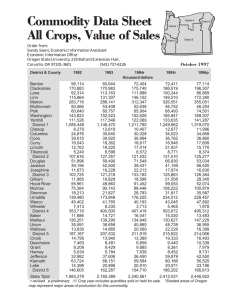
103-2 ?TMFNT of flf'FAN(1f;RAPNY OREGON STATE UNIVERSITY SCHOOL of SCIENCE AND DISTRIBUTION OF
advertisement

103-2 ?TMFNT of flf'FAN(1f;RAPNY NENAL EM R. rILLAMOOK BAY' SCHOOL of SCIENCE OREGON STATE UNIVERSITY FINAL REPORT: SPECIES COMPOSITION AND DISTRIBUTION OF MARINE NEKTON IN THE PACIFIC OCEAN OFF OREGON ATOMIC ENERGY COMMISSION CONTRACT AT(45-1)1726 Reference 63-26 September 1961-1963 L September 1963 SPECIES COMPOSITION AND DISTRIBUTION OF MARINE NEKTON IN THE PACIFIC OCEAN OFF OREGON Final Progress Report 1 September 1961 through l September 1963 Atomic Energy Commission Contract AT(45-1)1726 Department of Oceanography Oregon State University m" FINAL REPORT: SPECIES COMPOSITION OF MARINE NEKTON IN THE PACIFIC OCEAN OFF OREGON CONTENTS Page Summary ............................................................ Distribution and Ecology Section 1. Section 2. Preliminary Observations on the Distribution of Mesopelagic Fishes off Oregon .......................... 13 Species Composition and Distribution of Pelagic Cephalopods From the Pacific Ocean off Oregon, 1961-1963 .............................................. Section 3. Section 4. Notes on the Occurrence and Distribution of Macroplankton off the Oregon Coast ..................... 37 49 Vertical Distribution of the Numbers and Biomass of Mesopelagic Fishes with an Improved Isaacs-Kidd Midwater Trawl .......................................... 67 Radioecology Section 5. Notes on the Vertical Distribution of Zinc-65 Zirconium-95 from Oceanic Section 6. Section 7. and Animals ...................... Radioactivity and its Relationship to Oceanic Food Chains ................................................. Zinc-65 in Euphausiids as Related to Columbia River Water off Oregon ....................................... 81 91 107 Publications and Papers ............................................ 131 LIST OF FIGURES Follows page Figs. Section 1. 1. Location of midwater trawling stations off Oregon.......... 14 2. Catches of the four dominant mesopelagic fishes in the upper 200 m ................................................ 16 The variability of catches of the four dominant mesopelagic fishes ............................................. 18 Depth variations in the catches of three lantern fishes at night ................................................. 20 A temperature and salinity profile, 50 miles off Newport, Oregon, August 1961 ........................................ 22 6a. Average monthly catches of Lampanyctus leucopsarus ......... 26 6b. Average monthly catches of Diaphus theta ................... 26 3. 4. 5. Section 2. Location of midwater trawling stations off the Oregon coast ...................................................... 38 The relative abundance of pelagic cephalopods found off Oregon in midwater trawl samples ........................... 40 3. Size-frequency distribution of Abraliopsis ................. 46 4. Size-frequency distribution of Gonatus fabricii and Gonatus Larvae ..................................................... 46 1. 2. Section 3. 1. The distribution of adult and larval forms of similis for June, July, and August 1961 ................................ as recorded 58 2. Salp catch by the midwater trawl (1961-1963)... 60 3. Catches of two chaetognaths during January 1962 at three latitudes off Oregon ....................................... 64 Follows Section 4. Isaacs-Kidd midwater trawl showing the depth-distance recorder mounted on the depth depressor ................................ 70 Duplication of a trace made with the depth-distance recorder on a tow which sampled three depths within the upper 1000 m... 70 3. Histograms showing the number of fishes captured per 1000 m3 76 4. Histograms showing the wet weight of fishes collected per 1000 m3.... ................................................. 76 1. 2. Section 5. 1. Zn65 and Zr95-Nb95 contents in pelagic animals for day and night periods ................................................. 86 Section 6. 1. Comparison of gamma emitters from several trophic levels ...... 96 2. Concentrations of gamma emitters from several trophic levels.. 98 3. Comparison of spectra of euphausiids and copepods from the same sample ................................................... 100 Comparison of spectra of euphausiids and sergestids from the same sample .................... .............................. 100 4. Section 7. 1. Gamma-ray spectra of particulate material in two Oregon rivers 112 2. Zinc-65 in euphausiids and salinity distribution during summer 1961 ................... 112 3. Same as Figure 2 for November 1961 ............................ 112 4. Same as Figure 2 for January 1962 ............................. 112 5. Same as Figure 2 for March-April 1962 ......................... 114 6. Same as Figure 2 for July-August 1962 ......................... 114 FINAL REPORT: SPECIES COMPOSITION OF MARINE NEKTON IN THE PACIFIC OCEAN OFF OREGON SUMMARY This report summarizes the results of the research contract, "Species Composition and Distribution of Marine Nekton in the Pacific Ocean off Oregon" (AT(45-1)1726) for the two-year period from September 1, 1961, to September 1, 1963. INTRODUCTION Since it has been found that animals can accumulate and concentrate radioisotopes from their environment, their role in the biological transport of nuclides in the ocean has become of great interest. Nektonic or swimming animals (fishes, squids, shrimps, euphausiids, etc.) are known to undertake extensive vertical migrations, often through density gradients of the thermocline or halocline and into surface waters, and the movements of these pelagic animals may be more important than simple physical processes in the transport and distribution of radioisotopes in the ocean. Lateral movements of these animals may also be important in determining the fate of radionuclides introduced into the sea. However, virtually nothing is known about the extent of lateral movements of small, oceanic nekton. These animals are basic forage for higher consumers such as salmon and albacore. It is apparent, therefore, that there is a need to broaden our understanding of the ecology, distribution, and move- ments of these ubiquitous swimming animals that are intermediate in the oceanic food chain. Study of the oceanic nekton off Oregon was of interest for several reasons. First, no detailed studies of the oceanic fauna in this region 2 of the northeastern Pacific Ocean had ever been made. Little was known, therefore, about the species composition Secondly, the Pacific or distribution of marine nekton. Ocean off Oregon and Washington, because of the presence of the Columbia River, provides an exceptional opportunity for cooperative ecological from Hanford reactors both biologically This provides and radiological research. on this river maintains Neutron activation a "steady state" source of important and biologically inert isotopes in the ocean. a unique natural situation for a radioecological study, the results of which have applicability to the accidental or intentional disposal of radioactive materials at sea. Information on species of animals and their distribution is a prerequisite for interpretation of radioecological data. SAMPLING PROGRAM In order to provide data on the distribution of oceanic nekton, and more specifically, on spatial and temporal variations in distribution and species composition, a systematic sampling program was established in which standard collections were made routinely at predesignated sta- tions from 15 to 165 miles offshore along three latitudes off Oregon during every season of the year. This intensive sampling program was designed so that the distribution of animals could be compared between (1) stations across the continental slope, (2) latitudes, and (3) seasons of the year. In addition, collections at successive depths down to 1000 m were made routinely at a station over the outer continental slope, providing data on depth distribution and vertical migration. During the 3 two-year period of the contract, 44 cruises of the R/V ACONA were made in support of this program. A total of 353 samples with an Isaacs-Kidd For midwater trawl and 38 samples with a meter plankton net were taken. details on sampling and methods, see Section I. OBJECTIVES The primary objectives on the contract as stated in the original proposal and renewal request were as follows: (A) Characterization of the species composition of oceanic marine nekton and macroplankton off Oregon. (B) Distribution of animals, specifically spatio-temporal differences related to (1) depth and diurnal vertical migration,, (2) distance from shore, (3) latitude, and (4) seasons of the year. (C) Collections and identification of oceanic animals for radioanalyses and assisting in the interpretation of resulting data. SUMMARY OF RESULTS This section outlines the realization of the objectives stated above. A. Species Composition The major groups of oceanic marine nekton and macroplankton were studied taxonomically. The results represent the first detailed study of the species composition of these animals off the Oregon coast. Such data are basic to further research on radioecology or distributional analyses. 4 Taxonomic data for the collected material are summarized in the respective results sections: FISHES (Section 1) CEPHALOPODS (Section 2, Reprint 2) MACROPLANKTON EUPHAUSIIDS (Section 3A) NATANT SHRIMPS (Section 3B) SALPS (Section 3C) MEDUSAE, SIPHONOPHORES, (Section 3D) and CHAETOGNATHS A new species of squid and peculiar gelatinous egg masses of fish were described from the collected material (Reprint 1 and 2). B. Distribution of Nektonic Animals Study of variations in the distribution or abundance of oceanic animals has several inherent difficulties. The very nature of the open ocean environment imposes limitations on any sampling program. Numerous and repeated samples, desirable from a statistical standpoint, are made impractical due to the expense of ship operations. Patchy distribution of many marine organisms also leads to large variances, resulting in statistical analyses which suffer from insensitivity. Furthermore, available sampling devices for nektonic animals are at best only semiquantitative. These sampling problems were anticipated (see original proposal), and some attempt was made to measure the degree of variability encountered during repeated tows. For instance, coefficients of variation calculated for the catches of certain common fishes collected during one night at one station revealed a wide range of variability; high coefficients suggested patchy distribution or schooling activity (see Section 1). Thus, reliable assessment of differences in distribution of such oceanic populations, unless pronounced, require continued, long-term study. Despite these difficulties, however, it was possible to characterize cer- tain features of the distribution of nekton off Oregon. Seasonal Occurrence and Variations 1. Species Composition No marked seasonal variations were observed in the fundamental species composition of mesopelagic fishes (Section 1) or oceanic cephalopods (Section 2) collected off Oregon during the study period. The same dominant species prevailed in the upper 200 m in most oceanic samples. This is in contrast with neritic areas or large nekton where distinct seasonal changes may result from the migratory habits of many species. Seasonal variations in the concentration of Zn65 in euphausiids off Oregon were much less than the seasonal variations observed in Columbia River plume waters as determined by lowered salinities. This too may be inter- preted as evidence for a rather stable population of oceanic animals (see Section 7), 2, Seasonal Differences in Relative Abundance Although no marked seasonal changes in the species composition of nekton were evident in the ocean off Oregon, seasonal variations in the catches of common species of fishes and squids were noted. In both cases, highest catches were made during summer months, especially at stations 6 over the inner continental slope. The seasonal differences for the two dominant species of mesopelagic fishes (Lampanyctus leucopsarus and Diaphus theta) were generally significant statistically (see Section 1). The two dominant species of oceanic squids (Gonatus fabricii and Abraliopsis sp.) were six to eight times more common in collections during the summer than other seasons of the year (Section 2). In some cases these seasonal dif- ferences may be caused by increased availability of animals due to currents. In the case of Abraliopsis, however, the increased catches in the summer were accounted for by large numbers of young animals. This was also the case with the shrimp, Sergestes similis (Section 3B). Consequently, these seasonal maxima could be explained by seasonal breeding and recruitment. This indicates the importance of analyzing the size-structure of the populations at different seasons. The study of seasonal variations provided some interesting indications, but no general conclusions can be drawn on the consistency of these trends due to the limited data. Geographic Distribution and Variations 1, Latitudinal Distribution In general, little difference was apparent in the species composition or structure among the various east-west lines of stations, In the fishes, as with the euphausiids and salps, there were indications that the species diversity increased to the south, but there was no evidence for large latitudinal movements (See Section 1, 3A, and 3C). Variations in the relative abundance of certain nekton associated with distance from shore were suggested from the catch statistics. Highest catches of fishes and cephalopods were made at nearshore oceanic stations 7 during the summer months, whereas, during other seasons catches along the station lines were more uniform (Section 1, Fig. 6). Another rather con- sistent feature was the notable lack of oceanic nekton at the shallow water stations off Newport (Section 1). Since this was apparent during the current patterns of various seasons, it suggests that these animals are able to influence their own geographic position and remain in deep water by laterial movements. Depth Distribution and Vertical Migration 1. Day-Night Differences Within the Upper 200 m Distinct differences in the number of mesopelagic animals during day and night periods within the upper 200 m were found. For example, few mesopelagic fishes were taken during daylight hours within these Differences in the upper upper waters, but they were abundant at night. depth ranges of the dominant lantern fishes were noted by trawling to successive depths within the upper 100 m. From these data, it was con- cluded that the four common species of fishes migrate vertically through the density gradients of the halocline; moreover, some species cross the thermocline as well (for details, see Section 1). 2. Vertical Distribution Within the Upper 1000 m Comparison of successive tows to 200, 500, and 1000 m depths indi- cated that some species of oceanic nekton do not commonly migrate into epipelagic waters at night (Section 1, 2, 3A, and study is required to learn the 3B). However, more extent, if any, of migrations below 500 m. 8 Adaptation of the Lamont Multiple Plankton Sampler (MPS) as an openingclosing cod-end collecting unit for the midwater trawl, along with other modifications of the trawl, provided quantitative data on the depth distribution of oceanic animals. Section 4 summarizes these results giving first estimates for the vertical distribution of mesopelagic fishes. This study indicated that. (1) Most of the collected biomass occurred within the upper 500 m; average biomass between 500 and 1000 m was comparatively low on a gramper-cubic-meter basis, (2) During the day, highest catch number (g/m3) usually occurred at mid-depths (200-500 m), whereas, during the night, there was a large increase in the number and biomass collected in the upper 200 m. These differences are evidence that a large portion of the biomass is involved in vertical migration from mid-depths into upper waters. There was no apparent difference in the day and night biomass collected below 500 m. (3) A higher biomass per square meter of surface (down to 1000 m) collected at night appeared to be due to the larger average size of fish captured in the night collections. C. Radioecology of Oceanic Animals Studies of the radioecology of oceanic animals off Oregon were the joint result responsible of AT(45-1)1726 and AT(45-1)1750. for the The former contract was collection of samples necessary for radioanalyses. As a result of this cooperative effort, a large number of macroplankton (euphausiids, salps, shrimps, copepods) and fishes) Reprints animals, including and nekton (squids, have been studied by gamma-ray spectrometry (Sections 6 & 7; 4, 5, and 6), 9 A Some of the results of these studies are included in this report. preliminary study of the vertical distribution of Zn65 and Zr95-Nb95 from oceanic animals collected at different depths is found in Section 5. This section indicates that the amount of Zr95-Nb95 in the animals collected decreased rapidly with depth, The Zn65 content, on the other hand, was nearly the same in animals from all depths within the upper 1000 m, Calculation of the amount of Zn65 in the animals per 1000 m3 of water filtered illustrated a definite flux of radiozinc into surface waters during the night, This movement indicates the importance of the vertical migration of animals in affecting the distribution of this radionuclide off Oregon, Comparison of the gamma-ray spectra from various levels of the oceanic food chain (Section 6) revealed that fission products, such as Zr95-Nb95 and were concentrated only by herbivores and were dis- criminated against by the carnivores, This conclusion is further supported by the fact that the Zr95-Nb95 content of deep water animals, which are mainly predators, was also low (Section-5). Seasonal and geographic variations in the.Zn65 content of Euphausia pacifica are discussed in.Section 7. Noteworthy is the conclusion that in spite of seasonal differencesinthe position'of the Columbia River plume, the radiozinc concentration in euphausiids remained fairly constant throughout the year. uncertain. Th reasons for this stability are at present Diurnal migration of euphausiids, seasonal changes of currents, and a long biological half-life of Zn65 causes. are suggested as contributory PRELIMINARY OBSERVATIONS ON THE DISTRIBUTION OF MESOPELAGIC FISHES OFF OREGON by William G. Pearcy ABSTRACT Over 200 collections made throughout the year with a midwater trawl to various depths down to 1000 m and along three latitudes off Oregon provided preliminary data on species composition, sampling variability, daily vertical migrations, depth distribution, and seasonal and geographic variations of mesopelagic fishes. About 40 species of mesopelagic fishes were collected. Half the number of species were present in 0-200 m collections, half in deeper collections. Those frequently occurring only in collections to 500 or 1000 m are listed as lower mesopelagic fishes. Those found above 200 m at night, or upper mesopelagic fishes, dominated most collections. were largely the lantern fishes. These Differences in upper depth distributions of the dominant lantern fishes were evident at night; all penetrated the halocline, and some crossed the density gradient of the thermocline as well. Although no clear seasonal changes in species composition were apparent high catches of dominant fishes during the summer and low catches during other seasons suggested seasonal changes in relative abundance, perhaps due to movements across the continental slope. Mesopelagic fishes were common over the continental slope but were rare over the shelf, that depth limits their horizontal distribution. The diversity indicating of meso- pelagic fishes, as well as the catches of several species, increased from north to south, 12 INTRODUCTION Knowledge of the ecology of animals of the open oceans is very limited. This is