AN ABSTRACT OF THE THESIS OF
advertisement

AN ABSTRACT OF THE THESIS OF
Kamal Issa Masoud Al-Malah for the Degree of Doctor of
Philosophy in Chemical Engineering Presented on June 17,
1993
Title: A Macroscopic Model for Apparent Protein Adsorption
Equilibrium at Hydrophobic Solid-Water Interfaces.
Redacted for Privacy
Abstract Approved
Joseph McGuire
Redacted for Privacy
Robert D. Sp ull
A predictive model was developed to describe apparent
protein adsorption equilibrium at solid-water interfaces.
Dimensional analysis was used to express adsorbed mass in
terms of the following macroscopic properties: the partial
molar area occupied by protein at the interface, AP; the
work of adhesion per unit area, W,; the minimum surface area
cleared by an adsorbing protein, Ac; the Gibbs free energy
of unfolding, AGm"; the partial molar volume of protein in
solution, Vp; and the apparent equilibrium concentration,
C.
Considering only adsorption at hydrophobic interfaces
in the absence of specific electrostatic effects and
biochemical interactions, an adsorption mechanism was
constructed to visualize protein adsorption equilibrium.
The proposed mechanism consists of two steps, the first
being reversible arrival of the native protein molecule to
the interface.
molecule.
The second step is unfolding of the adsorbed
Arrangement of dimensionless groups comprised of the
factors enumerated above was then performed with reference
to the proposed mechanism.
The model was then tested using
experimentally measured isotherms for a number of proteins.
A general agreement in adsorbed mass was found between the
model and the experiments, indicating that at hydrophobic
surfaces AGmud and 147Ac play a major role in governing the
course of adsorption.
mass, r
,
Also the plateau value of adsorbed
was found to be directly related to A.
It was
found, however, that at hydrophobic, low-energy solid-water
interfaces, protein adsorption equilibrium can be
represented by a one-step mechanism for modeling purposes if
equilibrium spreading pressure measurements are used to
estimate protein molecule interfacial energy.
The model was applied to both simple, single domain
globular proteins as well as to proteins of more complex
structure.
Concerning multi-domain proteins, the most
thermolabile domain was observed to play the major role in
initial events contributing to surface-induced unfolding.
All parameters but Ac served as input to the model;
computer-generated values of Ac obtained from simulation
varied within an order of magnitude (about 100 to 400
A2 /molecule),
indicating that only a small portion of the
protein molecule need enter the interface in order for
adsorption to proceed. For single-domain proteins,
variations in Ac can be related to particular properties of
the protein, such as molecular weight and flexibility.
A Macroscopic Model for Apparent Protein Adsorption
Equilibrium at Hydrophobic Solid-Water Interfaces
by
Kamal Issa Masoud Al-Malah
A THESIS
Submitted to
Oregon State University
in partial fulfillment of
the requirement for the
degree of
Doctor of Philosophy
Completed June 17, 1993
Commencement June 1994
APPROVED:
Redacted for Privacy
Assoc
Scien
sor of Bioresource Engineering, and Food
logy in charge of major
Redacted for Privacy
Assistant Professor of Chemical
major
ngineering in charge of
Redacted for Privacy
Head of Department of Chemical Engineering
Redacted for Privacy
Dean of Graduake School
Date thesis is presented
June 17, 1993
Typed by the researcher for
Kamal Issa Masoud Al-Malah
TABLE OF CONTENTS
PAGE
1.
INTRODUCTION
1
2.
LITERATURE REVIEW
4
2.1 PROTEINS
4
2.1.1
PROTEIN STABILITY
5
2.1.2
HYDROPHOBIC INTERACTIONS
8
PROTEIN CONFORMATION
2.2 PROTEIN ADSORPTION
10
MODEL DEVELOPMENT
20
3.1 DIMENSIONAL ANALYSIS
20
2.1.3
3.
3.1.1
QUANTIFYING THE FACTORS IN
EQUATION (2)
3.2 A SIMPLE MECHANISM FOR PROTEIN ADSORPTION
13
22
35
3.3 LIMITATIONS ON THE APPLICABILITY OF THE
MODEL (EQ. 30)
45
4.
RESULTS
47
5.
DISCUSSION
84
5.1
PARAMETERS a AND A,
5.1.1
VALUE OF a AT LOW-ENERGY INTERFACES
VALUE OF a AT HIGH-ENERGY INTERFACES
5.2 MOLECULAR INFLUENCES ON A,
5.1.2
85
85
86
87
5.3 AN APPROXIMATE ESTIMATE FOR A, AS A
FUNCTION OF MW AND Z
90
5.4 IMPLICATIONS ASSOCIATED WITH THE MODEL
APPLICATION TO MULTI-DOMAIN PROTEINS
5.5 THE AFFINITY AND EXTENT OF ADSORPTION
6.
7.
94
100
5.5.1
ADSORPTION AFFINITY
100
5.5.2
THE EXTENT OF ADSORPTION
102
5.5.3
A NOTE ON TEMPERATURE EFFECTS
103
CONCLUSIONS
RECOMMENDATIONS
105
BIBLIOGRAPHY
108
107
APPENDICES
APPENDIX
APPENDIX
APPENDIX
1
124
2
126
3
127
LIST OF FIGURES
FIGURE
3.1.1
PAGE
The equilibrium contact angle, 0, formed
between a solid/liquid (SL) interface and
a liquid vapor (LV) interface.
3.1.2
23
The flexibility parameter (Z) correlated
with AGfold/RT for a-Lac,
(3 -Lag, BSA, and
Lyso.
3.2.1
27
A schematic depicting protein adsorption
equilibrium.
4.1
39
Protein adsorption isotherms (T =27 °C) on a
hydrophobic silicon surface, fitted to
Eq.
(30). Ref: Suttiprasit and McGuire
(1992)
4.2
56
.
Protein adsorption isotherms (T =52 °C) on a
hydrophobic silicon surface, fitted to
Eq. (30). Ref: Suttiprasit and McGuire
(1992)
4.3
57
.
Protein adsorption isotherms on hydrophobic,
butylated quartz slides, fitted to Eq.
(30).
Ref: Shibata & Lenhoff (1992).
4.4
61
Human Lysozyme adsorption isotherm (T =25 °C)
on hydrophobic DDS silica slides, fitted to
Eq.
4.5
(30).
Ref: Horsley et al.
(1987).
64
Plasminogen adsorption isotherm (T =25 °C) on
a hydrophobic MDA polyurethane surface,
fitted to Eq.
et al.
(1992).
(30).
Ref: Woodhouse
67
List of Figures (Continued)
4.6
HSA adsorption isotherms (T=25°C) on
different hydrophobic surfaces, fitted
to Eq.
4.7
(30).
Ref: Winterton et al.
(1986).
Protein adsorption isotherms (T =24 °C) on
hydrophobic siliconized glass, fitted to
Eq. (30).
Ref: Absolom et al. (1987).
4.8
70
74
Fbrgn adsorption isotherm (T =25 °C) on a
hydrophobic quartz surface, fitted to
Eq.
4.9
(30).
Ref: Nygren & Stenberg (1988).
Protein adsorption isotherms (T =25 °C) on
hydrophobic polystyrene latices, fitted to
Eq. (30).
Ref: Arai & Norde (1990).
5.3.1
Isotherms of Fig.
5.3.3
81
(4.1) plotted with the
isotherm-specific Ac value and a value of
a = 0.0.
5.3.2
78
91
Isotherms of Fig.
(4.1) constructed with the
same Ac value (A, = 192 A2) and a value of
a = 0.0.
92
Ac correlated with MW and Z.
93
LIST OF TABLES
TABLE
4.1
4.2
PAGE
The regressed values of a and Ac, and the
regression parameters for the model (Eq. 30)
48
Thermal properties of proteins used in
evaluating AGmfdd (equation 13), with
standard errors shown in parentheses.
51
4.3
The specific volume (Vp), molecular weight
(MW), and the partial molar area of protein
upon adsorption, Ap, for each of the proteins
examined in this study.
53
4.4
The Gibbs free energy of unfolding, protein,
solid, and water surface energies, and the
work of adhesion for proteins used in Figs.
(4.1) and (4.2).
Source of isotherms:
Suttiprasit and McGuire (1992).
58
4.5
The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant,
K, and the regression parameters for the
model (Eq. 30). Source of isotherms:
Suttiprasit and McGuire (1992).
59
4.6
The Gibbs free energy of unfolding, protein,
solid, and water surface energies, and the
work of adhesion for proteins used in Fig.
(4.3).
Source of isotherms: Shibata &
Lenhoff (1992).
62
4.7
The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant,
K, and the regression parameters for the
model (Eq. 30).
Source of isotherms:
Shibata & Lenhoff (1992).
62
4.8
The Gibbs free energy of unfolding, protein,
solid, and water surface energies, and the
work of adhesion for protein used in Fig.
(4.4).
Source of isotherm: Horsley et al.
(1987).
4.9
65
The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant,
K, and the regression parameters for the
model (Eq. 30).
Source of isotherm:
Horsley et al. (1987).
65
List of Tables (Continued)
4.10
The Gibbs free energy of unfolding, protein,
solid, and water surface energies, and the
work of adhesion for protein used in Fig.
Source of isotherm: Woodhouse et al.
(4.5).
(1992) .
4.11
4.12
4.13
4.14
4.15
4.16
4.17
The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant,
K, and the regression parameters for the
Source of isotherm:
model (Eq. 30).
Woodhouse et al. (1992).
68
68
The Gibbs free energy of unfolding, protein,
solid, and water surface energies, and the
work of adhesion for protein used in Fig.
Source of isotherms: Winterton et al.
(4.6).
(1986).
71
The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant,
K, and the regression parameters for the
Source of isotherms:
model (Eq. 30).
Winterton et al. (1986).
72
The Gibbs free energy of unfolding, protein,
solid, and water surface energies, and the
work of adhesion for proteins used in Fig.
(4.7).
Source of isotherms: Absolom et al.
(1987).
75
The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant,
K, and the regression parameters for the
model (Eq. 30).
Source of isotherms:
Absolom et al. (1987).
76
The Gibbs free energy of unfolding, protein,
solid, and water surface energies, and the
work of adhesion for protein used in Fig.
(4.8).
Source of isotherm: Nygren and
Stenberg (1988).
79
The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant,
K, and the regression parameters for the
model (Eq. 30).
Source of isotherm:
Nygren and Stenberg (1988).
79
List of Tables (Continued)
4.18
4.19
The Gibbs free energy of unfolding, protein,
solid, and water surface energies, and the
work of adhesion for proteins used in Fig.
Source of isotherms: Arai and Norde
(4.9).
(1990).
82
The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant,
K, and the regression parameters for the
Source of isotherm: Arai
model (Eq. 30).
and Norde (1990).
83
NOMENCLATURE
a
A function constant that defines the functionality
of r versus Cul (Eq. 19).
Ac
Minimum surface area (i.e., "hole area") cleared by
an adsorbing protein molecule in order to anchor
itself to the interface
(m2 /mol) or (k/molecule).
Partial molar area occupied by species i at the
interface
a,
(m2/mol) or (A2/rnolecule).
The average interfacial area occupied by an amino
acid residue
Ap
(A2/molecule).
Partial molar area occupied by protein at the
interface as defined by either Eq. (16) or (17)
(m2/mol) or (A2/molecule)
b
A constant such that r
.
/ b is the initial slope of
a plot of r versus Cry
(mg/1).
BSA
Bovine Serum Albumin
Cat
Apparent equilibrium concentration (mol/m3)
or
(mg/1) .
ChA
Chymotrypsinogen A
ACp
The difference in heat capacity between the unfolded
and folded states
(kJ/mole
K).
f
Fractional surface coverage
Fbrgn
Fibrinogen.
GN
Partial molar free energy of native protein
(kJ/mol).
(dimensionless).
GU
Partial molar free energy of unfolded protein
(kJ/mol).
AG,,
The standard Gibbs free-energy change of adsorption
(kJ/mol).
AGwom
Change in partial molar free energy of protein upon
unfolding
AHaB
(kJ/mol).
The standard enthalpy change of adsorption
(kJ/mol).
AHD
The enthalpy of denaturation (kJ/mol).
AM'
The non-hydrophobic enthalpic contribution to the
free energy change (Eq. 14)
(kJ/mol amino acid
residue).
HSA
Human Serum Albumin.
IgG
Immunoglobulin G.
K
The proportionality or the overall equilibrium
constant for the protein adsorption process
(dimensionless).
Ka
The equilibrium constant for the reversible arrival
step
(dimensionless).
kB
Boltzmann constant
Ku
The equilibrium constant for the reversible,
(1.38 X 10-23 J/molecule
surface-induced unfolding step
a-Lac
a-Lactalbumin.
f3 -Lag
fl-Lactoglobulin.
Lyso
Lysozyme.
MSE
The mean of square errors.
MW
Molecular weight.
K).
(dimensionless).
Myog
Myoglobin.
NA
Avogadro's number
Plmgn
Plasminogen.
PS
Polystyrene polymer.
PVC
Polyvinylchloride polymer.
RiboA
Ribonuclease A.
Silas
Silastic polymer.
AS.
Multiplied by T gives the non-hydrophobic entropic
(6.02217 X 1023 molecule/mol).
contribution to the free energy change (Eq. 14)
(kJ/mol amino acid residue
K).
T
Temperature
TH
The temperature at which the enthalpic contribution
(K).
to the free energy change (Eq. 14) is purely non-
hydrophobic
T,
(K).
The melting point temperature of protein at which
the thermogram peak reaches its maximum
Ts
(K).
The temperature at which the entropic contribution
to the free energy change (Eq. 14) is purely non-
hydrophobic
V;
(K).
Partial molar volume of species i in solution
(e/mol).
VP
Partial molar volume of protein in solution (m3/mol).
Ws
Work of Adhesion as defined by equation (3)
Z
The surface coordination number of the lattice as
defined by equation (11)
(Z = 2 + w).
(J/m2)
.
GREEK SYMBOLS
a
The proportionality constant that relates the Gibbs
free energy of unfolding in the bulk to that at the
(dimensionless).
interface
Adsorbed amount of protein (mol/m2) or (gg/cm2).
The plateau value of adsorbed amount of protein
(mol/m2) or (gg/cm2).
The ratio of the volume occupied by all atoms making
up a molecule to the total volume occupied by the
molecule itself
(dimensionless).
An ith independent dimensionless group that comprises
a multiplication of some dimensional variables or
parameters
'Y12
(dimensionless).
The interfacial free energy of any pair combination
of two phases from the protein (P), water (W), and
solid (S) phases as defined by equation (7)
'Yps
(mJ/m2).
The interfacial free energy between protein and
solid
(mJ/m2)
.
'Ypv
The interfacial free energy of protein (mJ/m2).
'Ypw
The interfacial free energy between protein and
water
(mJ/m2)
.
'Ysv
The interfacial free energy of solid (mJ/m2).
'Ysw
The interfacial free energy between solid and water
(mJ/m2)
'Ywv
.
The interfacial free energy of water (mJ/m2).
0
The equilibrium contact angle formed between a
solid- liquid (SL) interface and a liquid-vapor (LV)
interface as defined by equation (8)
g,
(°)
The equilibrium spreading pressure measured at the
protein solution-vapor interface and corresponds to
the plateau region of g, = f(Ceq), as defined by
equation (10)
(mJ/m2)
.
no
= kB T / a, (mJ/m2).
n
A generic symbol used to designate a mathematical
relationship among the pertinent variables.
w
The flexibility parameter of the polymer chain
(0
Lc. w <
A Macroscopic Model for Apparent Protein Adsorption
Equilibrium at Hydrophobic Solid-Water Interfaces
1. INTRODUCTION
Protein adsorption is involved in a number of areas in
biology, medicine, food and pharmaceutical processing, and
biotechnology.
In the food and pharmaceutical industries,
proteins can play a major role in the fouling of membrane
surfaces used in biomolecular fractionation, and fouling of
heat exchange surfaces due to their heat sensitivity and
high content in some fluid foods.
Additionally, protein
behavior at both air/water and oil/water interfaces can play
a major role in stabilizing colloidal food systems, foams,
and emulsions.
In biomedicine, protein adsorption is of great concern;
deciphering the mechanism of plasma protein interactions
with blood-contacting devices and the subsequent activation
of coagulation pathways and platelet adhesion is the key
problem in developing nonthrombogenic biomaterials
(Winterton et al., 1986; Woodhouse et al., 1992).
Surfaces
in the mouth like tooth enamel, the mucosa, and the gum are
exposed to a variety of proteins, which are the normal
2
constituents of salivary secretions.
These salivary
proteins are thought to mediate bacterial colonization which
can then initiate dental and gum diseases (Martensson et
al., 1993).
Another area of interest in biomedicine is the
interaction of water-soluble, crystalline proteins of the
eye lens with plasma membrane.
Recent studies suggest that
all three crystallines (a-, /3 -, and 7-crystallins) may
become partially associated with the plasma membrane during
aging and cataractogenesis (Matsuno et al., 1991).
A great deal of effort has been devoted to studying
the different factors that influence adsorption.
The
question of how these factors interact is undoubtedly
complex, and a comprehensive model of protein adsorption is
not available.
The important factors affecting adsorption
can be classified under one or more of the following three
areas: 1.
protein characteristics: including isoelectric
point, net charge and charge distribution, 3-D structure in
solution, placement and nature of hydrophobic patches, and
conformational variability; 2.
surface properties:
including topography and heterogeneity, electrical
potential, composition, water binding, and hydrophobicity;
and
3.
medium conditions: including pH, temperature, ionic
strength, equilibrium concentration, hydrodynamics, and
buffer type.
The objective of this Ph.D. research was to selectively
incorporate some of the above-mentioned factors into a
3
macroscopic model that would quantitatively describe protein
adsorption equilibrium (i.e., predict adsorption isotherms)
for a given protein-surface-medium system.
A two-step
adsorption mechanism was constructed to visualize protein
adsorption equilibrium.
The first step is reversible
migration of the native protein molecule to the interface.
The second step is unfolding of the adsorbed molecule.
The
energies of the two steps were quantified according to the
Gibbs free energy changes for each.
