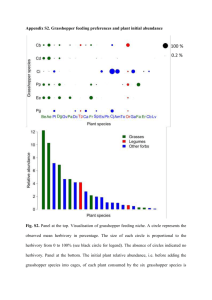
Grasshopper - Vegetation Relationships on Montana Grasslands by NORMAN LEWIS ANDERSON
advertisement

Grasshopper - Vegetation Relationships on Montana Grasslands by NORMAN LEWIS ANDERSON A thesis submitted to the Graduate Faculty in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY in Entomology Montana State University © Copyright by NORMAN LEWIS ANDERSON (1962) Abstract: The general distribution of 65 grasshopper species on Montana grasslands east of the Rocky Mountains is recorded. This distribution is discussed primarily from the standpoint of known food-plant preferences of grasshopper species involved. The results of detailed investigations of grasshopper-vegetation relationships on 105 native grassland and abandoned field study sites are presented and discussed. It was concluded that vegetation plays an important role in the selection of areas of occupancy by grasshoppers within a grassland habitat but is not a causative agent responsible for initial changes in grasshopper abundance. GRASSHOPPER - VEGETATION RELATIONSHIPS ON MONTANA GRASSLANDS by NORMAN LEWIS ANDERSON, JR. A thesis submitted to the Graduate Faculty in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY in Entomology Approved: . I X/'/ ^ Heady Major Department Chairman, EyaAining Committee D e a n , Graduate/Division MONTANA STATE COLLEGE Bozeman, Montana June, 1962 ACKNOWLEDGMENT The writer is deeply grateful to Doctor J 9 H. Pepper, Head,' and Professor E 9 B 9 Hastings of the Department of Zoology and Entomology, Montana State College, for their stimulating discussions, interest, and encouragement during all phases of the investigation. Sincere thanks are extended to Doctors Wo E 9 Booth, J 9 H 9 Rumely, and J 9 C . Wright, Department of Botany and Bacteriology, Montana State College, for help in plant identification and for field aid in making various vegetation measurements. iv TABLE OF CONTENTS LIST OF TABLES 'O 0 0 0 9 0 0 0 0 » ^ O » t > O 6 4 O O O « a O O O 0 « # t t O 0 O O o a O 6 * O 4 0 9 O O vi O LIST OF FIGURES... ...................................... ABSTRACT 0 0 0 0 0 9 0 0 0 0 0 INTRODUCTION o 0 0 0 9 0 0 9 9 0 0 0 0 0 0 0 9 0 9 9 0 0 0 0 0 0 0 0 9 0 0 9 9 0 0 0 0 0 vii .viii o e o o o o o o o o e d o o o e o o c o o o o o o e o o e o e e o o o o e o o o o e o o o METHODS OF STUDY... I 0 0 0 0 0 9 0 0 0 0 0 0 0 , 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 9 0 0 GRASSHOPPER SPECIES DISTRIBUTION. 0 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Acrolophitus hirtipes- (Say)..... .. Aeloplus -turnbulli turnbulli (Thomas) Aerochoreutes carlinianus carlinianus (Thomas) Aerbpedellus clavatus (Thomas) Ageneotettix deorum TScudder). Amphitornus coloradus (Thomas). Arphia conspersa Scudder Arphia pseudonietana pseudonietana (Thomas) Aulocara elliotti (Thomas) Boopedon nubiIum'(Say). Brachystola magna (Girard) o o o o o o o o o o o o o Bradynotes obesa (Thomas) Bruneria brunnea (Thomas) Bruneria sordida (McNeill) Camnula pellucida (Scudder) Ghorthippus longicornis (Latreille). Chortophaga viridifasciata (De Geer) Circotettix rabula rabula Rehn and Hebard CordlIlacris crenulata (Bruner) Cratypedes negle'ctus (Thomas) Dactylotum picturn TThomas) Derotmema haydenii haydenii (Thomas) Dissosteira Carolina (Linnaeus) Drepanopterna femoraturn (Scudder) Encoptolophus sordidus costalis (Scudder) Eritettix simplex tricarinatus (Thomas) Hadrotettix trifasciatus (Say) Hespefotettix viridis pratensis (Scudder) Hlppiscus rugosus (Seudder) Hypochlora alba (Dodge) MeIanoplus alpinus (Scudder) Melanoplus bilituratus (W a l k e r . Melanoplus bivittatus (Say)................... Melanoplus bowditehi eanus (Hebard) Melanoplus bruneri (Scudder) 0 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 ' 0 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 9 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0-00099 0 0 0 0 0 0 0 0 0 0 0 9 0 0 O O O O O O 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 9 0 0 0 O 0 0 9 0 9 0 0 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 9 O O O O O . 0 0 0 0 0 9 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 0 19 20 20 21 21 21 22 22 23 O O O O O O 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 0 9 0 0 9 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 9 9 0 0 0 0 0 0 00 0 0 0 0 0 0 0 0 0 0 0 0 9 0 9 0 0 0 0 0 0 9 0 9 0 0 0 0 0 0 0 0 0 9 9 0 9 12 12 13 13 14 14 15 15 15 16 17 17 IS IS IS 0 0 0 0 0 0 0 0 9 0 0 0 0 12 23 0 0 0 0 ' 23 24 24 25 25 25 26 27 27 27 Y Melanoplus confU.SU.S (S c"u.dd6ir)« » •» Melanoplus dctwsOXii (Scu.dd.03r) @000*000****00*0*00 Melanoplus differentialis (Thomas ......... Melanoplus femur-rubrum femur-rubruim (Be Geer)...... Melanoplus Kladstoni (Scudder).......... Melanoplus infantilis (Scudder)................•.... Melanoplus keeleri luridus ( D o d g e ) • Melanoplus kennicotti (Scudder Melanoplus montanus (T h o m a s Melanoplus occidentalis occidentalis (Thomas)...... Melanoplus oregonensis (Thomas)............... Melanoplus __________ packardii (Scudder)................ Mermiria maculipennis macelungi (Rehn)............... Metator pardalinus (Saussure)............ Neopodismopsis abdominalis (Thomas).. =........... Opei a -0bsoura (Thomas).............................. Orphulella pelidna pelidna (Burmeister)............. Orphulella speciosa (Scudder)...................... Phlibostroma quadrimaculatum (Thomas)............... Phoetaliotes nebrascensis ( T h o m a s Pseudopomala brachyptera ( S c u d d e r . Psoloessa delicatula delicatula (Scudder)........... Schistocerea lineata (Scudder)...................... Spharagemon collare (Scudder)....................... Spharagemon equals (Say) ...... Trachyrhachys kiowa kiowa (T h o m a s Trimerotropis bruneri (McNeill)........... Trimerotropis campestris (McNeill).................. Trimerbtropis cincta ( T h o m a s Ianthippus corallipes (Haldeman).................... o ) ADDITIONAL RESULTS AND DISCUSSION SUMMARY 0 0 0 0 0 0 . o o . o . * o o o o # o e c * o o o o © . . . . . . . . . . . . . . . . o e c o e o o t i o o o o o e o o o o o o o o o o o o e e o o o o o o o o o o o LITERATURE CITED..... . » 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2S 2^ 29 30 30 30 31 31 32 32 32 33 33 34 34 33 35 36 36 37 38 38 39 39 40 41 41 41 42 42 43 66 63 LIST OF TABLES TABLE I A summary of study site vegetation measurements TABLE II Per cent frequency of occurrence of grasshopper species in numbers greater than I per sq yd on Native Grassland Study Sites. (Perennial Grass-Cover Classes).... TABLE III Per cent frequency of occurrence of grasshopper species in numbers greater than I per sq yd on Abandoned.Field Study Sites. (Perennial Grass-Cover. Classes).... TABLE IV, Per cent frequency of occurrence of grasshopper species in numbers greater than I per sq yd on Native Grassland Study Sites. (Total Foliage-Cover Classes)...... TABLE V Per cent frequency of occurrence of grasshopper species in numbers greater than I per sq yd on Abandoned Field Study Sites. (Total Foliage-Cover Classes)....... TABLE VI Per cent frequency of occurrence of grasshopper species in numbers greater than I per sq yd on Native Grassland Study Sites. (Litter-Cover Classes)..... . TABLE VII Per cent frequency of occurrence of grasshopper ■ species in numbers greater than I per sq yd on Abandoned Field Study Sites. (Litter-Cover Classes TABLE VIII Vegetation on 4 permanent square-meter quadrats at the OW Ranch, Decker, Montana.............. - vii LIST OF FIGURES 1 FIGURE I. Study Site Locations FIGURE 2. Vegetative Types.......... ...................... 5 FIGURE 3 * Seasonal Fluctuations in the Grasshopper Population of One Area As'Measured in 1958...... 45 > ...... ....................4 viii.- ABSTRACT The general distribution of 65 grasshopper species on Montana grasslands east of the Rocky Mountains is recorded. This distribution is discussed primarily from the standpoint of known food-plant preferences of grasshopper species involved. The results of detailed investigations of grass­ hopper-vegetation relationships on 105 native grassland and abandoned field study sites are presented and discussed. It was concluded that vegetation plays an important role in the selection of areas of occupancy by grasshoppers within a grassland habitat but is not a causative agent responsible for initial changes in grasshopper abundance. I INTRODUCTION The history of grasshoppers on grasslands is largely one of fluctuating numbers. In a particular area the appearance of these, insects in low numbers for a period is followed occa­ sionally by an extreme increase in the population density. When such increases in several localities coincide the desig­ nation "grasshopper outbreak" is often assigned to the total area. Even during these periods, however, there are areas within such a region in which populations remain.low. As yet, such population fluctuations are not predictable as to time or .location. One of the environmental factors which appears to be of importance in influencing grasshopper distribution and numbers is vegetation, but critical understanding of grasshoppervegetation interrelationships is not fully revealed in the literature. For example, there are differing opinions con­ cerning the importance of vegetation composition from the standpoint of food. Morse (1899) stated that grasshoppers "...are voracious, their food being general in character rather than special, a question of quantity rather than qual­ ity." Since that time, however, food-plant preferences have been recognized and Isely (1938) found that, under cage condi­ tions, those species which are selective feeders will starve if they are deprived of their host-plants even in the presence of abundant fresh vegetation, palatable to other grasshopper 2 species. On the other hand, Williams (1954) found that "...when grasshoppers were deprived of water as well as food for 4& hours, they could be induced to feed on a number of succulent plants which previously they would only slightly nibble." Other workers (Clark 1947a, Rubtzov 1934, Strohecker 1937, Urquhart 1941) have generally concluded that particular food-plants are not limiting and that other environmental factors such as the amount of green feed and cover, micro­ climatic conditions, sunlight, moisture, and rate of evapo­ ration are more important. On grassland areas of Russia, workers (Bei-Bienko 1930, Nefedov 1931, Rubtzov 1932, 1934) have found direct correla­ tions between the abundance of certain grasshopper species and the taxonomic constitution and amount of plant cover. Kachaeva Sc Rubtzov (1935) state that "...in different habitats a decrease in the number of species and an increase in numbers of individuals of grasshoppers were parallel to a decrease in the number of species in plant associations and in the per- . centage of plant cover." In England, Clark (1940) found that: "Height and density of the vegetation are probably the most important features for grasshoppers." Australia: On the other hand, Clark (1947) found that in "The amounts of shelter vegetation and green feed are of greater importance than the amount of bare ground (or total plant cover)...". In this case, "shelter height", as - 3 distinguished from absolute plant height was defined as " the height of that part of a single plant or aggregate which cast practically unbroken shade on the ground." The main objectives of the study reported here were two­ fold: (i) to determine the distribution of grasshoppers on the grasslands of Montana east of the Rocky Mountains, and (ii.) to learn something of the role vegetation may play in determining grasshopper distribution and numbers. METHODS OF STUDY Study sites were located within specific grassland areas east of the Rocky Mountains (Fig. I). At the start of the study it was believed that a general map of vegetation types, such as that of He!burn, Edie, & Lightfoot (1962) (Fig. 2), would suffice in the selection of areas for detailed study. It was found upon using this map that a number of distinctive grassland communities existed within each of the broad vegeta­ tion-type categories used. It was felt that these commu­ nities differed enough from one another to warrant independent consideration from the standpoint of grasshopper-vegetation relationships. Consequently, soil survey and geological for­ mation maps were consulted extensively in selecting as many different soil types and resultant grassland communities for study as possible. Thirteen grassland community types, including abandoned fields, were subsequently recognized. fig I STUDY SITE LOCATIONS SOURCE Vegetative Rangeland Types In Montana, Montana Agricultural Experim ent Station, Bozeman, Montana, 1949. fig 2. VEGETATION TY PES (FROM://£!BURN, ED/E, 8 UGHTFOOT, 1962) With the exception of abandoned fields, these vegetation types were recognized primarily on the basis of the .dominant species present. A detailed vegetation analysis and description of these types and the study sites within them is the subject of a forthcoming paper. designated as follows: The recognized types investigated may be (I) Festuca scabrella Type, including study sites I and 2; (2) Festuca idahoensis Type, including study sites 3-7; (3) Agropyron spieatum Type, including study sites 8-12; (4) KbeTeria-Stipa-Agropyron Type, including study sites 13-22; (5) Stipa-Agropyron-Bouteloua Type, including study sites 23-35; (6) Bouteloua-Carex-Stipa Type, including study sites 36-46; (7) Bouteloua-Stipa Type, including study 47-58; (8) Buchloe-Bouteloua-Agropyron Type, including study sites 59-62; (9) Bouteloua-Agropyron Type, including study sites 63-70; (10) Agropyron smithii Type, including study sites 71-75; ( H and 12) Artemisia tridentata and Sarcobatus vermiculatus Types, including study sites 76-79; and (13) Abandoned Fields, including study sites 80-105. Within each site selected for study a detailed investi­ gation was made of the vegetation and the grasshoppers pres­ ent. A "point-plot" method of studying vegetation was employed at each study site. A set of 10 steel surveyor's arrows, 14 % 3/l6 in, were held in place 4 in apart along a wooden board. This apparatus was placed 50 times at random within each site.■ Each "hit" of a plant taxon at ground level - I - by an arrow-point was recorded and contributed to the measure of basal cover and percentage composition by each taxon. At a few sites the estimate of percentage composition was based upon the establishment of 4-sq-ft list quadrats in which indi­ viduals of each plant species were counted and recorded. When an arrow struck litter it was recorded and contributed to the measure of the amount of ground covered by litter. Total foliage cover was measured by recording any touch by plant foliage on an arrow above its point. A summary of the vege­ tation measurements for all study sites is presented in Table I. Field records of species occurrence, taken over a period of 12 years, were used in establishing the general grasshopper species distribution. At some of the study sites it was pos­ sible to obtain quantitative data on grasshopper populations present by use of bottomless cages placed at night. This "night cage" method of sampling has been described previously by Anderson & Wright (1952) and Anderson (1961). At most study sites, however, grasshopper population density was merely estimated and recorded along with a list of species observed. Approximate order of species abundance was deter­ mined from sweep-net collections. The identification of all grasshopper species was based primarily upon the reference collection at Montana State College. TABLE I. A summary of study site vegetation measurements. % Composition % Basal Cover Study Site I. 2. 3. 4« 5. 6. 7. 8. 9. 10. 11. 12. 13. 14« 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 2$. 26. 27. 28. Grasses "" & ' Sedges Forbs & ’' Shrubs 41 30 33 34 Total 16 20 16 71 59 48 41 52 33 31 33 36 21 19 32 6 26 15 20 29 5 4 11 • 4 6 5 19 14 4 5 2 5 6 I 35 34 19 26 14 15 8 14 21 26 44 19 10 11 19 5 5 22 17 16 14 15 25 24 25 20 14 16 36 18 18 17 17 24 • ' 36 28 31 25 33 30 40 23 20 22 23 25 Grasses & ' Sedges 58 44 29 36 15 42 67 67 48 76 73 50 76 43 85 81 70 86 79 79 42 52 90 79 92 76 75 97 Forbs : : % Foliage Cover % Litter Shrubs : 42 56 71 64 85 58 33 33 52 24 27 50 24 57 15 19 30 14 21 21 58 48 10 21 8 24 25 3 77 . 47 79 37 36 72 8 21 42 24 11 16 65 21. 20 16 27 15 11 4 44 48 56 20 33 30 53 17 100 66 69 65 31 66 32 28 . 39 21 8 19 49, 37 27 15 47 35 3 . 21 22 25 65 8 2 26 50 15 TABLE I. (Continued) 29, 30, 31. 11 40 32. 33: 34. 35. 75 65 15 36. 37. 38. 39. 40. 41. 42. 43. 44. 45. . 46. 47. 48. 49. 50. 51. . 52. 53. 5k. 73 17 8 5 2 I 6 3 6 I 3 8 3 7 7 6 8 I 2 I 4 3 7 8 3 2 11 10 7 39 20 71 3 7 5 I 3 3 48 58 38, 32 29 21 11 39 17 25 51 19 69 31 26 35 9 17 71 51 21 52 56. 57. 58. 5960. 6l. 62. 63. 36 41 23 38 33 28 48 53 77 66 21 61 44 33 32 39 83 90 97 97 94 95 87 97 91 29 72 ' 14 46 24 31 59 77 85 72 20 95 71 98 96 87 32 30 38 .16 25 74 53 32 62 80 59 74 . 43 46 '24 41 36 81 87 93 56 67 96 98 66 84 91 66 95 85 89 95 93 92 61 17 10 3 3 6 5 13 3 9 28 23 15 28 19 13 5. 2 4 13 7 44 33 4 2 34 16 9. ■ 34 5 . 15' 11 5 7 8 53 6 8 28 60 81 9% 65 39 54 43 17 66 67 2 20 22 14 15 12 8 2 3 . I 2 10 50 46 6 48 23 2 20 33 I 5 3 I I 2 32 62 3 3 7 8 2 2 3 I 2 2 I 2 I 13 I 6 I 2 24 23 12 15 I 41 4 18 6 2 1 vO I TABLE I, (Continued) 22 64» 65. 66. 13 61 67. 68: 69. 70. 71. 72. 73. 74. 75. 76. 77. 78. 79. 80. 81. 82. 83. 84. 85. 86* % ' 88. 89. 90. 91. 92. 93. ' 64 72 34 19 11 12 9 12 30 9 13 3 4 42 19 25 6 14 18 13 14 17 21 23 29 12 56 94« 15 95. 96. 97. 98. 23 69 92 64 I 23 96 5 9 18 70 73 I 65 82 10 ■ 19 .24 2 3 11 11 3 32 22 20 23 2 36 53 43 13 15 20 23 33 41 34 23 27 44 55 21 19 13 25 9 I 46 I I 18 22 19 27 24 3 27 I 42 I 6 I 25 27 43 22 15 56 36 59 42 24 70 98 65 87 99 88 64 44 4 27 13 I 12 36 56 86 14 78 44 53 91 ' 22 20 37 14 14 95 40 54 25 52 41 60 99 95 99 55 52 33 95 36 99 99 95 98 56 47 9 80 63 86 86 5 60 46 75 48 59 40 I 5 I 45 48 67 5 64 I I 5 2 2 8 33 9 23 55 48 35 62 51 2 2 I I I 80 19 17 22 33 37 45 31 43 5 7 40 26 35 9 5 64 89 8 41 31 2 23 28 15 20 65 76 32 54 35 58 38 76 75 85 65 2 41 91 23 17 50 16 5 50 2 2 2 20 78 79 50 TABLE I. (Concluded) 99. 100. 101. 102. 103. 104; 105. 29 31 22 13 6 9 2 22 27 6 27 31 3 25 ' 51 58 28 40 37 12 27 57 54 79 16 75 7 43. 46 21 67 84 25 93 33 51 85 54 51 58 35 28 60 79 55 39 5 2 I H H I 12 GRASSHOPPER SPECIES DISTRIBUTION The following grasshopper species were encountered during the course of the study. Unless otherwise indicated, each species was recorded in field notes during the past 12 years from all counties east of the Rocky Mountains. Acrolophitus hirtipes (Say) Field notes record the presence of this species in every county except Beaverhead. It was found on study sites 3, 5 » 19, 29, 31, 34, 39, 50, 59, 72, 73, 74, and 96. A. hirtipes was never found in high numbers and seemed to be restricted to areas of sparse vegetation. In accordance with previously recorded food-plant preferences (Griddle 1933, Anderson & Wright 1952) it was always associated with the presence of members of the plant family Boraginaceae. Aeoloplus turnbulli turnbulli (Thomas) This species was recorded as being present on study sites 63, 65, 72, 73, 76, and 77« numerous. At no time was it considered The usual situation was to find from one to several pairs congregated in a relatively small area and invariably associated with members of the plant family Chenopodiaceae (Griddle 1933, White & Rock 1945, Anderson & Wright 19.52). Because of the low numbers usually present and the localized distribution of this species it is possible that A. turnbulli was present on more of the study sites than recorded. It is rather surprising that it was not recorded from a number of the abandoned fields in which members of the Ghenopodiaceae were commonly encountered. Aerochoreutes carlinianus carlinianus ■(Thomas ) A* carlinianus was recorded from study sites 10, 29, 39, 72, 73, and 74. Although Griddle (1.933) and Brooks (1953) record this species as one which feeds mainly on Agropyron smithii there was no indication of any association with a particular plant or group of plants. On the contrary, the adults were most commonly found during the heat of the day hovering over bare ground and rocky outcroppings. Aeropedellus clavatus (Thomas) A. clavatus was one of the most commonly found species, occurring in numbers greater than I per sq yd in both native grassland areas and abandoned fields. It was listed from study sites 2-6, 8, 9, 13-20, 23-29, 31, 39, 41, 42, 47-49, 51, 59-61, 71-74, 76, 79, 82, 85, and 86. It may not have been encountered on some sites since it is an early-hatching species which has normally oviposited and disappeared by midJuly. Although there appears to be some association between this species and Koeleria cristata it is believed that the association is one involving the availability of green, succu­ lent grass which is satisfied by K. cristata early in the season. A. clavatus is known to feed on several grass species (Anderson & Wright 1952). — li). — Ageneotettix deorum (Scudder) A. deorum was listed on study sites 4» 12-21, 28-31# 33, 34, 37, 39, 40, 43, 44, 46, 47, 49, 50, 53, 54, 56-67, 70-75, 80-85, 87-89, 93, 95-101, and 104. It occurred in numbers greater than I per sq yd on both native grassland and abandoned fields. It appeared to inhabit a variety of habi­ tats but is most common and often abundant in areas of short grass and bare ground. Dry conditions are particularly favorable to this species (Allred 1941)« From the standpoint of food-plant preference A. deorum can be considered a grassfeeding species with little or no specific plant preference (Griddle 1933; Anderson & Wright 1952). It includes consider­ able dry material in its diet as an adult and can be in fairly high numbers without inflicting measurable damage to green vegetation (Anderson 1961). Amphitornus coloradus (Thomas) This species was recorded from study sites 4, 8, 9, 11, 12, 14, 15, 17-20, 28-33, 36, 37, 39, 44, 50, 55-57, 59, 64-66, 68, 70-74, 76, 80-82, 87, 88, 9#, and 100. At times it was found in numbers greater than I per sq yd on both native grassland and abandoned field study sites. This grass- feeding species (Griddle 1933, Ball et al. 1942, Anderson & Wright 1952, Brooks 1958) was most common in areas where there was a mixture of Bouteloua gracilis and Stipa comata. It not only feeds on these two grass species but also climbs upon and - 15 - 'remains inactive on the taller S . comata for long periods'. Arphia conspersa Scudder A. conspersa was listed on study sites 29, 32, 37-39, 55, 56, 60, 61, 66, 63, 70, 72-74, 79, 91, and 102. It over­ winters in one of the later nymphal instars and appears as an adult early in the spring. -Consequently, it may have ovi­ posited and disappeared before many of the study sites were established. This grass-feeding species was not found associ­ ated with any specific food-plants and has not been noted in abundance. It appeared to be most common in dry upland situ­ ations on sandy soil and often in association with open stands of Pinus ponderosa. Arphia pseudonietana pseudonietana (Thomas) A. £0 pseudonietana was listed in low numbers from study sites 7, 12, 17, 29, 33, 34, 36, 39, 41, 46-43, 52-55, 57, 66, 63, 72-74, 33, 39, 94, and 95. Although this spring-hatching species is one of the grass-feeders (Anderson & Wright 1952), it was not found associated with a specific food-plant or group of food-plants. A. p. pseudonietana and A. conspersa appeared to occupy similar habitats in Montana with A. p. pseudonietana being more commonly encountered at lower eleva­ tions and most numerous in areas of mixed short- and mid-grass vegetation. Aulocara elliotti (Thomas) This species was listed from study sites 9, 11, 14-13, - 16 - 22, 29, 30, 37, 39, 40, 44, 50, 54, 56, 57, 64-66, 71-75, 83, 84, 87, 89, 91, 93-95, 100, 102, and 104, A. elliotti may be considered the most abundant and injurious of the species found oh Montana grasslands year after year. During the study period it appeared in numbers greater than I per sq yd on both .native grassland and abandoned field sites. It is a grass- feeding species (Griddle 1933, Ball et al. 1942, Pfadt 1949, Anderson & Wright 1952, Brooks 1958) which shows definite preference for Agropyron spp. It is often found to be very abundant in stands of Agropyron smithii or in fields planted to Agropyron cristatum. It may also be numerous in areas, where A. smithii is but a minor part of the total vegetation complex. The local distribution of A. elliotti on native grasslands or medium-aged abandoned fields is usually gov­ erned by the distribution of A. smithii, with the greatest concentrations found where the density of A. smithii is highest. Boopedon nubiIurn (Say) B. Montana. nubilum has been encountered on few occasions in During the past 12 years it has been found in all counties north of the Missouri River and east of the eastern edges of Toole, Pondera and Teton counties. It has also been found in counties south of the Missouri River which lie east of Wheatland county. 15 and 71. It was listed from only 2 study sites, According to Ball et al. (1942) it is one of the - 17 - most ’’common and conspicuous of Arizona grassland species” and was found to be "more common in areas of Andropogon, Aristida, and other tall grasses„ than in areas of curly mesquite (Hilaria) and Bouteloua’’« Limited observations in Montana indicate that it is a grass-feeding species which is associated with dense stands of Agropyron smithii growing in moist depressions or swales. At no time was B„ nubiIum abundant or found commonly over a very large area. Brachystola magna (Girard) This species was rarely encountered during the study. It was not listed for any of the study sites but was recorded in field notes from Yellowstone, Big Horn, Rosebud, Ouster, Prairie, Dawson, and Richland counties. The few B. magna. observed were found along highway borrow-pits or in areas of sparse grass and forbs such as Helianthus annuus, Sphaeralcea eoccinea, and Monolepis nuttalliana. Bradynotes obesa (Thomas) This rare species was recorded in field records from only 3 areas. These were southeast of Red Lodge, Carbon county, south of Livingston, Park county, and west of Dell, Beaverhead county. It was listed on study site 11. In every instance it was found where the vegetation was sparse and of low stature on an outwash fan at the base of a mountain range. No field observations were made which would associate this species with any plant or group of plants. Bruneria brunnea (Thomas) B. brunnea was recorded from all counties as far east as Blaine county on the north and western Big Horn county on the south. It was also found in the northeast corner of the state in Daniels, Sheridan, and Roosevelt counties = It was . listed from study sites 2, 8, 1$, 20, k 5 , 47» 58, 69, 82, 88, and 98. It occurred in numbers greater than I per sq yd in a few grassland sites but not on abandoned fields. This grass- feeding species (Griddle 1933, Brooks 1958) was found to be most abundant on upland areas characterized by the presence of Koeleria cristata, Stipa comata, and Agropyron smithii with an understory of Carex filifolia, Bouteloua gracilis, and Selaginella densa. Bruneria sordida (McNeill) This species was noted on only two occasions when it was listed for study sites 10 and 11 in Beaverhead county. specimens were seen at these locations. Few In both instances the vegetation was sparse and of low stature on an outwash fan at the base of a mountain range. Camnula pellucida (Scudder) C . pellucida was recorded from study sites 2, 4, 7 , 80-82, and 86, occurring in numbers greater than I per sq yd on some abandoned field sites but in lower numbers on all native grassland sites. This species is listed by Griddle (1933) as a grass-feeding species which also feeds on "a few - 19 - broad-leaved plants"* He and other Canadian workers have con­ sidered Ce pellucida to be one of the most serious pests in cropland areas of the prairie Provinces. On Montana grazing lands and abandoned fields its local distribution is usually predictable due to a close association with Poa spp., in particular Poa pratensis. It has also been noted on occasion in fields planted to Agropyron cristatum but has not been found generally common in fields of small grain. Exceptions to this have been noted in the extreme northeastern Montana counties of Sheridan, Daniels, and northern Roosevelt where C . pellucida has often been found in grain fields and also has earned the common name "roadside grasshopper" by which it is often referred to in Canada. Ghorthippus longicornis (Latreille). This species has been found on few occasions from Glacier, Toole, Liberty, and Hill counties on the north, and Beaverhead, Madison, Broadwater, Gallatin, Park, Meagher, Sweetgrass, Stillwater, Carbon, and Yellowstone counties on the south. It was listed from study site I. C . longicornis is listed.among the grass-feeding species by Griddle (1933) and Brooks (1958). It was found at elevations between $,000 and 7,000 ft in Montana and usually in areas where seeping water or flowing springs had created wet conditions. 20 Chortophaga viridifasciata (Be Geer) C e viridifasciata was recorded for Beaverhead, Madison, Gallatin, Broadwater,: Park, Meagher, and Big Horn counties = It was listed from study sites 7, 29, 39, and 72-74. This grass-feeding species (Griddle 1933, Brooks 1953) appears as adult early in the spring, oviposits, and disappears by midJuly which may account for it not being listed more often. It appeared to occupy habitats similar .to those of Arphia cpnspersa and generally could be found also early in the spring in lush vegetation along the banks of slow moving streams. Here the green color phase appeared to be more common than the brown. It was never found in abundance. Circotettix rabula rabula Rehn and Hebard This species was recorded in field notes from all coun­ ties south of the Missouri River except Cascade, Chouteau, Judith Basin, Fergus, and Petroleum. North of the Missouri River it was recorded from all counties east of Hill county. It was listed from study sites 10, 12, 17, 29, 39, 50, 72-74, 76, 79, and 103« C_« r. rabula is listed among the mixed- feeding species by Griddle (1933) and Brooks (1953). In Montana, the adults appear to prefer bare spots and eroded areas and often are seen hovering over these areas on warm days as has been mentioned for Aerochoreutes carlinianus. Both of these species are difficult to catch in a sweep-net and were often listed on the basis of sight and sound records. 21 C . r . rabula was never found in abundance« Cordillacris crenulata (Bruner) Field notes record this species south of the Missouri River from all counties except Jefferson, Meagher, Broadwater, Judith Basin, and Fergus„ North of" the Missouri River it was recorded from all counties east of Toole county. It was listed from study sites 8, 10-12, 23, 28, 46, 53? 57? 62, 64? and 6?. C.. crenulata is a grass-feeding species which occurs most commonly, at times abundantly, in the short-grass plains areas. It is especially common on Bouteloua gracilis, Buchloe dactyloldes, and Carex'filifolia. Cratypedes negleetus (Thomas) This species was found in low numbers on.foot-hills areas east of the Rockies in all counties as far as Blaine county on the north and Yellowstone county on.the south. listed from study sites 8, 12, 17, and 50. It was also This species is listed by Griddle (1933) among those which feed on several species of broad-leaved plants. Brooks (1958) says "the spe­ cies is primarily a grass-feeder". It was found to be most common in areas of dense, tall vegetation comprised of a mix­ ture of grasses and of forbs such as Oxytropis spp., Astragalus spp., and Lupinus spp. Dactylotum pictum (Thomas) This species was not listed from any of the study sites. Only a few individuals were recorded in field notes from - 22 low-stature vegetation in southeastern Big Horn, Rosebud, Powder River, Ouster, Dawson, Richland, and Roosevelt coun­ ties. Those few specimens noted were on rocky areas of sparse vegetation. Derotmema haydenii haydenii (Thomas) h. haydenii was listed from study sites 2 9 , 3 9 , 72-74, D. 83, 94, and 99• This species is very localized in its distri­ bution but was found to be numerous.in some small areas. It appeared to be a forb-feeding species (Anderson- & Wright 1952) which preferred areas of dry, sparse vegetation. It was most commonly encountered in areas of highly compacted soil such as may be found in a dry stream bed, the mounds of an old prairie dog town, or a heavily grazed area on heavy clay soil. Dissosteira Carolina (Linnaeus) This species was recorded from study sites 50, 72-74, 77, and 94* It is listed among the mixed-feeding species by Griddle (1933) which "prefers broad-leaved plants". As an adult it frequently congregates on dusty, barren areas such as a dirt road or fallowed field and may thus occur in high numbers within a small area. D. Carolina appeared to spend little time feeding as an adult. It has strong flying capa­ bilities which prevent it from being captured readily by sweeping and it may have been overlooked on a number of the study sites; - 23 Drepanopterna femoratum (Scudder) Field notes show D. femoratum present in all counties except Madison and Jefferson. It was also listed from study sites 1$, 28, 29, 33, 34, 37, 47, 54, 57, 63-65, 71-75, 93, 94, 96, 97, 101, and 104. It occurred in numbers greater than I per sq yd on some native grassland sites but not on aban­ doned fields. This grass-feeding species shows a preference for Agropyron smithii (Anderson Sc Wright 1952). Although the distribution of this species appeared to be partly correlated with the presence or absence of A. smithii, it was definitely not as wide-spread in distribution as is A. smithii. As a later instar nymph and adult it congregated and was most numerous in the dry plains areas of heavy clay soil. 'Eneoptolophus sordidus costalis (Scudder) This species was listed from study sites 31, 36, 63, 66, 72-74, 94, 96, and 100. grassland habitats. It was found in low numbers in dry It has been listed among the grass- feeding species (Griddle 1933, Anderson & Wright 1952, Brooks 1958) and seemed to be most common on poorly drained clay soils dominated by Agropyron smithii. Eritettix simplex tricarinatus (Thomas) This species was listed from study sites 12, 13, 16, 27, 29, 32, 33, 35, 39# 54, 56, 6l, 66, and 72-74. First instar nymphs were generally found between 15 August and I September and in numbers great enough to make this species dominant at TT — 24 — that time. The adults which appear early in the spring appar­ ently disappear early in the season since only an occasional individual adult was noted after mid-July. Limited feeding observations indicated that-E. s_. tricarinatus is a grassfeeding species. It was found to be most abundant in short- grass areas dominated by Bouteloua gracilis and was noted feeding on this grass. Hadrotettix trifasciatus (Say) H. trifasciatus was recorded from study sites 15, 29, 39, 50, 57, 67, 70-74, 76, 77, #2, #7, 89, and 95. was common but never found in high numbers. This species It is listed among those species which feed on broad-leaved plants by vari­ ous workers (Griddle 1933, Isely 1938, Ball et al. 1942) and has been observed to include considerable amounts of dry plant material in its diet (Anderson & Wright 1952). H. trifascia­ tus was most commonly found in areas of sparse, dry, lowstature vegetation but did not appear to be associated with a specific plant or group of plants. Hesperotettix viridis pratensis (Scudder) H. y. pratensis was listed from study sites 5, 8, 12, 14, 17, 21, 29, 39, 40, 50, 57, 70, 72-74, 81^84, 89, 94, 95, and 100. It occurred in numbers greater than I per sq yd on some native grassland sites but not on abandoned field sites. Although this species was frequently encountered, its local distribution was closely associated with particular ' - 25 food-plants. It was most abundant in association with Gutierrezia sarothrae. Other plants with which it was occa­ sionally found associated were Solidago spp., Haplopappus spp., and Chrysothamnus spp. Hippiscus rugosus (Scudder) Field records show H. rugosus present in Big Horn, Rosebud, Custer, and Powder River counties. from study sites 29, 39, and 74. It was recorded This species has been noted in very low numbers on gravelly slopes which support tall grasses such as Agropyron spicatum,.Andropogon scoparius. and Boutelo.ua curtipendula. It was previously observed that, while these grasses appear to-be preferred by the adults, the early instar nymphs confine their feeding to the low-growing Bouteloua gracilis (Anderson & Wright 1952). Hypochlora alba (Dodge) H. alba was listed for all counties north of the Missouri River and east of the Rocky Mountains. South of the Missouri River it was listed for all counties east of Meagher. It also appeared on records for study sites 15, 28, 29, 33, 39, 65, and 72-74. This species was found to be extremely local in distribution and invariably associated with Artemisia Iudoviciana. Melanoplus alpinus (Scudder) A few field notes record this species from Beaverhead, Madison, Gallatin, Park, Sweetgrass, Carbon, and Glacier rt_ M T - 26 counties. The only study site for which it was listed is 2. The few specimens observed have been in short vegetation with interspersed open areas at elevations exceeding 6,000 ft. Brooks (195&) says this species feeds on grass in Canada.. Melanoplus bilituratus (Walker) l/ M.. bilituratus was listed from study sites 3-5, 7-9, 11, 13-17, 21, 22, 28-30, 32, 33, 37, 39, 40, 50, 54, 56, 57, 64, 65, 70-76, 80-87, 91-97, and 101-105. It occurred in numbers greater than I per sq yd on some study sites in both native grassland and abandoned fields. This is probably the most common species found on Montana grasslands and occurs in a greater variety of habitats than any other species. It is considered to be a mixed-feeding species which shows a pref­ erence for forbs as an adult (Anderson & Wright 1952). This preference for broad-leaved plants is reflected in the local distribution of M. bilituratus. During the study it was found to be most abundant in areas of degenerated grassland where the perennial grasses were depleted and annual grasses and annual and perennial forbs were abundant. It was particularly" abundant on abandoned fields in the earlier annual grass and l/ This species, formerly known as Melanoplus atlanis. M. mexicanus mexicanus. and, since 1958, as M. bilituratus is now named Melanoplus sanguinipes sanguinipes (F.) in a recent paper by Gurney (1962). - 27 forb or perennial forb stages of succession. Melanoplus bivittatus (Say) M. bivittatus was listed from study sites 3 , 7 , 10, 33, 39, 69, 71-74, 80-82, 92, 94, 103, and 105. It occurred in numbers greater than I per sq yd on some abandoned field sites but in lower numbers on native grassland. This species is a mixed-feeder which shows a preference for forbs. While it was most common and abundant in cropland areas, it was also found associated with forbs, on grasslands. It was not numer­ ous on these grassland areas but was often abundant and one of the dominant species present on abandoned fields in the early stages of plant succession. Melanoplus bowdltchi canus (Hebard) This species was recorded from all counties east of Toole county on the north and Madison county on the south. It was listed from study sites 22, 29, 39, 72-74, 76, 88, 98, and 100. M. b. canus shows a very strong feeding preference for Artemisia cana (Griddle 1933, White & Rock 1945, Anderson & Wright 1952, Brooks 1958), and its occurrence was correlated with the distribution of A. cana within an area. Melanoplus bruneri (Scudder) This species was recorded from mountain meadows above 6,000 ft as far east as Carbon county. being present on any of the study sites. It was not listed as It was noted to be abundant at 2 locationsj one on the west side of the Crazy Mountains and the other on the east slopes of the Crazy Mountains. Both areas were similar to those described by .Kreasky (I960). Kreasky observed that this species thrived "in weedy habitats in which Lupinus sericeus (lupine) is usu­ ally one of the dominant plants” . He noted further that it fed primarily on L. sericeus but turned to Phleum pratense when L. sericeus was depleted. Brooks (1953) says "the species is a mixed feeder, preferring grasses". Griddle (1933) says "its food habits are quite diversified" and states further: "This insect is an inhabitant of shrubby areas in which there are open spaces. We have found it most frequently in openings among mixed'woods in which its chief plant asso­ ciates were Amelanchier, Symphoricarpos, Arctostaphylos and a few coarse grasses or herbaceous plants". Melanoplus confusus (Scudder) This species was recorded in field notes from all coun­ ties except Glacier and Beaverhead. It was listed from study sites 3-6, 8, 9, 12-15, 17, 20, 26, 27, 29, 31, 33, 37, 39, 41, 50, 6l, 71-74, 8l, 82, 85, 88, and 98« It was present in numbers greater than I per sq yd on some native grassland ,sites but in lower numbers on abandoned fields. M. confusus hatches early in the spring and appears as an adult before any others of this genus. It occurred commonly in some areas until mid-July and only an occasional straggler remained by mid-August. Both Griddle (1933) and Brooks (1958) consider it — 29 to be a mixed-feeder in Canada. — Griddle, however, states that it "prefers grass", while Brooks states that it "prefers forbs". Feeding observations in Montana indicate that it pre­ fers forbs and, although it is widespread in its distribution, it was found to be most abundant in dry, sandy areas occupied by forbs and low-stature shrubs. Melanoplus dawsoni (Scudder) .. This species was listed from study sites 2-5, 13, 17, 23, 24, 29, 39, 41, 70-74, Si, 82, .84," and 85- It occurred in numbers greater than I per sq yd on some native grassland sites but in lower numbers on abandoned fields. M= dawsoni is one of the latest species of the genus to hatch in the spring. It was commonly encountered and, at times, abundant in open sandy areas with a cover of low-growing forbs and shrubs such as Symphoricarpos spp., Rosa spp., Solidago spp., Aster spp., and Lupinus spp. It feeds mainly on these broad­ leaved plants in Montana (Anderson & Wright 1952). Melanoplus differentialis (Thomas) This species was noted in counties east of Stillwater county, south of the Missouri River, and counties east of Valley county on the north. only. It was listed from study site 74 Although this species occurs most commonly in cropland areas, occasional individuals are found on grassland. The only feeding observation of M- differentialis on grassland was at study site 74 where it fed on Psoralea tenuiflora. - 30 - Melanoplus femur-rubrum femur-rubrum (Be Geer) This species was listed from study site 74 only. It has been found to be most abundant in irrigated cropland areas and was found only in low numbers on grassland areas * This mixed- feeding species appeared to be confined in its grassland dis­ tribution to moist areas such as occur around livestock drinking sites and about the banks of slow moving streams. Melanoplus gladstoni (Scudder) This species was listed from study sites 9, 1$, 30, 44, 54, 55, 68, 72-74, 84, 89, 92, 94, 95, 100, and 10$. In Canada, Griddle (1933) considered M= gladstoni to be a grassfeeder while Brooks (1958) states that it is a "general mixed feeder". The few observations on its feeding in Montana indi­ cated that it feeds on both forbs and grasses. It was never found abundantly but. was most commonly encountered in dry areas of grasses such as Bouteloua gracilis, Agropyron smithii, Stipa cpmata, and Koeleria cristata along with a mixture of low-stature forbs. Melanoplus infantilis (Scudder) This species was listed from study sites 8, 10, 12, 17-25, 27, 28 , 30, 31, 33-36, 41-43, 45-55, 58, 59, 63-66, x 68-70, 72-74, 83, 84, 86, 88, 90, 91, and 97-103. It occurred in numbers greater than I per sq yd on some native grassland and abandoned field sites. Griddle (1933), Isely (1944), and Anderson & Wright (1952) considered M= infantilis to be a - 31 grass-feeding species. Brooks (1958) states that it is a "mixed feeder, preferring grasses". This species was found to be widespread and, in some areas, numerous on Montana grass­ lands. It was nearly .always among the species inhabiting short-grass sites dominated by Bouteloua gracilis and the sedge Carex filifolia. It was also found in areas dominated by taller grasses such as Agropyron smithii, Stipa cpmata, and Koeleria cristata but generally in lower numbers than on the short-grass areas. Melanoplus keeleri luridus (Dodge) M. k„ luridus was recorded in field notes from counties north and east of Yellowstone county. sites 17, -18, and 74° It was listed for study The few individuals of this species which have been noted appeared to be confined to ravines and gravelly hillsides where a variety of forbs and low shrubs were present. Griddle (1933) lists it among the mixed-feeding species => The few feeding observations on this species in Montana have found it to be restricted to several forbs. Melanoplus kennicotti (Scudder) Only 3 records were.made of the presence of this species, all at elevations above 5,000 ft. It was seen south of Livingston, Park county, north of Red Lodge, Carbon county, and on study site 2 in Glacier county. tions were made in Montana. No feeding observa­ In Canada, however. Griddle (1933) has listed it among the grass-feeding species while - 32 Brooks (195$) states that "it definitely prefers forbs but will eat grasses and sedges". Melanoplus montanus (Thomas) A few specimens of this species were recorded above 6,000 ft in Gallatin, Park, and Glacier counties. from study site I. It was listed The few individuals observed have been in areas of dense vegetation but with an apparent preference for open spots within the habitat. No observations on feeding habits of this species were made. Griddle (1933) lists it among the species which feed on broad-leaved plants. Melanoplus occidentalis occidentalis (Thomas) This species was listed from study sites 5, 6, 9, 11-14, 16, 31, 34, 37, 43, 50, 57, 65, 71-74, $3, and $7. It occurred in numbers greater than I per sq yd on some native grassland and abandoned field sites. M. 0 . occidentalis was found to be most common and at times abundant in the dry grassland areas of the state. It was often one of the domi­ nant species along with Aulocara elliotti in areas of Agropyron s m i t h i i a food-plant which it prefers (Griddle 1933)<> It is not, however, a strict .grass-feeder but has been observed to include various forbs in its diet. It appeared to be restricted somewhat in its distribution by an apparent preference for areas of sandy loam soil. Melanoplus oregonensis (Thomas) ■A few field notes record the presence of this species - 33 above 5,000 ft in Gallatin, Park, and Sweetgrass counties. It was not listed from any of the study sites„ instances, it was found in very low numbers. In most However, on one flat, gravelly area near the northeast end of the Crazy Mountains it was the dominant species. The vegetation of the area was a mixture of Festuca idahoensis and many forbs such as Lupinus sp., Arnica sp., Senecio sp., Taraxacum sp., Geranium sp., and Gaillardea sp. The few observations on feeding by this species indicated a definite preference for broad-leaved plants. Melanoplus packardii (Scudder) M. packardii was listed from study sites 4# 7, 8, 11, 12, 1—I , 20-22,, 2914, 17:, IS3 39, 40, 43, 44, 50, 54, 57, 60, 69, 70, 72--74, 80-85, 87, 89, 92, 94, 95, 100:, 103,- and 105. Some sites on both native grassland and abandoned fields harbored numbers of this species greater than I per sq yd. It was found to be most abundant in areas of sandy or gravelly soil and particularly on abandoned fields in early stages of plant succession. It is considered to be a general-feeder (Anderson & Wright 1952) but in grassland areas showed a definite pref­ erence for members of the Leguminosae. The blossoms of vari­ ous forbs especially are selected for feeding by this grasshopper. Mermiria maculipennis macclungi (Rehn) This species was recorded in field records- from all - 34 - counties north and east of Yellowstone county. from study sites 74 and 76. It was listed It was never found in high num­ bers and was restricted in its distribution to sandy and rocky hillsides where tall grasses such as Bouteloua curtipendula and Galamovilfa IongifoIia were common. ■ Late in the season it commonly occurred in deep coulees in association with tall grasses present there. Metator pardalinus (Saussure) M. pardalinus.was listed from study sites 9, 12,.14, 15, 17, 28, 29, 31, 37, 47, 50, 51, 54-56, 64-66, 68,, 71-75,.82, 83, 86, 88-90, 93-95, 98, 100, 103, and 104. It occurred in numbers greater than I per sq yd on some native grassland sites but in lower numbers on abandoned fields. This species was found most commonly in the grasslands of the eastern part of the state. Although it will feed on a variety of grasses, it showed a preference for Agropyron smithii. When numerous, it appeared to be gregarious and as a later instar nymph and adult was found congregated in large groups. Local movement of the intact group was common (Anderson & Wright 1952). Neopodismopsis abdominalis (Thomas) This species was recorded in low numbers from Gallatin, Park, Sweetgrass, Meagher, and Glacier counties. listed for study site I. It was The few individuals observed have been in dense vegetation of high mountain meadows. According to Brooks (1958) "the species feeds on several species of grasses and sedges”„ Qpeia obscura (Thomas) This species was recorded from all counties north of the Missouri River« South of the Missouri, it was found in all counties east of Wheatland county. It was listed from study sites 15, 13, 23, 29, 31, 33, 36, 39, 47, 54-56, 63-65, 63, 72-74, and 96. It occurred in numbers greater than I per sq yd on some native grassland sites. 0. obscura was confined in its distribution to areas of dry, short grass. This grass- feeding species has been observed to hatch after the mid-part of June in Montana and to be most numerous in areas of Bouteloua gracilis, Carex filifplia, and Buchloe dactyloides. In areas where it could feed on any of a number of available grasses it showed a definite preference for B 0 gracilis-. This is more in accord with the findings of Ball 'et al. (1942) and Isely (1946) for Texas and Arizona than with the Canadian reports of Griddle (1933) and Brooks (1953) who state that 0. obscura prefers Agropyron .smithii. Orphulella pelidna pelidna (Burmeister) This species was not listed from any of the study sites. A few field notes from Madison, Gallatin, Park, Stillwater, Carbon, and Yellowstone counties record it to be present in low numbers. In every case it was associated with a wet situ­ ation such as an irrigated p a s t u r e a swampy site, or along a stream bank. Orphulella speciosa (Scudder) O. speciosa was not listed from any of the study sites = It was recorded in field notes from Yellowstone, Big Horn, Rosebud, Wibaux, Dawson, Richland, Roosevelt, Sheridan, Daniels, and Valley counties. It was generally found in very low numbers among tall grasses on dry, sandy or gravelly hill­ sides. In one area, however, it was found in abundance; this area was the south-facing slope of the narrow valley which separates the Wolf and Rosebud Mountains east of Lodge Grass, Big Horn county. Here it was associated with both Andropogon gerardi and Bouteloua curtipendula upon which it was feeding. In other areas grasses upon which it was observed feeding included Andropogon scoparius, Agropyron spicatum, Agropyron smithii, and Stipa comata. 0. speciosa is included among the grass-feeders by both Griddle (1933) and Brooks (195$)« Rhlibostroma quadrimaculatum (Thomas) P. quadrimaculatum was listed from study sites $, 10, 12, 17, 1$, 20, 21, 27-31, 34-62, 64, 66-70, $4, $7, 3$, 90, and 9$. It occurred in numbers greater than I per sq yd on some native grassland sites but in lower numbers on abandoned fields. P. quadrimaculatum is most common on the dry grass­ lands of eastern Montana. Invariably it was found associated, at times in abundance, with Bouteloua gracilis or, when pres­ ent, Buchloe dactyloides. or both. This was consistently - 37 - found to be the case even in areas where B.'gracilis consti­ tuted but a low percentage of the total vegetation and occurred only in small scattered mats. It is considered to be a grass-feeding species by Griddle (1933), Isely (1944), and Brooks (195$)• Phoetaliotes nebrascensis (Thomas) P. nebrascensis'was listed from study sites 15, 23, 2$, 31, 33, 36, 41, 49, 54, 63, 65, 66, 74, 75, $2, 36, 96, 97, and 101. It occurred in numbers greater than I per sq yd on some native grassland sites, but in lower numbers on abandoned fields. Isely (1944) lists this species among those with mixed-feeder mandibles. In Canada, it was listed by Griddle (1933) as eating mostly grass and by Brooks (195$) as "a very general grass-feeder". Whelan (193$) found in Nebraska it was "mostly on grass, one on weeds, and one on Solidago rigida". In Montana it was found to be a grass-feeding species with a definite preference for Agropyron smithii (Anderson & Wright 1952). It was found to be very localized in its distribution within an area. Early in the season it was in dense stands of A. smithii growing on flat areas. As these stands dried there was movement to adjacent coulees and waterways where the grasses remained green and succulent and in which dense stands could be found. Its local distribution thus appeared to be greatly influenced by the density and succulence of the grass vegetation. This has been substantiated by observations of - - Shotwell (193S) who states that it was "usually found in low­ lands where the vegetation is lush" and by Gantrall (1943) who found it only in "those areas of hydrophytic environment that are covered with heavy growth of commingled Galamagrostis canadense and Carex". Pseudopomala brachyptera (Scudder) This species appeared in collections from Chouteau, Sweet Grass, Petroleum, Musselshell, Big-Horn, and Rosebud counties. It was listed for study site 74. P. brachyptera occurred in very low numbers and was confined to lush, vigorous growths of grasses along waterways and in coulees. On several occa­ sions it was observed feeding on tall, succulent Agropyron smithii„ Its restriction to moist habitats elsewhere has been reported by Walker & Urquhart (1940) in Ontario and by Cantrall (1943) in Michigan. Psoloessa delicatula delicatula (Scudder) This species was listed from study sites 11, 16, 20-22, 29, 35, 37-39, 43-46, 52-62, 66-68, 70, 72-74, 86, 88, 90, 91, 98, 99, 102, and 103° P . .d„ delicatula overwinters as a nymph and appears as an adult early in the spring. Few adults remain after the middle of July so that it could easily be overlooked in collections after that date and until a new hatch appears in the fall. It is most common in short-grass areas but does not appear to be limited in its distribution by any single plant or group of plants. According to Brooks - 39 (1958) this "subspecies feeds on several species of grasses and sedges such as Stipa spp., Garex spp., Agropyron spp., or Festuca spp. Phlox hoodii Richards is also readily eaten". Schistocerca lineata (Scudder) S . lineata was recorded in field notes from scattered areas within all counties east of Fergus county and was listed from study site 74. It was encountered most frequently in shaded, moist ravines or moist upland flats where there was a dense stand of Glycyrrhiza lepidota and some low-stature shrubs. S . lineata was found by Griddle (1932, 1933) to be definitely associated with G. lepidota in Canada and to have fed upon species of the genera Astragalus, Lathyrus, and Vicia. Brooks (1958) also reports that in Canada it is a "forbs-feeder, preferring licorice root (Glycyrrhiza sp.)". On the other hand. Ball (1936) and Ball et al. (1942) found it feeding on the crowns of Bouteloua curtipendula in Arizona and indicated that it fed on low plants and used tall plants as shelter. At one location in Montana, S . lineata was found most commonly either perched on Rosa sp. or eating G. lepi­ dota. Adults were also observed feeding on Solidago rigida and Symphoricarpos orbiculatus very sparingly but were never seen feeding on any of the grasses (Anderson & Wright 1952). Spharagemon collare (Scudder) This species was listed from study sites 20, 29, 40, and 72-74. Although S . collare was common in grassland areas, it — Zj-O — appeared in low n u m b e r s I t seemed to feed on both forbs and grasses with some display of preference for grasses. No par­ ticular association with any plant or group of plants was noted. Spharagemon equale (Say) This species appeared to be more abundant on Montana grasslands than Spharagemon collare» It was listed from study sites 5, 10, 17, 20, 21, 29, 30, 32, 34, 39, 40„ 43, 46, 53-55, 65% 67, 63, 70, 72-74, 33, 39, 92, 97, 99, 101, 103, and 105. The nymphs and adults were common on dry grassland areas but dad not appear to be associated with any particular plant or group of plants. Griddle (1933) placed it among those species which feed on broad-leaved plants but with a preference for the Gruciferae. Brooks (1953) also points to its preference for the Cruciferae but considers it to be a "mixed feeder on range grass". On the basis of mandibular morphology, Isely (1944) placed S . equale among the forb­ feeding grasshoppers. Treherne & Buckell (1924), however, reported that it caused considerable damage to "bunch grass". ' Iu Montana it has been observed feedipg on both grasses and forbs with no indication of preference for either (Anderson & Wright 1952)o It was noted that the adults often congregated in small groups on vegetation-free areas. When disturbed, they flew away, but within a short time reassembled upon the same areas. - 41 Trachyrhachys kiowa kiowa (Thomas) This species was listed from study sites 10, 15, 17# 23-30, 35, 40, 43, 44, 46, 49, 50, 53, 54, 56, 57, 6l, 63-65, 67, 72-74, 87, 89, 90, and 95. It occurred in numbers greater than I per sq yd only on some native grassland sites. T. k. kiowa was generally most common in areas of Bouteloua gra­ cilis, a preferred food-plant (Ball et al. 1942, Anderson & Wright 1952). An exception to this was noted on a clay-soil flat dominated by a sparse stand of Agropyron smithii. On this area, located approximately 2 mi- east of study site 72, it was the dominant species and was feeding on A. smithii as readily as on the scattered patches of B. gracilis present. In another area it was associated with Buchloe dactyloides as well as B. gracilis. Trimerotropis bruneri (McNeill) Field notes record this species from all counties east of the Rocky Mountains except Glacier county. study sites 22, 43# 50, 72-74. It was listed from T. bruneri was found in very low numbers on dry grassland areas with low cover. There was no indication of association with a particular plant or group of- plants. A few feeding observations indicated that it pre­ ferred forbs. Trimerotropis campestris (McNeill) T. campestris was listed from study sites 7, 8, 10, 12, 17, 22, 29, 39, 45, 47, 49, 50, 58, 65, 66, 69, 72-74, 89, 92, - 42 - 95, and 105. This was the most common species of the genus on dry grassland areas and occurred in numbers greater than.I per sq yd on some native grassland sites. Both Griddle (1933) and Brooks (195#) considered it to be a mixed-feeding species in Canada. The observations in Montana agree with this but there was no indication that it showed a preference for grasses as Brooks stated. Trimerotropis cincta (Thomas) A few individuals of this species were recorded in field notes from Musselshell, Yellowstone, Big Horn, Rosebud, and southeastern Garbon counties. 72-74. It was listed from study sites There were insufficient observations on this species to come to any conclusions concerning feeding-habits or asso­ ciations with vegetation. Those individuals encountered on rare occasions were found in rocky locations occupied by a very sparse stand of vegetation. Xanthippus corallipes (Haldeman) X. corallipes was listed from study sites 2, 6, 7, 9, 12, 14, 15, 18, 22, 24, 29, 31, 35, 37, 39, 41, 43, 45, 48, 52, 57-59, 66, 69, 71-74, 84, 90, 99, and 103. The records of X. corallipes make no distinction as to the subspecies observed. This species overwinters as a nymph and appears as an adult early in the spring. Scattered individuals were found until mid-August but, in general, the majority had dis­ appeared by mid-July. Fall populations of newly-hatched - 43 I nymphs were in numbers greater than I per sq yd on some native grassland sites but in lower numbers on abandoned fields. It was found in a variety of habitats but most commonly on sandy areas of low-stature vegetation. Both the nymphs and adults were observed to feed on forbs and grasses. However, feeding on grasses was more common by nymphs than by the adults. The adults included more dry plant debris and forbs in their diet but, in comparison to the nymphs, spent little time in feed­ ing. ADDITIONAL RESULTS AND DISCUSSION The preceding discussion has indicated a relationship between the occurrence of most grasshopper species and their known food-plant preferences. Nevertheless, it was noted early in the study that the observed distribution and abun­ dance of grasshoppers were influenced by factors beyond the mere availability of food. Grasshopper populations were generally low during the period that the study sites were established. Consequently, it is realized that the grasshopper-vegetation relationships observed and measured during this period of comparatively low grasshopper numbers may be altered considerably during periods of high populations. The importance of time has been mentioned previously in regard to the relationship between certain grasshopper species TTT - 44 - life-histories and their presence or absence at the time a study site was visited„ Grasshopper species abundance is likewise related to the time of sampling. This relationship is brought out in the typical seasonal population curves of Figure 3« The data from which Figure 3 was established were obtained in 1958 from a small area of less than 2 acres on flat topography. This study area was of comparatively uniform vegetation consisting primarily of Stipa comata, Agropyroh smithii, Bouteloua gracilis, Koeleria cristata, Poa secunda, Garex eleocharis, and numerous forbs such as Artemisia frigida, Sphaeralcea coccinea, Liatris punctata, and Vicia sparsifolia. Detailed studies of vegetation and the local grasshopper distribution were made and will be presented in a later paper. For the purposes of the present paper, however, it should be mentioned that the quantitative estimates of the grasshopper populations present on the study area' were made by use. of the "night cage" method of sampling. The seasonal variations in the grasshopper populations of this area are typical of those measured in previous Montana studies which employed the use of "night cages" (Anderson & Wright 1952, Anderson 1961). The wide differences in numbers measured from one sampling time to the next result in part from using a random sampling method when the distribution of grasshoppers is not random. This non-random distribution becomes particu­ larly evident in studies of local damage to vegetation 45 -- TOTAL POPULATION YAR - MELANOPLUS - AGENEOTETTIX OEORUM OUADRIMACULATUM number per square -- PHLIBOSTROMA INFANTILIS JULY FIG. 3. AUGUST SEASONAL POPULATION FLUCTUATIONS OF ONE AREA IN AS | SEPTEMBER THE GRASSHOPPER MEASURED IN 1958. (Anderson 1961) . It may- be attributed to non-random plant distribution and seasonal changes in the physical structure of the vegetation. An important point to note from Figure 3 is the apparent rapid decline in grasshopper numbers soon after the peak numbers are reached following hatching. This.has been recorded for every occasion in which "night cages" have been used and cannot be attributed to grasshopper movement. The absence of extensive local movement on the part of most grasshopper species inhabiting an area has been demon­ strated indirectly by control measures which have effected forage protection for more than one season on pastures sur­ rounded by high grasshopper populations (Anderson 1961). More direct evidence has come from two separate field experiments involving marked adults. The first experiment was conducted on a gently sloping area dominated by Agropyron smithii and Stipa comata. Six plastic containers, lg" x 3 z " x 7", were buried to ground level within a radius of I yd. On 2? July i960 the containers were filled with a bran bait which had been tagged with phos­ phorus -32. The bran was kept moist and the grasshoppers were allowed to feed upon it until 27 August i960 when the experi­ ment was terminated. Grasshoppers captured by the "night cage" method of sampling and by sweep-net collections both on the baited and surrounding areas were monitored for radio­ activity. - 47 During the month-long.sampling period most of the radio­ active grasshoppers captured were within 20 ft of the feeding site. Nine radioactive female Aulocara elliotti were col­ lected at greater distances. Five of these were found within 100 ft, 2 between 140 and 160 ft, I at 200 ft, and I at 280 ft, from the feeding site. One radioactive male A., elliotti was obtained 112 ft from the feeding site. Six radioactive Helanoplus bilituratus females were found beyond 20 ft of the feeding site. ft. Five of these were within 60 ft and I at 128 Three radioactive M. bilituratus males were encountered within 100 ft of the feeding site. Nine radioactive Melanoplus packardii females were also found within 100 ft of the feeding station. males were captured. Fifteen radioactive M. packardii Five of these were within 100 ft, 7 between 100 and 180 ft, I at 305 ft, and I at 385 ft from the bait. Four radioactive Hadrotettix trifasciatus females were collected beyond a 20 ft radius. Three of these were between 60 and 80 ft, and I at 112 ft from the feeding site. The only radioactive H. trifasciatus male encountered was 68 ft from the feeding station. Two radioactive Melanoplus occidentalis males were encountered more than 20 ft from the feeding site; I at 40 ft and the other at 95 ft. One radioactive Camnula pellucida female was found 435 ft from the bran among other £. pellucida females which had congregated on a small area during oviposition. — I+B — The second experiment was conducted from 15 July - 20 August 1961 in a shallow basin of approximately 2 acres which was dominated by Agropyron smithii. In this experiment adult Aulocara elliotti were marked with a non-toxic "locust marking" paint manufactured by Imperial Chemical Industries, Ltd., London. During the experimental period some congre­ gation of adults was noted in the area. No marked adults were observed more than $0 yards from the point of release, with the majority of the marked individuals remaining within a radius of 20 yards. „In general it was noted that males were found farther from the release site than were females. The non-random distribution and limited movement of grasshoppers are indications of some relationship existing between vegetation composition and grasshopper distribution. This relationship is not clear especially where numbers of grasshoppers are concerned. Gangwere (1961) has concluded from his work on food selection by Orthoptera that "...food probably plays only a minor part in the insects' habitat selection". From studies of Australian grasshoppers, Andrewartha & Birch (1954, p. 592) state: "The supply.of food was therefore quite unrelated to the numbers of grasshoppers and it cannot in any sense be construed as a ’density-depend­ ent factor*.” Previous studies in Montana (Anderson & Wright 1952) showed that all grasshopper species observed displayed a - 49 - certain amount of food-plant preference. The degree of' selec­ tivity was found to vary greatly between species; some being comparatively omnivorous while others are highly selective. It could be assumed that the composition of vegetation may play an important role in determining the local distribution of grasshoppers. Such was indicated in the discussion of species occurrence and appears likely particularly where a species is highly selective as to food-plants. Such species were found only where the composition of the vegetation included their preferred food-plants. On the other hand, specific food-plants were often found in varying degrees of abundance without those grasshopper species which prefer them being present. _ The composition of the vegetation also appears to have an effect upon the distribution and abundance of those grass­ hopper species which are rather general grass- or forb-feeders not limited in preference to a single plant or to a few plant speciesi Numerous observations on grasshopper populations have shown the grass-feeding as well as forb-feeding species ’ "s: ■ to be most abundant in areas where their preferred foods were readily available. These observations have been substantiated by the data presented in Tables II and III. In general, the grass-feeding species listed in Table II were most numerous where perennial grasses comprised more than 1+0% of the vegetation. The appearance of such forb-feeders as TABLE II. Per cent frequency of occurrence of grasshopper species in numbers greater than I per sq yd on Native Grassland Study Sites. 0-20 * Aeropedellus claVatus * Agehedtettix"deorum * -Amphitornus coloradus * Aulocara elliotti * Bruneria brunnea * Drepanopterna femoraturn ** Hesperotettix viridis *** Melanoplus bilituratus ■***Melanoplus cohfusus ** Melanoplus dawsoni Melanoplus infantilis *** Melanoplus occidentalis Melanoplus packardii * Metator pardalinus * Opeia obscura * Phlibostroma quadrimaculatum * Phoetaliotes nebrascensis * Trachyrhachis kiowa *** Trimerotropis campestris Xanthippus corallipes * Primarily grass-feeding species 'I'* Primarily forb-feeding species *** Mixed-feeding species 50 21-40 33 17 50 50 17 17 17 17 17 41-60 ’ 50 25 13 25 13 13 13 38 25 13 . ■13 13 13 61-80 ■ 18 24 18 18 6 6 6 12 6 35 12 12 6 18 6 9 6 81-100 25 30 20 15 15 10 10 5 50 10 10 15 90 15 15 TABLE III. Per'cent"frequency of occurrence of grasshopper'species in numbers greater than I per sq yd on Abandoned Field Study Sites. ■ Perennial Grass-Cover Classes, in % 81-100 41-60 61-80 21-40 0-20 * * * * * *** Aeropedellus clava'tus Ageneotettix deorum Amphibornus coloradus Aulocara ellidtti Camnula pellucida Melanoplus bilituratus • Melanoplus bivittatus •**Melanoplus infantilis *** Melanoplus occidentalis ** Melanoplus packardii 33 17 100 40 33 100 17 ' 67 50 20 80 20 ■ 67 67 33 . 33 17 50 20 33 * Primarily grass-feeding species ** Primarily forb-feeding species *** Mixed-feeding species 100 - 52 - Hesperotettix viridis and Melanoplus packardii in the higher classes of perennial grass-cover can also be explained on the basis of the feeding habits of these 2 species. H. viridis was restricted primarily to Gutierrezia sarothrae and M. packardii to members of the Leguminosae occurring on native grasslands. The abundance of Aeropedellus clavatus and Melanoplus occidentalis in the low-percentage perennial grasscover classes is not so easily explained in light of their ,observed feeding habits. Both appear to prefer grass although M. occidentalis has also been observed to feed on forbs. Pos­ sibly the presence of A. clavatus may be explained most easily on the basis of availability of green succulent grass early in the season when this early-hatching species is most abun­ dant. The appearances of the remaining species listed in Table II are in accord with their observed feeding habits. As is shown in Table III, the number of species found to comprise the dominant grasshoppers on abandoned fields was relatively small. In accord with observed grasshopper feeding habits, fields which had been abandoned for several years and had reverted to a point of dominance by perennial grasses were those which harbored the highest populations of the grassfeeding species. Conversely, those abandoned fields in the earlier stages of plant succession in which annual or peren­ nial forbs and annual grasses were dominant (the 2 lowest per­ ennial grass-cover classes) harbored the highest populations - 53 - of forb-feeding species. It should be noted, however, that 3 species considered to be grass-feeders were found among the dominants of the 2 lowest perennial grass-cover classes. As has been pointed out, Ageneotettix deorum is a grassfeeding species which apparently displays no preference for any particular grass. It has been shown that it also devours large quantities of dry plant debris which is found on the ground. With the possible exception of fallen grass seeds, no particular component of the debris seems to be preferred. This debris-feeding habit may, therefore, be responsible for the widespread distribution of A., deorum both on abandoned fields and native grasslands. The distribution of the other two grass-feeding species on recently abandoned fields was found to be definitely asso­ ciated with areas of preferred food-plants. Aulocara elliotti was found in patches of Agropyron smithii or, on some older fields, Agropyron cristatum. with Poa pratensis. Camnula pellucida was associated It was observed that P. pratensis often accounted for only a very small percentage of the total vege­ tation of an abandoned field. It invariably grew in small, dense patches varying in size from a few to several hundred square feet. It was within these localized areas that C . pellucida was found. Usually this species had eaten the P. pratensis nearly down to ground level and it was on these heavily damaged, heavily sodded areas that aggregations of C. - 54 - pellucida were found during mating and oviposition. One may note from the above discussion that it has been possible to relate grasshopper distribution and abundance, to some degree, with the availability of specific food-plants. Again it should be pointed out that these specific food-plants may be present in varying amounts without those grasshopper species which prefer them being present. In other words, something beyond the mere availability of food appears to be involved in both habitat selection and abundance of grass­ hoppers on Montana grasslandsi In a review of the "factors affecting food selection in Orthoptera, Gangwere (1961) has stated: "Climate, as well as microclimate, exert an influence on the feeding behavior of Orthoptera. Air and ground level heat, humidity, and wind act separately and together, both on the feeding insects and on the vegetation or animals on which they feed. The effects of these agents are modified by features of the topography and by density of .the plant cover, which determine the degree to which the insects are sheltered." No measurements of the modifying effects of plant cover on the microclimate were made in the present study. Numerous opportunities, however, arose during this study to compare the numbers of grasshoppers on grassland grazed by livestock during the spring and summer with the numbers in an adjoining pasture which was grazed dur­ ing the winter only. The amount of bare ground in each - 55 - pasture was often nearly the same but invariably the height of the vegetation in the winter pastures was greater; i.e., more ground was shaded. Grasshopper populations were generally found "to be inversely proportional to the plant height and amount of shading. The relationships between grasshopper populations and measurements of total foliage cover of the study sites .tend to substantiate these observations (Table IV). Total foliage cover, as measured on the study sites, is in effect an estimate of the amount of shade cast by the vege­ tation. As is indicated by the data presented in Table IV, grasshopper populations on native grassland sites were found to occur less frequently where the per cent total foliage cover was greater than 40. Although some species are listed under total foliage-cover classes greater than 40%, it should be noted that they were in low- numbers and barely qualified / under the "greater than I per sq 'yd" category. It should- also be pointed out that the abundance of grasshoppers never exceeded I per sq yd in areas dominated by either Artemisia tridentata or Sarcobatus vermiculatus during observations over the 12 years covered by this study. Three species which were always found in numbers too low to qualify for inclusion in Table IV were Chorthippus longicornis. MeIanoplus montanus, and Neopodismopsis abdominalis. It is of interest that these species occurred in mountain TTTTT TABLE I V . Per cent frequency of occurrence of grasshopper species in numbers greater than I per sq yd on Native Grassland Study Sites. ■- Total Foliage-Cover Classes, in $ 0-20 Aeropedellus clavatus Ageneotettix deorum Amphitornus cbloradus Aulocara ellibtti Bruneria brunnea Drepanopterna femoraturn Hesperotettix viridis Melanoplus bilituratus Melahoplus cohfusus Melanoplus dawsbrii Melanoplus infantilis Melanoplus oceidentalis Melanoplus packardii Metator pardalinus Opeia obscura Phlibostroma quadrimaculatum Phoetaliotes nebrascensis Trachyrhachis kibwa " Trimerotropis campestris Xanthippus corallipes 30 33 17 27 3 13 7 17 13 7 47 17 7 10 10 33 10 17 3 21-40 41-60 25 25 42 8 8 13 25 8 13 ' 25 25 17 61-80 13 13 13 17 17 8 8 13 13 13 81-100 - 57 - meadows where the total foliage cover always .exceeded 60%, ■ The relationship between grasshoppers occurring on aban­ doned fields and the total foliage cover is not as evident as on the native grassland sites. Those species listed in Table V appeared to be affected to a greater extent by the composi­ tion of the vegetation on abandoned fields than by the total foliage cover. At each of the study sites a measurement of the amount of ground covered by litter was made. The. results of attempting to relate the presence of grasshoppers to the amount of litter will be found in Tables VI and VII. It was noted that there was usually a direct correlation between the amount of ground covered by litter and the total foliage cover. Therefore, with the possible exception of Ageneotettix deorum and Melanoplus bilituratus. 2 species which devour large quan­ tities of dry plant debris, it is not possible to point to any direct effects which the amount of litter may have on grass­ hopper species distribution or abundance. It is generally recognized that the vegetation of grass­ land areas has been and continues to be modified or replaced either in its entirety or in part. These changes nearly always appear to be the result of a disturbance by man or domestic animals through cultivation or the practice of grazing, in combination with the prevailing climatic condi- ■ tions. TTT Such changes in vegetation have been considered by a TABLE V. ' Per cent" frequency of occurrence of grasshopper species in numbers greater than I per sq yd on Abandoned Field Study Sites. 0-20 Aeropedellus clavatus Ageneotettix'deorum Amphitornus colbradus Aulocara ellidtti Camnula pellucida Melanoplus bilituratus .Melanoplus bivittatus Melanoplus infantilis Melanoplus oc-cidentalis Melanoplus packardii 25 25 50 25 50 25 25 25 50 Total Foliage-Cover Classes, in % 21-40 41-60 61-80 81-100 17 33 100 25 17 33 83 33 25 33 25 50 25 50 25 100 TABLE'VI. Per cent frequency of occurrence of grasshopper species in numbers greater than I per sq yd on Native Grassland Study Sites. Litter-Cover Classes, in % 0-20 Aeropedellus clavatus Ageneotettix deorum Amphitornus coloradus Aulocara elliotti Bruneria brunnea Drepanopterna femoratum Hesperdtettix viridis Melanoplus bilituratus Melanoplus confusus Melanoplus dawsoni Melanoplus infantilis Melanoplus occidentalis Melanoplus packardii Metator pardalinus Opeia obscura Phlibostroma quadrimaculatum Phoetaliotes nebrascensis Trachyrhachis kiowa Trimerotropis campestris Xanthippus corallipes 26 36 7' 29 13 7 16 10 3 39 19 7 7 10 29 7 16 3 21-40 29 14 21 7 14 14 . 14 7 21 14 7 7 7 7 41-60 61-80 81-100 100 20 40 20 40 40 40 20 20 20 20 20' 20 50 TABLE V T I . Per cent frequency of occurrence of grasshopper"species in numbers greater than I per sq yd on Abandoned Field Study Sites. Litter-Cover Classes, in % 21-40 41-60 61-80 0-20 Aeropedellus clavatus Ageneotettix deorum Amphitornus coloradus Aulocara ellibtti Camnula pellucida Melanoplus bilituratus Melanoplus bivittatus Melanoplus infantilis Melanoplus oceidentalis Melanoplus packardii 81-100 33 33 50 33 50 17 33 33 67 33 33 ' 50 100 33 17 33 33 33 17 50 ' 33 67 100 67. 33 - '61 - number of investigators to be of extreme importance from the standpoint of increases in grasshopper populations in many areas. The importance of abandoned land as a possible source of grasshopper infestations has been recognized by various workers (Fletcher 1902, Ball 1937, and others). In fact, weedy abandoned land was pointed to (Parker et al. 1955) as one of the factors responsible for the upsurge of Melanoplus bilituratus in the Great Plains area from 1934 to 193&. Shotwell (1953) summarized the situation thus: "In the Dakotas in the thirties when one-third of the wheat was taken out of production, this species built up to the biggest out­ break of grasshoppers seen in modern times." The effect of heavy livestock-grazing on grasshopper populations in the grasslands of British Columbia has been studied by Treherne & Buckell (1924) and was recognized earlier by Buckell (1921). He states: "At the. time when the Riske Greek range was covered with Bunch-grass, and in fact, up until the last few years when the increase in cattle and horses has kept the range continually eaten down, the grass­ hoppers were never in sufficient numbers to be noticed by the settlers. Several of the older ranchers claim that grass­ hoppers were not present on the ranges in the early days, but have come suddenly during the last few years. There is little doubt that this statement in its substance is incorrect. It - 62 is probably that all the injurious species, now found, were present in the years gone by, but in far smaller numbers and ■ have only increased but slightly from time to time." From his observations in England, Clark (194$) says that grasshoppers are "...significantly influenced by the effects of common methods of agriculture on soil and vegetation; dis­ turbance of soil by ploughing, or various types of grazing are important factors determining their occurrence, and the most favorable habitats observed here have been agriculturally derelict or useless land. Abundance of grasshoppers on agri- • cultural land in this country, as in others, is an index of poor soil or of poor farming, though here it does not carry the penalties of mass outbreaks that it may do elsewhere." Overgrazing has been pointed to as an important factor in the creation of optimum habitat conditions for the increase of grasshoppers in Russia (Bei-Bienko 1930, Rubtzov 1932, 1933)o Many workers in this country (Branson 1942, Coyner 193$, 1939, Dibble 1940, Parker 1937, Smith 1940) have asso­ ciated an increase in grasshoppers with heavy grazing by live­ stock. The major portion of this study has been concerned primarily with the existing vegetation as it was encountered at the time a particular area was visited. In addition, year to year vegetation changes were measured on a large wintergrazed pasture of the OW Ranch in southeastern Big Horn - 63 - county. The topography and vegetation of this area have been described previously by Anderson & Wright (1952). The data (Table VIII) were collected.from 4'permanent quadrats, each I sq meter in area, which were examined yearly some time between 15 June and 30 June. During the 8 years, changes in vegeta­ tion which were measured on these 4 quadrats appeared to be typical of the observed changes over a vast portion of the grazing lands in southeastern Montana. Grasshopper popula­ tions on the study area, as well as in most of the south­ eastern part of the state, had dropped to less than I per s q . yd by 1951 following a period of 4 years during which they reached "outbreak" numbers. When the 4 permanent quadrats were established in 1953, the grasshopper populations were below I per sq yd and remained at this low level until 196l. In 1961 certain portions of the study area, and of south­ eastern Montana in general, again harbored populations greater than I per sq yd. The important point here is that, during the period of low grasshopper numbers, great changes took place in the composition and physical structure of the vege­ tation without accompanying changes in the number of grass­ hoppers present. (See Table VIII) It is concluded from the findings reported in this paper and from field observations during the past 12 years that the composition and physical structure of the vegetation on grass­ lands of Montana play an important role in the determination TABLE VIII. Vegetation on 4 permanent square -meter quadrats at the OW Ranch > Decker, Montana. Total Number of Plants or **Total Basal Area (sq mm) Quadrat" Number and Plant Taxa I Agropyron smithii Alyssum alyssoides Androsace sp". Bromus japoriicus 'Camelina sativa Helianthus ahnuus Lappula redowskii Lepidium perfbliatum Plahtago purshii Salsola kali Tragopogon dubius *1IiSchedonnardus paniculatus **Sporobolus cryptandrus 1^ilI iStipa comat a II Agropyron smithii AlySsum alyssoides Bromus japonicus Carex eleocharis Lappula redowskii Polygonum douglasii Tragopogon dubius Viola nuttallii **Poa secunda 1I0IiStipa viridula :1953 1954 1955 1956 195S 1959 I960 1961 644 96 455 2670 12 I 221 526 255 1400 50.6 2 660 155 374 9 172 9 11 6 2 4 3 39 I I 5 7 14 13 5 I 2 90 -1 14 •I 19 1441 112 1032 602 S IS 45 2 27 10 16 25 1115 4 2 643 1200 814 4 I I 3 4 10 I I I 46 7 13 818 73 181 3 998 143 4 I 53 ■ I I 28 55 TABLE VIII. (Concluded) III ..... Agropyrdri smithii Bromus japonicus' Calochortus aIbicaulis Chenopodium sp. Cryptanthe kelseyana Festuca octifIora Helianthus arinuus Lappula reddwskii Oenothera" albicaulis Plantago purshii Sysimbrium loeselii Tradeseantia occidentalis ^Antennaria dimorpha. **Bouteloua gracilis **Poa secunda **Stipa comata ■IV -Agrdpyrori smithii Alyssum alyssoides Bromus japonicus Carex eleocharis Collinsia parviflora Festuca octiflora Lappula redowskii Plantago purshii Psoralea argophylla Psoralea tenuiflora Sphaeralcea coccinia Sysimbrium loeselii Tragopogon dubius ^i=Artemisia frigida **Koeleria cristata ;!;*Poa secunda **Stipa comata 1953 1954 1955 1956 1958 1959 I960 1961 15 34 B 2 39 223 20 .939 .39 276 43 7 2 I 2 I 2 17 I I 20 19 5 395 I 20 13 7 290 14 244 220 213 93 5 . 66 I 3 I 7 6 694 66 4 2 61 I 109 14 I I ' I 3 I 43 106 4 I 5 3 I I I I I I 2 I 60 106 I I 31 6 147 I 2 . 5 39 3 495 I 7 272 7 24 364 41 20 236 & 12 I 1 624 3 6 I 44 12 50 I I 473 7 236 19 129 10 137 3 170 174 119 54 46 3 106 57 5 76 2 293 3 431 506 32 107 I A I 347 226 16 46 I 26 27 23 - 66 - of areas of occupancy by grasshoppers. It is concluded fur­ ther that the composition and structure of vegetation are not responsible for initial grasshopper population increases or declines. This does not negate the importance of proper man­ agement of grazing lands as a deterrent to economic damage from grasshoppers (Pepper 1955). It does deny, however, that vegetation changes resulting from land use, effects of cli­ mate, or both, are in and of themselves responsible for grass­ hopper "outbreaks". The above conclusions are consistent with the observed mosaic patterns of grasshopper distribution on grassland areas 'I ; ! during periods of high as well as low population numbers. j These conclusions are also consistent with the findings that grasshoppers were often absent from habitats which appeared to be particularly favorable to them from the standpoint of vegetation. Above all, these conclusions are consistent with a concept which recognizes the grasshopper population as a dynamic variable which is not merely a pawn of its external ; environment. SUMMARY • The general distribution of 65 grasshopper species on the grasslands of Montana east of the Rocky Mountains is recorded and discussed. Detailed studies of grasshopper- vegetation relationships were made on 105 native grassland L - 67 ■and abandoned field sites. Measurements of vegetation including taxonomic composition, basal cover, foliage cover, and the amount of ground covered by litter were.made at each study site. Accompanying grasshopper population densities were estimated and the species present were recorded. Time as a factor influencing the results of this study is discussed with regard to grasshopper life-histories and changes in population density during a season. Two field experiments involving marked individuals are described. These indicate that grasshopper movement is not an important factor in seasonal changes of a population on a particular area. The following conclusions resulted from this study: 1. The taxonomic composition of the vegetation, as related to feeding-habits, is important in determining the local dis­ tribution and abundance of most grasshoppers. 2. The physical structure o£ the vegetation is important in determining the local distribution and abundance of most grasshoppers. The highest populations were found where the per cent total foliage cover was below 40. 3• The amount of ground covered by litter has little or no effect on local grasshopper distribution and abundance. 4. Vegetation plays an important role in the selection of areas of occupancy by grasshoppers within a grassland habitat but is not a causative agent responsible for initial changes in grasshopper abundance. - 68 LITERATURE CITED Allred, B. ¥. 1941. Grasshoppers and their effect on sage­ brush on the Little Powder River in Wyoming and Montana. Ecology 22: Anderson, N. L. 387-392. 196l. Seasonal losses in rangeland vegeta­ tion due to grasshoppers. - - Jour. Econ. Ent. 54: 369- 378. Anderson,■N. L. & J. C . Wright. 1952. gations on Montana range lands. Grasshopper investi­ Mont. Agr. Exp. Sta. Tech. Bull. 486. ' Andrewartha, H. G. & L. C . Birch. abundance of animals. :: ; 5" Ball, E. D. Jour. ________ _ . Econ. 1954. The distribution and Univ. Chicago Press. \ Food plants of some Arizona grasshoppers. 1936. Econ. Ent. 1937. Chicago: 679-684. Problem of the range grasshopper. Ent. 30: Jour. 904-910. __________, E . R . Tinkham, R . Flock & C . T . Vorhies. 1942. The grasshoppers and other Orthoptera of Arizona. Arizona Agr. Exp. Sta. Tech. Bull. 93. Bei-Bienko, G. T . 1930. The zonal and ecological distribu- \ ^%ion of Acrididae in the West Siberian and Zaisan Plains. (In Russian with English summary.) Leningr. I: Branson, F . A. T Bull. Plant Prot., 51-90. 1942. A preliminary report on the insect - 69 - . orders found in various grassland habitats in the vicinity of Hays, Kansas„ Brooks, A. R. Trans. Kans. Acad. Sci. 45: 1958. Acridoidea of Southern Alberta, Saskatchewan, and Manitoba (Ortho^tera). 90: 1S9-194. Can. Ent. Supplement 9. Buckell, E„ R. 1921. The influence of locusts on the ranges of British Columbia, 51st Ann. Rep. Ent. Soc. Ontario, ,1920: 23-29. Cantrall, I. J. 1943. The ecology of the Orthdptera and Dermaptera of the George Reserve, Michigan. Misc. Publ. Hus. Zool. Univ. Mich. 5^. Clark, E. J. hoppers. Clark, L. R. 1948. Studies in the ecology of British grass­ Trans. R. Ent. Soc. Lond. 2£: 1947. 173-222. An ecological study of the Australian plague locust (Chortoicetes terminifera Walk.) in the Bogan-Macquarie outbreak area, N . S . W. Coun. Sci. Ind. Res. (Australia) Bull. 226. __________0 1947a. Ecological observations on the small plague grasshopper, Austroicetes cruciata (Sauss.), in the Trangie District, Central Western New South Wales. Coun. Sci. Ind. Res. (Australia) Bull. 228. Coyner, W. R. 1938. Insect distribution and seasonal suc­ cession in overgrazed and normal grasslands. M. S . thesis, Univ. Okla., Norman. Unpubl.' - 70 - __________. 1939o A report of the effect of overgrazing on the Acrididae„ Griddle, N„ Proc. Okla. Acad, Sei= Ig.; 1932„ Scud, Can. __________, The life-history of Schistocerca lineata Ent0 64: 1933 o 9$-102o Studies in the biology of North American Acrididae development and habits„ Proc0 World's Grain Exhibition and Conference, Canada 2: Dibble, C 0 B 0 1940. in Michigan. Fletcher, J e $3-85° Grasshoppers, a factor in soil erosion Jour. Econ0 Ent. 23: 1902. 474-494« 498-499« Canadian Dept, Agr. Gent0 Exp0 Farm Rep0, Entomologist and Botanist„ Ann. Rep. on Exp0 Farms for 1901« Gangwere, S. K 0 1961. Orthoptera. Gurney, A e B. A monograph on food selection in Trans0 Am. Ent„ Soc. $7: 1962. 67-230. On the name of the migratory grass- . hopper of the United States and Canada, Melanoplus sanguinipes (F.) (Orthoptera, Acrididae). Soc. Wash. 75: 189-192. Helburn, N 0, M 0 J 0 Edie & G0 Lightfoot„ maps. Proc0 Biol0 Bozeman: 1962. Montana in Res. and End. Found. at Mont. State Coll. Isely, F. B 0 193$. The relations of Texas Acrididae to plants and soils. ______ . 1944« Ecol. Monogr. 8: 551-604« Correlation between mandibular morphology and food specificity in grasshoppers. Ann. Ent= Soc. - 71 - Am. 37: __________„ 47-67. 1946. Differential feeding in relation to local distribution of grasshoppers. Kachaeva, Z 0 F 0 & I. A 0 Rubtzov, Ecology 27: 1935. 128-138, Geobotanical descrip­ tion of the reservation areas of Acrididae in the Balangsk steppe. (In Russian). Vost., Sibiri, 2: Kreasky, J 0 B 0 Trud. Zasch0 Rast. 278-288. I960.' Extended diapause in eggs of high- altitude species of grasshoppers, and a note on foodplant preferences Melanoplus bruneri. . Am,- 53: Morse, A. P. locusts. Nefedov, N, I, Ann. Ent. Soc0 436-438. 1899» The distribution of the New England Psyche 8: 1931. 315-323. On the degree of the infestation of the soils of Troitzk Steppe-Forest Reserve with eggpods of Arcyptera microptera, F 0 W 0 (In Russian with English summary,) 8: Bull, Inst, Rech,, Biolo Perm, 1-17. Parker, J 0 R, 1937. The influence of man and cultivation on the grasshopper problem in the United States, No. 14 (Cairo): I, L, C. Appendix 14. __________, R. C, Newton & R. L. Shotwell. 1955 = Observa­ tions on mass flights and other activities of the migra­ tory grasshopper. U 0S 0D 0A 0 Tech, Bull. 1109. Pepper, J 0 H 0 1955. The ecological approach to management of - 72 insect populations„ Pfadt, Ro E . 1949= Jour. Econ= Ent= 48: 451-456. Food-plants, distribution, and abundance of the big-headed grasshopper, Aulocara elliotti (Thos,)= Jour. Kans. Ent= Soc. 22_: Rubtsov, I= A= 1932. 69-74° The habitats and conditions of grass­ hopper fs outbreaks in East Siberia. (In Russian with English summary.) Bull. Plant Prot= (I Ent=) 3.: __________. 1933 o Contribution to the ecology of injurious • grasshoppers in East Siberia. (In Russian=) Rast= Vost., Sibiri,' 1933: __________. . 1934« 8-97° 1938. Bull. Ent= Res. 25.: __________= 339-348. Species and distribution of grass­ hoppers responsible for recent outbreaks. ■ Ent= 31.: Trud, Zasch= Fertility and climatic adaptations in Siberian grasshoppers. Shotwell, R= L. 33-130. Jour= Econ= 602-610 = 1953° The use of sprays to control grasshoppers in fall-seeded wheat in Western Kansas = U =S=D=A =, B=EoP=Q=, E -8680 Smith, C= Co 1940= The effect of overgrazing and erosion upon the biota of the mixed grass prairie of Oklahoma. Ecology 21: Strohecker, H= F= 381-397° 1937° An ecological study of some Orthoptera of the Chicago area. Treherne, R= C= & E = R= Buckell= British Columbia= Ecology 18: 1924° 231-250. The grasshoppers of Dom= Can. Dept= Agr= Bull= 22, n ° ser= J - rB Urquhart, -F, A. 1941= An ecological study of the Saltatoria of Point Pelee, Ontario. Sero 50° Univ0 Toronto Studies, Biol. 1-91= Walker, E. M 0 & F 0 A 0 Urquhart. of Orthoptera in Ontario. Whelan, D 0 B. prairie. 193$. 1940. Mew records and notes Can. Ent. 72: 15-1$. Orthoptera of an eastern Nebraska Jour. Mans. Ent. Soc. 11: White, R= M. & P. J. G. Rock. 1945. 3-6. A contribution to the knowledge of the Acrididae of Alberta. Sci. Agr. 25.: 577-596. Williams, L. H. 1954= The feeding habits and food preference of Acrididae and the factors which determine them. Trans. R. Ent0 Soc. Bond. 105: 423-454= I MONTANA STATE UNIVERSITY LIBRARIES 3 I762 10005 27 3 _Z)3 %