particularly true of the small nektonic organisms, such, as fish, squid, and shrimp that are intermediate in the food web between small plankton and larger carnivores. ubiquitous in the oceans of the world. These micronektonic animals are As nekton, they are capable of sustained movements, independent of currents, in horizontal or vertical directions. The occurrence of vertical migrations of micronekton has been well documented since the CHALLENGER Expedition (Brauer, 1906; Murray and Hjort, 1912; Beebe and Vander Pyl, 1944; Tucker, 1951; Bainbridge, 1961; and others). Horizontal movements, on the other hand, although described for many of the larger epipelagic nekton, have not been adequately studied for the smaller mesopelagic nekton. Mesopelagic animals are de- fined as those distributed between 200 m and 1000 m depth during the day. (For classifications of epipelagic, mesopelagic and.bathypelagic zones, see Hedgpeth (1957).) The studies of mesopelagic fishes by Taning (1918). Barham (1956), and Fast (1960), revealed seasonal differences in the catches of certain species of lantern fishes that suggested' horizontal movements of these populations. Whether such movements are of a general nature for mesopelagic animals, how these movements are affected, how far they extend, and how they are related to the ecology of the populations are questions of real interest to the biological oceanographer. A purpose of this study on the distribution of midwater fishes off Oregon is to examine possible changes in the catches of mesopelagic fishes which may reflect seasonal movements across the continental slope, that 13 transitional area of the oceanic region the neritic bility, region is gross features approached. where depth decreases rapidly as Species composition, sampling varia- of depth distribution and vertical migrations are also discussed. Aron (1959, 1962) conducted extensive midwater trawling studies in the eastern North Pacific and contributed much to our knowledge of the zoogeography of oceanic animals. His collections extended over a wide geographic area but were limited to shallower depths and the summer and fall seasons. The present study entails systematic sampling of a rela- tively small area of the Pacific Ocean to greater depths on a year-round basis; it represents the largest number of collections to depths below 500 m and during the winter season for this area of the Pacific. No previous studies have surveyed this area. In view of their swimming capacity, nektonic animals may be important agents in the distribution and transport of radioisotopes. Detectable quantities of radioisotopes, such as zinc-65 induced in low levels by the Hanford nuclear reactors on the Columbia River, have been found in micronekton such as lantern fishes, sergestid prawns, and euphausiids off Oregon (Osterberg, 1962; Osterberg, et al., in press). These micronekton are capable of crossing density. gradients such as the thermocline and halocline, which normally inhibit mixing by physical processes. migrations of these animals may make them available as forage Inshore for commer- cially important species, which in turn may make these radioelements available to humans. If the open oceans are used for disposal of radio- active materials, obviously a more comprehensive understanding of nekton ecology and behavior is essential (Ketchum, 1960). 14 METHODS Micronekton were sampled with a six-foot Isaacs-Kidd midwater trawl (Isaacs and Kidd, 1953; Aron, 1962), A total of 228 collections were made on cruises between June 1961 and August 1962. Collections were made during various seasons of the year at stations located 15, 25, 45, 65, and between 85 and 165 miles offshore along three parallels of latitude off (1) the mouth of the Columbia River (35 collections), (2) Newport (46 collections), and (3) Coos Bay (29 collections) (Fig. 1). An oblique tow to 200 m depth was made at each station with the midwater trawl. The net was lowered rapidly until 730 m of wire were out and then retrieved at a constant speed of 30 meters per minute while steaming on a given compass course at six knots, Where the depth of the bottom would not permit tows to 200 m, shallow tows were made with a reduced rate of retrieval and an increased towing speed. Thus, the speed of the net through the water and the total time for the tow were approximately the same regardless of the depth of the water, along the three east-west series were made at night. All collections Usually the inshore stations (15-65 miles offshore) were sampled during one evening, and.the remaining station was sampled the following night. One hundred eighteen tows were made at a station 50 miles off Newport over the outer edge of the continental slope (encircled station in Fig. 1). These tows included (1) repeated tows in the upper 200 m over periods varying from 6 to 48 hours to evaluate sampling variability and daily changes in relative abundance, and (2) successive tows to.various depths, generally 200, 500, and 1000 m, to gain information on vertical distribution, I 126° 124° WASHINGTON ` ASTORIA OREGON NEWPORT CALIFORNIA 128° Figure 1. 126° 124° Location of midwater trawling stations off Oregon. Numbers designate the distance in miles from the, coast. The circle 50 miles off Newport, Oregon, includes the location of replicate tows and tows to various depths. l" The relationship between the maximum depth of the trawl and the length of the towing wire was determined with either a bathythermograph or depth gauge attached to the The results, given trawl. below, showed that a wire length of approximately four times the desired depth was required. maximum depth m wire desired depth 730 2000 4000 200 m 500 m 1000 m No, obs. 39 3 3 .(average) maximum depth variation Standard Deviation = 8.1 193 533 1007 430-600 (range) 960-1080 (range) For tows to 500 m the trawl was fished between 500 and 200 m (2000-730 m- wire) for about 30 minutes and for twos to 1000 m between 1000 and 500 m (4000-2000 wire) for one hour. Upon reaching the upper depth interval, the trawl was retrieved at the rate of 50-70 meters of wire per minute. All collections were made from the R/V ACONA. Since the main winch is located forward, the wire was secured in a towing block on the stern to facilitate a constant heading while towing. Geographic position at the start and end of each tow, total duration of the tow, course, speed, and sea and weather conditions were recorded for all collections. Samples were preserved with formalin at sea. Later, all fishes and other nekton were sorted, identified and measured in the laboratory ashore. SPECIES COMPOSITION Over 40 species of fishes were identified from the collections (Table 1). Mesopelagic fishes dominated the catches in both number and variety. Myctophidae, Melanostomiatidae, and Gonostomatidae were most abundant. Three species of Myctophidae, or lantern fishes, accounted for 76 percent of the total catch: Lampanyctus leucopsarus (45 percent), 16 Diaphus theta (21 percent) and Tarletonbeania crenularis (10 percent). Both L. leucopsarus and D.,theta occurred in over 80 percent of all collections (excluding day tows within the upper 200 m and tows in neritic waters). Tactostoma-macropus, a melanostomiatid, composed approximately 8 percent of the total catch. These fishes represented the four most abundant species found in our collections. Although numerically unimportant, epipelagic fishes were also collected, Included were such oceanic species as the saury, Cololabis saira, and neritic species as smelt, cod, etc. However, relatively few collections were made over the continental shelf, and fishes over the slope, especially those migrating into the upper 200 m at night, were sampled preferentially. Although numerous larval fishes were collected, they are not considered in this paper. DAY-NIGHT VARIATION AND SAMPLING VARIABILITY Variations in the numbers of the four dominant mesopelagic fishes collected during night and day periods are illustrated in Figure 2. The positions of all these collections are encompassed by the circle in Figure 1, Diurnal (diet) differences are obvious; catches of these fishes within the upper 200 m were high during the night compared with the day, While differences between day and night catches were clear, there was no evident trand in the catches of fishes during the night that indicated major variations associated with time. Since catches soon after sunset or before sunrise were not consistently lower than those made around midnight, it is believed that fish ascended quickly to the upper 200 m shortly after sunset and remained within this region until shortly NIGHT DAY 0400 HOUR 0800 1200 0000 2000 1600 0400 vffiz 13-14 July 1961 T. macropus 0 1 0 0 26 0 0 0 0 0 0 0 5 0 L. leucopsarue 0 T. crenularis D, theta 3 2 0 5 14 5 1 0 0 0 0 10 2 3 1 1 1 0104 17-18 July 1961 T. macropue L. leucopsarus T. crenularis D. theta 0 0 3 2 1 1 0 8 5 6 0 0 4 30 1 0 0 3 13 9 3 1 1 15 25 9 7 11 3 8 11 2 23-24 Jan, 1962 T. macropue T. D. 0 4 0 2 3 5 0 1 1 0 3 2 5 3 0 L. leucopsarus crenularis theta 0 24-25 Jan. 1962 T. macropus 0 0 0 0 L. leucopearua T. crenularis D, theta 1 2 2 0 0 10 10 7 9 7 1 2 3 1 8 6 3 2 1 2 1 1 9 6 8 8 2 0 15 2 4 4 9 2 3 7 4 7 5 5 1041 April 1962 T. macropus 0 L. leucopearus 0 5 2 0 0 2 9 0 0 0 0 2 1 2 T. crenularis .D. theta 11-12 April 1962 T. macropue L. leucopsarue T. crenularis D. theta Figure 2. 1 9 3 10 7 12 4 9 2 7 5 9 2 6 2 0 2 1 1 1 0 2 1 1 4 3 2 2 2 1 2 5 2 10 10 10 Catches of the four dominant mesopelagic fishes in the upper 200 m 50 miles off Newport, Oregon. 17 Table I. Fishes collected during midwater trawling studies off Oregon MES0PELAGIC Bathylagidae Bathylagus ochotensis Schmidt, 1938 B. milleri Jordan and Gilbert, 1898 B. pacificus Gilbert, 1890 Opisthoproctidae Macropinna microstoma Chapman, 1939 Bathylychnops exilis Cohen, 1958 Alepocephalidae Talismania bifurcata (Parr, 1951) Searsidae Holtbyrnia polycoeca (Parr, 1937) Idiacanthidae Idiacanthus antrostomus Gilbert, 1890 Myctophidae Ilierops crockeri (Bolin, 1939) H. thompsoni (Chapman, 1944) Myctophum californiense Eigenmann and Eigenmann, 1889 Tarletonbeania crenularis (Jordan and Gilbert, 1880) Diaphus theta Eigenmann and Eigenmann, 1890 Lampanyctus nannochir (Gilbert, 1890) L. leucopsarus (Eigenmann and Eigenmann, 1890) L. ritteri Gilbert, 1915 L. regalia (Gilbert, 1891) Ceratoscopelus townsendi (Eigenmann and Eigenmann, 1889) Scopelarchidae Neoscopelarchoides dentatus Chapman, 1939 Gonostomatidae Cyclothone signata Garman, 1899 C. microdon (Gunther, 1878) C. pallida Brauer, 1902 Paralepidae C. acclinidens Garman, 1899 C. spp. Danaphos oculatus (Garman, 1899) Cetomimidae Lestidium ringens (Jordan and Gilbert, 1881) 1 sp. Sternoptychidae Argyropelecus lynchus Garman, 1899 A. intermedius Clarke, 1877 A. pacificus Schultz, 1961 Melanostomiatidae Bathophilus flemingi Aron and McCrery, 1958 Tactostoma macropus Bolin, 1939 Malacosteidae Aristostomias scintillans (Gilbert, 1915) Nemichthyidae Nemichthys scolopaceus Richardson, 1848 Anoplogastridae Anoplogaster cornuta (Valenciennes, 1833) Melamphaidae Poromitra crassiceps (Gunther, 1878) Oneirodidae 2 spp. Chauliodontidae Chauliodus macouni Bean, 1890 NERITIC ------- EPIPELAGIC ---------- Petromyzontidae Entosphenus tridentatus (Gairdner, 1836) A N D O THE R S Scorpaenidae several spp. Engraulidae Engraulis mordax Girard, 1854 Cottidae Scorpaenichthys marmoratus (Ayres, 1854) Osmeridae Agonidae Agonopsis emmelane (Jordan and Starks, 1895) Thaleichthys pacificus (Richardson, 1836) Scomberesocidae Cololabis saira (Brevoort, 1850) Gad idae Microgadus proximus (Girard, 1854) Coryphaenoididae 1 sp. Icosteidae Icosteus aenigmaticus Lockington, 1880 Liparidae Nectoliparis pelagicus Gilbert and Burke, 1910 Anarrhichadidae Anarrhichthys ocellatus Ayres, 1855 Zoaricdae Lycodapus mandibularis Gilbert, 1915 18 before dawn. Therefore, though fishes may have migrated vertically within the upper 200 m region during the night, all tows during this period were By assumed to have sampled the same mesopelagic population of fishes. assuming that these night tows represent replicate samples, a basis is provided for estimates of sampling variability, This is useful to eval- uate subsequent spatial and temporal differences and to gain insight on the patchiness of distribution of mesopelagic fishes, Figure 3 shows the average catch of each of the four species plotted against the respective variance (calculated from the data given in Figure 2, excluding series in which less than four night collections were made). If the distribution were random (Poisson) the variance would be approximately equal to the mean, and points would be grouped near the 450 diagonal, dispersion, To estimate the departure from randomness, coefficients of s?/x (Blackman, 1942) were calculated, and the significance of their deviation from unity compared to the expression 1 2 2n (n-1)2 where n is the number of samples (Holme, 1950). The large range of the 16 coefficients calculated for these four species (from 0,3 to 12,3) indicates a lack of a consistent dispersion pattern. Eleven coefficients of dispersion did not significantly depart from unity and are clustered near the 450 diagonal; although this suggests a random distribution, these occur at low population densities where the sample size is probably too small for reliable estimates of the actual distribution (Cassie, 1959), Five coefficients were significantly greater than would be ex- pected from a randomly distributed population, These large values 99 115 5.7 A CO N 0 / / / / O T. macropus L. Ieucopsarus L T. crenularis D. theta T 5 10 15 20 - - 25 MEAN (x) Figure 3. The variability of catches of the four dominant mesopelagic fishes. The 45° dashed line denotes random (Poisson) distribution. 19 occurred when average catches were high, suggesting aggregation (over-dispersion) at high densities for all four species. A noteworthy example of such patchy distribution was the catch of 95 T. crenularis in one tow, compared with the usual 2 or 3 per collection, Aggregation at high densities has been reported for many including marine plankton (Barnes and Marshall, natural populations, 1951). These indications of aggregations distribution of the other, may reflect the actual spatial animals, but it is also possible that inconsistencies in sampling, such as failure to sample all depths equally, may lead to such conclusions. Since aggregation and high densities were both appa- rent only from the summer catches, seasonal changes in spatial distribu- tion, perhaps related to schooling or breeding activity, are suggested. Although there is insufficient data on schooling and spatial distribution of mesopelagic fishes, small schools of lantern fishes have actually been observed from a bathysphere by Beebe (1934) or a bathyscaphe by Peres (1958), Variability of catches of certain myctophids off California was.. also attributed to their schooling behavior by Beebe and Vander Pyl (1944). Aron (1962) reported that the difference in the total number of fishes caught between repeated midwater collections was always less than a factor of two. This suggests a more random distribution of fishes and much less sampling variability than noted in our study above. The apparent disparity between the two studies may be because different areas were sampled and different sampling techniques were used (Aron fished his trawl at one depth for 30 minutes). Also the total catch of fish may vary less than the catch of individual species. 20 DEPTH DISTRIBUTION Although opening-closing devices are necessary for a detailed analysis of depth distribution, some generalizations are possible from a study of collections with non-closing nets to successive depths or during periods of daylight and darkness, Depth distribution of the catches of three different lantern fishes (Fig, 4) illustrates the percentage of the total, number of a species collected in tows to various depths, Depths indicated are depths to which tows were made and not necessarily depths at which fish were captured, Despite these limitations, the data clearly demonstrate dif- ferences in vertical distribution for different species. For example, most Tarletonbeania crenularis were collected at the surface and were less abundant in tows below 10 m depth, This species, incidentally, is commonly collected with dip nets under night-lights off Oregon, On the other, hand, neither Diaphus theta nor Lampanyctus leucopsarus were cap- tured at the surface but were found in highest numbers below 10 m; D. theta was most abundant in tows to 10-25 m and L, leucopsarus in tows to 25 -30 m., Corroborative data on the upper depth distribution of these species at night are given by other workers, For example, Aron (1959, 1962) noted peak abundance of L, leucopsarus and D, theta below 30 m off the coast of Washington and in regions south of 50°N, but within the upper 30 m in more northerly regions; he also observed T. crenularis at the surface under night-lights, Tucker (1951) caught numerous L. leucopsarus in a night tow to 37 m in the Bering Sea, and Fast (1960) considered that the upper range of the adults of this species was about 50 m in 0 10 20 30 40 - 50 a- W 100 Figure 4. Depth variations in the catches of three lantern fishes at night, 50 miles off Newport, Oregon, as a percentage of the total catch of each species (number in parentheses). 21 Monterey Bay, California, Temperature and salinity profiles (Fig. 5) for the area where collections were made to successive depths show a thermocline located between 10 and 20 m and a halocline located between the surface mixed layer and 100 m. The halocline, usually with its base at about 200 m depth, is a permanent The oceanographic feature in this area of-the Pacific (Fleming, 1958). thermocline is well developed only in the summer (Tully, et al., 1960; These density gradients result in high stability of the Tabata, 1961). water above 200 m. Nevertheless, the dominant mesopelagic fishes migrate through the base of the halocline and into the upper waters during the night (see Fig. 2). D. theta and T. crenularis apparently migrate through the thermocline as well, Thus these gradients do not necessarily act as barriers to the mesopelagic fishes which swim vertically through, at least one density gradient during their daily migrations into near surface waters at night. Depth distribution within the mesopelagic region was examined using data from successive tows to 200, 500 and 1000 m. Half the total number of species of mesopelagic species was collected in night tows from 0-200 m. These fishes that penetrated the halocline and invaded epipelagic waters at night are considered upper mesopelagic species; they were largely Myctophidae. Species which were mainly restricted to tows to 500 m or below and were uncommon above 200 m are considered lower mesopelagic species; they were mostly Gonostomatidae, Chauliodontidae, Bathylagidae, and some Myctophidae. Some common lower mesopelagic species are listed in Table 2, which gives numbers caught in tows to the different depths. Note that 22 Table 2. Catches of Some Lower Mesopelagic Fishes at the Stations 50 miles off Newport, Oregon. Depth of Tows: Number of Tows: 0-200 m (46) 0-500 m (17) 0-1000 m (12) Bathylagus milleri 0 5 4 Bathylagus pacificus 0 3 40 Searsidae 0 1 5 Cyclothone microdon 0 44 152 Cyclothone signata 0 119 119 Chauliodus macouni 11* 43 41 Lampanyctus nannochir 0 2 20 Lampanyctus regalis 1 7 12 Neoscopelarchoides dentatus 0 4 7 Poromitra crassiceps 0 1 14 * Mostly small specimens. TEMPERATURE °C 6.0 8,0 10.0 12.0 34.00 32.00 140 16.0 3600 SALINITY %bo Figure 5. A temperature and salinity profile, 50 miles off Newport, Oregon, August, 1961. 