The model served to
explain the Langmuir-type pattern of adsorption equilibrium
isotherms commonly observed.
Its applicability was tested
by comparison with experimentally-measured isotherms
describing protein interaction with hydrophobic solid
surfaces.
4
2. LITERATURE REVIEW
The following review provides a basis for understanding
the theoretical development used to construct the model
presented in this dissertation.
Emphasis is placed on basic
protein biophysics, and the importance of solid surface
hydrophobicity and protein thermodynamic stability on
adsorption.
2.1 PROTEINS
Proteins are biological macromolecules synthesized in
cells for specific functions.
They are high molecular
weight polyamides that adopt exquisitely complex structures.
This complexity is characterized by different levels of
structure:
primary, secondary, tertiary, and quaternary.
Primary structure (Stryer, 1988) refers to the amino acid
sequence itself, along with the location of disulfide bonds
(i.e., covalent connections between two amino acid residues
within the protein molecule). Secondary structure refers to
the spatial arrangement of amino acid residues that are near
one another in the linear sequence.
Alpha-helices and beta-
sheets are typical examples of secondary structure.
5
Tertiary structure refers to the spatial arrangement of
amino acid residues that are far apart in the linear
sequence.
If a protein has two or more polypeptide chains,
each with its exclusive primary, secondary, and tertiary
structure, such chains can associate to form a multi-chain
quaternary structure.
Hence, a quaternary structure refers
to the spatial arrangement of such subunits and their
interaction.
2.1.1 PROTEIN STABILITY
Protein molecules are stabilized by different
intramolecular forces that play a key role in maintaining
protein structure.
In addition to the planar peptide bond
that constitutes the backbone of the molecule and the
possible presence of disulfide linkages, there are
intramolecular forces, though smaller in magnitude than a
covalent bond, that are just as important.
These
intramolecular interactions include the so-called
hydrophobic bonds, hydrogen bonds, and dispersion and
electrostatic attractive forces.
Disulfide bonds are
thought (Fasman, 1989) to stabilize proteins by reducing the
conformational entropy of the unfolded chain.
Statistical
treatments have proposed (Fasman, 1989) that the
destabilization of the unfolded state depends on the length
6
of the loop formed by a single cross link.
Creighton (1983)
found that for a given loop size, the most effective cross
link may be found between groups that are rigidly held in an
optimum orientation by the folded structure.
Hydrogen bonding forces are considered to be one of the
major contributors to the largely temperature-independent
part of the enthalpy of stabilization.
Due to their small
size and electropositivity in covalent bonds, hydrogen atoms
are easily brought into close proximity to electronegative
atoms.
The resulting interaction energy is intermediate
between the energies of van der Waals contacts and covalent
bonds.
Hydrogen bonds are very common in proteins and are
partly responsible for the a-helix and fl-sheet stabilities.
Hydrogen bond partners are exchanged during folding.
Intramolecular bonds are formed at the expense of
intermolecular hydrogen bonds with water.
The importance of dispersion or van der Waals forces in
protein stability hinges on differences in packing in the
folded and unfolded states (Fasman, 1989).
Klapper (1971)
and Chothia (1975) found that, upon folding, water is
expelled to the relatively open bulk phase, and the atoms
that form the protein core become as tightly packed as good
molecular crystals.
The difference in packing density
between the folded and unfolded states is expected to alter
the distribution of interatomic distances, which in turn may
affect the van der Waals interactions.
7
Proteins are polyelectrolytes since ionizable groups
from amino acid side chains and terminal amino acids can
participate in an acid-base equilibrium with the solvent.
Ionizable groups are not generally distributed randomly over
protein surfaces, reflecting their individual structural and
functional roles.
The charged groups in proteins (Barlow &
Thornton, 1983) are, on the average, surrounded by those of
opposite sign, and significantly more oppositely-charged
than like-charged groups are separated by a distance less
than 4.0 A.
This suggests that the charges, including the
a-helix dipoles, contribute to the stability of the protein.
It also suggests that electrostatic interactions are heavily
involved in short-range interactions (i.e.,
less than the
van der Waals interatomic distances).
The problem of how
much energy the charge interactions actually contribute to
the protein is a more complex issue (Thornton, 1982).
Hydrophobic interactions (Andrade, 1985) are basically
entropically driven, largely due to order/disorder phenomena
in the surrounding water.
Current estimates of amino acid
hydrophobicity are based on the measured free energies of
transferring side chains from water to organic solvents,
where the latter presumably simulate the polarity of the
protein interior.
The following subsection is dedicated to
explain in a more detail the concept of those hydrophobic
interactions and why they are thought to be entropically
driven.
8
2.1.2 HYDROPHOBIC INTERACTIONS
To illustrate the notion of hydrophobic interactions
being entropically driven and the idea of measuring the
degree of hydrophobicity of an amino acid residue by its
free energy of transfer from water to a hydrocarbon solvent,
AGuandu, the following development from Schulz and Schirmer
(1979) is presented here.
If N denotes the native state of a protein chain and U
denotes the randomly unfolded state of that chain, and
considering a composite system made of polypeptide chain and
solvent, the total free energy difference between forms N
and U becomes:
-RT1nK=-RT1n[N]/[U] =
OG,
=
+ AHNent - ThiSchain - TAS,www
Furthermore, suppose the unfolding/folding mechanism of a
polypeptide chain in water can be simulated by the phase
separation of a hydrocarbon (mineral oil) from water.
This
is accomplished by assigning the U state to the monodisperse
solution of oil in water, and the N state to separated
phases i.e., an oil drop on the water surface.
ASoil (SN -
Su) is negative because a monodisperse solution (U) is less
ordered than separated phases (N).
AHoil (HN - Hu) is
positive because in the N-state most oil molecules are
surrounded by their kind, whereas in the U-state all of them
are surrounded by water molecules.
The interactions between
9
oil molecules are only dispersion forces and hence weak.
On
the other hand, the interactions between oil and water
molecules are stronger because the strongly polar water
molecules induce dipoles in the neighboring oil molecules,
giving rise to an appreciable electrostatic term.
So AHD
favors the monodisperse solution, nevertheless, it is
relatively small.
It was experimentally shown (Schulz and Schirmer, 1979;
Kauzmann, 1959) that AS,,,,t (SN - Su), which is water in the
above example, is positive which favors the N-state (i.e.,
phase separation).
This indicates that the order of the
water molecules in the U-state (monodisperse oil solution)
is higher than that in the N-state.
Therefore, water
molecules surrounding an apolar molecule are characterized
by a higher degree of order at this polar/apolar interface,
where they assume a locally ordered, quasisolid structure (a
"cage-like" structure, clathrate, or iceberg structure) with
some loss of H-bonding capacity.
This also results in a
negative
All,,t,
the magnitude of which, however, is
relatively small.
In summary, AHD and ASS favor the U-
state whereas AHNe, and AS,,,Nua favor the N-state.
As phase
separation between oil and water is thermodynamically more
stable than the monodisperse case, it turns out that AS,,,Neat
is the predominant driving force that underlies the process
10
of phase separation in this case.
The effect of AS,,,b,, is
usually referred to as a hydrophobic or entropic effect.
Although an analogy has been drawn between a
polypeptide chain and a hydrocarbon, one should keep in mind
that unlike an oil, the polypeptide chain contains both
polar and nonpolar moieties.
But the probability of finding
polar groups on the exterior of a protein molecule is larger
than that in the interior of protein.
Moreover, the
nonpolar amino acid residues are more likely to exist in the
interior of the protein (in globular proteins, in
particular) rather than on the exterior (Andrade, 1985;
Kauzmann, 1959; Schulz and Schirmer, 1979).
This is
consistent with the thought that entropically-driven,
hydrophobic interactions tend to minimize the free energy of
the system by reducing the interfacial area between the
nonpolar moieties and the aqueous medium.
2.1.3 PROTEIN CONFORMATION
When a hydrophobic surface like that of air or a
hydrophobic solid is brought into contact with a protein
solution, an environment supporting unfolding of the intact,
hydrophobic core of a protein to establish new noncovalent
contacts with the interface is created.
Unfolding at the
interface is often referred to as surface denaturation.
11
Protein denaturation involves a conformational change.
Upon unfolding, the polypeptide chain becomes less compact
and more highly solvated, and much more flexible.
Protein
denaturation is a highly cooperative reaction, and general
molecular stability depends on environmental conditions such
as temperature, pressure, pH, ionic strength, and the
concentration of specific ligands, stabilizers, and
denaturants in solution.
Dill et al.
(1989) studied the thermal stability of
globular proteins.
Two factors are important in quantifying
the temperature dependence of globular protein folding: 1)
the conformational entropy of the chain; and 2) the heat
capacity change effected by the hydrophobic effect.
Folding
is driven by a negative free energy change accompanying
clustering of the hydrophobic residues into a globular
structure and is opposed by a positive free energy change
due to loss of conformational entropy upon folding.
Cold
denaturation is driven principally by the weakening of the
hydrophobic interactions, but thermal denaturation is driven
principally by the gain of conformational entropy in the
unfolded chain.
Privalov et al.
myoglobin.
(1986) studied cold denaturation of
The disruption of the native protein structure
both on cooling and on heating was characterized as
proceeding in an "all-or-none" manner, with a significant
and similar increase of the protein heat capacity, but with
12
inverse enthalpic and entropic effects: the enthalpy and
entropy of the protein molecule decrease during
cold-denaturation and increase during heat - denaturation.
Concerning proteins with a multi-domain structure, such
domains may act independently during thermal denaturation,
but with some degree of cooperativity among the different
subunits (Privalov and Medved, 1982; Koteliansky et al.,
1981).
Honeycutt and Thirumalai (1992) demonstrated the
existence of metastable states in the folding/unfolding
pathways, using a stochastic dynamics method (Honeycutt and
Thirumalai, 1992) to simulate the processes of folding and
unfolding.
These metastable states are characterized by
several free energy minima separated by barriers of various
heights such that the folded conformations of a polypeptide
chain in each of the minima have similar structural
characterisitics (namely, the gross appearance and radius of
gyration) but have different energies from one another.
They suggested that the formation of the more stable form
depends largely on the method of preparation; the initial
conditions determine the kinetics of formation of the more
stable form.
The lesson that can be learned from their
study is that although the unfolding or folding reaction may
be thermodynamically favorable, energy barriers exist along
the reaction coordinate, which hinder the protein from
13
reaching a configuration with the lowest free energy,
instead, "trapping" it into a metastable state.
2.2 PROTEIN ADSORPTION
In general, adsorption involves migration of a
substance from one phase to the surface of an adjacent
phase, accompanied by its accumulation at the interface
(Slejko, 1985).
Adsorption is a result of the binding
forces between individual atoms, ions or molecular regions
of an adsorbate and the adsorbent surface.
These binding
forces or interactions vary in magnitude from the weak van
der Waals-type of attraction contributing to physical
adsorption, to the strong covalent bonds in chemisorption.
Polymer adsorption in general and biopolymer adsorption in
particular show a range of binding energies depending on the
type of forces present in the interface.
Polymer adsorption
differs drastically from that of small molecules.
This is
basically due to the large number of conformations that a
macromolecule can adopt, both in the bulk solution and at
the interface.
Moreover, the entropy loss or gain
associated with a given flexible polymer can be
substantially greater than that for small molecules or
relatively stiff molecules (Parfitt & Rochester, 1983).
14
A thermodynamic approach was proposed by De Feijter et
al. (1987) to describe the adsorption of nonionic, flexible
polymers at solid surfaces.
Their approach relied on a
pseudo-lattice model (quasi-crystalline model), the cells of
which may accommodate a solvent molecule,
segment.
or a polymer
Each macromolecule was considered to consist of m
identical segments of which a fraction, f is adsorbed
directly to the surface (i.e., f m cells of the surface
layer are occupied by one adsorbed polymer).
Their approach
led them to conclude that the polymer adsorption isotherm
would exhibit a high affinity character; i.e., high
adsorption at very low bulk concentration, with almost
immediate plateau attainment.
Their approach was also
expected to apply to proteins, even though proteins are
considered to be somewhat rigid structures, exhibiting some
net charge.
Their conclusion may explain in part, however,
why protein isotherms generally assume plateau values at
relatively low concentration.
Lin et al. (1991) studied adsorption-desorption
isotherm hysteresis exhibited by 0-lactoglobulin A on a
weakly hydrophobic surface.
They found that the desorption
isotherm at pH 6.0 overlapped with the adsorption isotherm,
and the adsorption-desorption process of 0-lactoglobulin A
under this condition could be characterized by a fully
reversible Langmuir model.
The desorption isotherm at pH
4.5, however, did not coincide with the adsorption isotherm,
15
giving rise to hysteresis.
This would suggest that protein
adsorption experiments carried out under mild conditions of
pH at relatively hydrophilic surfaces may be treated with
the assumption that reversible equilibrium exists between
the bulk and interface.
Arnebrant et al.
(1987) studied the temperature
dependence of adsorption for a-lactalbumin and
0-lactoglobulin on chromium surfaces.
They observed that
the curves for 0-lactoglobulin at 25, 66, 70, and 73°C were
rather similar.
It was only when the temperature exceeded
the denaturation temperature (79°C) of /3- lactoglobulin, that
they could observe a significant difference in the isotherm.
This would suggest that the adsorbed mass of protein does
not significantly depend on temperature, as long as the
temperature at which the adsorption experiment was carried
out lies below the melting point temperature of the protein
(or the irreversible denaturation temperature.
If the
temperature exceeds that of the melting point of protein,
the surface activity of protein will depart from that
exhibited in its native form.
Lu et al. (1991) calculated the solvation interaction
energies for protein adsorption on hydrophilic and
hydrophobic polymer surfaces.
The solvation interactions
(repulsive hydration and attractive hydrophobic
interactions) were calculated for lysozyme, trypsin,
immunoglobulin Fab (antigen-binding fragment consisting of
16
the light chain and half of the heavy chain, with a
molecular weight of 50,000 daltons), and hemoglobin.
The
average solvation interaction energy was found to vary from
-259.1 to -74.1 kJ/mole for the four proteins at hydrophobic
polymer interfaces (polystyrene, polyethylene, and
polypropylene), whereas on hydrophilic surfaces
(poly(hydroxyethyl methacrylate) and poly(vinyl alcohol))
the average solvation interaction energies were greater than
zero.
These calculations illustrate the importance of
attractive, hydrophobic interactions between proteins and
polymer surfaces in adsorption.
Matsuno et al. (1991) studied the interactions of 7­
crystallins with silica, methylated silica, and diphenyl
silica.
They used different techniques to examine the
secondary and tertiary structural alterations that took
place upon adsorption on these silica surfaces exhibiting
different degrees of hydrophobicity.
Based on a comparison
between conformations of free and surface-bound protein as a
function of electrostatic and hydrophobic character of both
the protein and the adsorbent surface, they demonstrated
that: 1) protein destabilization on hydrophobic surfaces is
greater than that on more hydrophilic surfaces; 2)
detectable conformational changes tend to increase as the
surface hydrophobicity increases; and 3) subtle structural
differences among proteins can play an important role in
17
determining differences in protein stability and structure
upon adsorption.
Wei et al. (1991) examined the role of protein
structure in surface tension kinetics at the air-water
interface and demonstrated that the intrinsic,
conformational stability is an influencing factor in protein
surface activity at low bulk concentrations.
At high bulk
concentrations, surface hydrophobicity was highly correlated
with the observed surface tension kinetics.
Surface tension
kinetics in this context refers to the rate of change of
liquid surface tension, yLv, for the protein solution in
contact with air.
Kato and Yutani (1988) correlated the surface activity
of six mutants of tryptophan synthase a-subunits with their
stability, as measured by their free energy of denaturation
in water (411.77ma).
One measure of surface activity was given
by the air-liquid surface tension of a mutant solution.
These results are in line with the previous examples and
demonstrate the importance of conformational stability in
predicting the difference in the adsorptive behavior among
proteins.
Incidentally, it serves as another example
suggesting that the surface tension of protein, 7pv, is
related to its intrinsic conformational stability.
Horsley et al. (1987) studied human and hen lysozyme
adsorption on hydrophobic, negatively-charged, and
positively-charged silica.
On the average, human lysozyme
18
was found to adsorb in larger amounts than did hen lysozyme.
They attributed the difference in adsorptive behavior to the
thermal lability of the molecules; human lysozyme is more
susceptible to thermal denaturation, hence to surface
denaturation, than is hen lysozyme.
Hunter et al. (1991) studied the coadsorption and
exchange of lysozyme and 0-casein at the air-water
interface.
The air-water interface was used because it was
described as representing the simplest model hydrophobic
surface for studying protein adsorption.
Their results
suggested that electrostatic interactions do no play a major
role in determining exchange behavior at the air-water
interface; moreover that the flexibility of both the
adsorbed and displacing molecules are more important than
intermolecular interactions in determining whether exchange
occurs.
This work serves as an example of the general
observation that electrostatic interactions play a minor
role in dictating the adsorptive behavior of proteins at
hydrophobic interfaces.
In summary, then, the following general statements can
be made regarding protein adsorption.
The surface activity
of a protein is a cumulative property influenced by many
factors, including size, shape, charge, surface
hydrophobicity, and thermodynamic stability.
However, based
on abundant experimental observations, a sample of which
were cited here, the influence of only two of these factors
19
(surface hydrophobicity and conformational stability)
can be
considered, under controlled but actually quite relevant
circumstances, as effectively governing the adsorption
process.
Protein adsorption exhibits a diversity in
behavior from one surface to another and from one protein to
another.
This diversity results from the complexity of the
protein structure itself and from the many variables on
which protein adsorption depends.
A comprehensive model for
any aspect of protein adsorption that takes all these
variables into account is completely lacking in the existing
literature and much work is needed to reach that goal.
20
3. MODEL DEVELOPMENT
3.1 DIMENSIONAL ANALYSIS
The useful result of a dimensional analysis of protein
adsorption equilibrium would be a starting point for
determination of a functional relationship among the
variables thought to be pertinent to the process.
The
relationship can be expressed in the following compact form:
(11111:12/1131114/ '
ni) =
0.0
(1)
where the A represent independent dimensionless groups of
some dimensional variables or parameters (factors) which are
measurable or can be expressed in terms of other measurable
quantities, and Si defines this mathematical relationship
among the dimensionless II groups.
There are two matters involving choice in this
analysis.
The first is that of initial quantities (in the
case of protein adsorption, this refers to factors
influencing the process of adsorption).
This choice
identifies the factors considered to be important, and those
that may be neglected.
The second is the choice of the
final, dimensionless II groups.
The first choice requires a thorough understanding of
protein adsorption and a comprehensive survey of the
21
pertinent literature.
Consequently, a sufficient portion of
the coherent literature was reviewed and the following
relationship is suggested:
0 (
We
*A
c ,
r * Ap
A Gunfold
,
VP *
Ceg )
=
0.0
(2)
where
r
= adsorbed amount of protein
(mol/m2)
,
CN
= apparent equilibrium concentration
(mol/m3)
,
Ap
= partial molar area occupied by protein
at the interface
V,
(m2/mol)
,
= partial molar volume of protein in solution
(re/mol),
AG"tbid
= change in partial molar free energy of protein
upon unfolding
Ac
(J/mol),
= minimum surface area cleared by an adsorbing
protein molecule in order to anchor itself to
the interface
(m2/moi) ;
and
W,
= work of adhesion
The published results that led to selection of the
factors incorporated into Eq.
of this dissertation.
(2) are detailed in section 2
22
3.1.1 QUANTIFYING THE FACTORS IN EQUATION (2)
Work of Adhesion, W,
The work of adhesion is generally defined as the energy
required to separate a unit area of interface into two
phases.
Applied to protein adsorption, W, can be written as
follows:
W8
Y SW + Y PW
Y Ps
(3)
where yww is the interfacial free energy between solid and
water, 7rw is the interfacial free energy between protein
and water; and ws is the interfacial free energy between
protein and solid.
These three interfacial energies can each be defined by
an equation expressed in the following compact form:
a (Y Pill Y SV)
(4)
Y PW = 11 (Y PW Y WV)
(5)
Y sw = 0 (Y sv, Y wv)
(6)
Y PS =
where the functional relationship, n, represents an equation
of state that relates the interfacial energy between two
23
phases to the interfacial energies between each of those
phases and a third phase; in the case of Eqs. (4)-(6), the
third phase is vapor.
Neumann et al. (1974) empirically
obtained an explicit formulation for II:
(FY17.3
Y12
SrY7) 2
1
1
0 . 015
(7)
(Y13Y23) 2
where subscripts 1 and 2 denote any pair combination of two
phases from the protein (P), water (W), and solid (S)
phases, as written in Eqs. (4)-(6), and subscript 3 denotes
the third phase, which is vapor in all of the above cases.
Young's equation (1'sv = Tn.
cos 0) introduces 0
(1, the equilibrium contact angle formed between a solid-
liquid (SL) interface and a liquid-vapor (LV) interface.
The contact angle 0 defines the characteristic orientation
Figure 3.1.1 The equilibrium contact angle, 0, formed
between a solid/liquid (SL) interface and a liquid/vapor
(LV) interface.
24
of a liquid-vapor interface with reference to a solid
surface it contacts, as illustrated in Fig. 3.1.1.
equation can be combined with Eq.
Young's
(7) to yield, for a
specific solid/liquid contact:
1
cos 0
(0 . 015ysv-2 .00) (YsvYLO 2 +YLV
(8)
1
Y 1,1,[0 . 015 (YsvYLO 2 -1]
Eq.
(8) should be taken as generic; i.e., subscript S refers
to any solid phase, including protein, that is in contact
with a liquid phase.
Eq.
(8) therefore allows evaluation of
both -ysv and 7", as both 0 and -yLv are readily measurable.
Thus, Eq.
(7) should be solvable for the interfacial
energies of Eqs. (4)-(6).
However, evaluation of the surface energy of pure
protein with contact angle methods is not trivial.
It is
arguably impossible to determine a relevant value of 7" by
measuring contact angles on a prepared, solid protein
"surface".
For example, liquid penetration into a solid
protein surface would probably be unavoidable, as such a
surface would probably be both porous and hygroscopic.
In
this work, the value of -ypv was approximated by the
following equation:
YPV = Y LV
7C e
(9)
25
where ire (mJ/m2) represents the equilibrium spreading
pressure measured at the protein solution-vapor interface
and corresponds to the (concentration-independent) plateau
region of ve = f(Cat), i.e., a saturated interface, and
ryLv
represents the surface tension of the protein-free solution.
Spreading pressure is a measure of the reduction in surface
energy as a result of adsorption at an interface.
Eq.
(9) should give a less ambiguous value of -ypv than
that attainable by contact angle methods.
van Oss and Good
(1988) experimentally determined protein interfacial energy
using contact angle methods.
They concentrated a protein
solution by ultrafiltration on an anisotropic cellulose
acetate membrane, then carried out contact angle
measurements on the resultant protein layer.
Depending on
the degree of hydration of the resultant protein layer, the
value of 'ypv was determined.
Therefore, the value of lfpv
obtained, in essence, represented the surface tension of a
hydrated protein layer (formed in this case on a cellulose
acetate membrane) measured at the protein layer-air
interface and that value of -ypv is thus consistent in
principle with that given by Eq.
(9).
The value of 'ypv obtained by contact angle methods,
however, is subject to perturbations induced by the solid
support itself, and it rather reflects the degree of
orientation of the water molecules at the periphery of the
hydrated protein layer (van Oss and Good, 1988).
26
Unfortunately, relevant spreading pressure data is not
available for many proteins of interest, so another means to
calculate 7r, is needed.
Singer (1948) developed an equation of state that
relates Ire to measurable parameters:
n e = n 0 ( Zln(1
2
f
)-1n(1
f) )
(10)
where 1r0 = kB T / a, (kB is the Boltzmann constant; T is
temperature and a, is the average interfacial area occupied
by an amino acid residue); f is the fractional surface
coverage; and Z is the surface coordination number of the
lattice (Z = 2 + w where w refers to the flexibility of the
polymer chain).
For a completely rigid chain Z = 2, and for
a completely flexible chain Z = 4, thus 2 S Z < 4 (or 0
w < 2).
The value assigned to the parameter a, is 15 A2 per
amino acid residue (Ter-Minassian-Saraga, 1981; Damodaran
and Song, 1988).
Parameter Z can be correlated with the
thermal stability of the protein molecule (AGmmd) using
surface pressure data available for a-lactalbumin, /3­
lactoglobulin, bovine serum albumin (Suttiprasit et al.,
1992), and lysozyme (Uraizee and Narsimhan, 1991).
The resulting equation is:
Z = 2.0
+2.0
* exp( -0.135 A Gunfold
RT
27
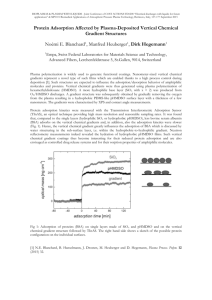
Figure 3.1.2 shows the value of Z as given by Eq.
(11)
compared to individual values of Z estimated by Eq.
the above-mentioned proteins.
(10) for
The value of fractional
surface coverage, f, should be fixed at a value equivalent
to monolayer film coverage.
As a result of the mathematical
difficulty that arises in Eq.
(10) when f approaches 1.0,
however, the value of f was set equal to the maximum
possible value such that Eq. (10) is sensitive to variations
in parameter Z.
Hence, a value of 0.92 was chosen by
equating the experimentally-based plateau value of
equilibrium spreading pressure, ire, with Eq.
(10).
4.0
aLac
3.8
3.6
Z / Spread. Pressure Data
Z /Eq. (11)
3.4
3.2
N
3.0
2.8
2.6
2.4
BSA
2.2
Lyso
a Lag
2.0
i
0
5
10
15
20
25
30
AGunfold / RT
Figure 3.1.2 The flexibility parameter (Z) correlated
with AGunfoki/RT for a-Lac, /3 -Lag, BSA, and Lyso.
28
A question may arise regarding the decision to define
protein surface energy (7pv) in terms of the hydrophobicity
of its intact core (expressed by AGm0, rather than in
terms of its effective "surface" hydrophobicity.
The
surface hydrophobicity of native protein, expressed
as -ypv
but measured in aqueous solution by a number of techniques
including contact angle, cell adhesion (in the absence of
electrostatic effects), and as derived from adsorption
experiments, is found to change only slightly from one type
of protein to another (van Oss et al., 1981; Neumann et al.,
1983).
Thus, 7pv was estimated using Eq.
(9), considered to
yield a better index of protein surface activity, and being
in line with the suggestion that a protein with a higher
tendency to unfold should exhibit higher surface activity
(i.e., greater reduction of the air-water surface tension
for a given protein solution (Suttiprasit et al., 1992).
Protein/vapor interfacial energy is thus measurable
with equations (9) through (11), and solid surface energy by
equation (8).
In practice, however, as the surface energy
of the solid in question approaches that of water, Eq.
yields anomalous results.
(7)
A set of equations of state was
developed by Neumann et al. (1980) that allows calculation
of interfacial energy under such circumstances.
Appendix 1
includes the Fortran program (Neumann et al., 1980) used in
this work to determine interfacial energy based on the
values of pertinent, input solid and liquid interfacial
energies.
29
The Gibbs Free Energy of Unfolding, AGmfom
The Gibbs free energy of unfolding, AGmfold, connotes the
partitioning of a protein molecule between two
conformations: native and unfolded.
AGmftm measures the
difference between the partial molar free energy of the
macromolecule in its unfolded or denatured state (U), and
its native, folded state (N).
AGfoid = GU
(12)
GN
This two-state model for folding and unfolding has been used
by a number of investigators (Tanford,
1970; Privalov &
Khechinashvili, 1974; Privalov, 1979; Velicelebi
&
Sturtevent, 1979; Becktel & Schellman, 1987) to study the
thermal stability of proteins in order to measure what are
known as protein stability curves.
It should be mentioned,
however, that Eq. (12) was restricted to one class of
proteins in this treatment.
This class includes compact,
single-domain, globular proteins undergoing a one-step (all­
or-none) reversible phase transition between two
thermodynamically defined conformations (native and
unfolded).
Privalov (1976) suggested that in a living
system the transition from the structureless state (U) to
the native state (N) should be reversible; the observed
irreversibility is caused by secondary phenomena including
aggregation, isomerization of prolyl residues, and
separation of chains, but the transition itself is in
principle reversible.
Dzakula and Andjus (1991) have
30
recently improved the accuracy of the two-state model by
expressing AGmfmd as a function of temperature. Their
proposed equation for AGmemd is as follows:
AHD(Tm
A Gunfold
T)
ACp(T
Tm
Tm) 2
2 Tm
(13)
where AHD (kJ/mole) is the enthalpy of denaturation
evaluated at the melting point temperature, Tm; and ACp
(kJ/mole
K) is the difference in heat capacity between the
unfolded and folded states.
These parameters are
conveniently determined by conducting a differential
scanning calorimetry (DSC) analysis of the given protein.
The melting point temperature, Tm (K), is estimated from the
temperature at which the thermogram peak reaches its
maximum, provided that the rate of heating is sufficiently
low.
AHD is estimated as being equal to the area of the
endotherm peak, and ACp is viewed as the difference in the
asymptotic heat capacity values on either side of the
endotherm peak (Dzakula and Andjus, 1991).
In the absence
of DSC data, an equation of state developed by Murphy et al.
(1990) can be used to define AGmw (per mole of amino acid
residue), as a function of temperature:
A Gunfold =
H*-TAS*-EACp[(T-TH)-T
)
(14)
where AH* (J/mole amino acid residue) and TAS* (J/mole amino
acid residue) represent the non-hydrophobic, enthalpic and
entropic contributions to the free energy change,
31
respectively; TH and Ts are the temperatures at which AH is
equal to AH* and AS equal to AS*, respectively; and ACp is
the heat capacity change upon unfolding.
Murphy et al.
According to
(1990), the heat capacity term on the right
hand side of Eq. (14) represents the hydrophobic,
destabilizing contribution to the free energy of folded
proteins.
TH and Ts are approximately equal for proteins
and they assume a value of 112 ± 2.4 °C (Murphy et al.,
1990).
AS* for proteins was found to be equal to 18.1 ± 1.0
J/(mole amino acid residue
K) (Murphy et al., 1990).
A
typical value for ACp of 50.0 ± 10.0 J/(mole amino acid
residue
K) can be assumed, based on experimental
measurements of changes in heat capacity upon denaturation
(Zhukovskii and Rovnov, 1989). Equating experimentally-based
values of AGmfom obtained from Eq.
(13) with Eq.
(14) allowed
estimation of a protein-independent AH* (J/mole amino acid
residue).
Appendix 2 shows results of that procedure for
nine proteins considered in this work; an average value of
6.076 ± 0.155 kJ/(mole amino acid residue) was obtained for
The Partial Molar Area of Protein, Ap
Since estimates of the partial molar volume of protein
in solution, Vp, are readily available (Sober,
1970; Pilz &
Czerwenka, 1973; Bull & Breese, 1973; Fasman, 1976;
Durchschlag, 1986;
Bendzko et al., 1988) only the
32
difference between the minimum surface area cleared by an
adsorbing molecule, Ac, and the partial molar area, AP, need
be discussed in support of Eq.
(2).
Based on experimental
studies (Andrade, 1985; Macritchie, 1978; Ter-Minassian-
Saraga, 1981) on protein adsorption at air-water and solid-
water interfaces, Ac changes only slightly from one type of
protein to another, and is independent of molecular size.
Moreover, there is no equation of state available that
relates Ac to any surface or protein property.
For these
reasons, Ac was allowed to be a computer-generated
parameter, rather than an input variable for the model of
Eq.
(2).
It was anticipated that this parameter would not
dramatically change from one type of protein to another,
however, in accordance with previous experimental
observations.
Ap is the partial molar area of protein, and was
evaluated such that protein molecules in the neighborhood of
the interface are spherical and hexagonally close-packed.
The following equation was originally suggested (Dabrowski
et al., 1987; Suri, 1970) with reference to adsorption from
simple organic mixtures on solid surfaces, provided that
molecules in solution exist in a closest-hexagonal packing:
Ai =1.091(Dri(ArA)113
(15)
where Ai (m2/mole) is the partial molar area of species i at
the solid/liquid interface, V, (m3/mole) is the molar volume
of species i in solution at the given temperature, and NA is
33
Avogadro's number.
The suggestion that we can apply the
same equation to protein adsorption would need some
justification.
One important point refers to our confining
the applicability of the two-state model of folding and
unfolding to compact, single-domain, globular proteins.
This can be considered as a sufficient condition for
fulfilling one criterion of Eq. (15), that being the
existence of spherically-shaped molecules in solution.
Whether proteins in solution can exhibit a closest-hexagonal
type of packing is an important matter as well.
Regarding
the nature of the protein interior, Klapper (1971)
demonstrated that on the average, the ratio of the volume
occupied by all atoms making up a protein to the total
volume occupied by the protein molecule itself (E) is about
0.747, compared to a E value of about 0.765 for a typical
compound assuming a closest hexagonal packing of identical
spheres.
Water assumes a E value of 0.363, whereas other
organic solvents, e.g., cyclohexane and carbon
tetrachloride, assume E values of 0.438.
With that, Eq.(15)
is considered applicable to protein in solution, so that it
becomes:
Ap = 1. 091 ( Vp)
2/3 (NA) 1/3
(16)
34
Even if we consider a protein that is spherical in
shape and exists in closest-hexagonal packing, deviation
from Eq.
(16) should be expected, depending on the degree of
surface unfolding encountered, and Eq.
(16) provides only an
estimate for the surface area of protein at an interface.
It is worth mentioning here, though, that the value of
Ap predicted by Eq. (16) was found to correlate to a large
extent with the corresponding close-packed, "end-on"
adsorption plateau value experimentally observed.
As a
qualitative rule in protein adsorption studies (Andrade,
1985), a protein monolayer with an end-on conformation
presumably prevails when adsorption occurs from solution at
high concentration, whereas a "side-on" conformation­
monolayer is observed at low protein concentration.
Although practically it is quite impossible to draw a line
between low- and high-concentration regions, the
concentration interval between Cal = 0.1 mg/ml and Ceci = 1.0
mg/m1 may be suggested as the diffuse line between low- and
high-concentration regions as far as side-on and end-on
conformations are concerned (Wei et al., 1990).
Using non­
linear regression as a tool to minimize the difference
between the regressed value of Ap as given by an equation
similar to Eq. (16), and that calculated from the known
geometry of a given protein (Appendix 3), the following
equation was developed to predict the value of Ap for a
protein monolayer with a side-on conformation:
35
Ap= 2.717(Vp)2/3(
(17)
In any case, descriptions of the nature of proteins and/or
their behavior in solution being analogous to that of
organic liquids has been encountered in more than one
technical paper (Klapper, 1970; Tanford, 1970; Creighton,
1983; Baldwin, 1986; Fersht, 1987; Murphy, 1990; Nicholls,
1991) .
The final step in this dimensional analysis involves
the arrangement of the dimensionless II groups, requiring an
awareness of the physical laws that underlie the adsorption
process.
The physical meaning of each term and how it
influences the adsorption process can be visualized with a
suitable adsorption mechanism.
3.2 A SIMPLE MECHANISM FOR PROTEIN
ADSORPTION
In general, the pattern of protein adsorption
equilibrium isotherms at solid/liquid interfaces assume
either a Langmuir-type or Freundlich-type shape.
The
Langmuir-type model is described by the following equation:
36
r-
r
max
c
(18)
eq
( b + Ceg )
where Ceq (mg/1) is the apparent equilibrium concentration;
r
(Ag/cm2) is the plateau value; and b (mg/1) is a
constant such that r
versus C.
/b is the initial slope of a plot of r
On the other hand, the Freundlich model is
described by the following equation:
r=a
(Ceq) b
(19)
where a and b are function constants that define the
functionality of r versus C.
Consequently, the Langmuir­
type isotherm shows a steep initial slope followed by
attainment of a plateau at high concentration, while the
Freundlich isotherm shows a monotonic increase in r with C.
