinsic and intrinsic factors, and their ultimate function in the hippocampus circuit. Arising from a local neural stem cell population, new neurons progress through several stages of maturation, ultimately integrating into the adult dentate gyrus network. The increased appreciation of the full neurogenesis process, from genes and cells to behavior and cognition, makes neurogenesis both a unique case study for how scales in neuroscience can link together and suggests neurogenesis as a potential target for therapeutic intervention for a number of disorders. L I. II. III. IV. V. VI. INTRODUCTION NSCS AND THE NEUROGENIC NICHE DEVELOPMENT AND MATURATION... REGULATION OF NEUROGENESIS FUNCTION OF NEUROGENESIS TECHNOLOGY FOR STUDYING THE... 991 993 997 999 1009 1015 I. INTRODUCTION The neurogenesis field has made remarkable progress in the past two decades (82, 111, 124, 370). Less than 20 years ago, the neuroscience community’s appreciation of adult neurogenesis was quite limited. Despite having been characterized using a range of techniques, the fact that young neurons continue to be incorporated into the adult brain was not widely accepted until the mid 1990s (128, 150). Several factors precipitated this shift. First, immunohistological techniques for labeling dividing cells with nucleotide analogs [e.g., bromodeoxyuridine (BrdU)] and protein markers that were specific to neurons (e.g., NeuN), coupled with confocal imaging, allowed the confident identification of adult-born neurons. Second, these tracing techniques revealed that the incorporation of young neurons did not simply occur at a persistent low rate of residual development; rather, it was heavily regulated by behavioral factors such as stress, age, exercise, and enrichment. Finally, the positive identification of adult-born neurons in the human hippocampus, and even in aged individuals, demonstrated that this phenomenon was potentially relevant to human cognition (95). Neurogenesis has been described in several brain regions of multiple species. It was identified early on in songbirds (253) and has since been observed in other bird species, reptiles, and fish (372). In mammals, neurogenesis appears to be considerably more limited, with robust levels limited to the dentate gyrus (DG) region of the hippocampus and the olfactory bulb (OB). New neurons have also been reported in other areas, with the neocortex (52, 120, 126) and hypothalamus (168) attracting the most attention, although the extent of neurogenesis in these areas remains controversial (273). It is worth noting that the existence of OB neurogenesis [in which maturing neurons migrate from a stem cell population in the subventricular zone (SVZ)] in humans is itself controversial, with differing reports having been presented in recent years (35, 75, 288). Human neurogenesis in the DG is generally accepted, although the levels of new neuron production have only been characterized in limited cases (95, 315). Both a microcosm of neural development and a never-before-appreciated form of neural circuit plasticity, neurogenesis has recently attracted investigations from perspectives ranging from molecular neuroscience studies of the genetic regulation of young neurons to studies of the computational and behavioral implications of neurogenesis at circuit scales. In this review, we focus primarily on the process of DG neurogenesis (FIGURE 1), summarizing the full process from the origins of new neurons as neural stem cells (NSCs) residing in the subgranular zone (SGZ) through their maturation into fully functional granule cells (GCs). We describe the regulation of the neurogenesis process, which is 0031-9333/14 Copyright © 2014 the American Physiological Society 991 AIMONE ET AL. >4 wk 3 wk 2 wk 1 wk Basket cell type 2 3 days type 1 Astrocyte to CA3 Proliferation Differentiation Survival FIGURE 1. Illustration of the development of dentate gyrus granule cells from stem cells to fully mature neurons. New neurons arise from two populations of primitive cells, the slowly dividing type 1 cells, also known as radial glial cells, and the more rapidly amplifying type 2 neural progenitor cells. Over the next few weeks, cells differentiate into neurons, slowly developing dendritic arborizations and axonal projections. Between 2 and 3 wk of age, new neurons begin to receive excitatory input from cortical perforant path axons, and by 4 – 8 wk, their physiology and anatomy begin to approach those of fully mature neurons. influenced by molecular, network, and behavioral sources. We then review the current view of the role of neurogenesis in the DG’s function in learning and memory, ending with a description of current and future techniques for studying new neurons. A. Characterization of Neurogenesis It is important to briefly summarize the methods for tracing new neurons, as the techniques used to label a dividing and maturing cell often influence the nature and interpretation of the experiments described below. The most common method for labeling dividing cells involves the incorporation of a traceable molecule into DNA. Because DNA synthesis is generally limited to mitosis, at least at measurable levels, it has been used as a marker of neurogenesis. The first 992 neurogenesis studies utilized tritiated (3H) thymidine (10), permitting the radiographic tracing of cells that had been born at the time of injection. In the 1990s, another thymidine analog, BrdU, was developed because this molecule could be detected using immunohistology. This development was a vital advance since immunolabeling could permit fate identification of dividing cells through the colabeling of other markers, such as the neuronal marker NeuN or the glial marker glial fibrillary acidic protein (GFAP). BrdU labeling remains widely used, as are its sister molecules IdU and CldU (for its iodide and chloride equivalents, respectively), in large part because it can measure both proliferation rates and survival. It is worth noting that BrdU, despite its usefulness, is not an ideal marker for several reasons, including potential toxicity, nonspecificity (damaged cells may incorporate it), and histological limitations (172). Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS Other immunohistological markers can be used to identify proliferating cells; for neurogenesis work, the most widely used is Ki67 (245). Similarly, there are several markers for specific stem cell phases, such as Sox-2 and nestin, which will be briefly described below. It is important to note that, while these protein markers are specific and do not require preceding injections, they are also transient and not particularly suitable for long-term labeling or quantification. Genetic tracing studies, such as cre-lox systems in which a transiently activated promoter permanently labels a neuron with a marker such as lacZ or green fluorescent protein (GFP), are increasingly used for long-term quantification (140, 250). Finally, the most common approach is retroviral labeling of dividing cells with a marker such as GFP. Retroviral labeling is fairly sparse, which limits quantification but can provide highly accurate birthdating, can show full morphologies of new neurons (371), and is suitable for imaging in live slices for electrophysiology studies (343). Several of these methods will be described at greater length later in the text. B. Neurogenesis in Humans For obvious reasons, measuring neurogenesis in humans is considerably more difficult. While human neurogenesis was originally confirmed using BrdU (95), the sample sizes were too low for thorough quantification, and subsequent studies using histological markers left some doubt regarding the overall levels of neurogenesis in humans (299, 300). Recently, Frisen and colleagues (315) described a technique whereby rates of new neuron birth in humans could be estimated by taking advantage of increased radioactive carbon in the atmosphere due to above-ground nuclear testing (315). For roughly 15 years, levels of 14C increased steadily above historic levels; thereafter, levels decayed steadily due to international agreements to limit above-ground testing. Upon entering food supplies, 14C incorporates into DNA in a many similar to that seen in the original [3H]thymidine studies. This technique enables estimates of overall levels of neurogenesis, showing a turnover rate of GCs at ⬃1.75% a year. Notably, subsequent studies with this technique did not observe new neuron turnover in the OB; however, it may be possible that new neuron incorporation rates there were not well suited for this technique. Interestingly, Frisen and colleagues (96) did observe new neurons in the human striatum, which has not been widely observed in rodents but has in other mammals such as rabbits (44, 206). II. NSCS AND THE NEUROGENIC NICHE The ability of NSCs to self-renew, proliferate, and differentiate into all cell types of the nervous system makes understanding these cells and the factors that regulate this process critical to developing potential therapies for a number of neurodegenerative diseases. In the adult brain, neurogenesis is well documented to continue throughout life in only two regions: the SVZ of the lateral ventricles and the SGZ of the DG. Although additional areas have been reported to support adult neurogenesis, we do not focus on those studies in this review. In both regions, NSCs give rise to neural progenitor cells (NPCs), which are limited in proliferation and differentiate into neurons or glia (111). NPCs from the SVZ migrate along the rostral migratory stream and supply newborn neurons for the OB; those in the SGZ migrate a short distance into the GC layer of the DG and integrate into the existing circuitry of the hippocampus. There has been ongoing debate regarding which cell population within the SGZ is the “true” NSC population, if such a thing can be appropriately defined (41). Considerable evidence exists from experiments in mice leveraging promoters from genes such as Gfap, Nestin, Sox2, and Mash to perform lineage analysis (93, 314, 321). These promoters each appear to label slightly different, though overlapping, populations of cells, including both radial glia cells (RGCs), often referred to as type 1 cells, and nonradial type 2 cells. Several studies have demonstrated that RGCs can give rise to type 2 cells through asymmetric division; however, it is debated whether RGCs can replicate through symmetric division (41). It is also not known whether type 2 cells can revert to an RGC state. However, it is known that from type 2, cells generate what appears to be a more fate-committed intermediate progenitor cell (IPC) population, sometimes referred to type 3 cells, which has a distinct expression profile, including transcription factors such as Tbr2, and appears constrained to a future neuronal fate (137). It is unclear when and at what rates astrocytes arise from this process as well, with competing hypotheses existing about whether glia are an end product of neuronal differentiation (93) or a separation differentiation path for multipotent cells (42), although it appears that the gliogenesis process may be equivalently complex (46). Notably, it does not appear that oligodendrocytes are a product of this process in vivo (42), although they may arise from a separate pool of progenitors (149). Since which neurons in the above process are indeed selfrenewing and multipotent is still contested, in this review we broadly define NSCs as those cells that are capable of self-renewal, albeit slowly, and broadly multipotent. Likewise, we refer to NPCs as those cells that proliferate quickly and remain capable of becoming glia and neurons but are limited in the number of progeny they can produce. In addition, with regard to neurogenesis and its regulation, it is often useful to consider three higher level processes: cell proliferation, neuronal differentiation, and cell survival (FIGURE 1). Each aspect is critical to the overall levels of neurogenesis. For example, NPC proliferation occurs in other regions of the adult brain, but the cells do not differentiate into neurons, becoming glia instead. However, NPCs isolated from these nonneurogenic regions, such as Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 993 AIMONE ET AL. cortex and optic nerve, in the adult brain, retain the potential to become neurons in vitro following treatment with fibroblast growth factor fibroblast growth factor-2 (FGF2), indicating that extrinsic factors play a major role in stimulating NPCs to differentiate into neurons (262). Further supporting the idea of a neurogenic microenvironment is the finding that the adoption of neuronal cell fates by NPCs under normal physiological conditions is limited in the adult brain to a few regions such as the SVZ and SGZ, but transplantation of NPCs from the SVZ into ectopic regions of the adult brain results in gliogenesis, in which NPCs become oligodendrocytes and astrocytes (298). In contrast, NPCs from a nonneurogenic region, such as the spinal cord, can differentiate into neurons when transplanted into the DG, supporting the idea that external cues from the local microenvironment promote neuronal differentiation of NPCs (306). The SVZ and SGZ represent neurogenic niches or local microenvironments that permit and support neurogenesis. Studies have identified key components and factors of the neurogenic niche in a number of ways, including in vitro coculture experiments and in vivo manipulations of neurogenesis. While there are a number of underlying similarities between the two environments, in this review we focus on the critical elements relevant to the hippocampal neurogenic niche. A. Microglia The neurogenic niche supports and promotes neurogenesis through both secreted factors and cell-cell contact. Microglia are the resident macrophages and primary immune cells of the brain, and they have a multitude of functions, ranging from phagocytosis to neuroprotection. In the adult brain, microglia in the resting state continuously survey the local environment, with their dynamic processes interacting with a number of cell types, including astrocytes, neurons, and endothelial cells (249). In the adult DG, microglia are found distributed evenly in the hilus and along the border of the GC layer (360). In the adult hippocampus, resting microglia are associated with GC death (277); more specifically, interactions between microglia and NPCs regulate neurogenesis via phagocytosis (310). Most newborn cells in the adult DG fail to survive past the first week (159, 310); eventually, the number of surviving newborn cells stabilizes after 3– 4 wk (159, 310, 326). The decline in newborn cells within the first week is largely due to apoptotic cell death, which is mediated by microglia, as their processes are localized and engulf apoptotic BrdU-positive cells (310). These studies investigating the behavior of resting and unchallenged microglia demonstrate the active role these cells play in monitoring and maintaining the local environment of an unperturbed adult brain. The contribution of microglia to the neurogenic niche appears to depend largely on their secretion of cytokines and 994 chemokines. Additionally, it is the balance of proinflammatory and anti-inflammatory signaling that is critical to whether or not the environment supports or prohibits neurogenesis (28, 49, 61). Classically activated microglia secrete the cardinal proinflammatory cytokines, including tumor necrosis factor-␣ (TNF-␣) and interleukin-6 (IL-6), that can inhibit NSCs from differentiating into neurons in favor of astrocytes (61). However, depending on the environment, individual cytokines can exhibit proneurogenic or antineurogenic properties. For example, transforming growth factor- (TGF-) is a cytokine secreted by microglia that is known to activate both proinflammatory and antiinflammatory pathways, and its expression has been shown to both induce (28) and inhibit neurogenesis (48), illustrating its ability to positively and negatively regulate this process. Depending on the mode of activation, activated microglia can release proinflammatory or anti-inflammatory cytokines, modulating the immune response supporting or suppressing neurogenesis (49, 233). In in vitro studies, microglia are most commonly activated with lipopolysaccharide (LPS). LPS is an endotoxin from the gram-negative bacterial cell wall that activates microglia through Toll-like receptor-4 to induce both morphological changes and release of cytokines and nitric oxide (37, 243). Coculturing LPS-activated microglia or conditioned media (CM) with NPCs decreases neuronal differentiation of NPCs (49, 233). In contrast, both IL-4- and interferon (IFN)-␥-stimulated microglia induce neuronal differentiation of NPCs, illustrating the potential proneurogenic effects of activated microglia (49). Interestingly, it is not only the mode of activation but also a temporal component that determines whether activated microglia induce or inhibit neurogenesis. For instance, coculturing acutely activated microglia with LPS (after 24 h of LPS treatment) or CM with NPCs decreases neuronal differentiation and increases glial differentiation (50, 233). NPCs also exhibit increased TUNEL staining when cocultured with CM from LPS-activated microglia, illustrating the potent effects of acutely activated microglia on NPC activity (233). However, CM from chronically activated microglia with LPS (after 72 h of LPS treatment) does not significantly decrease neuronal differentiation of NPCs (50), likely due to a difference in secreted cytokine profiles from acutely and chronically activated microglia. While acutely activated microglia release more proinflammatory cytokines, including IL-1␣ and IL-6, chronically activated microglia release more anti-inflammatory proteins, like IL-10 and PGE2 (50). Further studies have evaluated the activity of microglia by manipulating levels of neurogenesis. Two of the most common ways to increase neurogenesis are through running and exposure to an enriched environment (EE), which appear to Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS increase neurogenesis via different mechanisms enhancing cell proliferation and cell survival, respectively. The effect of running on cell proliferation is robust, as depicted in the increase in the numbers of BrdU cells in the DG with running (257, 350). However, exercise does not result in an increase in the overall number of microglia in the DG or increase the percentage of activated microglia, illustrating that the overall microglia profile remains unaffected by running (257). Additionally, running does not modulate the expression of MHCII or cytokines in the DG, suggesting that the effect of running on neurogenesis is not mediated by microglia (257). While running does not increase the number of total microglia or level of gene expression, running does influence the interaction between microglia and NPCs. Microglia isolated from runners induce more neurosphere formation of NPCs in vitro compared with microglia isolated from nonrunner mice (350). Fractalkine, whose receptor CX3CR1 is found on microglia (21), is critical in maintaining microglia in the resting, inactivated state (60). In vivo, CX3CR1-deficient mice, which have disrupted fractalkine signaling, exhibit decreased numbers of BrdU-positive cells as well as Dcx-positive cells, indicating a decrease in overall neurogenesis (21, 350). Infusion of a blocking antibody targeting CX3CR1 in running mice generated fewer neurospheres compared with wild-type runner mice, suggesting that the resting state of microglia is necessary for normal NPC activity and is an integral component of the neurogenic niche (350). Studies have also used aged animals to investigate the role of microglia in the neurogenic niche. While hippocampal neurogenesis occurs throughout adulthood, there is a significant decline in the rate of neurogenesis with increasing age. Aged animals have significantly less NPC proliferation, neuronal differentiation, and newborn neuron survival compared with younger animals (43, 134, 173). Age-related changes are also seen in microglia, and increased levels of inflammation and oxidative stress, both of which strongly inhibit neurogenesis, are observed in aged animals (182). In addition, microglia profiles of aged animals include more activated microglia as well as higher levels of TNF-␣ and IL-6 secretion upon activation, resulting in elevated levels of proinflammatory cytokines in the brain (203, 251). Aged animals have lower levels of anti-inflammatory signaling, such as the chemokine fractalkine. Intracerebroventricular infusion of fractalkine into aged animals restores hippocampal cell proliferation, suggesting that the presence of fractalkine helps to provide a noninflammatory environment supportive of neurogenesis. This phenomenon appears to be mediated by IL-1, as blocking CX3CR1 increases IL-1 protein levels in the hippocampus (21). These data further highlight the sensitivity of neurogenesis to the balance of pro- and anti-inflammatory cytokines and chemokines. In addition to inflammation, microglia play a key role in modulating levels of oxidative stress by producing the antioxidant glutathione (136, 198). Microglia from aged mice have significantly lower glutathione levels than microglia from young mice (251), and oxidation products are found in microglia in aged mice (133), suggesting that microglia are less effective in attenuating oxidative stress in aged mice. These data illustrate how changes in the activity of microglia with age potentially contribute to the age-related decline in neurogenesis by altering the neurogenic niche. B. Astrocytes In addition to microglia, NPCs interact with other cell types present in the neurogenic niche (FIGURE 1). Astrocytes represent one of the major contributors to the neurogenic niche, as coculturing with astrocytes, both directly and without contact, induces neuronal differentiation of NPCs (27, 256, 313). The relationship between astrocytes and NPCs depends on a number of factors, including gene expression of the astrocytes. Coculturing astrocytes with neurospheres has revealed the importance of GFAP and vimentin in mediating cell-cell contact between these two cell types (358). Wild-type neurospheres differentiated into neurons at a greater rate when cocultured with astrocytes without GFAP and vimentin expression, indicating that these intermediate filament proteins negatively regulated neurogenesis in vitro (358). This inhibitory effect may be mediated by Jagged1 and Notch signaling pathways, as Jagged1 mRNA is significantly decreased in astrocytes without GFAP and vimentin (358). Other proteins in hippocampal astrocytes, such as ephrins, have also been identified to play a role in regulating neurogenesis through cell-cell contact (18). Specifically, ephrin-B2 expressed by hippocampal astrocytes induced neuronal differentiation of NPCs by activating -catenin signaling, and shRNA-mediated knockdown of astrocytic ephrin-B2 resulted in reduced hippocampal neurogenesis (18). The region of origin of astrocytes plays a significant role in their ability to promote neurogenesis of NPCs. Astrocytes isolated from the adult hippocampus but not spinal cord promote neuronal differentiation of NPCs in coculture through both soluble factors and cell-cell contact (313). While astrocytes derived from the hippocampus and cortex are able to induce neuronal differentiation, those isolated from the hippocampus are more effective, suggesting hippocampal astrocytes release more potent neurogenic factors (256). Analyzing gene expression from astrocytes isolated from different regions of the central nervous system has identified a number of key factors. Two of these factors released by neurogenesis-promoting astrocytes are IL-1 and IL-6, which are proinflammatory cytokines that, in combination with other factors such as vascular cell adhesion molecule-1 Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 995 AIMONE ET AL. (VCAM-1) and interferon-induced protein 10 (IP-10), increase neuronal differentiation from NPCs (27). While proinflammatory cytokines generally inhibit neurogenesis, this inhibition depends largely on the concentration of the cytokines. More importantly, it is the combination of factors secreted by astrocytes into the local microenvironment that produces the neurogenic niche (27). In contrast, astrocytes isolated from the spinal cord express more insulin-like growth factor binding protein-6 (IGFBP-6), decorin, and enkephalin, which negatively regulate neurogenesis (27). One factor that is continuously expressed in the hippocampus throughout adulthood is Wnt3a, representing a promising key factor of the neurogenic niche (197). Wnt3a is produced by astrocytes, and -catenin signaling is stimulated in NPCs, indicating a possible pathway by which astrocytes directly regulate NPC activity (197). The Wnt signaling pathway has been shown to regulate NPC proliferation and differentiation in adult neurogenesis in vitro and in vivo (175, 197, 356). Wnt modulates adult neurogenesis by upregulating NeuroD1 expression, which is an essential transcription factor in the generation of GCs and neuronal differentiation from NPCs (175). In response to Wnt, NPCs increase expression of LINE-1, a retrotransposon that is important in NPC survival, demonstrating that Wnt regulates multiple facets of adult neurogenesis in the hippocampus (175). Other components of the