23 relatively few individuals were taken within the upper 200 m, notwithstanding the much larger number of collections in this upper region. Single specimens of Talismania bifurcata, Macropinna microstoma, and unidentified species of Cetomimidae and Oneirodidae were collected in tows to 1000 m and are thought to be lower mesopelagic forms. More collections of these and other species are needed with opening and closing nets to establish vertical ranges and changes in depth distributions during day and night periods. GEOGRAPHIC AND SEASONAL VARIATIONS Collections of mesopelagic fishes along the three latitudinal series of stations in the open ocean off Oregon revealed a similar species composition during all seasons of the year. L. leucopsarus, D. theta, T. crenularis and T. macropus predominated in nearly all collections within the upper 200 m at night regardless of season of year or latitude. Thus, a single community of upper mesopelagic fishes was suggested by the absence of drastic changes in the occurrence of species. Some differences in distribution were apparent, however. trends were related to latitude. Several The total number of mesopelagic species collected increased at the stations to the south (Table 3), indicating a higher diversity of the mesopelagic fish community at the lower latitudes off Oregon. Moreover, the relative abundance and frequency of occurrence of several species of fishes were clearly higher off southern Oregon than off northern Oregon. These data suggest that Oregon is the northern limit and zoogeographically a transitional area for some mesopelagic animals (see also Aron, 1959, 1962). 24 Table 3. (A) Total Number of Mesopelagic Species Collected and (B) The Number/Occurrence of Four Species at the Three Series of Stations off the Oregon Coast. Columbia River (A) Total Number of Species Newport Coos Bay 10 13 17 (26) (46) (29) (B) Bathylagus ochotensis 2/1 9/8 23/14 Hierops crockeri 3/3 7/4 30/14 0 1/1 4/3 Number of Tows Myctophum californiense Lampanyctus ritteri 22/10 33/15 70/20 25 Collections along parallels of latitude across the continental slope permit an examination of possible changes in the diversity of mesopelagic fishes with increasing distance from shore. In general both the total number of species and the average number of species per collection were lowest at the inshore stations and highest at an intermediate distance from shore (Table 4). Since Aron (1959) found that the number of species of fishes decreased in oceanic water with distance from the Washington coast, together these two studies suggest that the greatest diversity of mesopelagic fishes may occur.,4over or just beyond the outer continental slope. Mesopelagic fishes were usually collected from the inshore stations, A across the continental slope, to the stations over 100 miles offshore. notable exception, however, was the rarity of mesopelagic species both in number and kind, at the inshore stations off Newport (see Table 4 and Fig. 6). Although a considerable number of fishes were caught at stations located 15 and 25 miles from shore along the northern and southern series, they were rarely caught at the same distances off Newport despite numerous collections. The salinity at these stations off Newport is not greatly modified by freshwater runoff; in fact, the inshore stations off the Columbia River were most neritic in regard to reduced surface salinity. In addition, mesopelagic fishes were absent from the inshore stations off Newport even during periods of upwelling when the characteristics of the water near the surface were more typical of-deeper, offshore waters. An obvious difference among the inshore stations is the depth of water. Off Newport the water is comparatively shallow. Here the 200 m depth contour occurs about 25 miles offshore, but it is found inshore of the 15-mile 26 Table 4. Total Number of Species and the Average Number of Species per Tow Collected at Various Distances from Shore off Oregon. Distance Offshore (n.m.) Columbia River Newport Total Av./Tow Total Av./Tow Coos Bay Total Av./Tow 15 3 1.8 1 0.1 7 4.0 25 8 3.3 3 1.1 8 5.4 45 7 4.5 10 4.8 13 7.0 65 6 4.2 11 4.8 6 5.5 65 4 4.0 9 5.2 11 5.7 Diaphus Lam panyctus leucopsarus 46. 1 I'M i NEWPORT COOS BAY 44-37'N 43' 2I' M 46' COOS SAY NEWPORT ASTORIA - COLUM81A R1"E.R ASTORIA-COLUMBIA RIVER 45. 2:'N 44' 37 N N 28 23 14 JULY AUG NOV DEC. 3 0 1- JULY 12 43 24 16 C B e = 12 2 0 0 0 3 DEC 0 7 I- 5 5 w - 0 1 2 FEB. 0 iI I MAR. to 2 0 4 1 8 -S 2 0 UJ ! FES 7 C 0 0 8 7 JAN 7 7 UJ 0 I 5 2 0 a L. I 16 1, 2 13 110 6 NOV 84 JAN. 0.1 AUG 22 LU W 52 20 22 37 Q 7 6 0 O e 0 0 0 O O 35 W APR. p APR 3 3 3 2 7 MAY 0 6 MAY 0 is 14 JUNE 1 O 2 JUNE 3 12 JULY 0 0 JULY 13 0 23 22 15 AUG. 2 25 45 65 85-165 0 15 0 25 AUG 45 65 85-165 MILES FROM SHORE 0 IS 25 45 65 85-165 0 15 25 45 65 85-165 0 t5 25 45 65 85-165 MILES FROM SHORE Figure 6a. Average monthly catches of Lampanyctus leucopsarus at stations various distances from shore along three latitudes off Oregon. b. Average monthly catches of Diaphus theta at stations various distances from shore along three latitudes off Oregon. 0 15 25 45 65 95165 27 Thus, depth appears station at the latitudes of the other station series. to be important in limiting the inshore distribution of these fishes, perhaps by affecting vertical migrations so that preferred light intensity is simply not available during the day when the depth is less than 200 m, a depth which interestingly conforms to the arbitrary division between oceanic and neritic provinces of the ocean, This is evidence that these fish are truly mesopelagic in distribution. In order to examine seasonal changess,irt the distribution and abundance of mesopelagic fishes, catches of the most common species were plotted for various months of the year. These are illustrated for Lampanyctus leucopsarus and Diaphus theta in Figures`).6a and 6b. at inshore stations off Newport, Besides low catches seasonal variations are apparent. catches are usually highest during the summer. summer periods that were sampled and for all The This was the case for both latitudes, with the exception of the northern stations where peak catches of L. leucopsarus occurred during the winter. The number of fish collected over the continental slope during the summer was compared with that for the other months,-after use of an X + 1/2 transformation (Bartlett, 1947). The average catch was significantly higher during the summer khan during tie other seasons for D. theta at all latitudes and for L. leucopsarus off Coos Bay (Table 5). Such variations in numbers could be produced by selectivity of the trawl combined with seasonal changes of population size-structure from recruitment or mortality. A comparisQn between the size-frequency distri- butions for summer and winter periods of both D. theta and L. leucopsarus, however, indicated that fish of various sizes contributed to the high summer catches and that they were not solely a result of increased 28 Table 5. Comparison of Average Summer (July-Sept.) Catches With Average for Other Months for Two Species. Data are Included From Stations 15-65 Miles Offshore, Where Depth of Water Exceeds 200 m, at Three Latitudes off Oregon. t L. leucopsarus D. theta .98 3.02 d.f. P 21 >0.3 21 ( .01 Coos Bay Newport Columbia River t d.f. t 1.49 20 >0.1 2.49 20 <.05 d.f. P 3.41 22 <.01 3.79 22 <.01 29 vulnerability of young fish. Seasonal differences in bathymetric distri- bution may also affect catches if varying proportions of the populations migrate vertically into the upper 200 m during different seasons of the year. Such a trend was not evident from the collections to 1000 m during various seasons. As data is limited, neither seasonal changes in popula- tion size-structure nor depth distribution can be excluded from the analysis of these catch fluctuations, and both should be studied more thoroughly as they may reveal salient features of the oceanic ecology of these animals. Nevertheless, they do not appear to furnish a complete explanation to the observed seasonal variations. Hence, horizontal movements of the popula- tions may also be important. Only a few other studies on mesopelagic fishes have permitted comments on seasonal changes in distribution. Those of Barham (1956) and Fast (1960), which were conducted in Monterey Bay, California, are of special interest since they were concerned with the species which are also common off Oregon. Barham noted marked seasonal variations in the pattern of sonic scattering layers and in the catches of certain midwater animals. For ex- ample, Diaphus theta, collected in high numbers in mid-winter when it was thought to be a dominant sound scatterer, disappeared in the spring and reappeared in the summer. Fast reported high catches of Lampanyctus leucopsarus in Monterey Bay in the winter with a reduction of the population by about one-half between February and May. This was followed by a subsequent increase in the first two age groups to high numbers by the following February. Finally, Taning (1918) found that large individuals of Lampanyctus maderensis were absent from the eastern Mediterranean in the summer but were common during other seasons. These observations suggest 30 that seasonal changes in distribution of mesopelagic fishes may be influenced by population movements. Horizontal movements may occur in two ways: passively, by transport with prevailing currents; or actively, by swimming of the micronekton independently of currents. Seasonal variations in depth distribution of these mesopelagic animals, as observed by Taning (1918) and Barham (1956), could result in translocation of the populations if current velocities or directions vary at different depths. The importance of vertical distri- bution in influencing horizontal transport of plankton and micronekton is well recognized (Hardy and Gunther, 1935; Mackintosh, 1937; Brunn, 1958). To explain the fluctuations in number of L. leucopsarus in Monterey Bay, Fast (1960) elaborated a hypothesis based on the seasonal oceanographic periods and associated changes in current directions. His highest catches were made during the winter when prevailing winds are from the southeast and the northward flowing Davidson Current is found along the coast of California and Oregon. The major reduction of the population occurred in the spring after surface winds had shifted to a northwesterly direction and upwelling was evident. Fast attributed the high relative abundance to the concentrating effect of the Davidson Current and the low relative abundance the the dissipating effect of offshore drift during the upwelling period. Since he stated that the major differences between these two periods were due to surface currents, the lantern fish presumably migrated vertically into upper waters and resided there long enough to be transported by these superficial currents. This hypothesis of passive transport of mesopelagic fishes was not supported by the results off Oregon, where the highest catches appeared 31 during the summer, which is the period of upwelling, and lowest catches The fluctuations were not similar to appeared during the other seasons. those found by Fast either in respect to time of year or oceanographic periods. Other hydrographic differences between these areas (see Sverdrup, et al., 1942) may be involved. The collections in Monterey Bay, California, were made over a submarine canyon (a depth of about 900 fathoms), which may act as a concentrating basin (Fast, 1960), whereas the collections off Oregon were made in the waters off the unprotected coast, often where the depth was less than 900 fathoms. Only some northern stations off the Columbia River were over a submarine canyon. Clearly, before the effect of currents on the distribution of these animals can properly be assessed, more knowledge is needed on the seasonal variations of currents and water masses at all depths within the vertical ranges of these species. It is possible that active migrations of these micronekton may be a cause of the seasonal fluctuations. Virtually nothing is known about lateral movements of small oceanic animals. In general, extensive geo- graphic migrations of nektonic animals on a seasonal basis as evidenced for larger epipelagic fishes (e.g., Sette, 1950; Neave and Hanavan, 1960; Clemens, 1961) are unexpected for mesopelagic micronekton because of their relatively small size and slow swimming speed also because seasonal variations of physical factors, such as temperature, are only pronounced in the epipelagic zone. Small scale movements, however, across the continental slope may be entirely feasible. 32 ACKNOWLEDGMENTS The author is grateful to William Aron, M. Laurs, C. E. Bond, D. M. Cohen, R. L. Wisner, B. N. Kobayashi, R. H. Gibbs, Jr., and A. Ebeling who aided in identification of fishes; to L. D. Calvin for suggestions on statistical procedure, and to L. Hubbard and the captain and crew of the R/V ACONA for conducting the trawling operations at sea. This study was sponsored by the Atomic Energy Commission, Contract No. AT(45-1)1726. Ship operations were supported by grants from the National Science Foundation and Office of Naval Research. REFERENCES Aron, William. 1959, Midwater trawling studies in the North Pacific. J. Limnol. Oceanogr. 4:409-418. ------- 1962. The distribution of animals in the eastern North Pacific and its relationship to physical and chemical conditions. J. Fish. Res. Bd. Canada 19:271-314. Bainbridge, Richard, 1961. Migrations. In: Waterman, T. H. (Ed.) The physiology of crustacea Vol. II, Academic Press, New York. Barham, E. G. 1956. The ecology of sonic scattering layers in the Monterey Bay area, California. Doctoral Dissert. Ser. Publ. No. 21, 564, University Microfilms, Inc., Ann Arbor, Mich., 182 pp. Barnes, Harold and S. M. Marshall. 1951. On the variability of replicate plankton samples and some applications of "contagious" series to the statistical distribution of catches over restricted periods. Biol. Assoc. 30:233-263. J. Mar. 33 S. 1947. The use of transformations. Beebe, William. 1934. Half mile down. Bartlett. M. Biometrica 3:39-52. Harcourt, Brace and Co., N.Y., 344 pp. ------, and Mary Vander Pyl. 1944. Eastern Pacific expeditions of the Pacific Myctophidae (Fishes). New York Zoological Society, XXXIII. Zoologica 29:59-95. Blackman, G. E. 1942. Statistical and ecological studies in the distriAnn. Bot. N.S. 6:351-370. bution of species in plant communities. Brauer, August. 1906. Wiss. Ergebn. Brunn, A. F. Die Tiefsee-Fishce. I. VALDIVIA, XV:1-420. On the restricted distribution of two deep-sea fishes, 1958. Borophyrne a o on and Stomias colubrinus. Cassie, R. M. Systematischer Teil. J. Mar. Res. 17:103-112. Microdistribution of plankton. 1959. N. Z. J. Sci. 2: 398-409, Clemens, H. B., 1961. The migration, age and growth of Pacific Albacore (Thunnus germo), 1951-1958. Calif. Dept. Fish and Game, Fish. Bull. 115:1-128. Fast, T. N. 1960. Some aspects of the natural history of Stenobrachius leucopsarus Eigenmann and Eigenmann. University Microfilms, Inc., Mic. 60-6729, Ann Arbor, Mich., 107 pp. Fleming, R. H., 1958. Pacific Ocean. Notes concerning the halocline in the northeastern J. Mar. Res. 17:158-173. Hardy, A. C. and E. R. Gunther. 1935. The plankton of the South Georgia whaling grounds and adjacent waters, 1926-27. 1-146. Discovery Rept. 11: 34 Hedgpeth, J. W. Hedgpeth, J. Vol. I. Holme, N. A. Classification of marine environments. 1957, S. In. (Ed,) Treatise on marine ecology and paleoecology, Geol. Soc. Amer. Mem. 67. 1950, - a Population-dispersion in Tellina tenuis Da Costa. J. Mar. Biol. Assoc. 29: 267-280. Isaacs, J. D. and L, W. Kidd, 1953. Isaacs-Kidd midwater trawl. Scripps Instit. Oceanogr, Ref. 53-3, 21 pp. Ketchum, B. H. Oceanographic research required in support of 1960, radioactive waste disposal. Disposal of radioactive wastes. In: Int. Atomic Energy Agency, Vienna, 1960. Mackintosh, N. A, plankton, The seasonal circulation of Antarctic macro 1937. Discovery Rept, 16:365-412. Murray, John and Johan Hjort, 1912. The depths of the ocean. London, Macmillan, 1-821 pp. Neave, Ferris and M. G. Hanavan, 1960, Seasonal distribution of some epipelagic fishes in the Gulf of Alaska region. J. Fish. Res. Bd. Canada, 170221-233. Osterberg, Charles, 1962. Zn65 content of saips and euphausiids. Limnol. Oceanogr. 7:478-479, ------, W. G. Pearcy and H. C. Curl, Jr. relationship to oceanic food chains. Peres, J. M. 1958, In press, Radioactivity and its Submitted to J. Mar. Res. Trois plongees dans le canyon du Cap Sicie, effectuees avec le bathyscaphe F.N,R.S. III, de la Marine Nationale. Bull. Instit. Oceanogr. Monaco, No, 1115:1-21. Sette, 0. E, 1950, Biology of the Atlantic Mackeral (Scomber scombrus) of North America, Pt. II, U.S. Fish Wildl, Serv. Migrations and Habits. 51(49)°251-358. Fish. Bull., 35 Sverdrup, H. U., M. W. Johnson, and R. H. Fleming. their physics, chemistry and biology. Tabata, Susumu. 1961. The Oceans: 1942. Prentice-Hall, N.Y., 1087 pp. Temporal changes of salinity, temperature, and dissolved oxygen content of the water at Station "P" in the northeast Pacific Ocean, and some of their determining factors. J. Fish. Res. Bd, Canada, 18:1073-1124. Taning, A. V, 1918. and Myctophum). Mediterranean Scopelidae (Saurus, Aulopus, Chloropthalmus Rep. Danish Oceanogr. Exped. Mediterranean II, Biol. A. 7, pp. 1-154. Tucker, G. H. 1951. Relation of fishes and other organisms to the scattering of underwater sound. J. Mar. Res. 10:215-238. Tully, J. P., A. J. Dodimead, and Susumu Tabata. 1960. An anomalous increase of temperature in the ocean off the Pacific Coast of Canada through 1957 to 1958. J. Fish. Res. Bd. Canada 17:61-80. 37 SPECIES COMPOSITION AND DISTRIBUTION OF PELAGIC CEPHALOPODS FROM THE PACIFIC OCEAN OFF OREGON, 1961-1963 by W. G. Pearcy INTRODUCTION To my knowledge there are no previous reports on the cephalopods in the Pacific Ocean off Oregon. Much of our present knowledge about the species composition and distribution of cephalopods of the Pacific Ocean is the result of the cruises of the ALBATROSS, Steamer of the U. S. Fish Commission, made during the late nineteenth and early twentieth centuries. ALBATROSS collections along the west coast of North America were mainly at stations off California and Central America or off Alaska; but comparatively few collections were made off Oregon (Townsend, 1901). Neither Holye (1904) nor Berry (1912) list any cephalopods taken at ALBATROSS stations off Oregon or Washington. Clearly, data on pelagic cephalopods is desirable to enable comparisons of fauna and generalization on zoogeographic distribution, METHODS A total of 385 collections made between June 1961 and July 1963 with a six-foot Isaacs-Kidd midwater trawl from the R/V ACONA, provided most of the data on pelagic cephalopods collected off the Oregon coast. Collec- tions were made at a series of stations located 15, 25, 45, 65, and between 65 and 165 miles offshore along three latitudes off Oregon (Fig. 1) during the night within the upper 200 m (depth permitting). Generally, the 38 stations off the central Oregon coast (Newport) were sampled every month, whereas, the other stations were sampled bimonthly. In addition to the 0-200 m epipelagic collections at various distances from shore, tows were made to provide data on the depth distribution of cephalopods. Over one hundred collections were made to three successive depths(200, 500, and 1000 m) at a station 50 miles off Newport, Oregon, over the outer edge of the continental slope. (For more details on methods see Section 1,) Midwater trawl collections were preserved with formalin at sea. Cephalopods were removed, sorted, identified and dorsal mantle length (DML) measured in the laboratory ashore. Besides the midwater collections, cephalopods were collected occasionally with dip nets under lights at night. Some squid were also captured using otter trawls, RESULTS The families and species of pelagic cephalopods collected off Oregon are listed in Table 1. These include 17 species, representing 12 families. Seven of these species represent new distributional records for the northeastern Pacific Ocean, Only eight are included in Berry's (1912) "Review of the Cephalopods of western North America," a review based mainly on bottom collections, The additional species included in this paper clearly indicate the value of midwater trawling in any survey of cephalopod fauna. Although most of these species were collected in the midwater trawl, several were captured by other methods. For example, the two Moroteuthis robusta examined (DML 650 and 1350 mm) were captured in otter trawls off WASHINGTON 1- 165 85 65 45 25(15 ill, 19 AS roR I A OREGON 85 165 65 15 NEWPORT 85 165 "- 45 25 15 / 128° Figure 1. 