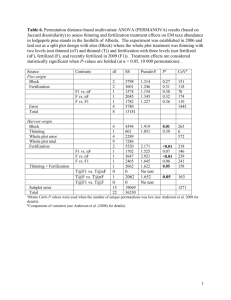
Document 13478414
advertisement

Response of native species to variable nitrogen, phosphorus, and potassium fertilization on mine soils by Philip John Hertzog A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science in Land Rehabilitation Montana State University © Copyright by Philip John Hertzog (1983) Abstract: Eastern Montana is a region in which coal mining in the past decade has resulted in the destruction of soil systems and vegetation. State law requires this land be reclaimed with the establishment of permanent, diverse, predominantly native plant communities. Fertilization is one management technique that may help to meet this legal requirement. This study evaluated the effects of several fertilization treatments on the establishment of vegetation on mine land. A native species mixture was seeded on cover-soiled, regraded mine spoils in the fall of 1981 at Colstrip, Montana. The following spring, the site was fertilized with 24 treatments of N (0, 14, 28, and 56 kg N/ha), P (0, 112, and 168 kg P/ha), and K (0 and 28 kg K/ha) in factorial combination. Vegetational establishment was evaluated by measuring plant density, aerial biomass, canopy cover, and frequency by plant class and species. In addition, species diversity, evenness, and richness were calculated for each treatment. After one growing season, P was the only fertilizer element to significantly affect vegetational establishment. Regardless of the level of N and K, P fertilization at 112 and 168 kg P/ha decreased density, aerial biomass, canopy cover, and frequency of warm season grasses. Legume aerial biomass and canopy cover were reduced by fertilization at 112 kg P/ha. The reduction of these two plant classes may be due to P fertilization increasing the competitive effect of other plant classes. Phosphorus fertilization increased the aerial biomass and canopy cover of annual forbs and annual grasses. Species diversity, evenness, and richness varied over the study site, but were not affected by fertilization. It was recommended that P fertilization not be used the first growing season due to its negative effect on warm season grass and to an extent legume establishment. Nitrogen and K fertilization were not necessary for plant establishment under conditions of this study. RESPONSE OF NATIVE SPECIES TO VARIABLE NITROGEN, PHOSPHORUS, AND POTASSIUM FERTILIZATION ON MINE SOILS by Philip John Hertzog A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science in Land Rehabilitation MONTANA STATE UNIVERSITY Bozeman, Montana March 1983 m a in l/b, ■ ii APPROVAL of a thesis submitted by Philip John Hertzog This thesis has been read by each member of the thesis committee and has been found to be satisfactory regarding content, English useage, format, citations, bibliographic style, and consistency, and is ready for submission to the College of Graduate Studies. _£k Date Chairperson, Approved for the Major Department ^iliolS3 Date Head, Major Department Approved for the College of Graduate Studies Date Graduate Dean ill STATEMENT OF PERMISSION TO USE In prese n t i n g this thesis in partial fulfillment of the requirements for a master’s degree at Montana State University, I agree that the Library shall make it available to borrowers under rules of the Library. Brief quotations from this thesis are allowable without special permission, provided that accurate acknowledgment of source is made. Permission for extensive quotation from or reproduction of this thesis may be granted by my major professor, or in his absence, by the Director of Libraries when, in the opinion of either, the proposed use of the material is for scholarly purposes. Any copying or use of the material in this thesis for financial gain shall not be allowed without my written permission. Signature, V ACKNOWLEDGEMENTS Behind all graduate students there stands a host of individuals, whose invaluable support makes the successful completion of a research study possible. To name all these individuals is impossible, but there are those who deserve recognition. Dr. Steve Young, my graduate committee chairman, deserves special recognition for his patience, technical advice, and encouragement. Dr. Frank Munshower and Dr. Paul Kresge provided advice on content and writing style. Bob Rennick and Patrick Platenberg supplied invaluable help in data collection, methodology, and plant identification. Dennis Neuman and Chris LeVine provided assistance on soil analyses. Ron ThorsoneS technical knowledge of computers and programming saved innumerable hours of statistical analyses. The typing skills of Jeanne Blee, as well as her knowledge of word processing were greatly appreciated. Finally, a special thanks goes to the Western Energy Company which provided financial support for this project. -X vi TABLE OF CONTENTS Table of Contents........ « . . . ............................ . vi List of Tables ............................................ .. . .viii List of Figures. . . . . . . . . . . . . . . ............ .... Abstract.................................... ............. .. Introduction ...................... . .............. . . . . . . x xii I ^ Zt ■=? \£> CO Literature Review. .............................................. Vegetational Responses to Fertilization . . . . . . .......... Botanical Composition........ ............. .. Cool Season Grasses.......... .. . . . ................ Warm Season Grasses. .................................. Sedges .......... 10 Legumes. .................. 10 Forbs................ 11 Shrubs and Half Shrubs .................. 11 Factors Affecting Vegetation Response to Fertilization. . . . 11 Biological . . . . ..................... 12 Nutrient Interactions.............. 12 Soil Properties. ......................................... 14 Management Techniques. .................. . . . . . . . 17 Nutrient Cycling in Mine Land ................................. 19 Materials and Methods..................... Experimental Design . . . . . . . . . . . .......... . . . . Vegetation Sampling . . . . . . ........ Density...................... Aerial Biomass ........ . . . . . . . . . . . . . . . . Canopy Cover . . . . . . . . . ........................ Frequency. . . . . . . . . . . . . . . . . . . . . . . . Diversity. . . ........................................ Evenness . . . .......... . . . . . . . . . . . . . . . Richness . . ................ . Soil Sampling . . . . . . . . . . . . . . . ........ . . . . Statistical Analysis. ............ Vegetation . . . . . . . . . . . . . . . .......... . . Soils............ 23 23 27 27 29 29 31 31 32 32 32 34 34 36 Study Site Description ................ . . . . . . . . . . . . . 37 Location. .................. . „ . ............ ........... 37 Topography. . ............................ 37 Climate .................................. . 38 Vegetation............ 39 G e o l o g y .................................. . . ......... .. . S o i l s .................. 40 40 vii Results and Discussion . . . . . . . . . Introduction. . . . . . .......... Phosphorus. ....................... Cool Season Perennial Grasses. Warm Season Grasses. . . . . . Annual Grasses . . . . . . . . Annual Forbs . . . . . . . . . Biennial and Perennial Forbs . Legumes. . . . . . . . . . . . Shrubs . . . . . . . . . . . . Total Vegetation . .......... Plant Community Development. . Residual Effects ............ Nitrogen. . ............... . . . . Potassium . . . . . . . .......... Nutrient Interactions . . . . . . . Adequacy of Regression Models . . . 45 45 46 47 48 55 58 61 62 68 69 70 76 78 86 87 88 Recommendations 90 S u m m a r y .......................................... 94 Appendices . . . Appendix A: Appendix B: Appendix C: . . . . . .......................... . . . . . . 97 Soil Profile Description................ . 98 Analysis of Variance Tables .............. 100 Vegetational Data by Plant Species.......... . . 106 Literature Cited . . . . . . . . . . . . . . .................... 111 viii LIST OF TABLES Table I. Seeding mixture and rates used on fertilization study? . 24 Table 2. Fertilizer treatment combinations................. . . 26 Table 3. Cover classes used for canopy cover sampling .......... Table 4. Mean monthly temperature, precipitation, and deviations from the long term norm, Colstrip, Montana, 1981-82. . . 39 Table 5. Baseline soil data of the study site, October, 1981. . . 42 Table Table Table 6 . Vegetational statistics of cool season perennial grasses by fertilization treatment, 1982 ............ . 31 48 7. Vegetational statistics of warm season grasses by fertilization treatment, 1982. . ............ ........ 49 8 . Mean percent frequency of plant species by P fertilization.rate, July, 1982 ...................... 54 . 9 . Vegetational statistics of annual grasses by fertilization treatment, 1982. . . . .................. 58 Table 10. Vegetational statistics of annual forbs by fertilization treatment, 1982......................... 59 Table IT. Vegetational statistics of biennial forbs by fertilization treatment, 1982. . ...................... 62 Table 12. Vegetational statistics of perennial forbs by fertilization treatment, 1982............. 63 Table 13. Vegetational statistics of legumes by fertilization treatment, 1982. . . . . . . . . . . . ................ 64 Table 14. Vegetational statistics of shrubs by fertilization treatment, 1982. .............. 69 Table 15. Vegetational statistics of total vegetation by fertilization treatment, 1982. ............ . . . . . . 70 Table 16. Mean diversity, evenness, and richness indices by . fertilization treatment, 1982......... 73 Table 17. Mean soil extractable P content by P fertilizer rate (ppm), October, 1982 .............................. 76 Table ix Table 18, Mean soil NOg-M content by N fertilizer rate (ppm), October, 1982.......... 82 Table 19. Mean percent soil organic matter content by N fertilizer rate, October, 1982. . . . . . . ................ . . . 84 Table 20. Mean soil total N content by N fertilization rate, October, 1982. . ......................... ............ 86 Table 21. Mean soil C/N ratios by N fertilization rate, October, 1982. . . . . . ............................ . Table 22. Soil profile description of the study site . . . . . . . 86 99 Table 23. Analysis of variance for density by plant class, May and July, 1982 ........................... 101 Table 24. Analysis of variance for aerial biomass by plant class, July, 1982.............................. 102 Table 25. Analysis of variance for canopy cover by plant class, July, 1982 . .................................... 103 Table 26. Analysis of.variance for extractable P content of soils, October, 1982 ........................ . . . . . 104 Table 27. Analysis of variance for NOg-N content of soils, October, 1982. . . . ............... 104 Table 28. Analysis of variance for organic matter content of soils, October, 1982 . . . . . ...................... . 104 Table 29. Analysis of variance for total N content of soils, October, 1982.......................................... 105 Table 30. Mean plant species density by fertilization treatment, May, 1982. ................................. 107 Table 31. Mean plant species density by fertilization treatment, July, 1982. . . . . . . . .................. 108 Table 32. Mean percent canopy cover of plant species by . fertilization treatment, July, 1982. . . . . . . . . . . 109 Table 3 3 . Percent frequency of plant species by fertilization treatment, October, 1982 .......... . . . . . . . . . . 110 X LIST OF FIGURES Figure I.Field design. .... ......................................25 Figure 2. Vegetation sampling design for each experimental p l o t ...................................... 28 Figure 3. Preliminary sampling to estimate number of frames n e e d e d .................. 30 4. Study site location ................................ . 37 Figure Figure Figure Figure Figure Figure 5. Volumetric soil water content . . . ................... 43 6 . Mean density of warm season grasses in response to P fertilization .................... .. 50 7. Mean aerial biomass of warm season grasses in response to P fertilization . . . . . ................ 52 8 . Mean canopy cover of warm season grasses in response to P fertilization .................... 53 9« Mean aerial biomass of annual grasses in response to P fertilization............... 56 Figure 10. Mean canopy cover of annual grasses in response to P fertilization . ; .'. ........ .. 57 Figure 11. Mean canopy cover of annual forbs in response to P fertilization ...................... . . 60 Figure 12. Mean aerial biomass of legumes in response to P ferti l i z a t i o n .......... ................... .. . 65 Figure 13. Mean canopy cover of legumes in response to P fertilization ...................................... 66 Figure 14. Mean aerial biomass of total vegetation in response to P fertilization ............................. 71 Figure 15. Mean canopy cover of total vegetation in response to P fertilization . . . . . ................ 72 Figure 16. Composition of plant community as affected by P fertilization in terms of aerial biomass. . . . . . . 74 Figure 17. Composition of plant community, as affected by P fertilization in terms of canopy cover............ ■. 75 xi Figure 18. Mean soil extractable P content response to P fertilization . . . . ........ . . . . . . . . . . • 77 Figure 19. Mean soil NOg-N content response to N fertilization . . . . . . ................ . 81 . 83 .... Figure 20. Mean soil NOg-N content response to N fertilization for block I and 2 data........ ............ .. . . . xii ABSTRACT Eastern Montana is a region in which coal mining in the past decade has resulted in the destruction of soil systems and vegetation. State law requires this land be reclaimed with the establishment of permanent, diverse, p r e d o m i n a n t l y native plant communities. Fertilization is one management technique that may help to meet this legal requirement. This study evaluated the effects of several fertilization treatments on the establishment of vegetation on mine land. A native species mixture was seeded on cover-soiled, regraded mine spoils in the fall of 1981 at Colstrip, Montana. The following spring, the site was fertilized with 24 treatments of N (0, 14, 28, and 56 kg N/ha), P (0, 112, and 168 kg P/ha), and K (0 and 28 kg K/ha) in factorial combination. Vegetational establishment was evaluated by measuring plant density, aerial biomass, canopy cover, and frequency by plant class and species. In addition, species diversity, evenness, and richness were calculated for each treatment. After one growing season, P was the only fertilizer element to significantly affect vegetational establishment. Regardless of the level of N and K, P fertilization at 112 and 168 kg P/ha decreased density, aerial biomass, canopy cover, and frequency of warm season grasses. Legume aerial biomass and canopy cover were reduced by fertilization at 112 kg P/ha. The reduction of these two plant classes may be due to P fertilization increasing the competitive effect of other plant classes. Phosphorus fertilization increased the aerial biomass and canopy cover of annual forbs and annual grasses. Species diversity, evenness, and richness varied over the study site, but were not affected by fertilization. It was recommended that P fertilization not be used the first growing season due to its negative effect on warm season grass and to an extent legume establishment. Nitrogen and K fertilization were not necessary for plant establishment under conditions of this study. I INTRODUCTION Since the late 1960'S and early 1970'a, America's need for energy has increased, decreased. while reliability of foreign sources of energy has Shortages of oil and gas during the 1970's resulted in long lines at gas stations and higher prices for all petroleum products. The Arab oil embargo during the early 1970's demonstrated the vulnerability of the United States to political blackmail. In order to prevent the United States from falling victim to the demands of oil and gas exporting countries, the Nixon administration initiated policies to achieve energy independence. Succeeding administrations adopted this goal of achieving energy independence for the United States. An integral part of the government's policy for achieving energy independence was the encouragement of the development of western coal fields. Eastern Montana is one western.region where coal development has expanded in the past decade. The Fort Union formation, a geological unit that encompasses much of eastern Montana, contains vast deposits of coal. Furthermore, the Powder River Region, southern extension of the Fort Union Formation, contain 240 billion tons of sub-bituminous coal a is estimated to (Packer 1974). Several strip mining operations have opened or expanded in eastern Montana during the past decade in response to the energy needs of the country. One such operation is Western Energy Company's Rosebud Mine located at Colstrip, Montana. Coal mining has taken place in the Colstrip area over the past fifty years, but in the last 10.years the 2 operation has expanded considerably. Large area strip mining operations are expected to continue at Colstrip for the next several decades. Strip mining of coal has resulted in the destruction of plant communities and disruption of soils. The Federal Surface Mining Control and Reclamation Act of 1977 required this land be reclaimed to a use equal to, or higher than, the use prior to mining. ^At Colstrip much of the mine land was formerly rangeland. The Montana Strip and Underground Mining Act of 1979 required the establishment On mine land of a suitable, permanent, diverse, vegetational cover consisting primarily.of native species capable of feeding livestock and wildlife, withstanding grazing pressure, regenerating under natural.conditions prevailing at the site, and preventing soil erosion to the extent achieved prior to mining. Establishment of diverse, predominantly native plant communities is the goal of revegetation efforts at Colstrip (Coenenberg 1982). In order to achieve this goal, mine spoils are graded to approximate original contour and covered with material suitable for plant growth usually consisting of pre-mine topsoil and subsoil. Establishment of diverse, native plant communities on mine soils may be affected by several factors. of the plant growth medium. One factor is the nutrient status Hodder et al. (1971) noted nutrient deficiencies on. mine spoils at Colstrip could be corrected using fertilization. : By limiting nutrient deficiencies, vegetational. productivity of mine land can be increased to its maximum potential. Little information is available in the literature concerning the 3 effects of fertilization of mine soils on the establishment of native plant species. In particular, information is lacking on how fertilization affects the various components of newly established plant communities such as warm season grasses, or biennial forbs. understanding of how fertilization establishment is important. affects initial An vegetation The initial composition of the plant community may determine the successional direction the community will take. It is important to choose the fertilization regime that achieves the desired revegetation goals. The objective of this study was to determine the first year response of a native species mixture to twenty-four fertilization treatments of nitrogen, phosphorus, and potassium in factorial combination on cover-soiled, regraded mine spoils at Colstrip. The objective of this study was met by evaluating the various plant classes that comprised the newly established plant community. A fertilization treatment that improves establishment of the various plant components, provides high diversity for the overall plant community, and increases plant productivity to levels that meet post mine land uses; should be deemed an acceptable management practice. Management practices that adversely affect the establishment of one or more of these components could lower diversity of the total community to unacceptable levels. In addition, a management practice that increases the dominance of one species over others, could decrease plant community diversity. 