Wnt pathway, including Dickkopf-1 and secreted frizzled-related protein 3, have been recently implicated in the regulation of neurogenesis by aging and behavior (143, 297). Glial metabolism appears to be a key modulator of adult hippocampal neurogenesis. Selective inhibition of glial metabolism can be accomplished by treating cells with fluorocitrate, which inhibits glial aconitase, leading to reduced levels of glutamine (106). While it is unclear if there are direct effects on NPCs, fluorocitrate treatment results in a decrease in NPC proliferation both in vivo and in vitro, suggesting that a critical metabolite produced by astrocytes may be necessary for NPC proliferation (58). ATP from astrocytic metabolism serves as a signaling molecule to increase NPC proliferation in vitro and in animal models with defective astrocytic ATP release (58). Behaviorally, animals deficient in astrocytic ATP release exhibit depression-like behavior, which can be reversed with the administration of ATP (59), supporting the link between adult neurogenesis and depression-like behavior (283). Studies have further investigated the role of astrocytes in the neurogenic niche using aged animals to evaluate how agerelated changes in astrocytes correlate with their low levels of neurogenesis. The levels of hippocampal growth factors that are known to promote neurogenesis, including insulin growth factor-I (IGF-I) and vascular endothelial growth factor (VEGF), are significantly reduced in aged rats compared with young rats (36, 305). While there are conflicting 996 reports about whether levels of FGF-2 or its receptor are decreased in the hippocampus of aged rats, it is clear that this pathway is affected negatively with age (36, 305). The overall density of hippocampal astrocytes, which are immunopositive for GFAP, remains the same throughout adulthood; however, the density of astrocytes that express FGF-2 declines with age in the DG as well as other regions of the hippocampus, suggesting that the local environment for NPCs progressively becomes less supportive of neurogenesis with age (305). The altered profile of astrocytes in the aged hippocampus suggests that astrocytes are a major contributor to maintaining a neurogenic niche, and the decline in their secreted, proneurogenic factors correlates with the decline in hippocampal neurogenesis with age. Interestingly, brain-derived neurotrophic factor (BDNF) is a neurotrophin that regulates hippocampal neurogenesis (194, 263). TrkB, a high-affinity receptor for BDNF, is expressed on NPCs (194), and levels of BDNF are highest in the hippocampus and hypothalamus in rat brains (154). Studies have demonstrated that hippocampal astrocytes minimally produce BDNF (281, 282), and BDNF is highly localized within the nucleus of GCs of the DG (154). While BDNF protein levels in the hippocampus do not decline with age, as do the previously discussed factors (38, 153, 178), its receptor TrkB decreases with increasing age in the hippocampus of rats (74, 311). There is also evidence that serum levels of BDNF in humans are correlated with hippocampal volume and memory, as illustrated by the lower levels seen in older people with smaller hippocampi and poor memory (94). C. Vasculature In addition to secreting factors to NPCs, astrocytes are tightly linked physically with endothelial cells, wrapping their endfeet around blood vessels (FIGURE 1). The vasculature represents an abundant source of extrinsic factors that can modulate adult neurogenesis. Illustrating this possibility, NPCs can be found in clusters in close proximity to blood vessels (264, 304). The physical location of NPCs in relation to blood vessels suggests that NPCs may receive critical factors from the vasculature to stimulate proliferation, neuronal differentiation, and survival (264). Endothelial cells secrete soluble factors that increase NPC proliferation, neurogenesis (303), maturation, and migration, in part by secreting BDNF (191). The intimate relationship between NPCs and the vasculature is much more evident in animals with access to a running wheel. As discussed earlier, running is a potent stimulator of adult hippocampal neurogenesis, which is accompanied by an increase in cerebral blood flow in the DG (266). The number of blood vessels does not increase with running, but the surface area covered by blood vessels is significantly increased in the DG (344). The increase in the Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS density of blood vessels of runners is specific to the DG and not the entire hippocampus (66), a finding that is consistent with the increased cerebral blood flow observed in running mice (266). Although the number of blood vessels remains stable in the DG with running, there is evidence of angiogenesis in the molecular layer of the hippocampus with running, because newborn cells colabeled with an endothelial cell marker, rat endothelial cell antigen-1, are observed in this region (92). Angiogenesis and neurogenesis rely heavily on the circulating growth factors and nutrients supplied by the vasculature. VEGF is a factor that has been shown to promote proliferation of not only endothelial cells but also NPCs (145). Expression of VEGF and its receptors is not limited to endothelial cells but also expressed in NPCs, neurons, and glial cells. VEGF mRNA and protein are expressed in the hippocampus, colocalizing with GFAP and NeuN (57, 196). VEGF-induced neurogenesis is mediated predominantly through the Flk-1 receptor rather than Flt-1. Expression of Flk-1, but not Flt-1, is robust in the SGZ, and Flk-1 colocalizes with BrdU-positive cells in the SGZ (57, 145). Adult rat NPCs express both VEGF and Flk-1, and VEGF stimulates neurogenesis via Flk-1 in vitro (292). In addition to local signaling, VEGF upregulation in the periphery is essential to the increase in neurogenesis induced by running, illustrating its broad range of influence in modulating neurogenesis (98). III. DEVELOPMENT AND MATURATION PROCESS OF ADULT-BORN NEURONS In the adult SGZ, NSCs give rise to NPCs, which in turn generate immature neurons. Immature neurons migrate a short distance to the granule cell layer (GCL). Adult-born neurons follow the same outside-in layering pattern as developmentally born neurons in DG, with adult-born neurons preferentially staying in the inner or middle of the GCL and cells born in embryonic and postnatal stages of development found in the outer GCL (219). Extracellular matrix molecules, such as the Reelin signaling pathway (45), tyrosine kinase receptors (70), and chemoattractive and repulsive signals (362), may play important roles in the migration of adult-born neurons. While the role of extracellular matrix molecules in neurodevelopment has been extensively studied, a recent study investigated the effects of manipulating Reelin and Disabled-1 (Dab1) on newborn cell migration and dendritic maturation in the hippocampus (327). In mice that overexpressed Reelin under the Ca2⫹/ calmodulin-dependent protein kinase II (CaMKII) promoter, 2-wk-old neurons displayed greater dendritic branching and arborization, indicative of faster maturation, compared with those found in wild-type littermates. Consistent with this finding, conditional deletion of Dab1 in newborn neurons resulted in aberrant dendritic extensions and shorter, fewer processes. Disturbed migration and integration are often identified in pathological conditions such as epilepsy, schizophrenia, and neurodegenerative diseases (63). A. Morphological Maturation of Adult-Born Neurons It takes roughly 2 mo for newborn neurons to reach morphological maturity (FIGURE 1). Although no significant structural differences are observed between fully mature adult-born and perinatal-born neurons, the maturation process is delayed in the adult (259, 371). It is likely that differences in the local niche between the adult and neonatal brain contribute to the different maturation rate. In the adult hippocampus, by roughly 1 wk after differentiation, newborn neurons can be seen extending their apical dendrite into the molecular layer, and spines are first detected in the dendrite at 16 days post division (dpd). Spine density significantly increases between 21 dpd and 28 dpd (371), which corresponds to a critical period for neurogenesisrelated learning and memory (325). The number of spines appears to reach a plateau by 56 dpd. Furthermore, during these initial 2 mo, the spines remain highly plastic, dynamic, and regulated by neural activity (371). Axon mossy fibers reach to the CA3 region before the first dendritic spine is detected (371) (FIGURE 2). The thin axon fibers are first found in the hilus at 10 dpd and continue growing until they reach to CA3 (371). Axonal boutons in hilus are significantly smaller than those in the CA3 region (330), consistent with what has been classically observed of mossy fibers. Synapses arising from adult-born neurons can be identified by electron microscopy (EM) as early as 17 dpd onto postsynaptic targets in the CA3 and the hilus (330). These axon boutons form synapses on thorny excrescences and dendritic shafts of neurons in CA3 and hilus, respectively. The overall size of these mossy terminals, the number of presynaptic vesicles, and the number of activated zones continue to increase until roughly 75 dpd, when they are comparable to those of perinatal-born neurons. In addition, some thin axon fibers form en passant synapses onto the dendrites of GABAergic interneurons. At a population level, synapses from 3- to 4-mo-old neurons were found to be functional by use of Channelrhodopsin-2 (ChR2) (330). A subsequent study using a similar optogenetic technique demonstrated that the maturation of mossy fiber axonal contacts onto CA3 from young neurons had a roughly similar time course as dendritic maturation, with newborn mossy terminals passing through several stages of increased plasticity (129). Notably, the spine formation process of adult-born neurons appears to be different from that of perinatal-born neurons in that adult-born neurons preferentially target preexisting synapses, and little is known about the underlying mechanisms (332). One hypothesis is that glutamate spillover may play a chemoattractive role and induce filopodia growth Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 997 AIMONE ET AL. EC2 EC3 CA1 SUB EC5 CA3 DG EC2 DG CA3 EC3 Pyramidal neuron Hilar interneuron MOPP cell Granule cell - newborn Garnule cell - adult Basket cell Mossy cell EC2 neuron EC3 neuron EC5 SUB CA1 FIGURE 2. Anatomy of hippocampal circuit into which new neurons integrate. Neurogenesis is localized to the dentate gyrus (DG) region, where only excitatory granule cells are continually produced throughout life. The DG has a complex local circuitry, with both inhibitory interneurons and excitatory feedback neurons (mossy cells) participating in the network’s behavior. Granule cells in the DG project to the CA3 region, which in addition to a robust recurrent connection then projects to the CA1 region. The CA1 then projects back to the entorhinal cortex and subiculum regions, closing the “hippocampal loop.” towards active synapses (331, 332). Local synaptic activity may induce glutamate release and activate glutamate receptors in filopodia, which induce new filopodia to target the existing synapse (331). Recent evidence suggests that local astrocytes may also contribute to this targeting process (171). Three-dimensional EM analysis shows that both synaptic outputs (axon boutons) and inputs (dendritic spines) from adult-born neurons form intermediate structures on which multiple synaptic partners are shared at a single synaptic structure (i.e., multisynapse boutons) (330, 332). As neurons mature, the proportion of multisynapse bouton structures decreases. This structural difference parallels the differences between the physiologies of immature and mature GCs. Newborn neurons display a high input resistance (97), receive less inhibition (193), and have been shown to exhibit considerably greater synaptic plasticity (115, 295). Because of these unique properties, young neurons are likely to be more excitable than mature neurons (97, 231, 232), and thus, in response to presynaptic inputs, the synapses formed by newborn neurons in the multisynapse boutons may be more 998 dynamic than the existing synapses. The increased dynamics of young synapses may allow new protrusions to enlarge and subsequently lead to the eventual retraction of spines from the mature neurons (331). B. Physiological Maturation of Adult-Born Neurons For integration into the DG circuit, adult-born neurons follow the same process as perinatal neurons: silent synapse, GABA excitatory response, glutamatergic inputs, and GABA inhibitory response (97). The amino acid GABA is the major inhibitory neurotransmitter in the adult brain (170). However, GABA plays very different roles in NPCs and their progeny (114). The presence of functional GABA receptors is observed as early as in type 2 progenitor cells (333). After neuronal fate is determined, yet prior to receiving synaptic inputs, adult-born neurons are tonically activated by GABA released at extrasynaptic sites (81, 114). GABA’s tonic effects are able to Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS regulate the excitability threshold for neurons (117, 351), including reducing its own tonic effect, enhancing longterm potentiation (LTP) and improving performance on hippocampus-dependent spatial learning (19, 69). In mature cortical and hippocampal neurons, ␣5 or subunits are primarily components of extrasynaptic GABAA receptors (117, 351). Little is known about the major components of the extrasynaptic GABAA receptors in adult-born neurons and about how GABA’s tonic effects regulate their maturation, integration, and function. This gradual increase of inhibition, together with the changes in the intrinsic physiology of young neurons and the onset of glutamatergic synapse formation in adult-born neurons, sets up a critical period that is a unique feature of young neurons during their maturation (193). This critical period is particularly important for the function of newborn neurons in memory encoding (6, 82). Functional GABA and glutamate receptors are detected in 3-day-old immature neurons (97), but GABAergic synaptic events are not detected until neurons are a week old. Similar to what is seen in type 2 progenitor cells and neurons in developing brain, high [Cl⫺]i and expression of Na⫹-K⫹ transporter NKCC1 in immature neurons cause GABA to depolarize instead of hyperpolarize cells. This initial depolarizing GABA phenotype has been shown to be critical for the maturation of adult-born neurons (64, 114). Knocking down NKCC1 in immature neurons leads to significant defects in both GABAergic and glutamatergic synapse formation as well as in dendritic development of new neurons (114). Two possibilities may explain this phenotype: manipulation of NKCC1 expression may convert GABA from excitatory to inhibitory, which causes the defect in synapse formation and in morphology, or NKCC1 manipulation changes the [Cl⫺]i inside of the cell. [Cl⫺]i is very critical for cell function (144). [Cl⫺]i may act as an intracellular messenger to directly regulate expression of genes such as the GABAA receptor subunit (320) and KCC2 (141). Changing [Cl⫺]i itself may induce malformation of synapses and dendritic arborization. Different combinations of GABAA receptor subunits are involved in GABAergic tonic and phasic response (117). Developmentally decreasing [Cl⫺]i in a cell might contribute to the switch of GABA from its tonic to phasic effects (261, 320) as well as the activation of the glutamatergic synapse (352). As described above, the maturation of adult-born GCs generally mimics the neuronal maturation process observed in development, albeit at a slower time scale. The same holds true for the dynamics and regulation of gene transcription in maturing GCs. Many of the gene expression profiles observed in development are also evident in adult-born GCs, and these genetic configurations are integral to the regulation of the maturation process. Details concerning the transcriptional and genetic regulation of neurogenesis have recently been examined in depth elsewhere (100, 139, 156, 207, 237). These processes fit into the “control” processes underlying neurogenesis as described by Kempermann (156), and mechanistically would be expected to be similar between developmental and adult-born neurogenesis. Rather than fully repeating that information here, we refer the reader to those reviews and focus our discussion on several examples in depth, emphasizing the circuit and behavioral factors that regulate the proliferation and differentiation of new neurons that will be specific to the regulation and integration adult neurons. Glutamatergic inputs are detected in 3-wk-old adult-born neurons. Interestingly, glutamatergic currents in immature neurons display a lower threshold for the induction of LTP facing stimulation of the perforant pathway (PP) (115) in a specific time window. Developmentally regulated synaptic expression of NR2B containing NMDA receptors partially contributes to the mechanisms for LTP induction in immature neurons (115). Fast, perisomatic GABAergic inhibitory currents are recorded in neurons later than functional glutamatergic inputs. Adult-born neurons receive local inhibitory inputs from all three subregions of DG: molecular layers, GCLs, and hilus (193) (FIGURE 2). With maturation, inhibitory inputs to adult-born neurons gradually increase. This developmental change is due to an increase in the number of inhibitory synapses as well as their synaptic strength (193). IV. REGULATION OF NEUROGENESIS The regulation of neurogenesis can be targeted at several steps of the overall process. As described above, stem cells pass through several distinct morphologically and genetically identifiable stages, most notably a slowly dividing RGC stage and a more rapidly proliferating neural progenitor stage. Although it is still somewhat a matter of debate whether RGCs truly self-renew, it appears clear that cells of the morphologically more compact progenitor population are essentially fate-committed to become neurons after a few rounds of amplification at most (88, 93, 321). A. Network Level Regulation The substantial regulatability of NSCs and NPCs suggests that local circuit activity likely can affect proliferation and differentiation while in the stem cell state (FIGURE 3). Recently, Song et al. (314) demonstrated that nestin-expressing RGCs in the SGZ were responsive to GABA through ␥2 receptors (314). This GABA response appeared to be tonically active and originate from nearby, but not synaptically connected, parvalbumin (PV) basket cells. This GABA, presumably from spillover from GABAergic synapses, suppressed the symmetric proliferation of RGLs, instead biasing them to quiescence. Thus increased PV-basket cell ac- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 999 AIMONE ET AL. >4 wk 3 wk 2 wk 1 wk Basket cell 3 days type 2 type 1 Astrocyte to CA3 Proliferation Differentiation ? Survival ? GABA Glutamate Serotonin (5HT) Norepinephrine (NE) Acetylcholine (ACh) Dopamine (DA) FIGURE 3. Regulation of neurogenesis by local circuit factors. Each of the three stages of neurogenesis– proliferation, differentiation, and survival–is a target of regulation by network factors. Local GABA, the primary inhibitory neurotransmitter in the brain, appears to suppress proliferation while inducing differentiation and survival. Glutamate, the primary excitatory neurotransmitter, is necessary for proper survival as well. Modulatory neurotransmitters, which have been implicated in numerous mental health conditions, appear to regulate different parts of the process; for instance, serotonin induces proliferation, whereas acetylcholine is necessary for proper maturation and survival. tivity, which likely corresponds to overall increased DG activity, will lower proliferation of stem cells, whereas decreased DG activity will likely lower tonic GABA levels and increase proliferation. This direct effect of regulation of RGCs by GABA, and presumably inverse correlation of proliferation with overall network activity, is notable with respect to another recent study by Dranovsky et al. (88) showing that exercise and EE increase 1000 the number of neurons that derive from the RGC population (while preserving overall numbers of RGCs), whereas social isolation significantly increases the density of RGCs. The model proposed by the authors is that EE increases the proliferation of the intermediate NPC population and that social isolation increases the RGC population directly. As described by Song, the effect of GABA on RGCs is likely due to diffusion from nearby synapses. Diffuse GABA is Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS only an indirect measure of network activity, and it is not long after progressing from the RGC stage that NPCs and very young new neurons begin to receive more direct GABAergic and glutamatergic inputs (through NMDA receptors) (80, 114, 215, 217, 260, 326, 333, 353). Responses to GABA progress from tonic to “phasic” GABA roughly around the time of activation of DCX as a label and, when tracked using retrovirus, within the first several days post injection (97, 114). Notably, the response to GABA is depolarizing in NPCs and young neurons due to the distinctly high reversal potential of [Cl⫺] because of the preferential expression of the NKCC1 ion pump as opposed to the KCC2 ion pump (114). In NPCs, this depolarizing effect of GABA is necessary for the influx of [Ca2⫹] through voltage-gated calcium channels (333); in turn, calcium activity has been linked to the critical pro-neuronal differentiation gene NeuroD1 (80). Activation of local interneurons in hilus elicits a GABAA receptor-sensitive synaptic response in type 2 progenitor cells. High [Cl⫺]i in type 2 progenitor cells makes the effects of GABA depolarizing as opposed to hyperpolarizing. GABA mediates excitation in NPCs and subsequently induces calcium influx via voltagegated calcium channels, which promote NeuroD expression and lead differentiation towards a neuronal fate. Recently, it was reported that GABA released from PV-positive interneurons in the hilus promoted radial glia-like quiescent NSCs to actively divide (314). GABA can also provide a trophic effect in early development, such as promoting DNA synthesis in proliferating neurons (202) and cell motility of immature neurons (32, 216). Notably, the role of GABA in neuronal development has also been well characterized in the SVZ, where it is essential to regulating proliferation of progenitors (201). Once differentiation is complete and neuronal maturation begins, the young neurons begin to develop considerably more sophisticated connectivity to the local circuit, which in response regulates the maturation process (97, 179, 215, 217). GABA remains depolarizing in young neurons until roughly 2 wk of age (114), which roughly corresponds to the time in which glutamatergic spines develop on immature dendrites (371). Glutamatergic inputs become more and more important as immature neurons develop. Initially the primary influence of glutamate is through NMDA receptors, which are notable for their permeability to Ca2⫹. NMDA appears to be expressed in NPCs and has been shown in vitro to have a direct effect on internal calcium levels (80); however, physiology studies typically do not show any glutamatergic signaling in NPCs or very young neurons in slice (97, 333), which is not surprising given their localization to the SGZ, where few synapses are glutamatergic. By roughly 2.5 wk of age, young neurons do have dendrites with spines in the molecular layer and receive glutamatergic inputs. NMDA appears to be significant in these early stages of excitatory synapse formation (115, 326, 367), with NMDA activation involving the NR1 sub- unit being critical in the survival of young neurons passing through the 2- to 3-wk-old “critical period” (326). Notably, the requirement of NMDA for neuronal survival appears to be relative to the surrounding circuit; global blockade of NMDA in the circuit mitigates the effect of the conditional NR1 knockout to some extent (326). The NMDA-dependent survival of young neurons corresponds to the timing of EE’s effects on survival (325), as well as a particular period of increased plasticity in maturing neurons (115). 