Upon examining the pertinent literature, one recognizes that
interpretation of data with Langmuir-type isotherms is
extremely popular, even though no real benefit is gained by
determining the function constants in either Eq. (18) or
(19); i.e., they are not related to adsorption affinity in
any clear manner (Martensson et al., 1993; Woodhouse er al.,
1992; Nygren et al., 1992; and Ruzgas et al., 1992).
Thermodynamic equilibrium criteria are implemented here
to analyze the phase equilibrium between the bulk and
interface, and for simplicity the following two conditions
are imposed:
37
1)
Existence of reversible equilibrium between the bulk
phase and the interface; and
2)
Existence of a monolayer coverage of protein as the
upper limit for the extent of adsorption.
Protein adsorption is usually regarded as irreversible,
though, based on experimental observations (particularly at
hydrophobic interfaces).
Arnebrant and Nylander (1986)
found that sequential adsorption on hydrophilic surfaces is
characterized by a larger fraction of reversibly adsorbed
molecules than adsorption on hydrophobic surfaces.
Elwing
et al. (1987) found that fibrinogen is partly exchanged by
7-globulin only on the hydrophilic side of a surface
exhibiting a wettability gradient, whereas on the
hydrophobic side there appeared to be no exchange at all.
Shirahama et al. (1990) found that sequential adsorption on
hydrophilic silica occurs by displacement of preadsorbed
protein, whereas on a hydrophobic surface it is accompanied
by desorption of only a fraction of preadsorbed protein.
It
should be mentioned, however, that in all of these
experimental observations, although pure desorption, in a
strict sense, constitutes a very unlikely event, exchange
reactions between adsorbed and incoming proteins have been
observed to take place.
In a sense, the assumption of
reversible equilibrium serves as a tool to account for the
outcome of exchange reactions that yield an observable,
gross adsorption equilibrium.
38
Moreover, the possibilty that adsorbing protein may
form a multilayered film is quite real.
Although multilayer
formation is the rule at air-water interfaces (Khaiat and
Miller, 1969; De Feijter, 1978; Graham and Phillips, 1979a;
Graham and Phillips, 1979b; Hunter et al., 1990), monolayer
formation is the rule at solid-water interfaces
(Brash and
Lyman, 1969; Lee and Kim, 1974; Morrissey and Stromberg,
1974; Absolom et al., 1987; Jonsson et al., 1987; Lundstrom
et al., 1987; Mizutani and Brash, 1988; Arai and Norde,
1990; Ruzgas et al., 1992; Shibata and Lenhoff,
1992).
In
fact, it is anticipated that study of monolayer adsorption
of protein will shed light on understanding multilayer
formation at air-water interfaces.
Figure (3.2.1) is a schematic of the proposed
adsorption mechanism.
Once a protein molecule arrives at
the interface, it unfolds in an attempt to adapt to the new
microenvironment.
During the course of an adsorption
experiment and at a certain point on the reaction
coordinate, a pseudo, reversible phase equilibrium should
adequately represent the situation existing between the bulk
phase and the interface.
For the purpose of analysis this
state of equilibrium can be resolved into two major sub-
equilibrium states (Figure 3.2.1b):
1) an equilibrium between native protein in the bulk and
that at the hydrophobic interface, the attainment of which
is largely driven by the Gibbs free energy for adsorption
(or the work of adhesion); and
39
2) an equilibrium between adsorbed, native and unfolded
protein.
Attainment of this sub-state of equilibrium is
largely driven by the Gibbs free energy of unfolding.
K
IN
U
---,---,-7,,
, , ,, ,,,
Sur f ace
(a)
,,
-V
\
, , . ,
, , ,
Surf ace
u, D
, ,
, ,
(b)
Figure 3.2.1 A schematic depicting protein adsorption
equilibrium.
(a) the gross, pseudoequilibrium; (b) a two-
step approximation of the equilibrium consisting of
1) reversible arrival and 2) reversible unfolding.
40
In the absence of electrostatic effects on adsorption
and of specific biochemical interactions (e.g., receptor­
ligand) as well, the first sub-equilibrium state should be
effectively characterized by the work of adhesion between
protein and surface.
Absolom et al. (1983) used a
fundamentally similar approach to describe the adhesion of
bacteria to various polymeric low energy surfaces with good
success.
The model proposed here is limited to adsorption
at hydrophobic interfaces, so from a thermodynamic
standpoint, the surface energies of each of the interacting
phases should adequately describe the initial adsorption
event.
For the second sub-equilibrium state, the Gibbs free
energy of unfolding was selected to quantify this process,
involving exposure of the previously intact hydrophobic core
to the aqueous medium. Several investigators (Chothia, 1975;
Baldwin, 1986; Matsumura et al., 1988; Privalov and
Makhatadze, 1990) indicated that the Gibbs free energy of
unfolding correlates well with the surface area of non-polar
groups exposed upon unfolding; that is, the hydrophobic
stabilization is proportional to the reduction of the
surface area accessible to solvent on folding.
Wei et al.
(1990) indicated that, at low bulk concentrations, surface
tension kinetics reflected the conformational stability of
the protein; while at higher concentrations, surface tension
kinetics were more strongly correlated with the effective
hydrophobicity of the protein.
Norde (1992) indicated that,
41
with "rigid" proteins, intramolecular structural
rearrangements do not contribute to the adsorption process;
while, with "soft" proteins, the intramolecular structural
rearrangements result in a significant driving force for
adsorption, that proteins may even adsorb under the adverse
conditions of a hydrophilic, electrostatically repelling
surface.
One important point that should be made is that in
addition to the proposed mechanism of Figure 3.2.1, there
are other mechanisms incorporating arrival, unfolding, and
exchange reactions to describe adsorption equilibrium.
But
the Gibbs free energy is a state function; that is, no
matter what path is chosen to analyze the process, overall,
it is governed by the Gibbs free energy change between the
initial (i.e., native protein in solution) and final state
(i.e., unfolded protein at the interface).
A theoretical model can be developed to express the
function constants that appear in Eq.
(18) in terms of
selected molecular, solution, and surface properties.
As
indicated with reference to Eq. (18), the Langmuir-type
model is the most popular model used in the literature to
describe adsorption equilibrium isotherms at solid-liquid
interfaces.
The Langmuir-type isotherm is characterized by
a (usually steep) initial slope at low concentration
followed by plateau attainment at high concentration.
A
mathematical expression of this statement can be developed
as follows.
42
At low
concentration:
r A « VP Ceq
(20)
where r Ap represents dimensionless adsorbed mass (or
surface coverage); and Vp Cal a dimensionless concentration
(or the volume fraction of protein in solution).
Defining
the relationship in the form of an equation, it becomes:
r = K Vp Ce (A -)
(21)
g
where K, the proportionality constant between dimensionless
adsorbed mass and dimensionless concentration, is the
overall equilibrium constant for the protein adsorption
process.
At high concentration:
r
=
1
)
A;
(22)
If Equations (21) and (22) are combined, the following
equation is obtained which accounts for both regions:
1
A
=
(
K Vp Ceq
)
(23)
Rearranging Equation (23) yields:
r
,
1
1
Ap
K VP Ceg
(24)
43
Finally, the adsorbed mass can be expressed as:
r"
r
V1r
-1
P eg AP
1
(25)
+ VP Cog
As shown in Figure 3.2.1a, the overall equilibrium
constant can be defined as:
K
Lpr to
ads
[pr tn]
bulk
(26)
where the subscript N stands for native conformation and U
for unfolded conformation.
The superscripts indicate that
protein is either adsorbed (ads) or in solution, near the
interface (bulk).
Equation (26) can be written as:
K
UartnWs
[pr tn] Its
[pr tn]
[pr tn] bulk
where the first term on the right hand side of Eq.
(27)
(27)
represents Ku and the second term represents Ka therefore,
the overall equilibrium constant assumes the form:
K = Ka
.
(28)
Ku
Introducing the definition of an equilibrium constant as a
function of the Gibbs standard free energy Gialfor both Ku
and Ka:
K = exP
P Gunf old )
R
walic
exp 7?-7,
(29)
where a is the proportionality constant that relates the
Gibbs free energy of unfolding in the bulk to that at the
44
interface, at specific conditions of temperature, pH, and
ionic strength.
Here, the surface-induced unfolding process
for a given protein molecule is related to its intrinsic
conformational lability (stability) in solution, where the
latter is characterized by AGmd.
Andrade et al.
(1987)
suggested that data on the solution denaturation of proteins
may be important in estimating protein lability (stability)
and, together with information on the surface tension and
interfacial tension behavior of proteins, would help develop
hypotheses and correlations with the actual solid-liquid
interface behavior.
W, is equal to the negative of the
Gibbs standard free energy change per unit area (Hiemenz,
1986) of the reversible arrival step.
Multiplied by Ac, the
quantity VW essentially represents the negative of the
Gibbs standard free energy change for the reversible arrival
step, at constant temperature and pressure.
Finally, plugging the value of K into Eq.
(25), a model
that relates r to the pertinent variables is obtained:
VC
1
P eg AP
r
exp (
Gunfold-WaAC)
RT
(30)
45
3.3 LIMITATIONS ON THE APPLICABILITY OF
THE MODEL (EQ. 30)
The limitations on reliable application of the proposed
model basically stem from the set of conditions associated
with quantifying each term appearing in Eq. (30).
The
physical properties of the protein, solid surface, and
solution, required to reliably define the quantities
appearing in Eq. (30) are outlined here.
1.
Protein properties: compact, single-domain, globular
structure;
2.
Solid surface properties: homogeneous, hydrophobic
surface, not promoting any specific biochemical
interaction; and
3.
Solution properties: pH and ionic strength such that
the effect of electrostatic interactions between the
solid surface and protein on adsorption are minimal.
System temperature must be confined between a minimum
value around room temperature, and a maximum value of
55°C, or the irreversible denaturation temperature for
a given protein, whichever is smaller.
Although most of the protein adsorption isotherms
studied here were constructed at moderate temperatures (18­
27°C), the confinement made concerning the acceptable
temperature range emanates from two points:
46
a.
Such a temperature range is common for study of
proteins in solution in their native conformations; and
b.
The partial specific volume of protein does not
significantly change with temperature within that
range.
Bull and Breese (1973) studied the temperature dependence of
partial volumes of proteins and found that there exists a
temperature range from 25°C up to 45 - 50°C within which the
partial specific volume gradient with respect to temperature
assumes, on the average, a value of 3.42 x 104 ml/g
K and
at about 55°C an abrupt change in magnitude of the slope was
observed.
They suggested that this anomaly might be
attributed to a predenaturational stage connected to a
labilization of the native protein structure.
47
4. RESULTS
In order to simulate adsorption equilibrium according
to Eq.
(30) for comparison with experimental data, all
physical properties appearing in that equation must be known
for a given protein-surface contact.
In addition to protein
physical properties, the temperature at which the adsorption
experiment was conducted as well as the solid surface
hydrophobicity, expressed as 7sv, must be known.
Consequently, all pertinent data used to quantify the
protein and solid surface properties that appear in Eq.
(30)
were provided with temperature as input to a Fortran program
used to estimate the value of Ac using a nonlinear
regression method based on minimization of the difference
between the value of experimentally-based, adsorbed mass, r,
and that given by the model equation.
This Fortran program
was written for each set of isotherms tested, under the last
name of the first author.
In each case, the applicability
of equation (30) was examined using only those sets of
protein isotherms obtained from adsorption experiments
conducted on hydrophobic solid surfaces.
The determination
of adsorbed mass of protein is to some extent technique-
dependent; to preclude the possibility of such artifacts
interfering with the present analysis, each set of isotherms
was tested separately.
48
The other requirement imposed on this analysis concerns
model application only to compact, single-domain globular
proteins that attain monolayer coverages.
However, multi-
domain proteins including human fibrinogen, immunoglobulin
G, and plasminogen were used to demonstrate limits in the
applicability of Eq. (30).
With the required input data,
non-linear regression was carried out to determine the best
value for the area cleared by a protein molecule upon
adsorption, Ac.
From preliminary simulation results
allowing computer generation of both Ac and a, it was found
that the variation in a was statistically insignificant as
both a and Ac are highly correlated; hence one variable
should be fixed.
Moreover, as shown in Table 4.1, the
Table 4.1 The regressed values of a and Ac, and the
regression parameters for the model (Eq. 30). Fourteen
different isotherms (110 data points) were used as input.
95% confidence interval is shown in parentheses.
a
Ac
(A2/molecule)
0.000
192
(0.029)
(10)
0.01
195
(0.033)
(10)
0.05
207
(0.034)
(10)
0.1
(0.036)
(10)
0.5
(0.039)
(12)
222
344
Adj R2
MSE X 103
F (X)
(4g/cm2)2
0.745
9.34
1.0186
0.741
9.48
1.0331
0.729
9.94
1.0840
0.717
10.38
1.1317
0.685
11.54
1.2575
49
quality of regression is even improved as the value of
parameter a approaches zero.
For that reason and for other reasons explained in
detail in Section 5.1, the proportionality constant a, which
relates the Gibbs free energy of unfolding at the interface
to that in the bulk, was set equal to zero.
The affinity
constant, K, defined by Eq.
(29) is shown in Tables 4.4,
4.6, 4.8, 4.10, 4.12, 4.14, 4.16, and 4.18 with the
parameter a being equal to 0.0 in all cases studied.
Statistically, the goodness of fit of the model equation can
be quantified by the value of the mean square of errors
(MSE) in addition to the graphical representation of the
regressed data.
For simplicity, each of the following abbreviations
will be used for the names of the selected proteins:
a-Lac
a-Lactalbumin
/3 -Lag
/3- Lactoglobulin
BSA
Bovine Serum Albumin
ChA
Chymotrypsinogen A
Fbrgn
Fibrinogen
HSA
Human Serum Albumin
IgG
Immunoglobulin G
Lyso
Lysozyme
Myog
Myoglobin
Plmgn
Plasminogen
RiboA
Ribonuclease A
50
Table 4.2 shows the thermal properties of each protein
used to evaluate AGmkki according to Eq.
(13).
In each case,
the thermal properties were obtained using DSC analysis.
One point worth mentioning is that the values of thermal
properties shown in Table 4.2 correspond to DSC analyses
carried out under acidic pH values.
As was pointed out by
Privalov and Makhatadze (1990), the partial heat capacity
values of heat and acid-denatured proteins are
indistinguishable in the temperature range from 5 to 125°C.
Moreover, carrying out DSC analysis under alkaline pH values
results in a value of enthalpy of denaturation that is
larger than that recorded under acidic pH values (Castellino
et al., 1981; Privalov and Medved; 1982).
The reason for
that is thermal denaturation carried out under alkaline pH
values proceeds while accompanied by the aggregation of
denatured molecules, hence a higher value of enthalpy of
denaturation is encountered (Privalov and Makhatadze, 1990;
Novokhatny et al., 1984).
51
Table 4.2 Thermal properties of proteins used in
evaluating AGmkm(eguation 13), with standard errors shown
in parentheses.
Protein
a-Lac
MW
A HD
A Cp
(kJ/mole) (kJ/mol.K)
Tm
Ref.
(K)
184
4.0
(11)
(0.8)
36640
599
(19)
17.6
(dimer)
344.0
(dimer)
25.0
334.3
4
(4)
23000
557
10.8
333.1
e
340000
2259
62.8
318.9
f,g
HSA
69000
879
26.0
341.1
d,f
IgG
150000
1260
62.8
317.1
f,h
Lyso
14400
590
7.3
351.1
b,i,j
Myog
17800
254
10.4
335.1
k
Plmgn
94000
929
37.6
313.1
1
323.6
e
13-Lag
BSA
ChA
Fbrgn
14161
66267
799
RiboA
13680
407
5.0
a. Suttiprasit and McGuire (1992)
b. Pfeil (1981)
c. Pace and Tanford (1968)
d. Leibman et al. (1974)
e. Fujita and Noda (1991)
f. Zhukovskii and Rovnov (1989)
g. Privalov and Medved (1982)
h. Privalov (1976)
i. Dzakula and Andjus (1991)
j. Privalov and Makhatadze (1990)
k. Privalov et al. (1986)
1. Novokhatny et al. (1984)
312.7
a,b
a,c
a,d
52
Table 4.3 shows the specific volume (Vp), molecular
weight (MW), and partial molar area of protein upon
adsorption, Ap, for each protein examined in this study.
It
should be emphasized that a difference in the value reported
for molecular weight (MW), and to a lesser extent in the
value of specific volume (Vi), may exist from one source of
data to another.
In all cases studied, the values of MW and
Vp reported by the investigators who constructed the
isotherm for a given protein was considered first.
In
any event, this variation is negligible compared with the
value of MW itself.
Before presenting the experimentally-based isotherms,
the following points should be addressed:
1)
The protein-free solution surface tension,
levy,
is
essentially buffer-independent and assumes a value very
close to that of pure water (Suttiprasit et al., 1992;
Neumann et al., 1983).
A value of 'y
equal to 72.5
mJ/m2, measured at room temperature, was assumed for
all proteins studied, except for those that were
carried out at 52°C.
2)
Regarding isotherms carried out at 52°C (Suttiprasit
and McGuire, 1992), the effect of temperature on both
water and solid surface tension was handled by
considering the linear surface tension decrease with
increasing temperature.
53
Table 4.3 The specfic volume (Vp) , molecular weight (MW),
and the partial molar area of protein upon adsorption, Ap,
for each of the proteins examined in this study.
Protein
MW
Vp
(cm3/g)
Ap(end-on)
Ref.
(A2/molec) (cm2/4,g)
a-Lac
14161
fl-Lag
BSA
ChA
36640
0.729
725
3.08
1806
(side-on)
7.68
a,b
(dimer)
0.751
1393
2.29
a,b
66267
0.733
2037
1.85
a,b
23000
0.730
6.54
c
Fbrgn
340000
0.723
2498
(s de -on)
6003
1.06
14951
(side-on)
2.65
b,d
HSA
69000
0.733
2092
1.83
b
IgG
150000
0.739
3530
1.42
b,e
734
3.07
1828
(side-on)
7.65
Lyso
14400
Myog
Plmgn
RiboA
a.
b.
c.
d.
e.
f.
g.
h.
i.
0.730
b,f,g
17800
0.742
2129
(side-on)
7.20
g
94000
0.715
6298
(side-on)
4.03
h,i
13680
0.703
7.58
g
1723
(s de -on)
Suttiprasit and McGuire (1992)
Fasman (1976)
Pace (1983)
Privalov and Medved (1982)
Andrade and Hlady (1987)
Pilz and Czerwenka (1973)
Arai and Norde (1990)
Woodhouse et al. (1992)
Barlow et al. (1969)
54
For water, clyv/dT was taken as -0.152 mJ/m2
K, and
for the solid, dyw/dT was taken to be -0.1 mJ/m2
K
(Hiemenz, 1986).
3)
The data points, each of which represents the value of
adsorbed mass versus that of apparent equilibrium
concentration, were read from the pertinent figures
available in the literature.
Usually the data points
had been connected by a line, and that line was also
used to generate data points in cases where the number
of data points was low.
55
Figures 4.1 and 4.2 show experimental data of
Suttiprasit and McGuire (1992) curve-fitted to Eq. (30).
In
this case, a-Lac, 0-Lag, and BSA were adsorbed on
hydrophobic silicon surfaces at 27 and 52°C, from 0.01 M
phosphate buffer at pH 7.00.
Table 4.4 shows the values of
AGmwd, along with the interfacial energies of solid,
protein, and water, and the work of adhesion, lik, relevant
to the isotherms of figures 4.1 and 4.2.
Table 4.5 shows
the corresponding values of Ac, the affinity constant, K,
and the regression parameters obtained for the isotherms of
Figures 4.1 and 4.2.
56
0.6
0.5
0.4
/
0.3
................
ac Lac / Data
0.2
0.1
--­
0.0
I
0
200
400
I
600
aLac / Eq. (30)
Pa / Data
PLag Eq. (30)
BSA / Data
/
BSA / Eq.
(30)
I
800 1000 1200 1400 1600 1600 2000
Apparent Ceq (mg/1)
Figure 4.1 Protein adsorption isotherms (T=27°C) on a
hydrophobic silicon surface, fitted to Eq. (30). Ref:
Suttiprasit and McGuire (1992).
57
0.8
0.5
0.4
...... ........ ....................................
.................
0.3
ftLag / Data
0.2
- - - - - -­
13Lag / Eq. (30)
BSA / Data
BSA / Eq. (30)
0.1
0.0
.
1
0
200
400
1
1
.
1
.
1
.
600
800 1000 1200 1400 1600 1800 2000
Apparent Ceq (mg/1)
Figure 4.2 Protein adsorption isotherms (T=52°C) on a
hydrophobic silicon surface, fitted to Eq. (30). Ref:
Suttiprasit and McGuire (1992).
58
Table 4.4 The Gibbs free energy of unfolding, protein,
solid and water surface energies, and the work of adhesion
for proteins used in Figs. (4.1) and (4.2).
Source of
isotherms: Suttiprasit and McGuire (1992).
Protein
T
( °C)
AGmfold
'Ysv
(kJ/mol ) (mJ/e)
'YPV
'Ywv
Wa
(mJ/m2) (mJ/e) (10/e)
a-Lac
27
6.4
17.5
38.0
72.5
53.6
f3 -Lag
27
27.2
17.5
56.3
72.5
23.6
BSA
27
38.1
17.5
55.4
72.5
25.0
0-Lag
52
23.7
15.0
51.2
68.7
23.9
BSA
52
18.8
15.0
50.2
68.7
25.4
59
Table 4.5 The surface area cleared by an adsorbing protein
molecule, Ac, the affinity constant, K, and the regression
parameters for the model (Eq. 30). Source of isotherms:
Suttiprasit and McGuire (1992).
Protein
T
( °C)
Ac(A2/molec) Conforma(p-value)
tion
K x 104
MSE x103
a-Lac
27
72
(0.0001)
end-on
1.1 ± .4
1.5
/3 -Lag
27
158
(0.0001)
end-on
0.8 ± .2
0.8
BSA
27
158
(0.0001)
end-on
1.4 ± .4
2.0
fl-Lag
52
177
(0.0001)
end-on
1.2 ± .2
0.5
BSA
52
177
(0.0001)
end-on
2.2 ± .5
0.9
60
Figure 4.3 shows another set of isotherms (Shibata and
Lenhoff, 1992) measured at hydrophobic, butylated quartz
slides.
Total internal reflectance fluorescence (TIRF)
spectroscopy was used to assess the adsorbed mass of Lyso
and ChA.
The isotherm experiments were carried out at pH
7.0, with protein dissolved in a 0.01 M phosphate buffer
including 0.1 M NaCl.
Tables 4.6 and 4.7 show the relevant
thermodynamic data and regression results for this set of
isotherms.
61
0.16
0.14
0.12
..
0.10
........ .....................................................
0.08
0.06
ChA / Data
ChA / Eq. (30)
0.04
Lyso / Data
Lyso / Eq. (30)
0.02
0.00
0
1000
2000
3000
Apparent Ceq (mg /1)
4000
5000
Figure 4.3 Protein adsorption isotherms on hydrophobic,
butylated quartz slides, fitted to Eq. (30). Ref: Shibata
and Lenhoff (1992).
62
Table 4.6 The Gibbs free energy of unfolding, protein,
solid and water surface energies, and the work of adhesion
for proteins used in Fig. (4.3). Source of isotherms:
Shibata and Lenhoff (1992).
Protein
T
AG.Hd
(°C)
(kJ/mol )
1'sv
(10/1e)
(10/1e)
(10/m2)
l'Pv
W,
Lyso
20
62.5
35.0
66.3
8.6
ChA
20
40.9
35.0
57.7
19.2
Table 4.7 The surface area cleared by an adsorbing protein
molecule, Ac, the affinity constant, K, and the regression
parameters for the model (Eq. 30). Source of isotherms:
Shibata and Lenhoff (1992).
Protein
T
Ac(k/molec)
(°C)
(p-value)
Conformation
Lyso
20
379
(0.0001)
side-on
0.33
3.7
ChA
20
205
(0.0001)
side-on
1.63
0.4
K x 104 MSE x104
63
Figure 4.4 shows a human (milk) Lyso adsorption
isotherm recorded on hydrophobic silica slides treated with
dimethyldichlorosilane (Horsley et al., 1987).
Total
internal reflectance fluorescence (TIRF) spectroscopy was
used to assess the adsorbed mass of Lyso.
The isotherm
experiment was carried out at pH 7.4, with protein dissolved
in a PBS (0.013 M KH2PO4, 0.054 M Na2HPO4)
0.1 M NaCl.
buffer including
Tables 4.8 and 4.9 show the relevant
thermodynamic and regression data determined for this
isotherm.
64
0.24
0.22
0.20
0.18
0.16
0.14
0.12
0.10
0.08
0.06
0.04
0.02
0.00
0
500
1000
1500
2000
2500
3000
ApparentCeq(mal)
Figure 4.4 Human lysozyme adsorption isotherm (T=25°C) on
hydrophobic DDS silica slides, fitted to Eq. (30) . Ref:
Horsley et al. (1987)
.
65
Table 4.8 The Gibbs free energy of unfolding, protein,
solid and water surface energies, and the work of adhesion
for protein used in Fig. (4.4). Source of isotherm:
Horsley et al. (1987).
Protein
Lyso
T
AGmmd
'Ysv
7pv
WE,
( °C)
(kJ/mol )
(10/m2)
(10/e)
65.1
(10/10)
25
59.8
16.5
10.5
Table 4.9 The surface area cleared by an adsorbing protein
molecule, Ac, the affinity constant, K, and the regression
parameters for the model (Eq. 30). Source of isotherm:
Horsley et al. (1987).
Protein
T
( °C)
Lyso
25
Ac(A2/molec)
(p-value)
Conforma­
tion
272
(0.0001)
end-on
K x 104 MSE x104
0.10
3.9
66
Figure 4.5 shows a Plmgn adsorption isotherm recorded
on hydrophobic, methylene dianiline (MDA) polyurethane
(Woodhouse et al., 1992).
A radiolabeling technique using
1251 was used to measure the adsorbed mass of Plmgn in these
experiments.
The isotherm experiments were carried out at
pH 7.4, with protein dissolved in isotonic Tris buffer.
Tables 4.10 and 4.11 show the relevant thermodynamic and
regression data associated with the Plmgn adsorption
isotherm.
67
0.25
0.20
0.15
0.10
0.05
0.00
100
200
Apparent Ceq (mg/1)
300
Figure 4.5 Plasminogen adsorption isotherm (T=25°C) on a
hydrophobic MDA polyurethane surface, fitted to Eq.
(30).
Ref: Woodhouse et al. (1992).
68
Table 4.10 The Gibbs free energy of unfolding, protein,
solid and water surface energies, and the work of adhesion
for protein used in Fig. (4.5). Source of isotherm:
Woodhouse et al. (1992).
Protein
Plmgn
T
AG=d
'Ysv
( °C)
(kJ/mol )
(mJ/I112)
'Ypv
(mJ /m2)
(mJ/m2)
41.7
52.1
21.5
25
31.3
W.
Table 4.11 The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant, K, and the
regression parameters for the model (Eq. 30). Source of
isotherm: Woodhouse et al. (1992).
Protein
Plmgn
T
Ac(k/molec)
( °C)
(p-value)
25
229
(0.0001)
Conforma­
tion
side-on
K x 104 MSE x104
15.5
1.8
69
Figure 4.6 shows HSA adsorption isotherms on different
hydrophobic polymer surfaces studied by Winterton et al.
(1986).
An 1251 radiolabeling technique was used to measure
the adsorbed mass of HSA.
The isotherm experiments were
carried out at pH 7.4, with protein dissolved in a PBS
buffer.
The polymers used in HSA adsorption were
polystyrene (PS), polyvinylchloride (PVC), and silastic
(Silas).
Tables 4.12 and 4.13 show the relevant
thermodynamic and regression data for HSA adsorption
isotherms on these polymers.
70
0.7
0.6
0.5
.....
0.4
0.3
IV 0
I/ 0
HSA on PS /Data
HSA on PS / Eq. (30)
/go
0.2
:7
0.1
- -­
...................
o
HSA on PVC / Data
HSA on PVC / Eq. (30)
HSA on Silastic / Data
HSA on Silastic / Eq. (30)
0.0
a
0
50
100
150
200
Apparent Ceq (mg/1)
250
300
Figure 4 . 6 HSA adsorption isotherms (T=25°C) on different
hydrophobic surfaces, fitted to Eq. (30) Ref : Winterton et
al. (1986).
.
71
Table 4.12 The Gibbs free energy of unfolding, protein,
solid and water surface energies, and the work of adhesion
for protein used in Fig (4.6). Source of isotherms:
Winterton et al. (1986).
Protein
T
AG"d
( °C)
(kJ/mol)
Solid
Surface
(mJ/m2)
'Ysv
'YPv
W,
(mJ/m2) (mJ/m2)
HSA
25
40.3
PS
22.4
56.8
22.8
HSA
25
40.3
PVC
30.5
56.8
21.7
HSA
25
40.3
Silas
23.0
56.8
22.8
72
Table 4.13 The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant, K, and the
regression parameters for the model (Eq. 30). Source of
isotherms: Winterton et al. (1986).
Protein
T
( °C)
Ac(A2/molec)
(p-value)
Conformation
K x 104 MSE x103
HSA
25
206
(0.0001)
end-on
9.2
3.9
HSA
25
200
(0.0001)
end-on
3.9
0.6
HSA
25
186
(0.0001)
end-on
3.0
3.9
73
Figure 4.7 shows isotherms for several proteins that
were measured at hydrophobic, siliconized glass particles
(Absolom et al., 1987).
The isotherm experiments were
carried out at pH 7.2, with protein dissolved in a PBS
buffer of ionic strength 0.1 M.
The proteins used were IgG,
HSA, BSA, and Fbrgn. An 125I radiolabeling technique was used
to measure the adsorbed mass of these proteins.
Tables 4.14
and 4.15 show the relevant thermodynamic and regression data
for the set of isotherms shown in Fig. (4.7).
74
BSA / Data
BSA / Eq. (30)
Fbrgn / Data
Fbrgn / Eq. (30)
1.1
HSA / Data
HSA / Eq. (30)
IgG / Data
IgG / Eq. (30)
A
1.0
, ...
0.9
/
/
/
0.8
0.7
0.8
-
.009
...
1
0.5
-
I
.............................................
.......
...............
A
A
A.­
..
0.4
0.3
-
I
I/
I iA
0.2
is
0.1
+I
0.0
I
0
.
I
4000
.
8000
1
.
I
.
12000
I
.
1
16000
.
I
.
I
20000
Apparent Ceq (mg/1)
Figure 4.7 Protein adsorption isotherms (T=24°C) on
hydrophobic siliconized glass, fitted to Eq. (30). Ref:
Absolom et al. (1987).
75
Table 4.14 The Gibbs free energy of unfolding, protein,
solid and water surface energies, and the work of adhesion
for proteins used in Fig. (4.7). Source of isotherms:
Absolom et al. (1987).
Protein
T
AGwfom
7sv
W.
(kJ/mol)
7pv
( °C)
(mJ/m2)
(mJ/m2)
(mJ/m2)
BSA
24
37.3
18.7
55.4
25.1
HSA
24
39.6
18.7
56.6
23.2
IgG
24
39.6
18.7
56.6
23.2
Fbrgn
24
107.7
18.7
71.8
0.9
76
Table 4.15 The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant, K, and the
regression parameters for the model (Eq. 30). Source of
isotherms: Absolom et al., 1987.
Protein
BSA
T
Ac(A2/molec)
( °C)
(p- value)
24
52
Conforma­ K x 10-3 MSE x103
tion
end-on
0.02
1.8
(0.0003)
HSA
24
66
(0.0003)
end-on
0.04
4.1
IgG
24
122
(0.0001)
end-on
1.02
3.0
Fbrgn
24
3631
(0.0003)
end-on
3.24
29.7
77
Figure 4.8 shows a Fbrgn adsorption isotherm recorded
on a hydrophobic, quartz surface by Nygren and Stenberg
(1988).
The isotherm experiments were carried out at pH
7.2, with protein dissolved in 0.01 M PBS.
Ellipsometry and
nosorbent assay (ELISA) were both used to
bed mass of Fbrgn.
Tables 4.16 and 4.17
thermodynamic and regression data for this
#102
12-06-2007 10:36PM
Item(s) checked out to patron
E: Structural effects on enzymatic a
ODE: 120008771988
DATE: 03-05-08
E: Structural stability effects
on a
3DE: 120008753622
)ATE: 03-05-08
E: Macroscopic model for apparent pr
)DE: 120008829323
)ATE: 03-05-08
On-line Renewals at
ittp://oasi. urs'
,!u/patroninfo
m.
78
0.5
0.4
0.3
0.2
0.1
0.0
0
100
200
300
Apparent Ceq (mg /1)
400
500
Figure 4.8 Fbrgn adsorption isotherm (T=25°C) on a
hydrophobic quartz surface, fitted to Eq. (30). Ref: Nygren
and Stenberg (1988).
79
Table 4.16 The Gibbs free energy of unfolding, protein,
solid and water surface energies, and the work of adhesion
for protein used in Fig. (4.8). Source of isotherm:
Nygren and Stenberg (1988).
Protein
Fbrgn
T
AGm"d
( °C)
(kJ/mol )
25
104.8
'Ysv
'YPV
14,
(mJ/m)
47.0
(mJ ile)
(10/1e)
71.7
0.9
Table 4.17 The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant, K, and the
regression parameters for the model (Eq. 30). Source of
isotherm: Nygren and Stenberg (1988).
Protein
T
( °C)
Fbrgn
25
Ac(A2/molec)
(p-value)
Conforma­
tion
5771
(0.0001)
side-on
K x 104 MSE x103
20.0
1.9
80
Finally, a set of isotherms (Arai and Norde, 1990)
measured at hydrophobic polystyrene latices is shown in Fig.
4.9.
The isotherm experiments were carried out at pH 7.0,
with protein dissolved in 0.05 M PBS.
The adsorbed mass for
a given protein was estimated by the depletion method; i.e.,
the protein concentration in the bulk was determined before
and after conducting the adsorption scheme.
The following
proteins were examined: a-Lac; Lyso; Myog; and RiboA.
Tables 4.18 and 4.19 show the relevant thermodynamic and
regression data regarding isotherms that appear in Fig. 4.9.
81
0.16
0.14
.... =11111111.11...
0.12
......
0.10
., :..0.
../
..
...... ...
A
............................
0.08
0.06
A
All
AN
a-Lac / Data
a-Lac / Eq. (30)
ljao / Data
0.04
0.02
Lye° / Eq. (30)
Myog / Data
Myog / Eq. (30)
RiboA / Data
RiboA / Eq. (30)
A
0.00
1
0
.
1
.
1
.
1
.
1
.
1
.
1
1
.
1
.
1
.
1
100 200 300 400 500 600 700 BOO 900 1000
Apparent Ceq (mg/1)
Figure 4.9 Protein adsorption isotherms (T=25°C) on
hydrophobic polystyrene latices, fitted to Eq. (30). Ref:
Arai and Norde (1990).
82
Table 4.18 The Gibbs free energy of unfolding, protein,
solid and water surface energies, and the work of adhesion
for proteins used in Fig. (4.9). Source of isotherms:
Arai and Norde (1990).
Protein
T
( °C)
AG,Nd
(kJ/mol )
l'sv
Tpv
W,
(mJ /m2)
(10/1112)
a-Lac
24
7.2
37.0
39.8
(W/m2)
38.4
Myog
24
6.8
37.0
39.6
38.5
RiboA
24
27.1
37.0
49.7
27.1
Lyso
24
59.8
37.0
65.1
10.0
83
Table 4.19 The surface area cleared by an adsorbing
protein molecule, Ac, the affinity constant, K, and the
regression parameters for the model (Eq. 30). Source of
isotherms: Arai and Norde, 1990.
Protein
T
( °C)
Ac(A2/molec)
(p-value)
Conformation
K x 104
MSE x103
a-Lac
25
163
(0.03)
side-on
387.2
0.27
Myog
25
105
(0.0002)
side-on
1.8
0.37
RiboA
25
125
(0.0002)
side-on
0.38
0.84
Lyso
25
338
(0.0002)
side-on
0.37
0.88
84
5. DISCUSSION
The applicability of Eq. 30 is governed by the extent
to which its premises are satisfied, depending on properties
of the protein, solid surface, and solution selected for
study.
Discounting experimental inaccuracies, agreement
between the model and an experimentally-measured adsorption
isotherm would be expected for adsorption of a single-
domain, globular protein at a homogeneous, hydrophobic
surface.
Agreement is apparent in figures 4.1 through 4.6,
and to a lesser extent in figures 4.7 through 4.9.
The term
"agreement" here not only denotes coincidence between
adsorbed mass predicted by the model and that measured in
the adsorption experiment, but should also be taken to imply
that the computer-generated value for the minimum surface
area cleared by an adsorbing protein molecule, Ac, is
consistent with that expected for the selected protein.
The
importance of generating a reasonable value for Ac will be
discussed with reference to the model premises and to the
values obtained for Ac by application of the model as well
as by experimental measurement.
But first, some
justification should be provided regarding selection of some
of the input parameters to the model.
85
5.1 PARAMETERS a and Ac
5.1.1 VALUE OF a AT LOW-ENERGY INTERFACES
Simulation of Eq.
(30) would require knowledge of Ku
AG "" Vp, Ap (all four properties calculated according to
the methods of Section 3), a, and Ac.
These last two input
variables are either to be fixed at a particular system-
specific value or computer-generated.
Parameter a is the
proportionality constant that relates the Gibbs free energy
of unfolding in the bulk to that at the interface.
The
value of a was set equal to zero (i.e., fixed at a given
value) for the following reasons:
1)
Simulation results allowing computer generation of both
a and Ac showed that variation in a was statistically
insignificant in that both a and Ac were highly
correlated.
As was shown in Table 4.1, setting a equal
to zero further improves the goodness of fit of the
model when the sets of isotherms are lumped together;
2)
There is no equation of state to quantify Ac, so the
choice of fixing a while keeping Ac as a variable
allows comparison of generated Ac values with those
experimentally determined for different proteins;
3)
At similar, hydrophobic solid surfaces, the differences
in extent of unfolding experienced among proteins ought
86
to be explained by differences in Wo which was
estimated here as a function of ye and independent of
a.
In other words, in defining 7pv in terms of ire, the
effect of surface-induced unfolding has been implicitly
incorporated;
4)
Choosing a value of a less than unity indicates
enhanced hydrophobic interactions between the protein
and solid surface, which results in a less stable
surface-bound protein compared with the free protein in
the bulk.
This result was evidenced by Matsuno et al.
(1991) who found that the thermal stability of adsorbed
7-crystallins generally decreased with increasing
hydrophobicity of the surface when compared to the
proteins in solution; and
5)
As the number of computer-adjusted model parameters
increases in any event, the model sensitivity to
variations in experimental data becomes incidental,
with no definitive scientific justification.
5.1.2 VALUE OF a AT HIGH-ENERGY INTERFACES
In Section 3.2, it was shown that the overall
equilibrium can be resolved into two major sub-equilibrium
states (Fig. 3.2.1b):
1) an equilibrium between native protein in the bulk and
that at the hydrophobic interface, the attainment of which
87
is largely driven by the Gibbs free energy for adsorption
(or the work of adhesion); and
2) an equilibrium between adsorbed, native and unfolded
protein.
Attainment of this sub-state of equilibrium is
largely driven by the Gibbs free energy of unfolding.
In Section 5.1.1, it was shown that at low-energy
(i.e., hydrophobic) solid surfaces setting a equal to zero
is a reasonable assumption based on modeling considerations
and on the fact that expressing -ypv in terms of Ire
essentially takes care of the second sub-state of
equilibrium.
However, at relatively hydrophilic or high-
energy surfaces, the work of adhesion energy term would
presumably becomes less significant (i.e., smaller in
magnitude as -ysv approaches -Avv) as compared to that at
hydrophobic interfaces.
Moreover, with relatively minor
destabilization effects at hydrophilic surfaces (Matsuno et
al., 1991), the energy barrier to surface-induced unfolding
would presumably increase.
In other words, the second sub-
state of equilibrium would become more significant as the
surface becomes more hydrophilic.
5.2 MOLECULAR INFLUENCES ON Ac
Tables 4.5, 4.7, 4.9, 4.13, 4.15, and 4.19 show that
values of Ac obtained from simulation vary within an order
of magnitude, and there is no obvious correlation between
88
molecular weight and Ac.
be fixed at a given value.
One might conclude that Ac could
It should be noted, however,
that although changes in Ac are within an order of
magnitude, the 95% confidence interval associated with Ac
was found, on the average, to be equal to 5% of Ac (except
for the case of a-lactalbumin that is shown Table 4.19, for
which a confidence interval of 165 K2/molecule was
determined).
The strong dependence on Ac should be expected
given that Ac is the only computer-generated parameter
present in the model.
This being the case, an approximate
value of Ac ought to be given as a function of some
molecular properties of protein.
This will be shown in
detail in Section 5.2.
As seen in Tables 4.5, 4.7, 4.9, 4.13, 4.15, and 4.19,
Ac assumed values within a few hundred square angstroms
(about 100 to 400 A2), which is relatively small compared to
the cross-sectional area of a "typical" globular protein,
which varies from about 1000 to 10,000 A2 (Andrade, 1985).
This implies that only a small portion of the protein
molecule need enter the interface in order for adsorption to
proceed.
These results are also in agreement with those of
Damodaran and Song (1988), who found that the area cleared
by BSA to anchor itself to the air-water interface varied
from 50 to 135 A.
Ter-Minassian-Saraga (1981) suggested
that the "hole area" (i.e., Ac) formed by adsorbing protein
may be related to the water activity at the interface and
89
not to the size of the adsorbing molecule.
Macritchie
(1978) suggested that relatively small values of Ac indicate
that each adsorbed molecule is sufficiently flexible to
behave as a series of largely independent kinetic units;
adsorption is thus a function of segment behavior and
independent of molecular size.
There is no equation of state relating Ac to pertinent
protein, solid surface, and solution properties.
Nevertheless, two general trends may be inferred upon
comparison of W, and AGmfold (Tables 4.4, 4.6, 4.8, 4.12,
4.14, and 4.18) with corresponding values of Ac (Tables 4.5,
4.7, 4.9, 4.13, 4.15, and 4.19).
1)
These trends are:
The larger the value of WI, the smaller the value of
Ac (in other words, the more hydrophobic the solid
surface or protein "surface", the smaller the
interfacial area needed for the initial attachment);
and
2)
The larger the value of AGmfold, the larger the value
of Ac.
It should be mentioned, however, that these trends do not
exactly hold for every case studied.
As shown in Table 4.4, the effect of temperature
(picked up in part by AGmfold) on Ac is generally
insignificant.
For example, Ac for 0-Lag increased from 158
to 177 A2, and for BSA increased from 158 to 177 A2 when the
temperature was increased from 27 to 52°C.
A small increase
90
in Ac as a function of temperature was also reported for
adsorption of ovalbumin at the air/water interface, where Ac
increased from 150 to 170 A2 when the temperature increased
from 5 to 20 °C (MacRitchie, 1978).
5.3 AN APPROXIMATE ESTIMATE FOR Ac AS A
FUNCTION OF MW AND Z
As was pointed out in Section 5.2, the model is
sensitive to variations in Ac as it is the only computer-
generated parameter present in the model.
The strong
dependence on Ac can be visualized via comparison between
the isotherms constructed with an isotherm-specific Ac value
(Fig. 5.3.1) and the same isotherms with Ac held constant
(Fig. 5.3.2).
As can be seen from Figs. 5.3.1 and 5.3.2, the initial
slope (i.e., the affinity constant, K) of the plot is
sensitive to variations in Ac.
This being the case and as
there is no equation of state to define Ac, one might
attempt to providean estimate for Ac value in terms of
molecular properties.
Figure 5.3.3 shows Ac as a function
of molecular weight (MW) and the flexibility parameter (Z).
91
0.6
Ac = 158 A2
-
0.5
A-
0.4
Ac= 158 A2
Ac = 72 A2
0.3
............ ............
a-Lac / Data
0.2
------- a-Lac / Eq. (30)
n-Lag / Data
P-Lag / Eq. (30)
0.1
BSA / Data
BSA / Eq. (30)
0.0
.
0
1
200
.
1
400
.
.
1
600
1
800
.
1
1
1000 1200
.
1
1400 1600 1800 2000
Apparent Ceti (mg/1)
Figure 5.3.1 Isotherms of Fig. (4.1) plotted with the
isotherm-specific Ac value and a value of a = 0.0.
92
OA
0.5
0.4
0.3
aLac / Data
------ aLac / Eq. (30)
PLag / Data
0.2
PLag / Eq. (30)
0.1
BSA / Data
BSA / Eq. (30)
0.0
0
200
400
600
BOO 1000 1200 1400 1600 1800 2000
Apparent Ceti (mg/1)
Figure 5.3.2 Isotherms of Fig. (4.1) constructed with the
same Ac value (Ac = 192 A2) and a value of a = 0.0.
93
240
220
200
180
160
140
120
Ac = 28.05
{(MW)1/2 j Z2}0.504
100
80
.
80
20
I
I
90
40
(mw)1/2
50
60
Z2
Figure 5.3.3 Ac correlated with MW and Z. The adjusted R2
is 0.77 and the standard errors associated with the
preexponent and exponent terms are 8.5 and 0.09;
respectively.
94
It should be pointed out that lysozyme was excluded
from Fig. 5.3.3 because lysozyme assumed, on the average, an
Ac value equal to 330
A2
/molecule, which is relatively large
compared to Ac values of other proteins considered in this
study.
The reason lysozyme has a relatively high value of
Ac may be attributed to the point that lysozyme is an
exceptionally rigid and stable molecule as can be inferred
from its relatively high AGmfold value.
Another reason may be
related to orientational effects; i.e., the lysozyme
molecule consists of a single polypeptide chain containing
129 amino acid residues, and the hydrophobic and hydrophilic
side chains tend to occur in large clusters rather than
being randomly distributed throughout the polypeptide
(Keshavarz and Nakai, 1979).
Both reasons may contribute to
the point that lysozyme needs a relatively large hole area
to anchor to the interface.
5.4 IMPLICATIONS ASSOCIATED WITH THE
MODEL APPLICATION TO MULTI-DOMAIN
PROTEINS
Before discussing the implications associated with
extending application of the model to multi-domain proteins,
the difference between a single-domain and multi-domain
protein should be addressed.
During thermal denaturation, a
95
single-domain protein is one that undergoes a one-step (all­
or-none) reversible phase transition between two
thermodynamically defined conformations (native and
unfolded).
Small, globular proteins such as mygolobin,
ribonuclease, cytochrome C, a-chymotrypsin, a-lactalbumin,
fl-lactoglobulin, and lysozyme are categorized under this
class of proteins.
On the other hand, if the protein
molecular weight is relatively large the molecule can often
be resolved into a number of somewhat independent domains.
Using DSC analysis, the presence of independent kinetic
units, which also exhibit some degree of cooperativity,
can
be discerned.
In particular, the thermogram of a multi-
domain protein is usually resolved into a set of peaks, if
DSC analysis is carried out at acidic pH values
(Novokhatny
et al., 1984).
Multi-domain proteins considered in this
work include human and bovine serum albumin (Kragh-Hansen,
1981), immunoglobulin G (Tischenko et al., 1982),
plasminogen (Novokhatny et al., 1984), and fibrinogen
(Privalov and Medved, 1982).
As far as the application of the model to multi-domain
proteins is concerned, the central problem lies in how to
define AGmfold for such proteins, which in turn affects the
estimation of both W, and the computer-generated parameter
Ac.
One way to tackle the adsorption of multi-domain,
complex proteins is to use the so-called domain approach
(Andrade et al., 1990; Suttiprasit et al., 1992).
The
domain approach considers a complex protein molecule like
96
BSA, IgG, Plmgn, or Fbrgn as being constructed of functional
and structural domains generally identified as regions of
relatively high packing density that are calorimetrically
independent (Andrade et al., 1990).
As far as adsorption is concerned, the interfacial
behavior of a complex protein could possibly be considered
as largely dominated by the interfacial activity of only one
domain or even a sub-domain (Andrade et al., 1990).
Although the conformational changes of the protein at the
interface is ultimately governed by the stability of each of
the individual protein domains, the most thermolabile domain
(which may comprise several thermal cooperative fragments)
is expected to play a major role in the initial events
contributing to surface-induced unfolding reactions.
In this work, the domain approach was used in the case
of Fbrgn, Plmgn, and IgG adsorption.
For either HSA or BSA,
the thermogram exhibited only one peak (Suttiprasit and
McGuire, 1992; Leibman et al., 1975), i.e., as if they were
single-domain globular proteins.
The reason for that is
probably because the DSC analysis were performed at alkaline
pH values (pH 7.00 for BSA, and pH 6.00 for HSA) compared to
their isoelectric points (pI is about 5.0 to 5.1 for both
HSA and BSA).
Nevertheless, the notion of using the domain
approach is still applicable, should DSC analysis be
available for these multi-domain proteins.
97
The value of Ac obtained for each of these multi-domain
proteins is therefore governed by the extent to which the
domain approach is appropriate in each case studied, and the
reliability of such a domain-based AGmbm value as an index
of thermal stabilility of the entire protein.
For Fbrgn, although the model to a large extent is in
quantitative agreement regarding the extent of Fbrgn
adsorption (Figs. 4.7 and 4.8), Ac (Table 4.17) is one order
of magnitude larger than any of the other protein-specific
Ac values.
No value was found for Ac in the literature to
verify this result, so the large value of Ac may simply be
incidental, owing to the relatively large value of AG " for
Fbrgn (Table 4.14), which in turn yields a relatively small
value of W.
It should be noted that the area of the first
peak in the Fbrgn thermogram is approximately equal to twice
that of the thermolabile fragment, namely fragment DH with
MW 95,000 (Privalov and Medved, 1982).
As a result of the
geometrical symmetry of the Fbrgn molecule, two DH fragments
exist per molecule, which in turn results in a relatively
large value of AGmud.
One could argue using
112AGunfom to
represent only one DH fragment, nevertheless, these two DH
fragments show some degree of cooperativity as part of the
entire Fbrgn molecule, which makes them act as one
independent domain being made of two fragments.
One could ask why should not the value of Ac be
relatively large, as the human fibrinogen molecule was the
98
largest examined in this work.
The answer to this question
lies in the proposition that Ac is not correlated with the
molecular weight of adsorbing protein, and even with a huge
molecule like myosin (MW 850,000), a value of Ac equal to
145 A2 at the air-water interface has been reported (Ter­
Minassian-Saraga, 1981).
IgG is another molecule with a multi-domain structure
(Tischenko et al., 1982), and its thermal unfolding is known
to proceed in two separate stages: the first stage of
unfolding requiring 300 kcal/mol and the second stage 900
kcal/mol, with an ovarall enthalpy of 1200 kcal/mol
(Privalov, 1976).
These two transitions do not show
overlapping peaks on a thermogram, indicating two thermally
independent domains.
Only the first thermogram peak (300
kcal/mol enthalpy and T, equal to 317.1 K) was considered in
the calculation of AGmfold for two reasons.
The first regards
attention to the domain approach to the study of adsorption
of complex proteins with the thought that the first
thermogram peak represents the thermolabile fragment of the
IgG molecule; i.e., the interfacial behavior of IgG is
thought to be largely dominated by the interfacial activity
of this thermolabile fragment.
The second reason is based
on studies made by Chasovnikova et al. (1982), who found ye
for IgG monolayers equal to 16 mJ /m2; based on Eq.
yields a value of -ypv equal to 56.5 mJ /m2.
(9), this
Their results
compare well with the value of ire obtained using Eq.
(10),
99
and of ypv using Eq.
(9)
(Table 4.14), when obtained with a
value of AGmfold based on the 300 kcal/mol enthalpy of
denaturation.
Human plasminogen is also a molecule with a multi-
domain structure, though this molecule is not as large as
IgG or Fbrgn.
Plasminogen may designate any of the several
plasminogen genetic variants.
Examples are Lys-plasminogen
(MW = 83,000) and Glu-plasminogen (MW = 92,000).
Upon
limited plasminolysis, the intact plasminogen with a
glutamic acid residue at the N-terminal end (Glu­
plasminogen) loses its first 76 amino acid residues and is
converted into modified Lys-plasminogen, with a lysine
residue at the N-terminus.
The plasminogen molecule is
subdivided into more or less independent subunits, i.e.,
into structural domains (Novokhatny et al., 1984).
Novokhatny et al. (1984) suggested that two large domains of
Lys-plasminogen are indeed likely to be quite independent,
judging by calorimetric analyses showing thermal transitions
in two nearly separate temperature regions: the N-terminal
part of the molecule, including its first four fragments,
and the C-terminal part, which includes three subunits
forming miniplasminogen (residues Va1m2 to Asn790).
The first
thermogram peak, that represents the thermolabile part
(i.e., the four-fragment, N-terminal part) of the molecule,
was used for calculation of AGmw of Lys-plasminogen.
Since
human plasminogen studied by Woodhouse et al. (1992) has a
molecular weight of 94,000, which better matches with Glu­
100
plasminogen as opposed to Lys-plasminogen and since AGukm
used in Eq. (30) was entered on a molar basis, an adjustment
was made to the extensive, thermal properties of Lys­
plasminogen based on the molecular weight difference between
these plasminogen variants.
5.5 THE AFFINITY AND EXTENT OF
ADSORPTION
5.5.1 ADSORPTION AFFINITY
The affinity of a protein in single-component solution
for a given surface can be characterized by the affinity (or
equilibrium) constant, K, given by Eq.
(29).
This
equilibrium constant connotes the total driving force for
the process of adsorption.
Not only does it constitute the
driving force for arrival to the surface, but also for
surface unfolding, which differentiates protein adsorption
equilibrium from that of small molecules.
As defined by Eq. (21), K represents the initial slope
of a plot of r Ap versus Vp C.
In the absence of
electrostatic and specific biochemical interactions, the
affinity constant depicts the magnitude of hydrophobic
interaction between the protein and solid surface.
surface hydrophobicity of protein and solid were
The
101
characterized by -ypv and 7svr respectively.
The more
hydrophobic the protein and solid surface, the larger the
value of K.
This hydrophobicity-dependence of K can be seen
as a general trend in Table 4.5 (comparing 0-Lag and BSA at
52°C), in Table 4.7 (comparing Lyso ChA), and in Table 4.19
(comparing Myog and either RiboA or Lyso).
However, the
above-mentioned statement lacks consistency with respect to
the other isotherms studied.
There are at least three
possible reasons for this.
The first concerns the
reliability of experimental data.
The affinity constant
represents the initial slope of a Langmuir-type isotherm,
and the degree of uncertainty associated with determining
the initial slope is much larger than that associated with
determining the plateau value of the same isotherm,
excluding those isotherms carried out at very low
concentrations (Woodhouse et al., 1992, Nygren and Stenberg,
1988).
In fact, some isotherms (e.g., Lyso and RiboA in
Fig. 4.9) exhibit a plateau value over the entire
concentration range studied, which makes it difficult to fit
to any model other than r = rte.
Isotherms that exhibit a
high affinity character, with a plateau almost over the
entire range, should be reconstructed at lower bulk
concentrations.
But this would be accompanied by enhanced
potential for experimental artifacts (e.g., dilution
effects, diffusion-limitations, impurity effects, and
reliability of the instrumentation).
Second, the effect of
surface hydrophobicity largely manifests itself in an
102
increase in extent of adsorption (r
), which makes the
importance of the initial slope less significant.
More than
one group of investigators (Suttiprasit and McGuire, 1992;
Absolom et al., 1987; Elwing et al., 1986; Jonsson et al.,
1982) reported that the adsorbed mass of protein on
hydrophobic surfaces was greater than that on hydrophilic
surfaces.
Third, although the effect of -ysv is picked up by
lila, the effect of surface hydrophobicity extends beyond its
contribution to 141,:
it also influences the surface-induced
unfolding (Andrade, 1985; MacRitchie, 1978).
5.5.2 THE EXTENT OF ADSORPTION
Two factors affect the extent of adsorption, rte:
molecular size and the strength of hydrophobic interactions
between the protein and the solid surface.
As shown in
Figs. 4.1 through 4.9, the larger the molecular weight, the
larger the adsorbed mass.
If as in blood serum, molecular
weights among proteins vary by one or two orders of
magnitude, contribution to total adsorbed mass could be
dominated by larger molecules in part simply because they
are large (Suttiprasit and McGuire, 1992).
And, the
stronger the hydrophobic interactions, the larger the
adsorbed mass per unit area.
The effect of molecular size
on extent of adsorption has been taken into account using
Eqs. 16, 17, and 22, via the partial molar volume.
103
The effect of hydrophobic interactions appears in part
by virtue of K; i.e., given proteins of equal size, the
stronger the hydrophobic interaction, the larger the value
of K, the larger the adsorbed mass per unit area.
Toward minimization of the number of parameters
required as input to the model, no parameter was
incorporated in Eq.
(22) to account for surface-induced
unfolding effects on r
.
In future work, should Ac be
fixed or defined, one may incorporate a computer-generated
parameter that will account for surface-induced unfolding
effects on r
.
The goal in this work was to lay out a model complex
enough to describe protein adsorptive behavior at
hydrophobic, solid-water interfaces, but not so intricate as
to lose track of surface and molecular property influences
on the observed phenomena.
5.5.3 A NOTE ON TEMPERATURE EFFECTS
The effect of temperature on K can be seen by comparing
protein isotherms conducted at 52°C with those conducted at
27°C.
For both 0-Lag and BSA, K increased with temperature
(Table 4.5).
Applying the van't Hoff equation [ln K2 / K1 =
AHaddR(1/T2
1/T1) ] between state 1 (i.e., T1 = 27°C) and
state 2 (i.e., T2 = 52°C), the following results were
obtained:
104
0-Lag: AH,d, = 13.2 kJ/mol
BSA: AH,d, = 14.7 kJ/mol
The standard Gibbs free-energy change of adsorption, AG
can be evaluated at T = 27°C, using AG", = -RT1nK, for each
protein, such that
/3 -Lag: AG",=-22.4 kJ/mol=* TAS",=
BSA: AG,d,=-23.8 kJ/mol=* TASads= AHads
AGads =
35.6 kJ/mol
= 38.5 kJ/mol
Assuming AH,d, is constant over the temperature range
examined, AG", can be evaluated for each protein at T = 52 °C,
such that
0-Lag: AG,d8=-25.4 kJ/mol= TAS,Ls= AH,d,- AG", = 38.6 kJ/mol
BSA: AGads=-27.0 kJ /moles TASads= AH,d,- AG,d, = 41.7 kJ/mol
These results are in harmony with the notion that protein
adsorption at hydrophobic surfaces is entropically driven,
and these entropically-driven, hydrophobic interactions
between the protein and the silicon surface are favored with
increasing temperature.
105
6. CONCLUSIONS
1.
At hydrophobic interfaces, a general agreement between
Eq.
(30) and experimental data was observed for single-
domain globular proteins indicating that AGmfal and W, Ac
play a major role in governing the course of
adsorption.
2.
The values of Ac obtained from simulation vary within
an order of magnitude (i.e., values within a range of
about 100 to 400 V), indicating that only a small
portion of the protein molecule need enter the
interface in order for adsorption to proceed.
3.
Choosing a = 0 results in a computer-generated value
for Ac in quantitative agreement with those provided in
literature, and is in line with the concept of
hydrophobic interactions between the protein and solid
surface facilitating the surface-induced unfolding.
4.
Implementing the domain approach to characterizing
adsorption of multi-domain proteins resulted in Ac
values consistent with the notion that Ac is
independent of molecular weight,
indicating that the
interfacial behavior of a complex protein could be
considered as largely dominated by the interfacial
activity of only one domain; the most thermolabile
domain (which may consist of several thermal
106
cooperative fragments) appeared to play a major role in
the initial events contributing to surface-induced
unfolding.
5.
For fibrinogen, the large value of Ac obtained may
simply be incidental, owing to its relatively large
value of AG.Nd (which in turn yields a relatively large
value of -ypv, and relatively small value of WO.
107
7. RECOMMENDATIONS
For future work, the following two studies are
recommended. First, single-component protein adsorption
experiments should be conducted at high-energy surfaces,
where it is anticipated that a AGmfold and W, Ac are each
important as energy terms in quantifying the affinity or
equilibrium constant, K.
Moreover, it would be reasonable
to redefine the work of adhesion such that -ypv is based on
surface energetics of an intact protein, and not on the
equilibrium spreading pressure.
Ultimately, we may be able
to develop an equation of state for
Ac.
For a better
quantification of the effect of molecular properties on K,
conducting adsorption experiments at very low concentrations
is recommended in any event.
Second, binary-component adsorption experiments should
be conducted.
The overall equilibrium constant for the
binary mixture of proteins may at first be equated with the
product of each of the protein-specific values of K derived
from single-component tests.
Deviation from the equality
might be attributable to Vroman effects; i.e., relative
surface activities among adsorbing proteins that lead to the
eventual make-up of an adsorbed layer in multi-protein
systems.
108
BIBLIOGRAPHY
1.
Absolom, D.R., F.V. Lamberti, Z. Policova, W. Zingg,
C.J. van Oss, and A.W. Neumann (1983). "Thermodynamics
of Bacterial Adhesion", Appl. Environ. Microbiol., 46,
pp. 90-97, American Society for Microbiology.
2.
Absolom, D.R., W. Zingg, and A.W. Neumann (1987).
"Protein adsorption to polymer particles: Role of
Surface properties", J. Biomed. Mater. Res., 21, pp.
161-171, John Wiley & Sons, Inc.
3.
Andrade, J.D. (1985). "Surface and Interfacial aspects
of Biomedical Polymers", 2, Plenum Press, New York and
London.
4.
Andrade, J.D. and V. Hlady (1987). "Plasma Protein
Adsorption: The Big Twelve", Annals New York Academy of
Sciences, 516, pp. 158-172.
5.
Andrade, J.D., J. Herron, V. Hlady, and D. Horsley
(1987). "Simulation of Protein Adsorption. The
Denaturation Correlation", Croatica Chemica Acta, 60,
No. 3, pp. 495-503.
6.
Andrade, J.D., V. Hlady, A.-P. Wei, and C.-G. Golander
(1990). "A Domain Approach to the Adsorption of Complex
Proteins: Preliminary Analysis and Application to
Albumin", Croatica Chemica Acta, 63, No. 3, 527-538.
109
7.
Arai, T. and W. Norde (1990). "The behavior of some
model proteins at solid-liquid interfaces
1. Adsorption from single protein solutions", Colloids
and Surfaces, 51, pp. 1-15, Elsevier Science Publishers
B.V., Amsterdam.
8.
Arnebrant, T. and T. Nylander (1986). "Sequential and
Competitive Adsorption of fl-Lactoglobulin and K.-Casein
on Metal Surfaces", J. Coll. Interf. Sci., 111, No. 2,
pp. 529-533, Academic Press, Inc.
9.
Arnebrant, T., K. Barton, and T. Nylander (1987). J. of
Colloid and Interface Science, 119, No. 2.
10.
Baldwin, R.L. (1986). "Temperature dependence of the
hydrophobic interaction in protein folding", Proc.
Natl. Acad. Sci. USA, 83, pp. 8069-8072.
11.
Barlow, D.J. and J.M. Thornton (1983). "Ion-pairs in
Proteins", J. Mol. Biol., 168, pp. 867-885, Academic
Press Inc., London.
12.
Barlow, G.H., L. Summaria, and K.C. Robbins (1969).
"Molecular Weight Studies on Human Plasminogen and
Plasma at the Microgram Level", J. Biological
Chemistry, 244, No. 5, pp. 1138-1141, printed in U.S.A.
13.
Becktel, W.J. and J.A. Schellman (1987). " Protein
Stability Curves", Biopolymers, 26, pp. 1859-1877, John
Wiley & Sons, Inc.
110
14.
Bendzko, P.I., W.A. Pfeil, P.L. Privalov, and E.I.
Tiktopulo (1988). "Temperature-induced phase
transitions in proteins and lipids / Volume and heat
capacity effects", Biophysical Chem., 29, pp. 301-307.
15.
Brash, J.L and D.J Lyman (1969). "Adsorption of Plasma
Proteins in Solution to Uncharged, Hydrophobic Polymer
Surfaces", J. Biomed. Mater. Res., 3, pp. 175-189, John
Wiley & Sons, Inc.
16.
Bull, H.B. and K. Breese (1973). "Temperature
Dependence of Partial Volume of Proteins", Biopolymers,
12, pp. 2351-2358, John Wiley & Sons, Inc.
17. Castellino, F.J., V.A. Ploplis, J.R. Powell,
and D.K.
Strickland (1981). "The Existence of Independent Domain
Structures in Human Lysn-Plasminogen", J. Biol. Chem.,
256, No. 10, pp. 4778-4782.
18.
Chasovnikova, L.V., N.A. Matveyeva, and V.V. Lavrent'ev
(1982). " Study of the conformational stability of
immunoglobulin G in monolayers at the phase boundary
between aqueous NaC1 and octane", Biofizika, 27, No. 3,
pp. 435-440 (Biophysics, 27, No. 3, pp. 444-450),
printed in Poland.
19.
Chothia, Cyrus (1975). "Structural invariants in
protein folding", Nature, 254, pp. 304-308.
20.
Creighton, T.E. (1983). "An Empirical Approach to
Protein Conformation Stability and Flexibility",
Biopolymers, 22, pp. 49-58, John Wiley & Sons, Inc.
111
21.
Dabrowski, Andrzej, Mieczystaw Jaroniec, and Jarostaw
Oscik (1987). "Multilayer and Monolayer Adsorption from
Liquid Mixtures of Nonelectrolytes on Solid Surfaces",
in: Surface and Colloid Science,
(Egon Matijevic,
editor), 14, pp. 83-213, Plenum Press, New York and
London.
22.
Damodaran, S. and K.B. Song (1988). "Kinetics of
Adsorption of Proteins at Interfaces: Role of Protein
Conformation in Diffusional Adsorption, Biochimica et
Biophysica Acta, 954, pp. 253-264, Elsevier Science
Publishers B.V. (Biomedical Division).
23.
De Feijter, J.A., J. Benjamins, and F.A. Veer (1978).
"Ellipsometry as a Tool to Study the Adsorption
Behavior of Synthetic and Biopolymers at the Air-Water
Interface", Biopolymers, 17, pp. 1759-1772, John Wiley
& Sons, Inc.
24.
De Feijter, J.A., J. Benjamins and M. Tamboer (1987).
Colloid and Surfaces, 27, pp. 243-266.
25.
Dill, Ken A., Darwin O.V. Alonso, and Karen Hutchinson
(1989).
"Thermal Stabilities of Globular Proteins",
Biochemistry, 28, pp. 5439-5449, American Chemical
Society.
26.
Durchschlag, H.
(1986), in: Thermodynamic data for
biochemistry and biotechnology,
(H.J. Hinz, editor), p.
45.
112
27.
Dzakula, Z. and R.K. Andjus (1991). "Biophysical Models
of Protein Denaturation. I. An Improvement of the Model
of Two States", J. theor. Biol., 153, pp. 41-59,
Academic Press Limited, London.
28. Elwing, H., A. Askendal, and I. Lundstrom
(1987).
"Protein exchange reactions on solid surfaces studied
with a wettability gradient method", Progr. Colloid &
Polymer Sci., 74, pp. 103-107.
29.
Elwing, H., B. Ivarsson, and I. Lundstrom (1986), Bur.
J. Biochem., 156, 359.
30.
Fasman, G.D. (1976). "Handbook of Biochemistry and
Molecular Biology", 3"1 edition, Proteins - Vol. II,
CRC, Press, Cleveland, Ohio.
31.
Fasman, G.D. (1989). "Prediction of Protein Structure
and the Principles of Protein Conformation", Plenum
Press, New York and London.
32.
Fersht, A.R. (1987). "The Hydrogen Bond in Molecular
Recognition", Trends Biochem. Sci., 12, pp. 301-304.
33.
Fujita, Y. and Y. Noda (1991). "Effect of reductive
alkylation on thermal stability of ribonuclease A and
chymotrypsinogen A", Int. J. Peptide Protein Res., 38,
pp. 445-452.
34.
Graham, D.E. and M.C. Phillips (1979a).
"Proteins at
Liquid Interfaces. I. Kinetics of Adsorption and
Surface Denaturation", J. Coll. Interf. Sci., 70, No.
3, pp. 403-414, Academic Press, Inc.
113
35.
Graham, D.E. and M.C. Phillips (1979b). "Proteins at
Liquid Interfaces. II. Adsorption Isotherms", J. Coll.
Interf. Sci., 70, No. 3, pp. 415-426, Academic Press,
Inc.
36.
Hiemenz, P.C. (1986). "Principles of Colloid and
Surface Chemistry", 2nd edition, p. 296, Marcel Dekker,
Inc., New York and Basel.
37.
Honeycutt, J.D. and D. Thirumalai (1992). "The nature
of folded states of globular proteins", Biopolymers,
32, pp. 695-709, John Wiley & Sons, Inc.
38.
Horsley, D., J. Herron, V. Hlady, and J.D. Andrade
(1987). "Human and Hen Lysozyme Adsorption: A
Comparative Study Using Total Internal Reflection
Fluorescence Spectroscopy and Molecular Graphics", In:
Proteins at Interfaces: Physicochemical and Biochemical
Studies, J.L. Brash and T.A. Horbett, eds. ACS
Symposium Series 343, Washington, D.C., p. 290.
39.
Hunter, J.R., P.K. Kilpatrick, and R.G. Carbonell
(1990). "Lysozyme Adsorption at the Air/Water
Interface", J. Coll. Interf. Sci., 137, No. 2, pp. 462­
482, Academic Press, Inc.
40. Hunter, J.R., R.G. Carbonell, and P.K. Kilpatrick
(1991). "Coadsorption and exchange of lysozyme/O-casein
mixtures at the air/water interface", J. Colloid
Interface Sci., 143, No. 1, pp. 37-53, Academic Press,
Inc.
114
41.
Jonsson, U., B. Ivarsson, I. Lundstrom, and L. Berghem
(1982), J. Coll. Interf. Sci., 90, 148, Academic Press,
Inc.
42.
Jonsson, U., I. Lundstrtim, and I. Ronnberg (1987).
"Immunoglobulin G and Secretory Fibronectin Adsorption
to Silica.
The influence of Conformational Changes on
the Surface", J. Coll. Interf. Sci., 117, No. 1, pp.
127- 138, Academic Press, Inc.
43.
Kato, A. and K. Yutani (1988). "Correlation of surface
properties with conformational stabilities of wild-type
and six mutant tryptophan synthase a-subunits
substituted at the same position", Protein Engineering,
2, No. 2, pp. 153-156, IRL Press Limited, Oxford,
England.
44. Kauzmann, W.
(1959). "Some factors in the
interpretation of protein denaturation, Adv. Prot.
Chem., 14, pp. 1-63.
45.
Keshavarz, E. and S. Nakai (1979). "The relationship
between hydrophobicity and interfacial tension of
proteins", Biochim. et. Biophys. Acta, 576, pp. 269­
279, Elsevier, North-Holland Biomedical Press.
46. Khaiat, A., and I.R. Miller (1969). Biochim. Biophys.
Acta., 183, pp. 309.
47.
Klapper, M.H. (1971). "on the Nature of Protein
Interior", Biochim. Biophys. Acta, 229, pp. 557-566.
115
48. Koteliansky, V.E., M.A. Glukhova, M.V. Bejanian, v.N.
Smirnov, V.V. Filimonov, O.M. Zalite, and S.Yu.
Venyaminov (1981). "A study of the structure of
fibronectin", Eur. J. Biochem., 119, pp. 619-624.
49.
Kragh-Hansen, U. (1981). "Molecular Aspects of Ligand
Binding to Serum Albumin", Pharmacological Reviews, 33,
No. 1, pp. 17-53, The American Society for Pharmacology
and Experimental Therapeutics, printed in U.S.A.
50.
Lee, R.G. and S.W. Kim (1974). "Adsorption of Proteins
onto Hydrophobic Polymer Surfaces: Adsorption Isotherms
and Kinetics", J. Biomed. Mater. Res., 8, pp. 251-259,
John Wiley & Sons, Inc.
51.
Leibman, D.Ya., Ye. I. Tiktopulo, and P.L. Privalov
(1975). "Investigation of the conformational
conversions of serum albumin by the method of scanning
microcalorimetry", Biofizika, 20, No. 3, pp. 376-379
(Biophysics, 20, No. 3, pp. 379-382), printed in
Poland.
52.
Lin, S., R. Blanco, and B.L. Karger (1991).
"Adsorption-desorption isotherm hysteresis of
fl-lactoglobulin A with a weakly hydrophobic surface",
J. Chromatogr., 557, pp. 369-382.
53.
Lu, D.R., S.J. Lee, and K. Park (1991). "Calculation of
solvation interaction energies for protein adsorption
on polymer surfaces", J. Biomater. Sci. Polym. Ed., 3,
No. 2, pp. 127-147.
116
54.
Lundstrom, I.
(1985). "Models of protein adsorption on
solid surfaces", Progr. Colloid. & Polymer Sci., 70,
pp. 76-82.
55.
Lundstrom, I., B. Ivarsson, U. JOnsson, and H. Elwing
(1987). "Protein Adsorption and Interaction at Solid
Surfaces" in: Polymer Surfaces and Interfaces, (W.J.
Feast and H.S. Munro, editor), pp. 201-229, John Wiley
& Sons, Inc.
56.
Macritchie, F.
(1978). "Proteins at Interfaces" in:
Advances in Protein Chemistry, (C.B. Anfinsen, J.T.
Edsall, and F.M. Richards), 32, pp. 283-326, Academic
Press, New York, San Francisco and London.
57.
Matsumura, M., W.J. Becktel, and B.W. Matthews (1988).
"Hydrophobic stabilization in T4 lysozyme determined
directly by multiple substitutions of Ile 3", Nature,
334, pp. 406-410.
58. Matsuno, K., R.V. Lewis, and C.R. Middaugh
(1991). "The
interaction of 7-crystallins with model surfaces",
Arch. Biochem. Biophys, 291, No. 2, pp. 349-355.,
Academic Press, Inc.
59. Mizutani, T. and J.L. Brash (1988). "A Thermodynamic
Study of Albumin Adsorption onto Some Solid Surfaces",
Chem. Pharm. Bull., 36, No. 7, pp. 2711-2715.
60.
Morrissey, B.W. and R.S. Stromberg (1974). "The
Conformation of Adsorbed Blood Proteins by Infrared
Bound Fraction Measurements", J. Coll. Interf. Sci.,
46, No. 1, pp. 152-164, Academic Press, Inc.
117
61.
Murphy, K.P., P.L. Privalov, S.J. Gill (1990). "Common
Features of Protein Unfolding and Dissolution of
Hydrophobic Compounds", Science (Washing.), pp. 559­
561.
62.
Neumann, A.W., D.R. Absolom, D.W. Francis, S.N. Omenyi,
J.K. Spelt, Z. Policova, C. Thomson, W. Zingg, and C.J.
van Oss (1983). "Measurement of surface tensions of
blood cells and proteins", Annals New York Academy of
Sciences, pp. 276-298.
63. Neumann, A.W., O.S. Hum, and D.W. Francis (1980).
"Platelet Adhesion", J. Biomed. Mater. Res., 14, pp.
499-509.
64. Neumann, A.W., R.J. Good, C.J. Hope, and M. Sejpal
(1974). "An Equation-of-State Approach to Determine
Surface Tensions of Low-Energy Solids from Contact
Angles", J. of Colloid and Interface Science, 49, No.
2., pp. 291-304, Academic Press, Inc.
65.
Nicholls, A., K.A. Sharp, and B. Honig (1991). "Protein
Folding and Association: Insights From the Interfacial
and Thermodynamic Properties of Hydrocarbons",
Proteins, 111, pp. 281-296.
66.
Norde, W. (1992). "Energy and entropy of protein
adsorption", J. Dispersion Science and Technology, 13,
No. 4, pp. 363-377.
67.
Novokhatny, V.V., S.A. Kudinov, and P.L. Privalov
(1984). "Domains in Human Plasminogen", J. Mol. Biol.,
179, pp. 215-222, Academic Press Inc. Ltd., London.
118
68.
Nygren, H. and M. Stenberg (1988). "Molecular and
supramolecular structure of adsorbed fibrinogen and
adsorption isotherms of fibrinogen at quartz surfaces",
J. Biomed. Mat. Res., 22, pp. 1-11, John Wiley & Sons,
Inc.
69.
Nygren, H., M. Stenberg, and C. Karlsson (1992).
"Kinetics supramolecular structure and equilibrium
properties of fibrinogen adsorption at liquid-solid
interfaces", J. Biomed. Mater. Res., 26, pp. 77-91,
John Wiley & Sons, Inc.
70.
Pace, C.N. (1983). "Protein Conformations and Their
Stability", JAOCS, 60, No. 5, pp. 970-975.
71.
Pace, N.C. and C. Tanford (1968). "Thermodynamics of
the Unfolding of fl-Lactoglobulin A in Aqueous Urea
Solutions between 5 and 55", Biochemistry, 7, No. 1,
pp. 198-207.
72.
Parfitt, G.D. and C.H. Rochester (1983). "Adsorption
from Solution at the Solid/Liquid Interface, pp.
153-218, Academic Press, London.
73.
Pfeil, W. (1981). "Thermodynamics of proteins",
Biophysical Chemistry, 13, pp. 181-186.
74.
Pilz, I. and G. Czerwenka (1973). "The Partial Specific
Volume of Various Proteins and its Dependence on
Concentration and Temperature of the Solution", Die
Makromolekulare Chemie, 170, pp. 185-190.
119
75.
Privalov, P.L. (1976). "Thermodynamic Investigations of
Biological Macromolecules", Pure & Appl. Chem., 47, pp.
293-304, Pergamon Press.
76.
Privalov, P.L. (1979). "Stability of Proteins", Advan.
Prot. Chem., 33, pp. 167-241, Academic
Press, New
York, San Francisco and London.
77. Privalov, P.L. and G.I. Makhatadze (1990). "Heat
Capacity of Proteins. II. Partial Molar Heat Capacity
of the Unfolded Polypeptide Chain of Proteins: Protein
Unfolding Effects", J. Mol. Biol., 213, pp. 385-391,
Academic Press Limited.
78.
Privalov, P.L. and L.V. Medved (1982). "Domains in the
fibrinogen molecule", J. Mol. Biol., 159, pp. 665-683.
79. Privalov, P.L. and N.N. Khechinashvili (1974). "A
Thermodynamic Approach to the Problem of Stabilization
of Globular Protein Structure: A Calorimetric Study",
J. Molec. Biol., 86, pp. 665-684.
80.
Privalov, P.L., Yu V. Griko, and S. Yu. Venyaminov
(1986). " Cold denaturation of Myoglobin", J. Mol.
Biol., 190, pp. 487-498, Academic Press, Inc. (London)
81. Ruzgas, T.A., V.J. Razumas, and J.J. Kulys (1992).
"Sequential Adsorption of 7-Interferon and Bovine Serum
Albumin on Hydrophobic Silicon Surfaces", J. Coll.
Interf. Sci., 151, No. 1, pp. 136-143, Academic Press,
Inc.
120
82.
Schulz, G.E. and R.H. Schirmer (1979). "Noncovalent
Forces Determining Protein Structure", In: Principles
of Protein Structure, p. 27-45., Springer-Verlag, New
York and Heidelberg.
83.
Shibata, C.T. and A.M. Lenhoff (1992). "TIRF of Salt
and Surface Effects on Protein Adsorption.
1.
Equilibrium", J. Coll. Interf. Sci., 148, No. 2, pp.
469-484, Academic Press, Inc.
84.
Shirahama, H., J. Lyklema, and W. Norde (1990).
"Comparative Protein Adsorption in Model Systems", J.
Coll. Interf. Sci., 139, No. 1, pp. 177-187, Academic
Press, Inc.
85.
Singer, S.J. (1948). "Note on an Equation of State for
Linear Macromolecules in Monolayers", J. Chem. Phys.,
16, pp. 872-876.
86.
Slejko, F.L. (1985). "Adsorption Technology: A
Step-by-Step; Approach to Process Evaluation and
Application", pp. 1-6, Marcel Dekker, Inc., New York
and Basel.
87.
Sober, H.A. (1970). "Handbook of Biochemistry", CRC,
The Chemical Rubber Co., Cleveland.
88.
Stryer, L.
(1988). Biochemistry, W.H. Freeman and
Company, New York.
89.
Suri, S. K.
(1970). "Adsorption from Solution-
Association of Nitromethane Molecules at the
Interface", J. Coll. Interf. Sci., 34, No. 1, pp. 100­
102, Academic Press, Inc.
121
90.
Suttiprasit, P. and J. McGuire (1992). "The Surface
Activity of a-Lactalbumin, /3- Lactoglobulin and Bovine
Serum Albumin. II. Some Molecular Influences on
Adsorption to Hydrophilic and Hydrophobic Silicon
Surfaces", J. Colloid Interface Sci., 154, No. 2, pp.
327-336.
91.
Suttiprasit, P., V. Krisdhasima, and J. McGuire (1992).
"The Surface Activity of a-Lactalbumin, fl-Lactoglobulin
and Bovine Serum Albumin. I. Surface Tension
Measurements with Single-Component and Mixed
Solutions", J. Colloid Interface Sci., 154, No. 2, pp.
316-326.
92. Tanford, C. (1970). "Protein Denaturation. Part C.
Theoretical Models for the Mechanism of Denaturation",
Advan. Prot. Chem., 24, pp. 1-95.
93.
Ter-Minassian-Saraga, L. (1981). "Protein Denaturation
on Adsorption and Water Activity at Interfaces: An
Analysis and Suggestion", J. of Colloid and Interface
Science, 80, No. 2., pp. 393-401, Academic Press, Inc.
94.
Thornton, Janet M.
in Proteins",
95.
(1982). "Electrostatic Interactions
Nature, 295, pp. 13-14.
Tischenko, V.M., V.P. Zav'yalov, G.A. Medgyesi, S.A.
Potekhin, and P.L. Privalov (1982). "A Thermodynamic
Study of Cooperative Structures in Rabbit
Immunoglobulin G", Eur. J. Biochem. (FEBS), 126, pp.
517-521.
122
96.
Uraizee, F. and G. Narsimhan (1991). "A Surface
Equation of State for Globular Proteins at the Air-
Water Interface", J. Colloid Interface Sci., 146, No.
1, pp. 169-178.
97.
van Oss, C.J. and R.J. Good (1988). "Orientation of the
Water Molecules of Hydration of Human Serum Albumin",
J. Protein Chem., 7, No. 2, pp. 179-183, Plenum
Publishing Corporation, New York.
98. van Oss, C.J., D.R. Absolom, A.W. Neumann, and W. Zingg
(1981). "Determination of the surface tension of
proteins. I. Surface tension of native serum proteins
in aqueous media", Biochimica et Biophysica Acta, 670,
pp. 64-73, Elsevier/North-Holland Biomedical Press.
99. Velicelebi, G. and J.M. Sturtevant (1979).
"Thermodynamics of the Denaturation of Lysozyme in
Alcohol-Water Mixtures", Biochemistry, 18, pp. 1180­
1186.
100. Wei, A.-P., J.N. Herron and J.D. Andrade (1990). "The
Role of Protein Structure in Surface Tension Kinetics"
in: Biotherapie (D.J.A. Crommelin, editor), In press.
101. Winterton, L.C., J.D. Andrade, J. Feijen, and S.W. Kim
(1986). "Heparin Interaction with Protein-Adsorbed
Surfaces", J. Colloid Intreface. Sci., 111, No. 2, pp.
314-342., Academic Press, Inc.
123
102. Woodhouse, K.A., P.W. Wojciechowski, J.P. santerre, and
J.L. Brash (1992). "Adsorption of Plasminogen to Glass
and Polyurethane Surfaces", J. Colloid Interface Sci.,
152, No. 1, pp. 60-69., Academic Press, Inc.
103. Zhukovskii, A.P. and N.V. Rovnov (1989)."Change in Heat
Capacity on Denaturing of Proteins", Biophysics, 34,
No. 3, pp. 560-561, Pergamon Press plc.
APPENDICES
124
APPENDIX 1
PROGRAM GAMA12
C
C.. A PROGRAM TO CALCULATE:
C
1) GAMA12 (INTERFACIAL TENSION)
C
FROM VARIABLE INPUTS
C
1) GAM1V "INTERFACIAL TENSION OF PHASE 1" (mJ/m2)
C
2) GAM2V "INTERFACIAL TENSION OF PHASE 2" (mJ/1112)
C
***********************************************************
C
IMPLICIT DOUBLE PRECISION (A-H2O-Z)
C
C
5
93
94
95
96
97
98
CHARACTER*80 NSURF,OUTFILE
PRINT*,,****************************************,
PRINT*,'ENTER THE VALUES OF GAM1V AND GAM2V '
READ(*,*) G1V,G2V
IF (G2V.LE.0.0) GO TO 5
CALL EQS(G1V,G2V,G12)
PRINT*,' SURFACE TENSION OF PHASE 1 =
',SNGL(G1V)
PRINT*,' SURFACE TENSION OF PHASE 2 =
',SNGL(G2V)
PRINT*,' INTERFACIAL SURFACE TENSION G12 = ',SNGL(G12)
CONTINUE
STOP
END
SUBROUTINE EQS(SSV,SLV,SSL)
IMPLICIT DOUBLE PRECISION (A-H2O-Z)
A=0.0150
EPS = 0.0001
GAMSV=SSV
GAMLV=SLV
IF (GAMSV.LE.GAMLV) GO TO 93
GAMSV=SLV
GAMLV=SSV
IF (GAMLV.GT.30.0) GO TO 95
GAMSL=((DSQRT(GAMLV)-DSQRT(GAMSV))**2.0)/
1(1.0-A*DSQRT(GAMSV*GAMLV))
GO TO 99
IF (GAMLV.GT.50.0) GO TO 97
GAMSL=((DSQRT(GAMLV)-DSQRT(GAMSV))**2.0)/
1(1.0-A*DSQRT(GAMSV*GAMLV))
IF ((GAMSL-GAMSV).GT.0.0) GO TO 99
GAMSL=(((2.0-A*GAMSV)*DSQRT(GAMLV)-DSQRT((2.0-A*GAMSV)
1**2.0*GAMLV-4.0*(GAMLV-GAMSV)))**2.0)/4.0
GO TO 99
GAMS1=10.0D0
B=DSQRT(GAMLV)
GAMS = GAMS1
125
C = DSQRT(GAMS)
SLOPE = ((1.0-A*B*C)*(1.0-B/C)-(GAMS-2.0*B*C+GAMLV)
1*((-A/2.0)*B/C))/(1.0-A*B*C)**2.0
SLOPE1 = SLOPE + 1.0
U = 1.0-A*B*C/2.0+(A*(GAMLV**1.5)/2.0-B)/C
V = (1.0-A*B*C)**2.0
DU=-(A*B)/(4.0*C)-(A*(GAMLV**1.5)/2.0-B)/
1(2.0*(GAMS**1.5))
DV=-A*B/C+(A*B)**2.0
SLOPE2=(V*DU-U*DV)/V**2.0
GAMS1=GAMS-SLOPE1/SLOPE2
IF (ABS(GAMS1-GAMS).GT.EPS) GO TO 98
GAMS2=((DSQRT(GAMS1)-DSQRT(GAMLV))**2.0)/
1(1.0-A*DSQRT(GAMS1*GAMLV))
IF (GAMSV.LE.GAMS1) GO TO 94
IF (GAMSV.GE.GAMS2) GO TO 96
GAMSL = GAMS1+GAMS2-GAMSV
99
SSL=GAMSL
RETURN
END
126
APPENDIX 2
The values of A H', the non-hydrophobic enthalpy of a
protein, as estimated by equating Eq.
solving for A H'.
(13) with Eq.
(14) and
The average value of molecular weight of
an amino acid residue is assumed to be equal to 123
(Baldwin, R.L. 1986), and that of ACp to be equal to 50
J/mol amino acid residue
Protein
K (Zhukovskii & Rovnov, 1989).
Molecular
A H'
Weight
(kJ/mole)
(kJ/mol.res.)
691.739
5.992
1796.817
6.032
a-Lac
14200.
0-Lag
36640.
A Ha
(dimer)
B S A
66267.
3229.121
5.994
H S A
69000.
3363.049
5.995
Myog
17800.
864.906
5.977
RiboA
13680.
686.568
6.173
Lyso
14400.
754.056
6.441
ChA
23000.
1145.179
6.124
IgG
150000.
7264.512
5.957
Average
6.076 ± 0.155
127
APPENDIX 3
The molecular dimensions of proteins used to develop an
equation for ALI, (Eq. 17), for a monolayer with the side-on
conformation.
Molecular
Dimensions
Side-on
Weight
(A3)
(A2)
37 X 32
Protein
a-Lac
14161.
37 X 32 X 25
0-Lag
36640.
69.3 X 35.8
(dimer)
(two spheres)
B S A
66267.
140 x 38 x 38
140 x 38
H S A
69000.
115 x 40 x 40
115 x 40
Myog
17800.
44 x 35 x 25
44 x 35
RiboA
13680.
38 x 28 x 22
38 X 28
Lyso
14400.
45 x 30 x 30
45 x 30
Cytochrome C
11353.
37 x 25 x 25
37 x 25
40 x 38 x 36
40 x 38
235 x 44 x 44
235 x 44
69.3 X 35.8
Super Oxide
15534.
Dismutase
7-globulin
160000.