4 LITERATURE REVIEW Plants need at least 16 elements in order to maintain vigorous growth and remain healthy. These elements are carbon (C), hydrogen (H), oxygen (0), nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), sulfur (S), zinc (Zn), iron (Fe), manganese (Mn), copper (Cu), boron (B), molybdenum (Mo), and chlorine (Cl). Often one or more of these essential nutrients is lacking in a soil in amounts necessary for adequate plant growth.. Fertilization is used to correct nutrient deficiencies. In order to develop ah, effective fertilization program on mine land, one must understand how vegetation responds to fertilization, factors that influence plant response to fertilization, and elements of nutrient cycling. Vegetational Response to Fertilization Botanical Composition Several studies on both rangeland and mine land noted that .fertilization can change botanical composition of plant communities. Fertilization of rangelands with N increased yield and/or cover for Agroovron smith!! with corresponding decreases for Bouteloua gracilis (Goetz 1969, Power 1979, Rauzi 1978). These studies attributed the decreases in B. gracilis to competitive and shading effects of JL smith!!. Rogler and Lorenz (1957) noted the grazing of study plots may eliminate these competitive effects. a southeastern Alberta grassland with Fertilization of N or N plus P generally increased basal area of cool season perennial grasses and weeds, while decreasing warm season grasses and sedges (Johnson et al. 1967)» In 5 eastern Montana, a ten year study showed species composition varied as much among years as among fertilizer treatments (Wight and Black 1979). Generally, M f e r t i l i z a tion increased plant community composition of cool season grasses, reduced sedges, and above 33 kg N/ha reduced Bouteloua gracilis. at rates A study near Havre, Montana found annual fall application of N for 3 years increased production of total grass, Stina comata, and Asronvron Smithiif but decreased other grasses, rates above 336 kg N plus forbs, and Artemisia friaida at cumulative N/ha (Houlton 1975). Meyn et al. (1976) reported P fertilization of native range near Colstrip, Montana increased cover of annual grasses, forbs, and Gutierrezia sarothrae without affecting perennial grass production. In another study, Asroovron smithii and Stina viridula production were not affected by fertilization with rates of O to 640 kg N/ha, however the highest rate increased production of Bromus tectorum and Bromus ianonicus (White and Halvorson 1980). In Sidney, Montana no changes in plant community composition were found four years after a single application of N plus P in which the highest rates were 672 kg N/ha and 224 kg P/ha (Wight and Black 1978). The effects of fertilization on the botanical composition of newly established vegetation on mine land have also been studied.. A study at Decker, Montana compared seed mixtures consisting of all native species, all introduced species, and a combination of both species (Farmer et aj., 1974). Fertilization increased the grass production of all the mixtures on topdressed and irrigated plots, however, introduced species significantly outproduced the natives. 6 DePuit et al. (1978) reported on the fertilization of several seed mixtures that included native and introduced species seeded separately and in combination. Native species did best at the zero or lowest fertilization rates, but were outproduced by introduced species at higher rates. Coenenberg In another study at Colstrip, Montana, DePuit and (1979) found N plus P fertilization increased stand composition of Agropvron cristatum and Bromus inermis while decreasing legumes. Cool Season Grasses Fertilization with N or N plus P increased the response, establishment, yield, or density of several wheatgrasses (Agroovron spp.) on mine land and rangeland (Danielson et al. 1979, DePuit et al. 1978, Hodder et al. 1971, McGinnies and Nicholas T98O). Physiological changes in wheatgrasses due to fertilization have also been noted. Samuel et al. (1980) found N fertilization increased crude protein content of A. smith!!. Goetz (1975) found the same trends for A. smithil. and reported that addition of P with N fertilization slightly increased crude protein content. protein values, were much lower. increased average VJith only P application, crude Black (1968) reported fertilization crude protein prod u c t i o n of veget a t i o n in northeastern Montana on an JL cristatum site and a native range site composed of 55 percent A 8. smithii. Phosphorus fertilization also increased P content of plants regardless of N fertilizer rate. Nitrogen fertilization increased water use efficiency of these two sites 1.5 to 2.0 times, regardless of P rate. Fertilization affected 7 mycorrhizal development In A. trachveaulum (Danielson et al. 1979). Nitrogen plus P fertilizer increased the rate of endomycorrhizae development and the presence of vesicles within roots. Several studies examined the effects of fertilization on other cool season range grasses. Goetz (1969) found in southwestern North Dakota that N fertilization increased basal cover of Calamovilfa loncifolia. while cover of Stioa comata decreased. Fertilization decreased percent composition of Sa. comata in western Montana (Klages and Ryerson 1965)» and reduced its frequency in Colorado (Houston and Hyder 1975). In contrast, Sa. comata reached maximum yields at 336 kg N/ha (Wight and Black 1972), and reached maximum leaf lengths at 75 kg N/A in the Northern Great Plains (Goetz 1970). Stioa comata also increased in protein content with N fertilization (Goetz 1975). Black and Reitz (1969) found N plus P fertilization increased the seed yield and water use efficiency of Sjl viridula on a study conducted at Sidney, Montana. In contrast, Power (1979) found no effects on S. viridula due to fertilization, content increased. while Goetz (1975) found protein Wight and Black (1972) found Koeleria cristata yield unaffected by N and/or P fertilization, but other studies found it significantly reduced or eliminated (Houston and Hyder 1975, Power and Alessi 1971). Poa secunda generally responded to fertilization with increased yields (Baldwin et al. 1974), increased protein content (Goetz 1975), and increased leaf length (Goetz 1970). Houston and Hyder (1975) found heavy applications of N decreased yields of P. secunda about 88 percent. yields of Festuca Nitrogen plus P fertilization increased idahoensis in Oregon (Baldwin et al. 1974) 8 Nitrogen and N plus P fertilization increased yields of Festuca scabrella when applied at rates up to 1015 kg N/ha and 860 kg P/ha, while fertilization at levels of 350 kg N/ha alone or with 290 kg P/ha killed Es. rubra (Johnston et al. 1968). Fertilization also increased yields of Bromus inermis in this Canadian study. Warm Season Grasses Few studies have documented the response to fertilization of seed mixtures or plant communities consisting almost entirely of warm season grasses. Warnes and Newell (1969) concluded that fertilization during the first growing season had no benefit on establishment of Panicum virgatum. Sorghastrum Andropogon g e r a r d i i . and nutans, Schizachvrium Bouteloua ourtioendula. s c o p a r i u m. Nitrogen fertilization tended to increase biomass production of weeds, decreasing warm season grasses. while Fertilizer applied after the first forage yield from 10 to 243 percent depending on number of years after establishment, soil fertility, and growing season, moisture increased conditions during year of harvest. In another study, Andropogon gerardii. Panicum virgatum, Sorghastrum nutans, and Bouteloua ourtioendula were seeded and then fertilized two years after establishment with annual applications of N and alternate years with P (Rehm et al. 1972). Fertilization increased forage yields of all species in three out of four years, and at the end of four years A. gerardii and ourtioendula were the dominant species on all plots. Fertilization increased forage production and protein content of leaves on a sandy range site in Texas for Andropogon hallii. 9 SchizachvrAum acoparium. Aristida purpurea. Boufceloua Curfcipendulaf Sporobolus crvofcandruSf and other species (Pettit and Deering 1974). Bryan and McMurphy (1968) seeded several warm season species and then fertilized with N and P. Fertilization reduced density of Panicum vireatum due to competition from weeds. With weed control production increased on fertilized plots. E a. virgatum Andronogon gerardii. Eragrostis curvula. and Bothriochloa ischaemum var. ischaemum were not significantly affected by fertilization on the basis of stand establishment. Some studies have reported the effects of range fertilization on warm season grasses. Rauzi et al. (1968) found fertilization of native rangeland in Wyoming did not affect warm season grasses, and variation in yield was a function of time instead of treatment. In another Wyoming study, high levels of K fertilization decreased Buchloe dactvloides yields from 9 percent to a trace over a five year period (Rauzi 1978). Fertilization of true prairie near Manhattan, Kansas decreased W free extract and increased crude fiber, lignin, and ash of Androoogon gerardi and Schizachvrium scooarium (Allen et al. 1976). Bouteloua fertilization, gracilis generally but e x ceptions fertilization increased leaf length, responds exists. Goetz negatively (1970) found N but Wight and Black (1972) found no effects on B. gracilis from N plus P fertilization. North Dakota study, to In a western N plus P fertilization increased protein content of this plant (Goetz 1975). ■ Without N fertilization, applications of P decreased protein content below levels of unfertilized plants. This 10 occurred in the early part of the growing season, but later in the season protein contents were higher in the fertilized plants. Sedges Limited data existed for response of sedges (Carex spp.) to fertilization. Carex filifolia increased in basal cover with K fertilization at rates between 33 and 100 kg N/ha (Goetz 1969). Nitrogen fertilization also increased leaf length (Goetz 1970) and protein content (Goetz 1975). Legumes Several fertilization. studies found legumes n e gatively affected by Nitrogen fertilization reduced legume cover (DePuit et al. 1978), growth (Blaser and Brady 1950), and yield (Cooper 1975). Cooper (1969) stated that N fertilization decreased increasing the competitive ability of grasses. legumes by Epstein (1972) noted decreased nodule formation on legume roots under high levels of soil N. Blaser and Brady (1950) demonstrated that the addition of K to N fertilizer increased productivity of legumes. Addit i o n of P fertilizer is well known for its generally positive effects on legumes (Cooper 1969). Howard et al. (1977) found N plus P fertilization generally favored improved growth of alfalfa. Nitrogen fertilization reduced frequency of Astragalus shortianus and eliminated Lathvrus polymorphus (Houston and Hyder 1975). Fertilization with N and P reduced infection rate of mycorrhizae in Trifolium hvbridum though total infected root length remained unchanged (Danielson et al. 1979). 11 Forbs Kilcher et al. (1965) found for the first two years after a single application of W and/or P fertilizer, weed yields increased. In North Dakota, N fertilization increased basal cover of forb species (Goetz 1969). In Colstrip N, P, and K fertilization of seeded mine lands significantly increased forb production, with Salsola kali as the dominant species (Holechek 1976). Nitrogen and P fertilization reduced basal cover of Selaginella Oiensaf an undesireable species (Smoliak 1965). Fertilization of mixed grass plains with nitrogen increased frequency of Leoidium densiflorum. Chenooodium leotoohvllum. but decreased Phlox hoodii (Houston and Hyder 1975). Shrubs and Half Shrubs Nitrogen and N plus P fertilization increased yield of Atriolax canescens (Aldon et al. 1976, Aldon 1978, Howard et al. 1977)» but emergence and initial growth remained Springfield 1973, Aldon et al. 1975). unaffected (Aldon and Fertilization with N and/or P increased height and yield of Artemisia frigida (Goetz 1970, Wight and Black 1972), but all rates of N in another study reduced its frequency (Houston and Hyder 1975). Factors Affecting Vegetational Response to Fertilization Several studies identified factors that affect plant response to fertilization on both range and mine land. Goetz (1970) found that the vegetational response to fertilization varied by range site, season, plant species, and amount of fertilizer applied. factors that plant response affected included soil type, Other soil 12 fertility level, soil and air temperature, and amount and distribution of precipitation during the growing season (Rauzl et al. 1968). the purpose of this review, For factors that affect the response of vegetation to fertilization are broken into biological, nutrient interactions, soil properties, and management techniques. Biological The presence or absence of microorganisms can have an effect on vegetational response to fertilization. Microorganisms caused a number of physio-chemical changes on mine spoils including increasing the amount of available nutrients (Cundell 1977). true for N and P. This was especially Cundell (1977) and Mosse (1973) stated that vesicular arbuscular mycorrhizae may be important in P deficient soils by increasing the phosphate absorbing surface on roots of grasses and other perennials. Cundell (1977) also suggested azotobacteria may be important in the rhizosphere of plants growing on spoils low in nutrients. Nutrient Interactions Several studies have shown that an excess or deficiency of one nutrient in a soil system can change the response of vegetation to fertilization with other nutrients. The influence of N fertilization on increasing the P uptake by plants is well established (Riley and Barber 1971). Olson and Dreier (1956) found that N fertilization stimulated wheat and oat uptake of.P fertilizer over a wide range of soil conditions. Two studies on mixed grass prairie in North Dakota demonstrated that greater yields 13 occurred when both W and P were applied together, than when each applied as separate treatments (Lorenz and Rogler I972, I973). In a study evaluating response of vegetation to fertilization on selected native grassland sites in western Canada, combinations of N and P fertilizer produced greater biomass yields on all but two sites, when compared to N and P applied separately (Kilcher et al. 1965). Johnston et al. (1968) also found for both a seeded and native range site in western Canada, that N and P applied together produced greater total vegetation yields than when each was applied separately. Black and Wight (1972) found that though P fertilization by itself did not increase total protein content of the forage, when applied in combination with N it increased protein content approximately 30 percent. Nitrogen and P applied in combination also increased percent plant P content and recovery of N in total forage. In contrast, Goetz (1975) found P applied with N did not increase protein content of total vegetation, when compared to N fertilized separately. Riley and Barber (1971) stated that NHiJ-N was superior to NOg-N in stimulating P uptake by soybeans. Another study on cereal grain plants found NHiJ-N increased P uptake by the plants, but NOg-N had little effect (Rennie and Soper 1958). Interactions between nutrients other than those between N and P can affect vegetational response to fertilization. In corn, uptake of Zn appeared to be inhibited by applied P to the extent where levels of Zn critical for growth were reached (Langin 1962). Bains and Fireman (1964) found for five different species of crop plants, that an increase in exchangeable sodium (Na) in the soil generally increased uptake of Na, N, and Mo, and decreased uptake of Ca, K, S, Mg, Cu, Zn, B, and Cl. Soil Properties Several p r operties fertilization. of soils influence plant response to Some of these properties include soil moisture, soil temperature, topsoil depth, and soil pH. Bauer et al. (1978a) stated that moisture has an overriding effect on plant growth and yield, and on the amount of nutrient needed to correct a deficiency. In contrast, Klages and Ryerson (1965) hypothesized that soil fertility may be a greater limiting factor than moisture on total range production, even in coarse-textured, droughty soils. Greater soil moisture content increased plant response to N and/or P fertilization by increasing plant N uptake (Power 1967) and yields (Bauer et al. 1967, Smika et al. 1965, Wight and Black 1979). Nitrogen fertilization did not affect botanical composition of a rangeland site in.Montana during the years of adequate precipitation, however growth of weedy species was stimulated during years of low precipitation (Klages and Ryerson 1965). In a rangeland study, fall soil moisture had. the greatest influence on the vegetation yield of unfertilized and P fertilized plots, while June precipitation influenced the N and N plus P fertilized plots (Johnston et al. 1969). Lauenroth and Dodd (1979) found N fertilization and irrigation favored native legumes growing in the shortgrass prairie of northeastern Colorado, decreased. but in following growing seasons density On plots receiving only irrigation, of legumes legume density 15 remained high. Soil moisture content may affect vegetational response to W fertilization by a f fecting the process of nitrification. Nitrification involves the oxidation of NH14-N to NOg-N. Many forms of. N fertilizers contain NH14-N, and in order for N to become available to the plants, nitrification must take place. Any factor such as soil moisture content that influences nitrification, will affect the amount of N available to plants from ammonical fertilizers. Several studies found nitrification affected by the soil moisture content. Incubation studies by Parker and Larson (1962) revealed that greatest nitrification occurred at soil moisture tensions of 0.7 bars. As soil moisture tension decreased from 0.7 bars, decreased. For tensions above 0.7 bars, increased. Stanford and Epst e i n nitrification nitrification was (1974) discovered not highest nitrification rates occurred at moisture tensions between 0.3 to 0.1 bar in a study that investigated nine different soils of varying texture. In another study, maximum nitrification took place at soil moisture tensions between 0.5 and 0.15 bars (Miller and Johnston 1964). These authors concluded that deficient moisture at higher tensions, and poor aeration at lower tensions limited nitrification. In North Dakota soils under incubation, nitrification rates decreased as soil water contents decreased between 0.2 and 15 bars (Reichman et al. 1966). Soil moisture content may also have an effect on plant response to P fertilization. Beaton and Read (1963). reported that 2.0 bars of moisture tension favored uptake of P in oats, while lowest uptake 16 occurred at 0.4 bars. They also noted water soluble sources of P fertilizer were most sensitive in affecting uptake of P in plants when soil moisture content changed. Several response studies of indicated vegetation to soil N temperature fertilization can affect by the influencing In aerated soils, most nitrification occurs between 0° nitrification. and 35° C (Stanford et al. 1973). 45° C and O0 C. Nitrification ceases completely at In laboratory studies, Parker and Larson (1962) found that a 2° C increase in temperature caused an increase in the rate of nitrification in the 16-20° C range. Between 25° C and 30° C, changes in rate of nitrification were not as evident with small changes in temperature. Stanford et al. (1973) found nitrification increased two fold for each 10° C increase in temperature. Influences of soil temperature on vegetational response to fertilization have been observed. In wheat and barley, temperature was negatively correlated with yield responses from N fertilization (Bauer et al. 1967). In oats greatest uptake of fertilizer P in the mono and diammonium phosphate form occurred at 5° C, when compared to uptake at 16° or 27° C (Beaton and Read 1963). No significant differences in P uptake were noticed between the 16° or 27° C levels. Studies have nitrification. shown Generally, Og content of the soil can affect as O2 content increased from 0 to 20 percent, nitrification increased in curvilinear fashion (Amer and Bartholomew 1951). At least 0.2 to 0.4 percent O2 was needed for nitrification to occur in a soil. Depth of topsoil placement on coal mine spoils has been shown to 17 affect plant response to fertilization. When topsoil was placed at thicknesses of 0 , 2 , 6 and 12 inches on spoil material, fertilization increased the response of vegetation over controls, but the magnitude of increase varied with depth (ARS and NDAES 1977). experiments, total herbage, In greenhouse total root production, and total biomass of Agroovron intermedium increased as topsoil thickness above spoil material increased from 0 to 30 cm (McGinnies and Nicholas 1980). Fertilization increased total production an average of 89 percent over the non-fertilized treatments. Nitrogen fertilization also increased root mass an average of 46 percent in the topsoil and 87 percent in the spoil material. Soil pH has been found to affect the amount of soluble P in soils. Acid soils tended to increase the amount of HgPO^" in the soil, while soils of pH 7.0 and above had greater amounts of HPOjt- 2 (Tisdale and Nelson 1975). Phosphorus was generally most available to plants between pH’s of 5.5 and 7.0. .Management Techniques Timing of fertilization can affect vegetational response. Arizona, In desert grassland plots fertilized during the latter part of the rainy season increased grass production when compared to plots fertilized earlier in the season (Stroehlein et al. 1968). Latter fertilization also increased protein content of plants on two of the sites. Samuel et al. (1980) working in Wyoming found yield and protein content of plants increased linearly with fall applied N, .but increased non-linearIy with spring applications of N at the same 18 rates. Spring applied N also produced higher yields, crude protein contents, and frequency of grazing by cattle, than fall fertilization at 22 kg N/ha. No differences were found between the fall and spring applications at 34 kg N/ha. Source of fertilizer material may also affect the manner in which vegetation responds to fertilization. Beaton and Head (1963) measured short term P uptake by oats from several fertilizer sources. They found mono-ammonium phosphate produced the greatest P uptake and anhydrous dicalcium phosphate the least. In one long term experiment in which several sources of P fertilizer were used, the source causing the greatest uptake of P in plants varied with soil type and plant species (Ensminger and Pearson 1957). Power