65 /J ? COOS BAY CALIFORNIA 126° Location of midwater trawling stations off the Oregon coast. Numbers denote distance in nautical miles from shore. The circle at 50 miles off Newport includes the location of tows to various depths. 39 Table 1. List of Pelagic Cephalopods Collected from the Pacific Ocean off Oregon. Loliginidae Loligo opalescens Berry, 1911 Sepiolidae Rossia pacifica Berry, 1911 Onychoteuthidae Onychoteuthis banksi (Leach, 1817) Moroteuthis robusta (Dall) Verrill, 1876 Enoploteuthidae *Abraliopsis sp. Veranyidae *Octopoteuthis sicula Ruppell, 1844 Histioteuthidae Gonatidae Chiroteuthidae Meleagroteuthis holyei Pfeffer, 1900 Gonatus fabricii (Lichtenstein, 1818) G. magister Berry, 1913 *G. anonychus Pearcy and Voss, 1963 *Gonatopsis borealis Sasaki, 1923 Chiroteuthis veranyi (Ferussac 1835) Cranchiidae Galiteuthis armata Joubin, 1898 *Taonius pavo Lesueur, 1821 *Cranchia scabra Leach, 1817 Vampyroteuthidae *Vampyroteuthis infernalis Chun, 1903 Octopodidae Bolitaenidae Japetella heathi (Berry, 1911) * Not previously reported from the northeastern Pacific. 40 the northern Oregon or southern Washington coast in water greater than 150 m depth, This species has been reported from California (e.g., Smith, 1963) and Alaska (Dall; see Berry, 1912) but none have been reported previously off Oregon. Loligo opalescens, a common inshore myopsid of the west coast of North America, was collected in several otter trawl collections where depth was 100 m or less. It was absent from midwater trawl collections, which were mainly made offshore in deeper water. Rossia pacifica, another myopsid, was also taken mainly in otter trawl collections. Gonatus anonychus, a new species of gonatid squid, recently described by Pearcy and Voss (1963), was collected by dip netting under night-lights. The relative abundance of the cephalopods collected by midwater trawling from all stations and all depths is shown in Figure 2. Gonatus fabricii was by far the dominant species, comprising,about 38 percent of the total catch. Gonatus spp, was next in numberical importance. This group consisted of larval and juvenile individuals without sufficient differentiation of hooks for specific identification. They may comprise at least three separate species of Gonatus (see Table 1), but judging from the relative abundance of larger gonatids, are probably mostly G. fabricii. Abraliopsis sp. and Chiroteuthis veranyi were next in numerical abundance (Fig. 2). Four squids, Gonatus fabricii, Gonatus spp., Abraliopsis sp., and C. veranyi, comprised about 88 percent of the total numbers of pelagic cephalopods collected, clearly demonstrating their dominance in our midwater trawl collections off Oregon. lected, Only a few other species were col- PERCENTAGE OF TOTAL CATCH 0 1 Gonatus fabricii Gonatus s pp. 10 20 30 I I I Abra/iopsis sp. Chiroteuthis veranyi Japete//a heathi Gonatopsis borealis Octopodidae Teonius pavo Gonatus magister Ga/iteuthis armata Octopoteuthis sicu/a Onychoteuthis banksi 0 Q Rossia p aci fica Vampyroteuthis infernalis Cranchia scabra 0 Me%dgroteuthis ho/ye/ fl Figure 2. a The relative abundance of pelagic cephalopods found off Oregon in midwater trawl samples, expressed as percentage of the total number of animals collected. 41 Geographic Distribution No differences in the basic species composition of cephalopods was noted among the three latitudinal series of stations off Oregon. Similar species composition was indicated for all latitudes and most stations from 15 to 165 miles offshore. Pelagic cephalopods were rare, however, at the inshore stations off Newport, Oregon, where the depth of water than 200 m. was less A similar paucity of mesopelagic fishes was noted in neritic waters off Oregon (Section 1). Depth Distribution Some features of the depth distribution of the cephalopods collected are summarized in Table hour of towing (Table The number of dominant squid captured per 2. 2A) was greater in tows to 200 m than in deeper tows, particularly for Gonatus spp., Chiroteuthis veranyi and Abraliopsis sp. Most of these animals found in tows to 500 and 1000 m were probably Such differences caught while the trawl sampled through the upper 200 m. in catches suggest that the four dominant squids (G. fabricii, Gonatus spp., Abraliopsis sp., and C. tion. However, veranyi) are mainly epipelagic in distribu- most of the 0-200 m collections were made during the night. Comparisons of day and night catches of Gonatus fabricii and Abraliopsis (Table 2B) show that higher catches were made during the night than during the day. This indicates either daily vertical migration or better visual avoidance of the trawl during the day. these squids in tows to 500 m, on the other day. These opposite trends, hand, Catches of both were highest during the with an increase in catches at depth during 42 Table 2. Average Number of Pelagic Cephalopods Collected, per Hour of Trawling in Tows of Various Depths. 0-200 m 0-500 m 0-1000 m 72 tows 23 tows 24 tows Gonatus fabricii Gonatus spp. Chiroteuthis veranyi Abraliopsis sp. 0.5 1.2 0.5 0.7 0.5 0,2 0.1 0.1 0.1 0.1 0.1 0.1 (B) Day-night catches for two species: Day Night Day Night Species (A) Four dominant species: Gonatus fabricii 0.2 0,6 0.7 0.2 Abraliopsis sp. 0.0 0.7 0.2 0.1 (C) Some lower mesopelagic species: Japetella heathi Octopoteuthis sicula Galiteuthis armata Taonius pavo 0.03 0.00 0.00 0.01 0.21 0.04 0.04 0.14 0.14 0.02 0.05 0.06 43 the day, support the contention of diurnal vertical migrations of these species, Some of the cephalopods usually inhabited waters below 200 m. All the species listed in Table 2C were most abundant in deep collections. No large individuals were taken above 200 m even at night, suggesting that these have a basically mesopelagic (200-1000 m) distribution (also see Sasaki (1929) for G. armata). Vampyroteuthis infernalis was also taken only in deep tows to 1000 m and never in tows that went only to 500 m. Seasonal Variations Although no prominent changes in species composition were apparent over the two-year period for which samples have been analyzed, seasonal differences in relative abundance of pelagic cephalopods were striking. During the summer the number of animals magnitude higher than per tow was about an order of during the other months. Table 3 compares the average number of dominant species collected per tow for the summer months with the non-summer months. The catch of G. fabricii was about eight times higher during the summer, Gonatus and C, veranyi about six times higher, Such seasonal differences are of interest since they indicate marked changes in the availability of cephalopods. Such differences could be due to changes in susceptibility to capture or changes in actual abundance of cephalopods. In either case, the differences could be related to the life history of the squid. examined. Consequently, size-frequency distributions were 44 Table 3. Seasonal Occurrence of Common Cephalopods in 0-200 m Collections. Average Number per Tow July-Sept. Oct.-June (79 tows) (205 tows) Gonatus fabricii 4.9 0.61 Gonatus spp. 1.9 0.32 Chiroteuthis veranyi 0.71 0.20 Abraliopsis sp. 1.9 0.33 45 The size-frequency distributions for Abraliopsis (Fig. ing, They indicate that 3) are reveal- during the summer the catch of this species con- sisted mainly of small individuals (less than 30 mm the other seasons larger squid predominated. DML), whereas, during These differences indicate that breeding in Abraliopsis is probably restricted to one season. Catches of larger animals may be lower during non-summer seasons because of better escapement and lower abundance following early mortality, The relative the larval distribution of sizes of G. fabricii (assuming most of Gonatus are G. for the two periods (Fig. fabricii), on the other are very similar hand, suggesting that the summer increase in catch 4), is not merely the result of the recruitment of young. The fact that during July-September 21 percent of the animals were over 30 mm in mantle length, whereas only 7 percent were over 30 mm from October-June suggests an actual increase in the abundance of larger G. fabricii during the summer. Such variations may be due to changes in abundance caused either by ocean circu- lation or actual movement of the squid. DISCUSSION Some squid are notoriously fast swimmers; others are soft bodied and more planktonic than nektonic, Obviously such differences in swimming ability determine the catch composition by any sampling method. Many times during this study Onychoteuthis banksi (greater than 130 mm DML) were ob- served around night-lights, but they were rarely captured in midwater trawl collections made at about the same station and time. cephalopods collected in the midwater trawl were small. captured was a Gonatopsis borealis of 250 mm DML. Most of the The largest animal G. borealis and Gonatus 46 magister over 200 mm in mantle length were most common in the otter trawl collections and several large Moroteuthis robusta (up to 1350 mm DML) were taken by this method. The distribution of a species also effects its availability to sampling devices. Since most of the midwater trawl collections were made over the continental slope in the upper 200 m at night, obviously coastal, epibenthic or deep-sea species will not be adequately represented. For example, although Rossia pacifica was frequently collected with a bottom trawl over the shelf, it was rare in midwater trawl collections over the slope. Another species, Loligo opalescens, was only taken in inshore otter trawl collections. The dominant type of squid was collected by two independent sampling devices off Oregon--the midwater trawl and albacore tuna. Of the cephalopods found in the stomachs of 66 albacore collected during the summer of 1962, small gonatids also predominated. As predators often obtain effective samples of cephalopods (Clarke, 1963), this agreement was encouraging. Comparisons of the fauna found off Oregon with adjacent regions at present would be premature due to our lack of knowledge. Such comparisons, however, would be of particular interest as correlated with the oceanography of the North Pacific, As the West Wind Drift (Subarctic Current) approaches the North American continent, it divides to form the California Current and Alaskan Gyral off the coast of Oregon and Washington. Hence, the oceanic fauna from Oregon is suspected to show relationships with regions in the same current system to the west and to the south. The ocean off Oregon is a transitional region, consisting largely of modified Subarctic water. The proportion of equatorial water increases with depth and varies JULY-SEPTEMBER Abraiiopsis s p. J:ffi I0 Figure 3. ri 0--H n A= 20 30 40 SIZE (MANTLE LENGTH IN mm ) 50 Size-frequency distributions of Abraliopsis found in midwater trawl samples during the summer (July-September) and other seasons of the year (October-June). 60 25 t- Gonatus fabricii Gonotus larvae 20E a 15 - 10p OCTOBER - JUNE Gonotus 15t- fabricii Ek Gonotus larvae 10 5 0 Figure 4. 10 20 30 SIZE (mm) 40 50 60 Size-frequency distribution of Gonatus fabricii and Gonatus larvae found in midwater trawl samples during the summer (July-September) and other seasons of the year (October-June). 47 seasonally (Tibby, 1941; Rosenberg, 1962). These oceanographic features are useful in interpreting the distribution of oceanic cephalopods. For example, McGowan and Okutani (1963) reported that the two most abundant species of squid larvae off California (Abraliopsis sp, and Gonatus fabricii) were most frequently collected in waters that showed a high percentage of northern(Subarctic) origin (or a low percentage of equatorial water). These two squids, as we have seen, are also the most abundant in our catches. The capture of Vampyroteuthis infernalis off Oregon, a species predominantly found in the deep water of tropical and subtropical oceans and not known beyond 400 latitude (Pickford, 1946), also corresponds to the presence of a high proportion of equatorial waters found in deep waters off Oregon. REFERENCES Berry, S. S. 1912, A review of the cephalopods of western North America. Bull. U.S. Bur, Comm. Fish. 30:269-336. Clarke, M. R. 1963, Proc. XVI Int. Congress of Zool. 1:67. samplers, Hoyle, W. E. Information obtained by using squid predators as 1904, Reports on the Cephalopoda. Bull. Mus, Comp. Zool, 43(1):1-71. McGown, J. A, and T. Okutani, 1963, The distribution and abundance of the epipelagic decapod (Cephalopoda) larvae in the California Current. Proc. XVI Int. Congress of Zool. 1:68, Pearcy, W. G, and G. L. Voss. the northeastern Pacific, 1963, A new species of gonatid squid from Proc. Biol. Soc. Washington 76:105-112, 48 Pickford, G. E. 1946. chiate cephalopod. Vampyroteuthis infernalis Chun, an archaic dibranI. Natural history and distribution. Dana Rept. No. 29:1-40. Rosenberg, D. H. 1962. Characteristics and distribution off the Oregon coast. M.S. Thesis. of water masses Oregon State Univ. Library. 1929. A monograph of the dibranchiate cephalopods of the Sasaki, M. Japanese and adjacent waters. J. Fac. Agric. Hokkaido Univ. 20(Suppl.): 1-357. Smith, A. G. 1963. More giant squids from California. Calif. Fish and Game 49:209-211. Tibby, R. B. 1941. The water masses off the west coast of North America. J. Mar. Res. 4:112-121. Townsend, C. H. 1901, Dredging and other records of the United States Fish Commission Steamer ALBATROSS, with bibliography relative to the work of the vessel. Rept. of Comm., U.S. Commerce of Fish and Fisheries, Pt. 26:387-562. 1Fy NOTES ON THE OCCURRENCE AND DISTRIBUTION OF MACROPLANKTON OFF THE OREGON COAST A, EUPHAUSIIDS by J. F. Hebard This paper constitutes a preliminary examination of the species and distribution of euphausiids off Oregon. Euphausiid shrimp play an important role in the economy of the sea, both as food for the larger nektonic forms (fish, squid, etc.) and as consumers of the smaller plankton forms (small crustaceans and phytoplankton), Euphausiids are also known to undertake extensive vertical migrations; however, little is known about their lateral movements. often constituted the bulk of midwater trawl samples off Oregon. They Moreover, Euphausia pacifica is known to concentrate certain radionuclides and has been used as a standard in several radioecological studies (Osterberg, et al,, Section 6 and 7). Along the coast of Oregon, the environmental conditions vary during the year, Upwelling,of deep, cold oceanic water into the surface layers is brought about by the action of prevailing northwesterly winds during summer, Also, the Columbia River introduces large quantities of fresh water into the surface waters during this time of the year. It is not known what influence such environmental changes have in altering the distribution of euphausiid shrimp, Results Of the euphausiids collected (see previous sections for methods), a total of eight species were identified, Only three of these species were abundant; the other five were sporadic in occurrence and never were 50 abundant. These eight species are listed below in order of numerical abundance in midwater trawl collections. 1. Euphausia pacifica 2. Thysanoessa spinifera 3, Thysanoessa longipes 4, Nematoscelis difficilis 5. Tessarabrachion oculatus 6, Stylocheiron abbreviatum 7e Nematobrachion flexipes 8, Stylocheiron maximum Examination of the average relative abundance of the six common species in collections from the three latitudes, indicated a decrease in For some of these species numbers from south to north (Table 1), pacifica and T. spinifera) (E. there was also a trend toward decreasing abundance offshore. The catches of euphausiids varied with season. was about an order of magnitude ample, the The biomass of summer. highest during the E. pacifica summer, euphausiids, numerous in the winter than in on the other hand, was generally months, which constituted a major portion of the Hence, seasonal differences in the abundance of E. pacifica and other species may be related to annual cycles life history, in their as well as to changes in hydrographic conditions. Some general features of the distribution of these from Brinton for ex- This was mainly due to the presence of large (p 2.0 cm in length) catch during the summer more T. spinifera, (1962) are as follows: species, taken mainly 51 Table 1, Average Abundance of Euphausiids per Tow at Three Latitudes off Oregon. Columbia River Newport Coos Bay E. pacifica 2,4 3.3 3.8 T, spinifera 0.6 1,0 3,1 T, longipes 0,6 0.7 1.1 N, difficilis 0,1 0,2 0.9 T, oculatus 0,0 0,2 0,2 S, abbreviatum 0,0 0,0 0,2 52 Euphausia Pacifica - This species, which was abundant off the Oregon coast most of the year, is a subarctic and transitional water species found over most of the North Pacific Ocean and southward along the North American coast as far as Baja, California. The Davidson current, a subsurface, northward flowing, winter current along the coast of the western United States, may carry eggs and young E. pacifica northward, thereby maintaining the coastal populations of this species off,Oregon. This species is pri- marily found in the upper 300 m, but may occur in small numbers below 300 m in particular conditions. Thysanoessa spinifera - This species is primarily a neritic species found in the Gulf of Alaska and farther to the south along the coast of North America. According to Brinton, concentrations of this species have been associated by our own data, however. Vertically, T. spinifera is apparently limited to the upper 100 m. Thysanoessa longipes - This species is restricted to the North Pacific Ocean up to and including the American Arctic. The southern oceanic limit is at approximately 40°N but extends to northern California in the eastern North Pacific. Vertically, T. longipes is primarily a surface form. Nematoscelis difficilis - They are found in a belt like distribution across the North Pacific in the North Pacific Drift and in the California Current which flows south along the coast of North America. Its range in the coastal North American waters is more extensive than in the Western Pacific. It is rarely found in cold, upwelled water off Oregon, Washington, or northern California. Nematoscelis difficilis is most common above 140 m, Tessarabrachion oculatus - This species is confined to the subarctic North Pacific with a southward extension of the range of the west coast of 53 the United States, In vertical distribution, the adults are found at all depths sampled except at southern limits of the range where it disappears from the surface, Stylocheiron abbreviatum - These species have been found in the subtropical and tropical waters between the subtropical convergences (40°N and 40°S) with the exception of the eastern equatorial region, It is an offshore, warm water species in the region of the California Current. They are collected between 50 and 300 m depth and apparently do not undertake any vertical migrations, Nematobrachion flexipes - Although reported from the coastal waters of Alaska and British Columbia, this species is a tropical and subtropical species, Adults undertake some vertical migration being mainly at 280-700 m during the day and above 280 m at night, This species and S. abbreviatum, which are primarily subtropical species, are not normally found off Oregon. Thus they may be useful as indicators of the origin of coastal waters. Stylocheiron maximum - This is a large mesopelagic species found northward into the Gulf of Alaska as well as in the central and equatorial regions of the Pacific, Adults of this species are rarely found above 140 m depth and are caught in great numbers only below 500 m. REFERENCES Brinton, Edward, 1962. The distribution of Pacific euphausiids. Scripps Institution of Oceanogr. 8:51-270. Bull. 55 OCEANIC NATANTIA (PENAEIDEA AND CARIDEA) by Carl Forss B. Since 1961 the Department of Oceanography, Oregon State University, has undertaken an intensive study of the fauna and ecology of oceanic and mesopelagic waters off the coast of work several species of like forms) have been macurous obtained. Oregon. During the course of this decapod Crustacea (shrimps and shrimp- This paper is a preliminary report on some of the Natantia (Penaeidea and Caridea). 4.4 For details on the collection methods, see Section 1.' RESULTS The most common species of natant shrimp was Sergestes similis. It accounted for the bulk of specimens collected over the continental slope off Oregon, The catches of this species tended to decrease with distance from shore off Coos Bay and Astoria, However, an increase in the catches was noted offshore from Newport (Table 2), with the highest catch per tow (64) being found the farthest distance from the coast, S. similis occurred mainly in the upper 200 m collections and catches decreased rapidly with depth (Table 3). Hence this species, which has a transparent, membranous exoskeleton, was found near the surface in epipelagic waters, at least at night, S, similis is known to range throughout the north Pacific from Japan to Washington as far south as the Gulf of California, Most of the specimens of S. similis were comparatively small, ranging from 10 to 18 mm in carapace length (tip of rostrum to posterior margin of carapace) with the carapace length of 12 mm appearing most frequently, seasonal distribution pattern for S. similis has not been definitely A Table 2. Catches of Some Natant Decapod Crustacea off the Oregon Coast in 0-200 m Midwater Trawl Collections. Coos Bay Newport Mi les Off shore Species 15 25 45 Sergestes similis 35.0 19.0 20.0 Astoria Miles Of fshore 65 9.2 Mi les Of fshore 15 25 45 65 15 15.0 29.0 22.0 64.0 17.0 4.2 0.7 2.4 0.1 0.4 0 0 1.4 0 0 25 45 65 Gennadas sp. 0.1 0 0 0 0 0 0 0.1 Pasiphaea pacifica 2.7 0.2 0 0 0 1.4 0 0 Pandalus sp. 0 0 0 0 24 0.8 0 0 1.0 0 0 0 Hymenodora frontalis 0 0 0 0 0 0 0 0 0.8 0 0 0 625 501 376 4516 16 16 '17 70 Total number of species caught at each station Total number of hauls at each station 345 9 97 9 181 9 46 5 55 886 72 9 12 12 '12 19 8 57 Table 3. Catches of Oceanic Natantia in Tows to Various Depths at NH-50 0-200 m (118 tows) Species Number of Number Gennadas sp. 38 0.04 Number of (28 tows) Number 4543 5 13 366 0.3 9 Pasiphaea pacifica .03 1 Pandalus sp_ .07 2 .96 1.9 .03 Parapasiphaea sp. .07 Hymenodora frontalis Hymenodora gracilis Number of .03 Number tows tows tows Sergestes similis 0-1000 m 0-500 m (28 tows) 2 1 1.9 27 54 1 54 .60 17 Acanthephyra curtirostris .03 1 .14 4 Notostomus elegans .07 2 .03 1 Systellaspis cristata .03 1 Systellaspis affinis .03 1 Systellaspis braueri .17 5 58 established, but during the summer months it was noted that the larvae exceeded the adults in number, especially for the month of August (Fig, 1). This shows that the high numbers found in the summer were due to larval or juvenile animals, probably the result of breeding earlier in the year. It is interesting to note that Barham (1956) concluded from the abundance of young found the year around the S. similis spawned more than once a year. Pandalus sp, was only common at the inshore station off Newport, which is the only station over the continental shelf, They were rare Two species of Pasiphaea were identified, P. pacifica, which offshore. is common in catches, and P. chacei, Pasiphaea like Pandalus appeared to be an inshore species, occurring most. abundantly at station AH-20 (Table 2), Based on the catches to various depths (Table 3), it was concluded that several species of shrimp were mesopelagic (200-1000 m) in distribution, 1000 m. Gennadas was found almost exclusively in tows to depths to 500 or Both Hymenodora frontalis and the less frequent H. gracilis were also collected mainly in tows to 500 or 1000 m. The remaining oplophorids (Acanthephyra, Notostomus and Systellaspis) were found only in tows to 500 or 1000 m. 2000 Sergestes sirni/is LARVAE ADULT 1000 0 JUNE JULY AUGUST 1961 Figure 1. The distribution of adult and larvae forms of Sergestes similis for June, July and August 1961. 59 SALPS by Lyle Hubbard C. Five species of salps representing five genera were found off the Oregon coast. vagina These species are Helicosalpa virgula Tilesius, 1802, (Pallas, 1774) Salpa fusiformis Cuvier, (Vogt, 1854) 1804, Iasis zonaria and Pegea confoederata (Forskol, 1775). were based upon Yount (1954). Thetys Identifications Dr. Yount also verified our identifications from specimens sent to him. It should be noted that the data for 1961-1962 is based on counts from subsamples (split counts), while that for 1962-1963 is based on the number of salps from the entire sample (whole counts). Species Distribution The distribution of salps, both by the number of species and the number of individuals, appeared to vary with latitude. As shown in Table 4, the largest number of salps species per station during the two years was consistently found off Coos Bay (CH) or the southern stations. The second highest number of species were found off Newport (NH), while the lowest number of species was noted off Astoria (AH). On the basis of the number of salps per tow, Coos Bay was again the highest followed by Newport; the lowest number of salps per tow was found at the northern area or stations off Astoria. Thus there was a trend for more species and higher catches to the south of the Oregon coast, H. virgula was recorded only once in our tows. captured at CH-45 in March 1962, second year (1962-1963). Two individuals were It did not make an appearance in the 60 Thetys vagina was recorded at Coos Bay and Newport for 1962 and 1963 only. It did not appear off Astoria. Both solitary individuals and chains of the aggregate form were noted. Pegea confoederata made a single appearance off Coos Bay in 1961-62, but occurred off Newport and Astoria in 1962-63, failing to show on the Coos Bay stations. Iasis zonaria was found off Newport in 1961-62 and off Newport and Coos Bay the following year. It has not been caught off Astoria to date. Salpa fusiformis is the only form that has been found with regularity along all three lines of stations during both years. The echinate form of this species (S. fusiformis aspera Ihle,l911) has not been separated from the non-echinate form due to taxonomic difficulties. Seasonal Occurrence The abundance of salps off the Oregon coast (Fig. 2) seemed to vary by the season with peak numbers during the spring and summer months. Their occurrence the rest of the year was irregular. Peak abundance was noted between May and August for both years. The two years were not comparable on the basis of the number of salps collected: 1962-63 appeared to be a "salp" year compared with 1961-62. After September the number of salps generally decreased until the next May, sometimes not making an appearance for a month or more. Salps were noted early in March for both years followed by low numbers in April. On February 2, 1963, a large number (31) of Pegea confoederata were captured at NH-15. Helicosalpa virgula made its only recorded appearance in our catches in March 1962. Aside from the general salp peak in the 134 2592 b. 100 Cl. WHOLE COUNTS SPLIT COUNTS 1728 1 Solp count in, tow Tow -no salps o 3 0 Far w a 25 0 0 0 S 1961 Figure 2. N 0 J F M A 1962 M 411l1;1 J A 1 0 S 1962 Salp catch as recorded by the midwater trawl (1961-1963). J F 1, I M A' 1963 1111 1 M Distribution of Salps off the Oregon Coast as Determined by Midwater Trawling. Table 4. WHOLE COUNTS (July 1962--June 1963) SPLIT COUNTS (July 1961--June 1962) 15 25 45 65 65+ 15 25 45 65 65+ 0 0 123,60 0 0 0.42 4.47 2.83 0 0 No. tows 5 5 5 3 7 7 6 3 2 No, spp. 0 1 0 0 1 1 1 0 0 4.57 2.00 370,28 288.00 4.22 1.08 0.70 21.33 0 3 Stations, 1 AH- (Astoria) , Av. no./tow NH (Newport)Av. no./tow No, tows 7 7 8 7 6 9 12 10 12 No. spp. 0 1 1 1 1 2 1 1 4 Av. no./tow 180.80 161.60 171.20 234.00 52.00 15.00 12.00 19.80 0.40 0 No. tows 5 5 5 4 2 5 5 5 4 2 No. spp. 1 1 2 1 1 2 3 3 1 0 CH (Coos Bay) 62 spring and summer, February peak, especially and March must also be considered as a secondary for "rarer" species appearing off the Oregon coast. REFERENCES Yount, James.L. 1954. The taxonomy of the salp ideal (Tunicata) of the central Pacific Ocean. Pac. Sci., Vol. V171, No. 3, pp. 276-330. 63 D. MEDUSAE, SIPHONOPHORES. AND CHAETOGNATHS by R. W. Renshaw The medusae, siphonophores, and chaetognaths captured by the midwater trawl have been analyzed. The medusae and siphonophores, because of their gelatinous structure, were not sampled adequately and cannot be treated thoroughly in this report. A few generalized statements about the medusae will be made later in the paper. The siphonophores were represented off the Oregon coast by 13 species (Table 5). in seasonal There did not appear to be any indication of changes occurrence except for the condrophore Velella velella, which appeared off the coast in the spring. There was some evidence of differ- ences in vertical distribution, but without an opening-closing mechanism (which has recently been added to the trawl) any statement would be premature and must await further investigation. The chaetognaths were represented in the samples by 11 species (Table 5). Two species, Sagitta macrocephala, and Eukrohnia fowleri, are mesopelagic species and were only caught in 1000 m tows. of these two species had a red pigmented gut. All specimens For most species the data does not indicate seasonal change, or the data were too incomplete to show any change. An inverse relationship between Sagitta scrippsae and S. euneritica is suggested, which was correlated with oceanographic seasons. S. scrippsae is an oceanic species that is most abundant in the West Wind Drive and California Current. On the other hand, S. euneritica is a neritic species extending from Baja, California, to as far north as 46 14.5'N (approximately the Washington-Oregon border). Figure 3A compares the abundance of S. scrippsae and S. euneritica in our inshore collections, 15 miles off the Oregon coast, during January. S. euneritica appears to be much more abundant inshore than S. scrippsae, particularly at northern stations. This indicates more oceanic influence at the southern stations than the northern stations. A similar trend is sug- gested in Figure 1B where S. scrippsae is more abundant 45 miles offshore than S. euneritica, except at the northern station (46°14'N). At the intermediate station (44°38N) S. scrippsae is more abundant than it was at 15 miles offshore. These data suggest that at southern latitudes (43°20'N) there is a greater oceanic influence than at northern stations, Moving north the oceanic influence decreases and there is a spreading of the neritic environment further offshore. Figure 3A is for January and indicates little, if any, subarctic influence along the Oregon coast at this time. during the summer (July) scrippsae increased (Fig. abundance. S. euneritica decreased in abundance whereas S. 3B) and other subarctic species reached maximum For example, S. elegans, a subarctic chaetognath, was abundant during the summer and was virtually absent during winter months. medusae, Aglantha However, digitale, The another subarctic species, was also most abundant during the summer months (July and August). It was rarely cap- tured in the winter. The medusae were represented by 11 species (Table 5). Several other species may be present but the condition of the specimens is such that identification is impossible. Of those medusae that have been identified, Atolla vanhoeffeni and Periphylla periphylla were the most abundant. Without exception, they were captured on every tow. One medusae, b. 1000 0 S. euneritica 1000 S. scri ppsae 45 MILES 15 MILES 500 X L11 m z n 0 <j 430 20 N _ 440 38'N f 460 14` N 0 430 20' N 44° 38'N Figure 3. Catches of two chaetognaths during January 1962, at three latitudes off Oregon. (b) 45 miles offshore (a) 15 miles offshore 460'14! N 65 Table 5. A List of Species of Siphonophores, Chaetognaths, and Medusae Captured off the Oregon Coast. Siphonophora Vogtia spinosa Nectodroma dubia N. reticulata Lensia conoidea Sulculcolaria quadrivalvis Chelophyes appendiculata Physophora hydrostatica Bargmannia sp. Chuniphyes multidentata Muggiaea atlantica Nanomia bijuga Velella velella Chaetognatha Sagitta bierii S. decipiens S. euneritica S. elegans S. macrocephala S. minima S. scrippsae S. zetesios Eukrohnia bathypelagica E. fowleri E. hamata Medusae Aglantha digitale Colobonema sericeum Crossota rufobrunnea Pantachogon haeckeli Halistaura cellularia Aeguorea sp. Aegina citrea Solmissus marshalli Atolla vanhoeffeni A. wyvillei Periphylla periphylla 66 Colobonema sericeum appears to be amesopelagic species. It was only captured when the trawl net was fished to 500 or 1000 m. If plankton animals may be used as hydrographic indicators, the above data suggest that during the winter the influence of subarctic water is at a minimum, while in the summer months subarctic water is at a maximum off the Oregon coast. changes in water masses off Oregon. Such data are useful in showing Hence seasonal variation in the abundance of certain oceanic animals may be directly related to the circulation off Oregon. For instance, the high catches of fishes and squids in the summer may be affected by movement of water as well as movement of the animals themselves. 67 VERTICAL DISTRIBUTION OF THE NUMBERS AND BIOMASS OF MESOPELAGIC FISHES WITH AN IMPROVED ISAACS-KIDD MIDWATER TRAWL by W. G. Pearcy INTRODUCTION The Isaacs-Kidd midwater trawl or IKMT (Isaacs and Kidd, 1953) is recognized as one of the most useful devices for sampling small nekton and macroplankton over a wide range of depths in the ocean (Aron, 1962; Foxton, 1963). It has two serious inadequacies, however, for most eco- logical research; neither quantitative estimates of the density of catch or information on depth distribution are provided by the unmodified trawl. Quantitative estimates of pelagic animals require knowledge of the volume of water filtered by the sampler. This is complicated by the presence of three different mesh sizes in the IKMT, each having a dif- ferent filtration and selection characteristics (see Aron, 1959). Informa- tion on the depth of animals collected requres some means of opening closing the trawl. Several methods have been attempted for the IKMT. and A Leavitt-type releasing device was employed by Kelly (1958, 1961) in which the mouth of the trawl was initially pursed, and messengers actuated opening and closing of the cod-end of the trawl at depth. Disadvantages of this method are the notorious unreliability of the Leavitt system (Yentsch et al., 1962; Currie, 1962; Foxton, 1963) and the fact that some water is being filtered during descent of the trawl (Kelly, personal communication). 68 The most notable advance is the recent development of catch-dividingbucket (Currie, 1962; Foxton, 1963), which automatically diverts the catch from one cod-end net to another at a selected depth, thus giving two samples per tow. The cod-end nets are 6 feet long to provide flow of water through the bucket. In this paper several innovations to the Isaacs-Kidd midwater trawl were employed to study a specific problem: the general features of the vertical distribution of mesopelagic fishes within the upper 1000 m. The results are preliminary. METHODS Several modifications of the IKMT were made to improve it for quantitative sampling at various depth intervals. None of these improvements required any independent development, only the adaptation of products already commercially available. A Lamont Multiple Plankton Sampler (MPS) was adapted as a cod-end collecting unit for the midwater trawl. one a side, was used in this study. A scaled-down model, 13 inches Opening and closing operations took place at the preselected depths of 150, 500, and 1000 m during the descent of the trawl. This device employs a pressure-sensitive release mechanism which releases three rotating bars at preset depths, thus opeing and closing the nets and sampling within three successive depth layers (Be, 1962). The pressure-release system was calibrated by the manufacturer.* Lowering the device vertically (wire angle <5°) to various depths at sea showed the nets closed between 150-160 m, 480-520 m, and 940-960 m, respectively., * G. M. Mfg. Co., 12 East 12th St., New York, N. Y. 69 To reduce the problems associated with different mesh sizes in the trawl net, the entire body of the net was lined with a 3/8" (squared measure) knotless nylon. Hence, with exception of the #0 plankton net in the collecting nets of the MPS, only one mesh size was used. A depth-distance recorder, mounted on a stay in the mouth of the trawl (Fig. 1), gave estimates within each depth interval. of the distance the trawl traveled According to the manufacturer,** the error of the depth-distance recorders, determined by pressure testing in the laboratory, is t 3 percent of the maximum depth. sea gave comparable depth variations. Our field testing at The recorder has horizontal fins and is self-adjusting for pitch Figure 2 duplicates the trace from a typical tow. The trawl descends at 30 m of wire/minute while the ship is underway at about 5 knots. first net, The which is open on the way down, is closed and the second is opened at 150 m depth. The second net is closed and the third opened at 500 m, and the last net closed at 1000 m depth. It is important to note that the trace is approximately a straight line within each depth interval, 0-150, 150-500, and 500-1000 m; this indicates that nearly equal volumes of water were fished at all depths within each layer. (If the trawl does not fish all depths equally and animals are not distributed uniformly within depths, estimates of abundance may be seriously biased.) Assuming that the flow of water through the depth-distance recorder applies to the entire mouth of the trawl, then the volume of water filtered can be calculated within each depth interval as a product of the cross sectional area of the trawl (2.89m2, as estimated by planimetry ** Tsurumi Precision Instruments Co., Japan. Ltd., 1506 Tsurumi-Machi, Yokohama, 70 and triangulation) and the distance traversed within each depth layer. The most serious problem encountered during sea trials with the modified trawl was insufficient flushing of the contents of the trawl past the MPS into the collecting nets. After tows to 1000 m depth, the net, which ascends with the cod-end open, frequently had animals adhering to the trawl netting. They were articularly concentrated toward the end of the trawl just before the attachment of the MPS. This indicated that animals were not immediately passing through the trawl netting. Thus, animals accumulating in the trawl at one depth layer were a source of contamination for nets open at lower depths. The flushing of the midwater trawl was improved by the following modifications: (1) a knotless nylon liner was used instead of the knotted netting; (2) the midwater trawl net was tailored with a gradual taper to the MPS eliminating the looseness produced when the 1/2 m cod-end diameter was constricted over the MPS; (3) a plastic (Herculite) collar was added to the end of the trawl to further reduce pockets for accumulation of animals and furnish a better means of attaching the MPS to the trawl; and (4) to increase the flow of water through the MPS while sampling, a six-foot section of 3/8" knotless nylon was added between the MPS frame and the plankton nets. With these changes, contamination was markedly reduced. Providing sufficient flow of water through the opening-closing device will undoubtedly be a problem common to all cod-end units (see Foxton, 1963). Some contam- ination is unavoidable since time is required for organisms to pass through the net. It can be minimized when flow is accelerated or the time fished at each depth layer is prolonged, i.e., when the ratio, flushing time/ Figure 1. Isaacs-Kidd midwater trawl showing the depth-distance recorder mounted on the depth depressor. DISTANCE 2 3 4 5 MILES IN 6 7 8 9 10 II I- a W a 1 000 Figure 2. Duplication of a trace made with the depth-distance recorder on a tow which sampled three depths within the upper 1000 meters. 71 fishing for a net is low. time, The weight of the MPS was originally thought to pose an added flushing problem by sinking relative to the end of the trawl. However, the low specific gravity of the PVC frame and the hydrostatic pressure of the water maintained the MPS at the same level as the trawl. Operational success of the MPS-IKMT unit has been total of 30 separate tows to 1000 m attempted, samples, $1 samples from discrete depths were ures (10 percent) good. Out of a which should give 90 All of the fail- obtained. were because the deep net (500-1000 m) did not close. The presence of water inside the cylinder may have caused the failures so an addition at "0" ring was installed around the piston. Ridges in the cylinder, which also may have prevented a full stroke of the piston, were Since these minor structural changes, all nets have,closed. ground down. Collections with the MPS and IKMT for study of the vertical distri bution of biomass and numbers of fishes were generally made over a 24-48 hour period, providing both day and night collections for each depth layer. All the collections reported on here were made in the vicinity of 44°37'N, 125°15'W, 50 miles off Newport, Oregon, over the outer continental slope. Geographic fixes were recorded at the beginning of each tow and when the trawl was at each closing depth, assuming a wire/depth ratio of 4:1 (see Pearcy, Section 1). not available, On the January series, and the distances towed-were a depth-distance recorder was approximated, using the geo- graphic fixes and correcting for the difference between the position of the vessel and the trawl. Since Backus, Hersey (1956) and Mead (1963) found that the cable of the IKMT was nearly straight during towing, these differences were determined by simple triangulation. 72 After each tow the net was thoroughly washed by towing beside the vessel and by use of fire hoses. formalin in sea water. Collections were preserved at sea with Fishes were removed from the collections, weighed after removal of excess water (wet weight), identified, and counted. RESULTS The number and biomass of fishes collected per 1000 mJ within each depth interval are summarized as histograms for three series of collection made during different seasons during the year (Figs. 3 and 4). Each series includes collections made during day and night periods. A general pattern of bathymetric distribution of fish numbers is evident for each of the three series of collections (Fig. 3). During the day, maximum numbers were found at mid-depth (150-500) for all series, and relatively few fish were found in surface waters (0-150 m) or below 500 m. During the night, i.e., tows initiated after sunset, the maximum numbers of fishes occurred in surface waters. In all instances, catches in deep water (500-1000 m) were lowest. The diel diurnal variations within each series indicate an increase in the numbers found in surface waters at night, a feature that has been commonly observed in the past. But the corresponding decrease in the numbers at mid-depths has rarely been documented. Both changes are posi- tive evidence of vertical migration of these mesopelagic fishes within the upper 500 m. There are no consistent trends shown that suggest any diel change in numbers below 500 m, as numbers are uniformly low during both day and night periods. Thus, there is no evidence for vertical migration below 500 m from these data. 7-8 JANUARY 1963 TIME" 0753-1102 1249-1612 C.-150 1822-2134 0.6 150 - 500 2358 - 0309 (:7 2:0 0.5 500 - I OOC 05 0:5 28-30 APRIL 1963 V) x W TIME 0658-0857 07 0-150 1250 -f635 1_3 0.2 0.8 0.3 0.4 T 03 02 1 1 150-500 Z = 500-1000 I L CL W 8C7-2018 0030 - 0235 0.9 0 ?-8-30 AUGUST 1963 TIME' 0536-i324 1 630 - 1845 i 7'2-1920 2317-0;4C 00'5 - 0240 0-150 150-500 500-1000 0.9 0.4 1 Figure 3. I Histograms showing the number of fishes captured per 1000 m3 of water filtered within three depth intervals for collections over day and night periods. 7-8 JANUARY 1963 TIME: 0753-1102 1249-1612 X822-2134 0-150 2358 - 0309 1.8 50-500 38 28-30 APRIL 1963 TIME 0-150 065E-0857 125..-635 18C .0.5 I 0030-0235 0.7 23 .50-500 56 z 28-30 AUGUST 1963 TI ME ; C3+-1322 237 -C i 82 50C 0.4 I Figure 4. 84 08 001 5 - 024C 199 3f 56 76 05 86 1 Histograms showing the wet weight of fishes collected per 1000 m3 of water filtered within three depth intervals for collections over day and night periods. 73 Similar trends are apparent from the catches expressed as biomass of fishes per 1000 m3 for the various depths (Fig. 4). Usually the weight of fishes was highest at mid-depth during the day and lower in surface waters and deep waters. However, in some cases the trends are rather variable, night, for example, maxima appear at all depths. During the The lack of.a large surface maxima during the January series may have been caused by a_full moon which surpressed the ascent of fishes. The biomass in 500-1000 m collections was usually low and no..day-night differences were evident. The occurrences were evident. The occurrence of high biomass in deep water in the last series was surprising after the low values found in earlier collections. In both cases, these high weights shown for. 500- 1000 m in Figure 4 were caused by.a.few large fishes. Differences in the vertical-distribution of number and biomass are interesting. Diel variations were evident.in both the 0-150 and 150-500 m depth intervals in numbers, but only between 0-150 m in the biomass. Moreover, the decrease in the numbers of fishes at mid-depth between day and night periods was not evidence in fish biomass. Hence a decrease in number was apparent at night between 150 and 500 m, oddly enough with no corresponding decrease in weight. The standing crop of fishes per m2 surface area through a column 1000 m deep was calculated for all series by multiplying the average catch per m3 times the depth of the the three layers. shown on the 1000 3.3. interval and summing the values for This merely is a weighted average of the estimates m3 basis. The number of fish per m2 ranged from 0.6- 2 Biomass was between 1.1 and 7.6 g per m. These estimates, which 74 are considered first approximations, for the standing crop of mesopelagic are the only ones to my knowledge fishes. Of interest is the fact that the biomass of these small oceanic fishes is about the same as the estimates of biomass per m2 of pelagic fishes in the English Channel (Harvey, 1950). Day-night differences in average catches per m2 in Table 1 indicate that usually fishes were more numerous in night tows. fish biomass was collected per m2 at night. In all series more This indicates.tha't collec- tions during darkness are more effective in capturing mesopelagic fishes within the upper 1000 m. The day/night ratio of b.iomass was lower than the day/night ratio of numbers except for the January series, indicating that there ,was- a greater increase in weight than in numbers in the night samples. In other words, the average size of fish captured at night was larger than in the day (Table 1). This difference explains the seemingly paradoxical decrease in numbers at mid-depth with no apparent decrease in biomass. At night, larger fish made up a significant portion of the biomass though few in number. DISCUSSION It has been known for a long time that more micronekton and macro- plankton are collected during the night in upper ocean waters than during the day. nized: Two explanations for this phenomenon commonly have been recog- (1) vertical migration of animals below sampling depths during the day, and (2) better visual avoidance of nets during the daylight hours. If all depths through which the nets are towed are not sampled equally, 75 Table 1. Number and Weight of Mesopelagic Fishes Collected per m2 (to 1000 m depth) and Average Size of Fishes (g/m) for Three Series of Collections. January 7-8, April 28-30, Night Day Date: Day/Night 1963 No./m2 1.2 1.0 0.9 1.1. 1.10 G/m2 1.7 1.8 2.1 2.1 0.83 G/no. 1.4 1.8 2.3 1.9 No./m2 0.7 0.6 0.2 1.4 0.36 G/m2 1.8 1.1 1.2 6.3 0.22 G/no. 2.6 1.8 6.0 4.5 404 408 1963 August 28-30, 1963 No./m2 2.2 2.4 2.1 3.3 1.8 0.87 G/m2 1.3 7.6 2.6 5.2 7.4 0.61 G/no. 0.6 3.2 1.2 1.6 4.1 76 an added possibility exists, namely that day and night differences are merely an artifact due to preferentially sampling depths where animals are concentrated during the night. This latter possibility is minimized when all depths are sampled equally by a net, e.g., oblique tows. Several examples of these day-night differences found in oceanic animals of the northeastern Pacific are pertinent. Tucker (1951) noted that the large increase in the number of lantern fishes captured in surface waters during the night was not accompanied by an equivalent decrease in the numbers found deep waters off San Diego, California. He noted that the difference between the number of fishes captured during day and night periods may have been caused by ascent at night from depths below 600 m, the maximum depth of his McAlister (Tucker's) trawling. (1961) observed that the weight of plankton collected with half-meter nets from 300-500 m was about the same for both day and night periods, but five times more was collected in the upper 300 m during the night than during the day. Aron (1962) also found that night collections caught substantially more biomass than day collections. The results of King and Iverson (1962) on the catches of pelagic fishes by midwater trawling in the Central Pacific demonstrated that night collections took about four or five times the volume but only 2.5 times the number of fishes as the day catches. Such differences between volume/number ratios indicated that, on the average, larger fishes were captured at night. This is similar to the results I have reported. As both vertical migration and the ability of animals to avoid the collecting net are influenced by submarine light intensity, their effects on diel variations of catch are difficult to discriminate. This is 77 especially true if sampling is not carried out below the day depth of the migratory animals. In all of the above mentioned studies, sampling or less. was restricted to 600 m depths Therefore, it was recognized by these authors that the high catches at night could be caused by either migration of animals from below the maximum sampling depth or by net avoidance. If migration during the day to depths lower than 500 m is, the sole explanation of the differences in catch,, then we would expect to find an increase in the numbers and biomass of fishes collected between 500-. 1000 m during the day. substantial proportion intensity, even larger trawl than in overlying In most cases cited, this must necessarily be a of the animals. At these depths of,low.,light fishes should be'less-able to visually avoid the waters. Off Oregon, catches below 500 m were generally low in numbers: and biomass. There 500 and 1000 vals. were indications of diel variations in number between m, as were observed in the 0-150 and 150-500 m depth inter- Consequently, even though there was good evidence for vertical migrations above 500 m, the simplest explanation for the day-night differences in numbers and weight of fishes per m2 is better visual avoidance of the trawl within the upper 500 mduring the day, particularly by the larger fishes. SUMMARY (1) A new method is described for quantitatively sampling difference depth intervals with the Isaacs-Kidd midwater trawl. 78 (2) Diel vertical migration is reflected by the increase in the number of fishes collected per 1000 m3 from 0-150 m at night and the decrease from 150-500 m during the day. (3) There was an increase in the biomass of fishes captured at night in surface waters (0-150 m), but no- apparent differences below 150 m. (4) The biomass,and especially the number of fishes found below 500 m, were low compared to overlying waters. (5) There was no apparent day-night difference in the number and biomass of fishes below 500 m. biomass and number of fishes captured per m2_surface area .(6) The (to 1000 m depth) were almost always greater at night. fish also was greater at night. catches per The average size Thus, the major diel differences in can be most readily explained. by better visual.avoidance m2 of the trawl within the upper 500 m during the day than during the night. REFERENCES Aron, W. 1959. Midwater traveling studies in the North Pacific. Limnol. and Oceanogr..4:409-418.. ------- 1962. Some aspects of sampling the macroplankton. Rapp. Proc. Verb. Cons. Int. 1'Explor. Mer. 153:29-38. Backus, R. H. and J. B. Hersey. 1956. Echo sounder observations of midwater nets and their towing cables. Be, A. W. H. samplers. CurVie, R. I. 1962. Deep Sea Res. 3:237-241. Quantitative multiple opening-and-closing plankton Deep Sea Res. 9:144-151. 1962. Net closing gear. l'Explor. Mer. 153:48-54. Rapp. Proc. Verb. ons. int. 79 Foxton, P. An automatic opening-closing device for large plankton 1963. nets and midwater trawls. Harvey, H. W. 1950. Plymouth. J. Mar. Biol. Assoc. U. K. 43:295-308. On the production of living matter in the sea off J. Mar. Biol. Isaacs, J. E. and L. W. Kidd. Assoc. U. K. 29:97-137. Isaacs-Kidd midwater trawl. 1953. Scripps Inst. of Oceanogr. Ref. 53-5, 18 pp. Kelly, G. F. Woods Hole Laboratory, U.S. Fish In.annual report. 1958. and Wildlife Service, June 30, 1958, pp. 29-33. 1961. Vertical distribution of young redfish and A. M. Barker. in the Gulf, of Maine. Rapp. Proc. Verb. Cons. Int. 1'Explo-r. Mer. 150:220-233. King, J. E. and R. T. B. Iverson. 1962. Midwater trawling for forage 1951-1956. organisms in the Central Pacific. U.S. Fish and Wildlife Service Fish Bull. 62(210):271-306. McAllister, C. D. 1961. Zooplankton studies at the ocean weather station "P" in the Northeast Pacific Ocean. Mead, G. W. 1963. Observations on fishes caught over the anoxic waters of Carioca Trench, Venezuela. Tucker, G. H. 1951. Deep Sea Res. 10:251-257. Relation of fishes and other organisms to the scattering of underwater sound. Yentsch, C. J. Fish. Res. Bd. Canada 18:1-29. J. Mar. Res. 10:215-238. S., G. D. Grice and A. D. Hart. 1962. Some opening-closing devices for plankton nets operated by pressure, electrical and mechanical action. 153:59, 65. Rapp. Proc. Verb. Cons. Int. 1'Explor. Mer. 81 NOTES ON THE VERTICAL DISTRIBUTION OF ZINC-65 AND ZIRCONIUM-95 FROM OCEANIC ANIMALS by W. G. Pearcy and C. L. Osterberg INTRODUCTION The potential role of animals in modifying the distribution of radionuclides by their own movements has been clearly recognized. Migrations may result in a net transport of elements from areas of high concentration to areas of low concentration in horizontal and vertical directions. Vertical transport of radionuclides is of` singular importance because animals making daily vertical migrations are abundant in the oceans. Moreover, their movements frequently penetrate density gradients that impede physical mixing of surface and subsurface waters. Vertical transport by such animals may equal or exceed transport of radioisotopes by physical processes (Ketchum and Bowen, 1958). This paper includes some preliminary data on the concentrations of two radionuclides in marine animals from various depths: zinc-65 and zirconium-95-niobium-95. Zinc-65 is produced mainly from neutron activation of Columbia River water by nuclear reactors. a fission product entering the ocean,from fallout. Zr95-Nb95 is These radionuclides, as well as other gamma emitters, have been previously reported from pelagic animals off Oregon (Osterberg, et al., Sections 6 and 7). 82 METHODS Animals were collected with an Isaacs-Kidd midwater trawl (mesh size 1/2" stretch) using a Lamont Multiple Plankton Sampler as an opening-closing unit, thus providing samples from three depth intervals: 0-15Q m, 150-500 m, and 500-1000 m, The volume of water fil- tered was estimated from the cross-sectional area of the mouth of the trawl and the distance towed at each depth interval as determined from geographic fixes (see Section 4 for details). Four tows, two during the day and two during the night, 50 miles off the central Oregon coast (44°37`N, 125°15`W) during January 7-8, 1963, provided the material radioanalyses. Whole samples of animals, consisting of a mixed species composition, were weighed (wet-preserved weight), ashed at 450°C, and packed in 15 cc plastic counting tubes, reweighed and sealed. Samples were then placed in the well of a 5" x 5" Harshaw Nal crystal and analyzed with a gamma-ray spectrometer (512 channel nuclear data model 130A) according to the method of Perkins (1958) and Covell (1959). time was either 100 or 200 minutes, depending on sample size. Counting Counts were converted to picocuries per entire sample, per gram and per 1000 m3 water filtered. RESULTS AND DISCUSSION The data on the picocuries of Zn65 and Zr95^Nb weight and wet weight are given in Table lA and 1B. 95 per gram of ash The total varia- tion in the concentration of Zn65 was small, ranging from 11.9 to 83 42.9 pc/g ash and 0.4-2.1 pc/g wet weight. The average Zn65 content found in animals from the four series of collections for each depth interval, 0-150 m, 150 -500 m, and 500 -1000. m,; indicated on an ash weight basis and 1.1: 1.2: ratios of l 2.1: 1.0 on a wet weight basis. 1.0 This indicates little variation in the average concentration,of Zn65 in the animals with depth. most as much Zn65 Animals from deep water per gram as animals (500-1000 m) contained al- above 500 m. The concentrations of Zr95-Nb95 on the other hand, showed a much wider variation, 7.I-269'4 pc/g ash and 0.4-7.2 pc/g wet. weight.: Much of this variation appears to be related to the depth of capture of the animals. The ratios for Zr95-Nb95 with depth are 9.0: ash weight basis and 5,,4: 0.7: 1.1: 1.0 on an 1.0 on,a wet weight basis for the.,0--150 m, 150-500 m, and 500-1000 m collections, respectively. These differences indicate that the concentration of this radionuclide is about 5 to 9 times higher in the animals collected at the surface (0-150 m) than at the greater depths. Undoubtedly, some of the differences between the trends shown on an ash weight and wet weight basis are. due to the type of animals collected. For example, during the day surface catches often consisted of a higher percentage of gelatinous or watery forms (medusae, salps, chaetognaths); hence, the ash weight/wet weight ratio may be much:.:lower than at night when animals like euphausiids are common in surface waters. The differences in the concentration of these two gamma emitters with depth agree with the conclusion of Osterberg et al. (Section 6) that particulate fallout isotopes such as Zr9S-Nb95 showed prominent spectral peaks only in filter-feeding heribores and were not concentrated appreciably 84 Table 1. Concentration of Zn65 and Zr95-Nb95 in the Samples of Oceanic Animals from 0-150,, 150-500, and 500-1000 m in Depth, Collected January 7-8, 1963, 50 miles off Newport, Oregon. standard counting error. Zn65 and Zr95-Nb95/g ash Zn65 and Zr95-Nb95/g wet weight of sample. water filtered. g wet weight of animals per 1000 m A. B. C. (picocuries) A. Zn65/g ash 0-150 m 150-500 m 500-1000 m 42.9 42.4 19.9 18.9 0.8 1.3 10.5 53.8 11.9 5.8 1.2 1.2 39.8 44.2 14.8 2.0 1.2 1.2 36.3 28.3 34.1 1.0 0.8 1.3 128.5 30.4 8.4 7.0 1.1 2.8 138.9 19.1 15.4 5.0 1.0 1.3 55.7 17.1 34.8 1.0 0.7 1.4 Zr95-Nb95/g ash 0-150 m 150-500 m 500-1000 m B. 269.1 11.8 38.4 0.7 7.1 1.3 Zn65/g wet weight 0-150 m 150-500 m 500-1000 m 1.00 1.89 1.42 0.42 1.79 0.71 2.07 1.00 0.75 1.89 1.48 2.12 7.22 2.90 0.89 2.17 13.96 7.28 2.32 Zr95-Nb95/g wet weight C. 0-150 m 6.27 150-500 m 0:53 5.19 1.01 500-1000 m 0.51 0.50 0.43 0.78 1.20 7.97 3.30 6.66 8.57 2.62 g wet weight/1000 m3 1.08 6.89 1.66 85 by higher trophic levels. which is ionic in sea water Whereas Zn65, (Greendale and Ballou, 1954), was concentrated by all animals analyzed off Oregon. surface A lower Zr95/Zn95 ratio was expected in deep water than in water, therefore, because most of the deep animals were carnivores. The vertical distribution of Zn65 and Zr95-Nb95 per volume of water is graphically shown in Figure 1 for each of the three depth layers. concentration of these two radionuclides was calculated from: The pc/1000 m3 (pc/g ash)(total g ash)/1000 m3 filtered. The histograms ofZn65 (Fig. LA) During the day the total amount of face layers is very low. increase reveal several interesting features. Zn65 in midwater animals from the sur- At night, however, there is an order of magnitude Although Zn65 per gram wet in the Zn65 of these upper waters. weight may be somewhat higher in animals collected at the surface at night (Table 1B), this cannot explain the large increase in radiozinc. It is mainly due to the high biomass in the upper 150 m at night and the paucity of animals at this depth during the day (Table 1C). Consequently, much of the Zn65 measured appears to be incorporated in small nektonic and large planktonic animals that undertake vertical migrations into the upper 150 m at night. The vertical flux of Zn65 indicated here may actually be a low estimate because a full moon during the night period probably suppressed the ascent of animals during the night (see The trends of Zn65/ 1000 m3 Pearcy, Section 4). at mid-depths (15Q-500 though less are also correlated spectacular than those observed in surface waters, with diel variations of biomass The observed (Fig. 1). m), decrease in Zn65 at mid-depths during the night is expected if animals ascend from this layer into surface waters. The fact that 150-500 m biomass (g/ 1000 m3 86 in Table 1C) does not demonstrate an apparent decrease at night is most simply explained by the capture of larger animals during the night when their ability to avoid the trawl is reduced compared to periods of daylight (see Pearcy, Section 4). These larger animals captured at night in mid- depths appear to have a lower concentration of Zn65/ g wet weight (Table 1 and Appendix I) than those which migrate nightly into surface waters. The concentration of Zn65 in deep water (500-1000 m) is relatively low and uniform in Figure 1. The amount of Zn65 found in the midwater animals off Oregon calculated per m2 in a column 1000 histograms in Figure 1. m deep was 5.9, 6.2, 6.1, and 10.2 pc for the The variability of these values is less than that found between samples at any one depth. Histograms showing the vertical distribution of Zr 95-Nb 95 (Fig. 1) are more difficult to interpret due to the variability found among depths. Still, the Zr95-Nb95/1000 m3 demonstrates appreciably day-night difference in surface waters (0-150 m). tion of animals. This again may be related to vertical migra- The amounts of this radionuclide in the day collections, however, were 2.7 and 4.6 pc/m2 compared to 9.5 and 7.4 pc/m2 for collections during the night. night. Thus over twice as much was collected per m2 at Such a difference, as well as the lack of evidence for any in- crease in Zr95-Nb95 during the day at mid-depths, suggests that other factors may play a role in affecting the diel and vertical variations of this radionuclide. An explanation for this difference is that the availability of animals that accumulate Zr95-Nb95 may vary within the sampling period. For example, it has been noted that the biomass of animals per m2 captured DAY N I GHT Zn65/ IOOOm3 5l. 2 z .5 0.5 1 - 500 Co - 000 Zn65/m2 64 1 130 [1421 86 24 2.4 2:0, 4.9 5.9 6.2 6.1 10.2 108 Zr95 - Nb95 / 1000 m3 r .1-150 58 162 481 ,5C-500 3.6 80 3.7 T. 0.8 500 - 1000 c Zr95-Nb95/ mZ Figure 1. 2.7 I 4.6 176 65 zo 9.5 7.4. Zn65 and Zr95-Nb95 contents in pelagic animals for day and night periods, January 7-8, 1963, 50 miles off Newport, Oregon. 87 during the day in tows to 1000 m is often less than the biomass collected at night. In the case of fishes, this is due primarily to the presence of larger animals in night collections, animals which may avoid the trawl in the day. Similar trends with plankton, such as euphausiids, suggest that they also demonstrate day-night changes in availability, perhaps due to more effective escapement during the day. Thus day-night differences in the catch of herbivores like euphausiids, which have been found to accumulate particulate fallout radionuclides like Zr95-Nb95 more than carnivores (Osterberg, et al., Section 6), may be particularly important in affecting our estimates of Zr95-Nb95/1000 m3 Another possibility is that there may be diel differences in the actual amount of Zr95-Nb95 found in the animals. Zr95-Nb95, unlike Zn 5, is not metabolically active and probably passes directly through the digestive tract. (Chipman, 1958). for instance, was rapidly excreted by copepods As a result, animals actively feeding in upper waters at night may have peak Zr95-Nb95 concentration at this time of day. Although this suggestion is not corroborated by the variations g (Table 1), of Zr95-Nb95 it requires further study. SUMMARY Zr95-Nb 1. The concentrations of two gamma emitters, Zn65 and 95 were examined in pelagic animals collected at three depths, 0-150, 150-500, and 500-1000 m, using an opening-closing unit on a midwater trawl. 2. Zn65/g wet weight of sample was about the same at all depths, whereas Zr95-Nb9S/g was about five times higher above 150 m than in deeper water. 88 3. The Zn 65 in animals per 1000 m3 water filtered was highest in This the upper 150 m during the night and in mid-depths during the day. was mainly due to vertical migrations of animals. 4. The highest values for Zr95-Nb95/1000 m3 also were found during the night in surface waters. 5. The amounts of Zn65 and Zr95-Nb9S per m2 (to 1000 m depth) were 5.9-10.2 and 2.7-7.4 picocuries, respectively. REFERENCES COVELL, D. F. 1959. Determination of X-ray abundance directly from the total adsorption peaks. Anal. Chem. 31:1785-1790. GREENDALE, A. E. AND N. E. BALLOU. 1954. Physical state of fission product elements following the vaporization in distilled water and seawater. CHIPMAN, W. A. organisms. U. S. Navy Res. Def. Lab. Doc. 436, pp. 1-28. 1958. Accumulation of radioactive materials by fishery Proc. 11th Gulf & Caribbean Fish. Inst., KETCHUM, B. H. AND V. T. BOWEN. 97-110. 1958. Biological factors determining the distribution of radioisotopes in the sea. In: Proc. 2nd U. N. Int. Conf. on Peaceful Uses of Atomic Energy, 18:429-433. PERKINS, R. W. 1958. Gamma-ray spectrometric systems of analysis. In: 2nd U. N. Int. Conf. on Peaceful Use of Atomic Energy, 28: 455-461. 89 Concentration of Several Gamma Emitters in Nine Species of Mesopelagic Fishes Collected on December 7, 1962, at a Station 50 Miles off Newport, Oregon Appendix 1. K40 pc A. Zn65 std std error error pc Ru-Rh Zr95-Nb95 pc error std std std pc error pc error Upper Mesopelagic Fishes, collected above 200 m at night Lampanyctus ritteri pc/g ash pc/g wet 56.6 1.7 2.0 26.7 0.8 4.2 106.7 3.3 26.5 103.5 3.2 27.0 49.5 20.4 201.6 6.1 20.1 0.5 74.3 151.6 3.5 75.4 1.1 51.0 1.2 12.0 4.6 2.7 34.9 0.9 4.6 56.1 1.5 28.5 55.8 28.5 79.2 1.8 55.5 1.1 4.9 98.5 2.3 102.1 2.1 29.4 36.7 4.0 3.4 1.1 0.1 14.8 11.2 0.3 wt. Diaphus theta pc/g ash pc/g wet wt. 6.9 Lampanyctus leucopsarus pc/g ash pc/g wet 23.2 0.6 wt. Tactostoma macropus 44.9 pc/g ash pc/g wet wt. 1.0 34.4 pc/g ash pc/g wet wt. 0.7 7.4 151.4 4.1 8.0 8.2 79.6 1.8 90.5 1.8 2.3 66.7 2.4 2.5 5.0 1.5 30.5 121.8 2.8 129.3 29.8 30.9 2.6 Tarletonbeania crenularis pc/g ash pc/g wet 23.1 wt. 9.9 7.3 2.5 39.6 12.6 4.3 B. Lower Mesopelagic Fishes, collected in tows to below 500 m depth Chauliodes macouni 31.8 pc/g ash 0.6 pc/g wet wt. 72.2 1.4 3.7 51.9 1.0 8.4 142.4 2.8 52.7 174.8 3.5 54.0 Poromitra cristiceps 30.7 55.9 pc/g ash 0.9 pc/g wet wt. 80.5 1.3 6.7 25.1 0.4 11.1 256.0 4.1 52.6 144.5 2.3 45.3 Lampanyctus nannochir 33.3 14.5 pc/g ash pc/g wet wt. 0.7 43.0 0.9 3.6 40.2 0.9 8.4 214.7 4.7 52.9 197.5 4.4 53.8 Bathylagus sp. 25.0 pc/g ash 0.9 pc/g wet wt. 26.8 0.9 2.3 30.8 1.1 4.5 56.9 1.9 23.0 101.4 21.4 13.9 10.3 3.5 91 RADIOACTIVITY AND ITS RELATIONSHIPTO OCEANIC FOOD CHAINS by Charles Osterberg, William G, Pearcy,_-and Herbert Curl, Jr. ABSTRACT Gamma-ray spectra of some primary producers (single-celled plants), filter-feeding I, II, III, herbivores, and carnivores, and III-V, respectively, assigned to trophic levels were prepared from marine samples taken in the Pacific off Oregon during 1961-1962. These organisms had been exposed in their natural environment to both fission products from fallout and neutron-induced radionuclides from reactors on the Columbia River. Comparisons of spectra of organisms from different trophic levels, determined from stomach contents and the and Ce141 were concentrated by primary producers and by carnivores. ducers).- vores. literature, herbivores, but not Cr51 was abundant in only filtered samples (primary pro- Mn54, Co60, and Cs137 Zn65 show that Zrg5-Nb9S were found in only herbivores and carni was found in every marine organism examined. that the abundance of Zr95-Nb95 and in marine trophic level studies. in particular, We conclude may be useful Peaks due to these fission products are greatly reduced in spectra of predaceous animals, compared with spectra of herbivores. 92 INTRODUCTION The oceans receive a substantial share of radioactive fallout resulting from nuclear testing because of their large areas and the drainage they receive from the continents. Prevailing westerly winds carry tropospheric fallout from nuclear tests in northeastern Asia across the Pacific Ocean to North America. Levels in the environment are normally quite low, but certain fission products are accumulated by filter-feeding zooplankton (Osterberg, 1962b, 1963). Radionuclides are also introduced by nuclear reactors at Hanford, Washington. Many trace elements in the Columbia River water used to cool the reactors are activated by the intense neutron flux (Nelson, ed., 1961). These induced radionuclides are returned to the river, and ultimately portions of them enter the ocean. The presence of radioisotopes in the ocean off the Oregon coast and the supply of nekton and plankton available from our midwater trawl program have made it possible for us to investigate the presence of both fallout and neutron-induced radionuclides in oceanic food chains. The gamma-ray spectra of marine organisms from different trophic levels are compared to determine which radionuclides are passed through food chains, and which are discriminated against. Assignment of an organism to a particular trophic level is, in most cases, based on studies of stomach contents, supplemented by references from the literature. METHODS Phytoplankton and detritus were collected by passing surface sea water through a 5-inch membrane filter (0.65 microns), plus glass fiber 93 pre-filter (Gelman Instrument Company). The filters were ignited in an open crucible to destroy the membrane filter, the residue placed in a muffle furnace for an hour at 500°C and then ground with mortar and pestle before packing into counting tubes. Macroplankton and micronekton were collected in a 6-foot Isaacs. Kidd midwater trawl towed for 30 minutes from 200 m depth to the surface. Plankton and nekton samples were freeze-dried after formaldehyde preservation. Large or oily samples were further concentrated by aching in a muffle furnace. Dried and/or ashed concentrates of the entire animalsl/ including digestive tracts were then packed into counting tubes 1/ Salps and, of course, tuna liver were exceptions. Only the opaque interior "nucleus" (or digestive tract) of the salp was used, since the transparent outer portion was found to be low in radioactivity (Osterberg, et al., 1963). (Falcon Plastics, Item #2001).. Tubes, containing the prepared samples were placed in the well of the 5" x 5" NaI(TI) primary crystal of the Hanford, Washington, total absorption anticoincidence spectrometer (Perkins, et al., 1960) for analysis. Counting time was 30 minutes, with 30-minute back- ground subtracted. RESULTS Trophic Level Relationships The first trophic level in the pelagic environment consists of single-celled plants. feeding herbivores. The second trophic level consists of filterCarnivores compose the higher trophic levels, Most 94 food relationships are not simple food chains, but are more often complex webs. Since feeding animals are opportunists, the uncertainties of diet are great, particularly in the case of large predators. Nevertheless, food patterns do exist and some division into trophic levels is possible. Trophic Level I Phytoplankton and detritus were trapped on a membrane filter through which surface sea water was passed. Most animals were removed by pre- filtering through a #6 mesh net. Trophic Level II Euphausia pacifica. Euphausiids may feed on phytoplankton, small crustacea such as copepods, or detritus (Ponomareva, 1954; MacDonald, 1927; Marshall, 1954). Although Ponomareva (1954) noted that E. pacifica occasionally fed on crustacea, it is primarily a filter-feeding herbivore. Setae of the thoracic legs of our adult E. pacifica are about 20-40-} apart; thus, the filtration apparatus is equipped to collect most marine diatoms. Calanus cristatus, Salpa spp., and Clio pyramidata are pelagic zooplankton which feed on suspended particles, principally phytoplankton. Calanus are mainly herbivores (Marshall and Orr, 1955), as are the cavolinid pteropods (Marshall, 1954; Yonge, 1926). Salpa spp. are in- discriminate feeders and principally phytoplankton grazers (Marshall, 1954; Yount, 1958; Foxton, 1961). 95 Trophic Level III This carid prawn is particularly common near Pasiphaea pacifica. the mouth of the Columbia River. Stomachs of 31 individuals were examined. Contents consisted of fragments of animals which were apparently dismembered and masticated before ingestion. Chitinous remains of crustacea were noted, including mandibles and eyes similar to those of Euphausia ifica. Several cephalopod beaks were also found. These observations, like those on other oceanic prawns (Chace, 1940), indicate that adult P. pacifica are carnivores. Only a few observations on the feeding habits Sergestes similis. of this mesopelagic prawn have been made previously (Barham, 1956). Our examination of the stomachs of these common animals revealed them capable of ingesting whole zooplankton. Entire'copepods were noted as well as fragments of larger euphausiids and prawns. Several fish scales were also present. Lampanyctus leucopsarus. Thirty-four stomachs of this lantern fish, the dominant mesopelagic fish taken in midwater trawl collections off Oregon, were examined. It feeds largely on euphausiids, calanoid copepods, and amphipods. Tactostoma macropus. fish were examined. Stomachs of 52 specimens of ,this stomiatoid Many were empty. Euphausiids and sergestid prawns occurred most frequently, but about half the total stomach contents by volume was due to the presence of several lantern fish. T. macropus appears to be intermediate between trophic level III and IV. 96 Trophic Level III-V Thunnus alalunga. Several hundred migratory albacore tuna were captured during the summer of 1962, from 25 to 50 miles off the northern Oregon coast, and stomachs of 62 of the tuna were examined. empty or less than one-quarter full. Most were Cephalopods composed about 75 percent of the bulk of the stomach contents, fish about 18 percent, and crustacea about 5 percent. Radioanalyses Gamma-ray spectra of organisms from several trophic levels taken at the same time and location are shown in Figure 1. Trophic level I is represented by a membrane filter through which surface sea water has been passed. Although chlorophyll a was present (1.56 mg/m3), the data do not indicate the percentage of particulate organic matter. The low amount of potassium-40 suggests that only a small quantity of inorganic material (probably as clay particles) was present. The K40 level on this filter was nevertheless higher than in filter samples from further offshore. Cerium-141, ruthenium-103, and zirconium-95 - niobium-95,2/ which are 2/ Our figures generally show the peak due to Zr95-Nb95 simply as but the techniques used do not permit a separation of Zr95 from its daughter, Nb95. 106 There is also some uncertainty with regard to Ru103 and but our evidence indicates a preponderance of Ru103 in these samples. No attempt was made to differentiate between Ce141 and particulate in sea water (Greendale and Ballou, 1954), were trapped by the filter, whereas zinc-65, which is ionic in sea water, was low even 500 TROPHIC LEVEL I 400 Cr51 200 Membrane filter I 300 Z r95 - Nb95 Ce141 100 K40 0 1500 1400 TROPHIC LEVEL II+ 1300 TROPHIC LEVEL III+ Lampanyctus leucopsarus TROPHIC LEVEL M+ Pasiphaea pacifica 50 CHANNELS Figure 1. Comparison of gamma emitters from several trophic levels. Organisms for all four spectra were collected 15 miles off Astoria on 5-6 April 1962. All trophic level II and III animals were from the same trawl sample. 97 though abundant in waters near the mouth of the Columbia River (Osterberg, Zn65 ions bound to particles would appear on the 1962a). Only those filter. The chemistry of chromium in sea water is uncertain, but the abundance of Cr51 (from Hanford) on filter samples (Fig. 1) indicates that a fair portion is either particulate or associated with particles. At trophic level II, represented by Euphausia pacifica (Fig. 1), two striking changes in the gamma spectrum are observed: lower Cr51, and higher Zr95-Nb95 Zn65 than on the filter. relatively Ru103, The and peaks indicate that either the particulate radionuclides are picked up directly filter-feeding euphausiids while feeding or they by may adsorb to larger particles, which are eaten. Concentration of these radionuclides does not appear to be due to adsorption on the surface of the zooplankton, however (Osterberg, et al., 1963). The Cr51 particles are either too small to be filtered out by the setae of the euphausiid or are selectively discriminated against, although in some cases small amounts of Cr51 are found in euphausiids. The lantern fish, L. leucopsarus, represents trophic level III. Cerium-141 and Zr95-Nb 95 are discriminated against-compared with the preceding trophic level (Fig. 1), even though E. pacifica was abundant in the stomachs of the fish, but Zn65 is still present. The spectrum of the carid prawn Pasiphaea pacifica (Fig. 1) closely resembles that of the lantern fish, reinforcing the conclusion from stomach analyses that both the prawn and the lantern fish are principally carnivores. Zn65 was the most conspicuous gamma emitter in the spectra of both animals. 98 Albacore tuna, taken 30 miles off Astoria, 10 August 1962, represented the highest trophic level examined (III-V). The spectrum of a single ashed sample of tuna liver indicated the virtual absence of particulate fission products, with the possible exception of Ru103 Concen- tration of Zn65 was exceedingly high compared with other peaks of the spectrum. A preference for other cations was also shown, with peaks due to cobalt-60, potassium-40, manganese-54, and cesium-137. DISCUSSION Spectra in Figure 1 show only the relative abundance of the different isotopes, and cannot be compared quantitatively, since no allowance has been made for sample size, radioactive decay, and efficiency factors. However, normalization of the data to obtain absolute concentrations of the various isotopes indicates that the relative spectra demonstrate real trends (Fig. 2). E. pacifica often contributes the greatest biomass in our trawl collections. samples. Its abundance permitted radioanalyses of about 150 euphausiid The year-round availability of this macroplankter and its affinity for radionuclides make it a useful biological standard with which to compare other organisms. Since environmental radioactivity varies with location3/, comparisons ideally should be restricted to organisms taken at 3/ Short period variations in fission product levels in euphausiids from a single location are small. Nine consecutive tows, made over a period of 8 hours, 50 miles off Newport, 11-12 April 1962, show the following averages and standard deviations: Zr95-Nb95, 13.6 1:1.2 picocuries/gram 141 17.5 T 2.8 pc/g, dry weight. and Filter per liter Euphausia pacifica Lampanyctus leucopsarus Pasiphaea paci f i ca K 40 Zn65 Zr- Nb95 ` Ru'03 Cry' Ce&4t 0 Figure 2. 5 20 PICOCURIES / GRAM WET WEIGHT 10 15 25 All organisms were Concentrations of gamma emitters from several trophic levels. collected 15 miles off Astoria on 5-6 April 1962. Note different units for filter sample. 99 the same time and sample. place. Our comparisons were usually from the same trawl They show that certain copepods, salps, and pteropods also con- With euphausiids, these animals represent centrate Zr95-Nb95 and the bulk of oceanic herbivores in our trawl samples. Similarities in the spectra of two animals from trophic level II are seen in Figure 3. herbivores. These spectra are somewhat typical of our oceanic That is, prominent peaks due to Zr95-Nb95 and Ce141 appear in the spectra of salp, copepod, pteropod and euphausiid samples taken in late 1961 and throughout 1962. When spectra of organisms from higher trophic levels are compared with the spectrum of E. form a general pattern. pacifica, the details described for Figure 1 That is, in every case there was a reduction in the Zr95-Nb95 and Ce141 peaks in predators relative to those observed in euphausiids. This discrimination was noted in the dozen or so instances when direct comparisons from the same sample were forced by a large number of analyses possible, of high trophic isms that invariably were low in fallout peaks. and was rein- level marine organ- This experience prompted us to consider Sergestes similis a predator on the basis of its spectrum, which shows a marked reduction in fission products compared with that of E. pacifica (Fig. 4). Subsequent stomach analyses verified this predic- tion. Ruthenium-103, presumably an anion (Lowman, 1960), was the only obvious non-cationic species regularly found in trophic levels III-V. It was present in the tuna a cotton rat (Sigmodon liver, hispidus) and has been reported in-the liver of (Kaye and Dunaway, 1962), although Chipman (1960) observed little uptake of Ru106 from digestive tracts of 100 menhaden (Brevoortia tyrannus). Cesium-137, which we also found in tuna liver, has been observed to concentrate slowly in some marine fish (Chipman, 1958; Baptist and Price, 1962). Pendleton and Hanson (1958) show that Cs137 is particularly concentrated at higher trophic levels. The affinity for zinc by all trophic levels, with the possible exception of trophic level I, makes Zn65 by itself a poor indicator of feeding relationships. We have found Zn65 in the predaceous squid (Onychoteuthis banksi) in surface waters, and in sea pens (Pennatulacea) attached to the bottom at 700 fathoms. This ubiquity of Zn 65 makes it of singular interest. The prominence of Zn65 in marine organisms has been mentioned by Lowman (1960), who stated that zinc and other transition elements were concentrated while cesium and strontium were discriminated against in marine animals, this situation being reversed on land. Zn65 is a prom- inent isotope in fresh-water fish from the Columbia River (Davis, 1958). Joyner (1962) found that carnivorous, marine, lagoon fish concentrate Zn65 to a greater extent than do herbivores. However, the levels of Zn65 reported in oysters (Watson, et al., 1961) near the Oregon-Washington coast were much higher than we found in any oceanic planktonic or nektonic organisms from the same general area, indicating that trophic level itself is not the sole criterion of ability to concentrate Zn65 CONCLUSIONS The presence of the Hanford reactors on the Columbia River makes Zn65 the most common gamma emitter in marine coastal waters. organisms from the Oregon All marine animals examined had accumulated this isotope. Zr95 Ce141 140 120 TROPHIC LEVEL ]I+ 100 L Euphausia pacifica 80 Zn65 60 40 K40 I z D o U 20 A 0 Zr95 J 70 I I- 60 L Ce141 0 TROPHIC LEVEL II+ 50 I Calanus cristatus 40 30 Zn65 K40 20 10 0 0 100 50 CHANNELS Figure 3. Comparison of spectra of euphausiids and copepods from the same miles off Astoria on 6 June 1962. sample, collected 105 The scatter in the lower spectrum is due to small sample size. Zr95 600 TROPHIC LEVEL II+ 500 400 Zn65 Ce 141 300 1 200 F- z Euphausia pacifica Ru 103 100 M 0 0 0 Zn 65 TROPH 1 C LEVEL l+ Sergestes simitis Ce141 0 Ru 103 Zr95 50 100 CHANNELS Figure. 4. Comparison of spectra of euphausiids and sergestids from the same sample, collected 25 miles off Newport on 6 November 1961. 101 Cr51, from the same source, was abundant only in filter samples repre- senting the first trophic level, and was not common at the higher trophic levels. The presence of particulate fission products Zr95-Nb95 and Ce appears to be a good criterion for distinguishing whether an animal is Rarely are these radioisotopes prominent in herbivorous or carnivorous. the spectra of predaceous animals in the sea, and then only in small amounts as compared to the same isotopes in marine herbivorous animals collected simultaneously. The highest trophic levels in the ocean, as evidenced by our tuna sample, almost completely discriminate against particulate fission products, but concentrate cations. Despite the chemical competition from potassium, which is abundant in sea water, Cs137 was present in tuna liver and in a carid prawn, both representa- tives of higher trophic levels. Radioactivity in the marine environment varies greatly with time and location. Very likely changes in stable trace elements also occur, although comparable data do not exist. Future work on marine food chains should include both measurements, so that specific activities can be determined. However, these local differences are minimized in our data by intercomparing organisms from the same trawl sample. We conclude that particulate radioactive fallout will be concen- trated at the second trophic level by filter-feeding plankton, but very little of this radioactivity will be present in the animals commonly utilized in the diet of man. On the other hand, neutron-induced Zn65 is more likely to enter the human food chain, but is one of the more innocuous radioisotopes, since it decays principally by electron capture and few ionizing particles are emitted. The discrimination against 102 particulate fission products tends to make the higher trophic levels of the marine food chain excellent sources of food in the event of high levels of radioactive fallout. ACKNOWLEDGMENTS We thank R. W. Perkins of General Electric's Hanford Laboratories for making his equipment available to us, and for his assistance with certain technical aspects of the gamma-ray spectrometry. This research was carried out under grants 1726 AT(45-1)1750 and AT(45-1) with the Atomic Energy Commission, Nonr 1286(02) with the Office of Naval Research, and Grant G 23103 with the National Science Foundation. 103 REFERENCES BAPTIST, J. P. and T. J. PRICE. 1962. cesium-137 by marine fishes. Accumulation and retention of U.S. Fish and Wildlife Service Bull. No. 206, pp. 177-187. BARHAM, E. G. The ecology of sonic scattering layers in the 1956. Monterey Bay area, California. Doctoral Dissertation Scr. Publ. No. 21,564, Univ. Microfilms, Ann Arbor, Mich., 182 pp. CHACE, F. A. Plankton of the Bermuda Oceanographic Expeditions. 1940. The bathypelagic caridean crustacea. IX. CHIPMAN, W. A. 1958. Zoologica, 25:117-209. Biological accumulation of radioactive materials. Proc. of the First Annual Texas Conf. on the Utilization of Atomic College Station, Texas, pp. 36-41. Energy, 1958. ------- 1960. Biological aspects of disposal of radioactive wastes in marine environments. 1959. 2. Conf. on the Disposal of Radioactive Wastes, Vienna, International Atomic Energy Comm., 1960, pp. 3-15. DAVIS, J. J., et al. 1958. Radioactive materials in aquatic and terres- trial organisms exposed to reactor effluent water. Second U. N. Int. Conf. on the Peaceful Uses of Atomic Energy, 1958, 1958. FOXTON, P. 1-8, Geneva, pp. 423-428, 1961. Salpa fusiformis Cuvier and related species. Discovery Rept. 32:1-32. GREENDALE, A. E. and N. E. BALLOU. 1954. by R. Revelle and M. B. Schaefer. USNRDL Document No. 436, cited 1957. radiation on oceanography and fisheries. Res. Council, Publ. No. 551, p. 12. The effect of atomic Natl. Acad. Sci.-Natl. 104 JOYNER, TIMOTHY. Effects of the biological specificity of zinc on 1962. the distribution of zinc-65 in the fish fauna of a coral atoll lagoon. Ph.D. Thesis, University of Washington, Ill pp. (unpublished). KAYE, STEPHEN V. and PAUL B. DUNAWAY. 1962. isotopes by herbivorous small mammals. LOWMAN, FRANK. test site. Bioaccumulation of radioactive Health Physics 7:205-217.- Marine biological investigations at.the Eniwetok 1960. Proc. of the Sci. Conf. on the Disposal of Radioactive Wastes, 1959, 2, Vienna, International Atomic Energy Comm., pp. 106-138. MacDONALD, R. Food and habits of Meganyctiphanes norvegica. 1927. J.. Mar. Biol. Assoc. U. K., 14:785-794. MARSHALL, N. B. 1954. Hutchins and Co,., Aspects of deep sea biology. Ltd., London, 380 pp. MARSHALL, S. M. and A. P. ORR. 1955. Calanus finmarchicus (Gunnerus). The biology of a marine copepod Oliver and Boyd, Ltd., Edinburgh,. 188 pp. NELSON, I. C., Ed. 1961. Evaluation of radiological conditions in the vicinity of Hanford for 1960. Hanford Atomic Prod. Oper..Rept. No. HW-68435, 115 pp. OSTERBERG, CHARLES. 1962a. Zinc-65 content in salps and euphausiids. Limnol. and Oceanog. 7:478-479. ------- 1962b. Science 138:- Fallout radionuclides in euphausiids. 529-530. ------- 1963. Radioactivity in oceanic organisms. Oregon State University, 125 pp. Ph.D. Thesis, (unpublished). 1963. OSTERBERG, CHARLES, LAWRENCE SMALL AND LYLE HUBBARD. in large marine plankton as a function of surface 883 -884. area. Radioactivity Nature 197: 105 PENDLETON, R. C. and W. C. HANSON. 1958. components of an aquatic community. Absorption of cesium-137 by Second U. N. International Conf. on the Peaceful Uses of Atomic Energy, 1958, 18, Geneva, pp. 419-422. PERKINS, R. W., J. M. NIELSEN, and R. N. DIEBEL. 1960. Total absorption gamma-ray spectrometers utilizing anticoincidence shielding. Rev. Sci. Instru., 31:1344-1349. PONOMAREVA, L. A. 1954. Euphausiids of the Sea of Japan feeding,on Rept. of the Acad. Sci. of the U.S.S.R. (Doklady) copepoda. Zoology 98(1):153-154. (Translated by V. 0. Pahn). WATSON, D. G., J. J. DAVIS, and W. C. HANSON. 1961. organisms along the Oregon and Washington coasts. Zinc-65 in marine Science 133: 1826-1828. YONGE, C. M. 1926. pteropods. YOUNT, J. L. Ciliary feeding mechanisms in the thecosomatous J. Linn. Soc. Zool., 36:417. 1958. Distribution and ecologic aspects of Central Pacific Salpidae (Tunicata). Pacific Sci., 12:111-130. 107 ZINC-65 IN EUPHAUSIIDS AS RELATED TO COLUMBIA RIVER WATER OFF THE OREGON COAST Charles Osterberg, June Pattullo and William Pearcy Abstract Most of the zinc-65 in the northeast Pacific Ocean originates in the Columbia River as a operations. in the result of the Hanford, Washington, nuclear reactor The Columbia River is also the principal source of fresh water region, so that at able far at sea because fate of the river certain seasons the plume of the river is detect- of its low salinity. water at In an effort to determine the sea, euphausiids from the ocean off Oregon were used as biological monitors. The Zn65 content of the euphausiids was measured by gamma-ray spectrometry, and the relationship between Zn65 content and the salinity distribution over the area was examined. Some correla- tions were apparent, although seasonal changes of Zn65 content in euphausiids were not great. Zn65 levels remained fairly high off Oregon even when Columbia River water was not evident as a low salinity plume. This indicates that water from the Columbia River, which accumulates as a plume off Oregon in the summer, influences the Zn65 content in animals in the area throughout the year. The long half-life of Zn65 in the mixed layer, the diurnal migra- tions of euphausiids, and the seasonal reversal of currents all may contrib- ute to the maintenance of this reservoir of coast. Zn65 in the ocean off the Oregon 108 INTRODUCTION The outflow of the Columbia River dilutes the salinity of the surface water in certain sections of the, northeast Pacific Ocean,. forming a shallow lens of low salinity water which over-rides the more dense oceanic water. This lens of water is known as the plume of the Columbia River, and, by definition, consists of water having a salinity of 32.5 0/00 or less. In summer, prevailing winds and currents carry this less saline, water southward, sometimes forming a pool approximately 100,000 square miles in area off the Oregon-California coast. In winter, surface waters are mixed to a greater depth, and Columbia outflow is diminished, while coastal rivers attain maximum flow. At this time the plume becomes smaller and less distinct and appears to be driven northward by the prevailing winds (Barnes et al. 1957; Anderson et al. 1961, 1962; Budinger et al. 1963). Water from the Columbia River is used to cool the nuclear reactors at Hanford, Washington. The intense neutron flux in the reactors activates certain trace elements in the coolant water. Stable zinc-64, for example, captures a neutron to become radioactive zinc-65. An estimated 38 curies per day of Zn65 passes Vancouver, Washington, en route to the sea. This rate of replenishment and the 245-day half-life of Zn65 lead to an equilibrium value of 14,000 curies of this isotope in the Pacific Ocean as a result of Hanford activities (Nelson ed. 1961). Zinc-65 has been found in sessile organisms along the Oregon-Washington coasts (Watson et al. 1961), in marine plankton at some distance from shore (Osterberg 1962, 1963), and in marine sediments near the Columbia River (Osterberg et al. 1963B). Zinc-65 in sea water is more difficult to detect because of the low concentrations that exist. It is the affinity of marine 109 organisms for zinc that makes them useful monitors of Zn65 The constant low-level radioactivity in the Columbia River and the inherent sensitivity of radiotracer techniques invite an attempt to trace river water at sea by using marine animals which concentrate Zn65 to readily measurable levels. If Zn65 is a characteristic unique to Columbia River water, and if marine organisms accurately reflect the radioactivity of their environment, then Zn65 in animals should be a good indicator of the presence of Columbia River water. This hypothesis is explored, using Euphausa pacifica as a monitor of Zn65. Zinc-65 con- centration in this abundant macroplankter is correlated with salinity distribution in an effort to delineate the seasonal distribution of Columbia River water in the ocean. METHODS Euphausiids were collected by using a six-foot Isaacs-Kidd midwater trawl at three east-west series of stations extending 5 to 165 nautical miles offshore from the Columbia River (Astoria), Newport, and Coos Bay. In one case, an additional series was made off Brookings, Oregon. Oblique tows from 200 meters to the surface, averaging 30 minutes duration, were made at night. The bulk of the samples usually consisted of euphausiids, with Euphausia pacifica predominating. Of the 194 midwater trawl collections throughout the period of study, 132 contained sufficient E. pacifica for radioanalysis (i.e., at least 40 g. wet weight). E. pacifica were separated from the preserved samples, freeze dried, and ground with mortar and pestle. Approximately 13 cc of the sample were 110 packed into 15 cc plastic spectrometer counting tubes, corked and sealed Samples were counted using General Electric's total absorp- with paraffin. tion anticoincidence spectrometer (see Perkins et al. 1960) at Hanford Laboratories, Washington. Data reduction Counting time was 30 minutes. and statistical analyses were made using an IBM 1620 computer. Filter samples from rivers were obtained with 5-inch diameter membrane filters (0.65 micron) and glass prefilters (Gelman Instrument Co.). Two filter units from each site were packed into each 15 cc counting tube. Samples (500 ml) of the cation exchange resins, which were used with river water, were counted in the well of a 9-inch crystal (Perkins 1961). Variability of Zn65 in Euphausiids Euphausiids concentrate Zn65 (Osterberg paring seasonal or geographic variations of short-term variability of Zn65 1962). Zn65, However, before com- some knowledge of the in E. pacifica is desirable. Euphausiids were taken in a series of nine repeated tows over a 16-hour period at a station 50 miles off Newport, 11-12 April 1962. The mean radiozinc content was 11.0 picocuries/gram dry weight, with a standard deviation of 1.5 pc/g. This indicates a fairly consistent concentration of Zn65, and variations which are small compared to seasonal or geographic trends. The Columbia River as a Source of Before the Zn65 Zn65 content of E, pacifica can be considered indicative of Columbia River water, it must be shown that fall-out comparatively insignificant amounts in this region. Zn65 occurs in Although Zn65 is not a fission product, it is associated with nuclear testing (Lowman 1960). 111 To check for Zn65 in euphausiids from areas relatively free from the effects of the Columbia River water, euphausiids found off Alaska and off San Diego Results show that a low- and Los Angeles, California, were analyzed.!/ The existence level background of Zn65 from fall-out does appear to exist. of a widespread background sence in of Zn65 has been further confirmed by its pre- a variety of foods in the eastern United States (Murthy et al. 1959), in fish and prawns in India (Chhabra and Hukkoo 1962), and in polar cod (Boreogadus saida) from Arctic waters (Osterberg and McCauley lab notes However, the quantity of Zn65 normally occurring off Oregon far exceeds that found in other areas of the world, except near nuclear testing sites. Additional evidence on the relative lack of fall-out Zn65 is furnished by comparisons of the radioactivity in the Columbia and Willamette Rivers, which join near Portland. late matter filtered detectable Fission products were present in the particu from the waters of both rivers, only in the Columbia River (Fig. 1). but Zn65 was readily There is a possibility that fall-out Zn65 might be in a form chemically different from the radiozinc induced in the Columbia River. Therefore, filtrates from both rivers were passed through cation exchange columns (Dowex-50) to remove Zn solution in the water. in This experiment also showed that the Zn65 peak was prominent only in the spectrum of Columbia River water. Since the Willamette River (and all of the other Oregon rivers that we have sampled-2/) is relatively free from Zn65, we conclude that most of this isotope found in the Oregon coastal region was produced at Hanford. This situation is 1/ We thank Frank Hebard, U.S. Fish-and Wildlife Service, for the Alaska sample, and Edward Brinton, Scripps Institution of Oceanography, for those collected off California. 2/ Nehalem, Nestucca, Siletz, Suislaw, Umpqua, Marys, N. Santiam, S. Santiam, Yamhill, Clackamas, Deschutes, Malheur, Owyhee, Burnt, Snake, John Day, Metolius, Rogue, Coquille, Alsea, and McKenzie Rivers. 112 subject to change, however, since each nuclear detonation is a passible new source of fallout RESULTS If Zn65 in Euphausiids and Related Salinity Distributions Zn65 in euphausiids reflects water at is sea, expected. the distribution of Columbia River then a negative correlation between Zn65 and surface salinity Figures 2-6 show amounts of Zn65 in euphausiids and salinity distributions at approximately the time when the euphausiids were collected. During July-August 1961 (Fig. 2), siids from Newport than in the sample from Columbia River. was more abundant in euphau- Zn65 Astoria, at the mouth of the Lower values, but relatively uniform with distance from shore, were found in euphausiids taken off Coos Bay. The salinity data, taken a month earlier, indicate a tongue of relatively fresh surface water extending at least as far south as the California border (salinity 32.5 0 /oo). This shallow lens of water is the plume of the Columbia River (Anderson et al. 1962, Budinger et al. 1963). During November 1961 (Fig. 3), low Zn65 values were found everywhere except at the station nearest the mouth of the Columbia River. from this station the addition, the south. (1962), