LYMPHOCYTE RESPONSES TO THE POLYPHENOL, OENOTHEIN B, AND THE
INFLUENCE OF AGE ON THESE RESPONSES
by
Andrew Gordon Ramstead
A dissertation submitted in partial fulfillment
of the requirements for the degree
of
Doctor of Philosophy
in
Immunology and Infectious Diseases
MONTANA STATE UNIVERSITY
Bozeman, Montana
April 2014
© COPYRIGHT
by
Andrew Gordon Ramstead
2014
All Rights Reserved
ii
DEDICATION
For the family, friends, and teachers who encouraged me to pursue this goal.
iii
ACKNOWLEDGMENTS
I first have to thank my advisor, Dr. Mark Jutila, for all of the help provided every
step of the way. I also want to thank you for having the trust and confidence in me to
pursue my ideas and make this project my own. Thanks also to my graduate committee,
Dr. David Pascual, Dr. Jovanka Voyich-Kane, Dr. Mark Quinn, and Dr. Patrick Hatfield,
for all of your suggestions and advice along the way. I want to thank all of the members
of the Jutila lab, both past and present, for all of the assistance and support during my
time as a member of the lab. Thank you to Kimberly Todd, Kerri Jones, Dr. Jim
Berardinelli, Larissa Jackiw, Deann Snyder, the Bozeman Deaconess Hospital Labor and
Delivery staff, and the Johnson Family Livestock Facility staff, whose efforts provided
me with the samples needed to perform these experiments. Finally, I have to thank my
family, Mom, Dad, and Katie, who encouraged and supported me during this process
when things were going well and when times were difficult. Thanks to all of you.
Without any of you, this dissertation would not have been possible.
iv
TABLE OF CONTENTS
1. INTRODUCTION TO DISSERTATION .............................................................. 1
2. BACKGROUND .................................................................................................... 7
Innate Lymphocytes in Innate Immunity ................................................................ 7
Innate Lymphoid Cells ............................................................................................ 8
Natural Killer Cells ..................................................................................... 9
Effect of Aging on NK Cells .................................................................... 11
Bovine NK Cells ....................................................................................... 14
Effect of Aging on Bovine NK Cells ........................................................ 17
Innate T Cells ........................................................................................................ 19
Memory αβ T Cells ................................................................................... 20
Generation and Characterization of Memory αβ T Cells..................... 20
Innate Activity of Memory T cells....................................................... 22
Effect of Aging on Memory αβ T Cells .................................................... 23
γδ T Cells .................................................................................................. 25
Effect of Aging on γδ T Cells ................................................................... 32
Bovine γδ T Cells ...................................................................................... 34
Effect of Aging on Bovine γδ T Cells....................................................... 36
Plant-derived Agonist Activity on Innate Lymphocytes ...................................... 37
Immunotherapy Targeting Innate Lymphocytes ....................................... 37
NK Cells.................................................................................................... 38
αβ T Cells .................................................................................................. 39
γδ T Cells .................................................................................................. 41
Oenothein B .............................................................................................. 42
Factors Influencing Polyphenol Stimulation of Lymphocytes ................. 46
3. OENOTHEIN B, A CYCLIC DIMERIC ELLAGITANNIN ISOLATED
FROM EPILOBIUM ANGUSTIFOLIUM, ENHANCES IFNγ
PRODUCTION BY LYMPHOCYTES................................................................ 50
Contribution of Author and Co-authors ................................................................ 50
Manuscript Information Page ............................................................................... 51
Abstract ................................................................................................................. 52
Introduction ........................................................................................................... 52
Materials and Methods .......................................................................................... 55
Ethics Statement.......................................................................................... 55
Isolation of Oenothein B ............................................................................. 56
Human and Bovine Peripheral Blood Mononuclear Cell Preparations ...... 56
Flow Cytometric Analysis and Cell Sorting of
Bovine and/or Human PBMCs ................................................................... 57
v
TABLE OF CONTENTS CONTINUED
IL-18 Activation Assays ............................................................................. 58
K562 Assay ................................................................................................. 59
Measurement of IFNγ ................................................................................. 59
Statistical Analysis ...................................................................................... 60
Results and Discussion ......................................................................................... 61
Oenothein B Activates Human and Bovine Lymphocytes ......................... 61
Oenothein B Primes Bovine PBMCs to Respond to IL-18 ........................ 64
Presence of CD335+ Cells is Essential for
Oenothein B Priming to IL-18 .................................................................... 66
Oenothein B Induces IFNγ Production by Human Innate Lymphocytes ... 71
Acknowledgments................................................................................................. 79
4. AGING INFLUENCES THE RESPONSE OF T CELLS TO
STIMULATION BY THE ELLAGITANNIN, OENOTHEIN B ........................ 80
Contribution of Author and Co-authors ................................................................ 80
Manuscript Information Page ............................................................................... 81
Abstract ................................................................................................................. 82
Introduction ........................................................................................................... 83
Materials and Methods .......................................................................................... 85
Ethics Statement.......................................................................................... 85
Isolation of Oenothein B ............................................................................. 85
Human and Bovine Peripheral Blood Mononuclear Cell Preparations ...... 86
Cell Sorting of PBMCs ............................................................................... 86
T cell Activation Assays ............................................................................. 87
Flow Cytometric Analysis of Bovine and/or Human PBMCs .................... 88
Measurement of IFNγ and GM-CSF........................................................... 88
Statistical Analysis ...................................................................................... 89
Results ................................................................................................................... 90
Oenothein B Stimulates and Induces IFNγ
Production by Bovine Adult T Cells ........................................................... 90
Oenothein B Enhances IFNγ Production by Adult Bovine
CD45RO+ αβ and γδ T Cells ...................................................................... 91
Oenothein B Enhances IFNγ Production by Purified
Adult Bovine T Cells .................................................................................. 93
Differentiation of Calf T Cells into CD45RO+ T Cells is
not Sufficient to Enhance Oenothein B-induced IFNγ Production ............. 95
Oenothein B Enhances IFNγ Production by Human Adult T Cells, but
not Cord Blood T Cells ............................................................................... 99
Oenothein B Induces More Cytokine Production by Human Memory
T Cells Than Naïve T Cells ...................................................................... 102
Discussion ........................................................................................................... 104
vi
TABLE OF CONTENTS CONTINUED
Acknowledgments............................................................................................... 112
5. COMPLEX ROLE OF γδ T CELL-DERIVED CYTOKINES
AND GROWTH FACTORS IN CANCER ........................................................ 113
Contribution of Author and Co-author ............................................................... 113
Manuscript Information Page ............................................................................. 114
Abstract ............................................................................................................... 117
Introduction ......................................................................................................... 117
γδ T Cell-associated Factors Which Enhance Anti-tumor Immunity ................. 119
γδ T Cell-associated Factors Which Suppress Anti-tumor Immunity ................ 122
Conflicting Role of γδ T Cell-derived IL-17 on Tumor Immunity..................... 126
Potentially Underappreciated Role of γδ T Cell-derived Growth
Factors in Tumor Immunity ................................................................................ 128
Expression of Growth Factors in Human γδ T Cells ................................ 129
Influences on Differential Cytokine Secretion by γδ T Cells
in Tumor Studies ................................................................................................. 130
Conclusions ......................................................................................................... 131
Acknowledgments............................................................................................... 134
Author Disclosure Statement .............................................................................. 134
6. CONCLUSIONS................................................................................................. 135
7. FUTURE STUDIES............................................................................................ 140
REFERENCES CITED ............................................................................................. 147
vii
LIST OF TABLES
Table
Page
5.1 Select recent reviews for γδ T cells and cancer ...................................................... 120
6.1 Summary of oenothein B-induced IFNγ production by lymphocytes ...................137
viii
LIST OF FIGURES
Figure
Page
2.1 Pharmacological manipulation of mevalonate pathway and isopentenyl
pyrophospate accumulation. ..................................................................................... 29
2.2 Inhibition of B16 tumor growth by oenothein B in a γδ T cell-dependent
manner.. ..................................................................................................................... 45
2.3 Oenothein B reduced morbidity during influenza infection. ..................................... 47
3.1 Oenothein B induces IL-2Rα or CD69 on bovine and human lymphocyte
subsets. ..................................................................................................................... 62
3.2 Oenothein B induces IL-2Rα or CD69 on bovine and human NK cells. .................. 63
3.3 Oenothein B induces CD25 on human T cells. .......................................................... 64
3.4 Oenothein B primes bovine PBMCs to respond to IL-18. ......................................... 67
3.5 Effect of monocyte and γδ T cell depletion on oenothein B-priming of
bovine PBMCs. ........................................................................................................ 69
3.6 Oenothein B primes bovine CD335+ cells to respond to IL-18. ............................... 70
3.7 IFNγ production by human lymphocytes in response to oenothein B. ...................... 73
3.8 Priming of human NK cells to IL-18 by oenothein B. ............................................... 74
3.9 Priming of human NK cells to K562 cells by oenothein B........................................ 75
3.10 Oenothein B directly primes purified human NK cells to produce IFNγ ............... 76
4.1 Oenothein B induces IL-2Rα expression on bovine T cells from both
adults and calves. ....................................................................................................... 93
4.2 Oenothein B induces more IFNγ production by adult bovine T cells than
calf T cells. ................................................................................................................ 94
4.3 Oenothein B induces more IFNγ production by bovine CD45RO+ T cells
than CD45RO- T cells. .............................................................................................. 96
ix
LIST OF FIGURES CONTINUED
Figure
Page
4.4 Oenothein B enhances IFNγ production by purified bovine CD4+ T cells
from adults, but not calves ........................................................................................ 97
4.5 Enhancing CD45RO expression in vitro does not enhance oenothein Binduced IFNγ production by calf T cells.. ................................................................. 98
4.6 Oenothein B induces CD69 expression on human cord blood T cells. .................. 100
4.7 Oenothein B induces IFNγ expression on human adult, but not cord blood,
T cells.. .................................................................................................................... 101
4.8 Oenothein B induces GM-CSF expression on human adult, but not cord
blood, T cells. .......................................................................................................... 103
4.9 Oenothein B enhances more cytokine production by human CD45RO+
memory T cells than naïve T cells........................................................................... 105
5.1 Changes in B16 melanoma growth in γδ T cell-deficient mice compared
to wild-type mice over time..................................................................................... 116
5.2 Summary of the influence of γδ T cell-derived cytokines and growth
factors on tumor growth .......................................................................................... 133
7.1 5-Azacytidine does not enhance IFNγ production by oenothein Bstimulated bovine calves ......................................................................................... 141
7.2 Influence of oenothein B on NK cell degranulation in response to a tumor
cell line .................................................................................................................... 144
7.3 Oenothein B does not prime murine splenocytes to produce IFNγ in
response to IL-18 ..................................................................................................... 146
x
ABSTRACT
Innate lymphocytes, such as NK cells, play an important role in a number of human
diseases. Therefore, they have become popular target cells for new therapeutics. Studies
to identify materials that stimulate these cells have identified a number of molecules from
nutritional supplements which can activate and/or inhibit their activity. While the use of
nutritional supplements has potential to enhance the activity of innate lymphocytes to
help treat and prevent disease, there is still much which is currently unknown about the
activity of these compounds. Previous work has identified oenothein B, an ellagitannin
isolated from Epilobium angustifolium, to be a stimulant for myeloid cells. In this study,
we further examined the immunomodulatory effects of oenothein B and identified that it
also stimulates lymphocytes. We found that oenothein B induces the production of IFNγ
by NK cells and both αβ and γδ T cells. In addition to inducing the production of IFNγ by
NK cells alone, oenothein B also enhances IFNγ production in response to secondary
stimuli, such as IL-18 or a tumor cell line, in a synergistic manner. We also identified a
novel effect of aging on the lymphocyte response to oenothein B. The induction of IFNγ,
as well as GM-CSF, production by T cells in response to oenothein B increases in adults
compared to young individuals. The predominant producers of IFNγ in response to
oenothein B also shift from the NK cell population in young individuals to the T cell
population in adults. The increase in cytokine production in adult T cells correlates with
increasing numbers of CD45RO+ T cells, which produce more cytokines in response to
oenothein B than CD45RO- T cells. This influence of aging on the immune response to
oenothein B may be applicable to numerous nutritional supplements which stimulate
innate immunity and could be an important variable in the effectiveness of these
compounds in improving human health. In this dissertation, we identify age as an
important factor which should be taken into account when examining the effect of
polyphenols on innate immunity and could have important implications for optimizing
the benefit of immunomodulatory polyphenols.
1
CHAPTER ONE
INTRODUCTION TO DISSERTATION
A significant portion of research in immunology is directed towards the
identification of potential prophylactic treatments, which can enhance host defense
against infection. Many of these types of studies focus on vaccines, which expose the
host’s immune system to an antigen or antigens, stimulating a specific, long-lived
immune protection to a specific pathogen expressing those antigens. Vaccination is very
effective, but there are downsides to this type of approach for countering infectious
disease. Even in the best case, identification, testing, and applying a vaccine in humans
takes many years and cost 100s of millions of dollars1,2. Vaccines tend to be
monospecific, thus the time, effort, and funding has to be repeated for each pathogen that
new countermeasures are needed. Pathogens also mutate, which results in strains that are
resistant to previous vaccinations, such as with influenza and Hepatitis B3,4. Therefore,
new host strategies are needed for largely unforeseen and unknown pathogens at this
time, but ones that could be problematic in the future. The innate immune system
provides a broad spectrum of response to help counter a wide array of pathogens, and has
been targeted for the development of novel countermeasures against infectious disease.
Both prophylactic and post-exposure strategies involving innate immune stimulation have
been shown to be plausible to prevent or ameliorate infections in animal models5-7.
2
The innate immune system is the immune system’s first line of defense against
pathogens. It is sufficient for the clearance of many infections and its importance can be
seen with individuals who have deficiencies in specific components of their innate
immune system, as they become highly susceptible to infections8-10. An advantage of the
innate immune system is that it has multiple means of detecting pathogens that are far
less specific to any given pathogen when compared to the adaptive immune response.
These include pattern recognition receptors (PRRs), receptors that detect molecules
unique to microbes, as well as methods to detect cellular damage caused by pathogens11,
12
. This allows the innate immune system to rapidly respond to a broad range of
pathogens. By enhancing innate immune responses, protection against a broad spectrum
of pathogens could be strengthened. Hence, this strategy could potentially be used to
protect individuals from more than one pathogen for which there are no effective
vaccines.
One way innate immunity can be enhanced is through nutrition and numerous
plant-derived products are believed to enhance immunological health6, 13-15. Many studies
have investigated the role of nutritional supplements on immune function and a
significant percentage of this research has focused on plant polyphenols. Polyphenols are
plant metabolites important in plant defense against insects and other stresses. Many
nutritional supplements contain large amounts of polyphenols, and pre-clinical and
clinical studies suggest an overall health benefit for the consumption of these
3
compounds16-21. The major benefit ascribed to these compounds is anti-oxidant activity,
in which polyphenols bind to free reactive oxygen species (ROS)22. ROS molecules are
produced by numerous cells in the body to help combat infections; however, free ROS
has been described to promote cellular and DNA damage, leading to inflammation and
carcinogenesis23,24. Therefore, the consumption of polyphenols and other anti-oxidants
are believed to reduce inflammation and inhibit carcinogenesis.
In addition to their anti-oxidant properties, polyphenols are also thought to
modulate immune responses. Surveying the many studies on polyphenols, an interesting
contradiction in the data has emerged. Numerous studies describe an anti-inflammatory
property of polyphenols, demonstrating their benefit in multiple inflammatory disease
models19, 25, 26. In contrast, other studies describe an immune enhancing property for
polyphenols and these data suggest great potential for enhancing human and domestic
animal resistance to a variety of diseases through nutritional supplementation20, 27-30.
Interestingly, some compounds that are described as anti-inflammatory in some studies
are described as proinflammatory in others. For example, oligomeric procyanidins
isolated from unripe apple peels (APP) are capable of suppressing inflammation and
reducing pathology in a model of colitis, but are also capable of activating lymphocytes
and enhancing their proliferation and cytokine production19, 31, 32. Even within the colitis
model there is a dichotomy, as APP suppressed inflammatory cytokine production in
wild-type mice with colitis, but enhanced cytokine production in αβ T cell-deficient mice
4
with colitis19. Additional studies are needed which investigate factors that influence the
pro- and anti-inflammatory properties of these compounds, possibly skewing their
behavior toward one or the other. Results from such studies may help guide the
development of proper dosing regimens for polyphenols, maximizing their benefit while
minimizing potential deleterious effects.
In Chapter Two, background information relevant for this dissertation is provided.
This information describes several innate lymphocyte cell types and their role in a variety
of diseases in humans, mice, and cattle, focusing on γδ T cells, NK cells, and the innatelike activity of memory αβ T cells. The effects of aging on the distribution and activity of
these innate lymphocytes are also described. Finally, stimulation of these cells by plant
polyphenols, specifically oenothein B, a polyphenol extracted from the Epilobium
angustifolium plant, is characterized.
In Chapter Three, published data describing the influence of oenothein B on IFNγ
production by innate lymphocytes is presented. Previous work found that oenothein B
could stimulate macrophages, inducing the production of several cytokines28, 33.
However, the effect of oenothein B on lymphocytes had not been investigated. Certain
polyphenols, such as APP, promote lymphocyte proliferation and the production of
certain cytokines, such as Granulocyte macrophage colony-stimulating factor (GMCSF)32. However, little work has examined the ability for polyphenols to induce the
production of interferon-γ (IFNγ), an important cytokine produced by several innate
5
lymphocytes and critical for their ability to enhance host defense to a variety of different
pathogens and tumors. In this chapter, it is shown that, in addition to myeloid cells,
oenothein B also stimulates lymphocytes and primes bovine and human NK cells to
produce increased amounts of IFNγ in response to a cytokine, IL-18, or a tumor cell line.
Data is also provided which shows that oenothein B also stimulates human γδ T cells and
αβ T cells to produce IFNγ. These data adds to the collective knowledge of the
proinflammatory properties of oenothein B and provides an additional possible
mechanism for the proposed antitumor activity of this compound.
In Chapter Four, in a paper being submitted for publication, the influence of age
on immune cell responses to oenothein B is described. During studies of lymphocyte
stimulation by plant-derived polyphenols, including oenothein B, considerable variability
in responses from different human and bovine donors was observed. While this
variability could provide insights into which individuals could receive the most
immunological benefit from polyphenol-rich supplementation, the potential factors which
contribute to this variability are largely unknown. One possible variable which was
hypothesized to influence the response of innate immune cells to polyphenols is the age
of the individual.
Aging has a profound effect on the composition and effectiveness of the immune
system. This can be observed clinically, as the very young and very old are usually most
susceptible to infections. While aging is known to influence innate lymphocyte
6
distribution and responsiveness, the possible influence of age on immune cell
responsiveness to nutritional supplements is largely unknown. In this chapter, age is
shown to be an important variable in the response of human and bovine T cells to
stimulation by oenothein B. These data adds to the collective knowledge of the
immunostimulatory properties of polyphenols for individuals in different age groups and
suggest that age should be taken into account when examining the capabilities for certain
polyphenols to enhance immunity.
In Chapter Five, a review written on the complex role γδ T cells play in antitumor
responses is provided. This review focuses on the different cytokines and other soluble
factors produced by γδ T cells during antitumor responses and how these factors can have
conflicting roles in promoting or inhibiting tumor growth. This variable activity by γδ T
cells in tumors could influence the effectiveness of γδ T cell-stimulating polyphenols to
enhance antitumor responses and should be taken into account when examining the
effects of supplements on the antitumor activity of γδ T cells.
7
CHAPTER TWO
BACKGROUND
Innate Lymphocytes in Innate Immunity
Innate immunity encompasses a variety of phagocytic cells of the granulocytic
and myeloid lineages, such as neutrophils and macrophages. In contrast, cells of the
lymphoid lineage, such as T and B cells, are most often associated with adaptive
immunity. However, there are subsets of lymphocytes that respond rapidly to a variety of
pathogen-associated molecular patterns (PAMPs) and danger signals, participating as
first-line defenders of the immune system and are considered part of the innate immune
system.
The innate lymphocyte pool includes innate lymphoid cells (ILCs), innate T cells,
and innate B cells. Innate T cells include γδ T cells, NKT cells, and some memory αβ T
cells. Despite this diversity in cell types, many of these cells exhibit common behaviors.
These include the early, antigen-independent secretion of cytokines important for the
recruitment and activation of cells involved in both innate and adaptive immunity, as well
as the ability to lyse infected or malignant cells in an antigen-independent manner. The
roles of innate B cells during innate immunity have been reviewed elsewhere and are
beyond the scope of this dissertation34, 35. For this chapter, we will focus on two broad
categories of innate lymphocytes, ILCs and innate T cells.
8
Innate Lymphoid Cells (ILCs)
Innate lymphoid cells are innate immune cells of lymphoid lineage that are
distinguished from B and T cells due to their lack of somatic rearrangement of
immunoglobulin and T cell receptors (TCRs). They are widely distributed throughout the
body and are found in the blood, lymphatics, and tissues. One member of the ILC family,
NK cells, has been extensively studied over a number of years and is known to play an
important role in a wide variety of infections and other diseases. Other ILC populations
have only more recently been studied and there is still much that is currently unknown
about their development and activity, especially in humans. However, they appear to be
important at barrier surfaces of the body and are involved in lymphoid organogenesis,
tissue remodeling, antimicrobial immunity, and inflammation36-40.
ILCs have been categorized into several different subsets in recent years. These
subsets are based largely on their cytokine profile, which correlate with T helper (Th) cell
subsets. ILC1s are characterized by the expression of Th1 cytokines, such as IFNγ. ILC2s
express Th2 cytokines, such as IL-5 and IL-13, and are believed to play a role in allergic
inflammation and host defense against helminths. ILC3s express Th17 cytokines and
appear to be important for mucosal immunity by promoting the production of
antimicrobial peptides. The different subsets of ILCs and their roles in immunity have
been reviewed in detail elsewhere36-40. For this dissertation, we will focus on the best
studied member of the ILC family, the NK cell.
9
Natural Killer Cells
Human Natural Killer (NK) cells make up approximately 15% of peripheral blood
lymphocytes and are present in abundance in bone marrow, liver, lung, spleen, and, to a
lesser extent, secondary lymphoid tissues41-43. NK cells are identified in humans by the
expression of CD56 and lack of expression of CD3. In mice, they are identified by the
expression of markers NK1.1 or DX5. As the name suggests, NK cells were first
identified for their ability to kill (lyse) abnormal cells in an antigen-independent
manner44. NK cells lyse tumor cells, cells infected with viruses or bacteria, as well as
allogeneic cells in transplant patients, playing an important role in all of these
conditions45-47. NK cells are one of the body's primary defenders against cancer and low
NK-like cytotoxicity is associated with increased cancer risk, while high levels of tumor
infiltrating NK cells are associated with favorable outcomes in cancer patients48, 49.
The cytotoxic activity of NK cells is controlled by a balancing act of activating
and inhibitory signals mediated by a diverse and complex repertoire of receptors.
Inhibitory receptors on NK cells, which include many killer-cell immunoglobulin-like
receptor (KIR) family members in humans and Ly49 family members in mice, typically
recognize MHC class I (MHCI) expression on normal, healthy cells from their host.
MHCI expression is often downregulated on infected, malignant, and other stressed cells
("loss of self"), making them less capable of stimulating inhibitory receptors and more
susceptible to NK cell-mediated killing50, 51. In addition to loss of MHCI expression, NK
10
cells often do not recognize MHCI from allogeneic cells, an important obstacle in the
success of tissue and bone marrow transplants52.
In contrast to the loss of ligands which stimulate inhibitory receptors, infected and
malignant cells often express increased levels of ligands for activating NK receptors.
Some important NK cell activating receptors include the natural cytotoxicity receptor
family (NCRs), some Ly49 and KIR family members, LILR, CD2 family members,
NKG2 family members (except NKG2A), CD16, TRAIL, and FasL50,51,53. The activation
receptor NKG2D appears to play an especially important role in optimal recognition of
tumor and infected cells by NK cells and their subsequent activation54. NKG2D
recognizes the ligands MICA, MICB, and UL16 binding proteins (ULBPs) in humans, as
well as the ligands Rae1 and Mult1 in mice55.
When the balance of activating and inhibitory signals from a target cell favors
inhibition, the NK cell will leave the cell alone and move on to the next potential target.
When the balance of signals favors activation, the NK cell will proceed to kill the target
cell. Once a NK cell targets a cell for killing, the NK cell will induce apoptosis in the
target cell by several mechanisms, including perforin/granzyme-induced apoptosis, FasFasL-induced apoptosis, and TRAIL-induced apoptosis56. In addition to the killing of
tumor cells, NK cell cytotoxicity is important for clearance of intracellular pathogens, by
killing infected host cells and by direct antimicrobial activities57-59.
11
While the lytic activity of NK cells is very important for host defense, NK cells
have additional roles during immune responses. One of these roles is the rapid expression
of several cytokines upon activation, including IFNγ, which promotes the microbicidal
activity of macrophages and supports the recruitment and function of adaptive T cells,
such as cytotoxic T lymphocytes (CTLs). IFNγ secretion by NK cells is important for
optimal clearance of a variety of pathogens, including fungal (Cryptococcus neoformans,
Aspergillus fumigatus), bacterial (Francisella tularemia, Legionella pneumophila,
Staphylococcus aureus, etc.), viral (HSV, RSV, etc.), and protozoan pathogens57, 60. In
addition to its benefit during infectious disease, IFNγ expressed by NK cells restricts
tumor angiogenesis, the production of new blood vessels which feed the tumor, and
promotes adaptive antitumor immunity61, 62. The expression of IFNγ is critical for the
optimal benefit of NK cells in the defense against many tumors and infections, suggesting
that enhancing the production of this cytokine would benefit host defense in these types
of diseases60, 62.
Effect of Aging on NK Cells
Aging influences NK cell numbers, activity, receptor expression, and maturation.
First, while the NK cell percentage of lymphocytes does not change dramatically during
aging, the number of NK cells/mm3 of human blood is reduced dramatically from cord
blood to adult blood63. In general, NK cell function is skewed more towards cytokine
production than cytotoxicity in neonates compared to adults. Human cord blood NK cells
12
are less cytotoxic than adult NK cells, but can be induced to have similar cytotoxic
activity to adult NK cells by stimulation with IL-263. However, cord blood NK cells are
responsive and express more CD69 and IFNγ than adult NK cells in response to IL-12
and IL-18, more IFNγ in response to a cocktail of ssRNA, IL-2 and IFNα, and similar
IFNγ in response to a mitogen63-66. In addition to these in vitro studies, a study looking at
BCG vaccinated infants showed a significant contribution of NK cells to the IFNγ
response, being the most responsive cell type to ex vivo restimulation (based on IFNγ
production). However, during this same study, the authors found that CD8 T cells were
the primary IFNγ-producers in vaccinated adults, suggesting the impact of NK cells,
especially as it relates to IFNγ production, during BCG vaccination may decrease with
age67.
In addition to changes between neonates and adults, NK cell activity also changes
from young adults to the elderly. When comparing young human adults to older adults,
older NK cells have reduced activity, including reduced proliferation, IFNγ production,
and possibly cytotoxicity68. In a study in rats looking at the anti-metastatic activity of NK
cells, NK cells from prepubescent male and female rats had greater activity than cells
from mature rats, suggesting that age impacts the antitumor activity of NK cells69.
Altogether these data from cord blood, young adults, and older adults suggest that age is
an important variable when studying NK cell activity.
13
These age-related changes in human NK cell activity could partially be due to
changes in functional receptor expression important to these cells. The percentage of NK
cells expressing the activating receptor NKG2D increases from cord blood to adult blood,
while the percentage of cells expressing the inhibitory receptor NKG2A/CD94 decreases,
with the balance between NKG2A and activating NKG2 molecules skewed more towards
inhibition in neonates compared to adults70, 71. Other differences in surface receptor
expression between newborns and adults include the reduced expression of KIRs, LIR-1,
CD62L, CD54, CD8αα, granzyme B, and CD57 in newborns and reduced expression of
NKp46, NKp30, and CD161 in adults70, 71. The relatively high expression of the
inhibitory receptors NKG2A/CD94 and the lower expression of granzyme B in cord
blood NK cells could account for the reduced cytolytic activity of cord blood NK cells71.
Differential expression of FasL or TRAIL likely does not account for the reduced
cytolytic activity, as these molecules are expressed at similar levels on adult and cord
blood NK cells71. Furthermore, CD16 expression is similar between newborns and adults,
suggesting similar IgG-mediated antibody dependent cytotoxicity70. Altogether these
data suggest that age significantly changes receptor expression of NK cells, which could
lead to significant changes in functional activity.
Studies from mice suggest that the physiological environment may play an
important role in the reduced activity of NK cells in older individuals. NK cells from
aged mice do not expand and upregulate the expression of KLRG1 when stimulated in
14
vivo with PAMPs. However, when NK cells from old mice were adoptively transferred
into young mice, they regained responsiveness to PAMPs similar to NK cells from
younger mice. Conversely, when NK cells from young mice were transferred into older
mice, they had reduced activity compared to NK cells transferred back to young mice.
Immature CD27-expressing NK cells were more prevalent in the blood, spleen, and lungs
of aged mice compared to young mice. However, when bone marrow NK cells from aged
mice were transferred into young mice, they matured like young NK cells. Conversely,
bone marrow stem cells from young individuals did not have enhanced maturation when
transferred into aged mice. Treatment with IL-15/IL-15Ra brought the number of
Granzyme B and KLRG1-expressing NK cells in aged mice to similar levels as young
mice and brought target cell killing of aged NK cells back to similar levels as young
mice72. These data suggests that the reduced activity of NK cells in the elderly could
partially be due to reduced IL-15 expression in these individuals. A better understanding
of how the physiological environment in which NK cells reside changes with age could
provide insight into why NK cells from different age groups behave differently.
Bovine NK Cells
In addition to humans and mice, the role of NK cells in host defense has been
studied in other animal models, including cattle. It is believed that cattle can offer an
important complementary model for immunology and this could also be true for the study
of NK cells73. As with humans, NK cells play an important role in bovine immunology.
15
Bovine NK cells are identified by the absence of CD3 and the expression of CD335
(NKp46)74. While some T cells can express CD335, this marker is largely specific for NK
cells in cattle75. Bovine NK cells have been detected in the spleen, lung, liver, bone
marrow, a variety of lymph nodes, and blood, with NK cells usually ranging from 2-10%
of peripheral blood mononuclear cells (PBMCs)73. Cattle have similar numbers of lymph
node NK cells as humans, and more than what is found in mice76. This increase in lymph
node NK cells is believed to be the result of cattle and humans having greater microbial
exposure than mice, which are typically housed in cleaner environments, as microbial
exposure is believed to attract naive NK cells to the lymph nodes, where they acquire the
ability to respond to infections73. Bovine NK cells in the lymph nodes contain perforin
and can kill target cells, unlike human lymph node NK cells, which only acquire these
features after activation76, 77.
As with human and murine NK cells, bovine NK cells are cytotoxic and appear to
be an important innate source of IFNγ in cattle in response to several important bovine
pathogens. For example, bovine NK cells are stimulated by the mycobacterial proteins
ESAT-6 and MPP14, BCG-infected DCs, and M. bovis-infected macrophages to produce
IFNγ and kill infected cells in a cell contact-dependent manner78-80. In addition to
mycobacterium, bovine NK cells directly recognize the protozoan Neospora caninum and
purified, IL-2 stimulated NK cells produced IFNγ and killed infected fibroblasts81. NK
cells are also the major early IFNγ-producing cell in response to another important
16
protozoal infection, Babesia bovis82. The production of IFNγ by bovine NK cells can be
stimulated by several mechanisms, including contact with infected cells, cytokines (such
as IL-12), and by direct contact with the pathogen78, 81, 82. Although IFNγ production and
cytotoxicity appear to be important for NK cell-mediated host defense to many bovine
pathogens, these cells can also mediate host defense through other, yet not well
understood, mechanisms. For example, bovine NK cells reduce M. bovis replication in
macrophages in a cell contact-dependent manner which does not appear to be mediated
by IFNγ or the killing of infected macrophages80. Further studies are needed to elucidate
the full repertoire of effector functions available to bovine NK cells to protect hosts from
infection.
While receptor expression on bovine NK cells has not been studied as thoroughly
as in mice or humans, several studies have examined NK receptor expression in cattle73.
KIR genes have been identified in cattle and these KIRs appear to include both activating
and inhibitory receptors. These bovine KIRs comprise three Ig-domains and one ITIM
sequence, making them different from human KIRs, which typically have two Igdomains and two ITIM sequences83. It has yet to be determined whether or not bovine
KIRs bind to MHCI. In cattle, there are several NKG2A genes and one NKG2C gene, but
their functions are still unknown84. In addition, receptors for the MIC protein are found
on bovine NK cells and recombinant MIC activated these cells, suggesting that, as in
humans, bovine NKG2D is an activating receptor which recognizes stress ligands85. A
17
single Ly49 gene has also been found in cattle, but its ligand and possible function is
unknown83. While these studies suggest that bovine NK cell activity may be controlled in
a similar manner to those in mice and humans, additional studies are needed to examine
the control of NK cell activity in cattle. This knowledge could then be used to better
understand NK cell activity during infection in cattle.
Effect of Aging on Bovine NK Cells
Bovine NK cells have been studied in newborn calves through to adults, and their
numbers and activity changes during aging. One study found that NK cells are present at
a lower frequency in neonate PBMCs compared to older calves and adults, but that the
percentage of NK cells in PBMCs increases to older calf/adult levels after a couple of
weeks86. In another study, the numbers of CD335+ NK cells in the blood varied
significantly during the first few months of life, with NK cell numbers increasing over
the first 10-12 weeks then decreasing over the following 10 weeks87. The numbers then
underwent another rise and fall over the next 10 weeks, suggesting that calf NK cell
numbers can be highly unstable in the blood. In the intestinal track of cattle, NK cell
numbers increased with age in the jejunum over the first six months88. In general, it
appears that NK cell numbers in cattle increase with age from birth to young adulthood.
Although NK cell numbers are lower in very young calves, several studies
suggest that NK cells from neonate calves are functional and likely play a critical role in
host protection for calves during their first few weeks. In one study, NK cell activity was
18
compared between neonate (less than 8 days old) and older calves, and while neonate
calves had approximately one-third of the circulating CD335+ cells that the older calves
had, the NK cells from neonates proliferated more robustly in response to IL-2 or IL-15
and produced more IFNγ than NK cells from older calves after culturing in IL-15 and
stimulation with IL-12 and an antibody to NKp4689. Furthermore, BCG vaccination
works best when given to neonates compared to adults and it is believed that this is the
result of enhanced responses of neonatal NK cells, as well as other innate immune cells90.
This enhanced response by neonatal NK cells to BCG may be mediated through direct
interaction with BCG, DC-derived cytokines, or by cell-to-cell interactions with infected
DCs, as described earlier.
As with humans, aging may also affect the cytotoxic potential of bovine NK cells.
A greater percentage of bovine NK cells express perforin in young calves (1-6 weeks)
compared to adults. In calves less than or equal to 6 weeks old, the majority of perforin+
cells were CD335+ NK cells. However, in adults, the majority of perforin+ cells were
CD335- cells, likely T cells86. In addition, IL-15 expanded NK cells from neonates (less
than 8 days old) were more effective at killing a bovine epithelial tumor cell line than
those from older calves89. These data suggest that NK cell cytotoxicity may decrease with
age in young calves. This contradicts the previously described data seen in humans,
where cytotoxicity increased in adults compared to newborns. However, those human
studies were performed using cord blood NK cells, which may not fully reflect the
19
activity of peripheral NK cells in very young individuals. Altogether, these data suggest
that NK cells from very young calves are more responsive to certain stimuli than those
from older animals and that NK cell activity can change rapidly in cattle during aging,
even from neonate to older calves.
Innate T Cells
T cells are a major lymphoid cell population and represent probably the most
studied cell in the immune system. While they are typically associated with antigenspecific adaptive immunity, some subsets of T cells respond rapidly in an antigenindependent fashion during innate immune responses. Among these T cell subsets include
NKT cells, γδ T cells, and subsets of memory αβ T cells. Briefly, NKT cells are innate T
cells found in blood and various tissues which recognize lipids presented by CD1d, a
member of the CD1 family of MHC-like molecules, and are known for producing large
quantities of cytokines, including Th1, Th2, and Th17 cytokines. They play a prominent
role in the immune responses to several bacteria and also appear to have a role in viral
infections, tumor immunology, and perhaps some allergic and autoimmune disorders.
NKT cells and their role in disease have been reviewed in detail elsewhere91. For this
dissertation, we will focus on memory αβ T cells and γδ T cells.
20
Memory αβ T Cells
Generation and Characterization of Memory αβ T Cells
αβ T cells make up the largest proportion of T cells and express a TCR consisting
of the α and β chain. After development in the thymus, αβ T cells enter the periphery as
naive, mature T cells which traffic back and forth from the blood to lymph nodes,
mucosa-associated lymphoid tissues, and the spleen. These naïve T cells proliferate upon
activation, but do not have fully developed effector functions, such as cytotoxicity and
cytokine production. During a primary immunological insult, DCs ingest, process, and
transport antigens to regional lymph nodes, where they present those antigens on MHC
molecules to T cells in the T cell cortex. CD4+ T cells recognize antigenic peptides
presented on MHCII, while CD8+ T cells recognize peptides presented on MHCI. Naive
T cells, which recognize a presented antigen, will proliferate and differentiate into
effector T cells. In general, CD4+ T cells differentiate into cytokine producing helper T
cells (Th), which support or suppress other effector T cell functions and antibody
production, while CD8+ T cells differentiate into cytotoxic T lymphocytes (CTLs), which
produce IFNγ and kill target cells expressing their cognate antigen on MHCI92.
Depending upon the immunological environment, CD4+ T cells can differentiate into one
of four major Th subsets. The four major Th subsets are: Th1, which produce IFNγ and
promote macrophage and CTL activity, Th2, which produce IL-4, IL-5, and IL-13 and
promote allergic responses and helminth clearance, Th17, which produce IL-17 and
21
promote neutrophil recruitment and inflammation, and Treg, which produce IL-10 and
suppress various immune functions93.
After differentiation, some effector T cells traffic to the periphery and tissues,
where they help mediate pathogen clearance, while others remain in the blood and
lymphatic system. Upon clearance of the pathogen, the vast majority of the responsive
effector T cells undergo apoptosis. However, a subpopulation of these cells differentiates
into long-lived memory T cells. These memory T cells are able to rapidly respond to a
secondary exposure by their cognate antigen, providing enhanced protection to secondary
infections. Memory T cells can be divided into two major subsets, central memory T cells
(Tcm) and effector memory T cells (Tem). Tcm cells express the lymph node homing
receptors CD62L and CCR7, and are primarily found within the blood, bone marrow, and
secondary lymphoid organs. They produce lower amounts of effector cytokines (IFNγ,
IL-4, etc.) compared to Tem cells, but have longer telomeres and express more IL-2,
suggesting that they are important in the long-term maintenance of memory T cell pools.
Tem cells, on the other hand, lack the expression of CD62L and CCR7, and can be found
in nonlymphoid tissues, as well as the blood. They also have greater cytotoxicity and
effector cytokine production than Tcm cells94-96. While many Tem cells recirculate, some
take up permanent residence in tissues. These tissue resident memory T cells (Trm) are
characterized by the expression of CD103 and their tissue residence is promoted by the
downregulation of S1P1, the receptor for sphingosine-1-phosphate, which prevents them
22
from recirculating. This downregulation of S1P1 is promoted by TGFβ, IL-33, and TNF94,
97
. Many Trm cells localize to portals of entry into the body, such as the lung and
intestinal tract, which places them in position to respond to the environment, allowing
them to respond rapidly to secondary exposure with their cognate antigen upon its entry
into the body through the mucosa94. However, in addition to these antigen-specific
responses, they can also participate during innate immune responses in an antigenindependent, bystander manner.
Innate Activity of Memory T Cells
Antigen-independent innate immune responses by memory T cells have been
demonstrated in both mouse models and clinical studies. In humans, CD8+ memory T
cells specific for influenza, CMV, and EBV are activated during an early HIV infection98.
The activation of memory T cells during these bystander responses is antigenindependent, but is dependent upon inflammation. Among the important bystander effects
is the production of IFNγ in response to innate IL-12, IL-18, and other cytokines in the
absence of cognate antigen99, 100.
Memory CD8+ T cells also express many receptors typically associated with NK
cells. One of these NK receptors, NKG2D seems to be of particular importance during
bystander memory CD8+ T cell responses. Exposure to IL-12, IL-15, and IL-18 could
promote the development of cytotoxic CD8+ T cells from the memory T cell pool in the
absence of TCR signaling and these cells could kill infected cells in an NKG2D-
23
dependent manner. This process was shown to be important for early pathogen clearance
during Listeria monocytogenes infection in mice101. Furthermore, during influenza
infection in mice, non-specific memory T cells rapidly expanded and upregulated the
expression of NKG2D. Neutralizing NKG2D enhanced infection, suggesting that
memory T cells could also inhibit influenza infections in an NKG2D-dependent
manner102.
Altogether, these data suggest that conventional memory T cells contribute to
innate responses and should be considered members of the innate lymphocyte pool,
participating in innate responses by cytokine production and cell contact-dependent
killing of infected cells. It is currently not well understood just how important these
bystander effects are to innate immunity. However, they may play a critical role in
response to certain pathogens and further studies are needed to better understand the
importance of antigen-independent innate responses by memory T cells. These responses
could be very important in human immunology, as memory T cells make up a large
percentage of the human T cell pool, especially in adults.
Effect of Aging on Memory αβ T Cells
As with NK cells, aging has a profound effect on the memory T cell pool. As
individuals age, they become more exposed to various pathogens and other antigens,
which leads to an increased accumulation of memory T cells in older individuals103. In
young individuals, the majority of αβ T cells express naive cell markers, while in older
24
individuals, more T cells express the effector and memory T cell marker CD45RO104.
This expansion of memory T cells could potentially lead to an increase in antigenindependent responses by T cells in older individuals compared to younger ones during
infection. In support of this, a study comparing IFNγ production in cord blood and adult
human PBMCs to the parasite Trypanosoma cruzi and IL-15 found that IFNγ production
was exclusive to the NK cell population in neonates, but in adults, CD56+ and CD56- T
cells also produced early IFNγ105. Additional studies are warranted which examine how
innate immune responses to different pathogens change with age due to an increase in
memory T cell populations.
In addition to increased antigen exposure, aging also appears to promote the
development of memory T cells by additional mechanisms. Reconstituted T cells from
bone marrow transplants have a memory-dominated profile and produce IL-4, IL-5, and
IFNγ in aged recipient mice, but not younger recipient mice106. Furthermore, virtual
memory T cells, which are antigen-inexperienced T cells expressing memory T cell
markers, also expand with age107. Homeostatic cytokine expression may contribute to
these age-related microenvironmental effects on memory T cells. For example, IL-15
promotes virtual memory T cell development, as well as conventional memory T cell
proliferation and survival, but can also lead to the accumulation of highly activated,
hyporesponsive CD28- memory T cells in older individuals108, 109. Further studies are
necessary to identify additional homeostatic factors influencing the development and
25
activity of memory T cells in older individuals, as well as how these factors may
influence innate responses by memory T cells.
γδ T Cells
γδ T cells are an innate T cell subset, which express γ and δ TCR chains, and are
thought to play a role in the bridging of innate and adaptive immunity110. Their
contribution to T cell numbers in the blood is species-specific, as they are found in low
numbers in the blood of rodents and humans, but are found in higher numbers in the
blood of cows, sheep, and other hoofed animals111. The reason for this species variability
in γδ T cell levels in the blood is not entirely known. In addition to being found in the
blood, γδ T cells are found in high numbers in mucosal tissues at portals of entry into the
body, including the intestines, lungs, and genitourinary tract, and make up a large
percentage of intraepithelial lymphocytes (IELs) in the intestines of multiple species112.
Therefore, they are positioned to be among the first responders to invading pathogens.
Subsets of γδ T cells are primarily identified by their TCR diversity. In humans,
the primary subsets are the Vδ2 and Vδ1 subsets.Vδ2 γδ T cells make up the primary
subset in the blood, while Vδ1 γδ T cells make up the primary subset in the mucosal
tissues113, 114. In addition to TCR expression, subsets of γδ T cells in humans can be
further distinguished by the expression of CD45RA and CD27, which divide γδ T cells,
both Vδ2 and Vδ1, into four distinct subsets115. CD27+ CD45RA+ cells are described as
26
naive cells that have limited effector activity in terms of cytokine production and
cytotoxicity, but have potential for proliferation and differentiation. Upon stimulation
with IL-2 and the γδ T cell agonist isopentenyl pyrophosphate (IPP), these cells
differentiate into CD27+ CD45RA- cells. These cells are described as central memory
cells and have the greatest proliferation potential, as well as some cytokine producing and
cytotoxic activity. At this point, central memory cells can further differentiate into one of
two subsets. Upon additional stimulation with IL-2 and IPP, these cells differentiate into
CD27- CD45RA- cells, which are described as effector memory cells. They have less
proliferative potential than central memory γδ T cells, but have the most robust IFNγ
production of the four subsets. These cells appear to play an important role in immune
responses to M. tuberculosis, as TB-infected individuals have a lower percentage of this
subset compared to healthy controls and individuals with active TB infections have lower
percentages of this subset than those with inactive disease116. Individuals infected with
HIV also have reduced percentages of this subset. If central memory γδ T cells are
stimulated with IL-15 instead of IL-2, they differentiate into CD27- CD45RA+ cells.
These cells are terminally differentiated and have no proliferative capacity, but can
produce some IFNγ and have the greatest cytotoxic activity of the four subsets115.
Like NK cells, γδ T cells identify stressed or infected cells through a variety of
receptors111. These receptors include the γδ TCR itself, which recognizes proteins and
small molecules on the surface of target cells. While ligands for γδ TCRs are still being
27
identified, the Vδ2 subset responds to non-peptide phosphoantigens, which can originate
from endogenous or exogenous sources, in a γδ TCR-dependent manner117. Numerous
pathogens express phosphoantigens through the non-mevalonate biosynthesis pathway
and infection by pathogens that produce high levels of these phosphoantigens, such as M.
tuberculosis, results in a large expansion of Vδ2+ γδ T cells118, 119. In addition to
exogenous phosphoantigens, γδ T cells can be stimulated by the endogenous
phosphoantigen, isopentenyl pyrophosphate (IPP), which is a substrate in the mevalonate
pathway of biosynthesis119. When a cell is stressed, such as during malignancy or
infection, IPP accumulates and is then presented on the surface of the cell. The
recognition of infected or malignant cells by γδ T cells relies heavily on the recognition
of IPP presented on the surface of these target cells119, 120. Recognition of these
phosphoantigens by the γδ TCR requires antigen presentation and cell-to-cell contact;
however, professional antigen presenting cells are not necessary and many cells,
including γδ T cells themselves, can present phosphoantigens to the γδ TCR121. This
antigen presentation does not involve well described antigen presentation molecules, such
as members of the MHC or CD1 family, but instead appears to involve the Ig superfamily
protein, Butyrophilin 3A1121-123. The understanding of phosphoantigen presentation to γδ
T cells by Butyrophilin 3A1 is still in its infancy and is in the process of being better
understood.
28
In addition to infection and cellular stress, the accumulation of IPP can be
manipulated pharmacologically, allowing for the manipulation of human Vδ2 γδ T cell
activity (Figure 2.1). Bisphosphonates, which are used to prevent the loss of bone mass
during osteoporosis, inhibit the enzyme Farnesyl Pyrophosphate Synthase, which
promotes the conversion of IPP to downstream products in the mevalonate pathway. The
inhibition of this enzyme leads to the accumulation of IPP, which can then activate γδ T
cells124. In contrast, statins can prevent the accumulation of IPP by inhibiting the activity
of the enzyme HMG-CoA reductase, which is found upstream of IPP synthesis in the
mevalonate pathway. Statins have been shown to inhibit γδ T cell activation by
bisphosphonates, as well as by influenza-infected cells120, 125. Considering how many
patients take statins, the influence of this treatment on γδ T cell activity during infections,
cancer, and other inflammatory diseases should be further examined.
In addition to the γδ TCR, γδ T cells also express various other receptors which
contribute to their activation, such as NK receptors and various PRRs126-130. The NK
receptors NKG2D and the NCRs contribute to the recognition of tumor cells by γδ T cells
and the recognition of these NK receptors to their ligands can activate γδ T cells, as well
as act as co-receptors, enhancing γδ TCR-mediated responses127-130. Stimulation of
NKG2D on γδ T cells promotes degranulation and NKG2D is essential for the optimal
cytotoxic activity of human γδ T cells129, 130.
29
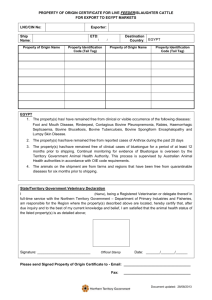
Figure 2.1 Pharmacological manipulation of mevalonate pathway and isopentenyl
pyrophospate accumulation. Statins inhibit HMG-CoA redutase, preventing the
conversion of 3-hydroxy-3-methylglutaryl-CoA to mevalonic acid and inhibiting the
production of isopentenyl pyrophosphate (blue). Bisphosphonates inhibit farnesyl
diphosphate synthase, resulting in an accumulation of isopentenyl pyrophosphate.
30
γδ T cells are believed to contribute to pathogen clearance during many
infections, including viral (influenza, CMV, HIV, etc.), bacterial (Staphylococcus aureus,
E. coli, Mycobacterium tuberculosis, etc.), and protozoan (Leishmania, Toxoplasma
godii) infections131. Like NK cells, γδ T cells can directly lyse infected cells in a cell
contact-dependent manner which can be mediated by the Fas-FasL, TRAIL, and
perforin/granzyme B pathways. All three of these pathways were shown to be involved in
the killing of influenza-infected human lung alveolar epithelial cells by Pamidronateactivated γδ T cells132. The antibody-dependent cell-mediated cytotoxicity (ADCC)
pathway, which is induced by the recognition of pathogen-specific antibodies by CD16,
can also enhance γδ T cell activity133.
In addition to their cytotoxic activity, γδ T cells also produce a variety of
cytokines and chemokines in response to infections, which is important for their
antimicrobial activity. IFNγ production by γδ T cells inhibits HCMV and influenza virus
propagation and is also believed to be critical in host resistance to Plasmodium
falciparum134-136. In addition to IFNγ, IL-17 and TNFα produced by γδ T cells promotes
the elimination of various bacteria and Plasmodium falciparum137-139. Finally, IL-17, as
well as IL-22, produced by γδ T cells induces the production of antimicrobial peptides
and recruits neutrophils to the site of infection, as seen in several infection models140-143.
Therefore, γδ T cells may be critical in promoting the optimal production of antimicrobial
peptides and recruitment of neutrophils to infected tissues.
31
In addition to enhancing innate immunity, γδ T cells also enhance the
development of adaptive immunity in several ways. They promote the maturation of DCs,
facilitate the development of αβ T cells, kill Tregs, and even migrate into secondary
lymphoid tissues to act as B helper T cells144-147. γδ T cells also have demonstrated
professional antigen-presenting cell activities and are capable of taking up, processing,
and presenting antigen from free viral particles and infected cells to both CD4 and CD8
αβ T cells148, 149. Certain studies even suggest that γδ T cells may be more efficient than
other APCs at cross-presentation of antigen to CD8+ cells148. These data support the idea
that γδ T cells serve as an important bridge between the innate and adaptive immune
response.
After pathogen clearance, γδ T cells may also be beneficial in the resolution of
inflammation and tissue recovery. In a murine model of influenza, γδ T cells were
prevalent at the site of inflammation late during infection, after clearance of virus,
suggesting that these γδ T cells were involved in the resolution of inflammation in this
model150. Consistent with this, studies from the early 2000's demonstrated that skin γδ T
cells in mice, known as dendritic epidermal T cells (DETCs), were necessary for optimal
wound healing, which was promoted by the production of Keratinocyte Growth Factor 7
(KGF-7) by these cells151. In addition, γδ T cells in the intestine of mice protect against
dextran sodium sulfate-induced inflammatory bowel disease and promote epithelial
turnover152. Human γδ T cells are also capable of producing several growth factors and
32
immunoregulatory proteins153, 154. These data suggest that γδ T cells, in addition to their
proinflammatory roles, may also promote tissue recovery and resolution of inflammation
in humans and animals.
In addition to infectious disease and tissue healing, γδ T cells also play an
important, but complex, role in cancer. γδ T cells lyse tumor cells from a variety of
cellular origins129. However, secreted factors produced by γδ T cells can either be
beneficial or detrimental to tumor responses and outcomes. The complex role γδ T cells
can play in response to cancer will be discussed in further detail in Chapter 5.
Effect of Aging on γδ T Cells
γδ T cells appear to mature throughout the lifetime of an individual until fully
differentiated effector cells dominate the γδ T cell pool in older individuals. In support of
this, a study in 2004 identified a naive subset of γδ T cells, which was described as
CD45RO-, CD27 bright, and CD11a dull. This subset made up the majority of both the
Vδ2 and Vδ1 γδ T cell subsets in cord blood and infants, but the presence of this subset
decreased with age and by approximately one year of age, it was only a small percentage
of Vδ2 cells, although it was still a large percentage of Vδ1+ γδ T cells. By adulthood, it
was only a small subset of both Vδ2+ and Vδ1+ γδ T cells155. Another study found that
the percentage of γδ T cells expressing CD27, identifying naïve and central memory γδ T
cells, decreased with age from young adult to older adults, while the percentage of CD27γδ T cells increased in these older individuals, further supporting the concept of
33
increasing maturation of γδ T cells over a lifetime156. It is not entirely known what drives
this maturation, but it could potentially be mediated by homeostatic cytokines, as well as
by increasing exposure to exogenous and endogenous γδ TCR ligands.
In addition to different maturation subsets, the percentage of γδ T cells in the
blood expressing the Vδ1 γδ TCR changes with age. Vδ1 γδ T cells make up a large
percentage of the peripheral γδ T cell population at the time of birth. However, over time,
the Vδ2 subset expands and becomes the predominant peripheral subset, while the
number of total Vδ1+ γδ T cells in blood decreases from cord blood to child to adult 155.
These data suggest that Vδ2+ cells expand more robustly than Vδ1+ cells over the
lifetime of an individual and this is believed to be due to exposure of phosphoantigenproducing pathogens.
As can be expected from the described subset changes in the γδ T cell pool with
aging, γδ T cell functions, including cytotoxicity, cytokine production, and proliferation
is also affected by aging. γδ T cells from elderly individuals express greater levels of the
activation marker CD69 than γδ T cells from young or middle age individuals, suggesting
a more activated phenotype, possibly a result of chronic inflammation found in the
elderly, a phenomenon known as inflammaging156. γδ T cells from infants express much
lower levels of perforin than adult γδ T cells, suggesting a reduced capability to lyse
target cells. IFNγ production by γδ T cells increases with age, as γδ T cells from children
had enhanced production compared to cord blood γδ T cells, but reduced compared to
34
adult γδ T cells155. Finally, the expansion of γδ T cells in response to phosphoantigens
and IL-2 decreases with age, except for the CCR7- subset of γδ T cells156. In summary,
maturation of γδ T cells during aging leads to differential effector functions, with IFNγ
production and cytotoxicity increasing with age, but proliferation decreasing with age.
Bovine γδ T Cells
Unlike in humans and mice, γδ T cells make up a large percentage of PBMCs in
cattle, especially in young calves. This is similar to other hoofed animals, such as sheep,
bison, and goats111. Bovine γδ T cells are broadly divided into subsets based on the
expression of WC1, which can be further characterized by three isoforms of WC1,
WC1.1, WC1.2, and WC1.3, with WC1.1 and WC1.2 expressed on largely nonoverlapping populations157. WC1+ bovine γδ T cells are found at high numbers in the
blood and accumulate at inflammatory sites, while WC1- γδ T cells accumulate in
mucosal tissues and splenic red pulp158. Overall, WC1- γδ T cells are believed to be more
immunoregulatory, while WC1+ cells are believed to be more inflammatory159, 160.
Unfortunately, the ligands recognized by the bovine γδ TCR have not been well
characterized, as bovine γδ T cells have minimal responses to the best known human γδ T
cell ligands, the phosphoantigens. However, many studies have examined the responses
of these cells to other stimulants. Bovine γδ T cells express a number of different PRRs,
including TLRs, scavenger receptors, C-type lectin receptors, and NOD receptors, which
allow bovine γδ T cells to respond to numerous microbial products111. For example,
35
stimulation with muramyl dipeptide, the NOD2 agonist, promoted proliferation and GMCSF expression of bovine γδ T cells161. Bovine γδ T cells may also be activated by NK
cell receptors. Transcripts specific for NKG2D, as well as additional NK receptors,
including KIR2DL1 and 2B4 (CD244), were measured in a bovine γδ T cell clone162.
Furthermore, a subset of bovine γδ T cells from the mesenteric lymph nodes and ileum of
cattle bind bovine MIC proteins and produce IFNγ in response to stimulation by MIC,
likely through NKG2D85. Finally, the NCR CD335, typically used as an NK cell marker,
can be upregulated on bovine γδ T cells in vivo by viral infection and CD335+ γδ T cells
produced robust amounts of IFNγ in response to IL-12 and IL-1875, 163.
Like NK cells, bovine γδ T cells appear to be important in the host response to
several important bacterial and viral pathogens to cattle. They are early producers of
IFNγ upon exposure to M. bovis, Brucella abortus, foot-and-mouth-disease (FMD), and
possibly salmonella enterocolitis, which activates infected macrophages111, 163. In
addition to the production of IFNγ upon exposure to FMD, γδ T cells also upregulate
CD335, perforin, and MHCII, lyse FMD-infected cells, and present antigen163. Despite
these data showing beneficial responses by bovine γδ T cells to counter infectious
disease, they may not always be beneficial during infection. For example, despite being
shown to produce IFNγ in response to M. bovis, bovine γδ T cells appear to be regulatory
during established M. bovis infections, as depletion of γδ T cells results in increased
antigen-specific proliferation of αβ T cells164. Therefore, further analysis of the role of γδ
36
T cells during infection in cattle is necessary to determine what drives an inflammatory or
regulatory γδ T cell phenotype during infection.
Effect of Aging on Bovine γδ T Cells
In cattle, γδ T cell numbers and activity has been compared between adults and
young calves. In young calves, γδ T cells can be close to 60% of peripheral mononuclear
cells165. During the first 5 months of life, the percentage of γδ T cells decreases slightly,
but absolute numbers stay relatively constant87. By 150 days of age, these percentages
decrease to adult levels (10-20%), but are still much higher than percentages in mice or
humans166. Despite decreases in percentages, absolute γδ T cell numbers in the blood
remain fairly constant from calf to adult167. In the small intestine, however, γδ T cells
numbers increase significantly with age88.
In addition to overall percentages, subsets of bovine γδ T cells in the peripheral
blood also change with aging. The percentage of CD8+ γδ T cells increases with age168.
Furthermore, in young calves, the WC1.1 population is the dominant population of
WC1+ γδ T cells. However, the percentage of WC1+ γδ T cells expressing the WC1.2
isoform increases with age until the percentage of cells expressing WC1.1 or WC1.2 is
about even by 500 days of age169. This change in WC1 subset distribution also affects γδ
T cell activity. In a 2007 study, IFNγ production was induced in WC1+ γδ T cells from
both adults (more than 2 years of age) and calves (less than 6 weeks old) by the use of IL12 and IL-18. The authors found that the percentage of WC1+ cells producing IFNγ was
37
increased in the calves compared to the adults by almost 10 fold170. This is consistent
with a decreasing percentage of WC1.1 γδ T cells in adults, as these cells produce more
IFNγ than WC1.2 γδ T cells169. Consistent with cytokine production, γδ T cells from 1
week-old calves were found to have enhanced proliferation compared to 5-6 month-old
steers upon stimulation with pokeweed mitogen. This was in contrast to CD4+ and CD8+
T cells, which had enhanced proliferation in the steer population. By 8 weeks of age, γδ T
cells from the calves had similar proliferation compared to the steers171. In summary, γδ
T cells from cattle appear to have enhanced proliferation and cytokine production in
younger individuals. However, these studies are few in number and may not reflect the
behavior of bovine γδ T cells to all stimulants. In addition, the effects of age on other
effector functions of γδ T cells, such as cytotoxicity and non-IFNγ cytokine production,
have not yet been examined and will likely be the focus of future studies.
Plant-derived Agonist Activity on Innate Lymphocytes
Immunotherapy Targeting Innate Lymphocytes
Because of their lytic activity, NK cells and γδ T cells have been targeted for
immunotherapy in cancer patients. During these therapies, cytokines and other agonists
are used to expand activated NK cell and γδ T cell numbers. Unfortunately, while these
types of therapies have had some success, these treatments have had limited efficacy in
cancer patients overall172, 173.
38
There are several potential explanations for the problems current immunotherapy
strategies to stimulate and expand human NK cells and γδ T cells have had in the clinic.
Use of cytokines has been problematic because of toxicity with high doses.
Phophoantigen-based therapies for γδ T cell immunotherapy only target the Vδ2 subset of
γδ T cells, ignoring the potential benefit of the Vδ1 subset, and there is an exhaustion
phenomenon that occurs with using phosphoantigens repeatedly172, 173. Therefore,
additional agonists for innate lymphocytes are needed which can enhance the activity of
these current therapies and reduce the amount of these stimulants needed. A number of
studies suggest that many plant extracts, some sold as nutritional supplements, could be a
good source of novel NK cell and T cell agonists.
NK Cells
Several studies suggest that polyphenol extracts can enhance NK cell activation,
proliferation, cytotoxicity, and cytokine production. The expression of CD69 on human
NK cells, as well as NK cell proliferation in response to IL-15, was enhanced in vitro by
polyphenols from unripe apple peels (APP)31. Increased cytotoxicity by murine and
human NK cells was observed upon in vitro treatment with the popular polyphenol
supplement resveratrol, as well as by the gall aqueous extract from Limoniastrum
guyonianum Boiss174, 175. In addition to its in vitro activity, an increase in NK cell
cytotoxicity was observed in resveratrol-fed mice176. A likely mechanism for the
increased cytotoxicity by resveratrol-treated NK cells was the enhanced expression of
39
perforin and NKG2D, which was induced on a human NK cell line177. To support these
observations made in vitro and in mice, clinical studies have found that the consumption
of polyphenol-rich fruit juices or foods may enhance NK cell activity. A recent example
showed that the consumption of a polyphenol-rich cranberry beverage may have
enhanced mitogen-induced proliferation by NK cells27. Another clinical study found
increased NK cell cytotoxicity in individuals given a polyphenol-rich purple sweet potato
leaves diet, similar to the results seen in resveratrol-fed mice178. In all, these data suggests
that treatment with immunomodulatory polyphenols have the potential to enhance NK
cell activity in the human population, which could protect individuals from a variety of
diseases.
αβ T Cells
Many of the studies which have examined the effect of polyphenols on αβ T cell
activity have demonstrated a suppressive effect. In fact, suppression of αβ T cells may be
critical for the observed benefit of polyphenols in some inflammatory disease models. In
support of this idea, work by Skyberg et al. demonstrated that the ability for oral APP
treatments to suppress inflammation in a dextran sodium sulfate model of colitis was lost
in αβ T cell-deficient mice, suggesting that the suppression of αβ T cell activity was the
primary benefit of APP treatment in this model19.
Still, in addition to these studies demonstating suppression of αβ T cells by
polyphenols, other studies suggest that certain polyphenols, at least under certain
40
circumstances, activate αβ T cells. For example, in contrast to the previously described
colitis study, APP, as it did with NK cells, enhanced resting human αβ T cell proliferation
in response to IL-1531. In addition to APP, lignin-like substances, which are high
molecular weight polymerized polyphenols, induced T cell proliferation and the
production of IFNγ, GM-CSF, and TNFα by murine CD4+ T cells in murine splenocytes,
with these T cells being the major source of IFNγ and GM-CSF in polyphenol-treated
splenocytes179, 180. When the mechanism for this activation of T cells by polymerized
polyphenols was examined, it was found that antigen presentating cells were required for
optimal cytokine production by T cells. Furthermore, blocking adhesion molecules
involved in T cell-APC interactions inhibited the cytokine production by T cells,
suggesting that cell-to-cell contact with APCs was required for the ability of polymerized
ligin-like polyphenols to induce cytokine production by CD4+ T cells180. Other
polyphenols which can enhance αβ T cell activity are green tea polyphenols, resveratrol,
and Cassia auriculata (CA)-derived polyphenols175, 181, 182. Similar to its effect on NK
cells, in vitro treatment of murine T cells with resveratrol promoted the production of
several cytokines, including IFNγ, by CD8+ T cells, as well as increased CTL
cytotoxicity175. Green tea polyphenols also enhanced CD8+ T cell activity, strengthening
their antitumor functions181. Finally, CA-derived polyphenols fed daily to aged rats (2426 months old) for 28 days increased T cell numbers and enhanced splenocyte
proliferation to LPS, suggesting that treatment with polyphenols may be very useful in
41
reducing the T cell hyporesponsiveness observed in elderly individuals182. Since αβ T
cells play such an important role in immunity, the influence of dietary polyphenols on
their activity should be better understood.
γδ T Cells
The localization of γδ T cells to mucosal tissues, especially the gut, put these cells in
ideal position to respond not only to pathogens, but to compounds taken orally or through
other mucosal routes. Therefore, it is no surprise that numerous studies have examined
the ability of numerous food products and nutritional supplements to influence the
activity of these cells. In one study, APP suppressed the development of oral sensitization
in mice and this suppression correlated with the expansion of γδ T cells in the IELs,
suggesting that polyphenol-induced expansion of γδ T cells may prevent the development
of food allergies16. In contrast to the immunoregulatory effects of APP-treated γδ T cells
proposed in this study, the colitis study by Skyberg et al. suggested that oral treatment of
APP may induce a proinflammatory response by γδ T cells. In that study, APP-treated αβ
T cell-deficient mice, where there was no observed immunosuppression induced by APP,
actually had increased cytokine production, including IFNγ, by mesenteric lymph node
lymphocytes compared to untreated αβ T cell-deficient mice. It is likely that γδ T cells
were a source of these cytokines, as their proportion amongst MLN lymphocytes in αβ T
cell-deficient mice increased fourfold19. In vitro studies in human PBMC cultures have
also found that APP upregulates CD69 and CD11b on human γδ T cells, promotes their
42
production of cytokines, and primes these cells to proliferate in response to suboptimal
concentrations of phosphoantigens31, 32, 183. Work by Daughenbaugh et al. showed that the
ability of APP to enhance cytokine production by γδ T cells came from its ability to
stabilize mRNA transcripts, allowing for rapid increased translation and generation of
protein32. It is possible that the stabilization of mRNA transcripts may be an important,
conserved feature of polyphenols, allowing them to enhance the production of a wide
variety of cytokines. In addition to APP, other polyphenol extracts activate γδ T cells,
including polyphenols from cocoa, tea, Cat’s claw bark, among others15, 17, 31.
Clinical studies in humans support some of the observations made in mice and
human cell cultures. In one study, a juice concentrate made up of several fruits and
vegetables increased γδ T cell numbers in human blood184. Consumption of a polyphenolrich tea beverage in another study primed γδ T cells to proliferate more robustly and
produce more IFNγ to ex vivo ethylamine challenge185. Finally, daily consumption of
cranberry polyphenols for 10 weeks enhanced γδ T cell proliferation, and possibly IFNγ
production, in response to ex vivo stimulation by PHA27. In all, these data suggests that
immunomodulatory food products containing polyphenols have the potential to enhance
γδ T cell activity in the human population.
Oenothein B
Oenothein B is a polyphenol extracted from the Epilobium angustifolium
(fireweed) plant. It is a cyclic ellagitannin with antitumor and antioxidant activity and
43
several studies over the past few decades have shown that it can also impact innate
immune cell activity. One of the more attractive features of this polyphenol is that
oenothein B is a pure compound with a known structure, unlike most polyphenol extracts,
which could allow for further therapeutic development. In the 1990's, this compound was
identified to inhibit tumor growth in vivo28, 29. This inhibition of tumor growth was not
believed to be caused by direct cytotoxicity to the tumor cells. Instead, oenothein B was
found to stimulate macrophages and the production of IL-1 by oenothein B-stimulated
macrophages was proposed to contribute to its observed antitumor effects in murine
models28, 29. However, this was not examined in detail. In 2009, work by Schepetkin et al.
supported these early studies and found that oenothein B activated both mouse and
human neutrophils and monocytes/macrophages. Among the activities enhanced by
oenothein B were intracellular calcium flux, the production of ROS, the activation of NFkB, and the production of pro-inflammatory cytokines, including TNFα and IL-633.
However, in contrast, subsequent studies have demonstrated an anti-inflammatory effect
of oenothein B. A 2011 study by Kiss et al. found that myeloperoxidase release and
production of ROS by activated neutrophils was inhibited by oenothein B186. A 2012
study demonstrated that oenothein B inhibited NO production, NF-κB activity, and the
production of IL-1β, IL-6, and TNFα by a murine macrophage cell line pretreated with
TLR2 and 4 agonists187. A third study found that oenothein B inhibited IL-1β and IL-6
production by activated DCs and a 2013 study showed that oenothein B inhibited
44
neuroinflammation in response to systemic LPS treatment25, 188. Altogether, these data
show that oenothein B has a complex influence on innate immune cells of the myeloid
and granulocyte lineages. One possible explanation for the discrepancies in these results
is the activation state of the cells at the time of treatment with oenothein B. For much of
the work by Schepetkin et al., unstimulated, resting cells were used33. However, in the
studies demonstrating suppression by oenothein B, the authors used activated cells186-188.
Therefore, oenothein B may have differential effects on activated and resting immune
cells. Surprisingly, despite all of these studies on the effect of oenothein B on neutrophils
and macrophages, the effect of this compound on lymphocytes had not been previously
studied. Therefore, we began to analyze the effect of oenothein B on lymphocytes.
Several plant-derived products which have been shown to inhibit tumor growth
are also known to stimulate innate lymphocytes, such as NK cells and γδ T cells176, 181, 185.
Therefore, we hypothesized that stimulation of innate lymphocytes could play an
important role in the antitumor properties of these types of compounds. When we tested
this hypothesis using oenothein B in a B16 melanoma tumor model, we saw a reduction
in the mean tumor weight in wild-type mice treated with oenothein B. However, in mice
that were γδ T cell-deficient, oenothein B appeared to enhance tumor growth (Figure
2.2). These data suggested that γδ T cells may have an important role in the antitumor
activity of oenothein B and that this compound can induce both anti- and protumor
effects.
45
In an alternative in vivo model, we tested the effect of oenothein B pre-treatment
in a model of influenza A infection. In a preliminary experiment, we found that
pretreatment with oenothein B intranasally reduced morbidity during influenza infection,
measured by weight loss, in a dose-dependent manner (Figure 2.3). In both of these
models, we hypothesized that oenothein B enhanced innate lymphocyte activity, which
accounted for the benefit of oenothein B treatment. Unfortunately, our animal systems
have not demonstrated the level of consistency required to fully test this hypothesis. This
prompted us to focus on in vitro analysis to identify factors which may lead to
inconsistencies in the immune-enhancing properties of oenothein B.
Figure 2.2 Inhibition of B16 tumor growth by oenothein B in a γδ T cell-dependent
manner. 10-14 week old C57BL/6 mice and γδ T cell-deficient mice were injected
subcutaneously with 5X105 B16 melanoma cells on day 0. 50μg oenothein B or saline
only was injected i.p. on days -1, 3, 7, and 11. On day 16, mice were humanely
euthanized and tumor weights were recorded. N = 4 for all groups.
46
Factors Influencing Polyphenol
Stimulation of Lymphocytes
Recent studies have already begun to examine what factors may influence the
ability of certain polyphenols to stimulate immune cells. One factor that appears to be
important is dose. In a 2001 study, which examined the effect of resveratrol on NK cell
and T cell effector responses, low doses (less than 2.5 ug/ml) of resveratrol enhanced NK
cell and T cell cytotoxicity, as well as the production of IFNγ, IL-2, and IL-4 by T cells.
However, higher doses (greater than 5 ug/ml) suppressed these responses175. This is
consistent with the study by Skyberg et al., as higher doses of APP was more effective at
suppressing colitis than lower doses19. Therefore, low amounts of polyphenols may favor
immunostimulation, while large amounts, especially over a period of time, favor
immunosuppression.
Another important factor in the immunomodulatory properties of polyphenols is
molecular size, as T cells from murine splenocytes produced IFNγ and GM-CSF when
stimulated by polymerized polyphenols with large molecular weights, but not their
corresponding monomers179. Furthermore, procyanidin oligomers, but not monomers,
stimulate γδ T cells15. In addition, the complete oenothein B molecule was necessary to
stimulate monocytes, while molecular subunits of this molecule failed to do so33.
Therefore, molecular stability and degradation in vivo may impact the
immunostimulatory properties of certain polyphenols. Importantly, while increasing
47
molecular size appears to have a significant effect on immunostimulation by polyphenols,
small molecular weight polyphenols still have potent antioxidant activity189.
Figure 2.3 Oenothein B reduced morbidity during influenza infection. Saline only or
indicated concentrations of oenothein B were administered intranasally. 24hrs later, mice
were infected intranasally with 1600 pfu of influenza strain PR8. Mice were weighed
daily until day 14. (A) Experimental schematic. (B) Daily mouse weights as a % of
starting body weight. N=5 for all groups.
In addition to molecular size, more subtle changes in polyphenol structure can
have profound impacts on their activity. One study found that methylated, but not
unmethylated, quercetin could enhance IL-1β production by monocytes in response to a
48
TLR2 agonist. In addition, not all methylated quercetin molecules had this activity, only
certain ones190. Since polyphenols can be methylated in vivo, this could be an explanation
as to why quercetin showed NK cell-enhancing activity in vivo, but not in vitro191. These
data suggest that small changes in polyphenol chemistry can have profound effects on
their immunostimulatory activity.
While several polyphenol-specific factors have been identified which influence
their ability to stimulate immune cells, very little research has been done to examine
immunological factors which could impact their activity. However, previously described
studies with oenothein B suggest that immune cells’ level of activation may be an
important variable, as oenothein B appeared to activate resting cells while suppressing
highly activated cells33, 186-188. Consistent with this, APP enhanced cytokine production in
αβ T cell-deficient mice with colitis. In these mice, colitis severity and proinflammatory
cytokine production in untreated mice was low compared to untreated wild-type mice,
suggesting that lymphocytes in these mice were not as activated, perhaps making them
more susceptible to activation by APP. However, in wild-type mice, where colitis was
more severe and lymphocytes produced more inflammatory cytokines, suggesting a
higher state of activation, treatment with APP was suppressive and inhibited cytokine
production19. Therefore, a common behavior of immunomodulatory polyphenols may be
the stimulation of resting immune cells, but the suppression of activated cells.
49
In addition to the activation state of immune cells, it was hypothesized that aging
could also influence immune responses to polyphenols, as it influences responses to
many other immune stimulants63, 65, 89 155, 156, 170, 171. For the rest of this dissertation, it is
first determined that oenothein B stimulates lymphocytes, activates NK cells, αβ T cells,
and γδ T cells, and promotes the production of IFNγ by these cells. Next, it is examined if
age is a human and animal variable which influences the response of these innate
lymphocytes to oenothein B, indeed showing that T cell responses to oenothein B are
affected by age. Finally, a literature review is provided of the role of γδ T cells in cancer,
which can either be beneficial or detrimental, as this may influence the effects of γδ T
cell-stimulating polyphenols in tumor models. These studies provide additional insight
into the immunomodulatory properties of oenothein B, as well as suggest additional
variables to be taken into account when examining the immunomodulatory properties of
oenothein B and, perhaps, other polyphenols.
50
CHAPTER THREE
OENOTHEIN B, A CYCLIC DIMERIC ELLAGITANNIN ISOLATED FROM
EPILOBIUM ANGUSTIFOLIUM, ENHANCES IFNγ PRODUCTION BY
LYMPHOCYTES
Contribution of Authors and Co-Authors
Manuscript in Chapter Three
Author: Andrew G. Ramstead
Contributions: Designed and performed experiments, prepared the manuscript.
Co-author: Igor A. Schepetkin
Contributions: Isolated oenothein B and edited the manuscript.
Co-author: Mark T. Quinn
Contributions: Isolated oenothein B and edited the manuscript.
Co-author: Mark A. Jutila
Contributions: Designed experiments, obtained funding, and edited the manuscript.
51
Manuscript Information Page
Andrew G. Ramstead, Igor A. Schepetkin, Mark T. Quinn (PhD), and Mark A. Jutila
(PhD)
Journal Name: PLoS ONE
Status of Manuscript:
___ Prepared for submission to a peer-reviewed journal
___ Officially submitted to a peer-reviewed journal
___ Accepted by a peer-reviewed journal
_X_ Published in a peer-reviewed journal
Published by: PLOS
Reference: PLoS ONE 7(11): e50546. doi:10.1371/journal.pone.0050546 (2012).
Supplemental material is available online under NIH public manuscript
#PMC3136559. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3136559/?tool=pubmed"
52
Abstract
Oenothein B is a polyphenol isolated from Epilobium angustifolium and other
plant sources, which has been reported to exhibit immunomodulatory properties.
Oenothein B is known to activate myeloid cells and induce the production of IL-1 and
other cytokines. However, its effects on lymphocytes are unknown. In this report, we
show that oenothein B stimulated innate lymphocytes, including bovine and human γδ T
cells and NK cells, resulting in either increased CD25 and/or CD69 expression. We also
demonstrate that oenothein B enhanced the production of interferon-γ (IFNγ) by bovine
and human NK cells alone and in combination with interleukin-18 (IL-18), a response not
observed with other commonly studied polyphenols. Furthermore, we demonstrate that
oenothein B enhanced the production of IFNγ by human T cells. Since IFNγ contributes
to antitumor, antibacterial, and antiviral cell responses, these data suggest an additional
mechanism that could account, at least in part, for the immune enhancing properties of
oenothein B.
Introduction
Nutritional supplements have been studied over many years for their ability to
treat and prevent disease, including cancer and infections. Polyphenols represent a group
of plant compounds found in many supplements that have been studied extensively for
their role in promoting human health. Numerous studies have focused on the antioxidant
53
properties of polyphenols; however, the antioxidant effects of nutritional polyphenols in
vivo are controversial192. In addition, there are numerous studies that demonstrate
biological activity of polyphenols beyond antioxidant activity, including modulating
enzyme activity193, receptor signaling194, and immunity31, 32, 33, 183.
Innate lymphocytes, such as NK cells and γδ T cells, play an important role in
host defense against cancer and various pathogens, and enhancing the activity of these
cells is an attractive option for immunotherapy195-197. Results by our group and others
have shown that some nutritional supplements are useful sources of novel agonists for
innate lymphocytes and that the use of these supplements may represent a novel strategy
to enhance the activity of these cells 6, 16, 31, 33, 183. For example, alkylamines from tea,
apples, and wine, polysaccharides from Acai fruit and Funtumia elastica bark, and other
plant components have been shown to activate and enhance the proliferation of γδ T
cells198-201. In addition, we have recently found that certain polyphenols, such as
oligomericprocyanidins (OPCs) from apple peel, also stimulate innate lymphocytes, from
different animals, including humans31. However, not all polyphenols are capable of
stimulating innate lymphocytes, and the size and structure of these compounds are
important for their immunomodulating properties15, 179.
NK cells and γδ T cells provide an early source of several cytokines, including
interferon-γ (IFNγ) and IL-17202-204. The production of IFNγ by lymphocytes is important
in immune defense against various tumors and infections and could provide a possible
54
mechanism for the antibacterial, antiviral, and antitumor properties proposed for certain
polyphenols205-207. However, the induction of IFNγ by polyphenols is poorly understood
or defined. In our earlier study of OPCs, we found no evidence for the induction of IFNγ
in innate lymphocytes. Conversely, we have detected some IFNγ production from human
PBMCs treated with oenothein B, a unique polyphenol with different structural and
immunological properties than OPCs33. Therefore, we investigated whether oenothein B
might induce IFNγ production in innate lymphocytes or, based on our earlier studies that
showed OPCs can enhance responses to secondary signals, possibly prime innate
lymphocytes to respond more robustly to known inducers of IFNγ, such as IL-18208.
Briefly, oenothein B is a dimeric, macrocyclicellagitannin isolated from
Epilobium angustifolium, as well as other plant sources. It has been studied for antitumor,
antiviral, antibacterial, antioxidant, proinflammatory, and anti-inflammatory properties28,
33, 186, 187, 211
. Oenothein B has been reported to inhibit inflammatory responses by
phagocytes induced by TLR agonists and other stimulants186, 187. However, in the absence
of additional stimulation, oenothein B promotes inflammatory responses by phagocytes.
In studies conducted in the early 1990’s, oenothein B was shown to reduce the growth of
several tumors in vivo and activate macrophages, promoting the production of IL-128.
Induced IL-1 production was proposed to be important in the antitumor properties of
oenothein B, although this has not been directly tested. We recently showed that
oenothein B induces the production of IL-1, as well as other proinflammatory cytokines,
55
including IL-6 and tumor necrosis factor α (TNFα), by monocytes33, responses not seen
with OPCs. In addition, we showed that substructures of oenothein B did not stimulate
phagocytes to the same extent as oenothein B33, suggesting an important role for the
complete structure in its immunological activity. To date, there are no reports on the
effects of oenothein B on lymphocytes. We now show that oenothein B stimulates innate
lymphocytes (γδ T cells and NK cells) and promotes their production of IFNγ. We also
describe a novel priming effect of oenothein B on NK cells, leading to enhanced IFNγ
production following IL-18 treatment. Finally, we describe a similar priming effect of
oenothein B in response to a tumor cell line.
Materials and Methods
Ethics Statement
All animal experiments were performed in accordance with National Institutes of
Health guidelines and approved by the Institutional Animal Care and Use Committee of
Montana State University (protocol identification: 2009-3, 2011-61). Human subjects
testing was performed in accordance with a protocol approved by the Institutional
Review Board of Montana State University (approval identification: MJ032609), and
written, informed consent was obtained from all individuals. No specific permits were
required for the described field studies involving E. angustifolium. According to the
Gallatin National Forest Office (Montana), collection of limited amounts of plant
56
materials for non-commercial, educational purposes does not require a permit. All plants
were collected from a National Forest and public land and no endangered or protected
species were collected.
Isolation of Oenothein B Oenothein B was isolated and identified as described
previously33. Briefly, fully blossomed E. angustifolium were collected and the dried plant
material (400g) was extracted with 80% methanol at room temperature for 3 days. The
combined extracts were concentrated, and any precipitates were removed by filtration
through a 0.22-μm filter. The filtrate was lyophilized to obtain the crude extract or
subjected to concentration and fractionation on a Sephadex LH-20 column (2.8 × 33 cm)
using 80% methanol as an eluent. The relevant fractions were pooled and evaporated to
dryness, re-chromatographed twice, and compound identification was performed by
NMR and mass spectrometry, as described33. Purity was determined to be >95% by
HPLC and mass spectrometry, as described33. A Limulus amebocyte lysate assay kit
(Cambrex, East Rutherford, NJ) was used to evaluate possible endotoxin contamination
in purified oenothein B. Purified oenothein B found to be free of endotoxin was stored at
-80°C until used in the functional assays described below.
Human and Bovine Peripheral
Blood Mononuclear Cell Preparations Whole blood was collected from 1- to 3month bull Holstein calves into sodium heparin tubes (BD Biosciences, San Jose, CA)
and from healthy human adult donors with ACD solution B anticoagulant tubes (BD
57
Biosciences). Mononuclear cells were separated from whole blood using Histopaque
1077 (Sigma-Aldrich, St. Louis, MO) or Ficoll-Paque™Premium (GE Healthcare,
Piscataway, NJ) for bovine and human cells, respectively, as previously described and
per the manufacturer’s instructions31. Additionally, bovine red blood cells were removed
by hypotonic lysis after Histopaque separation.
Flow Cytometric Analysis and Cell
Sorting of Bovine and/or Human PBMCs PBMCs were suspended in X-VIVO 15
serum-free medium or RPMI 1640 medium containing supplements and 10% FBS
(cRPMI (31)). Cells were then cultured with or without oenothein B at 37°C and 10%
CO2. Bovine cells were stained with antibodies against IL-2Rα (LCTB2A, VMRD),
CD335 (AKS1, AbDSerotec), γδ TCR (GD3.8212), or a bovine monocyte antigen
(BN180). Human cells were stained with antibodies against CD69 (FN50, Biolegend),
CD25 (M-A251, BD Pharmingen), CD3 (UCHT1, Biolegend), CD56 (MEM-188,
Biolegend and CM55B, eBioscience), CD8 (HIT8a, Biolegend), γδ TCR (11F2, BD
Biosciences), Vδ2 (B6, BD Pharmingen), or IL-18Rα (H44, Biolegend). All antibodies
were directly labeled or indirectly labeled using goat anti-mouse FITC, PE, or APC
(Jackson ImmunoResearch Laboratories, West Grove, PA). After staining, cells were
analyzed using a BD Biosciences FACSCalibur with high throughput sampling (HTS).
Removal of bovine CD335+ cells, γδ T cells, and monocytes from PBMC
preparations was performed using flow cytometric sorting. Briefly, bovine CD335+ cells,
58
γδ T cells, and monocytes were stained with monoclonal antibodies (mAb) against
CD335 (AbDSerotec), γδTCR (GD3.8), and monocytes (BN180), respectively. Negative
cells were then purified using a BD Biosciences FACSAria cell sorter to achieve >95%
purity. As a control, unsorted bovine PBMCs were collected from the FACSAria for
undepleted controls (controlling for the effects of the sorting procedure). Human NK
cells were also sorted. Briefly, NK cells were isolated by staining cell preparations with
CD3 (UCHT1, Biolegend) and CD56 (CM55B, eBioscience) and sorting CD3-/CD56+
cells using the FACSAria cell sorter to achieve >95% purity. After sorting, human cells
were allowed to rest overnight in cRPMI with 10% FBS at 37°C and 10% CO2 before
being used in the experiments described below.
IL-18 Activation Assays To test for priming effects by oenothein B, bovine and
human PBMCs were isolated and incubated in X-VIVO 15 medium at 37°C and 10%
CO2 in the presence of oenothein B (0-40 µg/ml) or medium only for approximately 24
hrs (bovine cells) or 48 hrs (human cells). Cells were then washed with Dulbecco’s PBS
and resuspended in X-VIVO 15 medium in the presence or absence of recombinant
human (rhu) IL-18 (R&D Systems, Minneapolis, MN). A fraction of the cells were then
incubated approximately 18 hrs, and the supernatant fluids were collected for IFNγ
quantification by ELISA (see below). Other cells were treated with brefeldin A
(eBioscience), incubated for 6 hrs, stained for intracellular IFNγ using anti-IFNγ
antibodies, and analyzed by flow cytometry (see below).
59
Sorted human NK cells were resuspended in X-VIVO 15 medium and plated in a
96-well plate at 5 X 104 cells/well. Cells were treated with oenothein B (20 µg/ml), rhu
IL-18 (100 ng/ml), both, or medium only. Cells were incubated for 24 hrs and supernatant
fluids were collected for IFNγ quantification by ELISA (see below).
K562 Assay K562 (chronic myelogenous leukemia) human cell line was from
American Type Culture Collection (Manassas, Virginia). Human PBMCs were isolated
and incubated in X-VIVO 15 medium at 37°C and 10% CO2 in the presence of oenothein
B (20 µg/ml) or medium only for approximately 24 hrs. Cells were then washed with XVIVO 15 and subsequently cultured in X-VIVO 15 in the presence or absence of K562
target cells. To measure soluble IFNγ, cells were co-cultured for 42 hours at 37°C and
10% CO2. Supernatant fluids were then collected for IFNγ quantification by ELISA (see
below). To measure intracellular IFNγ, cells were co-cultured for 24 hours at 37°C and
10% CO2 with brefeldin A added for the final 6 hours. IFNγ quantification was then
performed by flow cytometry (see below).
Measurement of IFNγ Enzyme-linked immunosorbent assays (ELISA) were used
to measure IFNγ in cell supernatant fluids. A bovine IFNγ kit (MABTECH, Cincinnati,
OH) and a human IFNγ kit (Biolegend ELISA Max) were used to perform ELISAs,
according to the manufacturer’s instructions. All measurements were performed in
duplicate or triplicate.
60
To measure IFNγ production by flow cytometry, leukocytes were isolated as
described above. Cells were treated with brefeldin A and incubated for 6 hrs at 37°C and
10% CO2. Bovine and human lymphocytes were stained as described above. Cells were
then fixed with 2% paraformaldehyde for at least 10 min, washed once with PBS + 2%
horse serum, and then washed once with 0.2% saponin (Sigma) in PBS + 2% horse
serum. Bovine IFNγ was detected using a PE-conjugated mouse IgG1 mAb against
bovine IFNγ (MCA1783E, ABD Serotec Inc., Raleigh, NC), whereas human IFNγ was
detected using a PE-conjugated mouse IgG1 mAb (clone 4S.B3, Biolegend). For isotype
controls, cells were stained with a PE-conjugated mouse IgG1 antibody (Biolegend).
IFNγ antibodies and isotype controls were resuspended in 0.2% saponin. Cells were
stained for 20 min at room temperature. After staining, cells were washed, then analyzed
using a FACSCalibur with HTS.
Statistical Analysis Statistical analyses were performed using Prism 4 (GraphPad
Software, San Diego, CA). The data were analyzed by Student’s paired t-test, Student’s
unpaired t-test, One-way ANOVA, or Two-way ANOVA as indicated.
61
Results and Discussion
Oenothein B Activates
Human and Bovine Lymphocytes
Previously, we and others have found bovine PBMCs to be a useful model for the
testing of novel innate lymphocyte agonists31, 213. The bovine model has also been used to
study infections by Mycobacterium species and Salmonella species since it better reflects
human diseases than rodent models214-216. To determine if oenothein B stimulated
lymphocytes, we first evaluated IL-2Rα expression as a marker for activation of bovine
PBMCs. IL-2Rα was upregulated on both bovine γδ T cells and NK cells after
stimulation with oenothein B (20-40 µg/ml) for 24 hours in vitro (Figure 3.1A and Figure
3.2). Doses and timepoints were based upon preliminary dose and kinetic analyses (data
not shown). We then examined if similar responses were seen in human PBMCs, using
CD69 expression as a marker for activation. In these studies, oenothein B stimulation for
2 days in vitro induced CD69 expression on human CD3+ T cells, γδ T cells, CD8+ T
cells, and CD3- CD56+ NK cells (Figure 3.1B and Figure 3.2) at similar doses known to
stimulate monocytes33. Within the human γδ T cell population, both Vδ2+ (major
circulatory subset) and Vδ2- (mainly Vδ1+ cells (217)) subsets were activated by
oenothein B (Figure 3.1B), which is similar to responses induced by OPCs31. In addition,
we also examined CD25 expression on human PBMCs. Interestingly, oenothein B
stimulation induced CD25 expression on T cells, but not NK cells (Figure 3.3).
62
Figure 3.1 Oenothein B induces IL-2Rα or CD69 on bovine and human lymphocyte
subsets. (A) Bovine PBMCs (105 cells/well) were treated with the indicated
concentrations of oenothein B in X-VIVO medium for 24 hrs, and IL-2Rα expression on
γδ T cells and NK cells was measured by multi-color flow cytometry. NK cells were
defined as non-γδ T cells that expressed CD335. The graphs represent pooled data from 3
individuals. Each treatment was analyzed in triplicate and error bars indicate SEM.
Significance compared to untreated cells (0 µg/ml) was determined by One-way ANOVA
with Bonferroni post-test. *p<0.05, **p<0.01, ***p<0.001 (B) Human PBMCs (105
cells/well) were treated with the indicated concentrations of oenothein B in cRPMI
medium for 48 hrs. CD69 expression on lymphocytes, which included CD3+ T cells,
CD8+ T cells, γδ T cells, and NK cells, was then measured by flow cytometry. The
graphs represent pooled data from 5 individuals. Each treatment was analyzed in
triplicate and error bars indicate SEM. Significance compared to untreated cells (0 µg/ml)
was determined by One-way ANOVA with Bonferroni post-test. *p<0.05, **p<0.01,
***p<0.001
63
Figure 3.2 Oenothein B induces IL-2Rα or CD69 on bovine and human NK cells. (A)
Bovine PBMCs (105 cells/well) were treated with 20 μg/ml oenothein B in X-VIVO
medium for 24 hrs, and IL-2Rα expression on NK cells was measured by multi-color
flow cytometry. Representative examples of two-color flow cytometry plots comparing
IL-2Rα staining on oenothein B-treated and untreated bovine NK cells (gated on CD335+
cells) from each animal are shown. (B) Human PBMCs (105 cells/well) were treated with
40 μg/ml oenothein B in cRPMI medium for 48 hrs. CD69 expression on NK cells (gated
on CD3- CD56+ cells) was then measured by flow cytometry. Representative examples
of two-color flow cytometry plots comparing CD69 staining on oenothein B-treated and
untreated human NK cells from each donor are shown.
64
Figure 3.3 Oenothein B induces CD25 on human T cells. Human PBMCs (105 cells/well)
were treated with the indicated concentrations of oenothein B in X-VIVO medium for 42
hrs. CD25 expression on lymphocytes, which included γδ T cells (CD3+/γδ TCR+), NK
cells (CD3-/CD56+), and αβ T cells (CD3+/γδ TCR-), was then measured by flow
cytometry. The graph represents pooled data from 5 individuals. Each treatment was
analyzed in duplicate and error bars indicate SEM. Significance compared to untreated
cells (0 µg/ml) was determined by One-way ANOVA with Bonferroni post-test. *p<0.05,
**p<0.01, ***p<0.001
Oenothein B Primes Bovine
PBMCs to Respond to IL-18
To examine the effects of oenothein B on IFNγ production in the bovine model,
bovine PBMCs were treated with oenothein B for two days and secreted IFNγ was
measured by ELISA. Similar to our studies on OPCs, we did not find significant amounts
of IFNγ produced by oenothein B-treated bovine PBMCs (data not shown). However, in
our original studies with OPCs, we found that OPC-treated γδ T cells had enhanced
responses to secondary signals, such as IL-2 and TCR agonists31. In addition, others have
65
found that feeding bovine calves polyphenols from pomegranate can enhance mitogeninduced IFNγ production by PBMCs30. Therefore, we hypothesized that oenothein B
might enhance or prime responses to an inducer of IFNγ.
As such, we tested if oenothein B treatment of bovine lymphocytes enhanced
responses to the IFNγ-inducing cytokine, IL-18. We also tested several well-studied
polyphenols, epigallocatechingallate (EGCG), resveratrol, curcumin, and theaflavin
digallate (TFDG), all of which are potent antioxidents, to determine if such a response
was a common property of polyphenols. When oenothein B–treated cells were
subsequently treated with suboptimal doses of IL-18, IFNγ production was greatly
enhanced compared to IL-18 or oenothein B alone (Figure 3.4). These data suggested that
oenothein B could prime immune cells for enhanced IFNγ production in response to low
doses of IL-18. Resveratrol and curcumin did not enhance IFNγ production in response to
IL-18, but rather appeared to suppress the response, which would be consistent with
previous studies describing their immunosuppressive properties218, 219. Both EGCG and
TFDG enhanced IFNγ production in response to IL-18 in one of the calves tested, but
their effect was not as consistent or as robust as oenothein B. The level of priming by
oenothein B and the amount of IFNγ produced varied between animals. It is likely that
these observed differences between the three calves were due to animal-specific
responses to oenothein B, as our preliminary studies with IL-2Rα suggested that PBMCs
66
from individual calves can respond differently to oenothein B. Based on these results, we
focused our subsequent studies on oenothein B and its effect on IFNγ production.
Presence of CD335+ Cells is Essential
for Oenothein B Priming to IL-18
After observing enhanced IFNγ production by bovine cells pre-treated with
oenothein B, we then determined which cells were important for this response. Since
oenothein B has been shown to be a potent monocyte agonist, we first examined if these
cells were essential for the priming responses. Monocytes were removed by flow
cytometric sorting, and the priming response was again evaluated. Priming responses
were still observed in monocyte-depleted PBMCs, although the level of priming was
reduced in two out of three experiments (Figure 3.5A). These results suggested that
monocytes likely contributed to the response in the mixed population, but were not
required for the response.
We then examined the importance of NKp46+ cells, since they are a major source
of IFNγ induced by IL-12 and IL-18 in bovine lymphocytes75. NKp46, also known as
CD335, is a NK cell marker, although it is expressed by other minor leukocytepopulations, including some γδ T cells75. To test the importance of these cells, we
depleted cells expressing CD335 from bovine PBMCs and found that nearly all of the
oenothein B-induced IFNγ priming response was absent compared to undepleted PBMCs
(Figure 3.6A).
67
Figure 3.4 Oenothein B primes bovine PBMCs to respond to IL-18. Bovine PBMCs
(105 cells/well) were treated with oenothein B (40 µg/ml and 20 µg/ml), EGCG (40
µg/ml and 20 µg/ml), resveratrol (50 µg/ml and 25 µg/ml), curcumin (40 µg/ml and
20 µg/ml), theaflavin digallate (50 µg/ml), or X-VIVO medium alone for
approximately 24 hrs. Cells were then washed and treated with 10 ng/ml rhu IL-18,
100 ng/ml rhu IL-18, or X- VIVO medium alone for approximately 24 hrs. After
incubation, soluble IFNγ levels were measured by ELISA. The data are expressed
as mean +/- SEM of three independent experiments. Statistical significance was
measured by Two-way ANOVA with Bonferroni post-test. ap<0.05 compared to
10ng/ml IL-18 only, bp<0.05 compared to 100ng/ml IL-18 only
68
Because CD335 is expressed on some γδ T cells75, we examined whether γδ T
cells contributed to the oenothein B-induced IFNγ response. Removal of γδ T cells
reduced, but did not eliminate, the priming response (Figure 3.5B). This result suggested
that, like monocytes, γδ T cells contributed to, but were not required for, the response and
further suggested that γδ TCR-/CD335+ cells were the primary source of IFNγ in these
assays. As a final approach to confirm these results, multi-color intracellular cytokine
analyses were performed. As shown in Figure 3.6, oenothein B-primed, IL-18-treated
CD335+ cells expressed IFNγ (Figure 3.6B and 3.6C). The percentage of CD335+ cells
was also enhanced by oenothein B (Figure 3.6C). However, this was likely due to
activated monocytes adhering to the sample plates and being removed from the CD335population rather than an expansion of CD335+ cells. Collectively, these data indicate
that CD335+ NK cells are the major source of IFNγ produced in response to oenothein B
and IL-18 in the bovine system.
69
Figure 3.5 Effect of monocyte and γδ T cell depletion on oenothein B-priming of bovine
PBMCs. Bovine PBMCs (105 cells/well) were depleted of (A) monocytes or (B) γδ T
cells and treated with 20 µg/ml oenothein B or X-VIVO medium alone for 24 hrs. Cells
were then washed and treated with 10 ng/ml rhu IL-18 or medium alone for 18 hrs. After
incubation, IFNγ levels in the supernatant fluids were measured by ELISA. The data are
expressed as mean +/- SEM of three independent experiments comparing depleted
PBMCs to un-depleted controls tested concurrently. All samples were tested in duplicate
or triplicate. Statistical significance was measured by Two-way ANOVA with Bonferroni
post-test. *p<0.05, **p<0.01, ***p<0.001
70
Figure 3.6 Oenothein B primes bovine CD335+ cells to respond to IL-18. (A)
Bovine PBMCs (105 cells/well) were depleted of CD335+ cells and treated with 20
µg/ml oenothein B or X-VIVO medium alone for 24 hrs. Cells were then washed
and treated with 10 ng/ml rhu IL-18 or medium alone for 18 hrs. After incubation,
IFNγ levels in the supernatant fluids were measured by ELISA. The data are
expressed as mean +/- SEM of three independent experiments comparing depleted
PBMCs to un-depleted controls tested concurrently. All samples were tested in
duplicate or triplicate. Statistical significance was measured by Two-way ANOVA
with Bonferroni post-test. *p<0.05, **p<0.01, ***p<0.001 (B) Bovine PBMCs
(105 cells/well) from a new calf were treated with the indicated amounts of
oenothein B or X-VIVO medium alone for 24 hrs. Cells were washed and treated
with 10 ng/ml rhu IL-18 or X-VIVO medium alone for 6 hrs in the presence of
brefeldin A. IFNγ production was measured by intracellular flow cytometry. The
data are expressed as mean+/- SEM. All samples were tested in triplicate.
Statistical significance was measured by Two-way ANOVA with Bonferroni posttest. *p<0.05, **p<0.01, ***p<0.001 (C) Representative examples of two-color
flow cytometry plots comparing IFNγ staining on CD335+ cells.
71
Oenothein B Induces IFNγ Production
by Human Innate Lymphocytes
In our previous studies with oenothein B, we showed that treatment of human
PBMCs with oenothein B promoted some IFNγ production, in contrast to the response we
observed with bovine cells33. However, potential sources of this cytokine were not
identified. We first confirmed our previous results in human PBMCs by cytokine ELISA
(Figure 3.7A). We then examined if lymphocytes were a source of this induced IFNγ.
Human T cells, including γδ T cells and CD8+ T cells, produced IFNγ in response to
oenothein B (Figure 3.7B and 3.7C). The percentages of NK cells positive for IFNγ
staining also increased in three out of five donors tested (Figure 3.7B), but minimal
staining was observed compared to that seen in T cells (Figure 3.7C). Thus, unlike the
bovine system, oenothein B-induced IFNγ production was not restricted to the human NK
cell population.
In the bovine system, NK cells were primed to produce enhanced IFNγ in
response to IL-18. We tested whether the same could be true for human NK cells. First,
multi-color flow cytometry showed that IL-18 receptor was increased on oenothein Btreated human NK cells in two out of five donors (Figure 3.8A). Similarly, further
analyses showed that in two, possibly three, of five human PBMC preparations, IFNγ
production was increased in oenothein B-primed, IL-18-treated human NK cells
compared to cells treated with oenothein B or IL-18 alone (Figure 3.8B and 3.8C).
72
We then tested if oenothein B could enhance IFNγ production in response to the
NK cell target leukemic cell line, K562. Others have shown that stimulation by K562
cells induces IFNγ secretion by NK cells and that this response can be enhanced by the
presence of a second stimuli220. Consistent with these reports, pretreatment of human
PBMCs with oenothein B enhanced IFNγ production in response to K562 cells compared
to untreated PBMCs (Figure 3.9A) andNK cells were the major cell population primed by
oenothein B for enhanced IFNγ production (Figure 3.9B).
NK cell numbers and activity can vary significantly from donor to donor. To
address whether the inconsistency seen in PBMC preparations in response to oenothein B
and IL-18 might be due to variable numbers of NK cells between PBMC samples or
variable influences by other cells within the mixed populations on the NK cells, human
NK cells were sorted, and then equal cell numbers were treated with oenothein B alone,
IL-18 alone, or a combination of both. IFNγ production was measured 24 hrs later by
ELISA. As shown in Figure 3.10, oenothein B alone directly induced IFNγ production by
NK cells and there was an increase in IFNγ production with the combined treatment in all
donors tested, although the amount of IFNγ produced varied between donors. This
variability in IFNγ production by NK cells has been observed in other studies and may
have a genetic component220, 221.
73
Figure 3.7 IFNγ production by human lymphocytes in response to oenothein B. (A)
Human PBMCs (105 cells/well) were treated with the indicated concentrations of
oenothein B or X-VIVO medium alone for 48 hrs, and soluble IFNγ levels in supernatant
fluids were measured by ELISA. The graph represents data from ten individuals, with
each sample plated in triplicate. Statistical significance was measured by One-way
ANOVA with Bonferroni post-test. *p<0.05, **p<0.01, ***p<0.001 (B) Human PBMCs
(105 cells/well) were treated with oenothein B or X-VIVO medium alone for 6 hrs in the
presence of brefeldin A. The percent of total CD3+ T cells, γδ T cells, CD8+ T cells, and
NK cells positive for IFNγ staining was then determined by flow cytometry. The graphs
represent data for five individuals, with each treatment analyzed in triplicate. Statistical
significance was determined by paired Student’s t-test. *p < 0.05, **p<0.01, ***p<0.001
(C) Representative examples of two-color flow cytometry plots comparing IFNγ staining
on oenothein B-treated and untreated human lymphocytes.
74
Figure 3.8 Priming of human NK cells to IL-18 by oenothein B. (A) Human PBMCs (105
cells/well) were treated with the indicated concentrations of oenothein B or X-VIVO
medium alone for 48 hrs, and expression of IL-18R on human NK cells was determined
by flow cytometry. Significance was determined by One-way ANOVA with Bonferroni
post-test. *p < 0.05, **p<0.01 (B) Human PBMCs (105 cells/well) were treated with 20
µg/ml oenothein B or X-VIVO medium alone for 48 hrs. Cells were washed with PBS
and treated with 50 ng/ml rhu IL-18 for 6 hrs in the presence of brefeldin A. IFNγ
staining [mean fluorescent intensity (MFI)] on human NK cells was determined by flow
cytometry. The graphs represent data from five individuals, with each treatment analyzed
in triplicate. (C) Representative examples of two-color flow cytometry plots comparing
IFNγ staining on gated human NK cells treated with 20 µg/ml oenothein B, 50 ng/ml
rhuIL-18, both, or X-VIVO medium alone. *p<0.05, **p<0.01, ***p<0.001
75
Figure 3.9 Priming of human NK cells to K562 cells by oenothein B. (A) Human PBMCs
(105 cells/well) were treated with 20 µg/ml oenothein B or X-VIVO medium alone for
approximately 24 hrs. Cells were then washed and co-cultured with or without K562 cells
at effector:target (E:T) ratios of 10:1 and 1:1 for approximately 42 hrs. After incubation,
soluble IFNγ levels were measured by ELISA. The data represent pooled results from
three donors and are expressed as mean +/- SEM. Samples were analyzed in duplicate.
Statistical significance was measured by Two-way ANOVA with Bonferroni post-test.
(B) Human PBMCs (105 cells/well) were treated with 20 µg/ml oenothein B or X-VIVO
medium alone for approximately 24 hrs. Cells were then washed and co-cultured with or
without K562 cells at an effector:target (E:T) ratio of 1:1 for approximately 18 hrs. After
incubation, brefeldin A was added to the culture for 6 hrs. IFNγ expression by NK cells
(CD3-/CD56+), T cells (CD3+), and others (CD3-/CD56-) was then measured by
intracellular flow cytometry. The data represent pooled results from two donors and are
expressed as mean +/- SEM. Samples were analyzed in duplicate. Statistical significance
was measured by One-way ANOVA with Bonferroni post-test. *p<0.05, **p<0.01,
***p<0.001
76
Figure 3.10 Oenothein B directly primes purified human NK cells to produce IFNγ.
Human NK cells (5 X104 cells/well) were sorted and treated with 20 μg/ml oenothein B,
100 ng/ml rhu IL-18, both, or medium alone. After 24 hrs, soluble IFNγ was measured by
ELISA. The graph represents soluble IFNγ levels in culture supernatant fluids from three
separate experiments with three different donors. Error bars indicate SEM. Each sample
was analyzed in triplicate. Significance was determined by One-way ANOVA with
Bonferroni post-test. *p < 0.05, **p<0.01, ***p<0.001
These data further suggest that, as with bovine cells, oenothein B treatment has the
potential to augment IFNγ production by human NK cells alone and in response to IL-18.
In addition, these data suggest that oenothein B can directly prime these cells to respond
to IL-18.
Collectively, our results show that, in addition to monocytes, oenothein B
stimulates subsets of bovine and human lymphocytes, including NK cells, CD8+ T cells,
and both Vδ2+ and Vδ2- γδ T cells, by upregulating IL-2Rα and/or CD69 on these cells.
We also show that oenothein B promotes the production of IFNγ by human lymphocytes,
specifically γδ T cells and CD8+ T cells. Furthermore, we demonstrate that IFNγ
77
production by NK cells can also be induced by oenothein B, although this response was
not as robust or consistent as that seen in T cells. Interestingly, differences in the capacity
of oenothein B to induce IFNγ production by T cells was observed between human and
bovine cells, as oenothein B alone did not directly induce significant IFNγ secretion by
bovine T cells as it did with human T cells. These data suggest that certain polyphenols
may exert species-specific effects and that immunomodulatory effects of polyphenols
demonstrated in one species may not always be conserved in other species. Thus, analysis
of the immunomodulating properties of polyphenols cannot rely solely on animal testing,
and a combination of animal and human cell testing is required to identify relevant,
conserved responses.
A possible explanation for some of the differences observed between human and
bovine T cells in these studies could be due to differences in ages, as young calves were
used for our bovine studies while adults were used for our human studies. It has been
shown that IFNγ secretion by T cells can increase with age, correlating with an increase
in CD45RO+ T cells222. Therefore, future studies could examine the effects of aging on
these responses. It is possible that lymphocyte responses to certain polyphenols in young
bovine calves are more reflective of those that might occur in children, suggesting a
potential new use for this animal model in the study of the effects of dietary polyphenols
on neonatal and adult lymphocytes.
78
A potentially important and conserved response to oenothein B is enhanced IFNγ
secretion following exposure to suboptimal IL-18 concentrations, which was observed in
both bovine and human NK cells. The synergistic effect of oenothein B and IL-18 for
enhancing IFNγ production by NK cells was observed in mixed PBMC cultures, NK celldepleted PBMCs, as well as sorted NK cells. Our earlier studies demonstrated that
oenothein B could induce IL-12 production by monocytes33, which others have found
synergizes with IL-18 to produce IFNγ223. Thus, this could provide an explanation for
oenothein B’s ability to enhance IL-18-induced IFNγ production in some of our
experiments; however, the enhanced production of IFNγ observed in sorted NK cell
cultures suggests a direct effect on NK cells by oenothein B. Additionally, oenothein B
enhanced IFNγ secretion in response to an NK cell target cell line, suggesting that the
ability of oenothein B to enhance IFNγ secretion is not restricted to IL-18, but also occurs
upon co-culture with tumor cell targets.
In conclusion, our results expand upon previous studies suggesting that oenothein
B stimulates innate and antitumor immunity, and further characterizes this activity,
suggesting that lymphocyte activation and IFNγ production may contribute to these
responses. The production of IFNγ by lymphocytes and other cells enhances antitumor
immunity by a number of mechanisms, and it will be important to examine whether
lymphocytes and/or IFNγ play an important role in the antitumor properties of oenothein
B in vivo. In addition, IFNγ production is a vital step in the host defense against
79
numerous pathogens, including viruses and intracellular bacteria. Therefore, our data also
suggest a potential mechanism whereby oenothein B could enhance antiviral and
antibacterial immunity in vivo. Thus, it will also be important to examine if oenothein B
enhances host defense against various pathogens whose clearance relies on lymphocyte
activity and IFNγ production. Further work is also necessary to identify the receptors and
signaling pathways involved in these immune stimulatory effects of oenothein B. Finally,
these studies suggest that oenothein B may be a promising candidate for therapeutic
development to supplement immunotherapies, especially those involving IL-18.
Acknowledgments
We would like to thank Dr. Robyn Klein (Department of Plant Sciences and Plant
Pathology, Montana State University, Bozeman, MT) for plant identification. We also
thank Larissa Jackiw for her assistance in FACS sorting, as well as Dr. Jodi Hedges and
Dr. Jeff Holderness for helpful scientific discussion.
80
CHAPTER FOUR
AGING INFLUENCES THE RESPONSE OF T CELLS TO STIMULATION
BY THE ELLAGITANNIN, OENOTHEIN B.
Contribution of Authors and Co-Authors
Manuscript in Chapter 4
Author: Andrew G. Ramstead
Contributions: Designed and performed experiments, prepared the manuscript.
Co-author: Igor A. Schepetkin
Contributions: Isolated oenothein B and edited the manuscript.
Co-author: Kimberley Todd
Contributions: Provided the cord blood.
Co-author: James Loeffelholz
Contributions: Provided the cord blood.
Co-author: Mark T. Quinn
Contributions: Isolated oenothein B and edited the manuscript.
Co-author: Mark A. Jutila
Contributions: Designed experiments, obtained funding, and edited the manuscript.
81
Manuscript Information Page
Andrew G. Ramstead, Igor A. Schepetkin, Kimberley Todd, James Loeffelholz, Mark
T. Quinn (PhD), and Mark A. Jutila (PhD)
Journal Name:
Status of Manuscript:
_ X_ Prepared for submission to a peer-reviewed journal
___ Officially submitted to a peer-reviewed journal
___ Accepted by a peer-reviewed journal
_ _ Published in a peer-reviewed journal
82
Abstract
Several plant extracts, including certain polyphenols, prime innate lymphocytes
and enhance responses to secondary stimuli. Oenothein B, a polyphenol isolated from
Epilobium angustifolium and other plant sources, is one these polyphenols. Oenothein B
enhances IFNγ production by both bovine and human NK cells and T cells, alone and in
response to secondary stimulation by cytokines and a tumor cell line. Innate immune cell
responsiveness is known to be affected by aging, but whether polyphenol responses by
these cells are also impacted by aging is not known. Therefore, we examined oenothein B
responsiveness in T cells from cord blood, young, and adult donors. We found that
oenothein B stimulates bovine and human T cells from individuals over a broad range of
ages, as measured by increased IL-2Rα and CD69 expression. However, clear differences
in induction of cytokines by T cells were seen. In human cord blood and bovine calves,
oenothein B was unable to induce IFNγ production by T cells. However, in human and
bovine adults, oenothein B induced IFNγ production by T cells and T cells were the most
prevalent IFNγ-expressing cell type. In addition, oenothein B induced GM-CSF
production by human adult T cells, but not cord blood T cells. Within the responsive T
cell population, we found that CD45RO+ memory T cells expressed more cytokines in
response to oenothein B than CD45RO- T cells, supporting the enhanced production of
cytokines by T cells in older individuals. In summary, our data suggest that the
83
immunostimulation of T cells by oenothein B is influenced by age, particularly with
respect to immune cytokine production.
Introduction
Plant polyphenols are known to modulate innate immunity. While many of these
compounds suppress inflammation, some can act as agonists for innate immune cells19, 25,
31, 33
. Lymphocytes, such as NK cells and T cells, which play an important role in host
defense against cancer and various pathogens, are stimulated by select polyphenols. For
example, polyphenols from tea, apples, cocoa, and others activate and enhance the
proliferation and cytokine production of γδ T cells and NK cells15, 17, 31, 32, 224.
Furthermore, these compounds are capable of antigen-independent priming of
lymphocytes, enhancing their responses to secondary stimuli, even in the absence of a
significant response to the compounds themselves31, 32, 224.
The production of IFNγ by lymphocytes is important during immune defense
against tumors and a large number of viral, bacterial, and fungal infections57, 60, 61, 134. In a
previous study, we found that oenothein B, a dimeric, macrocyclic ellagitannin isolated
from Epilobium angustifolium, enhanced IFNγ production by both NK cells and T cells
from bovine calves and humans, either alone or in combination with additional stimuli224.
During this study, the majority of IFNγ-producing cells in our bovine samples were
CD335 (NKp46)-expressing NK cells. We also found that oenothein B enhanced IFNγ
84
production by NK cells in our human samples. However, in contrast to our bovine data, a
non-NK cell population, specifically T cells (both γδ T cells and αβ T cells), produced
IFNγ in response to stimulation by oenothein B in human samples. Furthermore, this
oenothein B-responsive T cell population made up the majority of IFNγ-producing cells
in these human samples224. At this time, we did not investigate the basis for the difference
in the two systems, but felt age was likely a contributing factor. Specifically, the human
donors were all mature adults (at least 18 years old), while the bovine donors were young
animals (less than 6 months old). While numerous studies find age-related differences in
the response of immune cells to various agonists, the influence of age on the
immunostimulation activity of plant-derived polyphenols has not been well studied63, 89,
155, 170, 222
. This could have important health implications, as individuals with a broad age
range use polyphenol-containing supplements to support health and immunity and it is
currently unknown if these compounds are more effective for certain age groups.
Therefore, we examined what effect age had on the immunostimulation of T cells by
oenothein B in both humans and cattle. We show that oenothein B stimulated human and
bovine T cells from neonate, young, and adult donors, but that cytokine production by T
cells in response to oenothein B was enhanced in adults. Consistent with this, we found
that CD45RO+ T cells isolated from both cows and humans produced more IFNγ, as well
as GM-CSF, in response to oenothein B than CD45RO- cells. These data suggested that
85
the role of T cells in the modulation of immunity by oenothein B may be influenced by
age.
Materials and Methods
Ethics Statement
All animal experiments were performed in accordance with National Institutes of
Health guidelines and approved by the Institutional Animal Care and Use Committee of
Montana State University. Human subjects testing was performed in accordance with a
protocol approved by the Institutional Review Board of Montana State University, and
written, informed consent was obtained from all adult donors and cord blood donors.
Isolation of Oenothein B Oenothein B was isolated and identified as described
previously33. Briefly, fully blossomed E. angustifolium were collected and the dried plant
material (400g) was extracted with 80% methanol at room temperature for 3 days. The
combined extracts were concentrated, and any precipitates were removed by filtration
through a 0.22-μm filter. The filtrate was lyophilized to obtain the crude extract or
subjected to concentration and fractionation on a Sephadex LH-20 column (2.8 × 33 cm)
using 80% methanol as an eluent. The relevant fractions were pooled and evaporated to
dryness, re-chromatographed twice, and compound identification was performed by
NMR and mass spectrometry, as described33. Purity was determined to be >95% by
HPLC and mass spectrometry, as described33. A Limulus amebocyte lysate assay kit
86
(Cambrex, East Rutherford, NJ) was used to evaluate possible endotoxin contamination
in purified oenothein B. Purified oenothein B found to be free of endotoxin was stored at
-80°C until used in the functional assays described below.
Human and Bovine Peripheral
Blood Mononuclear Cell Preparations Whole blood was collected from 6-week
old bull Holstein calves, adult (>2 years old) Holstein cows, and adult (4-7 years old)
Angus and Angus X Hereford cows. All bovine blood was collected into sodium heparin
tubes (BD Biosciences, San Jose, CA). Whole blood from healthy human adult donors
was collected in ACD solution A anticoagulant tubes (BD Biosciences). Cord blood was
collected in sodium heparin anticoagulant tubes (BD Biosciences). Mononuclear cells
were separated from whole blood using Histopaque 1077 (Sigma-Aldrich, St. Louis, MO)
for bovine and human cells, as previously described and per the manufacturer’s
instructions31. Additionally, red blood cells were removed by hypotonic lysis after
Histopaque separation.
Cell Sorting of PBMCs Human CD45RO+ and CD45RO- T cells were isolated by
staining cell preparations with monoclonal antibodies (mAbs) against CD3 (UCHT1,
Biolegend) and CD45RO (UCHL1, eBioscience) and sorting using a FACSAria cell
sorter to achieve >98% purity. Staining with CD3 was used to distinguish CD45RO+ and
CD45RO- T cells from CD45RO+ and CD45RO- non-T cells. After sorting, human cells
87
were incubated overnight in cRPMI with 10% FBS at 37°C and 10% CO2 before being
used in the experiments described below.
Bovine T cells were isolated by staining cell preparations with mAbs against CD3
(MM1A, Washington State University and VMRD), CD4 (CC30), and γδ TCR
(GD3.8212) and sorting using a FACSAria cell sorter to achieve >98% purity. After
sorting, bovine cells were incubated overnight in cRPMI with 10% FBS at 37°C and 10%
CO2 before being used in the experiments described below.
T Cell Activation Assays To measure IL-2Rα or CD69 expression, bovine and
human PBMCs were isolated and incubated in cRPMI or X-VIVO 15 medium at 37°C
and 10% CO2 in the presence of oenothein B (0-40 µg/ml) or medium only for
approximately 24 hrs. IL-2Rα and CD69 expression was then analyzed by flow
cytometry (see below). Some samples were incubated for an additional 18 hrs in the
presence of brefeldin A (eBioscience) to quantify IFNγ and GM-CSF expression by flow
cytometry (see below). To measure IFNγ and GM-CSF secretion, bovine and human
PBMCs, as well as sorted human and bovine cells, were incubated in cRPMI or X-VIVO
15 medium at 37°C and 10% CO2 in the presence of oenothein B (0-40 µg/ml) or
medium only for approximately 24 hrs. Supernatant fluids were then collected for
quantification by ELISA (see below).
To promote differentiation into CD45RO+ T cells, bovine PBMCs were cultured
with 500 ng/ml Concanavalin A and 1 ng/ml recombinant human IL-2 in X-VIVO 15
88
medium at 37°C and 10% CO2 for 4 days. Medium was replaced after 4 days and PBMCs
were cultured 2 additional days in X-VIVO 15 medium only before stimulating with
oenothein B as described above.
Flow Cytometric Analysis of Bovine and/or Human PBMCs Bovine cells were
stained with mAbs against surface molecules IL-2Rα (LCTB2A, VMRD), CD3 (MM1A,
Washington State University and VMRD), CD45RO (GC42A1, VMRD), CD335 (AKS1,
AbDSerotec), and γδ TCR (GD3.8212). Human cells were stained with mAbs against
CD69 (FN50, Biolegend), CD3 (UCHT1, Biolegend), CD45RO (UCHL1, eBioscience),
CD56 (MEM-188, Biolegend and CM55B, eBioscience), and γδ TCR (11F2, BD
Biosciences). All mAbs were directly labeled or indirectly labeled using goat anti-mouse
FITC, PE, or APC secondary mAbs (Jackson ImmunoResearch Laboratories, West
Grove, PA) that were not cross-reactive to human or bovine antibodies. To control for
non-specific staining by secondary mAbs, cells labeled with both primary and secondary
mAbs were compared with cells labeled with secondary mAbs only. After staining, cells
were analyzed using a BD Biosciences FACSCalibur with high throughput sampling
(HTS).
Measurement of IFNγ and GM-CSF Enzyme-linked immunosorbent assays
(ELISA) were used to measure IFNγ and GM-CSF in cell supernatant fluids. A bovine
IFNγ kit (MABTECH, Cincinnati, OH), a human IFNγ kit (MABTECH, Cincinnati, OH),
89
and a human GM-CSF kit (Biolegend ELISA Max) were used to perform ELISAs,
according to the manufacturer’s instructions. All measurements were performed in
duplicate or triplicate.
To measure IFNγ and GM-CSF production by flow cytometry, leukocytes were
isolated, treated, and stained for cell surface molecules as described above. Cells were
then fixed with 2% paraformaldehyde for at least 10 min, washed once with PBS + 2%
horse serum, and then washed once with 0.2% saponin (Sigma) in PBS + 2% horse
serum. Bovine IFNγ was detected using a PE-conjugated mouse IgG1 mAb against
bovine IFNγ (MCA1783E, ABD Serotec Inc., Raleigh, NC). Human IFNγ was detected
using a PE-conjugated mouse IgG1 mAb (clone 4S.B3, Biolegend). Human GM-CSF
was detected using a PE-conjugated rat IgG2a mAB (clone BVD2-21C11, Biolegend).
For isotype controls, cells were stained with a non-specific, PE-conjugated mouse IgG1
antibody (Biolegend) or rat IgG2a antibody (Biolegend). IFNγ antibodies and isotype
controls were resuspended in 0.2% saponin. Cells were stained for 25 min at room
temperature. After staining, cells were washed and then analyzed using a FACSCalibur
with HTS.
Statistical Analysis Statistical analyses were performed using Prism 4 (GraphPad
Software, San Diego, CA). The data were analyzed by Student’s paired t-test, Student’s
unpaired t-test, One-way ANOVA, or Two-way ANOVA as indicated.
90
Results
Oenothein B Stimulates and Induces
IFNγ Production by Bovine Adult T Cells
To examine potential differences between adults and calves in relation to the
stimulation of T cells by oenothein B, we stimulated isolated PBMCs from Holstein bull
calves approximately 6-weeks old and adult Holstein cows at least 2 years old with
oenothein B. First, we compared oenothein B-induced activation of adult and calf T cells,
using IL-2Rα as an activation marker, as we have done in previous studies31, 224.
Treatment with certain polyphenols enhance T cell proliferation in response to IL-2,
likely due to the upregulation of IL-2Rα, which promotes the formation of the high
affinity IL-2R complex31. Previous studies showed that oenothein B enhanced IL-2Rα
expression on calf lymphocytes224. Similar to these results in calves, treatment with
oenothein B enhanced IL-2Rα expression on T cells from adult cows (Figure 4.1A).
However, oenothein B induced less IL-2Rα expression on adult CD3+ T cells compared
to calf T cells. CD3+ T cells were then separated into γδ T cells and αβ T cells and
examined for IL-2Rα expression. Interestingly, oenothein B induced less IL-2Rα
expression on adult γδ T cells compared to calves, but there was no significant difference
in IL-2Rα expression on adult and calf αβ T cells (Figure 4.1B and 4.1C). These data
suggests that oenothein B may be less effective at priming adult bovine γδ T cells to
91
respond to IL-2, compared to calf T cells, but equally as effective at priming adult and
calf αβ T cells.
Next, potential differences between adults and calves in relation to IFNγ
production by T cells in response to oenothein B were examined. Previous results found
that considerable IFNγ was produced by bovine calf PBMCs following oenothein B
treatment, plus IL-18. However, T cells (either αβ or γδ T cells) produced little, if any, of
the induced IFNγ. NK cells were consistently the major source of IFNγ in the calf
preparations224. The lack of significant IFNγ production by T cells from calves, with
non-T cells producing the majority of oenothein B-induced IFNγ , was repeated here
(Figure 4.2A and 4.2B). However, in contrast, T cells from adult animals produced
significantly more IFNγ than calf T cells (Figure 4.2A). Indeed, upon two color FACS
analysis, T cells made up the majority of IFNγ-producing cells in these adult samples
(Figure 4.2B). This result was exactly as we observed for adult human donors in our
previous study224. These data suggested that the ability of oenothein B to enhance IFNγ
production by bovine T cells increases with age.
Oenothein B Enhances IFNγ Production
by Adult Bovine CD45RO+ αβ and γδ T Cells
Next, it was determined if memory T cells were an important source of this
oenothein B-induced IFNγ in adult bovine T cells. Numerous studies have found that
memory T cells produce more IFNγ than naive T cells in response to both antigendependent and antigen-independent stimulation99, 100, 222. Furthermore, the percentage of
92
memory T cells in PBMCs increases with age, correlating with pathogen exposure over
the lifespan103, 222. Therefore, it was examined if T cells expressing the memory cellmarker CD45RO were an important source of oenothein-B induced IFNγ in adult cows
(4-7 year old cows). In these experiments, cells expressing IFNγ in these animals were
largely CD3+ T cells, consistent with our earlier results (Figure 4.3A and 4.3D).
Furthermore, cells expressing CD45RO produced more IFNγ than cells that did not
express CD45RO, suggesting that CD45RO+ T cells were the major IFNγ-producing
cells in response to stimulation with oenothein B in these animals (Figure 4.3B and
4.3D). These data suggested that IFNγ production by bovine T cells in response to
oenothein B could be enhanced by increasing numbers of antigen-experienced CD45RO+
T cells. However, not all CD45RO+ cells produced IFNγ and a small population of
CD45RO- cells did express IFNγ in response to oenothein B, suggesting that the linkage
of CD45RO expression to the capacity of oenothein B to stimulate IFNγ production in T
cells in adult cattle is not absolute. Finally, oenothein B was found to induce IFNγ
production by both γδ TCR+ and γδ TCR- cells (Figure 4.3C and 4.3D). These data
suggest that, consistent with studies with adult human PBMCs, oenothein B induces IFNγ
production by both αβ T cells and γδ T cells from adults224.
93
Oenothein B Enhances IFNγ Production
by Purified Adult Bovine T Cells
To determine if differences in IFNγ production between adult and calf T cells was
due to differences in the T cells themselves, or differences in the composition of PBMCs,
CD4+ T cells, γδ T cells, and other T cells (CD4- γδ TCR-) were sorted from
Figure 4.1 Oenothein B induces IL-2Rα expression on bovine T cells from both adults
and calves. Bovine PBMCs (105 cells/well) from calves (<12 weeks old) and adult cows
(>2 years old) were treated with the indicated concentrations of oenothein B or X-VIVO
medium only for 24 hrs and IL-2Rα expression on CD3+ T cells (A), αβ T cells (B), and
γδ T cells (C) was measured by multi-color flow cytometry. αβ T cells were characterized
as CD3+ cells that did not express γδ TCR. The graphs represent pooled data. Each
treatment was analyzed in triplicate and error bars indicate SEM. Significance compared
to untreated cells (0 µg/ml) and between calves and adults was determined by Two-way
ANOVA with Bonferroni post-test. *p<0.05, **p<0.01, ***p<0.001
94
Figure 4.2 Oenothein B induces more IFNγ production by adult bovine T cells than calf T
cells. (A) Bovine PBMCs (105 cells/well) from calves (<12 weeks old, N=4) and adult
cows (>2 years old, N=4) were treated with the indicated concentrations of oenothein B
in X-VIVO medium for 42 hrs with brefeldin A added for the final 18 hrs. The percent of
CD3+ T cells which were IFNγ + was measured by multi-color flow cytometry. The
graph represents pooled data. Each treatment was analyzed in duplicate and error bars
indicate SEM. Significance was determined by Two-way ANOVA with Bonferroni posttest. (B) Representative examples of two-color flow cytometry plots of adult and calf
bovine PBMCs treated with 20 µg/ml oenothein B or X-VIVO medium alone.
95
bovine adults and calves. When these purified cells were stimulated with oenothein B,
CD4+ T cells from adults expressed more IFNγ in response to direct stimulation by
oenothein B than those from calves (Figure 4.4A). Neither γδ T cells nor other T cells
from calves or adults expressed IFNγ in response to direct stimulation by oenothein B
(Figure 4.4B and 4.4C). These data suggest that oenothein B promotes greater IFNγ
expression by adult bovine T cells, at least CD4+ T cells, due to T cell-intrinsic
differences between adult and calf T cells.
Differentiation of Calf T Cells into
CD45RO+ T Cells is not Sufficient to
Enhance Oenothein B-induced IFNγ Production
CD45RO expression and IFNγ secretion is upregulated in T cells following
mitogen (ConA/IL-2) stimulation in vitro225, 226. As such, it was then determined whether
treatment with ConA to increase CD45RO expression, also increased the capacity of the
same cells to produce IFNγ following oenothein B stimulation. Bovine calf PBMCs were
stimulated with ConA and IL-2 for 4 days in vitro to simulate TCR stimulation and
promote differentiation into CD45RO+ cells. Control PBMCs were cultured without
ConA or IL-2. After 4 days, cells were allowed to rest in the absence of ConA/IL-2 for 2
additional days. As expected, treatment with ConA/IL-2 resulted in an increased
percentage of CD45RO+ cells compared to unstimulated cells (Figure 4.5A). However,
when these cells were subsequently re-stimulated with oenothein B, there was no increase
96
Figure 4.3 Oenothein B induces more IFNγ production by bovine CD45RO+ T cells than
CD45RO- T cells. Bovine PBMCs (105 cells/well) isolated from adult cows (n=2) were
treated with the indicated concentrations of oenothein B or X-VIVO medium alone for 24
hrs, with brefeldin A added for the final 6 hrs. The percentage of CD3+ (A), CD45RO+
(B), or γδ TCR+ (C) cells producing IFNγ was measured by multi-color flow cytometry.
The graphs represent pooled data. Each treatment was analyzed in duplicate and error
bars indicate SEM. Significance was determined by Two-way ANOVA with Bonferroni
post-test. (D) Representative examples of two-color flow cytometry plots of PBMCs
treated with 20 µg/ml oenothein B or medium alone. *p<0.05, **p<0.01, ***p<0.001
97
Figure 4.4 Oenothein B enhances IFNγ production by purified bovine CD4+ T cells from
adults, but not calves. CD4+ T cells (A), γδ T cells (B), and all other (CD4- γδ TCR-)
CD3+ T cells (C) were sorted from bovine adults (N=3) and calves (N=3), plated (104
cells/well), and treated with indicated concentrations of oenothein B or X-VIVO medium
alone. After 24 hrs, soluble IFNγ were measured by ELISA. The graphs represent pooled
data. Error bars indicate SEM. Each sample was analyzed in duplicate. Significance was
determined by Two-way ANOVA with Bonferroni post-test. ***p<0.001
98
Figure 4.5 Enhancing CD45RO expression in vitro does not enhance oenothein Binduced IFNγ production by calf T cells. Bovine PBMCs (105 cells/well) from calves
(<12 weeks old, N=3) were treated for 4 days with ConA (500 ng/ml) and IL-2 (1 ng/ml)
or X-VIVO medium only. After 4 days, medium was replaced with X-VIVO only for an
additional 2 days. PBMCs were then washed with X-VIVO and treated with the indicated
concentrations of oenothein B in X-VIVO medium for 42 hrs with brefeldin A added for
the final 18 hrs. (A) The percent PBMCs which were CD45RO+ was calculated in cells
not treated with oenothein B. Significance was determined by Paired T test. (B) The
percent of CD3+ T cells which are IFNγ + was measured by multi-color flow cytometry.
The graph represents pooled data. Each treatment was analyzed in duplicate and error
bars indicate SEM. Significance was determined by Two-way ANOVA with Bonferroni
post-test. **p<0.01
99
in the percentage of IFNγ-producing cells in the ConA/IL-2 cultured T cells compared to
T cells cultured in the absence of ConA/IL-2 (Figure 4.5B). These data suggest that the
increased frequency of CD45RO+ T cells alone is not sufficient to explain the increase in
IFNγ production by adult T cells in response to oenothein B.
Oenothein B Enhances IFNγ Production by
Human Adult T Cells, but not Cord Blood T Cells
To determine if the observations made with bovine lymphocytes also held true in
human lymphocytes, the ability of oenothein B to stimulate human adult and cord blood
T cells was tested. Oenothein B could activate cord blood T cells, as measured by
increased expression of CD69, although the activation of cord blood T cells was not as
robust as oenothein B-stimulated adult T cells (Figure 4.6). When human adult and cord
blood PBMCs were stimulated with oenothein B, adult PBMCs, but not cord blood
PBMCs, produced IFNγ (Figure 4.7A). Furthermore, oenothein B induced IFNγ
production by adult, but not cord blood, T cells (Figure 4.7B and 4.7C). These data were
consistent with our data comparing bovine calves and adults. In addition to IFNγ,
oenothein B induced GM-CSF production by adult human PBMCs and T cells, but not
cord blood PBMCs and T cells (Figure 4.8). Altogether, these data suggested that, while
oenothein B can activate both cord blood and adult T cells, oenothein B induces more
IFNγ and GM-CSF production by T cells from human adult mononuclear cell cultures
100
compared to neonate mononuclear cell cultures, supporting our observations in bovine
lymphocytes.
Human CD3+ T cells
***
***
200
175
**
MFI CD69
150
Adult
Cord Blood
125
100
75
50
25
0
0
20
40
μg/ml oenothein B
Figure 4.6 Oenothein B induces CD69 expression on human cord blood T cells. Human
blood mononuclear cells (5X104 cells/well) isolated from cord blood (n=3) and sexmatched adult donors (n=3) were treated with the indicated concentrations of oenothein B
or cRPMI medium only for 42 hrs and CD69 expression on CD3+ T cells was measured
by multi-color flow cytometry. The graphs represent pooled data. Each treatment was
analyzed in triplicate and error bars indicate SEM. Significance compared to untreated
cells (medium) and between cord blood and adult samples was determined by Two-way
ANOVA with Bonferroni post-test. **p<0.01, ***p<0.001
101
Figure 4.7 Oenothein B induces IFNγ expression on human adult, but not cord blood, T
cells. (A) Human blood mononuclear cells (5X104 cells/well) from cord blood (n=3) and
adult donors (n=3) were treated with the indicated concentrations of oenothein B or
cRPMI medium alone for 24 hrs, and soluble IFNγ levels in supernatant fluids were
measured by ELISA. The graph represents data from each sample, with each sample
plated in triplicate. Statistical significance between adults and cord blood was measured
by Two-way ANOVA with Bonferroni post-test. (B) Human blood mononuclear cells
(5X104 cells/well) isolated from a cord blood and adult donor were treated with the
indicated concentrations of oenothein B or X-VIVO medium alone for 24 hrs, with
brefeldin A added for the final 6 hrs. The percent of CD3+ T cells expressing IFNγ was
measured by multi-color flow cytometry. Each treatment was analyzed in duplicate and
error bars indicate SEM. Significance between cord blood and adult samples was
determined by Two-way ANOVA with Bonferroni post-test. (C) Representative examples
of two-color flow cytometry plots of human adult and cord blood PBMCs treated with 20
µg/ml oenothein B or medium alone. *p<0.05, **p<0.01, ***p<0.001
102
Oenothein B Induces More Cytokine Production
by Human Memory T Cells Than Naive T Cells.
In the bovine experiments described earlier, CD45RO+ memory T cells produced
more IFNγ than CD45RO- cells. To see if this was also true in humans, the ability of
oenothein B to stimulate human CD45RO+ and CD45RO- T cells was compared. When
human adult PBMCs were stimulated with oenothein B, CD69 expression was
upregulated on both CD45RO+ and CD45RO- T cells, with greater expression on
CD45RO- cells (Figure 4.9A).
In contrast to CD69, IFNγ and GM-CSF expression by T cells was enhanced in
CD45RO+ T cells compared to CD45RO- cells (Figure 4.9B). To further confirm these
human results, CD45RO+ and CD45RO- T cells were sorted from adult human donors.
When these purified cells were stimulated with oenothein B, CD45RO+ T cells expressed
more IFNγ and GM-CSF in response to direct stimulation by oenothein B than CD45ROT cells (Figure 4.9C). These data suggested that, as in the cow, oenothein B promotes
greater cytokine expression by CD45RO+ memory T cells, despite enhancing CD69
expression on both CD45RO+ and CD45RO- T cells. The sorted T cell data also suggest
that oenothein B enhances greater cytokine production by CD45RO+ T cells compared to
CD45RO- T cells even in the absence of macrophages and other non-T cell PBMCs.
103
Figure 4.8 Oenothein B induces GM-CSF expression on human adult, but not cord
blood, T cells. (A) Human blood mononuclear cells (5X104 cells/well) from cord
blood (n=3) and adult donors (n=3) were treated with the indicated concentrations
of oenothein B or cRPMI medium alone for 24 hrs, and soluble GM-CSF levels in
supernatant fluids were measured by ELISA. The graph represents data from each
sample, with each sample plated in triplicate. Statistical significance between
adults and cord blood was measured by Two-way ANOVA with Bonferroni posttest. (B) Human blood mononuclear cells (5X104 cells/well) isolated from cord
blood (N=2) and adult (N=2) donors were treated with the indicated concentrations
of oenothein B or cRPMI medium alone for 24 hrs, with brefeldin A added for the
final 6 hrs. The percent of CD3+ T cells expressing GM-CSF was measured by
multi-color flow cytometry. The graph represents pooled data. Each treatment was
analyzed in duplicate and error bars indicate SEM. Significance between cord
blood and adult samples was determined by Two-way ANOVA with Bonferroni
post-test. (C) Representative examples of two-color flow cytometry plots of human
adult and cord blood PBMCs treated with 20 µg/ml oenothein B or medium alone.
*p<0.05, **p<0.01, ***p<0.001
104
Altogether, these data are consistent with our bovine data, where activation of T cells, as
measured by increased expression of an activation marker, does not always correlate with
cytokine expression by oenothein B-stimulated T cells.
Discussion
Plant polyphenols have been studied extensively for numerous health benefits,
including enhanced immune activity. Polyphenols isolated from several sources activate
lymphocytes, including NK cells and T cells, promoting the production of various
proteins, including IFNγ32, 179, 224. The production of IFNγ by T cells is critical for
immune defense against many viral, bacterial, and other infections60, 205, 206. It is also
critical for defense against tumors207. Therefore, enhanced production of IFNγ by T cells
may play an important role in the proposed ability for certain polyphenols and other food
products to protect against infection and tumors. However, there is much that is still
unknown about the immunostimulatory properties of polyphenols, including what factors
may influence their activity.
105
Figure 4.9 Oenothein B enhances more cytokine production by human CD45RO+
memory T cells than naïve T cells. Human PBMCs (105 cells/well) isolated from adult
donors (n=3) were treated with the indicated concentrations of oenothein B or cRPMI
medium alone for 24 hrs, with brefeldin A added for the final 6 hrs. CD69 (A), as well as
IFNγ and GM-CSF (B) expression on CD3+ T cells was measured by multi-color flow
cytometry. The graphs represent pooled data from three individuals. Each treatment was
analyzed in duplicate and error bars indicate SEM. Significance between CD45RO+ and
CD45RO- CD3+ T cells was determined by Two-way ANOVA with Bonferroni posttest. (C) Human CD45RO+ and CD45RO- CD3+ T cells (105 cells/well) were sorted and
treated with indicated concentrations of oenothein B or cRPMI medium alone. After 24
hrs, soluble IFNγ and GM-CSF were measured by ELISA. The graphs represent pooled
data of soluble IFNγ levels in culture supernatant fluids from three separate experiments
with different donors and soluble GM-CSF levels from two separate experiments with
different donors. Error bars indicate SEM. Each sample was analyzed in triplicate.
Significance between CD45RO+ and CD45RO- CD3+ T cells was determined by Twoway ANOVA with Bonferroni post-test. *p < 0.05, **p<0.01, ***p<0.001
106
One factor that appears to be important for stimulation of T cell cytokine
production by polyphenols is the size of the polyphenol molecule. Work by Yamanaka
and coworkers found that stimulation of murine splenocytes by polymerized polyphenols
with large molecular weights, but not their corresponding monomers, enhances cytokine
production, including IFNγ and GM-CSF, by T cells179. Furthermore, we have found that
procyanidin oligomers, but not monomers, stimulate innate lymphocytes15. The
importance of molecular size is consistent with the activity of oenothein B, as it is a large
polyphenol and molecular subunits of this compound do not have the same
immunostimulatory capabilities33. An additional factor may be the chemical modification
of polyphenol structures, such as methylation. A recent study found that some methylated
quercetin molecules, but not unmethylated quercetin, could enhance TLR2-induced IL-1
production by a monocyte cell line. Furthermore, not all methylated quercetin molecules
enhanced IL-1 production, suggesting an exquisite level of chemical specificity in
modulation of innate immune cell activity by polyphenols190. It will be interesting to
determine if such chemical specificity also exists for immunostimulation of T cells and
other lymphocytes by polyphenols.
In this study, we propose that age may be an important human and animal factor
influencing the immunostimulatory properties of polyphenols. The immune system of
both humans and animals changes significantly as individual's age from birth to
adulthood to old age. Infants and young individuals have immature immune systems and
107
are more susceptible to a number of different diseases227, 228. T cells and other
lymphocytes change their behavior with aging70, 87, 170, 222. Subset distribution, receptor
expression, and other features of T cells differ between young individuals and adults, and
their responsiveness to various agonists also changes105, 155, 169, 170, 222. Our data suggest
that T cell responsiveness to large-molecular weight polyphenols can also change with
age.
Here, we show that T cells from both young and adult individuals are stimulated
by oenothein B, but differ in their responses. γδ T cells from calves express more IL-2Rα
in response to oenothein B than adult T cells, while αβ T cells from calves and adults
express similar levels of IL-2Rα in response to oenothein B. This suggests that oenothein
B may prime αβ T cells from both young calves and adults equally to IL-2-induced
proliferation, but may better prime γδ T cells from young calves. We also show that T
cells from both human and bovine adults secrete IFNγ and GM-CSF in PBMCs treated
with oenothein B. However, T cells from young individuals, either bovine calves or
human cord blood, produce less of these cytokines in response to oenothein B. This is
true for both αβ and γδ T cells. Altogether, these data suggest that the benefit of
oenothein B, and perhaps other polyphenols, on T cell activity may change with age,
possibly promoting enhanced T cell proliferation and expansion in young individuals
while promoting enhanced cytokine production in older individuals.
108
In addition, we show that CD45RO+T cells, in mixed cultures as well as sorted,
produce more IFNγ and GM-CSF in response to oenothein B than CD45RO- T cells in
both cows and humans. This could account, at least in part, for the enhanced cytokine
secretion by adult T cells, as CD45RO+ memory T cells accumulate with age as a result
of increased antigen exposure over time103. However, when we used ConA and IL-2 to
increase the percentage of CD45RO+ cells in calf PBMCs, IFNγ production by these T
cells in response to oenothein B was not significantly enhanced. Therefore, in addition to
an increased percentage of antigen-experienced T cells in adults, other factors in the adult
microenvironment may play an important role in enhancing cytokine production by adult
T cells in response to oenothein B. This would be consistent with our data that shows
that, even though CD45RO+ T cells express several times more cytokines in response to
oenothein B than CD45RO- cells, CD45RO- T cells from adults appear to produce more
IFNγ and GM-CSF in response to oenothein B than CD45RO- T cells from younger
individuals. However, an alternative explanation for the enhanced cytokine production by
adult CD45RO- T cells could be the loss of CD45RO expression on some activated T
cells, as CD45RO expression is known to sometimes drift229.
The mechanism for the enhanced antigen-independent cytokine production of
adult T cells compared to young T cells has yet to be determined. One possible
explanation, as oenothein B is known to stimulate myeloid cells, would be enhanced
IFNγ production by memory and adult T cells in response to myeloid cell-derived
109
cytokines, such as IL-12 and IL-18. However, our data with sorted T cells would suggest
that oenothein B can directly enhance cytokine production by adult and memory T cells.
Furthermore, our IL-2Rα and CD69 data suggest that naive T cells and T cells from
young individuals are just as responsive to stimulation by oenothein B as adult and
memory T cells, if not more so, so enhanced stimulation by oenothein B is not a likely
explanation. Another possible explanation involves methylation at cytokine promoter
sites in adult and memory T cells. Methylations at certain CpG sites within DNA are
known to prevent gene transcription. Studies have found that reduced methylation at sites
within the ifnγ promoter in memory T cells compared to naive T cells results in enhanced
ifnγ transcription and cytokine production in memory T cells. Furthermore, inhibiting
DNA methylation enhances IFNγ production by naive T cells230. In addition, another
study found that CD45RO- CD4+ T cells from adult T cells had reduced methylation at
the ifnγ promoter compared to cord blood T cells231. It is possible that reduced
methylation at cytokine promoter sites in adult T cells allow for enhanced IFNγ and GMCSF production by adult and memory T cells in response to oenothein B. However, this
would need to be tested experimentally.
In addition to their presence in blood, CD45RO+ memory T cells migrate into a
number of different tissues, including portals of entry into the body such as the lung and
gut94. This suggests that they may be prime targets for immunomodulatory food products.
While mostly studied for their antigen-specific responses, memory T cells can also play
110
an important role in innate immunity through antigen-independent, bystander effects.
These bystander effects include the early production of IFNγ, as well as the killing of
infected cells by NKG2D-dependent mechanisms99-102. Therefore, although they are
typically associated with adaptive immunity, they can also be characterized as innate
lymphocytes. Therefore, in addition to γδ T cells and NK cells, CD45RO+ memory T
cells may be another innate-like lymphocyte population whose numbers and activity
could be influenced by polyphenols and this should be taken into account when
examining enhancing effects of polyphenols on innate immunity. Further studies are
needed to determine what effect polyphenols have on CD45RO+ T cells and their activity
in vivo, as well as the possible importance of CD45RO+ T cells for certain polyphenols
to optimally enhance immunity to various pathogens and cancers. This could be an
important aspect that has been largely overlooked in mouse models, as mice are typically
housed in pathogen-free conditions and have low CD45RO+ memory T cell numbers
compared to humans and cows of a similar maturity level.
Our data further emphasizes the utility of the bovine system to study dietary
polyphenols effects on the immune system and, in particular, the influence of age on the
impact of dietary polyphenols. Studies comparing very young and adult individuals in
vivo are difficult, as experiments on human children are rarely done for obvious ethical
reasons and mice usually need to reach several weeks of age before they are large enough
to handle and treat. In addition, pathogen-free housing and the inbred nature of laboratory
111
mice are significant variables when comparing results to outbred animals and humans.
However, studies examining age-related changes of the bovine immune system are
common86, 87, 89, 170. Domestic cattle are also large enough to handle and feed at a very
young age and are often used in diet-influenced immunity studies. In one study
examining the effect of feeding polyphenol-rich pomegranate extracts to calves during
the first 70 days of age, the authors found that feeding pomegranate extract enhanced
PHA-induced IFNγ and IL-4 production by PBMCs30. It would be of great interest to
determine if T cells were the primary source of these cytokines and if using older animals
would have affected these results. Furthermore, this study and others suggest that bovine
lymphocytes are predictive for the influence of polyphenols on human lymphocytes31, 224.
Therefore, cattle may be a useful model for studying the role of age, particularly between
very young individuals to adults, on the immunomodulatory properties of food products,
such as polyphenols.
These data are the first to our knowledge to demonstrate differences in immune
cell responsiveness to polyphenols with age. Further work is required to determine if our
observations with oenothein B also holds true for other immunomodulating polyphenols,
especially those which stimulate T cells. In addition, further work is required to
determine if our observations also hold true in vivo. These data has implications for
optimizing the use of dietary polyphenols to enhance immunity in both humans and
112
animals and suggest that subject age should be taken into account when examining the
effect of consumption of polyphenol-rich supplements on immunity.
Acknowledgments
We would like to thank Dr. Robyn Klein (Department of Plant Sciences and Plant
Pathology, Montana State University, Bozeman, MT) for plant identification. We thank
Dr. James Berardinelli (Department of Animal and Range Sciences, Montana State
University, Bozeman, MT) and Kerri Jones for providing the bovine blood. We also
thank Larissa Jackiw for her assistance in FACS sorting, as well as Dr. Jodi Hedges for
helpful scientific discussion.
113
CHAPTER FIVE
COMPLEX ROLE OF γδ T CELL-DERIVED CYTOKINES
AND GROWTH FACTORS IN CANCER.
Contribution of Authors and Co-Authors
Manuscript in Chapter 5
Author: Andrew G. Ramstead
Contributions: Primary author of the manuscript.
Co-author: Mark A. Jutila
Contributions: Co-wrote and edited manuscript.
114
Manuscript Information Page
Andrew G. Ramstead and Mark A. Jutila (PhD)
Journal Name: Journal of Interferon and Cytokine Research
Status of Manuscript:
___ Prepared for submission to a peer-reviewed journal
___ Officially submitted to a peer-reviewed journal
___ Accepted by a peer-reviewed journal
_X_ Published in a peer-reviewed journal
Published by: Mary Ann Liebert, Inc.
Reference: J Interferon Cytokine Res. 2012 Dec;32(12):563-569.
Reprinted with permission from JOURNAL OF INTERFERON & CYTOKINE
RESEARCH, 2012, published by Mary Ann Liebert, Inc/. New Rochelle, NY.
115
During our preliminary experiments with the B16 melanoma tumor model, we
saw a reduction in the mean tumor weight in wild-type mice treated with oenothein B, but
an increase in mean tumor weight in γδ T cell-deficient animals. These data suggested
that γδ T cells may have an important role in the antitumor activity of oenothein B and
that this compound had both anti- and protumor effects. Unfortunately, inconsistencies in
our animal systems lead to inconsistencies in these results and an inability to fully test
these hypotheses. A major inconsistency in our animal system was the observed activity
of γδ T cells during B16 melanoma growth. In experiments conducted in 2010, salinetreated γδ-/- mice consistently had smaller tumors than saline-treated wild-type C57BL/6
mice (Figure 5.1A). This suggested that the γδ T cells in our model were pro-tumor and
potentially inhibited tumor immunity. However, in experiments conducted in 2011, B16
tumors grew larger in untreated γδ-/- mice compared to wild-type mice (Figure 5.1B).
Similar results were seen in 2012, when saline-treated γδ-/- mice consistently had larger
tumors than saline-treated wild-type mice (Figure 5.1C). Therefore, in our 2011 and 2012
experiments, γδ T cells appeared to have antitumor activities, opposite of what was
observed in 2010, but similar to what has been observed in the literature202. Altogether,
the γδ T cells in our mice appeared to shift from a protumor, regulatory phenotype to an
antitumor, inflammatory phenotype. This variability in γδ T cell responses may have had
a profound impact on the ability of oenothein B to influence tumor growth. This
contradictory γδ T cell behavior in the context of tumor immunology has been observed
116
in other murine studies, as well as in the clinic. In this chapter, I present a published
review of the literature examining the complex role of γδ T cells in cancer.
Figure 5.1 Changes in B16 melanoma growth in γδ T cell-deficient mice compared to
wild-type mice over time. (A) In 2010, C57BL/6 (N=20) and γδ T cell-deficient (N=15)
animals were injected with 5 X 105 B16 melanoma cells subcutaneously on day 0. Mice
were injected i.p. on days -1, 4, 7, and 11 with saline. Mice were euthanized on day 16
and tumor weights were recorded. (B) In 2011, C57BL/6 (N=10) and γδ T cell-deficient
(N=10) animals were injected with 1 X 106 B16 melanoma cells subcutaneously on day
0. Mice were euthanized on day 14 and tumor weights were recorded. (C) In 2012,
C57BL/6 (N=4) and γδ T cell-deficient (N=3) animals were injected with 5 X 105 B16
melanoma cells subcutaneously on day 0. Mice were injected i.p. on days -1, 4, 7, and 11
with saline as described in part A. Mice were euthanized on day 16 and tumor weights
were recorded. All mice were male and 8-14 weeks old. *** p < 0.001
117
Abstract
γδ T cells are innate lymphocytes, which recognize and kill a range of tumor cells and
are currently being explored as a target for tumor immunotherapy. However, γδ T cells
have a complex role in cancer and can promote, as well as inhibit, tumor growth. In
addition to tumor cell killing, γδ T cells express a number of cytokines and other soluble
factors in response to tumors. Soluble factors expressed by γδ T cells in these settings
include IFNγ, TNFα, IL-4, IL-10, TGFβ, IL-17, and a number of growth factors. These
factors have differing, sometimes opposing effects on anti-tumor immunity and tumor
angiogenesis, and likely contribute to the complex role of these cells in cancer. Here, we
review studies in both mice and humans examining differential cytokine secretion by γδ
T cells in response to tumors and tumor immunotherapy, and discuss the influence of
these γδ T cell-derived factors on tumor growth.
Introduction
Immunity has long been known to impact cancer development in a diverse
manner. Effective immune surveillance of cancerous tumors can suppress tumor growth,
but improper and/or prolonged immune activity can actually contribute to its initiation
and progression232, 233. Immune responses at tumor sites are a balancing act of
inflammatory and regulatory responses mediated by many different immune cells and
cytokines, many of which display dual functionality by both promoting and inhibiting
118
tumor growth233. Among the immune cells, γδ T cells may be important during the
establishment of the tumor microenvironment and development of tumor immunity.
Recently, the role of γδ T cells in tumor immunity has received considerable
attention and research. γδ T cells from both humans and mice infiltrate tumor sites, lyse
tumor cells, and prevent the growth of a variety of cancers202, 234-239. Tumor cell
recognition by γδ T cells is largely mediated through the recognition of membrane-bound
phosphoantigens, such as isopentenyl pyrophosphate (IPP), by the γδ T cell receptor
(TCR) and/or the recognition of stress ligands on the tumor cell through the TCR and
NKG2D196. Due to their anti-tumor activity, therapeutic strategies aimed at harnessing
and enhancing the anti-tumor properties of these cells have been developed and used in
the clinic196, 240. These therapies often require IL-2 combined with synthetic
phosphoantigens or bisphosphonates, such as zoledronate, which stimulate γδ T cells by
enhancing cellular accumulation of IPP 240-242. While these therapies show potential,
optimal results have not yet been achieved. Several recent reviews have examined the
anti-tumor activity of γδ T cells and their potential for immunotherapy (Table 1)196, 240,
243-247
.
Despite the evidence demonstrating anti-tumor responses by γδ T cells, the exact
role these cells play in cancer is not entirely clear. In mice, the absence of γδ T cells
sometimes leads to enhanced tumor growth, but in some cases leads to a reduction in
tumor burden202, 236, 248, 249. In human patients, infiltration of γδ T cells into the tumor is
119
associated with better prognosis in some cancers250, but not in others251. These data
suggest that, depending upon the tumor, γδ T cells can promote, inhibit, or possibly have
no significant effect on tumor growth. These differential roles are likely mediated, at least
in part, by the diverse repertoire of cytokines and other secreted factors that are induced
in these cells, which can be categorized as either inflammatory or regulatory252. A better
understanding of the diverse roles of γδ T cells and their secreted factors in cancer should
allow for better manipulation of these cells for immunotherapy. In this review, we will
summarize the literature with regard to different cytokines and other secreted factors
expressed by γδ T cells in response to tumors and examine how these factors could
impact tumor immunity and immunotherapy.
γδ T Cell-associated Factors Which Enhance Anti-tumor Immunity
γδ T cells are an important early source of the inflammatory cytokines IFNγ and
TNFα in many infections and other disease models253. The expression of IFNγ and TNFα
by γδ T cells is promoted by numerous stimuli, including TCR agonists, ligands to
NKG2D, and certain cytokines, such as IL-12 and IL-18130, 235, 254, 255. IFNγ and TNFα are
also important cytokines in anti-tumor responses and inhibit tumor growth through
several mechanisms, including the enhancement of anti-tumor immunity and inhibition of
tumor angiogenesis256, 257, 258. Human γδ T cells express IFNγ and TNFα upon exposure
120
to tumor cell lines of numerous origins235, 259, 260, suggesting these cytokines may play a
role in γδ T cell responses to tumors.
Table 5.1 Select recent reviews for γδ T cells and cancer
Topic
γδ T cell immunotherapy
Selected article
Hananni and others Trends Immunol 2012
Castella and others Cell Mol Life Sci 2011
Kalyan and others Curr Med Chem 2011
Yoshida and others Surg Today 2011
Gomes and others Cancer Research 2010
Tumor escape from γδ T cell
Capietto and others Cell Mol Life Sci 2011
attack
γδ T cell antigen presentation
Moser and Eberl Cell Mol Life Sci 2011
for cancer therapy
In mice, γδ T cells appear to be an important early source of tumor-induced IFNγ
and the expression of IFNγ is essential for optimal anti-tumor responses by these cells202,
238
. The early production of IFNγ by murine γδ T cells can enhance MHCI expression on
121
tumors as well as enhance CD8+ T cell responses202, 261 suggesting γδ T cells could be
important for augmenting downstream adaptive immune responses to tumors. These data
suggest that early IFNγ secretion by γδ T cells is important for some anti-tumor responses
in mice.
Results in humans suggest a far more complicated story in regard to the role of γδ
T cell-derived IFNγ and TNFα in anti-tumor responses. In cancer patients, the expression
of both IFNγ and TNFα by γδ T cells is modulated, but not always enhanced. Peripheral
γδ T cells from breast cancer patients produce enhanced amounts of TNFα compared to
healthy controls, which is thought to be beneficial262. However, peripheral γδ T cells
from patients with nasopharyngeal cancer and melanoma produce reduced amounts of
IFNγ and TNFα, which could contribute to defective anti-tumor immune responses in
these patients263, 264. Upon the removal of melanoma, IFNγ and TNFα expression by γδ T
cells is enhanced, suggesting that the reduced expression of these cytokines by γδ T cells
is mediated by tumor-associated factors, which benefit the tumor265. In support of this,
mesenchymal stem cells, which are commonly found in tumor microenvironments, were
shown to inhibit IFNγ and TNFα expression by peripheral γδ T cells through the
production of prostaglandin E2, which was induced by γδ T cell-derived IFNγ and
TNFα266. In cancer patients undergoing immunotherapy with zoledronate and IL-2, serum
levels of IFNγ increase after treatment267. This increase in IFNγ expression by γδ T cells
may be an important factor for successful γδ T cell immunotherapy, as clinical responses
122
to immunotherapy with zoledronate and IL-2 in one clinical trial correlated with
increasing numbers of an effector memory γδ T cell phenotype, which could produce
IFNγ241. However, in another clinical trial using infusions of zoledronate-activated γδ T
cells in multiple myeloma patients, IFNγ was not believed to be important for the antitumor activity even though serum levels of IFNγ increased after treatment268.
Collectively, these data suggest that the expression of IFNγ and TNFα is important in
certain cancers for anti-tumor responses by γδ T cells, and that downregulation of γδ T
cell-derived IFNγ and TNFα may help facilitate immune escape by tumors. However,
further studies are needed to better determine their importance in human patients,
particularly in response to immunotherapy.
γδ T Cell-associated Factors Which Suppress Anti-tumor Immunity
As mentioned above, γδ T cells may not always play a beneficial role in antitumor immunity. Instead, in some settings they likely have a regulatory role, suppressing
anti-tumor responses and enhancing tumor growth. This response is not species-specific,
in that immunosuppressive γδ T cells have been described in both mouse tumor models
and human cancers237, 248. Furthermore, their activity appears to be at least partially
mediated by certain cytokines.
In a study by Seo and coworkers269, murine γδ T cells infiltrating B16 melanoma
tumors after 5 days were shown to inhibit NK and NKT cell activity and express large
123
amounts of IL-4 and IL-10, but not IFNγ. The supernatant fluids from cultures of these
cells did not reduce NK and NKT cell cytotoxicity, but did reduce their proliferation,
suggesting that soluble IL-4 and IL-10 contributed to the inhibition of NK and NKT cell
activity by γδ T cells in this model. Additional studies supported this observation and
showed that γδ T cell-derived IL-4 and IL-10, as well as TGFβ, could inhibit anti-tumor
immunity and promote tumor growth in mice. For example, using the B16 melanoma
model, Hao and coworkers270 showed that the Vγ1 subset of murine γδ T cells promoted
tumor growth through the production of IL-4. These Vγ1 γδ T cells reduced the
expression of IFNγ and perforin within the tumor. Also, IL-4 inhibited the expression of
NKG2D and perforin by Vγ4 γδ T cells, which was important for the tumor-promoting
activity of these Vγ1 γδ T cells. Seo and coworkers248 showed that tumor-infiltrating γδ T
cells from MM2 tumor lesions in mice expressed IL-10 and TGFβ, but not IFNγ or IL-4.
γδ T cells isolated from the tumor lesions, as well as the spleens, of these MM2 tumorbearing mice inhibited the cytotoxic activity of NK cells and CD8+ T cells. Neutralizing
IL-10 and TGFβ inhibited some of the suppressive effects of these γδ T cells, suggesting
these cytokines participated in the suppressive activity of these cells. Depletion of these
γδ T cells by the use of a specific antibody enhanced anti-tumor immunity and reduced
tumor growth. Finally, a study by Ke and coworkers249 also described an
immunosuppressive function for γδ T cells in tumor responses, as γδ T cells suppressed
responses to an EL4 leukemia tumor cell line modified to express OVA, and IL-10
124
appeared to play a role in the suppression. Collectively, these data strongly suggest that at
least certain subsets of murine γδ T cells can express IL-4, IL-10, and TGFβ in response
to certain tumors, inhibiting anti-tumor immunity.
Immunosuppressive γδ T cells may also play an important role in human cancers.
In a study by Peng and coworkers237, the Vδ1 subset of tumor-infiltrating γδ T cells from
human breast cancer could suppress DC maturation and T cell effector functions, which
included proliferation, IL-2 secretion, and CD8+ T cell anti-tumor responses in a mouse
xenograft model. This suppressive activity was mediated, at least in part, by a soluble
factor or factors. The suppressive activity was present in isolated fractions with greater
than 100 kDa molecular mass and could be inactivated by heat, but not DNAse or
RNAse. However, the factors were not identified. When these cells were stimulated by
tumor cells and anti-CD3 antibody, they expressed cytokines typically associated with
pro-inflammatory responses, including IFNγ, GM-CSF, and IL-6, but not IL-1β, TNFα,
IL-12, IL-2, IL-4, IL-10, or TGFβ. These Vδ1 γδ T cells made up a large percentage of
tumor-infiltrating lymphocytes in breast and prostate cancer, suggesting that they may be
important in promoting an immunosuppressive microenvironment in these cancers.
However, Vδ1 γδ T cell-infiltration into necrotizing melanomas has correlated with
increased survival250, suggesting that the development of suppressive Vδ1 γδ T cells may
be specific for certain cancers. Even though the suppressive effects of these cells were
not mediated by IL-10 or TGFβ, these results resemble those found in mice by Seo and
125
coworkers248, where infiltrating γδ T cells suppressed the activity of CD8+ T cells by
secreted factors. Interestingly, stimulation of these suppressive breast cancer Vδ1 γδ T
cells by a TLR8 agonist could reverse the suppression of anti-tumor responses237.
Even though human γδ T cells may secrete different soluble factors than murine
γδ T cells, which suppress anti-tumor immunity, certain human peripheral γδ T cells do
express IL-4, IL-10, and TGFβ upon activation254, 271. In one study, culture of human γδ T
cells with IPP or Daudi lymphoma cells in vitro under Th2-polarizing conditions (rhIL-4,
anti-IL-12) resulted in reduced IFNγ and TNFα production and enhanced IL-4 production
by these γδ T cells254. In the absence of these polarizing conditions, γδ T cells primarily
secreted IFNγ. Furthermore, a study by Gaafar and coworkers262 showed that while γδ T
cells from breast cancer patients produced very little IL-4, expansion of these cells by
zoledronate and IL-2 led to increased production of IL-4 by these cells compared to
expanded γδ T cells from healthy controls. Therefore, IL-4, IL-10, and TGFβ production
by human γδ T cells may also play a role in suppressing anti-tumor responses, like they
do in mice. However, additional studies are needed to confirm this possibility.
Collectively, the results summarized above support the idea that certain human γδ
T cells, at least in some cancers, can behave as regulatory cells within the tumor
microenvironment and suppress anti-tumor responses and promote tumor growth, with
secreted factors important for their activity.
126
Conflicting Role of γδ T Cell-derived IL-17 on Tumor Immunity
In addition to their role in tumor responses, a renewed interest in γδ T cells has
also emerged due to the discovery that γδ T cells are an important innate source of IL-17,
particularly in the mouse. Secretion of IL-17 by murine and human γδ T cells is promoted
by TCR and pattern recognition receptor (PRR) stimulation, along with the cytokines IL1, IL-6, IL-23, and TGFβ (272). Previous studies describing the role of IL-17 on tumor
growth have had conflicting results, suggesting both pro-tumor and anti-tumor functions
for this cytokine273. Murine γδ T cells have been identified as a major source of IL-17 in
several tumor models, which are summarized below.
In some studies, a detrimental role for γδ T cell-derived IL-17 in tumor responses
has been suggested. Specifically, expression of IL-17 by tumor-infiltrating γδ T cells in a
model of fibrosarcoma in Balb/c mice promoted tumor angiogenesis and, subsequently,
enhanced tumor growth274. Consistent with this, others have found that IL-17 enhanced
the expression of vascular endothelial growth factor (VEGF), an important growth factor
in angiogenesis275. As such, the promotion of tumor angiogenesis may be an important
and detrimental function of IL-17+ γδ T cells. Importantly, the local tumor
microenvironment was important for the expression of IL-17 by these γδ T cells, as cells
from the tumor tissue had enhanced IL-17 production compared to normal skin and cells
from the spleen and draining lymph nodes of tumor-bearing mice did not increase IL-17
production. Furthermore, IL-6, TGFβ, and IL-23 were all involved in the promotion of
127
IL-17 by these γδ T cells. Another study examining lung metastasis showed that the
expression of IL-17 enhanced metastasis and reduced survival in experiments involving
the Lewis lung carcinoma model276. In these experiments, IL-17 was primarily produced
by γδ T cells and the secretion of IL-17 by γδ T cells was induced by IL-1. Enhanced
tumor growth in the lung induced by IL-17 may have been mediated by the reduced
potential of antigen presenting cells to promote Th1 immunity. However, based upon the
study by Wakita and coworkers274, angiogenesis may also have played a role. These data
suggest that IL-17 production by γδ T cells clearly promotes tumor growth in some
settings.
However, other studies in opposition to the results described above demonstrate a
beneficial role for IL-17+ γδ T cells in the inhibition of tumor growth. In a mouse model
of bladder cancer, treatment with Mycobacterium bovis BCG enhanced IL-17 expression
by γδ T cells, which was essential for optimal neutrophil recruitment into the tumor and a
reduction in tumor growth277. In another study with a number of different tumor models,
the early infiltration of IL-17-producing γδ T cells into the tumor bed of chemotherapytreated tumors was associated with the subsequent infiltration of IFNγ-producing CD8+ T
cells and the suppression of tumor growth278. In these experiments, both IL-17 and IFNγ
were necessary for the inhibition of tumor growth. Based on these results, it has been
proposed that immunotherapy aimed at polarizing γδ T cells to express IL-17 might be
useful in enhancing the efficacy of chemotherapy240. Interestingly, in both studies where
128
anti-tumor immunity was enhanced by γδ T cell-derived IL-17, other cells played an
important role for the beneficial response. In the bladder cancer study, neutrophils were
important, whereas in the chemotherapy study, IFNγ-secreting CD8+ T cells were
important. Therefore, it is possible that in the absence of these other responses, IL-17
production by γδ T cells could lose its benefit and, therefore, enhance tumor growth as
described above. Further studies are needed to better clarify the role of γδ T cell-derived
IL-17 on tumor growth and determine if γδ T cell production of IL-17 has relevance to
human cancers.
Potentially Underappreciated Role of γδ
T Cell-derived Growth Factors in Tumor Immunity
Tumors have been described as wounds that do not heal, and numerous growth
factors, including keratinocyte growth factor (KGF), play a role in their progression279. In
addition to pro- and anti-inflammatory cytokines, γδ T cells are a source of a number of
growth factors. This has been well defined in the mouse, where skin-associated γδ T cells
are a major source of KGF and are essential for optimal wound healing151. In humans, γδ
T cells produce transcripts and/or proteins for a number of growth factors, including
KGF, IGF-1, EGF, FGF-9, ANG, PDGF, and VEGF153, 154. Furthermore, in human
peripheral Vδ2 γδ T cells, the expression of FGF-9 is enhanced by IPP153. As such, the
expression of growth factors by tumor-infiltrating γδ T cells could potentially represent a
significant response that promotes tumor growth in some settings.
129
Expression of Growth Factors in Human γδ T Cells In a study by Shilbach and
coworkers154, human Vδ2 and Vδ1 T cells were expanded and found to produce a number
of growth factors, including IGF-1, EGF, PDGF, ANG, and VEGF. When these cells
were cultured with a neuroblastoma cell line, the Vδ1 cells produced reduced amounts of
these growth factors while Vδ2 cells produced slightly increased amounts. These data
prompted the authors to suggest that Vδ1 γδ T cells may be better at promoting antitumor responses to this type of tumor, partially due to their reduced expression of growth
factors. The expression of VEGF by γδ T cells, particularly in response to a tumor cell, is
intriguing as VEGF is vital for tumor angiogenesis, growth, and metastasis280. In addition
to direct VEGF expression by γδ T cells, KGF and FGF-9 are capable of promoting
VEGF expression in other cells in a paracrine manner281, 282. Therefore, γδ T cells may
also stimulate VEGF expression indirectly by the expression of other growth factors.
These data suggest that γδ T cells may participate in the production of growth factors
within the tumor microenvironment, functions that have not yet been attributed to γδ T
cells.
A recent clinical study examining the treatment of patients with zoledronate and
IL-2 observed an increase in VEGF levels in these patients in addition to an expansion of
γδ T cells and other immune cells267, supporting the possible role for γδ T cell-derived
growth factors in human cancer. Interestingly, the increase in VEGF was more
pronounced in patients with solid tumors compared to those with leukemia. It is unknown
130
whether γδ T cells played a direct role in this increase of VEGF production. However,
these data would be consistent with the previously discussed studies demonstrating that
activated γδ T cells express VEGF, as well as factors which can indirectly promote the
expression of VEGF. Importantly, elevated VEGF levels in these patients correlated with
a lack of success of the therapy. Even if γδ T cells were not important for this enhanced
VEGF expression, it appears to be an important obstacle to overcome in optimizing γδ T
cell immunotherapy. Further studies are warranted to determine if γδ T cells are an
important source of tumor-promoting growth factors in mice or humans.
Influences on Differential Cytokine
Secretion by γδ T Cells in Tumor Studies
Differential cytokine production and behavior by γδ T cells clearly is an important
variable in mouse studies examining the role of γδ T cells in cancer, but there are
important caveats to consider in defining these roles. Differences in mouse strain, age,
and other factors (source, housing, etc.) in these studies may influence γδ T cell cytokine
secretion and subset distribution, which could influence the effect of γδ T cells on tumor
growth in these experiments. For example, a study on West Nile Virus demonstrated that
the numbers and behavior of Vγ1 and Vγ4 γδ T cells in mice could vary with age283. In
addition, epidermal γδ T cells from Balb/c mice were shown to produce less IFNγ in
response to IL-12 and IL-18 than those from C57BL/6 mice284. Therefore, in mouse
studies examining the role of γδ T cells in cancer, it is likely important to further examine
131
γδ T cell responses and subsets within the specific mice used for the study in the absence
of tumor cells, as variations in these factors would likely lead to variable tumor responses
by the γδ T cells.
Conclusions
In response to tumor cells, γδ T cells produce a variety of cytokines that both
inhibit and enhance anti-tumor immune responses, which likely accounts for some of the
conflicting reports about the role of these cells in anti-tumor immunity (Figure 5.2).
Amongst these cytokines, IFNγ, and possibly TNFα, contribute to the ability of γδ T cells
to inhibit tumor growth. In contrast, the expression of IL-4, IL-10, TGFβ, other unknown
factors, and possibly growth factors, by γδ T cells suppress anti-tumor immunity and
enhance tumor growth. The expression of IL-17 by γδ T cells appears to have conflicting
effects on tumor growth, which could be dependent upon the type of cancer or other
factors, such as tumor-infiltration of other cell types or the use of chemotherapy. The
expression of these cytokines by γδ T cells influences downstream adaptive immune
responses to tumors, consistent with the described ability of γδ T cells to link innate and
adaptive immunity285. γδ T cell-derived IFNγ and IL-17 enhance CD8+ T cell responses,
while IL-10, TGFβ, and other γδ T cell-derived soluble factors inhibit them. Therefore, in
addition to their lytic activity, several studies suggest that the influence of γδ T cells on
132
adaptive immune responses to tumors is an important part of their role in anti-tumor
immunity.
Differential cytokine production by γδ T cells may also be important in γδ T cell
immunotherapy. Stimulation of γδ T cells with synthetic phosphoantigens or
bisphosphonates may only enhance γδ T cell-responses already influenced by the tumor
environment, beneficial or not, which could account for the variable effectiveness of
these therapies. Therefore, the identification of therapeutic options which enhance and
favor the production of beneficial anti-tumor cytokines and soluble factors by γδ T cells,
while minimizing or removing detrimental factors, may be key to unlocking the
maximum potential of γδ T cell immunotherapy. A good example of this concept can be
found in the study by Peng and coworkers237, where they were able to reverse the
immunosuppressive phenotype of tumor-infiltrating γδ T cells by stimulating them with a
TLR8 agonist. Other options may include the use of additional cytokines to further
enhance the anti-tumor activity of γδ T cells. For example, the addition of IL-18 to
zoledronate and IL-2 enhances IFNγ and TNFα expression by γδ T cells compared to
zoledronate and IL-2 alone286. The use of anti-VEGF and other anti-angiogenesis
therapies may inhibit any pro-angiogenesis responses induced by γδ T cells or γδ T cell
immunotherapy. Furthermore, chemotherapy might also have the potential to enhance the
effectiveness of γδ T cell immunotherapy, as discussed by Hannani and coworkers240. In
conclusion, in order to better understand the complex role of γδ T cells in cancer and
133
improve the effectiveness of γδ T cell immunotherapy, additional studies are needed that
examine the cytokine profiles of γδ T cells in response to tumors and immunotherapy, as
well as identify ways to best manipulate this profile for the benefit of the patient.
Figure 5.2 Summary of the influence of γδ T cell-derived cytokines and growth factors on
tumor growth.
134
Acknowledgements
Our studies are supported with grants from the National Institutes of Health (NIH)
(NCCAM AT0004986-01), NIH COBRE (P20 RR020185), M.J. Murdock Charitable
Trust, and The Montana State University Agricultural Experiment Station. We would like
to thank Dana Doney, Amanda Robison, and Dr. Jeff Holderness for critical review of the
manuscript.
Author Disclosure Statement
No competing financial interests exist.
135
CHAPTER SIX
CONCLUSIONS
Supplements containing plant polyphenols are consumed by many people to
improve health, with various benefits attributed to these compounds. It is important that
the proposed benefits of these molecules be studied for proof of benefit, as well as for
optimization of that benefit. Therefore, many studies have examined the effects of
polyphenols in more detail, but there is still much that is currently unknown.
In this dissertation, the immunomodulatory polyphenol oenothein B was found to
stimulate lymphocytes, including NK cells, γδ T cells, and αβ T cells, inducing the
expression of IL-2Rα, CD69, and IFNγ (Chapter 3). Oenothein B has been described to
have both pro- and anti-inflammatory effects on macrophages and neutrophils, but this is
the first study to examine its influence on lymphocytes. Interestingly, oenothein B primed
NK cells from both cattle and humans, in both mixed PBMC and purified cell cultures, to
produce enhanced IFNγ in response to a cytokine, IL-18, or a tumor target cell. This
priming effect on bovine and human NK cells by oenothein B does not appear to be a
common property of polyphenols, as similar doses of several other commonly studied
polyphenols did not affect IFNγ production by bovine NK cells in response to IL-18
(Chapter 3).
136
In addition to NK cells, T cells, both αβ and γδ, produced IFNγ in response to
stimulation with oenothein B (Chapters 3 and 4). Aging impacted oenothein B-induced
IFNγ production by T cells, as αβ and γδ T cells from bovine calves produced little IFNγ
in response to oenothein B, but αβ and γδ T cells from bovine adults produced
significantly more IFNγ than calf T cells upon treatment with oenothein B (Chapter 4,
Table 6.1). Similar results were seen in human PBMCs, as adult αβ and γδ T cells, but
not cord blood cells, produced IFNγ, as well as GM-CSF. In both cows and humans, the
CD45RO+ T cell population produced more cytokines in response to oenothein B than
CD45RO- T cells. This increase in cytokine production does not appear to be simply
related to oenothein B’s ability to stimulate these cells, as oenothein B upregulated IL2Rα and CD69 expression on T cells from both young and old individuals, as well as
both CD45RO+ and CD45RO- T cells (Chapter 4). These results could have important
implications for in vivo murine studies, as laboratory mice are often kept in specificpathogen free conditions and used at a young age, resulting in a relatively low number of
CD45RO+ memory T cells. If these cells are major cytokine-producing cells in response
to oenothein B in adults, this could potentially lead to differential effects on adult
immunity by oenothein B in mice compared to humans and cattle.
Like T cells, NK cell responses to oenothein B may also be impacted by aging, as
IFNγ production by NK cells in response to oenothein B appeared to be reduced in
bovine adults (Chapter 4, Table 6.1). IFNγ production in response to oenothein B was
137
Table 6.1 Summary of oenothein B-induced IFNγ production by lymphocytes
Induction of IFNγ Production by Oenothein B
Cattle
NK
αβ T
cells
cells
Adult
Calves
Adult
Calves
↑IFNγ
+
++
++
−
Human
NK
αβ T
cells
cells
↑IFNγ
Adult
+
Cord Blood Adult
−
++
γδ T
cells
Adult
++
Calves
−
γδ T
cells
Cord Blood Adult
−
++
Cord
Blood
−
dominated by NK cells in bovine calves, but was dominated by T cells in bovine and
human adults. Interestingly, these results are similar to what has been observed for IFNγ
responses in response to BCG vaccination in humans. Altogether, these data suggest that
a polyphenol's ability to enhance innate immunity and protection from disease could be
different for children and adults. For NK cell-mediated protection, oenothein B may be
more effective in younger individuals, while for T cell-mediated protection, especially if
cytokine production is important, oenothein B may be more effective in adults.
The production of IFNγ by NK cells and T cells is known to be important in these
cells' ability to limit tumor growth, through the enhancement of cellular cytotoxicity, the
reduction of tumor angiogenesis, and other mechanisms (Chapter 5). Therefore, these
data suggest a new potential mechanism of action for the proposed antitumor effects of
oenothein B. They also suggest a potential benefit for oenothein B to counter infectious
138
disease, as IFNγ production by NK cells and T cells are critical for the clearance of
numerous pathogens (Chapter 2).
In addition to age and the presence of CD45RO+ T cells, the phenotypic state of
γδ T cells may also influence the effects of oenothein B on innate immunity in vivo.
Oenothein B may have been effective in our early B16 melanoma experiments because γδ
T cells were of a more regulatory phenotype, as indicated by reduced tumor growth in
saline-treated γδ-/- mice. However, when γδ T cells were of a more inflammatory
phenotype in later studies, as indicated by enhanced tumor growth in γδ-/- mice, this
benefit may have been lost (Chapter 5). It is possible that oenothein B has a similar
activity as TLR8 agonists237, which can convert regulatory γδ T cells to more
inflammatory cells. This hypothesis would be consistent with the loss of benefit in γδ-/mice, as well as in later studies when γδ T cells did not need to be converted to an
inflammatory phenotype.
To our knowledge, the influence of age on the immunomodulatory effects of
polyphenols had not been previously studied, especially comparing children to adults.
While these types of studies may be difficult to perform in humans (ethical reasons), as
well as mice (difficult to use when very young), our data suggest that domestic cattle may
be a very useful model in studying the influence of age on the enhancement of immunity
by dietary polyphenols. These data has implications for optimizing the use of dietary
polyphenols in both humans and animals and suggest that subject age should be taken
139
into account when examining the effect of consumption of polyphenol-rich supplements
on immunity.
140
CHAPTER SEVEN
FUTURE STUDIES
While these data provide some insight into the effects of oenothein B on innate
lymphocytes and the influence of age on these effects, there is still much that needs to be
explored. For example, it is currently unknown why CD45RO+ T cells and T cells from
adults produce more IFNγ and GM-CSF in response to oenothein B. As discussed in
chapter 4, one possible explanation could involve methylation at cytokine promoter sites,
as ifnγ promoter sites in adult and memory T cells have been shown to have reduced
methylation compared to young and naïve T cells. As methylation at promoter sites is
known to inhibit gene expression, T cells with reduced methylation at the promoter sites
may be more capable of producing IFNγ and GM-CSF in response to oenothein B. This
could be tested by the treatment of T cells from young individuals with 5-azacytidine, as
5-azacytidine is incorporated into the DNA of proliferating cells and prevents
methylation. Therefore, co-treatment with 5-azacytidine could potentially enhance IFNγ
and GM-CSF production by T cells which normally do not produce these cytokines in
response to oenothein B. Preliminary experiments have currently not shown enhanced
IFNγ production by bovine calf T cells treated with 5-azacytidine in response to
oenothein B (Figure 7.1). Therefore, it is possible that reduced methylation is not
sufficient to explain the enhanced IFNγ production by adult T cells.
141
Figure 7.1 5-Azacytidine does not enhance IFNγ production by oenothein B-stimulated
bovine calves. αβ T cells (CD3+ γδ TCR-) (A) and γδ T cells (CD3+ γδ TCR+) (B) were
FACS sorted from bovine calves (N=2), plated (4 X 104 cells/well), and treated with
indicated concentrations of oenothein B or X-VIVO medium alone, with or without the
indicated concentrations of 5-Azacytidine. After 72 hrs, soluble IFNγ were measured by
ELISA. The graphs represent pooled data. Error bars indicate SEM. Each sample was
analyzed in duplicate.
142
However, additional experiments and experimental controls are needed to confirm these
preliminary results.
In addition, studies in this dissertation primarily focus on αβ T cells as a single
population, and do not examine potential differences between the CD4+ and CD8+ T cell
subpopulations in detail. Furthermore, the relative contribution of these T cell subsets, as
well as γδ T cells, to the overall production of IFNγ and GM-CSF by adult PBMCs
should be examined. This could be done by depleting CD4+, CD8+, or γδ T cells from
adult human and bovine PBMCs and comparing oenothein B-induced cytokine
production by PBMCs lacking these populations to whole PBMCs by ELISA. These
experiments could then determine which T cell subset was the most important for
oenothein B-induced cytokine production in adult PBMCs. It is likely that different
subsets of T cells will have different contributions in humans and cattle, as these species
have different compositions of these subsets.
Data in chapter 3 suggested that the production of IFNγ by bovine calf NK cells
could be enhanced by the presence of other mononuclear cells which did not produce
significant IFNγ themselves, such as monocytes and, possibly, γδ T cells. Another study
also demonstrated an important role for antigen presenting cells for cytokine production
by T cells in response to polymerized polyphenols180. Therefore, although purified NK
cells and T cells were shown in these studies to produce IFNγ in response to oenothein B
in the absence of other cells, the potential for other mononuclear cells to enhance
143
cytokine production by NK cells and T cells should be examined. This could be
examined by adding different FACS sorted mononuclear cell subsets to sorted NK cells
and T cells, looking for synergy in oenothein B-induced IFNγ production between
different cells. Furthermore, the importance of cell-to-cell contact or the secretion of
soluble mediators for this synergy could then be tested with transwell experiments.
In chapter 3, it was shown that oenothein B enhanced IFNγ production by human
NK cells in response to a tumor cell line, the K562 cell line. It was also hypothesized that
oenothein B could enhance tumor cell killing by NK cells. However, preliminary
experiments with K562 cells using the CD107a mobilization assay were inconclusive,
with some doses of oenothein B reducing CD107a mobilization on NK cells and another
enhancing it (Figure 7.2). As a result, it was decided to focus on the IFNγ response for
these studies. However, it is still possible that oenothein B enhances NK cell killing of
tumor cells. It is possible that K562 was not the ideal cell type to perform these
experiments due to its high susceptibility to NK cell killing, with more than 60% of NK
cells expressing CD107a on their surface in response to K562 cells in the absence of
oenothein B. Therefore, this experiment should be repeated using a broad range of tumor
cell lines with varying susceptibility to NK cell killing to determine the effect of
stimulation with oenothein B on NK cell-mediated killing of tumor cells.
144
% CD107a +
Human NK cells
90
80
70
60
50
40
30
20
10
0
***
***
*** **
*** ***
0
10
20
***
no K562
K562
40
μg/ml oenothein B
Figure 7.2 Influence of oenothein B on NK cell degranulation in response to a tumor cell
line. Human PBMCs (N=1) (105 cells/well) were treated with oenothein B or cRPMI
medium alone for 48 hrs. Cells were then washed and co-cultured for 4 hrs with or
without K562 cells at a 1:1 effector:target (E:T) ratio in cRPMI. CD107a expression on
the surface of NK cells (CD3- CD56+) was then determined by flow cytometry. Each
sample was analyzed in triplicate. Statistical significance was determined by Two-way
ANOVA with Bonferroni post-test. **p<0.01 ***p<0.001
Finally, there is currently no evidence to suggest that oenothein B enhances IFNγ
production in vivo, as it does in vitro. However, preliminary experiments suggest that
treatment with oenothein B can limit B16 melanoma growth, as well as reduce morbidity
to influenza A infection (Chapter 2). Future experiments are needed to first confirm
oenothein B’s ability to counter tumor growth and infectious disease in vivo. Experiments
should then determine if NK cells and T cells, as well as IFNγ, are critical for oenothein
B's ability to counter tumor growth and infectious disease. One study found that
145
treatment with Acai berry polysaccharides enhanced protection to pulmonary infection by
intracellular bacteria and that this enhanced protection was dependent upon the enhanced
production of IFNγ by NK cells and γδ T cells6. It is possible that oenothein B has similar
activity. In addition, these studies should also determine if oenothein B’s ability to
counter tumor growth and infectious disease in vivo is impacted by age or the presence of
CD45RO+ T cells, as suggested by these in vitro studies.
However, it is also possible that the mouse may not be the ideal model to test the
in vivo activity of oenothein B. Unlike human and bovine PBMCs, oenothein B induced
little IFNγ production by murine splenocytes in vitro, either alone or in combination with
IL-18 (Figure 7.3). In contrast, it appeared to inhibit IL-18-induced IFNγ production.
Therefore, it is possible that oenothein B does not have the same immunomodulatory
properties with mice as it does with humans and cattle. Therefore, as described
previously, domestic cattle may be a better in vivo animal model. Cattle have already
been shown to be an effective model for testing the in vivo immunomodulatory effects of
polyphenols, as feeding bovine calves polyphenols from pomegranate was found to
enhance mitogen-induced IFNγ production by PBMCs30. Similar experiments could be
performed which administer oenothein B, or oenothein B-containing plants, to calves
and/or adult cows. PBMC responses to IL-18, tumor cells, and other agonists could then
be compared between oenothein B-treated and untreated animals.
146
Figure 7.3 Oenothein B does not prime murine splenocytes to produce IFNγ in response
to IL-18. Spleens from 6 week-old (A) or 13 week-old (B) male human TL4 mice were
homogenized and mononuclear cells were isolated by Lympholyte M gradient.
Splenocytes (105 cells/well) were then treated with indicated concentrations of oenothein
B or X-VIVO medium only. After 24 hrs, cells were washed and treated with indicated
concentrations of recombinant mouse IL-18 or X-VIVO medium only. After 24 hrs,
soluble IFNγ levels in culture supernatant fluids were determined by ELISA. The graph
represents results from one experiment. Error bars indicate SEM. Each sample was
analyzed in triplicate. Significance compared to 0 µg/ml oenothein B samples was
determined by Two-way ANOVA with Bonferroni post-test. **p<0.01, ***p<0.001
147
REFERENCES CITED
1. André, FE. How the research-based industry approaches vaccine development and
establishes priorities. Dev Biol (Basel). 110, 25-29 (2002).
2. Pronker, E.S., Weenen, T.C., Commandeur, H., Claassen, E.H. & Osterhaus A.D.
Risk in vaccine research and development quantified. PLoS One 8(3), e57755
(2013).
3. Koel, B.F. et al. Substitutions near the receptor binding site determine major
antigenic change during influenza virus evolution. Science 342(6161), 976-979
(2013).
4. Lacombe, K. et al. High incidence of treatment-induced and vaccine-escape
hepatitis B virus mutants among human immunodeficiency virus/hepatitis Binfected patients. Hepatology 58(3):912-922 (2013).
5. Amador-Molina, A., Hernández-Valencia, J.F., Lamoyi, E., Contreras-Paredes, A.
& Lizano, M. Role of innate immunity against human papillomavirus (HPV)
infections and effect of adjuvants in promoting specific immune response. Viruses
5(11), 2624-2642 (2013).
6. Skyberg, J.A. et al. Nasal Acai polysaccharides potentiate innate immunity to
protect against pulmonary Francisella tularensis and Burkholderia pseudomallei
Infections. PLoS Pathog. 8(3), e1002587 (2012).
7. Finlay B.B. & Hancock, R.E. Can innate immunity be enhanced to treat microbial
infections? Nat Rev Microbiol. 2(6), 497-504 (2004).
8.
Orange, J.S. Natural killer cell deficiency. J Allergy Clin Immunol. 132(3), 515525 (2013).
9. Hambleton, S. et al. STAT2 deficiency and susceptibility to viral illness in
humans. Proc Natl Acad Sci U S A. 110(8), 3053-3058 (2013).
10. Béla, S.R. et al. Impaired innate immunity in mice deficient in interleukin-1
receptor-associated kinase 4 leads to defective type 1 T cell responses, B cell
expansion, and enhanced susceptibility to infection with Toxoplasma gondii.
Infect Immun. 80(12), 4298-4308 (2012).
148
11. Tang, D., Kang, R., Coyne, C.B., Zeh, H.J. & Lotze, M.T. PAMPs and DAMPs:
signal 0s that spur autophagy and immunity. Immunol Rev. 249(1), 158-175
(2012).
12. Kovach, M.A. & Standiford, T.J. Toll like receptors in diseases of the lung. Int
Immunopharmacol. 11(10),1399-1406 (2011).
13. Clinthorne, J.F., Beli, E., Duriancik, D.M. & Gardner, E.M. NK cell maturation
and function in C57BL/6 mice are altered by caloric restrition. J Immunol. 190(2),
712-722 (2013).
14. Maijó, M., Clements, S.J., Ivory, K., Nicoletti, C. & Carding, S.R. Nutrition, diet
and immunosenescence. Mech Ageing Dev. pii: S0047-6374(13)00133-4 (2013).
15. Holderness, J. et al. Response of gammadelta T Cells to plant-derived tannins.
Crit Rev Immunol. 28(5), 377-402 (2008).
16. Akiyama,H. et al. Dietary unripe apple polyphenol inhibits the development of
food allergies in murine models. FEBS Lett.579, 4485-4491 (2005).
17. Ramiro-Puig,E. et al. Intestinal immune system of young rats influenced by
cocoa-enriched diet. J Nutr. Biochem.19, 555-565 (2008).
18. Koláček, M. et al. Effect of natural polyphenols (Pycnogenol) on oxidative stress
markers in children suffering from Crohn's disease--a pilot study. Free Radic Res.
47(8), 624-634 (2013).
19. Skyberg, J. et al. Apple polyphenols require T cells to ameliorate dextran sulfate
sodium-induced colitis and dampen proinflammatory cytokine expression. J
Leukoc Biol. 90(6),1043-1054 (2011).
20. Snyder, D.T. et al. Oral delivery of oligomeric procyanidins in Apple Poly(R)
enhances type I IFN responses in vivo. J Leukoc Biol. In press (2014).
21. Tresserra-Rimbau, A. et al. Inverse association between habitual polyphenol
intake and incidence of cardiovascular events in the PREDIMED study. Nutr
Metab Cardiovasc Dis. pii: S0939-4753(14)00008-8 (2014).
22. Manganaris, G.A., Goulas, V., Vicente, A.R. & Terry, L.A. Berry antioxidants:
small fruits providing large benefits. J Sci Food Agric. In press (2013).
149
23. Rubio,V., Valverde,M. & Rojas,E. Effects of atmospheric pollutants on the Nrf2
survival pathway. Environ. Sci. Pollut. Res. Int.17, 369-382 (2010).
24. Iborra,M. et al. Role of oxidative stress and antioxidant enzymes in Crohn's
disease. Biochem. Soc. Trans.39, 1102-1106 (2011).
25. Okuyama, S. et al. Oenothein B suppresses lipopolysaccharide (LPS)-induced
inflammation in the mouse brain. 14(5), 9767-9778 (2013).
26. Jean-Gilles, D. et al. Inhibitory effects of polyphenol punicalagin on type-II
collagen degradation in vitro and inflammation in vivo. Chem Biol Interact.
205(2), 90-99 (2013).
27. Nantz, M.P. et al. Consumption of cranberry polyphenols enhances human γδ-T
cell proliferation and reduces the number of symptoms associated with colds and
influenza: a randomized, placebo-controlled intervention study. Nutr J. 12, 161
(2013).
28. Miyamoto, K. et al. Antitumor activity of oenothein B, a unique macrocyclic
ellagitannin. Jpn J Cancer Res. 84(1), 99-103 (1993).
29. Miyamoto, K. et al. Antitumor activity and interleukin-1 induction by tannins.
Anticancer Res. 13(1), 37-42 (1993).
30. Oliveira, R.A. et al. Effects of feeding polyphenols from pomegranate extract on
health, growth, nutrient digestion, and immunocompetence of calves. J Dairy Sci.
93(9), 4280-4291 (2010).
31. Holderness, J. et al. Select plant tannins induce IL-2Rαlpha up-regulation and
augment cell division in gammadelta T cells. J Immunol. 179(10), 6468-6478
(2007).
32. Daughenbaugh, K.F. et al. Contribution of transcript stability to a conserved
procyanidin-induced cytokine response in γδ T cells. Genes Immun. 12(5), 378389 (2011).
33. Schepetkin, I.A. et al. Immunomodulatory activity of oenothein B isolated from
Epilobium angustifolium. J Immunol. 183(10), 6754-6766 (2009).
34. Cerutti, A., Cols, M. & Puga, I. Marginal zone B cells: virtues of innate-like
antibody-producing lymphocytes. Nat Rev Immunol. 13(2), 118-132 (2013).
150
35. Baumgarth, N. Innate-like B cells and their rules of engagement. Adv Exp Med
Biol. 785, 57-66 (2013).
36. Hwang, Y.Y. & McKenzie, A.N. Innate lymphoid cells in immunity and disease.
Adv Exp Med Biol. 785, 9-26 (2013).
37. Spits, H. et al. Innate lymphoid cells--a proposal for uniform nomenclature. Nat
Rev Immunol. 13(2),145-149 (2013).
38. Walker, J.A., Barlow, J.L. & McKenzie, A.N. Innate lymphoid cells--how did we
miss them? Nat Rev Immunol. 13(2), 75-87 (2013).
39. Tait Wojno, E.D. & Artis, D. Innate lymphoid cells: balancing immunity,
inflammation, and tissue repair in the intestine. Cell Host Microbe. 12(4), 445457 (2012).
40. Spits, H. & Di Santo, J.P. The expanding family of innate lymphoid cells:
regulators and effectors of immunity and tissue remodeling. Nat Immunol. 12(1),
21-27 (2011).
41. Robertson, M.J. & Ritz, J. Biology and clinical relevance of human natural killer cells.
Blood 76(12), 2421-2438 (1990).
42. Fehniger, T.A. et al. CD56bright natural killer cells are present in human lymph
nodes and are activated by T cell-derived IL-2: a potential new link between
adaptive and innate immunity. Blood 101(8), 3052-3057 (2003).
43. Campbell, K.S. & Hasegawa, J. Natural killer cell biology: an update and future
directions. J Allergy Clin Immunol. 132(3), 536-544 (2013).
44. Kiessling, R., Klein, E. & Wigzell, H. "Natural" killer cells in the mouse. I.
Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity
and distribution according to genotype. Eur J Immunol. 5(2),112-117 (1975).
45. Wu, J. & Lanier L.L. Natural killer cells and cancer. Adv Cancer Res. 90,127-156
(2003).
46. Vankayalapati, R. et al. The NKp46 receptor contributes to NK cell lysis of
mononuclear phagocytes infected with an intracellular bacterium. J Immunol.
168(7), 3451-3457 (2002).
151
47. Hadad, U., Martinez, O. & Krams, S.M. NK cells after transplantation: friend or
foe. Immunol Res. In Press (2014).
48. Strayer, D.R., Carter, W.A. & Brodsky, I. Familial occurrence of breast cancer is
associated with reduced natural killer cytotoxicity. Breast Cancer Res Treat.
7(3),187-192 (1986).
49. Hsia, J.Y. et al. Prognostic significance of intratumoral natural killer cells in
primary resected esophageal squamous cell carcinoma. Chang Gung Med J.
28(5):335-340 (2005).
50. Pegram, H.J., Andrews, D.M., Smyth, M.J., Darcy, P.K. & Kershaw, M.H.
Activating and inhibitory receptors of natural killer cells. Immunol Cell Biol.
89(2), 216-224 (2011).
51. Jost, S. & Altfeld, M. Control of human viral infections by natural killer cells.
Annu Rev Immunol. 31, 163-194 (2013).
52. van der Touw, W. & Bromberg J.S. Natural killer cells and the immune response
in solid organ transplantation. Am J Transplant. 10(6), 1354-1358 (2010).
53. Wang, Y. et al. High expression of NKG2A/CD94 and low expression of
granzyme B are associated with reduced cord blood NK cell activity. Cell Mol
Immunol. 4(5), 377-382 (2007).
54. Bae, D.S., Hwang, Y.K. & Lee, J.K. Importance of NKG2D-NKG2D ligands
interaction for cytolytic activity of natural killer cell. Cell Immunol. 276(12),122-127 (2012).
55. Raulet, D.H., Gasser, S., Gowen, B.G., Deng, W. & Jung, H. Regulation of
ligands for the NKG2D activating receptor. Annu Rev Immunol. 31, 413-441
(2013).
56. Zamai, L. et al. Natural killer (NK) cell-mediated cytotoxicity: differential use of
TRAIL and Fas ligand by immature and mature primary human NK cells. J Exp
Med. 188(12), 2375-2380 (1998).
57. Scharton-Kersten, T.M. & Sher, A. Role of natural killer cells in innate resistance to
protozoan infections. Curr Opin Immunol. 9(1), 44-51 (1997).
58. Ma, L.L. et al. NK cells use perforin rather than granulysin for anticryptococcal
activity. J Immunol. 173(5), 3357-3365 (2004).
152
59. Biron, C.A., Nguyen, K.B., Pien, G.C., Cousens, L.P. & Salazar-Mather, T.P.
Natural killer cells in antiviral defense: function and regulation by innate
cytokines. Annu Rev Immunol. 17, 189-220 (1999).
60. Culley, F.J. Natural killer cells in infection and inflammation of the lung.
Immunology. 128(2), 151-163 (2009).
61. Wallin, R.P. et al. Angiostatic Effects of NK Cell-Derived IFN-γ Counteracted by
Tumour Cell Bcl-xL Expression. Scand J Immunol. 79(2), 90-97 (2014).
62. Smyth, M.J., Hayakawa, Y., Takeda, K. & Yagita, H. New aspects of naturalkiller-cell surveillance and therapy of cancer. Nat Rev Cancer. 2(11):850-861
(2002).
63. Le Garff-Tavernier, M. et al. Human NK cells display major phenotypic and
functional changes over the life span. Aging Cell. 9(4):527-535 (2010).
64. Nomura, A. et al. Functional analyses of cord blood natural killer cells and T
cells: a distinctive interleukin-18 response. Exp Hematol. 29(10), 1169-1176
(2001).
65. Eljaafari, F.M. et al. Potent induction of IFN-γ production from cord blood NK
cells by the stimulation with single-stranded RNA. J Clin Immunol. 31(4):728735 (2011).
66. Krampera, M., Tavecchia, L., Benedetti, F., Nadali, G. & Pizzolo, G. Intracellular
cytokine profile of cord blood T-, and NK- cells and monocytes. Haematologica.
85(7), 675-679 (2000).
67. Zufferey, C., Germano, S., Dutta, B., Ritz, N. & Curtis, N. The contribution of
non-conventional T cells and NK cells in the mycobacterial-specific IFNγ
response in Bacille Calmette-Guérin (BCG)- immunized infants. PLoS One.
8(10), e77334 (2013).
68. Camous, X., Pera, A., Solana, R. & Larbi, A. NK cells in healthy aging and ageassociated diseases. J Biomed Biotechnol. 2012, 195956 (2012).
69. Page, G.G., Ben-Eliyahu, S. & Taylor, A.N. The development of sexual
dimorphism in natural killer cell activity and resistance to tumor metastasis in the
Fischer 344 rat. J Neuroimmunol. 63(1), 69-77 (1995).
153
70. Guilmot, A., Hermann, E., Braud, V.M., Carlier, Y. & Truyens, C. Natural killer
cell responses to infections in early life. J Innate Immun. 3(3), 280-288 (2011).
71. Wang, Y. et al. High expression of NKG2A/CD94 and low expression of
granzyme B are associated with reduced cord blood NK cell activity. Cell Mol
Immunol. 4(5), 377-382 (2007).
72. Chiu, B.C., Martin, B.E., Stolberg, V.R. & Chensue, S.W. The host environment
is responsible for aging-related functional NK cell deficiency. J Immunol. 191(9),
4688-4698 (2013).
73. Boysen, P. & Storset, A.K. Bovine natural killer cells. Vet Immunol
Immunopathol. 130(3-4),163-177 (2009).
74. Storset, A.K. et al. NKp46 defines a subset of bovine leukocytes with natural
killer cell characteristics. Eur J Immunol. 34(3), 669-676 (2004).
75. Johnson, W.C., Bastos, R.G., Davis, W.C. & Goff, W.L. Bovine WC1(-)
gammadeltaT cells incubated with IL-15 express the natural cytotoxicity receptor
CD335 (NKp46) and produce IFN-gamma in response to exogenous IL-12 and
IL-18. Dev Comp Immunol. 32(8), 1002-1010 (2008).
76. Boysen, P., Gunnes, G., Pende, D., Valheim, M. & Storset, A.K. Natural killer
cells in lymph nodes of healthy calves express CD16 and show both cytotoxic and
cytokine-producing properties. Dev Comp Immunol. 32(7), 773-783 (2008).
77. Ferlazzo, G. et al. The abundant NK cells in human secondary lymphoid tissues
require activation to express killer cell Ig-like receptors and become cytolytic. J
Immunol. 172(3), 1455-1462 (2004).
78. Bastos, R.G., Johnson, W.C., Mwangi, W., Brown, W.C. & Goff, W.L. Bovine
NK cells acquire cytotoxic activity and produce IFN-gamma after stimulation by
Mycobacterium bovis BCG - or Babesia bovis-exposed splenic dendritic cells.
Vet Immunol Immunopathol. 124(3-4), 302-312 (2008).
79. Olsen, I. et al. Bovine NK cells can produce gamma interferon in response to the
secreted mycobacterial proteins ESAT-6 and MPP14 but not in response to
MPB70. Infect Immun. 73(9), 5628-5635 (2005).
80. Denis, M., Keen, D.L., Parlane, N.A., Storset, A.K. & Buddle, B.M. Bovine
natural killer cells restrict the replication of Mycobacterium bovis in bovine
154
macrophages and enhance IL-12 release by infected macrophages. Tuberculosis
(Edinb). 87(1), 53-62 (2007).
81. Boysen, P., Klevar, S., Olsen, I. & Storset, A.K. The protozoan Neospora
caninum directly triggers bovine NK cells to produce gamma interferon and to kill
infected fibroblasts. Infect Immun. 74(2), 953-960 (2006).
82. Goff, W.L., Storset, A.K., Johnson, W.C. & Brown, W.C. Bovine splenic NK
cells synthesize IFN-gamma in response to IL-12-containing supernatants from
Babesia bovis-exposed monocyte cultures. Parasite Immunol. 28(5), 221-228
(2006).
83. Dobromylskyj, M. & Ellis, S. Complexity in cattle KIR genes: transcription and
genome analysis. Immunogenetics. 59(6), 463-72 (2007).
84. Birch, J. & Ellis, S.A. Complexity in the cattle CD94/NKG2 gene families.
Immunogenetics. 59(4), 273-280 (2007).
85. Guzman, E., Birch, J.R. & Ellis, S.A. Cattle MIC is a ligand for the activating NK
cell receptor NKG2D. Vet Immunol Immunopathol. 136(3-4), 227-234 (2010).
86. Graham, E.M. et al. Natural killer cell number and phenotype in bovine peripheral
blood is influenced by age. Vet Immunol Immunopathol. 132(2-4), 101-108
(2009).
87. Kampen, A.H., Olsen, I., Tollersrud, T., Storset, A.K. & Lund, A. Lymphocyte
subpopulations and neutrophil function in calves during the first 6 months of life.
Vet Immunol Immunopathol. 113(1-2), 53-63 (2006).
88. Fries, P.N., Popowych, Y.I., Guan, L.L. & Griebel, P.J. Age-related changes in
the distribution and frequency of myeloid and T cell populations in the small
intestine of calves. Cell Immunol. 271(2), 428-437 (2011).
89. Elhmouzi-Younes, J., Storset, A.K., Boysen, P., Laurent, F. & Drouet, F. Bovine
neonate natural killer cells are fully functional and highly responsive to
interleukin-15 and to NKp46 receptor stimulation. Vet Res. 40(6), 54 (2009).
90. Hope, J.C., Sopp, P. & Howard, C.J. NK-like CD8(+) cells in immunologically
naïve neonatal calves that respond to dendritic cells infected with Mycobacterium
bovis BCG. J Leukoc Biol. 71(2), 184-194 (2002).
155
91. Brennan, P.J., Brigl, M. & Brenner, M.B. Invariant natural killer T cells: an innate
activation scheme linked to diverse effector functions. Nat Rev Immunol. 13(2),
101-117 (2013).
92. Williams, M.A. & Bevan, M.J. Effector and memory CTL differentiation. Annu
Rev Immunol. 25, 171-192 (2007).
93. Zhu, J. & Paul, W.E. CD4 T cells: fates, functions, and faults. Blood.
112(5),1557-1569 (2008).
94. Mueller, S.N., Gebhardt, T., Carbone, F.R. & Heath, W.R. Memory T cell
subsets, migration patterns, and tissue residence. Annu Rev Immunol. 31, 137161 (2013).
95. Sallusto, F., Lenig, D., Förster, R., Lipp, M. & Lanzavecchia, A. Two subsets of
memory T lymphocytes with distinct homing potentials and effector functions.
Nature. 401(6754), 708-712 (1999).
96. Masopust, D., Vezys, V., Marzo, A.L. & Lefrançois, L. Preferential localization
of effector memory cells in nonlymphoid tissue. Science. 291(5512), 2413-2417
(2001).
97. Skon, C.N. et al. Transcriptional downregulation of S1pr1 is required for the
establishment of resident memory CD8+ T cells. Nat Immunol. 14(12), 12851293 (2013).
98. Doisne, J.M. et al. CD8+ T cells specific for EBV, cytomegalovirus, and
influenza virus are activated during primary HIV infection. J Immunol. 173(4),
2410-2418 (2004).
99. Berg, R.E., Crossley, E., Murray, S. & Forman, J. Memory CD8+ T cells provide
innate immune protection against Listeria monocytogenes in the absence of
cognate antigen. J Exp Med. 198(10), 1583-1593 (2003).
100. Soudja, S.M., Ruiz, A.L., Marie, J.C. & Lauvau, G. Inflammatory monocytes
activate memory CD8(+) T and innate NK lymphocytes independent of cognate
antigen during microbial pathogen invasion. Immunity. 37(3), 549-562 (2012).
101. Chu, T. et al. Bystander-activated memory CD8 T cells control early pathogen
load in an innate-like, NKG2D-dependent manner. Cell Rep. 3(3), 701-708
(2013).
156
102.
Sckisel, G.D. et al. Influenza infection results in local expansion of memory
CD8(+) T cells with antigen non-specific phenotype and function. Clin Exp
Immunol. 175(1), 79-91 (2014).
103.
Farber, D.L., Yudanin, N.A. & Restifo, N.P. Human memory T cells: generation,
compartmentalization and homeostasis. Nat Rev Immunol. 14(1), 24-35 (2014).
104.
Falcão, R.P. & De-Santis, G.C. Age-associated changes of memory (CD45RO+)
and naive (CD45R+) T cells. Braz J Med Biol Res. 24(3), 275-279 (1991).
105.
Guilmot, A., Carlier, Y. & Truyens, C. Differential IFN-γ production by adult and
neonatal blood CD56+ natural killer (NK) and NK-like-T cells in response to
Trypanosoma cruzi and IL-15. Parasite Immunol. 36(1), 43-52 (2014).
106.
Timm, J.A. & Thoman, M.L. Maturation of CD4+ lymphocytes in the aged
microenvironment results in a memory-enriched population. J Immunol. 162(2),
711-717 (1999).
107.
Chiu, B.C., Martin, B.E., Stolberg, V.R. & Chensue, S.W. Cutting edge: Central
memory CD8 T cells in aged mice are virtual memory cells. J Immunol. 191(12),
5793-5796 (2013).
108.
Sosinowski, T. et al. CD8α+ dendritic cell trans presentation of IL-15 to naive
CD8+ T cells produces antigen-inexperienced T cells in the periphery with
memory phenotype and function. J Immunol. 190(5), 1936-1947 (2013).
109.
Chiu, W.K., Fann, M. & Weng, N.P. Generation and growth of CD28nullCD8+
memory T cells mediated by IL-15 and its induced cytokines. J Immunol.
177(11), 7802-7810 (2006).
110.
Nishimura, H. et al. Intraepithelial gammadelta T cells may bridge a gap between
innate immunity and acquired immunity to herpes simplex virus type 2. J Virol.
78(9), 4927-4930 (2004).
111.
Holderness, J., Hedges, J.F., Ramstead, A. & Jutila, M.A. Comparative Biology
of γδ T Cell Function in Humans, Mice, and Domestic Animals. Annual Review
of Animal Biosciences. 1, 99 -124 (2013).
112.
Hayday, A. & Tigelaar, R. Immunoregulation in the tissues by gammadelta T
cells. Nat Rev Immunol. 3(3), 233-242 (2003).
157
113.
Robak, E. et al. Lymphocyctes Tgammadelta in clinically normal skin and
peripheral blood of patients with systemic lupus erythematosus and their
correlation with disease activity. Mediators Inflamm. 10(4), 179-189 (2001).
114.
Toulon, A. et al. A role for human skin-resident T cells in wound healing. J Exp
Med. 206(4), 743-750 (2009).
115.
Pang, D.J., Neves, J.F., Sumaria, N. & Pennington, D.J. Understanding the
complexity of γδ T-cell subsets in mouse and human. Immunology. 136(3), 283290 (2012).
116.
Gioia, C. et al. Lack of CD27-CD45RA-V gamma 9V delta 2+ T cell effectors in
immunocompromised hosts and during active pulmonary tuberculosis. J Immunol.
168(3),1484-1489 (2002).
117.
Poupot, M. & Fournié, J.J. Non-peptide antigens activating human
Vgamma9/Vdelta2 T lymphocytes. Immunol Lett. 95(2), 129-138 (2004).
118.
Constant, P. et al. Stimulation of human gamma delta T cells by nonpeptidic
mycobacterial ligands. Science. 264(5156), 267-270 (1994).
119.
Morita, C.T., Jin, C., Sarikonda, G. & Wang, H. Nonpeptide antigens,
presentation mechanisms, and immunological memory of human
Vgamma2Vdelta2 T cells: discriminating friend from foe through the recognition
of prenyl pyrophosphate antigens. Immunol Rev. 215, 59-76 (2007).
120.
Jameson, J.M., Cruz, J., Costanzo, A., Terajima, M. & Ennis, F.A. A role for the
mevalonate pathway in the induction of subtype cross-reactive immunity to
influenza A virus by human gammadelta T lymphocytes. Cell Immunol. 264(1),
71-77 (2010).
121.
Morita, C.T. et al. Direct presentation of nonpeptide prenyl pyrophosphate
antigens to human gamma delta T cells. Immunity. 3(4), 495-507 (1995).
122.
Wang, H. et al. Butyrophilin 3A1 plays an essential role in prenyl pyrophosphate
stimulation of human Vγ2Vδ2 T cells. J Immunol. 191(3), 1029-1042 (2013).
123.
Vavassori, S. et al. Butyrophilin 3A1 binds phosphorylated antigens and
stimulates human γδ T cells. Nat Immunol. 14(9), 908-916 (2013).
158
124.
Roelofs, A.J. et al. Peripheral blood monocytes are responsible for gammadelta T
cell activation induced by zoledronic acid through accumulation of IPP/DMAPP.
Br J Haematol. 144(2), 245-250 (2009).
125.
Thompson, K. & Rogers, M.J. Statins prevent bisphosphonate-induced
gamma,delta-T-cell proliferation and activation in vitro. J Bone Miner Res. 19(2),
278-288 (2004).
126.
Wesch, D., Peters, C., Oberg, H.H., Pietschmann, K. & Kabelitz, D. Modulation
of γδ T cell responses by TLR ligands. Cell Mol Life Sci. 68(14), 2357-2370
(2011).
127.
Correia, D.V. et al. Differentiation of human peripheral blood Vδ1+ T cells
expressing the natural cytotoxicity receptor NKp30 for recognition of lymphoid
leukemia cells. Blood. 118(4), 992-1001 (2011).
128.
Nedellec, S., Sabourin, C., Bonneville, M. & Scotet, E. NKG2D costimulates
human V gamma 9V delta 2 T cell antitumor cytotoxicity through protein kinase
C theta-dependent modulation of early TCR-induced calcium and transduction
signals. J Immunol. 185(1), 55-63 (2010).
129.
Wrobel, P. et al. Lysis of a broad range of epithelial tumour cells by human
gamma delta T cells: involvement of NKG2D ligands and T-cell receptor- versus
NKG2D-dependent recognition. Scand J Immunol. 66(2-3), 320-328 (2007).
130.
Rincon-Orozco, B. et al. Activation of V gamma 9V delta 2 T cells by NKG2D. J
Immunol. 175(4), 2144-2151 (2005).
131.
Zheng, J., Liu, Y., Lau, Y.L. & Tu, W. γδ-T cells: an unpolished sword in human
anti-infection immunity. Cell Mol Immunol. 10(1), 50-57 (2013).
132.
Li, H. et al. Human Vγ9Vδ2-T cells efficiently kill influenza virus-infected lung
alveolar epithelial cells. Cell Mol Immunol. 10(2), 159-164 (2013).
133.
Couzi, L. et al. Antibody-dependent anti-cytomegalovirus activity of human γδ T
cells expressing CD16 (FcγRIIIa). Blood. 119(6), 1418-1427 (2012).
134.
Ninomiya, T. et al. Vgamma1+ gammadelta T cells play protective roles at an
early phase of murine cytomegalovirus infection through production of interferongamma. Immunology. 99(2), 187-194 (2000).
159
135.
Qin, G. et al. Type 1 responses of human Vγ9Vδ2 T cells to influenza A viruses. J
Virol. 85(19), 10109-10116 (2011).
136.
D'Ombrain, M.C., Hansen, D.S., Simpson, K.M. & Schofield, L. gammadelta-T
cells expressing NK receptors predominate over NK cells and conventional T
cells in the innate IFN-gamma response to Plasmodium falciparum malaria. Eur J
Immunol. 37(7), 1864-1873 (2007).
137.
O'Brien, R.L., Roark, C.L. & Born, W.K. IL-17-producing gammadelta T cells.
Eur J Immunol. 39(3), 662-666 (2009).
138.
McAleer, J.P. & Kolls, J.K. Mechanisms controlling Th17 cytokine expression
and host defense. J Leukoc Biol. 90(2), 263-270 (2011).
139.
Horowitz, A. et al. Cross-talk between T cells and NK cells generates rapid
effector responses to Plasmodium falciparum-infected erythrocytes. J Immunol.
184(11), 6043-6052 (2010).
140.
MacLeod, A.S. et al. Dendritic epidermal T cells regulate skin antimicrobial
barrier function. J Clin Invest. 123(10), 4364-4374 (2013).
141.
Shibata, K., Yamada, H., Hara, H., Kishihara, K. & Yoshikai, Y. Resident
Vdelta1+ gammadelta T cells control early infiltration of neutrophils after
Escherichia coli infection via IL-17 production. J Immunol. 178(7), 4466-4472
(2007).
142.
Dejima, T. et al. Protective role of naturally occurring interleukin-17A-producing
γδ T cells in the lung at the early stage of systemic candidiasis in mice. Infect
Immun. 79(11), 4503-4510 (2011).
143.
Li, Z., Burns, A.R., Miller, S.B. & Smith, C.W. CCL20, γδ T cells, and IL-22 in
corneal epithelial healing. FASEB J. 25(8), 2659-2668 (2011).
144.
Caccamo, N. et al. CXCR5 identifies a subset of Vgamma9Vdelta2 T cells which
secrete IL-4 and IL-10 and help B cells for antibody production. J Immunol.
177(8), 5290-5295 (2006).
145.
Sutton, C.E. et al. Interleukin-1 and IL-23 induce innate IL-17 production from
gammadelta T cells, amplifying Th17 responses and autoimmunity. Immunity.
31(2), 331-341 (2009).
160
146.
Huber, S.A. et al. γδ T lymphocytes kill T regulatory cells through CD1d.
Immunology. 131(2), 202-209 (2010).
147.
Schaerli, P. et al. CXC chemokine receptor 5 expression defines follicular homing
T cells with B cell helper function. J Exp Med. 192(11), 1553-1562 (2000).
148.
Meuter, S., Eberl, M. & Moser, B. Prolonged antigen survival and cytosolic
export in cross-presenting human gammadelta T cells. Proc Natl Acad Sci U S A.
107(19), 8730-8735 (2010).
149.
Brandes, M., Willimann, K. & Moser, B. Professional antigen-presentation
function by human gammadelta T Cells. Science. 309(5732), 264-268 (2005).
150.
Carding, S.R. et al. Late dominanace of the inflammatory process in murine
influenza by gamma/delta + T cells. J Exp Med. 172(4), 1225-1231 (1990).
151.
Jameson, J. et al. A role for skin gammadelta T cells in wound repair. Science.
296(5568), 747-749 (2002).
152.
Chen, Y., Chou, K., Fuchs, E., Havran, W.L. & Boismenu, R. Protection of the
intestinal mucosa by intraepithelial gamma delta T cells. Proc Natl Acad Sci U S
A. 99(22), 14338-14343 (2002).
153.
Workalemahu, G., Foerster, M. & Kroegel, C. Expression and synthesis of
fibroblast growth factor-9 in human gammadelta T-lymphocytes. Response to
isopentenyl pyrophosphate and TGF-beta1/IL-15. J Leukoc Biol. 75(4), 657-663
(2004).
154.
Schilbach, K., Frommer, K., Meier, S., Handgretinger, R. & Eyrich, M. Immune
response of human propagated gammadelta-T cells to neuroblastoma recommend
the Vdelta1+ subset for gammadelta-T-cell-based immunotherapy. J Immunother.
31(9), 896-905 (2008).
155.
De Rosa, S.C. et al. Ontogeny of gamma delta T cells in humans. J Immunol.
172(3), 1637-1645 (2004).
156.
Re, F. et al. Skewed representation of functionally distinct populations of
Vgamma9Vdelta2 T lymphocytes in aging. Exp Gerontol. 40(1-2), 59-66 (2005).
157.
Wijngaard, P.L. et al. Members of the novel WC1 gene family are differentially
expressed on subsets of bovine CD4-CD8- gamma delta T lymphocytes. J
Immunol. 152(7), 3476-3482 (1994).
161
158.
Pollock, J.M. & Welsh, M.D. The WC1(+) gammadelta T-cell population in
cattle: a possible role in resistance to intracellular infection. Vet Immunol
Immunopathol. 89(3-4), 105-114 (2002).
159.
Hedges, J.F., Cockrell, D., Jackiw, L., Meissner, N. & Jutila, M.A. Differential
mRNA expression in circulating gammadelta T lymphocyte subsets defines
unique tissue-specific functions. J Leukoc Biol. 73(2), 306-314 (2003).
160.
Meissner, N. et al. Serial analysis of gene expression in circulating gamma delta T
cell subsets defines distinct immunoregulatory phenotypes and unexpected gene
expression profiles. J Immunol. 170(1), 356-364 (2003).
161.
Kerns, H.M., Jutila, M.A. & Hedges, J.F. The distinct response of gammadelta T
cells to the Nod2 agonist muramyl dipeptide. Cell Immunol. 257(1-2), 38-43
(2009).
162.
Lahmers, K.K. et al. Comparative gene expression by WC1+ gammadelta and
CD4+ alphabeta T lymphocytes, which respond to Anaplasma marginale,
demonstrates higher expression of chemokines and other myeloid cell-associated
genes by WC1+ gammadelta T cells. J Leukoc Biol. 80(4), 939-952 (2006).
163.
Toka, F.N., Kenney, M.A. & Golde, W.T. Rapid and transient activation of γδ T
cells to IFN-γ production, NK cell-like killing, and antigen processing during
acute virus infection. J Immunol. 186(8), 4853-4861 (2011).
164.
Rhodes, S.G., Hewinson, R.G. & Vordermeier, H.M. Antigen recognition and
immunomodulation by gamma delta T cells in bovine tuberculosis. J Immunol.
166(9), 5604-5610 (2001).
165.
Davis, W.C. et al. Analysis of monoclonal antibodies specific for the gamma delta
TcR. Vet Immunol Immunopathol. 52(4), 275-283 (1996).
166.
Wilson, R.A., Zolnai, A., Rudas, P. & Frenyo, L.V. T-cell subsets in blood and
lymphoid tissues obtained from fetal calves, maturing calves, and adult bovine.
Vet Immunol Immunopathol. 53(1-2), 49-60 (1996).
167.
Ayoub, I.A. & Yang, T.J. Age-dependent changes in peripheral blood lymphocyte
subpopulations in cattle: a longitudinal study. Dev Comp Immunol. 20(5), 353363 (1996).
168.
Hogg, A.E. et al. Characterization of age-related changes in bovine CD8+ T-cells.
Vet Immunol Immunopathol. 140(1-2), 47-54 (2011).
162
169.
Rogers, A.N. et al. Function of ruminant gammadelta T cells is defined by WC1.1
or WC1.2 isoform expression. Vet Immunol Immunopathol. 108(1-2), 211-217
(2005).
170.
Price, S.J., Sopp, P., Howard. C.J. & Hope, J.C. Workshop cluster 1+ gammadelta
T-cell receptor T cells from calves express high levels of interferon-gamma in
response to stimulation with interleukin-12 and -18. Immunology. 120(1), 57-65
(2007).
171.
Foote, M.R. et al. Effects of age and nutrition on expression of CD25, CD44, and
L-selectin (CD62L) on T cells from neonatal calves. J Dairy Sci. 88(8), 27182729 (2005).
172.
Cheng, M., Chen, Y., Xiao, W., Sun, R. & Tian, Z. NK cell-based immunotherapy
for malignant diseases. Cell Mol Immunol. 10(3), 230-252 (2013).
173.
Fournié, J.J. et al. What lessons can be learned from γδ T cell-based cancer
immunotherapy trials? Cell Mol Immunol. 10(1), 35-41 (2013).
174.
Krifa, M., Bouhlel, I., Ghedira-Chekir, L. & Ghedira, K. Immunomodulatory and
cellular anti-oxidant activities of an aqueous extract of Limoniastrum guyonianum
gall. J Ethnopharmacol. 146(1), 243-249 (2013).
175.
Falchetti, R., Fuggetta, M.P., Lanzilli, G., Tricarico, M. & Ravagnan, G. Effects
of resveratrol on human immune cell function. Life Sci. 70(1), 81-96 (2001).
176.
Li, T., Fan, G.X., Wang, W., Li, T. & Yuan, Y.K. Resveratrol induces apoptosis,
influences IL-6 and exerts immunomodulatory effect on mouse lymphocytic
leukemia both in vitro and in vivo. Int Immunopharmacol. 7(9):1221-1231
(2007).
177.
Lu, C.C. & Chen, J.K. Resveratrol enhances perforin expression and NK cell
cytotoxicity through NKG2D-dependent pathways. J Cell Physiol. 223(2), 343351 (2010).
178.
Chen, C.M. et al. Consumption of purple sweet potato leaves modulates human
immune response: T-lymphocyte functions, lytic activity of natural killer cell and
antibody production. World J Gastroenterol. 11(37), 5777-5781 (2005).
179.
Yamanaka, D. et al. The effect of enzymatically polymerised polyphenols on CD4
binding and cytokine production in murine splenocytes. PLoS One. 7(4), e36025
(2012).
163
180.
Yamanaka, D. et al. Modulation of interferon-γ synthesis by the effects of ligninlike enzymatically polymerized polyphenols on antigen-presenting cell activation
and the subsequent cell-to-cell interactions. Food Chem. 141(4), 4073-4080
(2013).
181.
Mantena, S.K., Meeran, S.M., Elmets, C.A. & Katiyar, S.K. Orally administered
green tea polyphenols prevent ultraviolet radiation-induced skin cancer in mice
through activation of cytotoxic T cells and inhibition of angiogenesis in tumors. J
Nutr. 135(12), 2871-2877 (2005).
182.
John, C.M., Sandrasaigaran, P., Tong, C.K., Adam, A. & Ramasamy, R.
Immunomodulatory activity of polyphenols derived from Cassia auriculata
flowers in aged rats. Cell Immunol. 271(2), 474-479 (2011).
183.
Graff, J.C. & Jutila, M.A. Differential regulation of CD11b on gammadelta T
cells and monocytes in response to unripe apple polyphenols. J Leukoc Biol.
82(3), 603-607 (2007).
184.
Nantz, M.P., Rowe, C.A., Nieves, C. Jr. & Percival, S.S. Immunity and
antioxidant capacity in humans is enhanced by consumption of a dried,
encapsulated fruit and vegetable juice concentrate. J Nutr. 136(10), 2606-2610
(2006).
185.
Kamath, A.B. et al. Antigens in tea-beverage prime human Vgamma 2Vdelta 2 T
cells in vitro and in vivo for memory and nonmemory antibacterial cytokine
responses. Proc Natl Acad Sci U S A. 100(10), 6009-6014 (2003).
186.
Kiss, A.K. et al. Oenothein B's contribution to the anti-inflammatory and
antioxidant activity of Epilobium sp. Phytomedicine. 18(7), 557-560 (2011).
187.
Schmid, D. et al. Inhibition of NF-κB-dependent cytokine and inducible nitric
oxide synthesis by the macrocyclic ellagitannin oenothein B in TLR-stimulated
RAW 264.7 macrophages. J Nat Prod. 75(5), 870-875 (2012).
188.
Yoshimura, M. et al. Immunological effects of oenothein B, an ellagitannin
dimer, on dendritic cells. Int J Mol Sci. 14(1), 46-56 (2012).
189.
Lotito, S.B. et al. Influence of oligomer chain length on the antioxidant activity of
procyanidins. Biochem Biophys Res Commun. 276(3), 945-951 (2000).
164
190.
Lim, E.K. et al. Regiospecific methylation of a dietary flavonoid scaffold
selectively enhances IL-1β production following Toll-like receptor 2 stimulation
in THP-1 monocytes. J Biol Chem. 288(29), 21126-21135 (2013).
191.
Exon, J.H., Magnuson, B.A., South, E.H. & Hendrix, K. Dietary quercetin,
immune functions and colonic carcinogenesis in rats.Immunopharmacol
Immunotoxicol. 20(1), 173-190 (1998).
192.
Lambert, J.D., Sang, S. & Yang, C.S. Possible controversy over dietary
polyphenols: benefits vs risks. Chem Res Toxicol. 20, 583-585 (2007).
193.
Kiss, A., Kowalski, J. & Melzig, M.F. Induction of neutral endopeptidase activity
in PC-3 cells by an aqueous extract of Epilobium angustifolium L. and oenothein
B. Phytomedicine 13, 284-289 (2006).
194.
Wen, W., Lu, J., Zhang, K. & Chen, S. Grape seed extract inhibits angiogenesis
via suppression of the vascular endothelial growth factor receptor signaling
pathway. Cancer Prev Res (Phila). 1, 554-561 (2008).
195.
Bonneville, M. & Scotet, E. Human Vγ9Vδ2 T cells: promising new leads for
immunotherapy of infections and tumors. Curr Opin Immunol.18, 539-546
(2006).
196.
Gomes, A.Q., Martins, D.S. & Silva-Santos, B. Targeting γδ T lymphocytes for
cancer immunotherapy: from novel mechanistic insight to clinical application.
Cancer Res. 70, 10024-10027 (2010).
197.
Levy, E.M., Roberti, M.P. & Mordoh, J. Natural killer cells in human cancer:
from biological functions to clinical applications. J Biomed Biotechnol. 2011,
676198 (2011).
198.
Thompson, K., Rojas-Navea, J. & Rogers, M.J. Alkylamines cause Vγ9Vδ2Tcellactivation and proliferation by inhibiting the mevalonate pathway. Blood. 107,
651-654 (2006).
199.
Holderness, J. et al. Polysaccharides isolated from Açaí fruit induce innate
immune responses. PLoS One. 6, e17301 (2011).
200.
Graff, J.C. et al. Polysaccharides derived from Yamoa (Funtumia elastica) prime
γδ T cells in vitro and enhance innate immune responses in vivo. Int
Immunopharmacol. 9, 1313-1322 (2009).
165
201.
Percival, S.S., Bukowski, J.F. & Milner, J. Bioactive food components that
enhance γδ T cell function may play a role in cancer prevention. J Nutr. 138, 1-4
(2008).
202.
Gao, Y. et al. γδ T cells provide an early source of interferon gamma in tumor
immunity. J Exp Med. 198, 433-442 (2003).
203.
Bancroft, G.J., Schreiber, R.D. & Unanue, E.R. Natural immunity: a T-cellindependent pathway of macrophage activation, defined in the scid mouse.
Immunol Rev. 124, 5-24 (1991).
204.
Lockhart, E., Green, A.M. & Flynn, J.L. IL-17 production is dominated by γδ T
cells rather than CD4 T cells during Mycobacterium tuberculosis infection. J
Immunol. 177, 4662-4669 (2006).
205.
Wheelock, E.F. Interferon-like virus-inhibitor induced in human leukocytes by
phytohemagglutinin. Science. 149, 310-311 (1965).
206.
Virelizier, J.L. & Arenzana-Seisdedos, F. Immunological functions of
macrophages and their regulation by interferons. Med Biol. 63, 149-159 (1985).
207.
Tannenbaum, C.S. & Hamilton, T.A. Immune-inflammatory mechanisms in
IFNγ-mediated anti-tumor activity. Semin Cancer Biol. 10, 113-123 (2000).
208.
Srivastava, S., Salim, N. & Robertson, M.J. Interleukin-18: biology and role in the
immunotherapy of cancer. Curr Med Chem. 17, 3353-3357 (2010).
209.
Fukuchi, K. et al. Inhibition of herpes simplex virus infection by tannins and
related compounds. Antiviral Res.11, 285-297 (1989).
210.
Yoshida, T., Hatano, T. & Ito, H. Chemistry and function of vegetable
polyphenols with high molecular weights. Biofactors. 13, 121-125 (2000).
211.
Sakagami, H. et al. Cytotoxic activity of hydrolyzable tannins against human oral
tumor cell lines--a possible mechanism. Phytomedicine. 7, 39-47 (2000).
212.
Wilson, E., Walcheck, B., Davis, W.C. & Jutila, M.A. Preferential tissue
localization of bovine γδ T cell subsets defined by anti-T cell receptor for antigen
antibodies. Immunol Lett. 64, 39-44 (1998).
166
213.
Vesosky, B., Turner, O.C., Turner, J. & Orme, I.M. Gamma interferon production
by bovine gamma delta T cells following stimulation with mycobacterial
mycolylarabinogalactan peptidoglycan. Infect Immun. 72, 4612-4618 (2004).
214.
Allen, A.J. et al. Experimental infection of a bovine model with human isolates of
Mycobacterium avium subsp. paratuberculosis. Vet Immunol Immunopathol. 141,
258-266 (2011).
215.
Van Rhijn, I., Godfroid, J., Michel, A. & Rutten, V. Bovine tuberculosis as a
model for human tuberculosis: advantages over small animal models. Microbes
Infect. 10, 711-715 (2008).
216.
Costa, L.F., Paixão, T.A., Tsolis, R.M., Bäumler, A.J. & Santos, R.L.
Salmonellosis in cattle: advantages of being an experimental model. Res Vet Sci.
93, 1-6 (2012).
217.
Kenna, T. et al. Distinct subpopulations of γδ T cells are present in normal and
tumor-bearing human liver. Clin Immunol. 113, 56-63 (2004).
218.
Wu, S.L., Yu, L., Jiao, X.Y., Meng, K.W. & Pan, C.E. The suppressive effect of
resveratrol on protein kinase C theta in peripheral blood T lymphocytes in a rat
liver transplantation model. Transplant Proc. 38, 3052-3054 (2006).
219.
Hushmendy, S. et al. Select phytochemicals suppress human T-lymphocytes and
mouse splenocytes suggesting their use in autoimmunity and transplantation. Nutr
Res. 29, 568-578 (2009).
220.
Lion, E., Smits, E.L., Berneman, Z.N. & Van Tendeloo, V.F. Quantification of
IFN-γ produced by human purified NK cells following tumor cell stimulation:
comparison of three IFN-γ assays. J Immunol Methods. 350, 89-96 (2009).
221.
Korbel, D.S., Newman, K.C., Almeida, C.R., Davis, D.M. & Riley, E.M.
Heterogeneous human NK cell responses to Plasmodium falciparum-infected
erythrocytes. J Immunol. 175,7466-7473 (2005).
222.
Chipeta, J. et al. CD4+ and CD8+ cell cytokine profiles in neonates, older
children, and adults: increasing T helper type 1 and T cytotoxic type 1 cell
populations with age. Cell Immunol. 183,149-156 (1998).
223.
Robinson, D. et al. IGIF does not drive Th1 development but synergizes with IL12 for interferon-gamma production and activates IRAK and NFκB. Immunity. 7,
571-581 (1997).
167
224.
Ramstead, A.G., Schepetkin, I.A., Quinn, M.T. & Jutila, M.A. Oenothein B, a
cyclic dimeric ellagitannin isolated from Epilobium angustifolium, enhances IFNγ
production by lymphocytes. PLoS One. 7(11), e50546 (2012).
225.
Rothstein, D.M., Sohen, S., Daley, J.F., Schlossman, S.F. & Morimoto, C.
CD4+CD45RA+ and CD4+CD45RA- T cell subsets in man maintain distinct
function and CD45RA expression persists on a subpopulation of CD45RA+ cells
after activation with Con A. Cell Immunol. 129(2), 449-467 (1990).
226.
Reem, G.H., Cook, L.A., Henriksen, D.M. & Vilcek, J. Gamma interferon
induction in human thymocytes activated by lectins and B cell lines. Infect
Immun. 37(1), 216-221 (1982).
227.
Lewinsohn, D.A. & Lewinsohn, D.M. Immunologic Susceptibility of Young
Children to Mycobacterium tuberculosis. Pediatric Research. 63, 115–115 (2008).
228.
Ochoa, T.J. et al. Age-related susceptibility to infection with diarrheagenic
Escherichia coli among infants from Periurban areas in Lima, Peru. Clin Infect
Dis. 49(11), 1694-1702 (2009).
229.
Warren, H.S. & Skipsey, L.J. Loss of activation-induced CD45RO with
maintenance of CD45RA expression during prolonged culture of T cells and NK
cells. Immunology. 74(1), 78-85 (1991).
230.
Kersh, E.N. et al. Rapid demethylation of the IFN-gamma gene occurs in memory
but not naive CD8 T cells. J Immunol. 176(7), 4083-4093 (2006).
231.
White, G.P., Watt, P.M., Holt, B.J. & Holt, P.G. Differential patterns of
methylation of the IFN-gamma promoter at CpG and non-CpG sites underlie
differences in IFN-gamma gene expression between human neonatal and adult
CD45RO- T cells. J Immunol. 168(6), 2820-2827 (2002).
232.
Vakkila, J. & Lotze, M.T. Inflammation and necrosis promote tumour growth.
Nat Rev Immunol 4(8), 641-648 (2004).
233.
Chow, M.T., Möller, A. & Smyth, M.J. Inflammation and immune surveillance in
cancer. Semin Cancer Biol 22(1), 23-32 (2012).
234.
Fisch, P. et al. Gamma/delta T cell clones and natural killer cell clones mediate
distinct patterns of non-major histocompatibility complex-restricted cytolysis. J
Exp Med 171(5), 1567-1579 (1990).
168
235. Groh, V. et al. Broad tumor-associated expression and recognition by tumorderived gamma delta T cells of MICA and MICB. Proc Natl Acad Sci U S A
96(12), 6879-6884 (1999).
236.
Girardi, M. et al. Regulation of cutaneous malignancy by gammadelta T cells.
Science 294(5542), 605-609 (2001).
237.
Peng, G. et al. Tumor-infiltrating gammadelta T cells suppress T and dendritic
cell function via mechanisms controlled by a unique toll-like receptor signaling
pathway. Immunity 27(2), 334-348 (2007).
238.
He, W. Naturally activated V gamma 4 gamma delta T cells play a protective role
in tumor immunity through expression of eomesodermin. J Immunol 185(1), 126133 (2010).
239.
Bryant, N.L. et al. Preclinical evaluation of ex vivo expanded/activated γδ T cells
for immunotherapy of glioblastoma multiforme. J Neurooncol 101(2), 179-188
(2011).
240.
Hannani, D. et al. Harnessing γδ T cells in anticancer immunotherapy. Trends
Immunol 33(5), 199-206 (2012).
241.
Dieli, F. et al. Targeting human gamma
delta T cells with zoledronate and
interleukin-2 for immunotherapy of hormone-refractory prostate cancer. Cancer
Res 67(15), 7450-7457 (2007).
242.
Benzaïd, I. et al. High phosphoantigen levels in bisphosphonate-treated human
breast tumors promote Vgamma9Vdelta2 T-cell chemotaxis and cytotoxicity in
vivo. Cancer Res 71(13), 4562-4572 (2011).
243.
Castella, B., Vitale, C., Coscia, M. & Massaia, M. Vγ9Vδ2 T cell-based
immunotherapy in hematological malignancies: from bench to bedside. Cell Mol
Life Sci 68(14), 2419-2432 (2011).
244.
Kalyan, S., Wesch, D. & Kabelitz, D. Aminobisphosphonates and Toll-like
receptor ligands: recruiting Vγ9Vδ2 T cells for the treatment of hematologic
malignancy. Curr Med Chem 18(34), 5206-5216 (2011).
245.
Yoshida, Y., Nakajima, J., Wada, H. & Kakimi, K. γδ T-cell immunotherapy for
lung cancer. Surg Today 41(5), 606-611 (2011).
169
246.
Capietto, A.H., Martinet, L. & Fournié, J.J. How tumors might withstand γδ Tcell attack. Cell Mol Life Sci 68(14), 2433-2442 (2011).
247.
Moser, B. & Eberl, M. γδ T-APCs: a novel tool for immunotherapy? Cell Mol
Life Sci 68(14), 2443-2452 (2011).
248.
Seo, N., Tokura, Y., Takigawa, M. & Egawa, K. Depletion of IL-10- and TGFbeta-producing regulatory gamma delta T cells by administering a daunomycinconjugated specific monoclonal antibody in early tumor lesions augments the
activity of CTLs and NK cells. J Immunol 163(1), 242-249 (1999).
249.
Ke, Y., Kapp, L.M. & Kapp, J.A. Inhibition of tumor rejection by gammadelta T
cells and IL-10. Cell Immunol 221(2), 107-114 (2003).
250.
Bialasiewicz, A.A., Ma, J.X. & Richard, G. Alpha/beta- and gamma/delta TCR(+)
lymphocyte infiltration in necrotising choroidal melanomas. Br J Ophthalmol
83(9), 1069-1073 (1999).
251.
Inman, B.A. et al. Questionable relevance of gamma delta T lymphocytes in renal
cell carcinoma. J Immunol 180(5), 3578-3584 (2008).
252.
Bonneville, M., O'Brien, R.L. & Born, W.K. Gammadelta T cell effector
functions: a blend of innate programming and acquired plasticity. Nat Rev
Immunol 10(7), 467-478 (2010).
253.
Hao, J. et al. Current progress in γδ T-cell biology. Cell Mol Immunol 7(6), 409413 (2010).
254.
Wesch, D., Glatzel, A. & Kabelitz, D. Differentiation of resting human peripheral
blood gamma delta T cells toward Th1- or Th2-phenotype. Cell Immunol 212(2),
110-117 (2001).
255.
Paget, C., Chow, M.T., Duret, H., Mattarollo, S.R. & Smyth, M.J. Role of γδ T
cells in α-galactosylceramide-mediated immunity. J Immunol 188(8), 3928-3939
(2012).
256.
Talmadge, J.E., Tribble, H.R., Pennington, R.W., Phillips, H. & Wiltrout, R.H.
Immunomodulatory and immunotherapeutic properties of recombinant gammainterferon and recombinant tumor necrosis factor in mice. Cancer Res 47(10),
2563-2570 (1987).
170
257.
Lejeune, F.J., Liénard, D., Matter, M. & Rüegg, C. Efficiency of recombinant
human TNF in human cancer therapy. Cancer Immun 6:6 (2006).
258.
Lu, Y. et al. Responsiveness of stromal fibroblasts to IFN-gamma blocks tumor
growth via angiostasis. J Immunol 183(10), 6413-6421 (2009).
259.
Poggi, A. et al. Vdelta1 T lymphocytes from B-CLL patients recognize ULBP3
expressed on leukemic B cells and up-regulated by trans-retinoic acid. Cancer Res
64(24), 9172-9179 (2004).
260.
Halary, F. et al. Shared reactivity of Vdelta2neg gamma delta T cells against
cytomegalovirus-infected cells and tumor intestinal epithelial cells. J Exp Med
201(10), 1567-1578 (2005).
261.
Riond, J., Rodriguez, S., Nicolau, M.L., al Saati. T. & Gairin, J.E. In vivo major
histocompatibility complex class I (MHCI) expression on MHCI low tumor cells
is regulated by gammadelta T and NK cells during the early steps of tumor
growth. Cancer Immun 9, 10 (2009).
262.
Gaafar, A. et al. Defective gammadelta T-cell function and granzyme B gene
polymorphism in a cohort of newly diagnosed breast cancer patients. Exp
Hematol 37(7), 838-848 (2009).
263.
Argentati, K. et al. Reduced number and impaired function of circulating gamma
delta T cells in patients with cutaneous primary melanoma. J Invest Dermatol
120(5), 829-834 (2003).
264.
Puan, K.J. et al. Phenotypic and functional alterations of Vgamma2Vdelta2 T cell
subsets in patients with active nasopharyngeal carcinoma. Cancer Immunol
Immunother 58(7), 1095-1107 (2009).
265.
Provinciali, M., Re, F., Tucci, M.G., Ricotti, F. & Lattanzio, F. Persistent ex vivo
low number and functional in vitro recovery of circulating gammadelta T cells
after removal of a cutaneous primary melanoma. Scand J Immunol 72(2), 142-149
(2010).
266.
Martinet, L. et al. A regulatory cross-talk between Vgamma9Vdelta2 T
lymphocytes and mesenchymal stem cells. Eur J Immunol 39(3), 752-762 (2009).
267.
Kunzmann, V. et al. Tumor-promoting versus tumor-antagonizing roles of γδ T
cells in cancer immunotherapy: results from a prospective phase I/II trial. J
Immunother 35(2), 205-213 (2012).
171
268.
Abe, Y. et al. Clinical and immunological evaluation of zoledronate-activated
Vgamma9gammadelta T-cell-based immunotherapy for patients with multiple
myeloma. Exp Hematol 37(8), 956-968 (2009).
269.
Seo, N., Tokura, Y., Furukawa, F. & Takigawa, M. Down-regulation of
tumoricidal NK and NK T cell activities by MHC Kb molecules expressed on
Th2-type gammadelta T and alphabeta T cells coinfiltrating in early B16
melanoma lesions. J Immunol 161(8), 4138-4145 (1998).
270.
Hao, J. et al. Regulatory role of Vγ1 γδ T cells in tumor immunity through IL-4
production. J Immunol 187(10), 4979-4986 (2011).
271.
Kühl, A.A. et al. Human peripheral gammadelta T cells possess regulatory
potential. Immunology 128(4), 580-588 (2009).
272.
Ness-Schwickerath, K.J. & Morita, C.T. Regulation and function of IL-17A- and
IL-22-producing γδ T cells. Cell Mol Life Sci 68(14), 2371-2390 (2011).
273.
Alshaker, H.A. & Matalka, K.Z. IFN-γ, IL-17 and TGF-β involvement in shaping
the tumor microenvironment: The significance of modulating such cytokines in
treating malignant solid tumors. Cancer Cell Int 11, 33 (2011).
274.
Wakita, D. et al. Tumor-infiltrating IL-17-producing gammadelta T cells support
the progression of tumor by promoting angiogenesis. Eur J Immunol 40(7), 19271937 (2010).
275.
Liu, J. et al. IL-17 is associated with poor prognosis and promotes angiogenesis
via stimulating VEGF production of cancer cells in colorectal carcinoma.
Biochem Biophys Res Commun 407(2), 348-354 (2011).
276.
Carmi, Y. et al. Microenvironment-derived IL-1 and IL-17 interact in the control
of lung metastasis. J Immunol 186(6), 3462-3471 (2011).
277.
Takeuchi, A. et al. IL-17 production by γδ T cells is important for the anti-tumor
effect of Mycobacterium bovis bacillus Calmette-Guérin treatment against
bladder cancer. Eur J Immunol 41(1), 246-251 (2011).
278.
Ma, Y. et al. Contribution of IL-17-producing gamma delta T cells to the efficacy
of anticancer chemotherapy. J Exp Med 208(3), 491-503 (2011).
172
279.
Ceccarelli, S., Romano, F., Angeloni, A. & Marchese, C. Potential dual role of
KGF/KGFR as a target option in novel therapeutic strategies for the treatment of
cancers and mucosal damages. Expert Opin Ther Targets 16(4), 377-393 (2012).
280.
Saharinen, P., Eklund, L., Pulkki, K., Bono, P. & Alitalo, K. VEGF and
angiopoietin signaling in tumor angiogenesis and metastasis. Trends Mol Med
17(7), 347-362 (2011).
281.
Niu, J. et al. Keratinocyte growth factor/fibroblast growth factor-7-regulated cell
migration and invasion through activation of NF-kappaB transcription factors. J
Biol Chem 282(9), 6001-6011 (2007).
282.
Behr, B., Leucht, P., Longaker, M.T. & Quarto, N. Fgf-9 is required for
angiogenesis and osteogenesis in long bone repair. Proc Natl Acad Sci U S A
107(26), 11853-11858 (2010).
283.
Welte, T. et al. Role of two distinct gammadelta T cell subsets during West Nile
virus infection. FEMS Immunol Med Microbiol 53(2), 275-283 (2008).
284.
Sugaya, M., Nakamura, K. & Tamaki, K. Interleukins 18 and 12 synergistically
upregulate interferon-gamma production by murine dendritic epidermal T cells. J
Invest Dermatol 113(3), 350-354 (1999).
285.
Holtmeier, W. & Kabelitz, D. gammadelta T cells link innate and adaptive
immune responses. Chem Immunol Allergy 86, 151-183 (2005).
286.
Li, W. et al. Effect of IL-18 on expansion of gammadelta T cells stimulated by
zoledronate and IL-2. J Immunother 33(3), 287-296 (2010).