et al. (1973) reported greatest recovery of fertilizer N from corn tops occurred with NH^NOg when compared to other materials including calcium nitrate and urea. Power (1979) tested NH^NOg, urea formaldehyde, and three different, sulfur coated ureas, and found responses by vegetation varied with the fertilizer material used. In southeastern Montana, mulching had an affect on vegetational response to fertilization (Farmer et al. 1974). On spoils, fertilized mulch plots had greater grass yields than unmulched plots with the same fertilizer rates. On spoils covered with 8 inches of cover-soil, fertilization had no effect on seedling emergence of unmulched plots, while seedling density decreased on mulched plots. Method of placement of fertilizer in a soil system has an affect on how plants will respond to treatment. Injury to germinating seeds can be caused by placement of N and K fertilizer directly with the 19 seed (Tisdale and Nelson 1975)* This injury was due to restriction of available moisture or toxicity caused by an increase in concentration of soluble salts by the fertilizer. eliminated by other methods Injuries can be lessened or of fertilizer application such broadcasting or selection of non-ammonical fertilizer sources. et al. (1968) c o m p a r e d drill p l a c e m e n t of f e rtilizer broadcasting on sub-irrigated meadows in Nebraska. as Moore P w ith They found drill placement of fertilizer reduced plant density and yield when compared to the broadcast method. The decreases were attributed to the drying out of sod near the drill rows. Percent P in forage, root activity of legumes, and utilization of fertilizer P by plants was lower under drill treated sites. Incorporation of fertilizer P into the soil increased vegetation response when compared to P applied on the surface (Tisdale and Nelson 1975). immobile compared to other nutrients, Fertilizer P is relatively and incorporation allows plant roots to come into direct contact with fertilizer P. Nutrient Cycling in Mine Land The literature on nitrogen cycling in range and mine land is more extensive than that for P and K cycling. primarily on N cycling. This section focuses It is not intended to be a complete summary on nitrogen cycling, but the important aspects are covered. Addition of fertilizer N has been a v a i l a b l e N in soils shown to increase (Houston and Hyder 1975, Power plant 1972b). Fertilizer N enters and functions in the nitrogen cycle through various processes such as nitrification, plant uptake, and loss. 20 Soil organic matter is important in supplying plant available nitrogen through mineralization (Tisdale and Nelson 1975). On mine land, adequate amounts of organic matter are often lacking to provide sufficient nutrients for vegetational growth. Bauer et al. (1978b) noted that a characteristic common to all spoil material was the lack of organic matter. » In cases in which topsoiling practices were used, the occurrence of nutrient deficiencies may vary depending on the suitability and thickness of the applied cover-soil. The organic matter content of stockpiled topsoil may decrease with length of storage (Argonne National L a b o r a t o r y 1979). Parkinson (1979) suggested that addition of waste materials rich in cellulose, lignin, chitin, etc. could be a means of increasing organic matter content of spoil material, introduced. provided decomposing microorganisms were also Omodt et al. (1975) estimated it would take at least 350 years for organic matter on mine land spoils to naturally accumulate to levels found in undisturbed soils of western North Dakota. raise organic matter levels to I percent in mine spoils, To it was calculated 291 metric tons of manure/ha applied annually for a forty year period would be needed. Due to the impracticality of applying this much manure, it was concluded the salvaging and redistribution of topsoil on spoils would be more practical for maintaining organic matter levels of mine soils. Losses of N from the nitrogen cycle in mine land are divided into leaching* biological, gaseous, and geological. In Great Britian, nitrogen was the main nutrient lost by leaching on mine land (Marrs and Bradshaw 1980). Losses of N, P, Ca, and Mg were greater than 21 imputs from natural sources. of In the Northern Great Plains, leaching NOg-N was of little concern because of insufficient precipitation (Power 1972a). Power and Alessi (1971) found no accumulation of from leaching below 90 cm in a grassland system. NOg-N Young and Rennick (1982) noted supplemental irrigation on mine land at Colstrip caused leaching of NOg-N to occur. . The influence of irrigation on increasing NOg-N loss from mine land should not be ignored. Biological losses of plant available under certain circumstances. N can occur in mine land Berg (1980) noted addition of organic matter such as straw mulch could decrease plant available soil NOg-N. Reuszer (1957) stated that N was needed by microorganisms for the decomposition of added organic material. material contains If the added organic insu f f i c i e n t q u a n t i t i e s of N for its o w n decomposition, microorganisms will utilize indigenous or fertilizer N in the soil (Tisdale and Nelson 1975). An indication of whether or not organic matter contains sufficient quantities of N for its own decomposition is its carbon/nitrogen ratio (C/N). As a generalization microorganisms will utilize N from the soil to decompose organic matter with C/N ratios above 30. Organic matter with C/N ratios below 30, have sufficient N to meet the needs of microbial decomposers, while material with C/N ratios below 20 have excessive amounts of N that can be released into the soil system through m i crobial decomposition. Gaseous losses denitrification, of N can occur through three mechanisms: chemical reactions involving nitrites, losses of ammonia gas (Tisdale and Nelson 1975). and volatile Conversion of NHjj-N 22 to NOg-N occurred in exposed Palocene shales on mine lands,and the NOg-N subsequently lost possibly due to denitrification (ARS and NDAES 1 9 7 5 ). Urea applied via broadcast methods on Bromus inermis was suspected of being lost to the atmosphere, since only 4? percent of the applied N was accounted for in the soil and plants (Power et al. 19 7 3 ). Volatilization of NHjt-N placed on the surface of alkaline soils may also take place (Tisdale and Nelson 1975). Geological losses of fertilizer material may also occur in mine land. Ammonium added as fertilizer to spoil material in east central Texas was converted to non-exchangeable NHj1-N (Hons and Hossner 1979). The mine soil had a non exchangeable NH11-N retention capacity that ranged from 4.1 to 7.8 meq NHj1+ / 10O g, while lignite had a capacity of 46 meq NHj1V l O O g. 23 MATERIALS AND METHODS Experimental Design The study area was located in Mining Area A of Western Energy Company’s Rosebud Mine at Colstrip9 Montana9 and covered approximately 2,590 m 2. Spoil piles on the study site and adjacent areas were graded and leveled. 1981. Coversoil was placed on the leveled spoils in late September Average cover-soil depth was between 60 and 70 cm, but ranged from 46 to 100 cm. Cover-soil was placed on the site with two scrapers pulled by a four wheel drive tractor. have occurred during this process. Soil compaction may In addition, water was sprayed on the cover-soil after every few passes of the scraper for dust control. After placement of cover-soil, the area was chisel plowed twice to a depth of 25 cm and disced once on the contour in early October to alleviate compaction. topsoil and subsoil, 1976 (J. Cundiff, The cover-soil material, a mixture of both was obtained from a storage pile constructed in personal communication). Seedbed preparation commenced on November 3, 1981. At this time the study site was chisel plowed twice, disced and harrowed. The northern third of the study site was accidently ripped prior to seedbed preparation. To correct this situation, the ripped areas received an additional chisel plowing and discing. Three 35 by 23 m blocks were laid out in parallel with the long side of the blocks running approximately east to west. numbered from I to 3 running from south to north. Blocks were Figure I shows the 24 layout of blocks and experimental plots. separated each block. Within each block A buffer zone of 2.5 m twenty-four, 5 by 5 m plots were placed in four rows of six and separated by a I m buffer zone. Plots were numbered by rows within each block from south to north. Numbering began in each southwestern corner. block w i t h the plot located in the All plots were consecutively numbered from 1 to 72 starting with Block I. Each plot was broadcast seeded by hand on November 3, 1981 to achieve a fall dormant seeding. Table I lists the plant species and seeding rates used on all study plots. All species, except Astragalus cicer are considered native to the Northern Great Plains. A sheep’s foot cultipaeker was pulled over the site after seeding to ensure a firm seed bed. Table I. Seed mixture and rates used on fertilization study. Scientific name Common name Agroovron dasvstachvum A. smithii A. trachvcaulum Androoogon hallii Bouteloua curtioendula B„ gracilis Calamovilfa longifolia Panicum virgatum Stioa viridula Astragalus cicer Petalostemon ouroureum Atriolex canescens Critana thickspike wheatgrass Rosana western wheatgrass Revenue slender wheatgrass Sand bluestem Pierre sideoats grama Lovington bluegrama Goshen prairie sandreed Pathfinder switchgrass Lodorm green needlegrass Lutana cicer milkvetch Kaneb purple prairie clover Wytana fourwing saltbush total kg/ha pure live seed 3.5 4.7 3.6 6.8 1.1 0.8 1.1 2.1 2.4 4.3 3.8 3.6 37.8 # seeds/ m^ 118 118 118 118 118 118 65 118 118 118 237 43 1407 . Figure I. Field design. □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ □□□□ BLOCK 3 SCALE: BLOCK 2 BLOCK I 26 A randomized block design with three replications was used for this study. Twenty-four combinations of fertilization rates were tested and randomly assigned to plots within each block. The fertilizer treatments consisted of four rates of nitrogen (0, 14, 28, and 56 kg/ha), three of phosphorus (0, 112, and 168 kg/ha), and two of potassium (0 and 28 kg/ha) in complete factorial combination. lists the twenty-four fertilization treatments by codes. Table 2 Throughout the rest of this thesis, fertilization treatments will be referred to by these codes. Table 2. Fertilizer treatment combinations. Treatment NO PO NO Pl 12 NO P168 Pl 12 NO NO P168 NO PO N14 PO N14 Pl 12 KO0 KO KO K28 K28 K 28 KO KO Treatment N14 P168 P112 . N14 NI 4 P168 PO N14 N28 PO N28 Pl 12 N28 P168 N28 Pl 12 KO K28 K28 K28 KO KO KO K 28 N28 N28 N56 N56 N56 N56 N56 N56 Treatment P168 PO PO Pl 12 P168 P112 P168 PO K28 K28 KO KO KO K28 K28 K28 0Numbers following elemental designation refer to application rate (N = 0, 14, 28, and 56 kg/ha; P = 0, 112, and 168 kg/ha; K = O and 28 kg/ha). The source of nitrogen was ammonium nitrate (34-0-0). was applied in the form of triple superphosphate potassium in the f o r m of potassium chloride Phosphorus (0-44-0), (0-0-60). and The experimental plots were fertilized with their respective mixtures on April 29, 1982. plots by hand. The fertilizer was uniformly broadcasted over the Only one person fertilized plots in a block to ensure consistency of application within each block. 27 Vegetation Sampling A number of vegetational parameters were estimated on this study. These parameters included density, aerial biomass, canopy cover, frequency, diversity, evenness, and richness. Figure 2 illustrates the sampling scheme used in each plot. Three parallel transects were placed in each plot 1.5 m apart. The transects ran parallel to the east/west sides of the plot, and the two outer transects were placed I m from the edges. The west end of each transect was permanently marked. Density Plant density was counted on the experimental plots on May 21 and 22, and again between July 6 and 9, 1982. Six, 20 x 50 cm sampling frames were used in each plot during May. Transects I and 3 each had 3 frames located 1.5 m apart (Figure 2). Density was defined as the number of individual plants per sample frame. For species with a bunch or tillering growth habit, a plant was considered an individual if there was a gap of at least I cm between its base and the base of another plant. identifiable, Plants wer e counted on a species or placed into plant classes. basis w h e n Nomenclature for scientific names of plant followed U.S.D.A. (1982). The same sampling procedures for May were used for the July measurements except that 3 frames were added to Transect 2. Field observations in late June indicated six sample frames were not enough to include the majority of plant species found in each plot. Only approximately 50 percent of the species located in each plot were 28 Figure 2. Vegetation sampling design for each experimental plot. 2.5m Trx.3 SCALE N h 0m KEY: Aerial f Trx.1 Trx.2 I Canopy * Biomass Cover and Density I 29 found within the six sample frames. Preliminary sampling was conducted in late June to determine the number of sample frames needed to include the majority of plant species found on the study plots. The results of the preliminary sampling are shown in Figure 3. Cumulative number of plant species were graphed against number of sample frames. For the sample plot, 10 sample frames were optimal. The 10 frames included 12 of the 16 species found on the plot. Due to physical limitations of the study plot, only nine sample frames were used. Aerial Biomass Vegetation was clipped for aerial biomass between July 20 and 25, 1982 for cool season perennial grasses, annual grasses, forbs, Salsola kali, legumes, and shrubs. August 12 and 13, 1982. Warm season grasses were clipped on The vegetation was clipped at time of maximum aerial biomass production. A total of four, V U m ^ frames were used to sample each plot. Two frames were located on both Transects I and 2, and located 1.5 m apart ( Figure 2). level. Standing vegetation was clipped to within I cm of ground The clipped vegetation was oven dried at 67° C for 48 hours, and then weighed to the nearest tenth gram. Canopy Cover Canopy cover was recorded for all plots between July 20 and 25, 1982. . Nine 20 x 50 cm sampling frames were used for each plot. Three sample frames were located 1.5 m apart on each transect (Figure 2). Canopy cover was read for each plant species in a sample by methods 30 CUMULATIVE NUMBER OF SPECIES Figure 3. Preliminary sampling to estimate number of frames needed. t 0 4 8 12 NUMBER OF FRAMES (20 X 50 cm) 31 described by Daubenmire (1959). Daubenmire’s cover classes were modified for this study as listed in Table 3. Table 3. Cover classes used for canopy cover sampling. Cover class Range of percent canoDV cover I 2 ■ 3 4 5 6 7 8 9 10 11 12 13 14 0 5 10 20 30 40 50 60 70 80 90 95 0 > 5 - 10 . - 20 - 30 - 40 - 50 - 60 - 70 - 80 - 90 - 95 > 100 100 Midpoint of cover class 0 2.5 7.5 15 25 35 45 55 65 75 85 92.5 97.5 too Frequency Frequency was calculated from canopy cover data, and defined as the percentage of canopy cover sample frames in which a particular plant species occurred. Frequency for a plant species was based on a total of 27 sample frames located, in the three plots representing a particular fertilization treatment. Diversity Diversity was calculated for each fertilization treatment using the. Shannon Function. Pielou (1975) listed the Shannon Function as: . S H = Z Pilog P1 i=l where Pi is the proportion of the plant community belonging to the ith 32 species and S the number of species present in the sample. The portion of the total percent canopy cover of each plant species in a fertilization treatment represented the Evenness values. ■ Evenness was defined as the distribution of total plant community canopy cover among the individual plant species. The evenness index used was described by Odum (1971) as; e = H*/log Sf where H* is the calculated Shannon Funtion and S the number of species found in the sample. Richness Richness was defined as the total number of individual plant species found in each fertilization treatment. Richness was based on the species found in the canopy cover data for each treatment. Soil Sampling Samples for baseline soils characterization were taken on October 22 and 23, 1981. One composite sample was taken per block and broken into depth intervals of 0-30 cm, 30-60 cm, and 60-120 cm. Each composite consisted of fifteen, 4.8 cm diameter cores taken with a Giddings probe at randomly picked locations within each block. The composites for each depth interval and block were thoroughly mixed, placed in separate sample bags, and immediately frozen. After transport, the frozen samples were immediately thawed, air dried for 72 hours, then ground. Soil samples for each block and depth interval were analyzed for 33 percent total N1 NOg-N, extractable P and K, percent organic matter, electrical conductivity (EC), sodium absorption ratio (SAR), pH, exchangeable Ca, Mg, and Na; cation exchange capacity, and particle size. Total N was determined by the semimicro digestion/distillation method described by Black (1965). Kjeldahl Nitrate was analyzed at the Montana State University Soil Testing Laboratory which used procedures based on the work of Doner et al. (1973), Sims and Jackson (1971), and West and Ramachandran (1966). Extractable P and K; cation exchange capacity; exchangeable Ca, Mg, and Na; and SAR were determined by methods described by Sandoval and Power (1977). Sims and Haby (1970) described methods used for determining percent organic matter. An aliquot from a saturated soil paste for each sample was used for determining pH and EC. Particle size analysis was conducted using the hydrometer method described by Black (I965). In addition C/N ratios were calculated for each soil sample using percent total N and percent organic matter content. Organic C content was assumed to be 58 percent of the organic matter content (Brady 1974). Post fertilization soil sampling was conducted on October 11, 1982. Two sets of soil samples were taken. The first set involved compositing soil taken from plots containing all combinations of P and K fertilization for a specific level of N. This was done for all 4 levels of N in each block for the depth intervals of 0-15 cm and 1530 cm. This set of soil samples was analyzed for total N, NOg-N, and percent organic matter. ,The second set of samples consisted of compositing soil taken from plots containing all combinations of N and K for a specific level of P. This >ras done for all levels of P in \ 34 each block for the depth interval of 0-15 cm. analyzed for extractable P. This set of samples was Preparation and analysis of soil samples did not differ from procedures used for baseline soil sampling except that soil samples were taken with the use of an Oakfield probe, two cores were taken randomly from each of the appropriate plots to form the composite, and NOg-N was analyzed using methods described by Richards (1954). Haby and Larson (1976) found the two techniques of NOg-N analysis used on this study were highly correlated (r2=0.94). The use of the two techniques should not affect comparision of NOg-N levels. Soil moisture data were obtained with a neutron probe. McHenry (1963) described the theory and application of the neutron probe in measuring soil moisture. Two neutron probe access installed adjacent to the study site. One access tube was located north of the site, and the other to the south. installed to a depth of 160 cm. tubes were The access tubes were Neutron probe readings were taken once a month during the growing season at 15 cm intervals to a depth of 90 cm, then at 30 cm intervals to a depth of 150 cm. Field generated data were converted to percent volumetric soil moisture content by use of factory calibration equations. Statistical Analysis Vegetation Analysis of variance (ANOVA) methods were used to analyze the density, aerial biomass, and canopy cover data. For density the plant classes of cool season perennial grasses, 35 warm season grasses, annual grasses, annual forbs, biennial forbs, perennial forbs, legumes, shrubs, and total vegetation were analyzed. A five factor AWOVA consisting of time, blocks, N, P, and K was used to evaluate the data. density data. Two mean square errors were calculated for the The first error was calculated by pooling the sum of squares for all interactions involving blocks without time, and utilized in calculation of the F statistic of all main effects and nutrient interactions not involving time. calculated by pooling the sum The second error term was of squares for all interactions involving both time and blocks, and utilized in calculating the F statistic for time and all interactions involving time. Analysis of variance was conducted on aerial biomass data for cool season perennial grasses, warm season grasses, annual grasses, forbs, Salsoia kali, legumes, shrubs, and total vegetation. For canopy cover the plant classes of cool season perennial grasses, warm season grasses; shrubs, annual, biennial, and perennial forbs; and total vegetation were analyzed. legumes, A four factor ANOVA consisting of blocks, N, P, and K was conducted on the aerial biomass and canopy cover data. The error was calculated by pooling the sum of squares of all interactions involving blocks. The null hypothesis tested in the study was that the mean responses of a particular data parameter for a plant class are equal for all fertilization treatments. Significance was defined as the rejection of the null hypothesis with a probability greater than 95 percent (p<0.05). Rejection of the null hypothesis with a probability greater than 99 percent (p<0.01) was also noted. 