1. Neuromodulators and neurogenesis regulation In addition to the local circuit effects on neurogenesis, several of the numerous modulatory systems that have effects on hippocampal function have been shown to affect the proliferation and differentiation of NSCs and NPCs and the maturation of adult-born neurons (FIGURE 3). Of these, serotonin (5-HT) has been the most heavily studied, in part due to its relationship with depression and stress (121, 283). Attention to serotonergic effects on the NSC population was stimulated by the observation that 5-HT-selective reuptake inhibitor (SSRI) antidepressants such as fluoxetine could directly induce increased levels of proliferation (209). Subsequent studies showed that the putative 5-HT-increasing effects of SSRIs were consistent with a direct effect of 5-HT on proliferation that likely arises through the 5-HT1A receptor, with possible contributions through the 5-HT1B and 5-HT2A receptors (25, 271, 289). Serotoninergic fibers project to the DG from two distinct regions, the median raphe (MR) nucleus and the dorsal raphe (DR) nucleus (188). MR projections are known to provide substantial inputs to the varied interneuron populations in the hilus and throughout the hippocampus, often onto the ionotropic (unique among the monoaminergic systems) 5-HT3 receptors (110, 345). In contrast, the DR projections to the DG terminate preferentially in the SGZ region, with fairly small axonal terminations that appear to be positioned for diffuse release of 5-HT (169). As a result, it appears that the DR projection is best positioned to impact the neurogenesis process directly; however, the MR may well regulate local circuit activity through an indirect effect via local interneurons. The effects of the other major neuromodulatory systems [norepinephrine (NE), dopamine (DA), acetylcholine (ACh)] on neurogenesis are less well understood (346) (FIGURE 3). ACh is one of the more attractive candidates for a potential relationship, as the cholinergic systems are known to have a substantial relationship with the hippocampus and the DG in particular. Depletions of ACh (through lesions of basal forebrain cholinergic regions) lead to a reduction in neurogenesis (71, 148); however, high doses of selfadministered nicotine also decrease neurogenesis (1). The ␣7-nAChR receptor was shown to be essential for the proper maturation of adult-born GCs, with decreased survival of new neurons and reduced synaptic development in Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1001 AIMONE ET AL. those young neurons that survive in ␣7KO mice (55). Like ACh, NE has substantial impacts on the DG (130), and the limited studies with reference to neurogenesis suggest that it is required for normal neurogenesis; selective lesions of NEprojecting fibers decrease neurogenesis considerably (174) and increases in NE through blockade of NE-axon autoreceptor inhibition lead to increased proliferation and survival (278). One recent study using an in vitro assay suggests that the effects of proliferation may be through 2 receptors (218). Finally, the dopaminergic system, which is notoriously difficult to study in vivo, appears to have effects on neurogenesis as well; however, these studies have generally investigated the effects of antipsychotic drugs and Parkinson’s disease models on neurogenesis, so the direct effects are difficult to elucidate (346). It does appear that D2 receptors are involved in in vivo DG neurogenesis (365). Furthermore, it has been reported that DA can selectively impact the LTP potential of maturing neurons (238), so it is reasonable that DA could affect the maturation process of new neurons as well. Notably, these systems are all tightly coupled and known to be related to behaviors such as stress and exercise that are associated with neurogenesis modification (see below), so the absence of clear links is more indicative of the challenges in studying these systems in isolation in vivo than of an indictment of their functional importance. B. Regulation by Local Signaling In addition to the direct effects of neuronal activity on proliferation and differentiation, the DG network is capable of regulating neurogenesis through local astrocytes. Wnt signaling has previously been shown to play a critical role in the neuronal differentiation of NSCs (197). Wnt3a is secreted by astrocytes in the SGZ, whose factors have been shown to be capable of permitting neuronal differentiation (313). Through the canonical Wnt signaling pathway, -catenin acts as a transcriptional activator of TCF/LEF transcription factors that in turn regulate the expression of Wnt target genes. Two such targets are the transcription factors NeuroD1 and Prox1 (112, 151, 175, 183). Originally identified for its ability to activate the insulin gene promoter (246, 247), NeuroD1 has been shown to play a crucial role in neuronal differentiation (112, 183). NeuroD1 (also known as NeuroD and BETA2) is a basic helix-loop-helix (bHLH) transcription factor required for the survival and maturation of adult-born neurons. NeuroD1 null mice die from early-onset diabetes due to loss of expression of the insulin gene (246), so to study the role of NeuroD1 on adult neurogenesis, a transgenic mouse expressing a floxed NeuroD1 gene and ROSA26 YFP reporter was crossed with a tamoxifen (TAM)-inducible CRE recombinase under the transcriptional control of nestin (118, 177). A dose-dependent injection of tamoxifen allowed the conditional knockout of NeuroD1 in all nestin-expressing 1002 stem cells and their progeny. Compared with wild-type mice, in the NeuroD1 knockout animals, deletion of NeuroD1 did not alter the number of YFP-expressing recombined cells 6 days post TAM injection; however, 40 days after injection, the number of YFP-positive cells was significantly reduced (112). Morphological analysis revealed no difference in proliferation (YFP/Ki67) at 6 days, further supporting the idea that NeuroD1 is not required in early NPCs. However, reductions in both YFP/Prox-1 and YFP/ DCX at 40 days post TAM injection indicate a critical role for NeuroD1 during the later stages of differentiation/maturation. The dendritic length of YFP cells in NeuroD1 knockout mice was reduced as well. These data together highlight NeuroD1 as a critical intrinsic transcription factor that regulates survival and maturation of immature granule neurons as they become mature granule neurons. Another target of the Wnt signaling pathway is Prox1. Prox1 is unique in that its expression is restricted to the DG (180), whereas NeuroD1 is expressed in both neurogenic regions of the brain. Prox1 has previously been demonstrated to be a target of TCF/LEF signaling (151). In vitro analysis using ChIP as well as a luciferase reporter assay revealed two functional TCF/LEF sites within the Prox1 enhancer region. This finding was further confirmed in vivo. Knockdown of TCF/LEF transcription through a retrovirus expressing a dominant negative form of LEF (dnLEF) led to a reduction in colocalization with Prox1 compared with cells expressing a control GFP retrovirus. In addition, overexpression of a dominant active form of -catenin induced significant ectopic expression of the Prox1 protein. To determine the regulation of adult neurogenesis by Prox1, viral methods were used to either overexpress or inhibit Prox1 expression (151). In the knockdown studies, a lentivirus expressing an shRNA against Prox1 was injected into animals. Two weeks later, CldU was injected to label a newborn cell population, and 3 wk after CldU administration, a dramatic reduction in the number of CldU/DCXpositive cells was observed. Interestingly, however, the researchers did not observe a reduction in the number of NSCs expressing Sox2 and GFAP, indicating that Prox1 affected the number of newborn neurons (DCX) without altering the proliferative NSC populations. Overexpression of Prox1 into newborn cells, using a retrovirus, enhanced neuronal differentiation 1 wk after injection, as demonstrated by colocalization with DCX. To determine whether Prox1 played a role in the maintenance of mature GCs, animals were injected with BrdU to label a newborn cell population. Sixteen weeks later, when these neurons fully matured, the lentivirus shRNA Prox1 was injected to knock down Prox1 levels. Interestingly, there was no difference in the number of BrdU cells compared with the control animals. These data suggest that Prox1 is required for the neuronal differentiation of newborn neurons but not nec- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS essarily for the maintenance and survival of these neurons once they have reached maturity. C. Regulation by Extrinsic Factors Neurogenesis is a highly dynamic process and is regulated by many extrinsic factors as well as intrinsic factors (FIGURE 4). These external manipulations are known to both positively and negatively impact levels of neurogenesis throughout the life of mammals. Some of the most prominent positive regulators of hippocampal neurogenesis include EE, voluntary exercise, and diet (118, 234, 256, 257). In contrast, factors such as aging and stress have been shown to dramatically reduce levels of neurogenesis (126, 220). In either case, manipulations of these common elements of life play a significant part in the way that adult hippocampal neurogenesis is regulated. One important observation is that in contrast to the molecular and circuit level regulatory factors described above, the extrinsic regulation of neurogenesis has been challenging to quantify with the same level of precision. While running >4 wk 3 wk 2 wk 1 wk Basket cell 3 days type 2 type 1 Astrocyte to CA3 Proliferation Differentiation Survival Learning Running Enrichment Stress FIGURE 4. Regulation of neurogenesis by behaviors. Neurogenesis is regulated by many behavioral factors as well. Running is one of the most potent inducers of neurogenesis, targeting the proliferation of neural progenitor cells. Enrichment has a complementary effect, increasing the survival of neurons at a critical stage of their maturation. In contrast, stress is a severe negative regulator of new neuron birth, suppressing proliferation. The effects of learning are more complex, suppressing the neurogenesis process at some stages while increasing it at other stages. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1003 AIMONE ET AL. and stress were among the earliest known neurogenesis modulators and are among the most potent regulatory factors, studies focused on extrinsic regulation, particularly with regard to enrichment and diet, have been somewhat more challenging to interpret. One reason is that the variability between individual animals and their responses to the manipulation can be quite substantial. For example, 40 mice were housed together in one large EE and their exploratory activity was recorded with radiofrequency identification transponders over 3 mo. Despite the similar genetic background of all 40 mice, there was huge variability in their exploration of the environment. Furthermore, levels of neurogenesis correlated with the amount of exploratory activity of each individual mouse (108). This variability in exploration can be extrapolated to other manipulations, including running, diet, and stress. An additional complication is the variability in experimental design, which makes cross-laboratory comparisons challenging. The following sections describe how the continuous process of neurogenesis is actively regulated by experiences we encounter on a daily basis, with the caveat that most of these findings remain phenomenological with only limited insight into the underlying mechanisms. 1. EE and Running An EE can be defined as an environment that provides sensory, social, and motor stimulation. This environment may consist of an enlarged enclosure along with tunnels, huts, toys, running wheels, and other animals to provide more social interactions. One of the first studies to demonstrate the impact of EE on adult hippocampal neurogenesis labeled newborn cells with BrdU in mice that lived in an EE for 40 days. The researchers discovered that there was a small but significant increase in the number of surviving BrdU cells when animals were killed 4 wk later (160). Although there was no effect on cell proliferation in these animals, the mice that were exposed to the EE showed significantly higher numbers of total granule neurons in the hippocampus. In an effort to determine the long-term effects of EE, 10-mo-old mice were housed in an EE for roughly half of their life (10 mo) (158). The number of proliferating cells, as determined by BrdU 1 day prior to EE exposure, increased (although not significant), as did the neuronal differentiation of newborn cells. Cells labeled 4 wk prior to death were more than fivefold more likely to differentiate into a neuron in mice that lived in an EE compared with mice that were housed in a standard cage. In multiple instances, these enrichment-induced increases in neurogenesis levels were correlated with improved performance on hippocampus-dependent behavior such as in the Morris water maze (MWM). Living in an EE for 4 wk is associated with increases in neurotrophins and growth factors, including VEGF and BDNF, in the rodent hippocampus. In rats, there is a signif- 1004 icant increase in VEGF mRNA expression after 4 wk of living in an EE (57). The increase in hippocampal neurogenesis with exposure to an EE appears to be dependent on VEGF expression, as shRNA-mediated knockdown of VEGF in hippocampal neurons inhibits EE-induced neurogenesis (57). In mice, expression of BDNF mRNA is upregulated, but VEGF, nerve growth factor (NGF), and epidermal growth factor (EGF) are not observed after exposure to an EE for 4 wk (176). Upregulation of BDNF is due to histone modifications of the BDNF promoter (increased histone H3K4 trimethylation of BDNF P3 and P6 promoters, and decreased histone H3K9 trimethylation of BDNF P4 promoter and histone H3K27 trimethylation of BDNF P3 and P4 promoters). BDNF induces phosphorylation of mitogen/stress-activated kinase 1 (MSK1) (272). MSK1 is also activated by the ERK1/2 or MAPK signaling cascades and is involved in CREB and H3 phosphorylation, activation of the NFB pathway, and the regulation of immediate early genes (for review, see Ref. 17). There is recent evidence that MSK1 is involved in regulating hippocampal synaptic plasticity and not only baseline but also EE-induced adult hippocampal neurogenesis (72, 152). In a mouse model without functional MSK1, MSK1⫺/⫺ mice had less proliferation and fewer immature neurons in the DG compared with wildtype mice (152). After exposure to EE for 40 days, wild-type mice had increased cell proliferation and CREB phosphorylation, whereas these remained unchanged in MSK1⫺/⫺ mice (152). These data suggest that MSK1 is part of a critical signaling pathway involved in regulating EE-induced hippocampal neurogenesis. Running wheels were used in these particular EE paradigms, and it was later discovered that voluntary exercise, i.e., running, was one of the most salient features of the EE (340, 341). EEs are composed of multiple types of sensory enrichment and, although exposure to these environments has been shown to positively affect adult neurogenesis, it was unknown which component(s) contributed most to this effect. To answer that question, adult mice were assigned to random groups based on the different types of enrichment the animals experienced during exposure to an EE (341). These groups were termed learners, swimmers, runners, and enriched (without running wheel). Only the running and enriched groups of animals exhibited increased levels of neurogenesis. Consistent with previous findings, enrichment, even without a running wheel, had the biggest impact on the survival of newborn neurons 4 wk after BrdU labeling. Runners, however, displayed a dramatic increase in the number of proliferating cells, ultimately leading to an overall increase in the number of surviving newborn neurons at 4 wk post BrdU labeling. Both swimmers and learners showed no differences in either cell proliferation or cell survival compared with control animals. Interestingly, run- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS ning also had an influence on synaptic plasticity, specifically enhancing LTP in the DG of the hippocampus (340). Like enrichment, the increase in neurogenesis and enhanced LTP associated with running also correlated with improvement in hippocampus-dependent behavior such as in the MWM. More recently, several studies have suggested that the notable increase in neurogenesis observed with enrichment can be attributed mainly to physical activity associated with exercise (167, 239) (FIGURE 4). Animals were separated into four groups: control, enriched only, enriched running, and running. Both the enriched running and running groups were allowed free access to running wheels. Groups that were housed with running wheels (enriched running and running) demonstrated significantly higher levels of neurogenesis than controls, and this enhancement was positively correlated with an improvement in MWM performance (121). Although the enrichment-only group did not show enhanced levels of neurogenesis, they did demonstrate a faster habituation to the open field test compared with controls, suggesting beneficial effects of enrichment independent of neurogenesis. Additionally, one study has suggested that the beneficial effects of EE are independent of hippocampal neurogenesis (225). Following complete ablation of hippocampal neurogenesis through x-focal irradiation, irradiated mice exposed to an EE performed as well as sham control enriched animals on the MWM, suggesting that the previously observed enhancement of enriched mice in MWM behavior was not attributable to hippocampal neurogenesis. Although the extent to which EE influences adult neurogenesis is debatable, one recent study has discovered that it affects neurogenesis on an individual basis (109). Forty mice tagged with radiofrequency identification transponders were allowed to live in an EE for 3 mo. Interestingly, adult neurogenesis correlated positively with the exploration of individual mice (determined by “roaming entropy”). Mice that spent more time actively roaming the environment showed a correlation with increased neurogenesis levels, suggesting that the impact of EE is ultimately dependent on the mouse’s individual experiences within the environment. Although both enrichment and exercise have been shown to enhance levels of neurogenesis, they may work through different mechanisms. Enrichment has a greater influence on cell survival and neuronal differentiation, whereas running appears to affect cell proliferation (341). Enrichment and exercise increase neurogenesis to different extents (although this varies from study to study), but the application of both of these manipulations together results in a synergistic effect on neurogenesis (99). Although the enhancements of neurogenesis associated with both running and enrichment are consistent with increases in trophic factors such as BDNF and NGF, the underlying differences between the two manipulations remain unclear. It is clear, however, that the intense physical activity associated with exercise leads to dramatic changes in the vasculature and blood-brain barrier permeability (258). Due to this increase in angiogenesis, circulating hormones and growth factors are more readily delivered to the hippocampus through the permeable blood-brain barrier and ultimately promote the proliferation of new neurons in the DG. Circulating growth factors that are induced through exercise include VEGF (98), IGF-I (334), and FGF-2 (56), all of which have been shown to influence hippocampal neurogenesis. Furthermore, the effects of running and enrichment may also involve neuromodulators such as ACh and 5-HT, which we described above. More recently, in an animal model that lacked brain 5-HT, basal levels of adult neurogenesis were not affected by the permanent depletion of 5-HT, but it was required for exercise-induced neurogenesis (166). Many studies have revealed the underlying molecular mechanism of regulating neurogenesis by exercise and enrichment. Running has been shown to increase thickness (12) and metabolic capacity (221) in specific regions of cortex. Neural plasticity and electrical activity have also been associated with exercise. Running and EE-associated motor activities increase cerebral blood flow (364), blood-brain barrier permeability (301), and glucose metabolism (348). These changes combined might increase hormone and growth factor level, such as VEGF (145), GDNF, and BDNF (286). BDNF in particular has received considerable attention in neurogenesis regulation. Proliferation of NPCs in vivo is boosted by exogenous BDNF injection into the hippocampus (293). Together with NT3, BDNF promotes the differentiation and maturation of adult NPCs in culture. The expression of BDNF is regulated by neural activity and plasticity. Increasing BDNF mRNA and protein has been found to be associated with memory acquisition and consolidation (33, 184). Interestingly, the signaling of BDNF promotes the survival and maturation of GABAergic inhibitory neurons. A recent study indicates that BDNF might potentially link neural activity and GABA-mediated effects on neurogenesis (354). 2. Aging While there are clear benefits of manipulations such as exercise, enrichment, and diet, other external factors are consistent with a decline in hippocampal neurogenesis levels. The most dramatic and well-studied of these negative regulators is aging. Aging is consistent with significant reductions in cell proliferation, survival, and neuronal differentiation. With the use of BrdU to label dividing cells at a given time point, proliferation in the DG was significantly reduced in aged (21 mo) rats compared with middle-aged (6 mo) rats. Overall numbers of newborn neurons decreased roughly eight- to ninefold from middle-aged to aged rats, indicating the dramatic decrease in neurogenesis associated with aging (173). Another study observing cell proliferation in rats at broader time points indicated that this age-related decrease in neurogenesis was observed as early as 6 wk of Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1005 AIMONE ET AL. age (when compared with 2-wk-old rats) and persisted until 12 mo, where it began to level off. When DG volume was quantified in these animals, the volume increased from 2 to 6 wk and then remained relatively stable through 24 mo of age (134). When comparing cell proliferation and total DG volume during aging, the initial decrease in proliferation from 2 to 6 wk may be attributed to the fact that the brain has not yet fully developed, suggesting a peak in cell proliferation at this time. Similar observations have been noted in mice. Cell proliferation in mice decreases from 1 to 2 mo of age and progressively declines until 18 mo, where the significant decreases in proliferation are no longer observed (34, 43). Survival of these newborn labeled cells was dramatically reduced in 3- to 6-mo-old mice compared with 18-mo-old mice when BrdU was quantified 4 wk after labeling. Consistent with the decrease in cell survival, colocalization of BrdU and the neuronal marker NeuN was also substantially reduced in both 3- and 6-mo-old mice compared with 18-mo-old animals, indicating a decrease in neuronal differentiation with age. Interestingly, this decrease in neuronal differentiation was offset by an increase in glial differentiation (173, 344). The age-related decline in hippocampal neurogenesis is likely a result of a reduction in NSC population and/or activity. The sex-determining region Y-box 2 (Sox2) and bHLH gene Hes5 have been identified as NSC markers (204, 293). In aged rats, the Sox2⫹ population remains unchanged but proliferation rates are decreased, suggesting that a decrease in NSC activity is the major contributor to the reduced hippocampal neurogenesis with age (131). In contrast, examination of Hes5⫹ cells in aged mice revealed an overall reduction in the total number of Hes5⫹ cells, indicating that there is a significant decrease in the NSC population with age (204). Specifically, the largest decrease was found in the proliferative horizontal Hes5⫹ cell population in aged mice. While it is unclear from these two studies whether the NSC population remains intact, there is a consistent finding of decreased proliferative activity of NSCs in aged animals. A combination of intrinsic and extrinsic age-related changes is likely to contribute to decreased hippocampal neurogenesis in aged animals (see Ref. 185 for review). A recent study identified that Wnt signaling from astrocytes regulates the expression of survivin, which is important for cell proliferation, in NPCs (229). Additionally, as discussed earlier in this review, the neurogenic niche is altered with increasing age; microglia and astrocytes transition from anti-inflammatory and antioxidative to proinflammatory signaling pathways. Exercise and enrichment are capable of increasing the number of newborn neurons in the DG fivefold, and aging is associated with an eight- to ninefold decrease, demonstrating the extent to which external manipulations are capable of affecting adult hippocampal neurogenesis. Given the breadth with which changes in the external environment 1006 influence neurogenesis, it is interesting to see how these external factors interact with one another. The most widely studied interaction is how enrichment and exercise affect aging in animals. As previously mentioned, when 10-moold mice (a time when decreased neurogenesis is already present) lived in an EE for 10 more months, they exhibited (at the age of 20 mo) a fivefold increase in the number of newborn neurons compared with control animals (158). The improvement in neurogenesis levels was consistent with performance in the MWM task. Interestingly, even at advanced stages of aging, when the reduction in neurogenesis and cognitive decline are most severe, voluntary exercise has been shown to mitigate many of these deficits. Animals, 18 mo of age, that were allowed access to a running wheel for 45 days demonstrated a remarkable increase in cell survival and neuronal differentiation, ultimately leading to an increase in the total number of new neurons (344). Aged enriched animals exhibited improvements in the MWM. 3. Stress Along with aging, another extrinsic factor negatively regulating hippocampal neurogenesis is stress (FIGURE 4). In the laboratory setting, there are various types of stressors used to observe the effects of stress on adult neurogenesis. Acute stressors are defined as a single stressful event, and chronic stressors are multiple stressful events over a period of time; however, both types of stressors vary based on their method of application (physical stressor, social stressor, and odor stressor) and duration (for a review, see Ref. 296). Despite the variability in how stress is applied, the general consensus of these studies is that stress has a substantial effect on DG cell proliferation. Tree shrews were subjected to acute psychosocial stress, for 1 h, when two males were placed together to establish a dominance/subordinance hierarchy that caused stress for the subordinate animal (125). After the stressful experience, animals received a BrdU injection and were killed 2 h later to determine cell proliferation. Tree shrews that were exposed to psychosocial stress exhibited significantly reduced numbers of proliferating BrdUpositive cells. Similarly, adult marmoset monkeys were subjected to a resident-intruder psychosocial stress model and, from this one stressful experience, intruder monkeys demonstrated a significant decrease in cell proliferation (127). There is one study, however, that did not observe an effect of stress on cell proliferation. Rats placed in a similar acute psychosocial situation did not exhibit a decrease in cell proliferation but rather a decrease in neuronal survival (328). Experiments utilizing a chronic psychosocial stressor method have observed similar decreases in cell proliferation in tree shrews, rats, and mice (76, 77, 104). Just like acute stressors, one study has also identified an inverse effect of chronic stress on cell survival (268). Interestingly, some studies have found that when animals are put through behavioral tasks, such as the MWM, the stress associated with either extended training paradigms (20) or even the novelty Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS of the task itself (91) is sufficient to reduce levels of neurogenesis. Animals that were exposed to multiple mild stressors such as cage tapping/tilting, auditory exposure to predators, cages with water or damp bedding, or housing with temporary light fluctuations were assessed for levels of neurogenesis. Survival of BrdU cells was significantly decreased in response to chronic exposure of two random mild stressors, across multiple mouse strains. This decrease in cell survival was consistent with a decrease in neuronal differentiation as quantified by BrdU and NeuN colocalization (228). Although slight differences in stress application and species of animal may account for the variability observed, the effects of stress on cell proliferation and cell survival have been well documented. Stress is associated with a release of glucocorticoids into the bloodstream, which have been shown to modulate neurogenesis (53, 123) and potentially relate neurogenesis to its hypothesized role in mood (186). Following performance of an adrenalectomy to reduce circulating adrenal hormones in the bloodstream of rats, an increase in the number of GFAP-positive cells in the DG was observed (123). This increase was reduced when corticosterone was introduced through the drinking water, although not to the level of sham treated animals. In a similar experiment, rats were given subcutaneous injections of corticosterone and a significant decrease in the number of proliferating cells in the DG was observed (53). An adrenalectomy of these animals dramatically increased the number of proliferating cells. These experiments suggest a role for circulating glucocorticoids in the bloodstream and their negative regulation of adult neurogenesis, hinting at a possible mechanism by which stress can affect adult neurogenesis. Interestingly, one of the most potent positive regulators of adult neurogenesis, exercise, is also associated with an increase in glucocorticoids in the bloodstream; however, these seemingly contradictory occurrences could be partially explained by the massive upregulation of circulating growth factors accompanying exercise. This influx of circulating growth factors may overshadow the negative effects of stress-induced glucocorticoids. Finally, expression of glucocorticoid receptor (GR) is robust in the GCL and colocalizes with both NeuN and DCX (98). Additionally, glucorticoids may act directly on NPCs, as treatment with a GR agonist reduces cell proliferation in vitro. shRNA-mediated knockdown of glucocorticoid receptors in the hippocampus disrupted migration and positioning of newborn cells in the GCL, suggesting a critical role for glucocorticoid receptors in mediating the integration of newborn cells into the existing circuitry (105). 4. Diet Lifestyle choices such as exercise and diet have a tremendous impact on many aspects of brain function, including mood, energy metabolism, behavior, cognitive function, and hippocampal neurogenesis. Many studies have investi- gated which specific foods or compounds can provide a cognitive advantage or disadvantage. Rodents on a diet that lacks essential vitamins or minerals exhibit decreased hippocampal neurogenesis, which is accompanied by impaired learning and memory (for review, see Ref. 316). In contrast, rodents on a diet supplemented with polyphenols or omega 3 fatty acids show increased hippocampal neurogenesis as well as improved performance on cognitive tests (for review, see Ref. 316). While there is an overwhelming amount of literature on diet and cognitive function, there are many factors that need to be considered before drawing conclusions about the effect of diet on cognitive function. Many of these studies do not take into account which compounds are being absorbed, how they are metabolized, and which specific metabolite acts directly in the brain to improve cognition. Furthermore, it is unclear how studies in rodents can be applied to humans, specifically with regards to dose and if humans can consume a comparable amount of the compounds. For these reasons, it is even more difficult to comment on in vitro studies in which compounds are directly applied to neurons to determine the effects of cell survival or function. Given the complicated nature of these studies, here we review a handful of studies that focus on specific flavonoids or flavonoid-rich diets in rodents and humans to provide a preview of a burgeoning field of cognitive neuroscience research. Flavonoids are a subset of polyphenols that are found in many different foods such as berries, parsley, green tea, and cocoa (for review, see Refs. 31, 210). Epicatechin, found in high concentrations in tea and cocoa, has been shown to improve spatial memory retention in adult mice (342). When epicatechin is combined with running in adult mice, the cognitive improvement is more robust, and this combination is associated with an increase in spine density, suggesting that a synergy exists between diet and exercise (342). Identifying individual compounds that are effective in enhancing neurogenesis and cognitive function is of particular interest to developing dietary interventions, particularly with the elderly population. Cognitive functions decline with increasing age, and a number of studies have investigated the potential use of flavonoid consumption to prevent this decline. In elderly humans (70 –74 yr old), consumption of chocolate, wine, and tea is associated in a dose-dependent manner with better test scores on a battery of cognitive tests (254). A 10-yr study examined the cognitive function of subjects aged 65 or older as assessed with psychometric tests. Those with the greatest amount of flavonoid intake performed better at both the beginning and end of the study compared with those with the lowest intake. While the lowest flavonoid group scored 2.1 points lower compared with their baseline, the Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1007 AIMONE ET AL. highest flavonoid group scored 1.2 points lower on the Mini-mental status examination (MMSE), suggesting that flavonoid intake positively correlated with cognitive function (189). However, not all diets with high flavonoid content attenuate the cognitive decline associated with increasing age. Over a 5-yr study, subjects aged 65 or older who adhered to a more Mediterranean diet high in fruits and vegetable consumption exhibited a slower decline as assessed by the MMSE but not by other cognitive measures (102). In human studies, there are countless variables that cannot be controlled from one individual to the next, which makes it extremely difficult to determine the most influential factors in improved cognitive functions. As mentioned above, lifestyle choices, including physical and mental exercises, have a tremendous impact on cognitive performance and can vary extensively in these subjects. Additionally, individuals in these studies not only consumed different amounts of flavonoids but also from a variety of sources. While flavonoid intake is self-reported from an individual’s diet, the person is typically unaware of how the processing of food alters the quantity of flavonoids in a given vegetable or product, which makes it difficult to determine how much of which flavonoids had the greatest impact on cognition. berry-supplemented aged rats compared with control aged rats (62). Enhanced cell proliferation may be mediated by the increase in IGF-I signaling, which is a known regulator of neurogenesis, as a result of the blueberry diet (62). While this study also used a blueberry supplement in lieu of a specific flavonoid, the findings were of particular interest because they linked the effects of blueberries on cognition to hippocampal neurogenesis, identifying possible mechanisms such as neurogenesis and growth factor signaling through which blueberries modulate cognitive function. IGF-I and other growth factors activate signaling cascades such as MAPK and CREB, which are involved fundamentally in learning and memory processes. Hippocampal levels of CREB phosphorylation and downstream target genes such as BDNF and Bcl-2 decreased with age but could be rescued with administration of green tea catechins in aged mice (192). Similar age-related declines in hippocampal levels of phospho-ERK1/2 and phosphorylated Akt were reversed in aged rats on a blueberry-supplemented diet (359). These studies reveal potential signaling pathways and mechanisms that may be crucial in enhancing cognitive function and synaptic plasticity by the flavonoids and/or other compounds found in the diets. 5. Learning Human studies are complicated by nature and, while the majority of studies suggest flavonoids provide positive benefits to cognitive functions, it remains unclear which particular aspect of cognition is most improved with flavonoid consumption (208). Studies using rodents on specific flavonoid-rich diets report enhanced learning and memory. Changes in cognitive function are apparent in aged mice and their ability to complete hippocampus-dependent cognitive tasks efficiently. With age, hippocampal neurogenesis declines dramatically, and this decline is accompanied by a slower learning curve in the MWM task as the aged mice take longer to swim to the hidden platform compared with young mice (344). The effectiveness of supplementing the diet with flavonoid-rich extracts from berries or tea is evident in a number of motor and cognitive tasks in aged animals. For example, aged rats who received grape juice (10 –50%) for 6 – 8 wk exhibited improved performance on behavioral tests, including the hippocampus-dependent MWM, rod walking, and accelerating rotarod, all tasks that are normally impaired in aged rats (309). Consistent with this finding, diets supplemented with blueberry (1.86%), spinach (0.91%), or strawberry (1.48%) extract reversed the cognitive and motor deficits associated with age, as assessed by a working memory water maze paradigm and rod walk in aged rats (146). Aged rats on a blueberrysupplemented (2%) diet for 8 wk made fewer errors on a radial arm water maze task than those on a control diet. This improved performance corresponded to an increase in the number of newborn cells in the hippocampus of blue- 1008 Interestingly, what was initially one of the more controversial relationships between behavior and neurogenesis is the effect of learning on new neurons (FIGURE 4). While there is considerable research into the effects of new neurons on hippocampus-dependent learning (see sect. V), studies of the effects of learning itself on the proliferation and survival of new neurons initially showed mixed results (122, 340). This initial confusion likely stems from two things: 1) not all learning paradigms and tasks are equivalent, and 2) the effect of learning appears to be temporally complex, with learning promoting the survival of some young neurons while suppressing the survival of earlier-born neurons (87). Notably, the training alone does not appear sufficient; in a study of aged rats, only the subset of animals that learned the task showed the aforementioned effects on survival (89). Subsequent studies have illustrated that the effects of learning on neurogenesis are quite complex. Interestingly, the negative effects of learning on survival noted above appear to be necessary for performance on the task, suggesting the relationship between learning and survival is acutely important (90). Learning affects not only the survival of cells but also the maturation of newborn neurons, with training in MWM increasing the complexity of dendritic arborization and the spine density of adult-born GCs (187, 337). Indeed, this learning effect appears to influence what the neurons will eventually respond to (155, 319, 325). Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS V. FUNCTION OF NEUROGENESIS Despite its extensive characterization, the functional importance of adult neurogenesis, particularly in humans, remains a big question for the broader community. Although the existence of adult neurogenesis was generally accepted by the mid 1990s, the appreciation that adult-born neurons had a meaningful role, as opposed to being simply a functionless evolutionary artifact (a “neural appendix” of sorts), has taken much longer. Presently, the question is not whether neurogenesis is relevant for cognition in rodents; rather, the debate has shifted towards the scope of its function (e.g., is neurogenesis function equivalent to DG function?) and its relevance in primates. This sharp transition from skepticism about its relevance to ascribing it a key role in memory formation has arisen due to several factors, including theoretical and computational examinations of how new neurons would relate to the well-characterized DG network, focused behavioral studies designed to test clear hypotheses on function, and sophisticated knockdown approaches that can specifically and sometimes reversibly impair neurogenesis. Previous reviews have exhaustively described the numerous behavioral studies that have targeted neurogenesis (82). Here, we will take a different approach by explaining the major functional hypotheses that are currently being con- A sidered by the community, highlighting both the theoretical and computational justification for the hypotheses as well as the associated supporting behavioral evidence (5, 252) prior to describing the broader approaches to characterizing neurogenesis knockdown behavior. Notably, the following hypotheses are not exclusive of one another, because the unique time scales of neurogenesis maturation may allow new neurons to have different impacts on memory at different stages of their development. A. Hypothesis 1: Role for Adult Neurogenesis in Pattern Separation The primary function that has been most associated with adult neurogenesis is pattern separation (4, 285) (FIGURE 5). Broadly, pattern separation can be thought of as a network process whose outputs are less similar, or less overlapping to one another, than its inputs. 1. What is pattern separation? The concept of “pattern separation” has been used somewhat differently in different domains (i.e., computational, behavioral, and physiology), so it helps to define the concept more directly as it relates to the DG. Broadly speaking, a computational pattern separation function can be attribHypothesized Mechanisms B New learning directed to new neurons A B Higher DG activity without new neurons A Different Increased interference Same B FIGURE 5. Pattern separation theory for the neurogenic dentate gyrus. Left: illustration of the cognitive phenomenon of pattern separation. Two events, consisting of highly similar objects and configurations, can be learned to be different if neurogenesis is present in the DG, whereas without neurogenesis the memories will be the same. Right: potential mechanisms for how neurogenesis may improve pattern separation. Top right: having new neurons available can permit the second event to utilize new neurons instead of the same old neurons to encode the differences in contexts. Bottom right: an alternative mechanism is that reducing neurogenesis increases the baseline activity of mature granule cells, leading to higher statistical overlap (and thus interference) between representations. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1009 AIMONE ET AL. uted to almost any neural network, as individual neurons respond in different ways to different possible inputs. Behaviorally, pattern separation is more typically linked to discrimination tasks, which are straightforward to design metrics for, but are often difficult to attribute to specific regions. Electrophysiology studies of pattern separation are often in between these definitions; they can investigate the correlation of neuronal activity within a region but, because it is remains difficult to measure large populations simultaneously, one cannot fully investigate the computational mechanisms underlying the separation. The computational pattern separation function that has been typically associated with the DG is more specifically related to the hippocampus’ function in memory formation. Information provided by the cortex (which presumably is in the form of high level event and spatial representations) to the hippocampus is potentially highly correlated: two different contexts may be composed of many of the same objects and spatial features yet ideally should be stored as distinct memories. This decorrelation of cortical inputs to optimally form memories has been attributed to the DG for several reasons (223, 255, 335, 336). First, the large number of GCs relative to the principal cell numbers of the entorhinal cortex (EC) and CA3 is similar to the machine learning technique of projecting information into a higher dimensional space to better classify or separate it (e.g., support vector machines; Refs. 23, 82). Having more neurons available overall allows fewer neurons to be used to represent the same information, and sparse codes are ideally suited for reducing overlap (which can equate to interference) between DG outputs. Second, the high tonic inhibition arising from both feed-forward and feedback inhibitory populations and related low activity of GCs in vivo are well suited to achieve this sparse coding, supporting the idea that this form of separation is indeed used by the DG (147, 190). Finally, the sparsely projecting yet powerful mossy fiber output synapses of GCs are potentially well suited for utilizing a sparse, separated code during memory formation (135). The hypothesis that the DG as a whole has a pattern separation role has been further validated by behavioral studies in mice and rats, in which DG lesions impair spatial discrimination (116, 161, 222, 234), and in human imaging studies, in which the CA3/DG area appears to be particularly involved in the discrimination of similar visual objects (24). Not surprisingly, given this long-held view of the DG, there are numerous computational models that have examined the role of new neurons in pattern separation (29, 355, 361). It is worth noting that these models have typically not examined the role of new neurons in the aforementioned rationale for the DG in pattern separation (i.e., why a highly divergent, sparsely active network needs new neurons). Rather, in these models, neurogenesis reduces interference by directing new learning towards new neurons (a common 1010 theme through almost all neurogenesis models; Ref. 5), thus ensuring that new memories and old memories are encoded distinctly. In some respect, while this function can be related to the pattern separation theory, the mechanism somewhat contrasts with the fairly passive, classic DG pattern separation motif. One possibility is that these two views of pattern separation can be unified if the activation of “classic” separating mature neurons and directed learning on immature neurons were distinct in the timing regime (i.e., when they fire is as important as what they fire to) (276). 2. Evidence for a neurogenesis pattern separation function While the computational mechanisms explaining a neurogenesis/pattern separation link are not fully understood, behavioral data using different neurogenesis knockdown approaches have provided strong evidence of a connection between neurogenesis and tasks involving spatial discrimination. Several spatial discrimination tasks have been used, including the radial-arm maze (67, 291), fear context discrimination (244, 284), and the two-choice discrimination task (67, 73, 224). This latter task has become one of the more prominent tasks to examine spatial discrimination, as it uses a touchscreen operant chamber that permits a relatively large number of observations and typically shows that the discrimination of proximal (in space) choices is selectively impaired without neurogenesis. B. Hypothesis 2: Role for Adult Neurogenesis in Encoding Temporal Context While the pattern separation function above has a long history in the DG, a related potential function for new neurons represents a new perspective on DG and hippocampal memory formation. As mentioned previously, one observation of almost all computational models of adult neurogenesis is that new neurons are the primary recipients of learning (5). From one perspective, driving learning to young neurons can improve pattern separation by ensuring that new memories and old memories do not interfere with one another (361). However, this direct pattern separation role of new neurons is most evident when the process is considered from a temporal perspective. Specifically, although memories formed at different times would be separated because different populations of young neurons were involved, this separation would not apply for memories for novel events experienced at or around the same time (FIGURE 6). Notably, unlike pattern separation, the role of new neurons in encoding time is an added function of neurogenesis that is distinct from prior hypotheses of DG function. This potential role was arrived at independently by two groups using slightly different theoretical rationales and has been subse- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS Hypothesized Mechanism Time Overlapping young neurons form temporal association Different young neuron population creates temporal separation FIGURE 6. Temporal coding theory for the neurogenic dentate gyrus. Left: illustration of the cognitive phenomenon of temporal episodic coding. Two events with very little similarity occur within a few days of each other (an unexpected “visitor” coming and buying a new truck). The temporal proximity of these events is sufficient to lead to a relationship in their long-term episodic memories. In contrast, another event several months later (a major snow storm) would not be associated due to the time elapsed. Right: mechanism for temporal coding by new neurons. The two temporally close events would activate separate populations of mature granule cells (pattern separation) but would use overlapping populations of immature neurons (pattern integration). This overlap is time dependent; as time passes, the immature neuron population matures and another set of young neurons appears to encode future events. This effectively increases separation for events far apart in time. quently observed in several computational studies (6, 7, 30). In both cases, young neurons have been shown to be better suited to encode novel information than mature neurons, which have already been configured to encode familiar information. 1. Evidence for temporal separation and neurogenesis The big challenge for the temporal separation theory for neurogenesis is the lack of clear behavioral evidence for such a function. Perhaps due to its status as a novel type of memory, there are no direct animal behavior tests for this function similar to the spatial discrimination tasks that have been used to argue for a general pattern separation role. Nonetheless, there is some evidence that tasks that require integration of events that occur close in time require neurogenesis. For example, the hippocampus has long been known to be required for trace conditioning, a form of fear conditioning that uses a time delay between the conditioned stimulus (i.e., tone) and the unconditioned stimulus (i.e., eyepuff), and this task was one of the first behaviors for which a neurogenesis dependence was observed (307, 308). However, although a temporal component can be attributed to this and other tasks, it is important to note that these studies were not designed to examine the relationship of neurogenesis to temporal coding but rather to hippocampal memory in general. Notably, Morris et al. (235) have recently reported a time-dependent task that was motivated by this neurogenesis hypothesis. While they did not ablate new neurons, they observed that the DG as a whole is necessary for temporal associations over longer time scales (235). It remains to be seen if other specific tasks that investigate the role of time in behavior directly can be developed and if these tasks depend on neurogenesis. C. Hypothesis 3: Role for Adult Neurogenesis in Memory Resolution A somewhat different perspective on neurogenesis function relates to what new neurons encode as opposed to how new neurons encode (FIGURE 7). In addition to its pattern separation function, the other primary function attributed to the DG is referred to as conjunctive encoding (161, 255). Basi- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1011 AIMONE ET AL. Hypothesized Mechanism Mature neurons are sufficient to encode familiar features …but not novel features Young neurons are capable of encoding all inputs High resolution memory Low resolution memory FIGURE 7. Memory resolution theory for the neurogenic dentate gyrus. Left: illustration of different resolution memories. All events that are remembered have a number of features that are not encoded. In a “high-resolution” memory, more details about the event are remembered; while the memory may not be perfect, it is much higher fidelity. In a “low-resolution” memory, less about the original event is encoded within memory, perhaps focusing on a lower number of features or coarse representations (“tree” instead of “maple tree”). Right: mechanism for increasing memory resolution by neurogenesis. Mature neurons and young neurons encode different types of information; mature neurons are powerful at representing what they have encoded in the past, whereas young neurons are capable of encoding novel events. The combination of both populations facilitates the encoding of more features robustly within memories. cally, conjunctive encoding simply refers to the DG’s (and downstream hippocampus’s) integration of spatial and nonspatial information into memories. Rather than simply encoding place or objects, DG neurons likely encode a higher representation that combines the what and where (and potential when, see above) of experienced events for encoding into memory. The challenge of linking the varied forms of pattern separation described above, combined with the appreciation that the DG may be involved in a more complex role in memory formation, has led us to propose that the DG, aided in large part through its neurogenesis process, has a substantial role in dictating the “resolution” of memories that are encoded by the hippocampus (4, 347). The DG memory resolution hypothesis is as follows. First, the sparse representation of the DG population and strong inhibition suggest that mature GCs are highly selective to what they respond to (i.e., “tightly tuned”). In addition to making 1012 them excellent pattern separators, this high selectivity makes mature GCs very informative about those features that they respond to. When a mature neuron is activated, there is only a relatively sparse set of features that could have been responsible. As a result, when an event is encoded, the set of mature neurons activated provides a very “high-resolution” encoding of those familiar features that drove their activity. There is a notable downside to this high-resolution coding: if each mature GC only encodes a finite number of features, it stands to reason that there will be features that are not encoded well by any mature GCs. While there are a substantial number of GCs (in rodents the GC layer is one of the largest forebrain regions by number), it is unlikely that they can guarantee that any event that may be encountered will have mature GCs capable of encoding it. In contrast, young GCs are theoretically far less selective in response to the inputs. This lack of selectivity could be due Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS to several features, namely, their altered physiological properties (2, 97, 114, 232, 269) and differential excitatory and inhibitory connectivity (193, 371). Individually, this lack of selectivity reduces the information content of young neurons relative to their mature counterparts and, in an absolute sense, makes networks with young neurons less effective at pattern separation (6). However, as a population, the broad tuning of young neurons can form a powerful distributed code that can uniquely represent features, and because this code consists of low-information units, it is likely capable of representing any cortical input that may be provided. Alone, this combination of sparse (mature GCs) and distributed (immature GCs) coding schemes is a powerful approach to episodic memory encoding. However, improving the high-information sparse-coding representations over time is probably beneficial. Thus the memory resolution hypothesis predicts that low-information young neurons eventually become high-information mature neurons by virtue of their increased propensity for synaptic plasticity (115, 295). By focusing plasticity towards young neurons, the mature neurons can essentially fix their coding on familiar features indefinitely, with little interference over time. Notably, this directing of learning towards immature neurons to preserve the representations of mature neurons has been a key feature of numerous computational models of neurogenesis (5, 6, 13, 14, 361). 1. Evidence for a memory resolution function for neurogenesis The principal prediction of the memory resolution hypothesis is that mature neurons will be highly selective to what they respond to and that young neurons will be somewhat less discriminating when young, but that they will mature to represent events from their development. This prediction is difficult to validate directly due to the challenges associated with in vivo physiology in the DG and because there is no clear method to determine which features are salient to an animal at a given time. However, there have been numerous studies using immediate early gene (IEG) assessments of DG activity during experiences in EEs and behavioral tasks (9, 83, 155, 275, 319, 325) and of CA3 following neurogenesis disruption (248). The results from these IEG studies have been somewhat mixed and at times difficult to interpret for a number of reasons (3). In summary, it appears that young neurons do learn to encode what they are exposed to when young; however, the overall activity of both the young and mature populations is quite low and it is unclear how representative the IEG-labeled population is of the active population. From a behavioral level, the memory resolution hypothesis would suggest that animals without neurogenesis should show little effects on memory tasks that rely principally on familiar features; however, tasks that involve the flexible use of novel information would be predicted to suffer. There have not been many attempts to break down behavioral tasks in this manner while investigating neurogenesis; however, one recent study used diptheria toxin (DT) to ablate adult-born neurons that were thought of as previously specializing to the trained event, showing that the loss of already trained adult-born neurons did impair subsequent performance on those tasks (16). 2. Effect of neurogenesis reduction on behavior Given that the neurogenesis occurs in the hippocampus, a critical brain region for declarative memory in humans, it has long been thought that adult-born neurons should contribute to hippocampus-dependent learning and memory. With the use of methylazoxymethanol acetate (MAM), a DNA methylating agent, to ablate neurogenesis, it was first shown that hippocampal neurogenesis was critical for the formation of hippocampus-dependent association memories using a trace version of a fear conditioning protocol and was dispensable for hippocampus-independent memory using a delayed version of a fear conditioning protocol (307). However, later studies of the consequences of adult neurogenesis ablation using irradiation or other nonspecific methods showed rather inconsistent data, possibly due to the nonspecific nature of neurogenesis ablating approaches, the behavioral assays used, and variations in experimental conditions from study to study (82). The latest studies using more specific methods for neurogenesis manipulation and behavioral tasks aimed at DG-related functions have not only established a role for adult-born neurons in hippocampus-dependent learning and memory but also revealed several key functional features of adult neurogenesis (82). First, as described above, adult neurogenesis appears to be required for pattern separation, which is currently described as a key function of the DG. This critical role for adult neurogenesis was revealed by using tasks that tested animals’ ability to discriminate between closely located spatial positions or similar contexts (67, 73, 284). Recent molecular and physiological studies demonstrated that the DG can achieve pattern separation by using distinct population codes and different firing patterns to represent different environmental inputs (83, 190). It will be interesting to test whether such an event-specific activity pattern is affected by altering the rate of adult neurogenesis. In addition, tasks related to pattern separation have been noted to improve in response to genetic and behavioral manipulations that increase neurogenesis (73, 284). Second, adult neurogenesis is more critically involved in memory discrimination at the time of memory recall. Mice with adult-born neurons ablated after the acquisition of contextual or spatial memories showed an impaired behavioral performance upon recall of those memories (16, 129). Using a contextual fear conditioning paradigm, Tonegawa and colleagues (199, 274) recently reported that optoge- Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1013 AIMONE ET AL. netic activation of GCs responding to a contextual exposure episode was sufficient to create an artificial fear memory to the exposed context and that artificially activating those GCs subsequent to the fear memory formation was sufficient to induce a fear response in a neutral environment. Further studies are needed to determine whether adult-born GCs are involved in forming false memories. Third, adult neurogenesis is also required for the long-term consolidation of contextual memory. After the formation of the contextual fear memory, the recall of fear memory is initially dependent on the hippocampus but becomes independent of the hippocampus after several weeks (11). Kitamura et al. (164) showed that ablation of neurogenesis by irradiation prolonged the hippocampus dependency of contextual fear memory in mice, suggesting a role for neurogenesis in promoting system memory consolidation. Given that the hippocampus is considered to be a temporary storage site for memory, neurogenesis may serve as a mechanism for forgetting in the hippocampus through general retroactive interference, regardless of memory content (107). Fourth, adult-born neurons at different maturation stages may make different contributions to learning and memory, given the differences in their physiological characteristics. By reducing the number of adult-born DGCs at different maturation stages in Nestin-tk transgenic mice, it was found that immature adult-born DGCs but not their mature counterparts were required for learning and memory (85). Furthermore, specifically silencing the adult-born DGCs at 4 wk of age but not their 2- or 8-wk-old counterparts by optogenetic techniques after task acquisition resulted in impaired memory recall (129). Finally, blocking the output of both developmentally born and adult-born mature DGCs did not impair contextual discrimination in mice but caused a deficit in the recall of previously formed spatial or contextual memories when presented with an incomplete set of cues (244). These results led Tonegawa and colleagues (244) to propose that immature GCs and mature GCs were responsible for pattern separation and pattern completion, respectively. However, this hypothesis is challenged by the observations that ablation or inhibition of young adultborn GCs impaired spatial and contextual memory recall and by the finding that optogenetically inhibiting GC activity did not affect the recall of contextual fear memory (16, 129, 162). Future studies are needed to resolve this disparity. In addition to its role in learning and memory, the hippocampus has been implicated in emotional regulation. Instead of a unitary structure, the functional role of the hippocampus varies along its dorsoventral axis, with the dorsal and ventral parts involved in cognitive and emotional functions, respectively (26, 101). This hypothesis was supported by both differential connections between the hippocampus and other brain regions and distinct lesion effects, as well as 1014 a recent optogenetic characterization along the dorsoventral axis (162, 236, 322). Early studies of hippocampal neurogenesis also revealed a correlation between the rate of neurogenesis and emotional status, with increased neurogenesis being observed in antidepressant-treated animals and decreased neurogenesis in stressed animals, as discussed above. It has therefore been proposed that adult hippocampal neurogenesis could also be an important affective regulator (142). Although initial correlative studies provided experimental evidence for this hypothesis, subsequent studies reported data inconsistent with this hypothesis, suggesting that there is a rather complicated relationship between hippocampal neurogenesis and affective states in animal models. For example, the increase in NPC proliferation observed with antidepressants is dependent on the strain of mice studied: antidepressants only enhance NPC proliferation in 129SvEv and DBA mice but not in C56BL/6, balb/c, or A/J mice (78, 138, 227, 289). Furthermore, stress does not always cause a reduction in neurogenesis. It was recently reported that, instead of inhibiting neurogenesis, acute stress actually increased proliferation of NPCs in the dorsal hippocampus, leading to enhanced activity in newborn GCs and an improvement in fear extinction (163). Studies examining the causal relationship between hippocampal neurogenesis and affective regulation also suggest that hippocampal neurogenesis is not a master regulator of emotional status in rodents; instead, neurogenesis may influence the emotional circuitry indirectly. In the first studies addressing the causality of neurogenesis and affective regulation, Santarelli et al. (289) reported that hippocampal neurogenesis was required for the effectiveness of fluoxetine (an antidepressant of the SSRI class) in certain tests for anxiety- and depression-like behaviors, such as the novelty suppressed feeding task and the splash test. However, subsequent studies showed that the dependence of antidepressant efficacy on hippocampal neurogenesis was not found in other tests for anxiety- and depression-like behaviors, such as the open field test or forced swimming test (78). In addition, hippocampal neurogenesis was only involved in the efficacy of limited classes of antidepressants (267), probably because anxiety and depression have complex etiologies and different classes of antidepressants may have different therapeutic targets. A recent study showed that hippocampal neurogenesis also played a role in stress adaptation, which in turn could influence anxiety- and depression-like behavior in a distinctive way in different, commonly used behavioral tasks (312). In summary, the relationship between hippocampal neurogenesis and emotional regulation is rather complicated. Because of the heterogeneous nature of depression and poorly understood neural networks underlying the disease, it is difficult to pinpoint the exact mechanism for adult-born Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS GCs in affective regulation. It is possible that adult-born GCs are part of brain networks that underlie certain aspects of emotional disorders and are affected by certain antidepressants, making hippocampal neurogenesis indirectly involved in emotional regulation. For example, as discussed above, hippocampal neurogenesis is thought to be an important factor in pattern separation, so impaired neurogenesis may result in overgeneralization to emotional stimuli, which are found in certain types of anxiety disorders. Indeed, a recent study showed that there was a negative correlation between performance in a pattern separation task and the score received on the Depression Anxiety Stress Scales (302). Hence, it could be the hippocampal neurogenesis-mediated cognitive function that plays a role in affective regulation. 3. Relationship of neurogenesis and cognitive deficits in humans Given the challenges associated with directly measuring neurogenesis in humans, the role of new neurons in human cognitive disorders remains unknown. Nonetheless, the lower neurogenesis levels typically seen in aged or diseased mice suggest that human neurogenesis may be potentially lower in psychiatric and neurological conditions and, in turn, linked to impaired cognition. For instance, both stress and aging, which have dramatic effects on neurogenesis levels in mice, are well known to be associated with memory impairments in clinical human populations (40, 205). Likewise, patients who have undergone radiation therapy for brain tumors are known to experience cognitive and memory deficits. While radiation clearly has broad effects on neural tissue, it is notable that radiation remains one of the most potent techniques for experimentally ablating neurogenesis and that the observed cognitive deficits are consistent with functions linked to neurogenesis (270, 279). Despite the lack of direct evidence, there are several studies that have sought to link cognitive functions related to neurogenesis in rodents to human clinical conditions. In particular, the pattern separation function described above has been measured in humans and observed to be impaired in aged individuals. Furthermore, this age-dependent decline of function can be correlated with a regional fMRI effect specific to the DG/CA3 area (317, 366). In all, the observations that human cognition correlates with neurogenesisrelated behaviors support the contention that activities and interventions that improve neurogenesis may be useful strategies to alleviate cognitive deficits in humans. VI. TECHNOLOGY FOR STUDYING THE FUNCTION OF ADULT NEUROGENESIS As adult neurogenesis has become an increasingly well-accepted concept over the past couple of decades, focus has been directed toward developing new technologies to inves- tigate both the NSC development processes and the function of adult neurogenesis. There is also increasing interest in developing techniques to manipulate the levels of neurogenesis for both scientific and clinical purposes. Below we discuss the development of these new techniques designed to study adult neurogenesis. A. Regulating the Number of Adult-Born Neurons Adult hippocampal neurogenesis can be influenced by many factors, such as the genetic background of the animal, the age of the animal (185), mood (283), and housing environment (341). The number of adult-born neurons can be regulated at the levels of cell proliferation, differentiation, and survival. Recently, various strategies have been developed to disrupt or augment neurogenesis specifically in the hippocampus. These methods provide spatial and cellular specificity to manipulate neurogenesis in the adult brain. The Herpes simplex virus type 1 thymidine kinase (HSV-1 TK) is a phosphotransferase that phosphorylates a broad range of pyrimidine nucleosides and purine nucleosides, including thymidine, deoxycytidine, and ganciclovir (GCV), which is a synthetic analog of 2’-deoxy-guanosine (65). Phosphorylated GCV is further metabolized to its triphosphate, GCVTP, and incorporates into the nascent DNA, which induces chromosome breaks and sister chromatid exchanges (329) and thus causes cell apoptosis (280, 329). Combining GCV with a genetically expressed HSV-1 TK in adult-born neurons by use of a specific promoter such as GFAP and nestin (226, 363) has been used to specifically ablate adult-born neurons (85, 290). DT is another toxin that is widely used for lineage-specific ablation (265). DT is synthesized by Corynebacterium dipheriae (220). It has been adapted for expression in mammalian cells (220), and a single molecule might be sufficient to kill a cell. DT is a secretory precursor polypeptide and can be cleaved into two fragments, A (DTA) and B (DTB) chains, joined by a disulfide bridge. DTB binds to the receptor on the cell surface and is absorbed into the cells via endocytosis. Once DTA is transported into the cytoplasm, it catalyzes the transfer of ADP-ribose moiety of nicotinamide adenine dinucleotide (NAD⫹) to a modified histidine residue on elongation factor 2, resulting in termination of protein synthesis (68) and subsequent cell death. Ideally, the delivery of DT into NPCs by retrovirus could specifically ablate newborn neurons in the adult brain. Since mice and rats are naturally resistant to DT, the genetic expression of an identified monkey DT receptor (a membrane-anchored form of the heparin-binding EGF-like growth factor) (241) in NPCs in these species provides another approach that ablates newborn neurons in the adult brain induced by DT (47, 287). Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1015 AIMONE ET AL. Only a fraction of the cells generated in the adult brain survives more than several months because the majority (⬃60 – 80% in standard housing) undergoes apoptosis (54, 79). Therefore, antiapoptotic techniques can potentially be used to increase neurogenesis. A recent example of this approach is the use of a mouse line with a genetic knockout of the proapoptotic gene Bax in NPCs to selectivity increase adult neurogenesis (284). B. Mapping the Circuit of Adult-Born Neurons To date, most of the anatomical characterization of how new neurons physically connect to the existing circuitry has relied on circumstantial evidence from electrophysiology and the gross morphology of the neurons (97, 179, 214, 371). Developing more advanced techniques for dissecting the circuit of adult-born neurons is potentially very critical for fully understanding their integration and ultimately their function. A genetically modified rabies virus was developed recently to trace the monosynaptic input of particular cell types (357). It has been successfully used to map the circuits of the piriform cortical neurons (132, 230), amygdala (132), retinal ganglion cells (318), and motor neurons (368). Unlike traditional tracers, this genetically modified rabies virus is able to target specific cell types and is subsequently retrogradely transported to presynaptic neurons only across one synapse, thereby labeling direct connections onto the initially infected cells (51). Two modifications are made to generate this genetically modified rabies virus. 1) Rabies glycoprotein is necessary for a virus to retrogradely transport from target neurons to their presynaptic partners. Replacing rabies glycoprotein with fluorescent protein in a virus genome allows visualization of the presynaptic neurons and prevents the rabies virus from jumping across multiple synapses. 2) Pseudotyping rabies virus with EnvA then allows the rabies virus to target a specific cell type. EnvA is an avian sarcoma and leukosis viral envelope protein and can be used to guide virus infection specifically into cells that express the TVA viral receptor. Because TVA is only found in birds and not in mammals, introducing TVA into host cells in mice restricts EnvA pseudotyped rabies virus infection to only the particular cell type. The use of this technique to map the circuit of newborn neurons in adult hippocampus requires both the genetically modified rabies virus and help from the aforementioned techniques to target young neurons. To get rabies glycoprotein and TVA to be specifically expressed in newborn neurons, the Moloney murine leukemia retrovirus can be used to specifically target genes to nonquiescent NSCs, NPCs, and their progeny. Therefore, combining a retrovirus and a 1016 genetically modified rabies virus is an ideal tool to study the monosynaptic circuit of adult-born neurons (86, 195, 349). C. Imaging the Activity of Adult-Born Neurons Ultimately, fully understanding the function of immature neurons will likely require an ability to observe the activity of young neurons in the context of the functioning DG circuit during behavior. A recent technique gaining widespread use in rodents is large-scale calcium imaging of the neural circuits. Combining optical microscopy with a fluorescent [Ca2⫹] indicator can be used to record activity of population of neurons. Because action potentials trigger Ca2⫹ influx through voltage-gated calcium channels, changes of free [Ca2⫹] can be used as a reliable readout for neural activity. The most popular techniques to qualitatively measure [Ca2⫹] have been the use of either synthetic dyes or proteins that change their intensity of fluorescence upon calcium binding (212). Chemical indicators are often a derivative of BAPTA, an EGTA homolog (338). They have high selectivity for calcium, and their fluorescence significantly increases in response to binding calcium (339). Membrane-permeable indicators (369) or dextran-conjugated indicators (240) such as Oregon Green BAPTA-1 (OGB-1) and fluo 4 have been widely used in functional imaging in vivo. This technique has been used in slice studies of the DG (213). In one study, the DG was bathed with a calcium indicator and retroviruslabeled, adult-born, and embryonically born neurons could then be measured in the same experimental condition in vitro (213). Genetically encoded calcium indicators (GECIs) overcome several of the primary limitations of chemical indicators, such as cell type specificity. GECIs can be targeted to specific neuronal populations and are able to be stably expressed in cells over months. GECIs are based on fusions of fluorescent proteins and calcium buffers, such as calmodulin (CaM) (242) or troponin C (TnC) (211). When calcium binds to CaM/TnC, conformational changes induce changes in fluorescence intensity (242). Long-term expression of GECIs in adult-born neurons by a retrovirus or progenitor-specific promoter would allow the imaging of the activity of newborn neurons in behaving animals in vivo over long periods of time and would greatly facilitate the demonstration of what adult-born neurons encode. D. Controlling the Activity of Adult-Born Neurons Manipulating the activity of newborn neurons in the adult brain is required to further understand the function of adult neurogenesis. Inactivating or activating newborn neurons Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS by genetics or viral vector using optogenetic strategies (103) has the potential to directly investigate the impact of adultborn neurons on the local circuit. The most common optogenetic tools are Chlamydomonas reinhardtii channelrhodopsin-2 (ChR2) and Natronomonas pharaonis halorhodopsin (NpHR), which are microbial light-sensitive proteins. ChR2 is a cation channel that mainly allows Na⫹ to enter the cell and activate cells following exposure to blue light (479 nm). NpHR is a chloride pump that activates upon illumination with 580 nm yellow light and causes inhibition of neurons. The millisecond temporal scale of optogenetics allows control of neuronal activity in real time. The introduction of ChR2 to adult-born neurons by retrovirus has shown that newborn neurons generated in the adult brain are able to form functional synapses with their postsynaptic target CA3 pyramidal neurons (330). Direct activation using ChR2 of a group of neurons in the DG that are activated during fear conditioning has been shown to be sufficient to mediate the fear memory recall (200). It will be interesting to directly activate or inhibit newborn neurons by optogenetics during learning. In addition to optogenetics, other methods can be used to control neuronal activity. Tetanus toxin (TeTx) is a neurotoxin produced by the bacteria Clostridium tetani (165). TeTx suppresses neurotransmitter release by cleaving the synaptic vesicle-associated membrane protein VAMP2/synaptobrevin2 (294). Therefore, expressing TeTx in neurons will inhibit their output and inactivate their function. A similar method involves the use of the Drosophila allatostatin (AL) receptor (AlstR), which is a G protein-coupled inward rectifier K⫹ channel (39). Applying AL to AlstRexpressing neurons blocks generation of action potentials (181). Inducible and rapidly reversible AL-dependent silencing of a cell has been demonstrated in cortex and spinal cord in the anesthetized rat, mouse, and monkey (119, 323, 324). Finally, an engineered muscarinic receptor, hM3Dq, is specifically activated by a synthetic ligand, clozapine-Noxide (CNO), but not by its endogenous ligand, acetylcholine (15). CNO activates Gq signaling pathway in hippocampal neurons and induces burst firing (8, 113). E. Future of Neurogenesis Technology and Research While current technology has provided us many insights into how these new neurons arise and what they do, there is still much that we need to learn about these new cells. Ultimately, understanding the cellular nature of the neurogenesis process from NSC to GC as well the functions of adult-born neurons is going to require a combination of many of these approaches. For instance, externally regulatable genetic approaches manipulating how progenitors respond to their local environment and external cues will reveal much about the proliferation and differentiation processes. In short, while the technological advances of the last couple of decades have revealed most of what we know about adult neurogenesis, we may yet learn that we still know very little. ACKNOWLEDGMENTS We thank Mary Lynn Gage for editorial comments and assistance. Current address of G. D. Clemenson: Dept. of Neurobiology and Behavior, Univ. of California Irvine, Irvine, CA 92697. Address for reprint requests and other correspondence: F. H. Gage, Laboratory of Genetics, Salk Institute for Biological Studies, La Jolla, CA 92037 (e-mail: gage@salk. edu). GRANTS This work was supported by the James S. McDonnell Foundation, JPB Foundation, Mather’s Foundation, Glenn Center of Aging, and National Institutes of Health Grant R01MH090258-04. J. B. Aimone is supported by Sandia National Laboratories’ Laboratory Directed Research and Development (LDRD) program. Sandia National Laboratories is a multiprogram laboratory managed and operated by Sandia Corporation, a wholly owned subsidiary of Lockheed Martin Corporation, for the United States Department of Energy’s National Nuclear Security Administration under Contract DE-AC04-94AL85000. DISCLOSURES No conflicts of interest, financial or otherwise, are declared by the authors. REFERENCES 1. Abrous DN, Adriani W, Montaron MF, Aurousseau C, Rougon G, Le Moal M, Piazza PV. Nicotine self-administration impairs hippocampal plasticity. J Neurosci 22: 3656 – 3662, 2002. 2. Aimone JB, Deng W, Gage FH. Adult neurogenesis: integrating theories and separating functions. Trends Cogn Sci 14: 325–337, 2010. 3. Aimone JB, Deng W, Gage FH. Put them out to pasture? What are old granule cells good for, anyway? Hippocampus 20: 1124 –1125, 2010. 4. Aimone JB, Deng W, Gage FH. Resolving new memories: a critical look at the dentate gyrus, adult neurogenesis, and pattern separation. Neuron 70: 589 –596, 2011. 5. Aimone JB, Gage FH. Modeling new neuron function: a history of using computational neuroscience to study adult neurogenesis. Eur J Neurosci 33: 1160 –1169, 2011. 6. Aimone JB, Wiles J, Gage FH. Computational influence of adult neurogenesis on memory encoding. Neuron 61: 187–202, 2009. 7. Aimone JB, Wiles J, Gage FH. Potential role for adult neurogenesis in the encoding of time in new memories. Nat Neurosci 9: 723–727, 2006. 8. Alexander GM, Rogan SC, Abbas AI, Armbruster BN, Pei Y, Allen JA, Nonneman RJ, Hartmann J, Moy SS, Nicolelis MA, McNamara JO, Roth BL. Remote control of Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1017 AIMONE ET AL. neuronal activity in transgenic mice expressing evolved G protein-coupled receptors. Neuron 63: 27–39, 2009. 31. Beecher GR. Overview of dietary flavonoids: nomenclature, occurrence and intake. J Nutr 133: 3248S–3254S, 2003. 9. Alme CB, Buzzetti RA, Marrone DF, Leutgeb JK, Chawla MK, Schaner MJ, Bohanick JD, Khoboko T, Leutgeb S, Moser EI, Moser MB, McNaughton BL, Barnes CA. Hippocampal granule cells opt for early retirement. Hippocampus 20: 1109 –1123, 2010. 32. Behar TN, Li YX, Tran HT, Ma W, Dunlap V, Scott C, Barker JL. GABA stimulates chemotaxis and chemokinesis of embryonic cortical neurons via calcium-dependent mechanisms. J Neurosci 16: 1808 –1818, 1996. 10. Altman J. Are new neurons formed in the brains of adult mammals? Science 135: 1127–1128, 1962. 33. Bekinschtein P, Cammarota M, Igaz LM, Bevilaqua LR, Izquierdo I, Medina JH. Persistence of long-term memory storage requires a late protein synthesis- and BDNFdependent phase in the hippocampus. Neuron 53: 261–277, 2007. 11. Anagnostaras SG, Maren S, Fanselow MS. Temporally graded retrograde amnesia of contextual fear after hippocampal damage in rats: within-subjects examination. J Neurosci 19: 1106 –1114, 1999. 12. Anderson BJ, Eckburg PB, Relucio KI. Alterations in the thickness of motor cortical subregions after motor-skill learning and exercise. Learn Mem 9: 1–9, 2002. 13. Appleby PA, Kempermann G, Wiskott L. The role of additive neurogenesis and synaptic plasticity in a hippocampal memory model with grid-cell like input. PLoS Computat Biol 7: e1001063, 2011. 14. Appleby PA, Wiskott L. Additive neurogenesis as a strategy for avoiding interference in a sparsely-coding dentate gyrus. Network 20: 137–161, 2009. 15. Armbruster BN, Li X, Pausch MH, Herlitze S, Roth BL. Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc Natl Acad Sci USA 104: 5163–5168, 2007. 16. Arruda-Carvalho M, Sakaguchi M, Akers KG, Josselyn SA, Frankland PW. Posttraining ablation of adult-generated neurons degrades previously acquired memories. J Neurosci 31: 15113–15127, 2011. 17. Arthur JS. MSK activation and physiological roles. Front Biosci 13: 5866 –5879, 2008. 18. Ashton RS, Conway A, Pangarkar C, Bergen J, Lim KI, Shah P, Bissell M, Schaffer DV. Astrocytes regulate adult hippocampal neurogenesis through ephrin-B signaling. Nat Neurosci 15: 1399 –1406, 2012. 19. Atack JR, Bayley PJ, Seabrook GR, Wafford KA, McKernan RM, Dawson GR. L-655,708 enhances cognition in rats but is not proconvulsant at a dose selective for alpha5-containing GABAA receptors. Neuropharmacology 51: 1023–1029, 2006. 20. Aztiria E, Capodieci G, Arancio L, Leanza G. Extensive training in a maze task reduces neurogenesis in the adult rat dentate gyrus probably as a result of stress. Neurosci Lett 416: 133–137, 2007. 21. Bachstetter AD, Morganti JM, Jernberg J, Schlunk A, Mitchell SH, Brewster KW, Hudson CE, Cole MJ, Harrison JK, Bickford PC. Fractalkine and CX3CR1 regulate hippocampal neurogenesis in adult and aged rats. Neurobiol Aging 32: 2030 –2044, 2011. 23. Baker JL. Is there a support vector machine hiding in the dentate gyrus? Neurocomputing 52: 199 –207, 2003. 24. Bakker A, Kirwan CB, Miller M, Stark CE. Pattern separation in the human hippocampal CA3 and dentate gyrus. Science 319: 1640 –1642, 2008. 25. Banasr M, Hery M, Printemps R, Daszuta A. Serotonin-induced increases in adult cell proliferation and neurogenesis are mediated through different and common 5-HT receptor subtypes in the dentate gyrus and the subventricular zone. Neuropsychopharmacology 29: 450 – 460, 2004. 26. Bannerman DM, Deacon RM, Offen S, Friswell J, Grubb M, Rawlins JN. Double dissociation of function within the hippocampus: spatial memory and hyponeophagia. Behav Neurosci 116: 884 –901, 2002. 27. Barkho BZ, Song H, Aimone JB, Smrt RD, Kuwabara T, Nakashima K, Gage FH, Zhao X. Identification of astrocyte-expressed factors that modulate neural stem/progenitor cell differentiation. Stem Cells Dev 15: 407– 421, 2006. 28. Battista D, Ferrari CC, Gage FH, Pitossi FJ. Neurogenic niche modulation by activated microglia: transforming growth factor beta increases neurogenesis in the adult dentate gyrus. Eur J Neurosci 23: 83–93, 2006. 34. Ben Abdallah NM, Slomianka L, Vyssotski AL, Lipp HP. Early age-related changes in adult hippocampal neurogenesis in C57 mice. Neurobiol Aging 31: 151–161, 2010. 35. Bergmann O, Liebl J, Bernard S, Alkass K, Yeung MS, Steier P, Kutschera W, Johnson L, Landen M, Druid H, Spalding KL, Frisen J. The age of olfactory bulb neurons in humans. Neuron 74: 634 – 639, 2012. 36. Bernal GM, Peterson DA. Phenotypic and gene expression modification with normal brain aging in GFAP-positive astrocytes and neural stem cells. Aging Cell 10: 466 – 482, 2011. 37. Beutler B. Inferences, questions and possibilities in Toll-like receptor signalling. Nature 430: 257–263, 2004. 38. Bimonte HA, Nelson ME, Granholm AC. Age-related deficits as working memory load increases: relationships with growth factors. Neurobiol Aging 24: 37– 48, 2003. 39. Birgul N, Weise C, Kreienkamp HJ, Richter D. Reverse physiology in Drosophila: identification of a novel allatostatin-like neuropeptide and its cognate receptor structurally related to the mammalian somatostatin/galanin/opioid receptor family. EMBO J 18: 5892–5900, 1999. 40. Bishop NA, Lu T, Yankner BA. Neural mechanisms of ageing and cognitive decline. Nature 464: 529 –535, 2010. 41. Bonaguidi MA, Song J, Ming GL, Song H. A unifying hypothesis on mammalian neural stem cell properties in the adult hippocampus. Curr Opin Neurobiol 22: 754 –761, 2012. 42. Bonaguidi MA, Wheeler MA, Shapiro JS, Stadel RP, Sun GJ, Ming GL, Song H. In vivo clonal analysis reveals self-renewing and multipotent adult neural stem cell characteristics. Cell 145: 1142–1155, 2011. 43. Bondolfi L, Ermini F, Long JM, Ingram DK, Jucker M. Impact of age and caloric restriction on neurogenesis in the dentate gyrus of C57BL/6 mice. Neurobiol Aging 25: 333–340, 2004. 44. Bonfanti L, Ponti G. Adult mammalian neurogenesis and the New Zealand white rabbit. Vet J 175: 310 –331, 2008. 45. Bovetti S, Bovolin P, Perroteau I, Puche AC. Subventricular zone-derived neuroblast migration to the olfactory bulb is modulated by matrix remodelling. Eur J Neurosci 25: 2021–2033, 2007. 46. Brunne B, Zhao S, Derouiche A, Herz J, May P, Frotscher M, Bock HH. Origin, maturation, and astroglial transformation of secondary radial glial cells in the developing dentate gyrus. Glia 58: 1553–1569, 2010. 47. Buch T, Heppner FL, Tertilt C, Heinen TJ, Kremer M, Wunderlich FT, Jung S, Waisman A. A Cre-inducible diphtheria toxin receptor mediates cell lineage ablation after toxin administration. Nat Methods 2: 419 – 426, 2005. 48. Buckwalter MS, Yamane M, Coleman BS, Ormerod BK, Chin JT, Palmer T, WyssCoray T. Chronically increased transforming growth factor-beta1 strongly inhibits hippocampal neurogenesis in aged mice. Am J Pathol 169: 154 –164, 2006. 49. Butovsky O, Ziv Y, Schwartz A, Landa G, Talpalar AE, Pluchino S, Martino G, Schwartz M. Microglia activated by IL-4 or IFN-gamma differentially induce neurogenesis and oligodendrogenesis from adult stem/progenitor cells. Mol Cell Neurosci 31: 149 –160, 2006. 29. Becker S. A computational principle for hippocampal learning and neurogenesis. Hippocampus 15: 722–738, 2005. 50. Cacci E, Ajmone-Cat MA, Anelli T, Biagioni S, Minghetti L. In vitro neuronal and glial differentiation from embryonic or adult neural precursor cells are differently affected by chronic or acute activation of microglia. Glia 56: 412– 425, 2008. 30. Becker S, Wojtowicz JM. A model of hippocampal neurogenesis in memory and mood disorders. Trends Cogn Sci 11: 70 –76, 2007. 51. Callaway EM. Transneuronal circuit tracing with neurotropic viruses. Curr Opin Neurobiol 18: 617– 623, 2008. 1018 Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS 52. Cameron HA, Dayer AG. New interneurons in the adult neocortex: small, sparse, but significant? Biol Psychiatry 63: 650 – 655, 2008. 73. Creer DJ, Romberg C, Saksida LM, van Praag H, Bussey TJ. Running enhances spatial pattern separation in mice. Proc Natl Acad Sci USA 107: 2367–2372, 2010. 53. Cameron HA, Gould E. Adult neurogenesis is regulated by adrenal steroids in the dentate gyrus. Neuroscience 61: 203–209, 1994. 74. Croll SD, Ip NY, Lindsay RM, Wiegand SJ. Expression of BDNF and trkB as a function of age and cognitive performance. Brain Res 812: 200 –208, 1998. 54. Cameron HA, McKay RD. Adult neurogenesis produces a large pool of new granule cells in the dentate gyrus. J Comp Neurol 435: 406 – 417, 2001. 75. Curtis MA, Kam M, Nannmark U, Anderson MF, Axell MZ, Wikkelso C, Holtas S, van Roon-Mom WM, Bjork-Eriksson T, Nordborg C, Frisen J, Dragunow M, Faull RL, Eriksson PS. Human neuroblasts migrate to the olfactory bulb via a lateral ventricular extension. Science 315: 1243–1249, 2007. 55. Campbell NR, Fernandes CC, Halff AW, Berg DK. Endogenous signaling through alpha7-containing nicotinic receptors promotes maturation and integration of adultborn neurons in the hippocampus. J Neurosci 30: 8734 – 8744, 2010. 56. Campuzano R, Barrios V, Cuevas B, Asin-Cardiel E, Muela A, Castro JM, FernandezAyerdi A, Cuevas P. Serum basic fibroblast growth factor levels in exercise-induced myocardial ischemia more likely a marker of endothelial dysfunction than a marker of ischemia? Eur J Med Res 7: 93–97, 2002. 57. Cao L, Jiao X, Zuzga DS, Liu Y, Fong DM, Young D, During MJ. VEGF links hippocampal activity with neurogenesis, learning and memory. Nat Genet 36: 827– 835, 2004. 58. Cao X, Li LP, Qin XH, Li SJ, Zhang M, Wang Q, Hu HH, Fang YY, Gao YB, Li XW, Sun LR, Xiong WC, Gao TM, Zhu XH. Astrocytic ATP release regulates the proliferation of neural stem cells in the adult hippocampus. Stem Cells 31: 1633–1643, 2013. 59. Cao X, Li LP, Wang Q, Wu Q, Hu HH, Zhang M, Fang YY, Zhang J, Li SJ, Xiong WC, Yan HC, Gao YB, Liu JH, Li XW, Sun LR, Zeng YN, Zhu XH, Gao TM. Astrocytederived ATP modulates depressive-like behaviors. Nature Med 19: 773–777, 2013. 60. Cardona AE, Pioro EP, Sasse ME, Kostenko V, Cardona SM, Dijkstra IM, Huang D, Kidd G, Dombrowski S, Dutta R, Lee JC, Cook DN, Jung S, Lira SA, Littman DR, Ransohoff RM. Control of microglial neurotoxicity by the fractalkine receptor. Nat Neurosci 9: 917–924, 2006. 76. Czeh B, Michaelis T, Watanabe T, Frahm J, de Biurrun G, van Kampen M, Bartolomucci A, Fuchs E. Stress-induced changes in cerebral metabolites, hippocampal volume, and cell proliferation are prevented by antidepressant treatment with tianeptine. Proc Natl Acad Sci USA 98: 12796 –12801, 2001. 77. Czeh B, Muller-Keuker JI, Rygula R, Abumaria N, Hiemke C, Domenici E, Fuchs E. Chronic social stress inhibits cell proliferation in the adult medial prefrontal cortex: hemispheric asymmetry and reversal by fluoxetine treatment. Neuropsychopharmacology 32: 1490 –1503, 2007. 78. David DJ, Samuels BA, Rainer Q, Wang JW, Marsteller D, Mendez I, Drew M, Craig DA, Guiard BP, Guilloux JP, Artymyshyn RP, Gardier AM, Gerald C, Antonijevic IA, Leonardo ED, Hen R. Neurogenesis-dependent and -independent effects of fluoxetine in an animal model of anxiety/depression. Neuron 62: 479 – 493, 2009. 79. Dayer AG, Ford AA, Cleaver KM, Yassaee M, Cameron HA. Short-term and longterm survival of new neurons in the rat dentate gyrus. J Comp Neurol 460: 563–572, 2003. 80. Deisseroth K, Singla S, Toda H, Monje M, Palmer TD, Malenka RC. Excitation-neurogenesis coupling in adult neural stem/progenitor cells. Neuron 42: 535–552, 2004. 61. Carpentier PA, Palmer TD. Immune influence on adult neural stem cell regulation and function. Neuron 64: 79 –92, 2009. 81. Demarque M, Represa A, Becq H, Khalilov I, Ben-Ari Y, Aniksztejn L. Paracrine intercellular communication by a Ca2⫹- and SNARE-independent release of GABA and glutamate prior to synapse formation. Neuron 36: 1051–1061, 2002. 62. Casadesus G, Shukitt-Hale B, Stellwagen HM, Zhu X, Lee HG, Smith MA, Joseph JA. Modulation of hippocampal plasticity and cognitive behavior by short-term blueberry supplementation in aged rats. Nutr Neurosci 7: 309 –316, 2004. 82. Deng W, Aimone JB, Gage FH. New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory? Nat Rev Neurosci 11: 339 – 350, 2010. 63. Cayre M, Canoll P, Goldman JE. Cell migration in the normal and pathological postnatal mammalian brain. Prog Neurobiol 88: 41– 63, 2009. 83. Deng W, Mayford M, Gage FH. Selection of distinct populations of dentate granule cells in response to inputs as a mechanism for pattern separation in mice. Elife 2: e00312, 2013. 64. Chancey JH, Adlaf EW, Sapp MC, Pugh PC, Wadiche JI, Overstreet-Wadiche LS. Depolarization is required for experience-dependent synapse unsilencing in adultborn neurons. J Neurosci 33: 6614 – 6622, 2013. 65. Chen MS, Walker J, Prusoff WH. Kinetic studies of herpes simplex virus type 1-encoded thymidine and thymidylate kinase, a multifunctional enzyme. J Biol Chem 254: 10747–10753, 1979. 66. Clark PJ, Brzezinska WJ, Puchalski EK, Krone DA, Rhodes JS. Functional analysis of neurovascular adaptations to exercise in the dentate gyrus of young adult mice associated with cognitive gain. Hippocampus 19: 937–950, 2009. 67. Clelland CD, Choi M, Romberg C, Clemenson GD Jr, Fragniere A, Tyers P, Jessberger S, Saksida LM, Barker RA, Gage FH, Bussey TJ. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science 325: 210 –213, 2009. 68. Collier RJ. Diphtheria toxin: mode of action and structure. Bacteriol Rev 39: 54 – 85, 1975. 69. Collinson N, Kuenzi FM, Jarolimek W, Maubach KA, Cothliff R, Sur C, Smith A, Otu FM, Howell O, Atack JR, McKernan RM, Seabrook GR, Dawson GR, Whiting PJ, Rosahl TW. Enhanced learning and memory and altered GABAergic synaptic transmission in mice lacking the alpha 5 subunit of the GABAA receptor. J Neurosci 22: 5572–5580, 2002. 85. Deng W, Saxe MD, Gallina IS, Gage FH. Adult-born hippocampal dentate granule cells undergoing maturation modulate learning and memory in the brain. J Neurosci 29: 13532–13542, 2009. 86. Deshpande A, Bergami M, Ghanem A, Conzelmann KK, Lepier A, Götz M, Berninger B. Retrograde monosynaptic tracing reveals the temporal evolution of inputs onto new neurons in the adult dentate gyrus and olfactory bulb. Proc Natl Acad Sci USA 110: E1152–E1161, 2013. 87. Dobrossy MD, Drapeau E, Aurousseau C, Le Moal M, Piazza PV, Abrous DN. Differential effects of learning on neurogenesis: learning increases or decreases the number of newly born cells depending on their birth date. Mol Psychiatry 8: 974 –982, 2003. 88. Dranovsky A, Picchini AM, Moadel T, Sisti AC, Yamada A, Kimura S, Leonardo ED, Hen R. Experience dictates stem cell fate in the adult hippocampus. Neuron 70: 908 –923, 2011. 89. Drapeau E, Montaron MF, Aguerre S, Abrous DN. Learning-induced survival of new neurons depends on the cognitive status of aged rats. J Neurosci 27: 6037– 6044, 2007. 90. Dupret D, Fabre A, Dobrossy MD, Panatier A, Rodriguez JJ, Lamarque S, Lemaire V, Oliet SH, Piazza PV, Abrous DN. Spatial learning depends on both the addition and removal of new hippocampal neurons. PLoS Biol 5: e214, 2007. 70. Conover JC, Doetsch F, Garcia-Verdugo JM, Gale NW, Yancopoulos GD, AlvarezBuylla A. Disruption of Eph/ephrin signaling affects migration and proliferation in the adult subventricular zone. Nat Neurosci 3: 1091–1097, 2000. 91. Ehninger D, Kempermann G. Paradoxical effects of learning the Morris water maze on adult hippocampal neurogenesis in mice may be explained by a combination of stress and physical activity. Genes Brain Behav 5: 29 –39, 2006. 71. Cooper-Kuhn CM, Winkler J, Kuhn HG. Decreased neurogenesis after cholinergic forebrain lesion in the adult rat. J Neurosci Res 77: 155–165, 2004. 92. Ekstrand J, Hellsten J, Tingstrom A. Environmental enrichment, exercise and corticosterone affect endothelial cell proliferation in adult rat hippocampus and prefrontal cortex. Neurosci Lett 442: 203–207, 2008. 72. Correa SA, Hunter CJ, Palygin O, Wauters SC, Martin KJ, McKenzie C, McKelvey K, Morris RG, Pankratov Y, Arthur JS, Frenguelli BG. MSK1 regulates homeostatic and experience-dependent synaptic plasticity. J Neurosci 32: 13039 –13051, 2012. 93. Encinas JM, Michurina TV, Peunova N, Park JH, Tordo J, Peterson DA, Fishell G, Koulakov A, Enikolopov G. Division-coupled astrocytic differentiation and age-related Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1019 AIMONE ET AL. depletion of neural stem cells in the adult hippocampus. Cell Stem Cell 8: 566 –579, 2011. 94. Erickson KI, Prakash RS, Voss MW, Chaddock L, Heo S, McLaren M, Pence BD, Martin SA, Vieira VJ, Woods JA, McAuley E, Kramer AF. Brain-derived neurotrophic factor is associated with age-related decline in hippocampal volume. J Neurosci 30: 5368 –5375, 2010. 95. Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH. Neurogenesis in the adult human hippocampus. Nature Med 4: 1313–1317, 1998. 96. Ernst A, Alkass K, Bernard S, Salehpour M, Perl S, Tisdale J, Possnert G, Druid H, Frisen J. Neurogenesis in the striatum of the adult human brain. Cell 156: 1072–1083, 2014. 97. Esposito MS, Piatti VC, Laplagne DA, Morgenstern NA, Ferrari CC, Pitossi FJ, Schinder AF. Neuronal differentiation in the adult hippocampus recapitulates embryonic development. J Neurosci 25: 10074 –10086, 2005. 98. Fabel K, Tam B, Kaufer D, Baiker A, Simmons N, Kuo CJ, Palmer TD. VEGF is necessary for exercise-induced adult hippocampal neurogenesis. Eur J Neurosci 18: 2803–2812, 2003. 114. Ge S, Goh EL, Sailor KA, Kitabatake Y, Ming GL, Song H. GABA regulates synaptic integration of newly generated neurons in the adult brain. Nature 439: 589 –593, 2006. 115. Ge S, Yang CH, Hsu KS, Ming GL, Song H. A critical period for enhanced synaptic plasticity in newly generated neurons of the adult brain. Neuron 54: 559 –566, 2007. 116. Gilbert PE, Kesner RP, Lee I. Dissociating hippocampal subregions: double dissociation between dentate gyrus and CA1. Hippocampus 11: 626 – 636, 2001. 117. Glykys J, Mody I. Hippocampal network hyperactivity after selective reduction of tonic inhibition in GABA A receptor alpha5 subunit-deficient mice. J Neurophysiol 95: 2796 –2807, 2006. 118. Goebbels S, Bode U, Pieper A, Funfschilling U, Schwab MH, Nave KA. Cre/loxPmediated inactivation of the bHLH transcription factor gene NeuroD/BETA2. Genesis 42: 247–252, 2005. 119. Gosgnach S, Lanuza GM, Butt SJ, Saueressig H, Zhang Y, Velasquez T, Riethmacher D, Callaway EM, Kiehn O, Goulding M. V1 spinal neurons regulate the speed of vertebrate locomotor outputs. Nature 440: 215–219, 2006. 120. Gould E. How widespread is adult neurogenesis in mammals? Nat Rev Neurosci 8: 481– 488, 2007. 99. Fabel K, Wolf SA, Ehninger D, Babu H, Leal-Galicia P, Kempermann G. Additive effects of physical exercise and environmental enrichment on adult hippocampal neurogenesis in mice. Front Neurosci 3: 50, 2009. 121. Gould E. Serotonin and hippocampal neurogenesis. Neuropsychopharmacology 21: 46S–51S, 1999. 100. Faigle R, Song H. Signaling mechanisms regulating adult neural stem cells and neurogenesis. Biochim Biophys Acta 1830: 2435–2448, 2013. 122. Gould E, Beylin A, Tanapat P, Reeves A, Shors TJ. Learning enhances adult neurogenesis in the hippocampal formation. Nat Neurosci 2: 260 –265, 1999. 101. Fanselow MS, Dong HW. Are the dorsal and ventral hippocampus functionally distinct structures? Neuron 65: 7–19, 2010. 123. Gould E, Cameron HA, Daniels DC, Woolley CS, McEwen BS. Adrenal hormones suppress cell division in the adult rat dentate gyrus. J Neurosci 12: 3642–3650, 1992. 102. Feart C, Samieri C, Rondeau V, Amieva H, Portet F, Dartigues JF, Scarmeas N, Barberger-Gateau P. Adherence to a Mediterranean diet, cognitive decline, and risk of dementia. JAMA 302: 638 – 648, 2009. 124. Gould E, Gross CG. Neurogenesis in adult mammals: some progress and problems. J Neurosci 22: 619 – 623, 2002. 103. Fenno L, Yizhar O, Deisseroth K. The development and application of optogenetics. Annu Rev Neurosci 34: 389 – 412, 2011. 104. Ferragud A, Haro A, Sylvain A, Velazquez-Sanchez C, Hernandez-Rabaza V, Canales JJ. Enhanced habit-based learning and decreased neurogenesis in the adult hippocampus in a murine model of chronic social stress. Behav Brain Res 210: 134 –139, 2010. 105. Fitzsimons CP, van Hooijdonk LW, Schouten M, Zalachoras I, Brinks V, Zheng T, Schouten TG, Saaltink DJ, Dijkmans T, Steindler DA, Verhaagen J, Verbeek FJ, Lucassen PJ, de Kloet ER, Meijer OC, Karst H, Joels M, Oitzl MS, Vreugdenhil E. Knockdown of the glucocorticoid receptor alters functional integration of newborn neurons in the adult hippocampus and impairs fear-motivated behavior. Mol Psychiatry 18: 993–1005, 2013. 106. Fonnum F, Johnsen A, Hassel B. Use of fluorocitrate and fluoroacetate in the study of brain metabolism. Glia 21: 106 –113, 1997. 107. Frankland PW, Kohler S, Josselyn SA. Hippocampal neurogenesis and forgetting. Trends Neurosci 36: 497–503, 2013. 108. Freund J, Brandmaier AM, Lewejohann L, Kirste I, Kritzler M, Kruger A, Sachser N, Lindenberger U, Kempermann G. Emergence of individuality in genetically identical mice. Science 340: 756 –759, 2013. 109. Freund J, Brandmaier AM, Lewejohann L, Kirste I, Kritzler M, Krüger A, Sachser N, Lindenberger U, Kempermann G. Emergence of individuality in genetically identical mice. Science 340: 756 –759, 2013. 110. Freund TF, Gulyas AI, Acsady L, Gorcs T, Toth K. Serotonergic control of the hippocampus via local inhibitory interneurons. Proc Natl Acad Sci USA 87: 8501– 8505, 1990. 125. Gould E, McEwen BS, Tanapat P, Galea LA, Fuchs E. Neurogenesis in the dentate gyrus of the adult tree shrew is regulated by psychosocial stress and NMDA receptor activation. J Neurosci 17: 2492–2498, 1997. 126. Gould E, Reeves AJ, Graziano MS, Gross CG. Neurogenesis in the neocortex of adult primates. Science 286: 548 –552, 1999. 127. Gould E, Tanapat P, McEwen BS, Flugge G, Fuchs E. Proliferation of granule cell precursors in the dentate gyrus of adult monkeys is diminished by stress. Proc Natl Acad Sci USA 95: 3168 –3171, 1998. 128. Gross CG. Neurogenesis in the adult brain: death of a dogma. Nat Rev Neurosci 1: 67–73, 2000. 129. Gu Y, Arruda-Carvalho M, Wang J, Janoschka SR, Josselyn SA, Frankland PW, Ge S. Optical controlling reveals time-dependent roles for adult-born dentate granule cells. Nat Neurosci 15: 1700 –1706, 2012. 130. Harley CW. Norepinephrine and the dentate gyrus. Prog Brain Res 163: 299 –318, 2007. 131. Hattiangady B, Shetty AK. Aging does not alter the number or phenotype of putative stem/progenitor cells in the neurogenic region of the hippocampus. Neurobiol Aging 29: 129 –147, 2008. 132. Haubensak W, Kunwar PS, Cai H, Ciocchi S, Wall NR, Ponnusamy R, Biag J, Dong HW, Deisseroth K, Callaway EM, Fanselow MS, Luthi A, Anderson DJ. Genetic dissection of an amygdala microcircuit that gates conditioned fear. Nature 468: 270 –276, 2010. 133. Hayashi Y, Yoshida M, Yamato M, Ide T, Wu Z, Ochi-Shindou M, Kanki T, Kang D, Sunagawa K, Tsutsui H, Nakanishi H. Reverse of age-dependent memory impairment and mitochondrial DNA damage in microglia by an overexpression of human mitochondrial transcription factor a in mice. J Neurosci 28: 8624 – 8634, 2008. 111. Gage FH. Mammalian neural stem cells. Science 287: 1433–1438, 2000. 112. Gao Z, Ure K, Ables JL, Lagace DC, Nave KA, Goebbels S, Eisch AJ, Hsieh J. Neurod1 is essential for the survival and maturation of adult-born neurons. Nat Neurosci 12: 1090 –1092, 2009. 134. Heine VM, Maslam S, Joels M, Lucassen PJ. Prominent decline of newborn cell proliferation, differentiation, and apoptosis in the aging dentate gyrus, in absence of an age-related hypothalamus-pituitary-adrenal axis activation. Neurobiol Aging 25: 361–375, 2004. 113. Garner AR, Rowland DC, Hwang SY, Baumgaertel K, Roth BL, Kentros C, Mayford M. Generation of a synthetic memory trace. Science 335: 1513–1516, 2012. 135. Henze DA, Wittner L, Buzsaki G. Single granule cells reliably discharge targets in the hippocampal CA3 network in vivo. Nat Neurosci 5: 790 –795, 2002. 1020 Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS 136. Hirrlinger J, Gutterer JM, Kussmaul L, Hamprecht B, Dringen R. Microglial cells in culture express a prominent glutathione system for the defense against reactive oxygen species. Dev Neurosci 22: 384 –392, 2000. 158. Kempermann G, Gast D, Gage FH. Neuroplasticity in old age: sustained fivefold induction of hippocampal neurogenesis by long-term environmental enrichment. Ann Neurol 52: 135–143, 2002. 137. Hodge RD, Kowalczyk TD, Wolf SA, Encinas JM, Rippey C, Enikolopov G, Kempermann G, Hevner RF. Intermediate progenitors in adult hippocampal neurogenesis: Tbr2 expression and coordinate regulation of neuronal output. J Neurosci 28: 3707– 3717, 2008. 159. Kempermann G, Gast D, Kronenberg G, Yamaguchi M, Gage FH. Early determination and long-term persistence of adult-generated new neurons in the hippocampus of mice. Development 130: 391–399, 2003. 138. Holick KA, Lee DC, Hen R, Dulawa SC. Behavioral effects of chronic fluoxetine in BALB/cJ mice do not require adult hippocampal neurogenesis or the serotonin 1A receptor. Neuropsychopharmacology 33: 406 – 417, 2008. 139. Hsieh J. Orchestrating transcriptional control of adult neurogenesis. Genes Development 26: 1010 –1021, 2012. 140. Imayoshi I, Sakamoto M, Ohtsuka T, Takao K, Miyakawa T, Yamaguchi M, Mori K, Ikeda T, Itohara S, Kageyama R. Roles of continuous neurogenesis in the structural and functional integrity of the adult forebrain. Nat Neurosci 11: 1153–1161, 2008. 141. Inoue K, Furukawa T, Kumada T, Yamada J, Wang T, Inoue R, Fukuda A. Taurine inhibits the K⫹-Cl⫺ cotransporter KCC2 to regulate embryonic Cl⫺ homeostasis via the with-no-lysine (WNK) protein kinase signaling pathway. J Biol Chem 287: 20839 – 20850, 2012. 142. Jacobs BL, van Praag H, Gage FH. Adult brain neurogenesis and psychiatry: a novel theory of depression. Mol Psychiatry 5: 262–269, 2000. 143. Jang MH, Bonaguidi MA, Kitabatake Y, Sun J, Song J, Kang E, Jun H, Zhong C, Su Y, Guo JU. Secreted frizzled-related protein 3 regulates activity-dependent adult hippocampal neurogenesis. Cell Stem Cell 12: 215–223, 2013. 144. Jentsch TJ, Stein V, Weinreich F, Zdebik AA. Molecular structure and physiological function of chloride channels. Physiol Rev 82: 503–568, 2002. 145. Jin K, Zhu Y, Sun Y, Mao XO, Xie L, Greenberg DA. Vascular endothelial growth factor (VEGF) stimulates neurogenesis in vitro and in vivo. Proc Natl Acad Sci USA 99: 11946 –11950, 2002. 160. Kempermann G, Kuhn HG, Gage FH. More hippocampal neurons in adult mice living in an enriched environment. Nature 386: 493– 495, 1997. 161. Kesner RP. A behavioral analysis of dentate gyrus function. Prog Brain Res 163: 567–576, 2007. 162. Kheirbek MA, Drew LJ, Burghardt NS, Costantini DO, Tannenholz L, Ahmari SE, Zeng H, Fenton AA, Hen R. Differential control of learning and anxiety along the dorsoventral axis of the dentate gyrus. Neuron 77: 955–968, 2013. 163. Kirby ED, Muroy SE, Sun WG, Covarrubias D, Leong MJ, Barchas LA, Kaufer D. Acute stress enhances adult rat hippocampal neurogenesis and activation of newborn neurons via secreted astrocytic FGF2. Elife 2: e00362, 2013. 164. Kitamura T, Saitoh Y, Takashima N, Murayama A, Niibori Y, Ageta H, Sekiguchi M, Sugiyama H, Inokuchi K. Adult neurogenesis modulates the hippocampus-dependent period of associative fear memory. Cell 139: 814 – 827, 2009. 165. Kitasato S. Experimentelle Untersuchungen über das Tetanusgift. Medical Microbiol Immunol 10: 267, 1891. 166. Klempin F, Beis D, Mosienko V, Kempermann G, Bader M, Alenina N. Serotonin is required for exercise-induced adult hippocampal neurogenesis. J Neurosci 33: 8270 – 8275, 2013. 167. Kobilo T, Liu QR, Gandhi K, Mughal M, Shaham Y, van Praag H. Running is the neurogenic and neurotrophic stimulus in environmental enrichment. Learn Mem 18: 605– 609, 2011. 168. Kokoeva MV, Yin H, Flier JS. Neurogenesis in the hypothalamus of adult mice: potential role in energy balance. Science 310: 679 – 683, 2005. 146. Joseph JA, Shukitt-Hale B, Denisova NA, Bielinski D, Martin A, McEwen JJ, Bickford PC. Reversals of age-related declines in neuronal signal transduction, cognitive, and motor behavioral deficits with blueberry, spinach, or strawberry dietary supplementation. J Neurosci 19: 8114 – 8121, 1999. 169. Kosofsky BE, Molliver ME. The serotoninergic innervation of cerebral cortex: different classes of axon terminals arise from dorsal and median raphe nuclei. Synapse 1: 153–168, 1987. 147. Jung MW, McNaughton BL. Spatial selectivity of unit activity in the hippocampal granular layer. Hippocampus 3: 165–182, 1993. 170. Krnjevic K, Schwartz S. The action of gamma-aminobutyric acid on cortical neurones. Exp Brain Res 3: 320 –336, 1967. 148. Kaneko N, Okano H, Sawamoto K. Role of the cholinergic system in regulating survival of newborn neurons in the adult mouse dentate gyrus and olfactory bulb. Genes Cells 11: 1145–1159, 2006. 171. Krzisch M, Temprana SG, Mongiat LA, Armida J, Schmutz V, Virtanen MA, KocherBraissant J, Kraftsik R, Vutskits L, Conzelmann KK, Bergami M, Gage FH, Schinder AF, Toni N. Pre-existing astrocytes form functional perisynaptic processes on neurons generated in the adult hippocampus. Brain Struct Funct. In press. 149. Kang SH, Fukaya M, Yang JK, Rothstein JD, Bergles DE. NG2⫹ CNS glial progenitors remain committed to the oligodendrocyte lineage in postnatal life and following neurodegeneration. Neuron 68: 668 – 681, 2010. 172. Kuhn HG, Cooper-Kuhn CM. Bromodeoxyuridine and the detection of neurogenesis. Curr Pharmaceutical Biotechnol 8: 127–131, 2007. 150. Kaplan MS. Environment complexity stimulates visual cortex neurogenesis: death of a dogma and a research career. Trends Neurosci 24: 617– 620, 2001. 173. Kuhn HG, Dickinson-Anson H, Gage FH. Neurogenesis in the dentate gyrus of the adult rat: age-related decrease of neuronal progenitor proliferation. J Neurosci 16: 2027–2033, 1996. 151. Karalay O, Jessberger S. Translating niche-derived signals into neurogenesis: the function of Prox1 in the adult hippocampus. Cell Cycle 10: 2239 –2240, 2011. 152. Karelina K, Hansen KF, Choi YS, DeVries AC, Arthur JS, Obrietan K. MSK1 regulates environmental enrichment-induced hippocampal plasticity and cognitive enhancement. Learn Mem 19: 550 –560, 2012. 153. Katoh-Semba R, Semba R, Takeuchi IK, Kato K. Age-related changes in levels of brain-derived neurotrophic factor in selected brain regions of rats, normal mice and senescence-accelerated mice: a comparison to those of nerve growth factor and neurotrophin-3. Neurosci Res 31: 227–234, 1998. 154. Katoh-Semba R, Takeuchi IK, Semba R, Kato K. Distribution of brain-derived neurotrophic factor in rats and its changes with development in the brain. J Neurochem 69: 34 – 42, 1997. 155. Kee N, Teixeira CM, Wang AH, Frankland PW. Preferential incorporation of adultgenerated granule cells into spatial memory networks in the dentate gyrus. Nat Neurosci 10: 355–362, 2007. 156. Kempermann G. Seven principles in the regulation of adult neurogenesis. Eur J Neurosci 33: 1018 –1024, 2011. 174. Kulkarni VA, Jha S, Vaidya VA. Depletion of norepinephrine decreases the proliferation, but does not influence the survival and differentiation, of granule cell progenitors in the adult rat hippocampus. Eur J Neurosci 16: 2008 –2012, 2002. 175. Kuwabara T, Hsieh J, Muotri A, Yeo G, Warashina M, Lie DC, Moore L, Nakashima K, Asashima M, Gage FH. Wnt-mediated activation of NeuroD1 and retro-elements during adult neurogenesis. Nat Neurosci 12: 1097–1105, 2009. 176. Kuzumaki N, Ikegami D, Tamura R, Hareyama N, Imai S, Narita M, Torigoe K, Niikura K, Takeshima H, Ando T, Igarashi K, Kanno J, Ushijima T, Suzuki T. Hippocampal epigenetic modification at the brain-derived neurotrophic factor gene induced by an enriched environment. Hippocampus 21: 127–132, 2011. 177. Lagace DC, Whitman MC, Noonan MA, Ables JL, DeCarolis NA, Arguello AA, Donovan MH, Fischer SJ, Farnbauch LA, Beech RD, DiLeone RJ, Greer CA, Mandyam CD, Eisch AJ. Dynamic contribution of nestin-expressing stem cells to adult neurogenesis. J Neurosci 27: 12623–12629, 2007. 178. Lapchak PA, Araujo DM, Beck KD, Finch CE, Johnson SA, Hefti F. BDNF and trkB mRNA expression in the hippocampal formation of aging rats. Neurobiol Aging 14: 121–126, 1993. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1021 AIMONE ET AL. 179. Laplagne DA, Kamienkowski JE, Esposito MS, Piatti VC, Zhao C, Gage FH, Schinder AF. Similar GABAergic inputs in dentate granule cells born during embryonic and adult neurogenesis. Eur J Neurosci 25: 2973–2981, 2007. 201. Liu X, Wang Q, Haydar TF, Bordey A. Nonsynaptic GABA signaling in postnatal subventricular zone controls proliferation of GFAP-expressing progenitors. Nature Neurosci 8: 1179 –1187, 2005. 180. Lavado A, Oliver G. Prox1 expression patterns in the developing and adult murine brain. Dev Dyn 236: 518 –524, 2007. 202. LoTurco JJ, Owens DF, Heath MJ, Davis MB, Kriegstein AR. GABA and glutamate depolarize cortical progenitor cells and inhibit DNA synthesis. Neuron 15: 1287–1298, 1995. 181. Lechner HA, Lein ES, Callaway EM. A genetic method for selective and quickly reversible silencing of Mammalian neurons. J Neurosci 22: 5287–5290, 2002. 182. Lee CK, Weindruch R, Prolla TA. Gene-expression profile of the ageing brain in mice. Nat Genet 25: 294 –297, 2000. 183. Lee JE, Hollenberg SM, Snider L, Turner DL, Lipnick N, Weintraub H. Conversion of Xenopus ectoderm into neurons by NeuroD, a basic helix-loop-helix protein. Science 268: 836 – 844, 1995. 203. Lucin KM, Wyss-Coray T. Immune activation in brain aging and neurodegeneration: too much or too little? Neuron 64: 110 –122, 2009. 204. Lugert S, Basak O, Knuckles P, Haussler U, Fabel K, Götz M, Haas CA, Kempermann G, Taylor V, Giachino C. Quiescent and active hippocampal neural stem cells with distinct morphologies respond selectively to physiological and pathological stimuli and aging. Cell Stem Cell 6: 445– 456, 2010. 184. Lee JL, Everitt BJ, Thomas KL. Independent cellular processes for hippocampal memory consolidation and reconsolidation. Science 304: 839 – 843, 2004. 205. Lupien SJ, Maheu F, Tu M, Fiocco A, Schramek TE. The effects of stress and stress hormones on human cognition: implications for the field of brain and cognition. Brain Cognition 65: 209 –237, 2007. 185. Lee SW, Clemenson GD, Gage FH. New neurons in an aged brain. Behav Brain Res 227: 497–507, 2012. 206. Luzzati F, De Marchis S, Fasolo A, Peretto P. Neurogenesis in the caudate nucleus of the adult rabbit. J Neurosci 26: 609 – 621, 2006. 186. Lehmann ML, Brachman RA, Martinowich K, Schloesser RJ, Herkenham M. Glucocorticoids orchestrate divergent effects on mood through adult neurogenesis. J Neurosci 33: 2961–2972, 2013. 207. Ma DK, Marchetto MC, Guo JU, Ming GL, Gage FH, Song H. Epigenetic choreographers of neurogenesis in the adult mammalian brain. Nat Neurosci 13: 1338 –1344, 2010. 187. Lemaire V, Tronel S, Montaron MF, Fabre A, Dugast E, Abrous DN. Long-lasting plasticity of hippocampal adult-born neurons. J Neurosci 32: 3101–3108, 2012. 208. Macready AL, Kennedy OB, Ellis JA, Williams CM, Spencer JP, Butler LT. Flavonoids and cognitive function: a review of human randomized controlled trial studies and recommendations for future studies. Genes Nutr 4: 227–242, 2009. 188. Leranth C, Hajszan T. Extrinsic afferent systems to the dentate gyrus. Prog Brain Res 163: 63– 84, 2007. 189. Letenneur L, Proust-Lima C, Le GA, Dartigues JF, Barberger-Gateau P. Flavonoid intake and cognitive decline over a 10-year period. Am J Epidemiol 165: 1364 –1371, 2007. 190. Leutgeb JK, Leutgeb S, Moser MB, Moser EI. Pattern separation in the dentate gyrus and CA3 of the hippocampus. Science 315: 961–966, 2007. 191. Leventhal C, Rafii S, Rafii D, Shahar A, Goldman SA. Endothelial trophic support of neuronal production and recruitment from the adult mammalian subependyma. Mol Cell Neurosci 13: 450 – 464, 1999. 192. Li Q, Zhao HF, Zhang ZF, Liu ZG, Pei XR, Wang JB, Cai MY, Li Y. Long-term administration of green tea catechins prevents age-related spatial learning and memory decline in C57BL/6 J mice by regulating hippocampal cyclic amp-response element binding protein signaling cascade. Neuroscience 159: 1208 –1215, 2009. 193. Li Y, Aimone JB, Xu X, Callaway EM, Gage FH. Development of GABAergic inputs controls the contribution of maturing neurons to the adult hippocampal network. Proc Natl Acad Sci USA 109: 4290 – 4295, 2012. 209. Malberg JE, Eisch AJ, Nestler EJ, Duman RS. Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. J Neurosci 20: 9104 –9110, 2000. 210. Manach C, Scalbert A, Morand C, Rémésy C, Jiménez L. Polyphenols: food sources and bioavailability. The Am J Clin Nutr 79: 727–747, 2004. 211. Mank M, Santos AF, Direnberger S, Mrsic-Flogel TD, Hofer SB, Stein V, Hendel T, Reiff DF, Levelt C, Borst A, Bonhoeffer T, Hubener M, Griesbeck O. A genetically encoded calcium indicator for chronic in vivo two-photon imaging. Nat Methods 5: 805– 811, 2008. 212. Marban E, Rink TJ, Tsien RW, Tsien RY. Free calcium in heart muscle at rest and during contraction measured with Ca2⫹-sensitive microelectrodes. Nature 286: 845– 850, 1980. 213. Marin-Burgin A, Mongiat LA, Pardi MB, Schinder AF. Unique processing during a period of high excitation/inhibition balance in adult-born neurons. Science 335: 1238 – 1242, 2012. 214. Markwardt SJ, Dieni CV, Wadiche JI, Overstreet-Wadiche L. Ivy/neurogliaform interneurons coordinate activity in the neurogenic niche. Nat Neurosci 14: 1407–1409, 2011. 194. Li Y, Luikart BW, Birnbaum S, Chen J, Kwon CH, Kernie SG, Bassel-Duby R, Parada LF. TrkB regulates hippocampal neurogenesis and governs sensitivity to antidepressive treatment. Neuron 59: 399 – 412, 2008. 215. Markwardt SJ, Wadiche JI, Overstreet-Wadiche LS. Input-specific GABAergic signaling to newborn neurons in adult dentate gyrus. J Neurosci 29: 15063–15072, 2009. 195. Li Y, Stam FJ, Aimone JB, Goulding M, Callaway EM, Gage FH. Molecular layer perforant path-associated cells contribute to feed-forward inhibition in the adult dentate gyrus. Proc Natl Acad Sci USA 110: 9106 –9111, 2013. 216. Marty S, Berninger B, Carroll P, Thoenen H. GABAergic stimulation regulates the phenotype of hippocampal interneurons through the regulation of brain-derived neurotrophic factor. Neuron 16: 565–570, 1996. 196. Licht T, Goshen I, Avital A, Kreisel T, Zubedat S, Eavri R, Segal M, Yirmiya R, Keshet E. Reversible modulations of neuronal plasticity by VEGF. Proc Natl Acad Sci USA 108: 5081–5086, 2011. 217. Masiulis I, Yun S, Eisch AJ. The interesting interplay between interneurons and adult hippocampal neurogenesis. Mol Neurobiol 44: 287–302, 2011. 197. Lie DC, Colamarino SA, Song HJ, Desire L, Mira H, Consiglio A, Lein ES, Jessberger S, Lansford H, Dearie AR, Gage FH. Wnt signalling regulates adult hippocampal neurogenesis. Nature 437: 1370 –1375, 2005. 218. Masuda T, Nakagawa S, Boku S, Nishikawa H, Takamura N, Kato A, Inoue T, Koyama T. Noradrenaline increases neural precursor cells derived from adult rat dentate gyrus through beta2 receptor. Prog Neuro-psychopharmacology Biol Psychiatry 36: 44 – 51, 2012. 198. Lindenau J, Noack H, Asayama K, Wolf G. Enhanced cellular glutathione peroxidase immunoreactivity in activated astrocytes and in microglia during excitotoxin induced neurodegeneration. Glia 24: 252–256, 1998. 219. Mathews EA, Morgenstern NA, Piatti VC, Zhao C, Jessberger S, Schinder AF, Gage FH. A distinctive layering pattern of mouse dentate granule cells is generated by developmental and adult neurogenesis. J Comp Neurol 518: 4479 – 4490, 2010. 199. Liu X, Ramirez S, Pang PT, Puryear CB, Govindarajan A, Deisseroth K, Tonegawa S. Optogenetic stimulation of a hippocampal engram activates fear memory recall. Nature 484: 381–385, 2012. 220. Maxwell IH, Maxwell F, Glode LM. Regulated expression of a diphtheria toxin A-chain gene transfected into human cells: possible strategy for inducing cancer cell suicide. Cancer Res 46: 4660 – 4664, 1986. 200. Liu X, Ramirez S, Pang PT, Puryear CB, Govindarajan A, Deisseroth K, Tonegawa S. Optogenetic stimulation of a hippocampal engram activates fear memory recall. Nature 484: 381–385, 2012. 221. McCloskey DP, Adamo DS, Anderson BJ. Exercise increases metabolic capacity in the motor cortex and striatum, but not in the hippocampus. Brain Res 891: 168 –175, 2001. 1022 Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS 222. McHugh TJ, Jones MW, Quinn JJ, Balthasar N, Coppari R, Elmquist JK, Lowell BB, Fanselow MS, Wilson MA, Tonegawa S. Dentate gyrus NMDA receptors mediate rapid pattern separation in the hippocampal network. Science 317: 94 –99, 2007. 223. McNaughton BL, Morris RGM. Hippocampal synaptic enhancement and information storage within a distributed memory system. Trends Neurosci 10: 1987. 224. McTighe SM, Mar AC, Romberg C, Bussey TJ, Saksida LM. A new touchscreen test of pattern separation: effect of hippocampal lesions. Neuroreport 20: 881– 885, 2009. 225. Meshi D, Drew MR, Saxe M, Ansorge MS, David D, Santarelli L, Malapani C, Moore H, Hen R. Hippocampal neurogenesis is not required for behavioral effects of environmental enrichment. Nat Neurosci 9: 729 –731, 2006. 226. Mignone JL, Kukekov V, Chiang AS, Steindler D, Enikolopov G. Neural stem and progenitor cells in nestin-GFP transgenic mice. J Comp Neurol 469: 311–324, 2004. 227. Miller BH, Schultz LE, Gulati A, Cameron MD, Pletcher MT. Genetic regulation of behavioral and neuronal responses to fluoxetine. Neuropsychopharmacology 33: 1312– 1322, 2008. 228. Mineur YS, Belzung C, Crusio WE. Functional implications of decreases in neurogenesis following chronic mild stress in mice. Neuroscience 150: 251–259, 2007. 229. Miranda CJ, Braun L, Jiang Y, Hester ME, Zhang L, Riolo M, Wang H, Rao M, Altura RA, Kaspar BK. Aging brain microenvironment decreases hippocampal neurogenesis through Wnt-mediated survivin signaling. Aging Cell 11: 542–552, 2012. 230. Miyamichi K, Amat F, Moussavi F, Wang C, Wickersham I, Wall NR, Taniguchi H, Tasic B, Huang ZJ, He Z, Callaway EM, Horowitz MA, Luo L. Cortical representations of olfactory input by trans-synaptic tracing. Nature 472: 191–196, 2011. 231. Mongiat LA, Esposito MS, Lombardi G, Schinder AF. Reliable activation of immature neurons in the adult hippocampus. PloS one 4: e5320, 2009. 232. Mongiat LA, Schinder AF. Adult neurogenesis and the plasticity of the dentate gyrus network. Eur J Neurosci 33: 1055–1061, 2011. 233. Monje ML, Toda H, Palmer TD. Inflammatory blockade restores adult hippocampal neurogenesis. Science 302: 1760 –1765, 2003. 234. Morris AM, Churchwell JC, Kesner RP, Gilbert PE. Selective lesions of the dentate gyrus produce disruptions in place learning for adjacent spatial locations. Neurobiol Learning Memory 97: 326 –331, 2012. 235. Morris AM, Curtis BJ, Churchwell JC, Maasberg DW, Kesner RP. Temporal associations for spatial events: the role of the dentate gyrus. Behav Brain Res 256: 250 –256, 2013. 236. Moser MB, Moser EI. Functional differentiation in the hippocampus. Hippocampus 8: 608 – 619, 1998. 237. Mu Y, Lee SW, Gage FH. Signaling in adult neurogenesis. Curr Opin Neurobiol 20: 416 – 423, 2010. 238. Mu Y, Zhao C, Gage FH. Dopaminergic modulation of cortical inputs during maturation of adult-born dentate granule cells. J Neurosci 31: 4113– 4123, 2011. Young dentate granule cells mediate pattern separation, whereas old granule cells facilitate pattern completion. Cell 149: 188 –201, 2012. 245. Namba T, Mochizuki H, Onodera M, Mizuno Y, Namiki H, Seki T. The fate of neural progenitor cells expressing astrocytic and radial glial markers in the postnatal rat dentate gyrus. Eur J Neurosci 22: 1928 –1941, 2005. 246. Naya FJ, Huang HP, Qiu Y, Mutoh H, DeMayo FJ, Leiter AB, Tsai MJ. Diabetes, defective pancreatic morphogenesis, and abnormal enteroendocrine differentiation in BETA2/neuroD-deficient mice. Genes Development 11: 2323–2334, 1997. 247. Naya FJ, Stellrecht CM, Tsai MJ. Tissue-specific regulation of the insulin gene by a novel basic helix-loop-helix transcription factor. Genes Development 9: 1009 –1019, 1995. 248. Niibori Y, Yu TS, Epp JR, Akers KG, Josselyn SA, Frankland PW. Suppression of adult neurogenesis impairs population coding of similar contexts in hippocampal CA3 region. Nature Commun 3: 1253, 2012. 249. Nimmerjahn A, Kirchhoff F, Helmchen F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 308: 1314 –1318, 2005. 250. Ninkovic J, Mori T, Gotz M. Distinct modes of neuron addition in adult mouse neurogenesis. J Neurosci 27: 10906 –10911, 2007. 251. Njie EG, Boelen E, Stassen FR, Steinbusch HW, Borchelt DR, Streit WJ. Ex vivo cultures of microglia from young and aged rodent brain reveal age-related changes in microglial function. Neurobiol Aging 33: 195.e1–12, 2012. 252. Nogues X, Corsini MM, Marighetto A, Abrous DN. Functions for adult neurogenesis in memory: an introduction to the neurocomputational approach and to its contribution. Behav Brain Res 227: 418 – 425, 2012. 253. Nottebohm F. Why are some neurons replaced in adult brain? J Neurosci 22: 624 – 628, 2002. 254. Nurk E, Refsum H, Drevon CA, Tell GS, Nygaard HA, Engedal K, Smith AD. Intake of flavonoid-rich wine, tea, and chocolate by elderly men and women is associated with better cognitive test performance. J Nutr 139: 120 –127, 2009. 255. O’Reilly RC, McClelland JL. Hippocampal conjunctive encoding, storage, and recall: avoiding a trade-off. Hippocampus 4: 661– 682, 1994. 256. Oh J, McCloskey MA, Blong CC, Bendickson L, Nilsen-Hamilton M, Sakaguchi DS. Astrocyte-derived interleukin-6 promotes specific neuronal differentiation of neural progenitor cells from adult hippocampus. J Neurosci Res 88: 2798 –2809, 2010. 257. Olah M, Ping G, De Haas AH, Brouwer N, Meerlo P, Van Der Zee EA, Biber K, Boddeke HW. Enhanced hippocampal neurogenesis in the absence of microglia T cell interaction and microglia activation in the murine running wheel model. Glia 57: 1046 –1061, 2009. 258. Olson AK, Eadie BD, Ernst C, Christie BR. Environmental enrichment and voluntary exercise massively increase neurogenesis in the adult hippocampus via dissociable pathways. Hippocampus 16: 250 –260, 2006. 259. Overstreet-Wadiche LS, Bensen AL, Westbrook GL. Delayed development of adultgenerated granule cells in dentate gyrus. J Neurosci 26: 2326 –2334, 2006. 239. Mustroph M, Chen S, Desai S, Cay E, DeYoung E, Rhodes J. Aerobic exercise is the critical variable in an enriched environment that increases hippocampal neurogenesis and water maze learning in male C57BL/6J mice. Neuroscience 219: 62–71, 2012. 260. Overstreet Wadiche L, Bromberg DA, Bensen AL, Westbrook GL. GABAergic signaling to newborn neurons in dentate gyrus. J Neurophysiol 94: 4528 – 4532, 2005. 240. Nagayama S, Zeng S, Xiong W, Fletcher ML, Masurkar AV, Davis DJ, Pieribone VA, Chen WR. In vivo simultaneous tracing and Ca2⫹ imaging of local neuronal circuits. Neuron 53: 789 – 803, 2007. 261. Owens DF, Liu X, Kriegstein AR. Changing properties of GABA (A) receptor-mediated signaling during early neocortical development. J Neurophysiol 82: 570 –583, 1999. 241. Naglich JG, Metherall JE, Russell DW, Eidels L. Expression cloning of a diphtheria toxin receptor: identity with a heparin-binding EGF-like growth factor precursor. Cell 69: 1051–1061, 1992. 262. Palmer TD, Markakis EA, Willhoite AR, Safar F, Gage FH. Fibroblast growth factor-2 activates a latent neurogenic program in neural stem cells from diverse regions of the adult CNS. J Neurosci 19: 8487– 8497, 1999. 242. Nakai J, Ohkura M, Imoto K. A high signal-to-noise Ca2⫹ probe composed of a single green fluorescent protein. Nat Biotechnol 19: 137–141, 2001. 263. Palmer TD, Takahashi J, Gage FH. The adult rat hippocampus contains primordial neural stem cells. Mol Cell Neurosci 8: 389 – 404, 1997. 243. Nakamura Y, Si QS, Kataoka K. Lipopolysaccharide-induced microglial activation in culture: temporal profiles of morphological change and release of cytokines and nitric oxide. Neurosci Res 35: 95–100, 1999. 264. Palmer TD, Willhoite AR, Gage FH. Vascular niche for adult hippocampal neurogenesis. J Comp Neurol 425: 479 – 494, 2000. 244. Nakashiba T, Cushman JD, Pelkey KA, Renaudineau S, Buhl DL, McHugh TJ, Rodriguez Barrera V, Chittajallu R, Iwamoto KS, McBain CJ, Fanselow MS, Tonegawa S. 265. Palmiter RD, Behringer RR, Quaife CJ, Maxwell F, Maxwell IH, Brinster RL. Cell lineage ablation in transgenic mice by cell-specific expression of a toxin gene. Cell 50: 435– 443, 1987. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1023 AIMONE ET AL. 266. Pereira AC, Huddleston DE, Brickman AM, Sosunov AA, Hen R, McKhann GM, Sloan R, Gage FH, Brown TR, Small SA. An in vivo correlate of exercise-induced neurogenesis in the adult dentate gyrus. Proc Natl Acad Sci USA 104: 5638 –5643, 2007. 287. Saito M, Iwawaki T, Taya C, Yonekawa H, Noda M, Inui Y, Mekada E, Kimata Y, Tsuru A, Kohno K. Diphtheria toxin receptor-mediated conditional and targeted cell ablation in transgenic mice. Nat Biotechnol 19: 746 –750, 2001. 267. Petrik D, Lagace DC, Eisch AJ. The neurogenesis hypothesis of affective and anxiety disorders: are we mistaking the scaffolding for the building? Neuropharmacology 62: 21–34, 2012. 288. Sanai N, Tramontin AD, Quinones-Hinojosa A, Barbaro NM, Gupta N, Kunwar S, Lawton MT, McDermott MW, Parsa AT, Manuel-Garcia Verdugo J, Berger MS, Alvarez-Buylla A. Unique astrocyte ribbon in adult human brain contains neural stem cells but lacks chain migration. Nature 427: 740 –744, 2004. 268. Pham K, Nacher J, Hof PR, McEwen BS. Repeated restraint stress suppresses neurogenesis and induces biphasic PSA-NCAM expression in the adult rat dentate gyrus. Eur J Neurosci 17: 879 – 886, 2003. 269. Piatti VC, Davies-Sala MG, Esposito MS, Mongiat LA, Trinchero MF, Schinder AF. The timing for neuronal maturation in the adult hippocampus is modulated by local network activity. J Neurosci 31: 7715–7728, 2011. 270. Raber J, Rola R, LeFevour A, Morhardt D, Curley J, Mizumatsu S, VandenBerg SR, Fike JR. Radiation-induced cognitive impairments are associated with changes in indicators of hippocampal neurogenesis. Radiation Res 162: 39 – 47, 2004. 271. Radley JJ, Jacobs BL. 5-HT1A receptor antagonist administration decreases cell proliferation in the dentate gyrus. Brain Res 955: 264 –267, 2002. 272. Rakhit S, Clark CJ, O’Shaughnessy CT, Morris BJ. N-methyl-D-aspartate and brainderived neurotrophic factor induce distinct profiles of extracellular signal-regulated kinase, mitogen- and stress-activated kinase, and ribosomal s6 kinase phosphorylation in cortical neurons. Mol Pharmacol 67: 1158 –1165, 2005. 273. Rakic P. Neurogenesis in adult primate neocortex: an evaluation of the evidence. Nat Rev Neurosci 3: 65–71, 2002. 274. Ramirez S, Liu X, Lin PA, Suh J, Pignatelli M, Redondo RL, Ryan TJ, Tonegawa S. Creating a false memory in the hippocampus. Science 341: 387–391, 2013. 289. Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S, Weisstaub N, Lee J, Duman R, Arancio O, Belzung C, Hen R. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 301: 805– 809, 2003. 290. Saxe MD, Battaglia F, Wang JW, Malleret G, David DJ, Monckton JE, Garcia AD, Sofroniew MV, Kandel ER, Santarelli L, Hen R, Drew MR. Ablation of hippocampal neurogenesis impairs contextual fear conditioning and synaptic plasticity in the dentate gyrus. Proc Natl Acad Sci USA 103: 17501–17506, 2006. 291. Saxe MD, Malleret G, Vronskaya S, Mendez I, Garcia AD, Sofroniew MV, Kandel ER, Hen R. Paradoxical influence of hippocampal neurogenesis on working memory. Proc Natl Acad Sci USA 104: 4642– 4646, 2007. 292. Schanzer A, Wachs FP, Wilhelm D, Acker T, Cooper-Kuhn C, Beck H, Winkler J, Aigner L, Plate KH, Kuhn HG. Direct stimulation of adult neural stem cells in vitro and neurogenesis in vivo by vascular endothelial growth factor. Brain Pathol 14: 237–248, 2004. 293. Scharfman H, Goodman J, Macleod A, Phani S, Antonelli C, Croll S. Increased neurogenesis and the ectopic granule cells after intrahippocampal BDNF infusion in adult rats. Exp Neurol 192: 348 –356, 2005. 275. Rangel LM, Eichenbaum H. What’s new is older. eLife 2: e00605, 2013. 294. Schiavo G, Benfenati F, Poulain B, Rossetto O, Polverino de Laureto P, DasGupta BR, Montecucco C. Tetanus and botulinum-B neurotoxins block neurotransmitter release by proteolytic cleavage of synaptobrevin. Nature 359: 832– 835, 1992. 276. Rangel LM, Quinn LK, Chiba AA, Gage FH, Aimone JB. A hypothesis for temporal coding of young and mature granule cells. Front Neurosci 7: 75, 2013. 295. Schmidt-Hieber C, Jonas P, Bischofberger J. Enhanced synaptic plasticity in newly generated granule cells of the adult hippocampus. Nature 429: 184 –187, 2004. 277. Ribak CE, Shapiro LA, Perez ZD, Spigelman I. Microglia-associated granule cell death in the normal adult dentate gyrus. Brain Struct Funct 214: 25–35, 2009. 296. Schoenfeld TJ, Gould E. Stress, stress hormones, and adult neurogenesis. Exp Neurol 233: 12–21, 2012. 278. Rizk P, Salazar J, Raisman-Vozari R, Marien M, Ruberg M, Colpaert F, Debeir T. The alpha2-adrenoceptor antagonist dexefaroxan enhances hippocampal neurogenesis by increasing the survival and differentiation of new granule cells. Neuropsychopharmacology 31: 1146 –1157, 2006. 297. Seib DR, Corsini NS, Ellwanger K, Plaas C, Mateos A, Pitzer C, Niehrs C, Celikel T, Martin-Villalba A. Loss of Dickkopf-1 restores neurogenesis in old age and counteracts cognitive decline. Cell Stem Cell 12: 204 –214, 2013. 279. Rola R, Raber J, Rizk A, Otsuka S, VandenBerg SR, Morhardt DR, Fike JR. Radiationinduced impairment of hippocampal neurogenesis is associated with cognitive deficits in young mice. Exp Neurol 188: 316 –330, 2004. 280. Rubsam LZ, Davidson BL, Shewach DS. Superior cytotoxicity with ganciclovir compared with acyclovir and 1-beta-D-arabinofuranosylthymine in herpes simplex virusthymidine kinase-expressing cells: a novel paradigm for cell killing. Cancer Res 58: 3873–3882, 1998. 281. Rudge JS, Alderson RF, Pasnikowski E, McClain J, Ip NY, Lindsay RM. Expression of ciliary neurotrophic factor and the neurotrophins-nerve growth factor, brain-derived neurotrophic factor and neurotrophin 3-in cultured rat hippocampal astrocytes. Eur J Neurosci 4: 459 – 471, 1992. 282. Rudge JS, Pasnikowski EM, Holst P, Lindsay RM. Changes in neurotrophic factor expression and receptor activation following exposure of hippocampal neuron/astrocyte cocultures to kainic acid. J Neurosci 15: 6856 – 6867, 1995. 298. Seidenfaden R, Desoeuvre A, Bosio A, Virard I, Cremer H. Glial conversion of SVZderived committed neuronal precursors after ectopic grafting into the adult brain. Mol Cell Neurosci 32: 187–198, 2006. 299. Seress L. Comparative anatomy of the hippocampal dentate gyrus in adult and developing rodents, non-human primates and humans. Prog Brain Res 163: 23– 41, 2007. 300. Seress L, Abraham H, Tornoczky T, Kosztolanyi G. Cell formation in the human hippocampal formation from mid-gestation to the late postnatal period. Neuroscience 105: 831– 843, 2001. 301. Sharma HS, Cervos-Navarro J, Dey PK. Increased blood-brain barrier permeability following acute short-term swimming exercise in conscious normotensive young rats. Neurosci Res 10: 211–221, 1991. 302. Shelton DJ, Kirwan CB. A possible negative influence of depression on the ability to overcome memory interference. Behav Brain Res 256C: 20 –26, 2013. 283. Sahay A, Drew MR, Hen R. Dentate gyrus neurogenesis and depression. Prog Brain Res 163: 697–722, 2007. 303. Shen Q, Goderie SK, Jin L, Karanth N, Sun Y, Abramova N, Vincent P, Pumiglia K, Temple S. Endothelial cells stimulate self-renewal and expand neurogenesis of neural stem cells. Science 304: 1338 –1340, 2004. 284. Sahay A, Scobie KN, Hill AS, O’Carroll CM, Kheirbek MA, Burghardt NS, Fenton AA, Dranovsky A, Hen R. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature 472: 466 – 470, 2011. 304. Shen Q, Wang Y, Kokovay E, Lin G, Chuang SM, Goderie SK, Roysam B, Temple S. Adult SVZ stem cells lie in a vascular niche: a quantitative analysis of niche cell-cell interactions. Cell Stem Cell 3: 289 –300, 2008. 285. Sahay A, Wilson DA, Hen R. Pattern separation: a common function for new neurons in hippocampus and olfactory bulb. Neuron 70: 582–588, 2011. 305. Shetty AK, Hattiangady B, Shetty GA. Stem/progenitor cell proliferation factors FGF-2, IGF-1, and VEGF exhibit early decline during the course of aging in the hippocampus: role of astrocytes. Glia 51: 173–186, 2005. 286. Sairanen M, Lucas G, Ernfors P, Castren M, Castren E. Brain-derived neurotrophic factor and antidepressant drugs have different but coordinated effects on neuronal turnover, proliferation, and survival in the adult dentate gyrus. J Neurosci 25: 1089 – 1094, 2005. 1024 306. Shihabuddin LS, Horner PJ, Ray J, Gage FH. Adult spinal cord stem cells generate neurons after transplantation in the adult dentate gyrus. J Neurosci 20: 8727– 8735, 2000. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org REGULATION AND FUNCTION OF ADULT NEUROGENESIS 307. Shors TJ, Miesegaes G, Beylin A, Zhao M, Rydel T, Gould E. Neurogenesis in the adult is involved in the formation of trace memories. Nature 410: 372–376, 2001. 308. Shors TJ, Townsend DA, Zhao M, Kozorovitskiy Y, Gould E. Neurogenesis may relate to some but not all types of hippocampal-dependent learning. Hippocampus 12: 578 – 584, 2002. 309. Shukitt-Hale B, Carey A, Simon L, Mark DA, Joseph JA. Effects of Concord grape juice on cognitive and motor deficits in aging. Nutrition 22: 295–302, 2006. 327. Teixeira CM, Kron MM, Masachs N, Zhang H, Lagace DC, Martinez A, Reillo I, Duan X, Bosch C, Pujadas L. Cell-autonomous inactivation of the reelin pathway impairs adult neurogenesis in the hippocampus. J Neurosci 32: 12051–12065, 2012. 328. Thomas RM, Hotsenpiller G, Peterson DA. Acute psychosocial stress reduces cell survival in adult hippocampal neurogenesis without altering proliferation. J Neurosci 27: 2734 –2743, 2007. 310. Sierra A, Encinas JM, Deudero JJ, Chancey JH, Enikolopov G, Overstreet-Wadiche LS, Tsirka SE, Maletic-Savatic M. Microglia shape adult hippocampal neurogenesis through apoptosis-coupled phagocytosis. Cell Stem Cell 7: 483– 495, 2010. 329. Thust R, Tomicic M, Klocking R, Voutilainen N, Wutzler P, Kaina B. Comparison of the genotoxic and apoptosis-inducing properties of ganciclovir and penciclovir in Chinese hamster ovary cells transfected with the thymidine kinase gene of herpes simplex virus-1: implications for gene therapeutic approaches. Cancer Gene Ther 7: 107–117, 2000. 311. Silhol M, Bonnichon V, Rage F, Tapia-Arancibia L. Age-related changes in brainderived neurotrophic factor and tyrosine kinase receptor isoforms in the hippocampus and hypothalamus in male rats. Neuroscience 132: 613– 624, 2005. 330. Toni N, Laplagne DA, Zhao C, Lombardi G, Ribak CE, Gage FH, Schinder AF. Neurons born in the adult dentate gyrus form functional synapses with target cells. Nat Neurosci 11: 901–907, 2008. 312. Snyder JS, Soumier A, Brewer M, Pickel J, Cameron HA. Adult hippocampal neurogenesis buffers stress responses and depressive behaviour. Nature 476: 458 – 461, 2011. 331. Toni N, Sultan S. Synapse formation on adult-born hippocampal neurons. Eur J Neurosci 33: 1062–1068, 2011. 313. Song H, Stevens CF, Gage FH. Astroglia induce neurogenesis from adult neural stem cells. Nature 417: 39 – 44, 2002. 314. Song J, Zhong C, Bonaguidi MA, Sun GJ, Hsu D, Gu Y, Meletis K, Huang ZJ, Ge S, Enikolopov G, Deisseroth K, Luscher B, Christian KM, Ming GL, Song H. Neuronal circuitry mechanism regulating adult quiescent neural stem-cell fate decision. Nature 489: 150 –154, 2012. 315. Spalding KL, Bergmann O, Alkass K, Bernard S, Salehpour M, Huttner HB, Bostrom E, Westerlund I, Vial C, Buchholz BA, Possnert G, Mash DC, Druid H, Frisen J. Dynamics of hippocampal neurogenesis in adult humans. Cell 153: 1219 –1227, 2013. 332. Toni N, Teng EM, Bushong EA, Aimone JB, Zhao C, Consiglio A, van Praag H, Martone ME, Ellisman MH, Gage FH. Synapse formation on neurons born in the adult hippocampus. Nat Neurosci 10: 727–734, 2007. 333. Tozuka Y, Fukuda S, Namba T, Seki T, Hisatsune T. GABAergic excitation promotes neuronal differentiation in adult hippocampal progenitor cells. Neuron 47: 803– 815, 2005. 334. Trejo JL, Carro E, Torres-Aleman I. Circulating insulin-like growth factor I mediates exercise-induced increases in the number of new neurons in the adult hippocampus. J Neurosci 21: 1628 –1634, 2001. 335. Treves A, Rolls ET. Computational constraints suggest the need for two distinct input systems to the hippocampal CA3 network. Hippocampus 2: 189 –199, 1992. 316. Stangl D, Thuret S. Impact of diet on adult hippocampal neurogenesis. Genes Nutr 4: 271–282, 2009. 336. Treves A, Tashiro A, Witter MP, Moser EI. What is the mammalian dentate gyrus good for? Neuroscience 154: 1155–1172, 2008. 317. Stark SM, Yassa MA, Lacy JW, Stark CE. A task to assess behavioral pattern separation (BPS) in humans: data from healthy aging and mild cognitive impairment. Neuropsychologia 51: 2442–2449, 2013. 337. Tronel S, Fabre A, Charrier V, Oliet SH, Gage FH, Abrous DN. Spatial learning sculpts the dendritic arbor of adult-born hippocampal neurons. Proc Natl Acad Sci USA 107: 7963–7968, 2010. 318. Stepien AE, Tripodi M, Arber S. Monosynaptic rabies virus reveals premotor network organization and synaptic specificity of cholinergic partition cells. Neuron 68: 456 – 472, 2010. 338. Tsien RY. Fluorescence measurement and photochemical manipulation of cytosolic free calcium. Trends Neurosci 11: 419 – 424, 1988. 319. Stone SS, Teixeira CM, Zaslavsky K, Wheeler AL, Martinez-Canabal A, Wang AH, Sakaguchi M, Lozano AM, Frankland PW. Functional convergence of developmentally and adult-generated granule cells in dentate gyrus circuits supporting hippocampusdependent memory. Hippocampus 21: 1348 –1362, 2011. 339. Tsien RY. New calcium indicators and buffers with high selectivity against magnesium and protons: design, synthesis, and properties of prototype structures. Biochemistry 19: 2396 –2404, 1980. 340. Van Praag H, Christie BR, Sejnowski TJ, Gage FH. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc Natl Acad Sci USA 96: 13427–13431, 1999. 320. Succol F, Fiumelli H, Benfenati F, Cancedda L, Barberis A. Intracellular chloride concentration influences the GABAA receptor subunit composition. Nat Commun 3: 738, 2012. 341. Van Praag H, Kempermann G, Gage FH. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat Neurosci 2: 266 –270, 1999. 321. Suh H, Consiglio A, Ray J, Sawai T, D’Amour KA, Gage FH. In vivo fate analysis reveals the multipotent and self-renewal capacities of Sox2⫹ neural stem cells in the adult hippocampus. Cell Stem Cell 1: 515–528, 2007. 342. Van Praag H, Lucero MJ, Yeo GW, Stecker K, Heivand N, Zhao C, Yip E, Afanador M, Schroeter H, Hammerstone J, Gage FH. Plant-derived flavanol (⫺)epicatechin enhances angiogenesis and retention of spatial memory in mice. J Neurosci 27: 5869 – 5878, 2007. 322. Swanson LW, Cowan WM. An autoradiographic study of the organization of the efferent connections of the hippocampal formation in the rat. J Comp Neurol 172: 49 – 84, 1977. 343. Van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, Gage FH. Functional neurogenesis in the adult hippocampus. Nature 415: 1030 –1034, 2002. 323. Tan EM, Yamaguchi Y, Horwitz GD, Gosgnach S, Lein ES, Goulding M, Albright TD, Callaway EM. Selective and quickly reversible inactivation of mammalian neurons in vivo using the Drosophila allatostatin receptor. Neuron 51: 157–170, 2006. 324. Tan W, Janczewski WA, Yang P, Shao XM, Callaway EM, Feldman JL. Silencing preBotzinger complex somatostatin-expressing neurons induces persistent apnea in awake rat. Nat Neurosci 11: 538 –540, 2008. 325. Tashiro A, Makino H, Gage FH. Experience-specific functional modification of the dentate gyrus through adult neurogenesis: a critical period during an immature stage. J Neurosci 27: 3252–3259, 2007. 326. Tashiro A, Sandler VM, Toni N, Zhao C, Gage FH. NMDA-receptor-mediated, cellspecific integration of new neurons in adult dentate gyrus. Nature 442: 929 –933, 2006. 344. Van Praag H, Shubert T, Zhao C, Gage FH. Exercise enhances learning and hippocampal neurogenesis in aged mice. J Neurosci 25: 8680 – 8685, 2005. 345. Varga V, Losonczy A, Zemelman BV, Borhegyi Z, Nyiri G, Domonkos A, Hangya B, Holderith N, Magee JC, Freund TF. Fast synaptic subcortical control of hippocampal circuits. Science 326: 449 – 453, 2009. 346. Veena J, Rao BS, Srikumar BN. Regulation of adult neurogenesis in the hippocampus by stress, acetylcholine and dopamine. J Natural Sci Biol Med 2: 26 –37, 2011. 347. Vineyard CM, Aimone JB, Emmanuel GR. Neurogenesis in a high resolution dentate gyrus model. In: Biologically Inspired Cognitive Architectures 2012. New York: Springer, 2013, p. 371–372. 348. Vissing J, Andersen M, Diemer NH. Exercise-induced changes in local cerebral glucose utilization in the rat. J Cereb Blood Flow Metab 16: 729 –736, 1996. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org 1025 AIMONE ET AL. 349. Vivar C, Potter MC, Choi J, Lee JY, Stringer TP, Callaway EM, Gage FH, Suh H, van Praag H. Monosynaptic inputs to new neurons in the dentate gyrus. Nat Commun 3: 1107, 2012. 350. Vukovic J, Colditz MJ, Blackmore DG, Ruitenberg MJ, Bartlett PF. Microglia modulate hippocampal neural precursor activity in response to exercise and aging. J Neurosci 32: 6435– 6443, 2012. 351. Walker MC, Semyanov A. Regulation of excitability by extrasynaptic GABA (A) receptors. Results Probl Cell Differ 44: 29 – 48, 2008. 352. Wang DD, Kriegstein AR. GABA regulates excitatory synapse formation in the neocortex via NMDA receptor activation. J Neurosci 28: 5547–5558, 2008. 353. Wang LP, Kempermann G, Kettenmann H. A subpopulation of precursor cells in the mouse dentate gyrus receives synaptic GABAergic input. Mol Cell Neurosci 29: 181–189, 2005. 354. Waterhouse EG, An JJ, Orefice LL, Baydyuk M, Liao GY, Zheng K, Lu B, Xu B. BDNF promotes differentiation and maturation of adult-born neurons through GABAergic transmission. J Neurosci 32: 14318 –14330, 2012. 355. Weisz VI, Argibay PF. A putative role for neurogenesis in neuro-computational terms: inferences from a hippocampal model. Cognition 112: 229 –240, 2009. 356. Wexler EM, Paucer A, Kornblum HI, Palmer TD, Geschwind DH. Endogenous Wnt signaling maintains neural progenitor cell potency. Stem Cells 27: 1130 –1141, 2009. 357. Wickersham IR, Lyon DC, Barnard RJ, Mori T, Finke S, Conzelmann KK, Young JA, Callaway EM. Monosynaptic restriction of transsynaptic tracing from single, genetically targeted neurons. Neuron 53: 639 – 647, 2007. 358. Wilhelmsson U, Faiz M, de Pablo Y, Sjoqvist M, Andersson D, Widestrand A, Potokar M, Stenovec M, Smith PL, Shinjyo N, Pekny T, Zorec R, Stahlberg A, Pekna M, Sahlgren C, Pekny M. Astrocytes negatively regulate neurogenesis through the Jagged1-mediated Notch pathway. Stem Cells 30: 2320 –2329, 2012. 359. Williams CM, El Mohsen MA, Vauzour D, Rendeiro C, Butler LT, Ellis JA, Whiteman M, Spencer JP. Blueberry-induced changes in spatial working memory correlate with changes in hippocampal CREB phosphorylation and brain-derived neurotrophic factor (BDNF) levels. Free Radic Biol Med 45: 295–305, 2008. 360. Wirenfeldt M, Dalmau I, Finsen B. Estimation of absolute microglial cell numbers in mouse fascia dentata using unbiased and efficient stereological cell counting principles. Glia 44: 129 –139, 2003. 1026 361. Wiskott L, Rasch MJ, Kempermann G. A functional hypothesis for adult hippocampal neurogenesis: avoidance of catastrophic interference in the dentate gyrus. Hippocampus 16: 329 –343, 2006. 362. Wu W, Wong K, Chen J, Jiang Z, Dupuis S, Wu JY, Rao Y. Directional guidance of neuronal migration in the olfactory system by the protein Slit. Nature 400: 331–336, 1999. 363. Yamaguchi M, Saito H, Suzuki M, Mori K. Visualization of neurogenesis in the central nervous system using nestin promoter-GFP transgenic mice. Neuroreport 11: 1991– 1996, 2000. 364. Yancey SL, Overton JM. Cardiovascular responses to voluntary and treadmill exercise in rats. J Appl Physiol 75: 1334 –1340, 1993. 365. Yang P, Arnold SA, Habas A, Hetman M, Hagg T. Ciliary neurotrophic factor mediates dopamine D2 receptor-induced CNS neurogenesis in adult mice. J Neurosci 28: 2231– 2241, 2008. 366. Yassa MA, Mattfeld AT, Stark SM, Stark CE. Age-related memory deficits linked to circuit-specific disruptions in the hippocampus. Proc Natl Acad Sci USA 108: 8873– 8878, 2011. 367. Ye GL, Yi S, Gamkrelidze G, Pasternak JF, Trommer BL. AMPA and NMDA receptormediated currents in developing dentate gyrus granule cells. Brain Res 155: 26 –32, 2005. 368. Yonehara K, Balint K, Noda M, Nagel G, Bamberg E, Roska B. Spatially asymmetric reorganization of inhibition establishes a motion-sensitive circuit. Nature 469: 407– 410, 2011. 369. Yuste R, Peinado A, Katz LC. Neuronal domains in developing neocortex. Science 257: 665– 669, 1992. 370. Zhao C, Deng W, Gage FH. Mechanisms and functional implications of adult neurogenesis. Cell 132: 645– 660, 2008. 371. Zhao C, Teng EM, Summers RG Jr, Ming GL, Gage FH. Distinct morphological stages of dentate granule neuron maturation in the adult mouse hippocampus. J Neurosci 26: 3–11, 2006. 372. Zupanc GK. A comparative approach towards the understanding of adult neurogenesis. Brain Behav Evol 58: 246 –249, 2001. Physiol Rev • VOL 94 • OCTOBER 2014 • www.prv.org