36 Soils Post fertilization soil data taken in October 1982 were analyzed using ANOVA. P were each Total N, NOg-N, percent organic matter, and extractable analyzed using considered a replication. one factor ANOVA. Each block was Significance and testing of the null hypothesis were conducted in the same manner described for vegetation. 37 STUDY SITE DESCRIPTION Location The study site was located in Mining Area A of Western Energy Company's Rosebud Mine at Colstrip, Rosebud County, Montana (Figure 4). Colstrip is approximately 48 km south of Forsyth, MT at an elevation approximately 980 m above sea level (Meyn et al. 1976). The legal description of the study site was the center of the NW 1/4, SW 1/4, S.33, T.2N, R.41E of the Montana Principal Meridian. Figure 4. Study site location, Colstrip, Montana. C OLSTRIP ■ Tonography Resistance of geologic strata to erosion largely determines the topography of the Colstrip area (Skilbred 1979). The landscape is dominated by rolling prairies with alternating ridges, drainages, and sandstone bluffs. Most streams drain to the north and eventually flow into the Yellowstone River. 38 The study site was located on a north facing hilltop at an elevation of approximately 1036m above sea level. fairly level with The study site was convex slopes of 0 to. 3 percent. The slopes gradually increased in steepness from south to north across the study site. Climate Continental climatic conditions exist in the Colstrip area with cold winters and warm summers (Meyn et al. 1976). warmest month, ColstripeS July is usually the while January the coldest (Munshower and DePuit 1976). climate is semiarid with p r e c i p i t a t i o n a n n ually (N.O.A.A. an 1981). average of 40.1 The m a j o r i t y cm of of the precipitation comes in the form of rain during the months of April, May, and June. Mean long term annual and monthly precipitation and temperature data were obtained from N.O.A.A. (1981). Monthly precipitation and temperature data for the 1982 growing season were obtained directly from the U.S. Weather Service reporting station in Colstrip. Table 4 lists the monthly averages, and deviations from the long term averages for temperature and precipitation at Colstrip. During the study temperatures were below normal during April and May, while above normal during March,. June, July, and. August. Precipitation was below normal between the months of April and August, but above normal in March / 39 Table 4. Mean monthly temperature, precipitation and deviations from the long term norm, Colstrip, Montana, 1981-82. Mean temperature °C Month October November December January February March April May June July August 7.2 5.6 -11.1 -3.9 2.2 5.0 10.6 17.2 23.3 24.4 Deviation 0C -2.0 5.9 —5.0 -0.9 2.1 -2.1 -1.4 0.1 1.4 3.2 Mean precipitation cm 3.8 1.3 2.0 2.5 0.5 5.6 3.3 4.1 4.8 2.6 2.9 Deviation cm 1.2 -0.4 0.4 1.1 -0.9 3.7 -1.4 -2.2 -3.6 -0.4 —0.6 Vegetation Rangeland in eastern Montana is generally classified as mixed grass prairie association (Payne 1973). The major subtype of this association in the Colstrip area is ponderosa pine savannah. The dominant species in this subtype are Agropvron smithii, A. SPicatumf and Bouteloua gracillis. Ross and Hunter (1976) classified the climax vegetation of the Colstrip area as a complex of Silty and Clayey range sites. In addition the Forest-Grassland complex range site is also found in some locations surrounding Colstrip. Approximately 53 percent of the rangeland in Rosebud County is in good to excellent condition. Prior to placement of cover soil on the study site, Salsola kali and other annual forbs were the dominant species. After placement of cover soil in late September 1981, no germination or plants were observed on the site for the remainder of the year. 40 Geology Veseth and Montagne (1980) described the geologic history of eastern Montana including the Colstrip area. the last major Cretaceous sea, Following the retreat of soft nonmarine sediments spread over the plains and basins of eastern Montana. . During the early Tertiary period of the Paleocene epoch, creation of the Fort Onion Formation occurred. The Tongue River member encompasses the Colstrip area. of the Fort Union Formation The Tongue River member is composed of soft interbedded light yellow to yellowish gray lenticular sandstones; gray claystones and shale; thin dark carbonaceous shales; coal seams; and clinker beds. Clinker beds were formed by burning shallow coal beds baking overlying sediments into reddish beds of various hardness. Clinkers often form resistant caps on buttes and ridges of the Colstrip area. Soils On nearly level to moderately steep hills and plains, a mixture of Camborthids and Torriorthents are recognized (Schafer et al. 1979). A mine soil profile typical of the study site was described acc o r d i n g to Soil T a x o n o m y (Soil Survey Staff 1975), modifications for mine soils suggested by Schafer (1979a). profile description is listed in Table 22 of Appendix A. the study site were classified as Typic Ustorthents. with The soil The soils of Using unofficial soil series names from Schafer (1979a), the study site was placed into the Cow Creek Series. Evidence from the soil profile suggested that original topsoil and subsoil from the one lift cover-soil stockpiling operation did not thoroughly mix. Pockets of strongly contrasting dark material, possibly remanent of former A horizons were found scattered throughout lighter colored material which may have been the original subsoil. It is possible the nutrient status of these two materials differ, and may cause variation in vegetational growth within plots over the study site. Using criteria developed by Schafer (1979b), classified as Land Capability Class IV. the study site was This class is suitable for cultivated pasture and rangeland, but not suitable for row crops. The major limitation of the study site was due to lack of topsoil as defined as material from the original A horizon. Topsoil was not segregated from subsoil during the stockpiling operation. At least 15 cm of original topsoil would have been needed on top of the coversoil material in order for the site to have qualified as Class III. Baseline data for soils on the study site is listed in Table 5 by block and depth interval. Generally little variation properties occurred for each depth interval by block. in soil One exception was the sodium absorption ratio for the 0-30 cm interval in.Block 1 and the 30-60 cm interval in Block 3. The SAR values were higher than those found for other blocks and depth intervals, but not high enough to interfer with plant growth and development. Volumetric soil moisture content is graphed in Figure 5. In November 1981 and the following April, soil moisture content of the upper soil profile was generally low. By May and June, soil moisture Table 5. Baseline soils data of the study site, October, 1982 Soil parameter NOo-N (ppm) 0-30 Block 1 30-60 60-120 7.4 9.5 5.5 .052 .051 .047 1.6 0.9 0.9 10.0 18.2 11.1 2.0 2.0 0.6 87.0 116.0 122.0 17.19 28.87 24.88 2.80 5.32 4.53 0.18 0.10 0.06 8.0 11.0 11.1 7.79 7.49 7.63 1.08 0.87 0.91 44 48 49 18 23 23 29 33 33 loam loam loam 5.0 CO on O 8.2 13.0 Total N (*) .056 .059 Organic matter ( % ) 0.9 0.7 C/N 6.9 9.3 1.4 Extractable P (ppm) 2.2 Extractable K (ppm) 118.0 121.0 20.82 24.16 Exchangeable Ca (meq/IOOg) Exchangeable Mg (meq/IOOg) 4.82 3.83 0.12 Exchangeable Na (meq/IOOg) 0.02 10.2 Cation Exchange Capacity (meq/IOOg) 11.5 7.88 7.68 pH Sodium absorption ratio 3.2 0.92 % sand 43 43 % clay 23 23 % silt 34 34 loam Texture class loam Soil death (cm) bv block Block 2 0-30 30-60 60-120 0-30 1.2 18.3 1.2 98.0 19.31 3.48 0.10 8.6 7.81 1.22 46 23 31 loam Block I 30-60 60-120 8.0 8.6 .046 .055 0.8 0.9 8.4 11.3 2.8 1.7 108.0 108.0 30.52 22.09 4.87 4.71 0.16 0.09 9.8 10.3 7.54 7.85 0.98 2.27 44 46 23 23 31 33 loam loam 4.7 .041 1.2 17.0 1.2 88.0 11.07 3.43 0.16 7.2 7.55 0.84 46 20 34 loam Figure 5. KEY: Volumetric soil moisture content, 1981-82. NEUTRON PROBE A C C E S S TU B E , NORTH END NEUTRON PROBE A C C E S S TU BE, SOUTH END 0- 5/ 17/82 15 30' SOIL DEPTH (cm) 46- 6076 80 - • 120 " 160 - 4— +— h PERCENT VOLUME OF SOIL WATER 0- 8 / 12/82 1630 - SOIL DEPTH (cm) 46 60 76 - 80- \ 120 / H-----1-----1-----1 PERCENT VOLUME OF SOIL WATER 44 content near the surface had increased from precipitation events. the summer progressed, decreased As soil moisture content of the study site 45 RESULTS AND DISCUSSION Introduction The objective of this study was to determine first year.response of a native species mixture to 24 fertilization.treatments on mine soils at Colstrip, combinations of MT. N, The P, and treatments K. consisted Vegetational of factorial response to the fertilization treatments was evaluated by measuring density, aerial biomass, canopy cover, and frequency for the various plant classes comprising the newly established plant community. diversity, evenness, fertilization treatment. site at the end of and r i c hness w e r e In addition calculated for each Soil analyses were conducted on the study the first growing season to determine how fertilization affected the nutrient status of the soil. Results and Discussion are divided into sections by fertilizer element and related topics. The phosphorus, nitrogen, potassium, and nutrient interactions sections detail how fertilization affected the plant community and its components. . In addition, these sections include results and discussion on post fertilization soil analyses. The adequacy of regression model section discusses the limitations of the regression equations. Appendix B contains all the analysis of variance tables referred to in this study. Generally, experimental results are discussed in terms of plant classes. For the person interested in the results of a specific fertilization treatment on a particular species, Tables 30 to 33 of Appendix C contain this information listed by data parameter. 46 Phosphorus This section discusses the effects of P fertilization by plant classes, plant community development, and residual effects on the soil. Phosphorus fertilization affected the density, aerial biomass, and canopy cover response of several plant classes (Tables 23, 24, and 25; Appendix B). Regression analyses evaluated the influence of P on the plant classes signficantly affected, by fertilization. fertilization was defined as the independent variable while vegetational response as the dependent variable. quadratic models were evaluated. Level of P Both linear and Significance was defined as the rejection of the null hypothesis (all slope coefficients equal to zero) with a probability greater than 95 percent. test (LOF) described by Neter and Wasserman The lack of fit (1974) tested the appropriateness of regression models in.which slope coefficients proved significant. Lack of fit tests the null hypothesis that the expected value of the dependent variable is equal to the regression model. null Significance for the LOF test occurs with an acceptance of the hypothesis at a probability greater than 95 percent. For regression models that proved non significant for the LOF test, alternative models were tested until an appropriate model was fitted. Regression models for which slope coefficients an d appropriateness proved significant are illustrated in Figures 6 to 15. The corresponding p values determination illustrations, are also for the listed. t test and On each of coefficients the of regression the dot r e p r e s e n t s the m e a n r e s p o n s e for all 47 combinations of N and K fertilization with a specific level of P for the plant class and data parameter being estimated. The verticle line drawn through each mean response represents the 95 percent confidence interval that contains the mean response, and is based on the eight combinations of N and K fertilization associated with a particular level of P. Cool Season Perennial Grasses Table 6 summarizes the data collected for cool season perennial grasses. Phosphorus fertilization did not significantly affect the data parameters tested for this plant class (Table 23, 24, and 25; Appendix B). 33, Frequency also was unaffected by P fertilization (Table Appendix C). Meyn et al. (1975) stated that evidence from Colstrip and the literature indicated fertilization has unpredictable effects on perennial grasses during the first growing season. This unpredictability in cool season grass response may be due to variation in factors such as precipitation,' soil temperature, and site characteristics. Cool season grasses may not have responded to P fertilization due to climatic conditions. During April and May 198.2 average monthly temperatures were lower than the long term average for Colstrip (Table 4). Precipitation was also below normal for the months from April to August (Table 4). The lower temperatures and precipitation may have reduced growth rates of cool season perennial grasses. climatic c o n d i t i o n s such as w a r m e r If different t e m p e r a t u r e s and higher precipitation existed during the first season of growth, responses to 48 fertilization may have occurred for cool season perennial grasses. Table 6. Vegetational statistics of cool season perennial grasses by fertilization treatment, 1982. Fertilizer treatment NO NO NI 4 N14 N28 N28 N56 N56 NO NO NI 4 N14 N28 N28 N56 N56 NO NO N14 N14 N28 N28 N56 N56 P168 P168 P168 P168 P168 P168 P168 P168 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 P112 Pl 12 Pl 12 PO PO PO PO PO PO PO PO Density May plants/nr Density July plants/nr Aerial biomass kg/ha Canopy cover 100.0 105.6 90.0 95.0 95.6 83.9 69.4 98.3 106.7 80.0 81.7 95.6 103.9 103.3 77.2 89.4 81.7 94.4 115.6 88.9 63.3 90.6 100.6 138.3 100.7 85.6 74.8 98.5 94.1 84.8 55.9 80.0 87.8 64.4 76.3 79.6 90.4 107.8 75.6 94.8 81.9 81.9 105.9 87.0 70.4 85.6 91.5 109.3 327.6 319.6 226.8 222.4 222.8 262.8 347.6 261.2 234.0 255.6 246.0 258.0 338.8 217.6 210.4 164.0 212.0 105.6 202.8 306.4 165.2 170.8 169.6 243.6 30.9 26.0 20.0 19.4 18.0 27.4 16.4 20.6 23.1 16.2 20.4 27.1 24.8 27.7 21.0 23.7 13.2 12.2 15.4 20.0 21.6 16.4 18.6 23.1 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 % Warm Season Grasses Table 7 sumarizes the data collected for warm season grasses. Phosphorus fertilization significantly affected the density, aerial biomass, and canopy cover of this plant class (Tables 23* 24, and 25; Appendix B). During May, density of warm season grasses was zero, but by late June emergence occurred. A four factor ANOVA evaluated the effect P 49 fertilization had on the July density data (Table 23, Appendix B). Phosphorus proved significant for the July density data and tended to decrease grass density as fertilization increased (Figure 6). Little information existed in the literature on how fertilization affects emergence of warm season grasses. Welch et al. (1962) reported fertilization of this plant class in Texas did not affect emergence. Table 7. Vegetational statistics of warm season grasses by fertilization treatment, 1982. Fertilizer treatment NO NO N14 N14 N28 N28 N56 N56 NO NO N14 N14 N28 N28 N56 N56 NO NO NI 4 N14 N28 N28 N56 N56 P168 P168 P168 P168 P168 P168 P168 P168 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 PO PO PO PO PO PO PO PO KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 Density May plants/m^ 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 Density July plants/m2 11.9 14.1 5.2 10.4 13.3 11.1 1.5 5.6 5.9 3.7 23.7 8.5 8.5 11.1 8.9 17.4 22.2 19.3 29.3 18.9 13.7 18.9 10.0 17.8 Aerial biomass kg/ha 2.4 1.2 1.6 2.8 2.8 3.2 2.0 2.4 1.2 3.2 4.4 1.6 1.2 3.2 1.6 3.2 8.8 7.6 7.2 8.8 5.6 6.4 4.8 3.2 Canopy cover % 1.1 1.0 0.7 1.0 0.6 1.2 0.0 0.5 0.7 0.6 1.2 1.2 0.7 0.7 0.8 0.8 1.4 1.3 1.9 1.8 1.1 1.4 1.3 1.8 50 NUMBER OF PLANTS PER M SQ Figure 6. Mean density of warm season grasses in response to P fertilization, July, 1982. —0 . 0 5 9 X +• < I 8.47 0.0 I 0.38 PHOSPHORUS RATE ( k g / h a ) 51 It should be noted at this point that P fertilization did not significantly affect the first growing season density of other plant classes analyzed in this study (Table 23, Appendix B). Other studies agreed with these results (Aldon I978, DePuit et a I. 1978, Holechek 1976). Phosphorus fertilization tended to decrease the aerial biomass (Figure 7) and canopy cover (Figure 8) of warm season grasses. Several rangeland studies reported similar results (Johnston et al. 1967, Wight and Black 1979). Only P fertilization had an effect on the frequency of warm season grasses (Table 33, Appendix C). To better show this effect, frequency is listed in Table 8 by taking the mean of all combinations of N and K at a specific level of P for each plant species. of w a r m season grasses tended to decrease w i t h Frequency increased P fertilization. Identification of warm season grasses by species was not possible until mid August due to the lack of development in individual plants. As a result, green bristle grass (Setaria viridis), a warm season annual, was counted as a perennial. grass composed only a small In mid August, green bristle portion of the warm season grass population. Other species on the study site included sideoats grama (Bouteloua ourtipendula). blue grama (JBa. gracilis), and prairie sandreed (Calamovilfa longifolia). Increased competition from other plant classes as the result of P fertilization may have caused the adverse effects on warm season grasses. Emergence of warm season grasses did not occur until late 52 AERIAL BIOMASS ( k g /h a ) Figure 7. Mean aerial biomass of warm season grasses in response to P fertilization, August, 1982. Y = -0.027X p < 0.01 r eq + 6.28 0.64 0.70 t p < 0.01 Y =r 0 .0 0 0 2 X -0.059X +6.55 ------------------------- 1--------------- 1 112 P H O S P H O R U S RATE ( k g / h a ) 168 53 PERCENT CANOPY COVER Figure 8. Mean canopy cover of warm season grasses in response to P fertilization, July, 1982. Y = p < r eq. -0.006X + 1.46 0.01 = 0.63 P H O S P H O R U S RATE ( k g / h a ) Table 8. Mean frequency of plant species by P fertilization rate, July 1982 Plant class/soecies Cool season perennial grasses: Acronvron dasvstachvum A. smithii A. snicatum A. trachvcaulum A. trichonhorum BrPfflU? inermis Hprdeun .lutatu® Stina viridula Warm season grasses: Annual grasses: Avena fatua Bromus ianonicus B. tectorum Annual Forbs: Amaranthus sp. Camell na microcarpa Chenonodium album c. leptephYllum Descurainia pinnata D. gppbia Ellisia nvctelea Helianthus annuu? H. netiolaris P168 Pl 12 £Q 92.6 74.1 0.0 14.8 0.5 0.9 2.3 13.0 28.7 93.5 76.4 0.5 20.8 0.9 0.0 0.5 14.4 31.5 92.1 64.8 0.0 17.1 0.0 0.0 0.5 13.0 47.7 3.2 34.3 10.2 3.7 34.3 6.9 2.3 25.5 3.2 8.3 10.2 14.4 4.2 0.5 1.8 0.9 1.8 1.4 7.9 4.2 8.3 4.6 0.5 0.9 1.4 2.8 0.5 9.7 2.8 7.9 2.3 0.0 0.5 0.0 4.2 0.5 Plant class/snecies Kochia sconaria Lannula redowski Polygonum aviculare P. convolYUlU? P. rarnoaissimum Salsola kali SisYffltr'*nm ^ltlssifflUffl Sslanuip triflerun Vaccaria secetalis Xanthium strumarium Biennial forbs: Lactuca serriola Traconocon dubius Perennial Forbs: Ambrosia nsilostachva Taraxacum officinale Legumes: Astraealus cicsr A l S£-l Melilotus officinalis Petalostemon nurnureum Shrubs: Atrinlex canescens P168 Pl 12 0.5 0.5 0.0 0.5 53.2 55.1 6.0 5.1 7.4 6.9 100.0 100.0 0.0 1.9 2.8 5.1 1.4 1.9 0.0 0.5 20 0.5 0.0 50.9 2.3 6.5 99.5 0.0 5.1 0.0 0.0 8.3 6.0 6.5 5.1 10.2 7.4 0.5 0.0 0.5 0.0 0.5 0.5 87.5 0.5 23.6 18.1 82.4 0.0 22.7 18.5 85.2 0.0 22.7 24.1 22.7 19.0 16.2 55 June. By this time annual grasses and annual forbs had become established with aerial biomass and/or canopy cover significantly increased by P fertilization. On high P fertilized plots, these established plant classes may have outcompeted warm season grasses by decreasing available soil moisture through transpiration. The decreases in density and frequency indicated P fertilization affected distribution and emergence rather than individual plant growth* A decrease in the number of individual plants, rather than decreased growth of individual plants caused the reduction in aerial biomass and canopy cover. i Statistical analysis indicated a significant interaction of time x P fertilization affected warm season grass density response (Table 23, Appendix B). This interaction was expected. As discussed, P fertilization significantly affected density in July, but not in May due to lack of emergence. This difference in response to P fertilization between the two sampling times, resulted in the time x P interaction. Annual Grasses Table 9 summarizes the data collected for annual grasses. Phosphorus fertilization significantly affected the aerial biomass and canopy cover response of this plant class (Tables 23 and 24, Appendix B). Aerial biomass.(Figure 9) and canopy cover (Figure 10) tended to increase with P fertilization. Frequency of annual grasses, especially cheatgrass (Bromus tectorum) tended to also increase with P fertilization (Table 8). 56 Figure 9. Mean aerial biomass of annual grasses in response to P fertilization, July, 1982. Y = (0.028X p < 0.01 0.34 AERIAL BIOMASS (kg/ha) r sq. 4-4.38) PHOSPHORUS RATE (k g /h a ) 57 PERCENT CANOPY COVER Figure 10. Mean canopy cover of annual grasses in response to P fertilization, July, 1982. Y — 0 . 0 3 X +■ 4 . 11 p < 0 . 01 r sq. = 0 0.27 112 PHOSPHORUS RATE (kg /ha ) 1 68 58 Results of this study agreed with the literature for response of annual grasses to fertilization. increase in aerial Literature indicated annual grasses biomass and canopy cover with N or N plus P fertilization (Meyn et al. 1976, White and Halvorson 1980). Table 9. Vegetational statistics of annual grasses by fertilization treatment, 1982. Density May plants/m2 Fertilizer treatment NO NO N14 N14 N28 N28 N56 N56 NO NO NI 4 N14 N28 N28 N56 N56 NO NO NI 4 N14 N28 N28 N56 N56 P168 P168 P168 P168 P168 P168 P168 P168 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 PO PO PO PO PO PO PO PO KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 Density July plants/m2 5.6 5.0 2.8 5.0 4.4 4.4 3.3 3.9 9.4 2.8 6.1 6.7 3.3 2.2 1.7 3.9 3.9 3.9 0.6 3.3 2.8 1.7 2.8 5.6 5.6 0.7 2.6 5.2 2.2 0.4 2.2 6.3 4.8 1.5 0.4 6.7 5.6 5.6 2.2 3.7 1.1 5.9 0.4 1.1 1.5 1.9 1.9 5.2 Aerial biomass kg/ha Canopy cover 208.4 45.2 47.2 129.6 44.4 72.0 126.8 175.2 35.6 34.0 16.0 40.4 132.4 55.2 11.2 51.2 31.2 32.0 59.2 2.8 7.2 0.4 39.2 60.8 10.4 1.9 7.5 10.0 6.3 6.1 11.6 15.7 11.3 5.0 0.8 11.4 13.2 10.1 7.7 5.1 4.9 4.3 3.2 1.9 2.8 3.0 3.2 8.0 % Annual forbs Table 10 summarizes the data collected for annual forbs. Phosphorus fertilization significantly affected the canopy cover response of this plant class, but not the aerial biomass response of 59 Russian thistle Appendix B). (Salsola k a l i ) and forbs (Tables 23 and 24, Canopy cover of annual forbs tended to increase as P fertilization increased (Figure 11). Russian thistle, was the dominant annual forb and constituted approximately 40 to 60 percent of the community's total canopy cover (Table 32, Appendix C). Table 10. Vegetational statistics of annual forbs by fertilization treatment, 1982. Fertilizer treatment NO NO N14 N14 N28 N28 N56 N56 NO NO N14 N14 N28 N28 N56 N56 NO NO N14 NI 4 N28 N28 N56 N56 P168 P168 P168 P168 P168 P168 P168 P168 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 PO PO PO PO PO PO PO PO KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 Density May plants/m^ 25.6 23.3 31.1 21.1 20.6 20.6 45.0 15.6 17.8 26.1 23.3 13.3 21.1 28.9 30.6 17.8 23.9 31.7 22.2 24.4 27.8 11.7 21.1 28.3 Density July plants/m2 40.4 25.2 3 2 .2 37.8 34.8 28.9 41.9 36.7 35.2 35.6 37.8 31.1 34.4 59.6 31.5 28.5 34.1 41.5 41.9 35.9 33.0 27.4 26.7 40.0 SAKA* Aerial biomass kg/ha OF** Aerial biomass kg/ha Canopy cover 56 1320.8 2291.6 2622.0 1505.2 2962.8 1866.8 2203.2 2060.4 2520.8 1661.2 1961.2 2182.0 2913.6 1788.0 1998.4 1969.6 1698.0 1840.4 1713.6 994.0 2001.2 1510.8 2358.8 1875.6 307.6 57.6 124.8 132.8 129.2 414.4 216.8 389.2 66.8 290.8 201.6 197.2 149.2 217.6 186.4 76.0 32.8 129.2 178.0 107.2 85.6 183.2 92.8 217.2 67.1 85.7 101.6 82.7 92.1 75.4 105.5 83.2 93.1 91.0 82.0 74.1 75.7 80.6 92.5 87.4 83.9 73.4 70.1 75.4 77.0 57.7 73.7 76.3 •SAKA = Russian thistle. **0F = all forbs except Russian thistle, includes annual, biennial and perennial forbs. 60 PERCENT CANOPY COVER Figure 11. Mean percent canopy cover of annual forba in response to P fertilization, July, 1982. 80 — 60 - Y = 0.82X + r sq. = 40 73.93 0.29 - PHOSPHORUS RATE (kg /ha ) 61 Responses of annual forbs to P fertilization for canopy cover generally agreed with the literature, contradicted the literature. but aerial biomass data Several studies found N or N plus P fertilization increased canopy cover and aerial biomass of annual forbs, especially Russian thistle during the first growing season (Buchholz 1972, DePuit et al. 1978, Holechek 1976). Despite the lack of statistical significance, field observations indicated Russian thistle and other annual forbs had greatest aerial biomass on heavily P fertilized plots. indicated Methods described by Snedecor and Cochran (I98O) the number of sample frames used in this study was insufficient to detect changes of 25 percent in aerial biomass response of Russian thistle and other forbs. frames, With additional sample significant differences to P fertilization may have been detected. Biennial and Perennial Forbs Tables 11 apd 12 summarize the data collected for biennial and perennial forbs respectively. Phosphorus fertilization did not significantly affect the response of these plant classes for any of the data parameters measured (Tables 23, 24, and 25; Appendix B). Density, aerial biomass, and canopy cover of these forbs were relatively low on the study site. Biennial and perennial forbs occurred mainly on the eastern side of the study site. A grassland community established on previously mined land bordered the study site on the east side. Apparently these forbs invaded the study site from this revegetated area. 62 Table 11. Vegetatlonal statistics of biennial forbs by fertilization treatment, 1982. Fertilizer treatment NO NO N14 N14 N28 N28 N56 N56 NO NO N14 N14 N28 N28 N56 N56 NO NO N14 N14 N28 N28 N56 N56 P168 P168 P168 P168 P168 P168 P168 P168 Pl 12 Pl I2 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 PO PO PO PO PO PO PO PO KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 Density May plants/m2 Density July plants/.u2 0.0 0.0 1.1 0.0 0.0 0.6 0.0 0.6 0.0 0.6 0.0 0.0 0.6 0.6 0.6 0.0 1.1 0.0 0.6 0.0 0.6 0.6 0.0 1.1 3.0 0.4 1.5 0.7 0.7 1.1 1.9 2.2 1.1 1.1 0.4 2.6 1.9 1.5 1.9 0.0 0.4 2.2 1.5 0.0 3.0 2.6 1.9 2.2 Canopy cover % 0.7 0.2 1.9 0.0 0.3 1.7 1.4 0.4 0.7 0.6 0.1 0.9 0.6 0.3 1.1 0.0 1.9 1.3 2.0 0.1 2.5 0.8 1.4 2.5 Legumes Table 13 summarizes the data collected for legumes. Phosphorus fertilization significantly affected the aerial biomass and canopy cover response of this plant class (Tables 23 and 24, Appendix B). Aerial biomass decreased between the O and 112 kg P/ha rates, but increased between 112 and 168 kg P/ha (Figure 12). For canopy cover, legumes tended to decrease as P fertilization increased (Figure 13). Despite this linear trend, canopy cover was depressed at the 112 kg P/ha fertilization rate when compared to the O and 168 kg P/ha levels. 63 Table 12. Vegetational statistics of perennial forbs by fertilization treatment, 1982. Fertilizer treatment NO NO N14 NU N28 N28 N56 N56 NO NO NW NW N28 N28 N56 N56 NO NO NW NW N28 N28 N56 N56 P168 P168 P168 P168 P168 P168 P168 P168 Pl 12 Pl12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl12 PO PO PO PO PO PO PO PO Density May plants/m2 Density July plants/m2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 Canopy cover 56 0.0 0.0 0.0 0.0 o.o 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.1 0.1 0.0 0.0 0.0 0.0 0.0 Of the legumes on the study site, Cicer milkvetch (Astragulus cicer) made the largest contribution to density and canopy cover (Tables 30, 31» and 32; Appendix C). Phosphorus fertilization generally did not affect frequency of legumes (Table 8). 64 Table 13. Vegetational statistics of legumes by fertilization treatment, 1982. Fertilizer treatment NO NO NU NU N28 N28 N56 N56 NO NO NU NU N28 N28 N56 N56 NO NO NU NU N28 N28 N56 N56 P168 P168 P168 P168 P168 P168 P168 P168 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 PO PO PO PO PO PO PO PO KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 Density May plants/m2 Density July plants/m2 Aerial biomass kg/ha Canopy cover 55.6 45.6 53.3 44.4 46.7 45.0 45.0 55.6 37.2 43-3 32.8 45.0 44.4 41.7 41.1 33.3 52.2 48.3 80.6 32.2 31.1 51.7 56.1 46.7 59.6 37.0 45.9 54.1 38.5 48.9 40.0 54.4 39.3 60.4 31.1 46.3 43.0 45.2 37.0 44.8 55.9 49.6 82.6 36.7 35.9 53.7 42.2 67.8 53.2 60.4 36.4 19.6 58.8 54.8 49.6 49.6 52.0 26.4 14.4 30.0 35.2 26.8 32.8 23.2 27.6 39.2 51.2 33.2 59.2 76.4 56.0 66.4 7.4 4.7 4.9 3.7 3.2 8.8 4.6 5.7 4.6 3.7 5.4 5.5 4.5 3.8 5.0 3.9 6.6 4.1 10.0 5.5 7.3 8.2 6.8 10.4 % 65 AERIAL BIO M A SS (k g /h a ) Figure 12. Mean aerial biomass of legumes fertilization, July, 1982. Y — 0.0 0 3 X p < 0.05 I sq. = 2 - 0 .6 2 4 X in response f to P 5 1.16 0.33 60 40 20 t H-------------112 P H O S P H O R U S RATE ( k g / h a ) 168 66 13. Mean canopy cover of legumes fertilization, July, 1982. PERCENT C A NO PY COVER Figure Y = - 0 . 0 1 4 X +- 7 . 0 2 p < 0.06 r sq. a t in response to P 0.23 \--------------------- P H O S P H O R U S RATE ( k g / h a ) 1 67 Two factors may have affected the response of legumes to P fertilization. classes. The first factor was competition from other plant Phosphorus fertilization significantly increased the aerial biomass and canopy cover of other plant classes on this study. These plant classes may have outcompeted legumes for site resources as fertilization increased their aerial biomass and canopy cover. This competition may have resulted in the decrease in legumes at the 112 kg P/ha rate for aerial biomass and canopy cover. Studies at Colstrip indicated combinations of N and P fertilization of seeded species mixtures reduced canopy cover and aerial biomass of legumes as the result of competition from other plant classes (DePuit et al. 1978» Holechek 1976). Brown and Munsell (1956) reported P fertilization decreased legume composition in a legume/grass mixture by increasing the competitive ability of Kentucky bluegrass (Poa nratensish Decreases in individual plant growth, rather than a decrease in the number of plants accounted for the reduction in aerial biomass and canopy cover of legumes at 112 kg P/ha. Frequency and density of legumes did not change as P fertilization increased, indicating no effect on the number of individual plants. DePuit and Coenenberg (1979) noted fertilization with N and P decreased aerial biomass and canopy cover of legumes in a diverse species mixture by decreasing plant growth. The second factor a f fecting response of legumes fertilization may be the beneficial effects P has on legumes. to P Several studies noted P fertilization increased composition or yields of legumes in perennial grass mixtures (Cooper et al. 1969» Mott 19^3» 68 Synder et al. 1978). Aerial biomass and canopy cover of legumes may have been greater at 168 kg P/ha, than at 112 kg P/ha, because of the beneficial effect P has on legumes. Phosphorus at 168 kg P/ha stimulated growth of legumes as indicated by increased aerial biomass and canopy cover. This stimulation of legume growth at. 168 kg P/ha may have offset negative competitive effects from other plant classes found at 112 kg P/ha. DePuit and Coenenberg (1979) predicted the results found in this study. They noted that W with P fertilization decreased aerial biomass and canopy cover of legumes, but speculated that legumes would increase with higher levels of P. Shrubs Table 14 summarizes the data collected for shrubs. Phosphorus fertilization had no significant effects on the aerial biomass and canopy cover response of this plant class (Tables 23 and 24, Appendix B). Fourwing saltbrush was the only shrub on the study site, but was represented in all fertilization treatments. Literature indicated N and P fertilization increased yields of fourwing saltbrush (Aldon 1978, Aldon et al. 1976). The lack of response.of shrubs to P fertilization in this study could not be explained. Perhaps this lack of response is temporary and results from subsequent years sampling will differ. One growing season may. not be long enough for fourwing saltbush to respond to fertilization. 69 Table 14. Vegetational statistics of shrubs by fertilization treatment, 1982. Fertilizer treatment NO NO NI 4 N14 N28 N28 N56 N56 NO NO NI 4 NI 4 N28 N28 N56 N56 NO NO N14 N14 N28 N28 N56 N56 P168 P168 P168 P168 P168 P168 P168 P168 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 Pl 12 PO PO PO PO PO PO PO PO KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 Density May plants/m2 Density July plants/m2 2.8 0.6 2.8 1.7 2.8 2.8 2.2 3.3 1.1 1.7 0.6 2.8 2.2 3.9 0.0 1.1 3.3 2.8 1.7 0.6 3.9 2.8 1.1 1.1 1.5 1.1 4.4 3.7 1.9 3.3 1.9 1.1 2.2 1.1 0.4 2.6 2.2 3.7 1.1 1.1 3.0 1.5 1.5 1.1 1.1 1.5 1.9 1.1 Aerial biomass kg/ha 7.2 0.8 1.2 13.2 3.2 3.2 2.4 5.2 5.6 15.2 3.2 4.4 5.2 1.6 1.6 3.6 6.0 2.0 4.0 2.0 1.6 7.6 4.4 12.8 Canopy cover % 0.4 0.5 0.9 1.5 0.6 0.8 0.6 0.7 0.6 0.3 0.1 0.9 0.7 1.0 0.6 0.5 0.9 0.6 0.3 0.3 0.7 1.0 1.0 0.3 Total Vegetation Table 15 summarizes the data collected for total vegetation. Phosphorus fertilization significantly affected aerial biomass and canopy cover response of vegetation (Tables 23 and 24, Appendix B). Aerial biomass (Figure 14) and canopy cover (Figure 15) tended to increase with P fertilization. The results of this study for aerial biomass and canopy cover of total vegetation were expected. Several studies reported that P fertilization in the presence of adequate soil NOg-N increased aerial 70 biomass and canopy cover of total vegetation during the first growing season (Barnhisel and Evangelou 1981, DePuit et al. 1978, Farmer et al. 1974). Table 15. Vegetational statistics of total vegetation by fertilization treatment, 1982. Fertilizer treatment NO NO NI4 N14 N28 N28 N56 N56 NO NO N14 N14 N28 N28 N56 N56 NO NO NI4 NI4 N28 N28 N56 N56 P168 P168 P168 P168 P168 P168 P168 P168 Pl12 Pl 12 Pl12 Pl 12 Pl12 Pl 12 Pl12 Pl 12 PO PO PO PO PO PO PO PO KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 Density May plants/nr Density July PlantsZmei 189.4 180.0 181.1 167.2 170.0 157.2 165.0 177.2 172.2 154.4 144.4 163.3 175.6 180.6 151.1 145.6 166.1 181.1 221.1 149.4 129.4 158.9 181.7 221.1 223.0 164.1 166.7 210.4 185.6 178.5 145.2 186.3 176.3 167.8 170.0 177.4 185.9 234.8 158.1 190.4 198.5 201.9 263.0 180.7 158.5 191.5 175.9 243.3 Aerial biomass kg/ha Canopy cover 2226.8 2776.8 3060.0 2024.8 3424.0 2676.8 2948.8 2943.6 2916.4 2286.8 2446.8 2713.2 3574.8 2309.6 2442.0 2290.8 2016.4 2156.4 2216.8 1454.0 2325.6 1955.2 2725.6 2478.8 118.1 120.0 137.5 118.3 120.9 121.4 140.0 126.8 134.1 117.3 110.0 121.1 120.4 124.2 128.7 121.4 112.7 97.2 103.1 104.9 113.1 88.4 105.9 122.2 % Plant Communitv Develonment Table 16 lists the diversity, each fertilization treatment. evenness, and richness indices for Diversity ranged from .5056 to .8383, evenness from .3767 to .5925, and richness from 16 to 26. Despite large differences between maximum and minimum values of the indices, 71 AERIAL BIOMASS (kg/ha) Figure 14. Mean aerial biomass of total vegetation in response to P fertilization, July, 1982. 3400 - 3000 - Y = 3.6 I 3 X 4 - 2 1 7 9 P 0.01 < r sq. = 2600 - 2200 - 0.28 1800 4------ 1 0 112 PH O SPH O R U S RATE ( k g / h a ) 168 72 PERCENT CANOPY COVER Figure 15. Mean canopy cover of total vegetation in response to P fertilization, July, 1982. 0.12X + < r sq. 10#.# 0.01 o.so PHOSPHORUS RATE (kg /ha ) 73 fertilization did not affect diversity, evenness, or richness during the first growing season. Table 16. Diversity, evenness, and richness indices by fertilization treatment. Fertilizer treatment NO NO N14 N14 N28 N28 N56 N56 NO NO N14 N14 N28 N28 N56 N56 NO NO NU NU N28 N28 N56 N56 P168 P168 P168 P168 P168 P168 P168 P168 Pl12 Pl 12 Pl12 Pl 12 Pl12 Pl 12 Pl12 Pl 12 PO PO PO PO PO PO PO PO Diversity KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K28 KO K 28 KO K28 KO K28 KO K28 KO K28 Evenness Richness 26 19 26 24 20 26 23 26 26 22 19 24 25 24 25 20 22 26 20 16 16 21 22 23 .5925 .4705 .5305 .5264 .4908 .5485 .5136 .5553 .4915 .4641 .4791 .5687 .5601 .5327 .5063 .4949 .4771 .5075 .5241 .5500 .5764 .5146 .3767 .5776 .8383 .6016 .7506 .7266 .6386 .7761 .6993 .7857 .6955 .6230 .6127 .7849 .7830 .7353 .7077 .6439 .6404 .7181 .6818 .6623 .6940 .6804 .5056 .7866 If trends in vegetational responses observed during the first growing season due to P fertilization continue in subsequent growing seasons; diversity, fertilization. of plant and richness may be affected by P Figures 16 and 17 illustrate the percent composition classes fertilization evenness, in the on the community basis of significantly aerial biomass and affected canopy by P cover 74 Figure 16. Composition of plant community fertilization, aerial biomass. as affeced by P 100 CO CO < S O m 90 - OPC -j < OC LU < 80 - -J < IO lLL O LU O < X < < < < IZ LU O OC LU Q. 5 - ^WSGv LEG WSG, PHOSPHORUS RATE (kg/ha) eWSG = warm season grasses; LEG = legumes; AG = annual grasses; OPC** = other plant classes. eeOPC contains plant classes not significantly affected by P fertilization for aerial biomass response. 75 Figure 17• Composition of plant community fertilization, canopy cover. as affected by P CC LU > O O > CL O Z < O < HO HLL O UJ O < HZ LU O CL LU O- WSG PHOSPHORUS RATE (kg/ha) •WSG = warm season grasses; LEG = legumes; AG = annual grasses; AF = annual forbs; OPC** = other plant classes. eeOPC contains plant classes not significantly affected by P fertilization for canopy cover response. 76 respectively. Phosphorus fertilization decreased the percent composition of warm season grasses and to an extent legumes, while increasing annual grasses and annual forbs. The continued reduction of warm season grasses and legumes (at 112 kg P/ha) on P fertilized plots in subsequent growing seasons, could decrease plant community diversity when compared to non P fertilized plots. Only subsequent years' sampling will indicate if fertilization influences diversity and its components, evenness and richness. Residual Effects Table 17 lists the extractable P content of the study site soil. Phosphorus fertilization significantly affected the extractable P content of the soil (Table 26, Appendix B). increased with P fertilization (Figure 18). Extactable P content These results agreed with other studies (Lambert and Grant 1980, Young and Rennick 1982), Table 17. Mean soil extractable P content by P fertilizer rate (ppm), October, 1982 P level (kg/ha) 0-15 cm 0 2.2 112 168 17.2 27.4 77 Figure 18. Mean soil extractable P content in response to P fertilization, 0-15 cm, October, 1982. EXTRACTABLE PHOSPHORUS (ppm) Y = 0 . 1 4 8 X +■ 1 . 8 2 0.01 r sQ. — 0.87 30 - 10 - PHOSPHORUS RATE ( k g /h a ) 78 Vegetational responses to P fertilization should continue in subsequent growing seasons. Phosphorus fertilization greatly increased the extractable P content of the soil between October 1981 and 1982 (Tables 5 and 17). This P should be available for plant utilization in subsequent growing seasons. Tisdale and Nelson (1975) noted P applied by broadcast methods generally remains within a few centimeters of the soil surface. broadcasting. The current study applied P by The lack of m o b i l i t y of P should not prevent vegetational responses from occurring in subsequent growing seasons. Tisdale and Nelson (1975). stated that forage crops were able to utilize broadcast applied P by absorption through shallow roots. Nitrogen No significant differences in vegetational response due to N fertilization were found for density, aerial biomass, or canopy cover of the plant class tested (Tables 23, 24, and 25; Appendix B). These results contradicted other studies. McGinnies and Nicholas (1980) reported N fertilization increased total herbage an average of 89 percent on spoil material covered with various depths of topsoil. Several rangeland fertilization studies indicated various plant classes and species respond to N application (Cooper 1975, Goetz 1970, Power 1979, N a m e s and Newell 1969). Responses to P fertilization for aerial biomass and canopy cover indicated an adequate, supply of indigenous NOg-N in study site soils to meet plant needs during the first, growing season. Several studies have shown vegetation rarely responded to P fertilization unless At 79 adequate levels of N were present (Johnston et al. 1969, Lorenz and Rogler 1973, Wight and Black 1972), resu l t s similar vegetational to this study. response to N Power et al. (1974b) observed They concluded that lack of fertilization, and responses to P fertilization indicated indigenous soil NOg-N levels were adequate for plant needs. Compared with other studies at Colstrip, baseline soil analyses indicated high levels of indigenous N O g - N in the study site soil. Nitrate ranged from 7.4 to 8.2 ppm in the upper 30 cm of the soil profile, and 8.6 to 13.0 ppm between 30 and 60 cm (Table 5). Colstrip DePuit and Coenenberg (1979) found average At NOg-N in the top 30 cm of the soil profile to be 2.2 ppm, and 5.3 ppm between 30 and 60 cm in topsoiled spoils. Holechek (1976) found similar results at Colstrip in topsoiled spoils. Young and Rennick (1982) reported mean nitrate content ranged from 5.0 to 7,6 ppm for composites taken for several intervals between 0 and 15 cm of the soil surface on topsoiled, regraded mine soils. During October. 1981, percent organic matter content of the study site soil was generally less than that in native soils of the Colstrip area. Percent organic matter ranged from 0.8 to 0.9 percent over the study site in the upper 30 cm.of the soil profile (Table 5). Munshower et al. (1978) measured percent organic matter on eight grazing exclosures in the Colstrip vicinity at 10 cm increments from the soil surface to a depth of 40 cm. Organic matter hanged from 1.5 to 7.5 percent on the surface and from 0.8 depths to 3.1 for all other 80 The higher amounts of NOg-N and lower organic matter content in the study site soils may be due to cover-soil handling procedures. The cover-soil used on the study site was stockpiled in 1976, and applied to the site in October, 1981. During this time, organic matter may have undergone mineralization as indicated by high and low organic matter levels. Zap, NOg-N Data from the Indian Head Mine near North Dakota supported this conclusion (Argonne Nat. Lab. 1979). Nitrate levels in topsoil at the Indian Head Mine increased from 4 ppm after 10 months of storage to 12 ppm after 40 months. For the same time period organic N, a component of organic matter, dropped from slightly above 0.2 percent to slightly above 0.1 percent. Though NOg-N was adequate to meet plant needs during the first growing season, this may not be the case in subsequent years. uptake and leaching reduced growing season. Plant NOg-N levels in the soil during the first Nitrate levels in the upper 30 cm of the soil profile ranged from 3.9 to 6.8 ppm during October, 1982 (Table 18), compared to 7.4 to 8.2 ppm in October, 1981 (Table 5). Nitrogen fertilization significantly affected the soil NOg-N content in the upper 15 cm in October, 1982 (Table 27, Appendix B). Regression analyses evaluated the influence of N fertilization on soil NOg-N content in a manner similar to that described for vegetational data. Nitrate levels were highest on plots fertilized with 0 and 56 kg N/ha, but depressed at the two intermediate rates (Figure 19). Regression analyses conducted on soil NOg-N content of the upper 15 cm for Blocks I and 2 showed the same relationships between soil NOg-N and N fertilization, but with a higher correlation (Figure 20). Block 81 Figure 19. Mean soil NOg-N content in response to N fertilization, 0-15 cm, October, 1982. 0.003X < -0.137 X + 5.58 0.07 NITRATE (ppm) ®q. — 0 . 3 4 I---------------1-------------- \ NITROGEN RATE (kg/ha) I 82 3 soil NOg-N content data showed responses similar to Blocks I and 2, but with lower soil NOg=N contents (Figure 20). Regression analyses were not conducted on Block 3 data due to a lack of data points. The lower soil NOg-N content in Block 3 can be attributed to greater levels of aerial biomass and canopy cover of vegetation in this block when compared to Blocks I and 2. The increased growth in Block 3 may have been due to site characteristics and surface manipulation, rather than fertilization. The increased vegetational response in Block 3 appeared to lower soil NOg-N content by taking up greater quantities of NOg-N from the soil. Table 18. Mean soil NOu-N content by N fertilizer rate (ppm), October, 198? N level (ke/ha) 0 14 28 56 0-15 cm 15—RO cm 5.6 4.1 3.9 6.2 5.4 4.5 4.8 6.8 An interaction between levels of account for the depression of soil N/ha. As discussed, plant classes. N x P N O g - N content at the 14 and 28 kg P fertilization increased the growth of several This increased increased uptake of N and P fertilization may growth may have resulted NOg-N by plants from the soil. interaction and its effect on in an The existence of a NOg-N content of the, soil can not be determined without further analyses. 83 Figure 20. Soil NOg-N content in response to Blocks I and 2, October, 1982. KEY: , * N fertilization, N I T R A T E L EV EL S IN B L O C K S I AND 2 NITRATE LEVELS IN BLOCK 3 0.004X p < 0 .0 1 0.86 NITRATE (ppm) r eq. NITROGEN RATE (kg/ha) 84 The ability of the mine soil to replenish the soil MOg-N pool may be low= Organic matter content of the mine soil generally remained unchanged between October 1981 and 1982 (Tables 5 and 19), and was not .significantly affected by N fertilization (Table 28, Appendix B). As previously noted, organic matter content of the cover-soil on the study site was generally lower than that on rangeland soils of the Colstrip area. This.lower organic matter concentration indicated less material available for conversion to NOg-N via mineralization. As a result, N deficiencies may occur at the study site as the soil NOg-N pool becomes depleted. Plots in which soil NOg-N was lowered by a possible N x P interaction (as previously discussed), are likely to show signs of N deficiencies earlier than the other study plots. Table 19. Mean percent soil organic matter content by N fertilizer rate, October, 1982. N level (kg/ha) 0 14 28 56 0-15 cm 0.88 0.86 0.92 0.88 15-30 cm 0.88 0.80 0.92 0.90 The use of soil material as cover-soil after prolonged storage in stockpiles may have serious implications for the revegetation of mine lands. As indicated in this and the Indian Head Mine (Argonne Nat. Lab. 1979) studies, prolonged storage in stockpiles decreases the organic matter content of the cover-soil. With reduced organic matter content, the cover.soil may not be able to meet vegetational demands for N, and hence hinder plant community development and growth. 85 Though fertilization corrects nutrient deficiencies on a short term basis, a buildup of sufficient soil organic reserves is needed to maintain long term mine land productivity. Omodt et al. (1975) noted long periods of time and large amounts of nutrient imputs are needed to raise organic matter contents in mine soils. An alternative to the use of long term stored cover-soil is to direct haul or stockpile topsoil for minimal amounts of time. The organic matter in direct haul or short term stored topsoil does not tend to break down before application on a mine site. This material should contain higher levels of organic matter with a greater mineralization potential than long term stored cover-soil. It remains to be seen whether or not organic matter content of the cover-soil used on this study has been reduced to the point of significantly affecting mineralization potential. years' Only subsequent vegetation and soil sampling will provide information on the progress of nitrogen cycling establishment on the study site. Total N content of the study site soil in October 1982 is shown in Table 20. Nitrogen fertilization did not significantly affect total N content of the soils (Table 29» Appendix B). Table 21 lists the C/N ratios determined from total N and organic matter content. Nitrogen fertilization generally did not affect C/N ratios which were similar to levels prior to fertilization. 86 Table 20. Mean soil total N content by N fertilizer rate ($), October, 1982. M level (ka/ha) 0-15 cm 0 14 28 56 15-30 cm 0.05 0.04 0.04. 0.05 0.05 0.05 0.04 0.05 Table 21. Mean soil C/N ratio by.N fertilizer rate, October, 1982. 0-15 cm W level (ka/ha) 0 14 28 56 I5—30 cm 10.2 12.5 13.3 10.2 10.2 . 9.3 13.3 10.4 Potassium Potassium fertilization did not significantly affect response of the plant classes tested in this study for the data parameters analyzed (Tables 23, 24, and 25? Appendix B). The lack of vegetational response to K fertilization indicated sufficient quantities of indigenous soil K to meet plant needs over the study site during the first growing season. Other research supported this conclusion. Meyn et al. (1975) reported K adequate to low in overburden at Colstrip. In the Fort Union Formation only N and P are likely to be deficient for plant growth (Bauer et al. 1978a). Buchholz (1972) found indigenous soil K adequate for the growth of thickspike wheatgrass (Aeropyron dasvstachvum) on a site at Colstrip. X ' 87 Nutrient Interactions Statis t i c a l analyses interaction and N x K x indicated time interaction affected density response of annual forbs (Table 23» Appendix B). significant, a s ignificant N x P x K Since no main effects were the interactions were treated with caution... Fertilizer treatment N28 Pl 12 K28 had the highest mean density for annual forbs in July (Table 10). Mean density equaled 59.6 plants/m2 for this treatment, while it ranged from 25.2 to 41.9 plants/m2 for the other treatments. Russian thistle accounted for the increase in mean density of annual forbs in treatment N 2 8 P 1 1 2 K28 during July (Table 31» Appendix C). Experimental plots 15, 39, and 54 received the N28 Pl12 K28 fertilization treatment. Data indicated Plot 54 had higher densities for Russian thistle than the other two plots. located in a depression on the study site. areas drained and. temporarily precipitation events. ponded Plot 54 was Runoff from surrounding in Plot 54 during heavy The increased runoff into Plot 54 probably increased the soil water content of the plot, and allowed a larger number of Russian thistle plants to establish. Apparently, the soil moisture content of Plot 54, rather than fertilization resulted in the significant interactions. Significant interactions between N x P x K, and K x time existed for density data of legumes (Table 23, Appendix B). Fertilization treatment NI4 PO KO had the highest mean density during both May and July, (Table 13). and 82.6 Mean density oif treatment N14 PO KO equaled 80.6 plants/m2 for May and July respectively. The other treatments ranged in mean density from 31.1 to 56.1 plants/m2 in May, 88 and 31.1 to 67*8 plants/m2 in July. Experimental plots in which treatment NI4 PO KQ was applied, did not visibly differ from other plots on the study. No explanation existed for the significant interactions that occurred for legumes. These results for density data of legumes should be treated with caution, since no main effects proved significant. Adequacy of regression models The regression models significantly fitted for various plant classes on the basis of density, aerial biomass, canopy cover, and for post fertilization soil analyses are inadequate as predictive models (Figures 6 to 15, 18 to 20). Reasons for inadequacy in the regression models included lack of data points and high variability in the data. A large gap existed between the 0 and 112 kg P/ha fertilization rates. If P had been applied at rates between 0 and 112 kg P/ha, regression fit determination, may have been w h i c h . ranged better. from The 0.23 to the c o e f f icients 0.85* of indicated fertilization accounted for only a small to moderate amount of the variation in the data. Much of the unaccounted variation may be attributed to differences within blocks caused by differences in soil properties micro-climate, non-significant effects fertilization on vegetation, and lack of replications. of N and K In particular, the nature of the cover-soil may have caused within block variation. As discussed, pre-mine topsoil and subsoil were not thoroughly mixed during cover-soil handling operations. The pre-mine topsoil was scattered in pockets throughout the study site soil, and may have 89 differed considerably from the subsoil material in nutrient status. This difference in nutrient supplying ability may have caused differences in plant response within short distances on the study site. The small coefficients of determination pose no problems in interpreting results of this study. Statistically, the lack of fit test indicated the regression models’ appropriateness for defining the relationship between fertilization and vegetational response. The purpose of regression analyses in this study was to clarify trends caused by fertilization, rather than to create predictive models. Despite high variability unaccounted for by fertilization treatment, the regression models illustrated observable general trends that existed in the field. 90 RECOMMENDATIONS Concrete recommendations concerning fertilization could not be made on the basis of this study for several reasons. The data analyzed on this study constituted only one growing season. Data from subsequent growing seasons will be needed in order to effectively make recommendations. DePuit and Coenenberg (1979) noted considerable differences in vegetational response to fertilization between the first and subsequent growing seasons. In light of this information, vegetational response on the present study site could differ in following growing seasons. Montana State University's Reclamation Research Unit plans to continue monitoring this study in subsequent growing seasons. Another factor limited the ability to m ake concrete recommendations concerning fertilization of mine land at Colstrip. Phosphorus was applied by broadcast incorporated into the soil. methods rather Phosphorus is ain than being immobile element and / will not readily leach into the soil when applied on the surface. A leaching study found surface applied triple superphosphate (0-45-0) did not move below a depth of 10 cm, with the majority of extractable P located between 0 and 2.5 cm of the surface (Bjornson and Sims 1971). Incorporation mixes P fertilizer into the root zone, allowing greater potential contact of P with plant.roots. Vegetation utilizes incorporated P more effectively than broadcast applied P. crops, In forage twice as much P applied by broadcasting was needed to achieve the same vegetational responses from incorporation (Tisdale and Nelson 91 1975). The difference between broadcast and incorporation application methods of P could i n f luence fertilization recommendations. Incorporation of P is more economical than broadcast application due to a smaller amount of fertilizer material needed to achieve similar vegetational responses in the same growing season. information in the literature conc e r n i n g The lack of equilibration of P fertilization by broadcasting with incorporation methods, limited the ability to apply the' results of this study to native plant species on coal mine land. In order to determine the levels of P applied by incorporation for adequate establishment of native species on mine land, additional research is needed. Based on first year growing season data, indigenous NOg-N should not be fertilized with N. mine soils high in In this study, NOg- N content of the soil prior to fertilization was above 7.0 ppm for the upper 30 cm of the soil profile. Nitrogen fertilization in this study did not significantly affect vegetational response for the parameters measured. In contrast, DePuit and Coenenberg (1979) noted a one time application of 37 kg N/ha with 37 kg P/ha produced better vegetational establishment (of a native and introduced species mixture) than no fertilization on Colstrip mine soils averaging 2.2 ppm NO^-N. and Coenenberg’s results, study, interpreted in conjunction with the current imply that light applications of N may be beneficial when indigenous soil NOg-N levels are below 7 ppm. soil DePuit An exact indigenous N O g - N level at which N should be applied can not be determined. Variation in vegetational response to N fertilization caused by differences among sites for soil properties, microclimate, and 92 topography; and differences in yearly climatic conditions limits the ability to set a threshold level of soil NOg-N. Long term, careful observations over a number of years and variety of site conditions should provide the background for predicting soil NOg-N contents at which N fertilization is necessary. Based on first growing season data cover-soiled, regraded mine spoils at Colstrip should not be fertilized with P. Phosphorus fertilization adversely affected the establishment of the native plant community in this study by decreasing the density, aerial biomass, canopy cover, and frequency of warm season grasses. In addition P fertilization reduced aerial biomass and canopy cover of legumes at 112 kg P/ha. The revegetation goal at Colstrip is the establishment of diverse, predominantly native plant communities (Coenenberg 1982). Warm season grasses and legumes are important components of the plant community, and any management practice which reduces these two plant classes will decrease the diversity of the plant community. Even without P fertilization, it is unlikely the seed mixture used in this study will meet revegetation goals. Despite warm season grasses and canopy cover having their greatest aerial biomass and canopy cover composition at O kg P/ha, these plant classes represented only a small portion of the plant community (Figures 16 and 17). Regardless of fertilization treatment, only a small number of biennial forbs, perennial forbs, and shrubs established on the study site. The lack of warm season grasses, legumes, biennial forbs, perennial forbs, and shrubs in the community may reduce the diversity of the community below levels acceptable for revegetation goals. 93 Results of the present study may have application to a two phase seeding revegetation Colstrip. strategy currently used on mine soils at Two phase seeding involves initial seeding of hard to establish warm season grasses, forbs, legumes, and shrubs followed a year or two later with interseeding of cool season perennial grasses (Coenenberg 1982). Phosphorus fertilization may be inappropriate initially due to negetative effects on the establishment of warm season grasses and legumes. If data from subsequent growing seasons indicates P fertilization benefits plant community development and diversity, P application at time of cool season perennial grass interseeding may be more appropriate. Phosphorus and cool season perennial grasses could be placed simultaneously into the soil with a drill in alternating rows. Warm season grasses may also be benefited by this later fertilization. Rehm et al. (1972) noted H and P fertilization increased yields of warm season grasses when applied two years after establishment. They also found later fertilization did not adversely affect the botanical composition of warm season grasses in relation to cool season perennial grasses if warm season grasses initially dominated the community. In two phase seeding, warm season grasses should be one of the dominant plant classes after the first seeding phase. Based on first growing season data coversoiled, regraded mine spoils at Colstrip should not be fertilized with K. Potassium fertilization did not significantly affect vegetational response for any of the parameters measured. 94 SUMMARY This study tested the effects of M, P, and K fertilization on the initial establishment of a diverse native species seed mixture on coversoiled, regraded mine spoils at Colstrip, Montana, Experimental plots were established and seeded in the late fall of 1981, f ertilized in the spring of I982. rates of nitrogen (0, 14, 28, and Fertilization consisted of four and 56 kg N/ha), three rates of phosphorus (0, 112, and 168 kg P/ha), and two rates of potassium (0 and 28 kg K/ha) in complete factorial combination for a total of twenty-four treatment combinations. A randomized block design replicated the treatments three times. Density, aerial biomass, and canopy cover were estimated by plant class and species during the first growing season. Frequency of plant species, diversity, evenness, and richness were calculated from the canopy cover data for each fertilization treatment.. prior to fertilization, Baseline soil analyses were conducted arid selected soil parameters analyzed at the end of the first growing season. The density, aerial biomass, canopy cover, and post fertilization soil data were statistically analyzed. P hosphorus fertilization vegetational responses. created the only significant Lack of vegetational response to N and K fertilization was attributed to the presence of sufficient indigenous soil NOg-N and K to meet first year vegetation needs. Post fertilization soil analysis indicated that the level of soil NOg-N was affected by rate interactive effects from P. of N fertilization with possible Soil N 03 =N levels were reduced on some 95 treatments to levels at which signficant differences in vegetations! response to N fertilization may occur in subsequent growing seasons. Significant differences in vegetational response due to P can be expected to continue. Post fertilization soil analyses indicated P fertilization significantly increased level of extractable P in the soil. This increase in P is expected to be available to plants in subse q u e n t growing seasons. No s ignificant vegetational response due to K f e r t i l i z a t i o n differences should occur in in subsequent growing seasons based on results of other studies conducted at Colstrip. Except for warm season grasses, fertilization did not significantly affect density of the plant classes studied during the first growing season. Phosphorus fertilization reduced density of warm season grasses significantly possibly by increasing competition from other plant classes. Phosphorus fertilization significantly affected aerial biomass response of warm season grasses, annual grasses, legumes, and total vegetation. Phosphorus fertilization also affected Russian thistle response, but inadequate sampling prevented detection of significance. Phosphorus fertilization tended to increase the aerial biomass of annual grasses, decreasing warm Russian thistle, season grasses. and total vegetation, Fertilization at while 112 kg P/ha decreased aerial biomass of legumes when compared to the 0 and 168 kg P/ha rates. Canopy cover data showed the same results as aerial biomass except annual forbs were significantly increased by P fertilization. 96 Diversity, evenness, and richness appeared unaffected by fertilization treatment. In conclusion, the following recommendations were made oh the basis of the first g r o w i n g season v e g e t a t i o n data and post fertilization soil analyses. 1. Under the conditions of this study, N fertilization is not necessary for vegetations! establishment during the first growing season. Generally, mine soils with 7 ppm NOg-N or above need not be fertilized with N. 2. Phosphorus fertilization should not be used on a diverse seed mixture during the first growing season due to the adverse effects on warm season grass and to an extent legume establishment. 3. In a two phase seeding program, P fertilization may be more beneficial when applied one or two years after establishment of warm season grasses. Phosphorus could be applied simultaneously with cool season perennial grasses at time of interseeding. 4. Potassium fertilization is not necessary for the establishment of vegetation on mine soils at Colstrip. APPENDICES 98 APPENDIX A SOIL PROFILE DESCRIPTION I I I 99 Table 22. Soil profile description of the study site. Soil Series: Cow Creek8 Classification: Typic Ustorthents, fine-loamy88, mixed (calcareous), mesic. Location: Rosebud County, MT. Colstrip. Western Energy Co. Rosebud Mine, Area A, center of the NW1/4, SW1/4, S.33, T.2N. R.HIE. Physiographic position: upland. Topography: convex simple slopes, north aspect, 0-3%. Drainage: moderately well drained, moderate permeability. Erosion: uneroded. Vegetation: Agroovron son.. Salsola kali, annual forbs, Bromus son., and Atriolex canescens. Parent material: mine spoil and salvaged topsoil/subsoil. Sampled bvs P. J. Hertzog, May 21, 1982. Remarks: Mine spoils were covered with material during Fall 1981. This material had been stockpiled in 1976 and consisted of both topsoil and subsoil. Colors are for moist soil unless otherwise indipated. Al 0-93 cm (0-37 in.). Brown (IOYR 5/3) loam, light brown gray (2.5Y 6/2) dry; 30% IOYR 5/3 siltstone fragments possibly from former Bt horizon; massive; slightly hard (dry), friable (moist), slightly sticky and slightly plastic (wet); strongly effervescent; with clear irregular contrasting pockets of. former Al material comprising 18% of the horizon, very dark gray brown (IOYR 3/2) loam, dark gray brown to gray brown (2.5Y 4.5/2) dry; massive; hard (dry), friable (moist), slightly sticky and slightly plastic (wet); no effervescence; abrupt smooth boundary. Cl 93-1IO+ cm (37-43+ in.). Gray brown (2.5Y 5/2) loam, light gray brown to light gray (2.5Y 6.5/2) dry; 25% 2.5Y 7.5/2 soft siltstone fragments with 7.5 YR 5/6 coating covering 0-50%. of the fragment, 7% 7.5 YR 3/0 carbon fragments, 4% 7.5 YR 6/6 hard shale fragments, 1% 2.5 YR 6/4 soft sandstone fragments; massive; soft (dry), very friable (moist), slightly sticky and slightly plastic (wet); strongly effervescent. 8Based on unofficial soil series from Schafer (1979). 89Siltstone fragments disregarded in soil family textural determination, see Schafer (1979). APPENDIX B ANALYSIS OF VARIANCE TABLES Table 23. Analysis of variance for density by plant class, May and July 1982 Source Blocks Nitrogen Phosphorus Potassium NxP NxK PxK N x P x K Error (1)a df 2 I 2 I 6 3 2 6 46 CSPG" 9.28 1.78 25.09 76.27 160.68 118.20 5.96 65.28 74.96 WSG' 16.03* 5.48 33.84** 0.05 7.52 7.81 1.25 3.39 3.32 AG 21.12** 0.54 2.02 1.41 0.95 3.15 0.84 1.09 1.39 Time N x T PxT K x T N x P x T N x K x T P x K x T N x P x K x Error(2)b I 3 2 I 6 3 2 T 6 48 154.94* 14.03 0.04 2.45 12.73 8.35 10.28 10.66 24.77 434.52** 3.65 22.56** 0.03 5.01 5.21 0.83 2.26 2.57 2.01 0.47 0.05 0.71 0.92 0.05 0.51 0.67 0.85 "CSPG = cool season grasses; PF = perennial forbs; LEG = grasses, July 1981. •Includes Setaria viridis. a •Significant at p<0.05 ••Significant at p<0.01 ^Derived from pooling sum of bDerived from pooling sum of Mean Square AF BF 50.21* 1.19" 0.29 0.13 0.11 0.19 5.21 0.05 21.81 0.15 5.14 0.01 23.41 0.05 29.29 0.33 11.81* 0.27 344.45** 6.95 5.18 12.10 3.34 12.16* 1.09 7.14 3.94 3.34** 0.03 0.01 0.01 0.11 0.05 0.05 0.18 0.13 PF 0.001 0.001 0.001 0.0 0.002 0.002 0.003 0.002 0.002 LEG 59.23 15.80 85.56 2.05 17.50 54.22 35.22 94.54* 38.58 SHRB 0.06 0.61 0.43 0.004 0.50 0.34 0.65 0.11 0.48 TOTVEG 43.91 26.97 289.68 83.33 335.09 393.12 27.09 328.39 246.79 0.002 0.001 0.001 0.0 0.001 0.002 0.002 0.001 0.001 6.55 0.94 6.52 42.37* 2.07 11.33 1.74 9.39 7.38 0.07 0.23 0.11 0.0 0.22 0.15 0.07 0.06 0.13 882.97** 48.69 62.57 86.81 15.45 72.37 19.84 47.98 70.18 WSG? 26.71 9.13 56.40** 0.08 12.53 13.02 2.08 5.65 5.53 WSG = warm season grasses; AG = annual grasses; BF = biennialforbs; legumes; SHRB = shrubs; TOTVEG = total vegetation; WSG2 = warm season warm season annual. squares for all interactions with blocks, but without time. squares for all interactions with blocks and time. Table 24. Analysis of variance for aerial biomass by plant class/species, July 1982 Mean Square Source df CSPG " Blocks 2 1,814,890** Nitrogen 3 3,492 Phosphorus 2 142,653 Potassium I NXP WSG' AG 476.22** 42,336 36.00 12,077 SAKA 96,178,156** OF 836,762** 1,858,459 48,076 4,278,319 209,168 LEG SHRB 2,068.06 552.17 5,669.11 73.98 12,311.76* 8.00 T0TVEG 138,024,304** 2,543,431 556.22** 155,782** 6,651 3.56 1,800 11,183,535 205,012 200.03 329.39 9,056,984 6 66,009 35.78 30,265 1,290,269 76,187 3,038.12 231.70 1,567,070 NXK 3 17,767 4.89 29,276 3,246,183 96,910 229.49 93.69 2,437,766 PXK 2 17,102 4.22 568 63,251 1,824 938.09 1.56 146,050 NXPXK 6 38,342 20.67 33,327 2,971,942 196,715 1,826.96 323.19 2,044,146 Error 46 53,621 36.69 26,484 4,347,685 130,083 3,754.71 190.83 2,371,979 9,283,989* 102 "CSPG - cool season perennial grasses; WSG * warm season grasses; AG = annual grasses; SAKA = Salsola kal i ; OF = other forbs; LEG = legumes; SHRB = shrubs; TOTVEG = total vegetation. 'Includes Setaria vlridis. a warm season annual. **Signifleant at p <0.01. * Significant at p <0.05. Table 25. Analysis of variance for canopy cover by plant class, July 1982 Source Blocks df 2 CSPG" 5,499.34«e WSG • 55.33** AG 1,159.74 Nitrogen 3 211.02 Phosphorus 2 1,937.93 Potassium I 311.11 1.89 0.30 NxP 6 985.75 2.96 NxK 3 589.99 PxK 2 N x P x K Error 3.46 222.21 Mean Square AF BF 58,628.78*» 46.69 2,687.75 2.13 PF 0.03 0.0 59.88 0.01 5,796.06 40.50 0.0 436.88 2,508.03 4.34 1.22 632.75 1,211.68 23.87 1.34 26.86 6 346.79 0.29 46 659.96 3.59 26.54” 1,446.53* 10,777.35#e LEG 259.56 SHRB 2.11 TOTVEG 58,718.18»* 24.20 2.19 2,922.49 451.93* 1.13 23,364.76** 5.93 0.89 4,293.56 0.03 66.49 3.67 1,084.79 5.27 0.03 170.13 4.42 693.08 716.42 4.46 0.02 32.20 2.54 377.38 309.73 3,043.48 26.91 0.03 86.71 0.61 3,675.06 374.68 2,041.23 24.46 0.03 135.72 2.45 1,899.26 "CSPG = cool season grasses; WSG = warm season grasses; AG = annual grasses; BF = biennial forbs; PF = perennial forbs; LEG = legumes; SHRB = shrubs; TOTVEG = total vegetation. 'Includes Setaria vlridis. a warm season annual. •Significant at p<0.01 ••Significant at p<0.05 104 Table Table 26. Analysis of variance for extractable P content October, 1982. Source df Mean square 0-1*5 cm Blocks Ext. P Error 2 2 6 482.200 67.23 27. Analysis of variance for NOq-N content of of soils, soils, October, 1982 Mean sauare Source df Blocks N03„n Error 2 3 6 0-1*5 15-30 cm am 3.283 1.956 3.927° 0.745 ^Significant at p<0.05. Table 28. Analysis of variance for organic matter content October, 1982. of soils, Mean square Source df 0-15 cm 15-30 cm Blocks NOg-N Error 2 3 6 0.0019 0.0013 0.0088 0.0057 105 Table 29. Analysis of variance for total N content of soils, October, 1982. Mean sauare Source Blocks NOg-M Error ‘ df 0-15 cm 15-30 cm 2 3 6 0.00003 0.00003 0.00003 0.00006 106 APPENDIX C VEGETATIONAL DATA BY PLANT SPECIES Table 30. Mean plant species density (plants/m2) by fertilization treatment, May 1982. Cool season perennial grasses: Ware season grasses:* Annual grasses: Annual forbe: Polygonum convolvulus Salsola kali succulent forb** other forbs Biennial forbs: Tragopogon dublus Perennial forbs:* Legumes: Aatragalua clcer Melllotus officinalis Atrlplex canescens S S 5 $ I I S I S $ S S I 100.0 105.6 S a E I I 5 a S I S 2 £ S S' 5 2 I E 80.0 81.6 95.6 103.9 103.3 77.2 £ 95.0 95.5 83.9 69.5 S E S 90.0 E S 2 S I 98.4 106.7 2 S a 2 E £ £ S £ $ 89.4 81.7 94.4 115.5 g S £ g E £ S £ Z S 88.9 63.3 E £ I S E E £ £ £ 90.6 100.6 138.3 4.4 1.1 2.2 2.8 0.0 2.8 2.2 1.1 3.3 0.0 4.4 2.8 0.6 2.8 1.1 5.0 4.4 1.7 1.1 0.6 5.6 3.9 2.8 0.6 2.8 1.7 0.6 0.6 1.1 1.7 2.2 0.6 3.3 2.8 1.1 0.0 0.6 2.2 1.1 2.2 0.6 1.1 0.6 2.2 0.6 5.6 0.0 0,6 3.3 0.6 0.0 2.2 0.0 2.8 15.0 0.0 1.1 0.0 0.0 0.0 0.0 1.1 0.0 1.1 21.1 0.0 0.0 6.7 1.1 0.0 0.0 0.6 0.0 1.7 20.0 0.6 0.6 2.2 1.7 0.0 0.6 0.6 0.0 0.6 12.2 2.2 1.1 0.0 1.1 0.0 1.1 0.0 0.6 0.0 17.2 0.0 0.6 2.2 0.0 0.0 0.0 0.0 0.0 0.6 17.2 0.0 0.6 0.6 2.2 0.0 1.1 1.1 0.6 1.1 38.3 0.0 0.0 1.7 0.6 0.0 0.6 0.0 0.0 1.1 11.1 0.0 0.6 0.0 0.0 0.0 0.6 0.6 0.0 2.2 13.9 0.0 0.6 0.6 0.0 0.0 0.0 1.1 0.0 3.9 20.0 0.0 0.6 0.6 1.1 0.0 0.0 0.0 0.0 1.1 20.6 0.0 0.0 0.0 0.6 0.0 0.0 1.1 0.0 0.6 11.1 0.0 0.0 1.1 0.6 0.0 0.0 0.0 0.0 1.7 17.8 0.0 0.0 0.0 0.6 0.0 0.0 0.6 0.0 2.8 1.1 1.7 0.0 0.0 0.0 0.0 1.1 25.0 0.0 1.7 1.1 1.7 0.0 0.0 0.0 0.0 0.6 14.4 0.0 0.0 1.1 0.6 0.0 0.0 0.0 0.0 0.6 21.7 0.0 0.0 0.6 1.7 0.0 0.0 0.0 0.0 3.3 23.9 0.0 1.1 1.7 1.1 0.0 0.0 1.1 0.0 1.7 17.2 0.0 0.0 0.0 0.0 0.0 0.6 0.0 0.0 1.7 22.8 0.0 0.0 0.0 1.1 0.0 0.0 0.0 0.0 1.1 25.6 0.0 0.0 0.0 0.6 0.0 0.0 0.0 0.6 0.0 10.6 0.0 0.0 0.6 1.7 0.0 0.0 1.1 0.0 0.6 17.2 0.0 0.0 0.6 3.9 0.0 0.0 1.7 0.0 0.6 21.1 0.0 0.6 0.0 0.0 1.1 0.0 0.0 0.6 0.0 0.6 0.0 0.6 0.0 0.0 0.6 0.6 0.6 0.0 1.1 0.0 0.6 0.0 0.6 0.6 0.0 1.1 50.6 5.0 45.0 0.6 52.2 1.1 42.8 1.7 44.4 2.2 41.7 3.3 42.8 2.2 51.1 4.4 36.1 1.1 43.3 0.0 32.2 0.6 39.4 5.6 42.8 1.7 40.0 1.7 37.8 3.3 31.7 1.7 50.6 1.7 46.7 1.7 78.9 1.7 30.0 2.2 28.9 2.2 50.6 1.1 55.6 0.6 45.0 1.7 2.8 0.6 2.8 1.7 2.8 2.8 2.2 3.3 1.1 1.7 0.6 2.8 2.2 3.9 0.0 1.1 3.3 2.8 1.7 0.6 3.9 2.8 1.1 1.1 2.8 * Mo emergence observed for these plant classes. ** Unidentified forb, succulent in appearence Jl 0.6 Table 31. Mean plant species density (plants/m^) by fertilization treatment, July 1982. 3 5 Cool season perennial grasses: Agropyroo dasystachyue A. smlthli 2 E 3 ! E s 5 3 3 £ S S EsE £ 2 £ £ £ 68. I 61.9 55.9 77.8 69.6 54.8 41.5 62.6 63.3 49.6 47.0 51.9 59.3 81.9 46.3 61.1 67.8 69.6 91.9 69.3 26.7 22.2 17.8 15.9 21.1 25.9 11.9 14.8 23.0 13.3 22.2 24.1 29.6 24.1 22.2 26.7 7.8 10.4 11.5 14.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 3.3 0.7 0.0 2.2 1.1 2.2 0.7 1.1 0.4 0.4 4.4 1.5 1.5 0.7 1.9 4.8 4.8 0.7 1.5 2.6 0.0 0.0 0.0 0.0 0.0 0.7 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.7 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 2.2 0.7 1.1 2.2 2.2 1.1 1.9 1.1 1.1 1.1 2.2 1.9 0.0 1.1 5.2 1.9 0.7 1.1 1.1 0.4 11.9 14.1 5.2 10.4 13.3 11.1 1.5 5.6 5.9 3.7 23.7 8.5 8.5 11.1 8.9 17.4 22.2 19.3 29.3 18.9 A. trachycaulue A. trichopliorum Broisue lneneis Hordeua lubatua Stlpa vlridula Wara season grasses:* Annual grasses: Aveaa fatua Bromus laponlcus B- tacCorua Annual forbs: Amaranchus sp. Casielina alcrocarpa Chenopodlua album C. Ieptophyllua Descurainia plnnata D. sophia Elllsia nyctclea Bellanthua annuus H. petlolaris Kochla scoparla Lappula redovskiI Polygonum aviculare P. convolvulus P raaofllssiaua Saleola kail Slsyabrlua altisslaum Solanum trIflorum Vaccarla segetaJ-ls Xanthlum Btruiaarlum other annual forbs Biennial forbs: Lactuca aerrlola Tragopogon dublua Perennial forbs: Ambrosia psllostachya Legumes: Astragalus clcer A. sp. Melilotus officinalis Petslosteum purpureum Atrlplex caccKcens 1.5 * Include* Setarle vlrldia. a wan 3 0.0 5.2 0.4 0.0 0.7 0.0 0.0 1.1 1.5 0.0 4.8 0.4 0.0 1.1 1.1 0.0 0-4 0.0 0.0 1.5 0.7 0.0 4.1 2.2 0.0 3.0 1.9 0.0 1.5 0.0 0.0 0.4 0.0 0.0 5.9 0.7 0.0 4.1 1.5 0.4 5.2 0.0 0.4 1.9 0.0 0.0 3.0 0.7 0.0 1.1 0.0 0.4 5.2 0.4 0.0 0.4 0.0 0.0 1.1 0.0 0.4 0.7 0.7 2.2 0.7 0.7 1.1 1.9 0.7 0.4 0.0 1.5 0.7 1.5 2.6 0.0 1.1 3.3 1.1 0.4 0.0 0.7 0.4 1.1 0.0 0.4 0.4 1.5 0.4 1.1 0.0 0.0 0.0 0.0 0.4 0.4 0.0 0.4 0.0 0.0 1.5 0.4 1.1 1.5 0.0 1.1 0.0 1.9 0.0 0.7 0.4 0.7 0.4 0.4 1.1 0.4 0.0 1.1 0.7 0.0 0.7 0.0 0.0 0.7 0.4 0.0 0.4 0.0 0.4 0.4 0.4 0.0 0.0 0.4 0.4 0.0 0.0 1.5 0.4 0.4 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.7 0.4 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.4 0.0 0.0 0.4 0.4 0.4 0.0 0.0 0.0 1.1 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.7 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 5.2 2.2 3.3 3.7 3.3 4.1 4.8 4.4 4.1 4.1 1.9 3.3 3.0 5.2 5.2 2.6 2.6 7.8 6.3 1.1 0.4 0.0 0.4 0.7 0.7 0.4 0.4 0.0 0.4 0.0 0.4 0.0 1.1 1.9 0.0 0.0 0.0 0.0 0.4 0.0 1.9 0.0 1.1 0.4 0.4 0.0 0.4 1.1 0.7 0.0 0.0 0.0 0.4 1.1 1.1 1.1 0.4 0.4 0.7 0.0 28.5 20.7 24.1 25.6 28.9 21.1 33.3 24.4 27.0 28.9 34.8 23.7 27.0 46.7 19.3 23.0 29.6 22.6 31.5 33.3 0.4 0.0 0.0 0.0 0.0 0.0 0.7 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.4 0.4 1.1 0.4 0.4 0.0 0.4 0.7 0.0 0.0 1.1 0.7 1.9 1.5 1.1 0.4 2.6 0.7 0.7 0.4 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 £ £ 57.0 71.1 83.0 90.7 10.4 11.1 7.0 14.1 0.0 0.0 0.0 0.0 1.9 2.6 0.4 3.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 1.1 0.7 1.1 1.5 13.7 18.9 10.0 17.8 0.4 1.1 0.0 0.0 1.9 0.0 0.0 1.1 0.4 0.0 5.2 0.0 0.0 0.7 1.1 0.7 0.0 0.0 0.4 0.4 0.0 0.4 1.1 0.4 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 4.1 2.6 1.9 4.1 0.4 0.4 0.0 0.4 1.1 0.7 0.7 1.5 27.4 22.6 20.7 31.1 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 2.2 0.7 0.4 0.0 0.7 0.7 0.7 0.0 0.4 0.4 1.1 0.0 0.7 1.1 0.7 1.5 0.7 0.4 0.4 0.7 0.0 0.4 1.5 1.1 0.7 1.1 0.7 0.7 1.1 0.7 0.0 0.0 0.0 0.4 2.2 0.0 0.7 0.7 0.0 0.0 2.2 0.7 2.2 0.4 1.5 0.4 1.1 1.1 0.4 0.0 0.0 0.0 . 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 48.5 0.0 6.7 4.4 0.0 1.1 3.7 0.0 2.6 3.3 0.0 0.7 4.1 0.0 1.9 3.3 0.0 2.2 4.8 0.4 3.7 1.5 0.0 4.8 3.0 0.0 2.6 3.7 0.0 1.5 6.3 0.0 1.9 4.1 0.4 6.3 4.4 0.0 1.9 1.5 0.0 1.9 4.1 0.0 3.0 3.7 0.0 0.0 1.9 1.1 3.7 11.5 0.0 0.0 1.9 1.1 4.1 11.1 0.0 1.1 5.2 0.0 1.5 3.0 0.0 1.9 6.7 0.0 1.5 4.8 0.0 3.0 5.2 1.1 4.4 3.7 1.9 3.3 1.9 1.1 2.2 1.1 0.4 2.6 2.2 3.7 1.1 1.1 1.5 1.1 1.1 1.5 1.9 1.1 3.0 1.5 Table 32. Mean canopy cover of plant species by fertilization treatment, July 1982 Plant class/specIes Cool season perennial grasses: Agropyron dasystachyua A. ealthll Stipa virldula Warm season grasses:* Annual grasses: B. tectorus Cbenopodium album C. Ieptophyllua Deacuralttia plimata D. sophla Ulisla nyctelea Hellanthuc eraiuus H. patlolarla Kochla scoparla Lappula redovskl Polygonua aviculare P. convolvulus P. raaoslselaua Salsola kail Slsyabrlus alliesImum Solanuo trIflorxua Vaccarla segetalls Xanthlua Btrusariun Biennial forbs: Lactuca eerrlola Tragopogon dublus Perennial forba: Ambrosia patiostachya Taraxacum officinale Legumes: Petalosteus purpureua Atriplex canescens S S S $ S S S 5 S 2 S a I I I I S i I i S IS.2 12.8 12.2 13.7 13.3 11.8 0.0 0.0 0.7 1.1 0.0 0.0 0.0 0.0 0.6 0.0 0.6 0.3 1.1 1.0 0.0 6.9 1.5 O.Q 1.7 0.2 5.3 0.0 0.9 0.0 0.1 1.0 0.5 0.6 3.5 0.0 1.1 0.0 0.0 0.1 1.0 1.0 0.3 2.2 5.0 0.7 9.0 0.4 S g 11.3 15.0 11.0 12.2 14.4 5.3 11.8 3.9 6.8 7.7 0.0 0.0 0.0 0.0 0.0 0.8 0.2 0.6 1.1 0.2 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.6 0.3 0.8 0.3 0.7 0.6 1.2 0.0 0.5 0.7 0.0 5.5 0.8 0.0 6.0 0.1 1.4 2.1 7.5 10.9 2.7 2.6 0.1 9.5 1.7 a $ S a S g 3 I i 2 S 9.9 12.0 15.2 13.4 4.6 5.9 9.6 10.1 0.0 0.0 0.3 0.0 1.3 1.5 1.6 1.3 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.8 0.5 0.0 0.6 1.2 1.2 0.7 0.0 5.0 0.0 0.1 0.7 0.0 a i S 3 S a 1.5 6.1 0.1 S S S £ £ £ £ S a i £ $ £ S S 8.3 4.0 0.0 0.6 0.0 0.0 0.1 0.2 1.4 7.9 3.7 0.0 0.4 0.0 0.0 0.0 0.3 1.3 9.0 12.6 11.9 11.6 13.2 14.3 5.6 6.1 5.6 4.3 4.7 7.7 0.0 0.0 0.0 0.0 0.0 0.0 0.4 1.2 1.9 0.4 0.5 0.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.1 2.0 0.2 0.3 0.5 1.9 1.8 1.1 1.4 1.3 1.8 0.1 2.8 2.0 0.3 3.3 0.6 0.1 3.2 0.0 16.4 8.6 11.6 9.4 10.3 8.9 0.0 0.0 0.0 1.3 0.6 2.7 0.0 0.0 0.1 0.0 0.0 0.0 0.1 0.0 0.0 0.6 1.5 0.5 0.7 0.8 0.8 1.3 0.0 0.0 6.9 10.6 10.1 3.2 2.7 0.0 a 3 0.0 5.1 0.0 S I 0.0 1.9 0.0 1.7 1.1 0.0 S 0.0 2.9 0.1 0.2 0.1 0.3 0.3 0.2 0.2 0.0 0.5 0.2 0.0 0.0 0.4 0.2 0.3 0.6 0.0 0.3 0.6 0.4 0.0 0.0 0.3 0.2 0.1 0.3 1.0 0.0 1.0 1.9 0.9 0.6 0.7 0.0 0.1 0.1 0.1 0.1 0.1 2.0 0.3 0.0 0.0 0.0 0.0 0.6 0.1 1.5 1.4 1.7 1.0 0.7 1.8 0.2 2.2 0.6 1.5 2.2 1.0 0.9 0.1 0.1 0.5 0.4 2.2 0.0 0.1 0.4 0.1 0.0 0.2 0.1 0.1 0.1 0.2 0.2 0.1 0.3 0.1 0.1 0.1 0.2 0.1 0.0 0.2 0.0 0.6 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.6 0.0 0.0 0.0 2.1 0.0 0.1 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.2 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.2 0.1 0.1 0.0 0.1 0.2 0.1 0.1 0.0 0.0 0.0 0.4 0.1 0.0 0.0 0.1 0.0 0.0 0.0 0.1 0.0 0.3 0.0 0.1 0.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.9 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 11.0 12.3 17.2 10.5 16.3 10.7 10.6 10.9 14.8 11.9 10.6 8.2 10.1 10.5 11.6 8.5 8.6 17.5 8.5 13.0 10.4 5.7 0.1 0.0 0.2 0.4 0.2 0.4 0.9 0.0 0.2 0.1 1.2 0.0 0.7 0.4 0.7 0.0 0.1 0.1 0.0 0.0 0.0 0.1 2.0 0.3 4.7 0.1 0.9 0.1 0.0 2.2 0.7 0.8 0.0 0.3 0 6 0.7 0.6 2.6 0.1 0.1 0.2 0.0 0.6 0.2 SI. 3 72.8 76.5 66.8 72.8 61.3 86.4 66.5 75.6 74.7 69.3 63.2 60.6 65.6 76.7 74.7 72.5 53.2 60.6 59.6 66.1 50.9 0.6 0.0 0.0 0.0 0.0 0.0 2.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.2 0.2 0.0 0.1 0.0 0.1 0.1 0.1 0.0 0.0 0.2 0.3 0.3 0.3 0.1 0.6 0.0 0.0 0.0 0.0 0.3 0.0 0.0 1.8 0.0 0.3 0.0 0.0 0.0 0.3 0.0 0.0 0.0 1.7 0.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.6 2.2 0.4 a E S 0.0 7.7 0.3 0.2 0.3 0.0 1.5 0.5 0.1 0.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.2 0.0 0.0 0.0 0.0 0.0 0.0 8.3 9.0 0.0 0.2 0.4 2.4 63.8 61.9 0.0 0.0 0.1 0.2 0.0 0.0 0.0 0.0 0.6 0.2 0.2 0.0 0.9 1.0 0.0 0.0 0.2 0.1 0.4 1.3 1.0 0.4 0.1 0.3 0.2 0.6 0.2 0.4 0.0 0.1 0.3 0.7 0.3 0.3 0.2 0.1 0.5 0.6 0.0 0.0 0.0 1.8 0.4 0.9 0.6 1.4 0.1 0.0 1.6 0.9 0.6 0.3 0.7 0.7 0.2 2.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.1 . 0.0 0.0 0.0 0.0 0.0 O-O 0.0 0.0 0O-0O 2.2 0.0 0.6 0.4 3.2 0.0 4.6 0.9 3.0 0.1 1.4 0.2 4.1 0.0 1.4 0.3 3.3 0.0 0.9 0.4 2.8 0.0 0.3 0.6 2.9 0.0 1.9 0.6 3.2 0.0 1.7 0.6 3.4 0.0 0.7 0.4 2.5 0.0 0.7 0.6 2.3 0.0 2.3 0.4 3.2 0.0 0.5 0.2 5.6 0.0 0.3 0.6 3.0 0.0 0.6 0.5 8.3 0.0 0.7 0.9 4.0 0.0 1.2 5.5 6.9 4.9 9.1 1.0 0.6 0.5 0.9 0.6 0.3 0.3 0.7 4.0 0.0 2.9 0.6 0.4 4.1 0.0 0.5 0.2 0.5 4.1 0.0 0.4 0.5 0.9 2.6 0.0 0.5 0.6 VS 0.6 0.8 0.6 0.6 0.6 0.3 0.1 0.9 0.6 1.1 0.7 1.0 1.0 0.3 * Includes Setaria Tlrldis. a warm season annual. I I Table 33. Percent frequency of plant species by fertilization treatment, July 1982 S I Plant clase/specles Cool season perennial trachycaulua A. trlchopborue Broaus lnerals Hordeun Jubatua Stlpa vlrldula W a n e season grasses:* Annual Eatua Brotsos Iepoolcus 8 S 8 2 I I z I a I a 2 2 8 z S ; 5 96.3 81.5 3.7 14.8 92.6 100.0 92.6 85.2 25.9 18.5 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 s 8 8 3 S I I S 96.3 85.2 96.3 77.8 85.2 74.1 96.3 66.7 88.9 85.2 81.5 92.6 100.0 100.0 55.6 70.4 70.4 88.9 55.6 92.6 74.1 0.0 0.0 0.0 0.0 0.0 11.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 14.8 18.5 14.8 18.5 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 3.7 7.4 7.4 7.4 37.0 3.7 40.7 11.1 0.0 0.0 U.l 3.7 29.6 18.5 44.4 U . l U.l 25.9 3.7 25.9 40.7 81.5 7.4 3.7 14.8 0.0 0.0 U.l 22.2 44.4 0.0 11.1 0.0 0.0 0.0 18.5 0.0 22.2 0.0 3.7 0.0 11.1 0.00 18.5 E 5 5 I I E I E 8 Asaramhus sp. Caiaellna a l c r o c a r p a C h e n o p o d Iua al b u m Ieptophyllua Deocuralnla plnnaca 7.4 18.5 0.0 0.0 0.0 U.l 22.2 0.0 0.0 0.0 7.4 22.2 14.8 3.7 0.0 0.0 14.8 48.2 18.5 40.7 3.7 U.l 8 0.0 22.2 0.0 0.0 0.0 0.0 29.6 0.0 8 a « annuus petlolarls Kochla ecoparla L a p p u l a redovtfkl Polygomus avlculare P. convolvulus P. raaoslsslaua S a l sola kail Sisyabrlua a l l lsolsum Solanua Vaccarla Xanthlua Biennial trlflorua segetalis atrumarlua Eorbs Lsctuca : serrlola T r a g o p o g o n d u b Ius P e r e n n i a l Eor b * : Aabrosla pellostachya Taraxacum officinale Astragalus clear Melllotue officinalis Petaloeteum purpureua Atriplex canescene * Includes Seierlm vlrldia. 7.4 7.4 7.4 18.5 7.4 3.7 0.0 0.0 3.7 0.0 0.0 0.0 66.7 3.7 14.8 3.7 3.7 3.7 0.0 0.0 0.0 0.0 0.0 0.0 3.7 0.0 40.7 7.4 0.0 3.7 0.0 7.4 7.4 0.0 0.0 7.4 40.7 7.4 29.6 7.4 29.6 3.7 7.4 0.0 22.2 3.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 44.4 7.4 3.7 7.4 7.4 14.8 3.7 0.0 0.0 0.0 0.0 3.7 0.0 0.0 55.6 7.4 3.7 3.7 18.5 0.0 0.0 7.4 7.4 0.0 0.0 0.0 0.0 $9.3 7.4 14.8 7.4 0.0 0.0 0.0 0.0 3.7 0.0 0.0 48.2 7.4 3.7 0.0 0.0 0.0 0.0 0.0 7.4 0.0 0.0 0.0 7.4 7.4 0.0 0.0 0.0 0.0 0.0 0.0 3.7 3.7 0.0 7.4 0.0 0.0 0.0 14 8 7.4 7.4 0.0 3.7 7.4 0.0 0.0 7.4 3.7 14.8 14.8 7.4 0.0 0.0 0.0 00 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 88.9 0.0 33.3 22.2 85.2 0.0 11.1 7.4 85.2 0.0 14.8 18.5 96.3 0.0 18.5 25.9 81.5 0.0 22.2 14.8 92.6 0.0 22.2 37.0 88.9 3.7 29.6 7.4 7.4 18.5 29.6 29.6 14.8 33.3 22.2 0.0 22.2 U.l 0.0 U .l U.l 3.7 0.0 7.4 0.0 7.4 0.0 0.0 0.0 51.9 14.8 7.4 51.8 3.7 0.0 40.7 . 25.9 22.2 U . l 0.0 11.1 0.0 40.7 14.8 18.5 14.8 25.9 7.4 0.0 0.0 0.0 3.7 3.7 0.0 0.0 59.3 0.0 0.0 0.0 14.8 3.7 7.4 3.7 0.0 0.0 7.4 7.4 0.0 0.0 0.0 59.3 0.0 7.4 7.4 7.4 7.4 0.0 3.7 0.0 3.7 3.7 0.0 0.0 59.3 7.4 0.0 3.7 0.0 0.0 85.2 74.1 92.6 77.8 852 0.0 0.0 0.0 22.2 0.0 0.0 11.1 3.7 avara season annual. 18.5 0.0 0.0 0.0 0.0 0.0 0.0 18.5 33.3 3.7 7.4 48.2 U.l 44.4 29.6 37.0 *0.7 33.3 0.0 22.2 22.2 29.6 3.7 0.0 22.2 0.0 3.7 29.6 3.7 3.7 25.9 7.4 3.7 25.9 14 8 0.0 3.7 7.4 0.0 0.0 0.0 3.7 11.1 22.2 0.0 11.1 22.2 11.1 14.8 0.0 0.0 0.0 0.0 11.1 0.0 0.0 0.0 0.0 0.0 3.7 3.7 7.4 3.7 11.1 7.4 0.0 0.0 0.0 7.4 14.8 3.7 0.0 0.0 3.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 81.5 0.0 14.8 14.8 85.2 0.0 14.8 22.2 77.8 0.0 29.6 14.8 81.5 0.0 18.5 7.4 96.3 0.0 22.2 85.2 0.0 44.4 25.9 3.7 22.2 25.9 33.3 14.8 18.5 0.0 3.7 3.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 81.5 0.0 37.0 85.2 0.0 11.1 25.9 81.5 0.0 18.5 U .l 81.5 0.0 29.6 14.8 25.9 22.2 11.1 3.7 18.5 7.4 0.0 0.0 3.7 0.0 0.0 0.0 0.0 77.8 7.4 14.8 3.7 3.7 3.7 0.0 0.0 0.0 0.0 0.0 3.7 3.7 3.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 25.9 0.0 0.0 7.4 7.4 U. l U . l 7.4 0.0 44.4 0.0 3.7 0.0 0.0 7.4 3.7 7.4 3.7 51.8 0.0 11.1 0.0 3.7 0.0 0.0 0.0 0.0 E 14.8 70.4 0.0 3.7 3.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 29.6 0.0 0.0 0.0 0.0 11.1 3.7 0.0 0.0 3.7 3.7 0.0 96.3 81.5 0.0 11.1 0.0 0.0 0.0 11.1 14.8 0.0 *0.7 0.0 37.0 18.5 7.4 0.0 0.0 0.0 14.8 3.7 0.0 0.0 85.2 0.0 t4.8 0.0 0.0 0.0 0.0 3.7 0.0 0.0 0.0 74.1 7.4 3.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 37.0 40.7 0.0 0.0 0.0 7.4 14.8 3.7 0.0 18.5 3.7 0.0 3.7 3.7 0.0 0.0 0.0 0.0 0.0 25.9 63.0 0.0 18.5 18.5 96.3 0.0 22.2 37.0 85.2 0.0 22.2 11.1 29.6 22.2 U.l 3.7 0.0 3.7 3.7 0.0 7.4 22.2 7.4 3.7 U. l 100.0 100.0 100.0 100.0 100.0 100.0 100.0 100.0 0.0 96.3 63.0 0.0 0.0 0.0 0.0 0.0 7.4 0.0 0.0 0.0 3.7 0.0 0.0 U.l 7.4 3.7 11.1 96.3 63.0 0.0 14.8 3.7 0.0 3.7 0.0 3.7 0.0 0.0 3.7 55.6 7.4 7.4 7.4 3.7 0.0 0.0 0.0 3.7 0.0 0.0 0.0 40.7 7.4 92.6 70.4 0.0 0.0 S 0.0 96.3 63.0 96.3 59.3 S 29.6 25.9 S 0.0 S 0.0 0.0 s 77.8 74.1 5 3.7 14.8 29.6 Z S E I 5 E 44.4 37 8 E % S 44.4 £ E 8 S E 3.7 7.4 3.7 0.0 0.0 0.0 3.7 0.0 0.0 0.0 51.8 7.4 7.4 7.4 7.4 3.7 3.7 0.0 0.0 0.0 0.0 0.0 0.0 55.6 3.7 2.4 0.0 3.7 0.0 U . l U.l 100.0 100.0 100.0 100.0 100.0 100.0 100.0 100.0 100.0 100.0 100.0 U.l 0.0 59.3 25.9 3 £ s 5 5 U .l 3.7 0.0 0.0 0.0 0.0 3.7 0.0 3.7 0.0 3.7 7.4 7.4 0.0 U.l 7.4 0.0 0.0 0.0 3.7 0.0 0.0 0.0 0.0 18.5 0.0 0.0 0.0 18.5 55.6 0.0 55.6 3.7 U .l 7.4 3.7 0.0 0.0 3.7 0.0 7.4 0.0 0.0 0.0 37.0 3.7 7.4 40.7 14.8 14.8 7.4 7.4 11.1 0.0 0.0 0.0 0.0 0.0 0.0 88.9 0.0 29.6 29.6 85.2 0.0 29.6 18.5 74.1 0.0 18.5 29.6 92.6 0.0 29.6 22.2 11.1 14.8 25.9 U .l 0.0 3.7 7.4 96.3 100.0 100.0 100.0 0.0 0.0 0.0 0.0 0.0 3.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 U.l 55.6 7.4 14.8 100.0 0.0 7.4 0.0 0.0 110 Elllela nyctelea Helianthus 37.0 7.4 B- t e c t o r u a Annual Eorbs: H. a grasses: Aveua C. 5 grasses: Agropyron daaystachyua A. s a l t h l l A. a S LITERATURE CITED 112 LITERATURE CITED Agricultural Research Service and Worth Dakota Agricultural Experiment Station Research Staffs. 1975. Progress report, research on reclamation of strip-mined lands in the Northern Great Plains. Northern Great Plains Res. Center and N. D. Ag. Exp. Sta. Progress Rep. Fargo, N.D. 20 p. Agricultural Research Service and North Dakota Agricultural Experiment Station Research Staffs. 1977. Progress report, research on reclamation of strip-mined lands in the Northern Great Plains. Northern Great Plains Res. . Center and N. D. Ag. Exp. Sta. Progress Rep. Fargo, N.D. 26 p. Aldon, E. F. 1978. Reclamation of coal-mined land in the Southwest. J. Soil Water Cons. 33:75-79. Aldon, E. F. and H. W. Springfield. 1973. Revegetating coal mine spoils in New Mexico. U.S.D.A. For. Serv. Res. Note RM-245. Rocky Mt. For. and Range Exp.Sta., Fort Collins, Colo. 4 p. Aldon, E. F., H. W. Springfield, and G. Garcia.1975. Can soil amendments aid revegetation of New Mexico coal mine spoils? U.S.D.A. For. Serv. Res. Note RM-292. Rocky Mt. For. and Range Exp. Sta., Fort Collins, Colo. 7 p. , Aldon, E. F., W. H. Springfield, and D. G. Scholl. 1976. Fertilizer response of alkali sacaton and fourwihg saltbush grown oh coal mine spoil. U.S.D.A. For. Serv. Res. Note RM-306.. Rocky Mt. For. and Range Exp. Sta., Fort Collins, Colo. 4 p. Allen, L. J., L. H. Barbers, R. R. Schalles, C. E. Owensby, and E. F. Smith. 1976. Range burning and fertilization related to nutritive value of bluestem grass. J. Range Manage. 29:306-308. Amer, F. M. and W. V. Bartholomew. 1951. Influence of oxygen concentration in soil air on nitrification. Soil Sci. 71:215219. Argonne National Laboratory. 1979. Land reclamation program annual report, 1978. Available from National Technical Information Service, 5285 Port Royal Road, Springfield, VA 22161. Bains, S. S. and M. Fireman. 1964. Effects of exchangeable sodium percentage on growth and absorption of essential nutrients and sodium by five crop plants. Agron. J. . 56:432-435. Baldwin, D. M., N. W. Hawkinson, and E. W. Anderson. fertilization of native rangeland in Oregon. 27:214-216. 1974. High-rate J. Range Manage. 113 Barnhisel, R. I. and V. P. Evangelou. 1981. Role of phosphorus and water availability in reclaiming surface-mined spoils. Reel. News and Views. 1:1-3. Bauer, A., E. H. Vasey, R. A. Young, and J. L. Ozbun. 1967» Stored moisture best guide to nitrogen needed. N. D. Ag. Exp. Sta. Farm ■ Res. 24:15-24. Bauer, A., W. A. Berg, and W. L. Gould. 1978a. Correction of nutrient deficiencies and toxicities in strip-mined lands in semiarid and arid regions, pp. 451-466 in: Reel, of Drastically Disturbed Lands (F. W. Schaller and P. Sutton, eds.). ASA-CSSASSSA. Madison, NI. 742 p. Bauer, A., P. Nyren, G. Reichman, G. Gee and J. Gilley. 1978b. Fertilization of wheat, corn, and grass-legume mixture grown on reclaimed spoilbanks. N. Dak. Ag. Exp. Sta. Res. Rep. 67. N.D. State Univ., Fargo, N.D. 15 p. Beaton, J. D. and D. W. L. Read. 1963« Effects of temperature and moisture on phosphorus uptake from a calcareous Saskatchewan soil treated with several pelleted sources of phosphorus. Soil Sci. Soc. Amer. Proc. 27:61-65. Berg, W. A. 1980. Nitrogen and phosphorus fertilization of mined lands, pp. 20-1-20-8 in: Adequate Reclamation of mine lands? Symp. Soil Cons. Soc. of Amer. and WRCC-21. Billings, Mt. Bjornson, 0, R, and J, R. Sims. 1971. Movement of phosphorus from several fertilizer sources in Bozeman silt loam soil. Mt. Ag. Exp. Sta. Res. Rep. 14. Mt. State Univ., Bozeman, Mt. 10 p. Black, A. L, 1968. Nitrogen and phosphorus fertilization for production of crested wheatgrass and native grass in northeastern Montana. Agron. J. 60:213-216. Black, A. L. and L. L. Reitz. 1969. Row spacing and fertilization influences on forage and seed yields of intermediate wheatgrass, Russian wildrye, and green needlegrass on dryland. Agron. J. 61:801-805. Black, A. L. and J. R. Wight. 1972. Nitrogen and phosphorus availability in a fertilized rangeland ecosystem of the Northern Great Plains. J. Range Manage. 25:456-460. Black, C. A. 1965. Methods of Soil Analysis. Inc. Madison, W L 1572 p. Blaser, R. E. and M. C. Brady. associations. Agron. J. Amer. Soc. of Agron., 1950. Nutrient competition in plant 42:128-135. Brady, N„ C. 1974. The Mature and Properties of Soils. Pub. Co. New York. 639 p. Brown, B. A. and R. I. M unsell. 1956. as influenced by fertilization. 35 p. Bryan, G. G. and W. E. McMurphy. 1968. as influences on grass seedlings. MacMillan Clovers in permanent grassland Storrs Ag. Exp. Sta. Bui. 329. Competition and fertilization J. Range Manage. 21:98-101. Buchholz, J. 1972. Some effects of fertilization on revegetation of coal mine spoils in southeastern Montana. M. S. Thesis, Dept. Animal and Range Sci. Mt. State U., Bozeman. 51 p. Coenenberg, J. C. 1982. Practical considerations in establishing diverse plant community on minesoils. pp. B6-1-B6-20 in: Mt. Ag. Exp. Sta; Res. Rep. 194» Mt. State Univ., Bozeman, Mt. Cooper, C. S, 1975. Influence of harvest management and midseason nitrogen fertilization on yield, botanical composition, and crude protein content of grass-legume mixtures, Mt. Ag. Exp. Sta. Bui. 676. Mt. State Univ., Bozeman, Mt. 15 p. Cooper, C„ S., D. E. Baldridge, and C. W. Roath. management of irrigated pasture mixtures. 622. Mt. State Univ., Bozeman, Mt. 22 p. 1969. Selection and Mt. Ag. Exp. Sta. Bui. Cundell, A. M. 1977. The role of microorganisms in the revegetation of strip-mined land in the western United States. J. Range Manage. 30:299-305. Danielson, R. M., J. Zak, and D. Parkinson. 1979. Plant growth and mycorrhizal development in amended coal spoil material, pp. 912919 in: Ecology and Coal Resource Development II. CM. K. Mali, ed.). Pergamon Press. New York. 1091 p. Daubenmire, R. 1959. A canopy-coverage method of vegetational analysis. Northwest Sci. 33:43-64. DePuit, E. J. and J. G. Coenenberg. 1979. Response of revegetated coal strip mine spoils to variable fertilization rates, longevity of fertilization program and season of seeding. Mt. Ag. Exp. Sta. Res. Rep. 150. Mt. State Univ., Bozeman, Mt. 81 p. DePuit, E. J., J. G. Coenenberg, and W. H. Willmuth. 1978. Research on revegetation of surface mined lands at Colstrip, MT: Progress report, 1975-77. Mt. Ag. Exp. Sta. Res. Rep. 127. Mt. State Univ., Bozeman, Mt. p. 165. 115 Doner, H. E», M. G. Volz, and A. Pukite. 1973. Determination of nitrate by a chromotropic acid method, pp. 20-21 in: Summary of papers, Statewide conference on Adv. and Innovations in Proc. for Soil, Plant, and Water Analysis. Berkeley, CA. Ensminger, L. E. and H0 W. Pearson. 1957. Residual effects of various phosphates as measured by yields, P^2 uptake, and extractable phosphorus. Soil Sci. Soce Amer. Proc. 21:80-84. Epstein, E. 1972. Mineral Nutrition of Plants: Principals and Perspectives. John Wiley and Sons. New York. 412 p. Farmer, E. E., R. W. Brown, B. Z. Richardson, and P. E. Packer. 1974. Revegetation research on the Decker coal mine in southeastern Montana. U.S.D.A. For. Serv. Res. Paper INT-162. Ogden, UT. 12 p. Goetz, H. 1969. Composition and yields of native grassland sites fertilized at different rates of nitrogen. J. Range Manage. 22:384-390. Goetz, H. 1970. Growth and development of Northern Great Plains species in relationship to nitrogen fertilization. J. Range Manage. 23:112-117. Goetz, H. 1975. Effect of site and fertilization on protein content of hative grasses. J. Range Manage. 28:380-385. Haby, V. A. and Ri A. Larson. 1976. Soil nitrate-nitrogen analysis by the chromotropic acid procedure. Proc. 27th Annual Fertilizer Conf0 of the Pacific Northwest. Billings, Mt. Hodder, R. L., D. E. Reyerson, R. Mogen* and J. Buchholz. 1971. Coal mine spoils reclamation research project, Western Energy Company, Colstrip, Montana:progress report 1970. Mt. Ag. Exp. Sta. Res. Rep. 8. Mt. State Univ., Bozeman, Mt. 56 p. Holechek, J, L. 1976. Initial effects of different species mixtures and fertilizer rates on a mine spoils rehabilitation planting. M.S. Thesis, Dept. Animal and Range Sci. Mt. State Univ., Bozeman, Mt. 91 p. Hons, F. M. and L. R. Hossner. 1979. Soil nitrogen relationships in spoil material generated by the surface mining of lignite coal. Soil Sci. 129:222-228. Houlton, H. A. R. 1975. Range vegetation response to nitrogen fertilization in north central Montana, pp. 1-15 in: Proc. Range Fert. Symp., Mt. Ag. Exp. Sta. Res. Rep. 70. Mt. State Univ., Bozeman, Mt. 116 Houston, W. R0 and D« N» Hyder0 1975. Ecological effects and fate of N following massive N fertilization of mixed-grass plains. J0 Range Manage. 28:56-60. Howard, G, S., G, E0 Schuman, and F0 Rauzi, 1977. Growth of selected plants on Wyoming surface-mined soils and flyash. J. Range Manage. 30:306-310, Johnston, A., S, Smoliak, A. D0 Smith, and L0E. Lutwick0 1967. Improvement of southeastern Alberta range with fertilizers. Can. J, Plant Sci0 47:671-678, Johnston, A., A. D. Smith, L. E0Latwich, and S. Smoliak. 1968. Fertilizer response of native and seeded ranges. Can. J. Plant Sci. 48:467-472. Johnston, A., S. Smoliak, A, D. Smith, and L0E0 Lutwick0 1969. Seasonal precipitation, evaporation, soil moisture, and yield of fertilized range vegetation. Can. J. Plant. Sci, 49:123-128. Kilcher, M. R., S. Smoliak, W,A, Hubbard, A. Johnston, A. I. H. Gross, and E. V. McCurdy. 1965. Effects of inorganic nitrogen and phosphorus fertilizer on selected sites of native grassland in Western Canada. C^n. J0 Plant Sci. 45:229-237. Klages, M. G. and D0 E0 Ryerson0 1965. Effect of nitrogen and irrigation on yield and botanical composition of western Montana range. Agron, J. 57:78-81. Lambert, M. G0 and D. A. Grant. 1980. Fertilizer and lime effects on some southern North Island hill pastures. N. Z. J0 Exp. Ag. 8 :223-230 . Langin, E0 J,, R. C. Ward, R, A, Olson, and H. F. Rhoades. 1962. Factors responsible for poor responses of corn and grain sorghum to phosphorus fertilization: II. lime and P placement effects on P-Zn relations. Soil Sci. Soc. Am. Proc, 26:574-578. Lauenroth, W, K. and J. L. Dodd, 1979° Response of native grassland legumes to water and nitrogen treatments. J0 Range Manage, 32:292-294. Lorenz, R, J. and G. A, Rogler0 1972, Forage production and botanical composition of mixed prairie as influenced by nitrogen and phosphorus fertilization. Agron0 J0 64:244-249. Lorenz, R, J. and G0 A0 Rogler, 1973« Growth rate of mixed prairie in response to nitrogen and phosphorus fertilization. J. Range Manage. 26:365-368. 117 Marrs, R. H. and A1 D1 Bradshaw. reclaimed china clay wastes. Appl1 Eco1 17:727-736. 1980. III1 Ecosystem development on Leaching of nutrients. J1 McGinnies, W» J1 and P1 J1 Nicholas. 1980. Effects of topsoil thickness and nitrogen fertilizer on the revegetation of coal mine spoils, J. Environ. Qual1 9:681=685. McHenry, J. R1 1963. Theory and application of neutron scattering in measurement of soil moisture. Soil Sci1 95:294-307. Meyri, R, L., J1 L1 Holechek, and E1 S1 Sundberg1 1975. Short and long term fertilization requirements for reclamation of mine spoils at Colstrip, Montana, pp. 266-279 in: Proceedings of the Fort Union Coal Field Symposium. Vol. IIL Eastern Mt. College, Billings. Meyn, R. L., E1 S1 Sunberg, and R1 P. Young. 1976. Research on reclamation of surface mined lands at Colstrip, Montana: progress report 1975. Mt. Agr. Exp1 Sta1 Res. Rep1 101. Mt. StateUniv., Bozeman, Mt. 84 p. , Miller, R1 D1, and D1 D1 Johnson, 1964, The effect of soil moisture tension on carbon dioxide evolution, nitrification and nitrogen mineralization. . Soil Sci1 Soc. Am. Proc. 28:644-647.. Moore, A. N., E1 M. Brouse, and H1 F1 Rhoades. 1968. Influence of phosphorus fertilizer placement on two Nebraska sub-irrigated meadows. J. Range Manage. 21:112-114. Mosse, B. 1973: Plant growth responses to vesicular-arbuscular mycorrhiza. IV. in soil given additional phosphate. New Phytol. 72:127-136. Mott, G1 0. 1943. Effectiveness of fertilization and management in increasing yields of pastures in Indiana. Soil Sci. Soc. Amer1 Proc1 8:276-281. M unshower, F. F1 and E1 J1 DePuit1 1976. The effects of stack emmissions on the range resource in the vicinity of Colstrip, Montana. Progress report 1975. Mt. Ag. Exp1 Sta1 Res. Rep. 98, Mt. State Univ1 , Bozeman, Mt. 112 p. Muhshower, F. F1, E. J. DePuit, and D1 R1 Neuman. 1978. of stack emissions on the range resource in the Colstrip, Montana: Pre-operational description ecosystem. Mt. Ag. Exp1 Sta1 Res. Rep. 126. Mt. Bozeman, Mt. 196 p. The effects vicinity of of the range State Univ., 118 National Oceanic and Atmospheric Administration. 1981. Climatologi­ cal data: annual summary. U. S. Dept, of Com., Environ. Data and Information Service. Vol. 84. Meter, J1. and W. Wasserman. Richard D. Irwin, Inc. 1974. Applied Linear Statistical Models. Homewood, 111. 842 p. Odum, E. P. 1971. Fundamentals of Ecology. Saunders Co. Philadelphia. 547 p. 3rd edition. W. B. Olson, R. A. and A. F. Dreier. 1956. Nitrogen a key factor in the fertilizer phosphorus efficiency. Soil Sci. Soc. Am. Proc. 20:509-514. Omodt, H. W., F. W. Schroer, and D. D. Patterson. 1975. The properties of important agricultural soils as criteria for mined land reclamation. N. D, Ag. Exp. Sta. Bui. 492. N.D. State Univ., Fargo, N.D. 52 p. Packer, P. E. 1974. Rehabilitation potentials and limitations of surface-mined land in the Northern Great Plains. U.S.D.A. For. Serv. Gen. Tech. Rep. INT-14.' Intermt. For. and Range Exp. Sta., Ogden, UT. Parker, D. T, and W. E. Larson. 1962. Nitrification as affected by temperature and moisture content of mulched soils. Soil Sci. Soc. Am. Prbc. 26:238-242. Parkinson, D. 1979. Microbes, mycorrhizae, and mine soil. pp. 626642 in: Ecology and Coal Resource Development (M. K. Wall, ed.). Pergaraon Press, New York. 1091 p. Payne, G. F. 1973. Exp. Sta. Bui. Vegetative rangeland types in Montana. Mt. Ag. 671. Mt. State Unlv., Bozeman, Mt. 16 p. Pettit, R. D. and D. W. Deering. 1974. Yield and protein content of s a ndyland range forages as affected by three nitrogen fertilizers. J. Range Manage. 27:211-213. Pielou, E. C. 1975. York. 16.5 p. Ecological Diversity. John Wiley and Sons. New Power, J. F. 1967. The effect of m o i s t u r e on f ertilizer N immobilization in grasslands. Soil Sci. Soc. Am. Proc. 31:223226. Power, J. F. 1972a. Nitrate enrichment of soils and water under dryland in the Great Plains. pp. 119-136 in: Control of agriculture-related pollution in the Great Plains. Great Plains Agric. Counc. Publ. 60. Neb. Ag. Exp. Sta., Lincoln, Neb. 119 Power, J. F„ 1972b. Fafce of fertilizer nitrogen applied to a Northern Great Plains rangeland ecosystem. J. Range Manage. 25:367-371. Power, J. F. 1979. Use of slow release N fertilizers on native mixed prairie. Agron. J. 71:446-449. Power, J. F. and J. Alessi. 1971» Nitrogen fertilization of semi arid grasslands: plant growth and soil mineral N levels. Agron. J. 63:277-280. Power, J. F., J. Alessi, G. A. Reichman, and D. L. Grunes. 1973. Recovery, residual effects, and fate of nitrogen fertilizer sources in a semiarid region. Agron. J. 65:765-768. Power, J. F., W. 0. Willis, F. M. Sandoval, and J. J. Bond. Can productivity of mine land be restored in North Dakota? Ag. Exp. Sta. Farm Res. 31:30-32. 1974. N. D. Rauzi, F. 1978. High rates of nitrogen change composition of shortgrass rangeland in southeastern Wyoming. J. Range Manage. 31:366-370. Rauzi, F., R. L. Lang, and L. I. Painter. 1968. Effects of nitrogen fertilization on native rangeland. J. Range Manage. 21:287-291. Rehm, G. W., W. J. Moline, and E. J. Schwartz. 1972. Response of a seeded mixture of warm-season prairie grasses to fertilization. J. Range Manage. 25:452-456. Reichman, G. A., D. L. Grunes, and F. G. Viets, Jr. 1966. Effect of soil moisture on ammonifiestion and nitrification in two northern plains soils. Soil Sci. Soc. Am. Proc. 30:363-366. Rennie, D. A. and R. J. Soper. 1958. The effect of nitrogen additions on fertilizer-phosphorus availability. II. J. Soil Sci. 9:155-167» Reuszer, H. W. 1957. Composts, peats and sewage sludge, pp. 237-245 in: Soil, the 1957 Yearbook of Agriculture. U.S.D.A. 784 p. Richards, L. A. 1954. Diagnosis and improvement of saline and alkali soils. U.S.D.A. Ag. Handbook 60. 160 p. Riley, D. and S. A. Barber. 19 7 1. Effect of ammonium and nitrate fertilization on phosphorus uptake as related to root-induced pH changes on the root-soil interface. Soil Sci. Soc. Am. Proc. 35:301-306. Rogler, G. A. and R, J. Lorenz. 1957» Nitrogen fertilization of northern Great Plains rangelands. ,J. Range Manage. 10:156-160. 120 Ross, R. L. and H. E. Hunter. 1976. Climax vegetation of Montana, based on soils, and climate. U.S.D.A. Soil Cons, Serv., Bozeman, Mt. 64 p. Samuel, M. J., F. Rauzi, and R. H. Hart. 1980. Nitrogen fertilization of range; yield, protein content, and cattle behavior. J. Range Manage. 33$119-121. Sandoval, F. M. and J. F. Power. 1977. Laboratory methods recommended for chemical analysis of mined-land spoils and overburden in Western United States. U.S.D.A. Ag. Handbook 525. BI p. Schafer, W. M. 1979a. Mine soil resources in the Northern Great Plains; Characterization, evaluation, and potential. Ph.D. Dissertation. Mt. State Univ., Bozeman, Mt. 203 p. Schafer, W. M. 1979b. Guides for estimating cover-soil quality and mine soil capability for use in coal stripmine reclamation in the western United States. Reel. Review. 2:67-74. Schafer, W. M., G. A. Nielsen, D. J. Dollhopf, and K, Temple. 1979. Soil genesis, hydrological properties, root characteristics, and microbial activity of I to 50 year old stripmine spoils. EPA60017-79-100. 212 p. Sims, J. R. and V. A. Haby. 1970. Simplified colormetric determination of soil organic matter. Soil Sci. 112:137-141. Sims, J. R. and G. D. Jackson. 1971. Rapid analysis of soil nitrate with chromotropic acid. Soil Sci. Soc. Am. Proc. 35:603-606. Skilbred, C. L. 1979. Plant succession on five naturally revegetated strip-mined deposits at Colstrip, Montana. M. S. thesis. Mt. State. Univ., Bozeman, Mt. 128 p. Smika, D. E., H. J. Haas, and J. F. Power. 1965. Effects of moisture and nitrogen fertilizer on growth and water use by native grass. Agron. J. 57:483-486. Smoliak, S. 1965. Effects of manure, straw and inorganic fertilizer on northern Great Plains. J. Range Manage. 18:11-15. Snedecor, G. W. and W. G. Cochran. 1980. Statistical Methods. State Univ. Press, Ames, Iowa. 507 p. Soil Survey Staff. 1975. Soil Taxonomy. Ag. Handbook 436. 754 p. Standford, G. and E, Epstein. relations in soil. 1974. Iowa U.S.D.A. Soil Cons. Serv. Nitrogen mineralization - water Soil Sci. Soc. Am. Proc. 38:103-107« 121 S t a n d f ord, G„ M., M. H. Frere, and D. E. Schwaninger. 1973» Temperature coefficient of soil nitrogen mineralization. Soil Sci. 115:321-323. Stroehlein, J. L., P. R. Ogden, and B. Billy. fertilizer application on desert grasslands. 21:86-89. 1968. Time of J. Range Manage. Synder, G. H., A. E. Kretschmer, and J. B. Sartain. 1978. Field response of four tropical legumes to lime and superphosphate. Agron. J. 70:269-273. Tisdale, S. L. and W. L. Nelson. 1975. Soil F e r tility Fertilizers. 3rd ed. Macmillan Pub. Co. New York. 694 p. and United States Department of Agriculture. 1982. National list of scientific plant names. Soil Cons. Ser. SCS-TP-159. Veseth, R. and C. Montagne. 1980. Geologic parent materials of Montana soils. Mt. Ag. Exp. Sta. Bui. 721, Mt. State Univ., Bozeman, Mt. 117 p. Warnes, D. D. and L. C. Newell. 1969. Establishment and yield responses of warm-season grass strains to fertilization. J. Range Manage. 22:235-240. Welch, N. H., E. Burnett and E. B. Hudspeth. 1962. Effect of fertilizer on seedling emergence and growth of several grass species. J. Range Manage. 15:94-98. West, P. W. and T. P. Ramachandran. 1966. Spectrophotometric determination of nitrate using chromotrophic acid. Analytical Chemical Acta. 35:317-324. White, L M. and A. D. Halvorson. 1980. Nitrate levels in vegetative and floral tillers of western wheatgrass and green needle grass as affected by nitrogen fertilization. Agron, J. 27:143-148. Wight, J. R. and A. L. Black. 1972. Energy f i x a t i o n and precipitation use efficiency in a fertilized rangeland ecosystem of the Northern Great Plains. J. Range Manage. 25:376-380. Wight, J. R. and A. L. Black. 1978. Soil water use and recharge in a fertilized mixed prairie plant community. J. Range Manage. 3 1 :280 -282 . Wight, J. R. and A. L. Black. 1979. Range fertiliztaion: plant response and water use. J. Range. Manage. 32:345-349. 122 Williams, P. J. and J. E. Cooper. nitrification .in amended 13:533-543. 1976. Hitrogen mineralization and colliery spoils. J. Appl. Ecol6 Wilson, H. A. 1965. The microbiology of strip mine spoil. • Ag. Exp. Sta. Bui. 5061:5-44. W. Va. Young, S6 A. and R. B. Rennick. 1982« Establishment of seeded species oh coal surface minesoils following temporary irrigation. Mt. Ag. Exp. Sta. Res. Rep. 200. Mti State Univ., Bozeman, Mt. 59 P. MONTANA STATE U NIVERSITY LIBRARIES stks N378.H444@ Theses Response o f native species to variable n RL 3 1762 00169238 1 m i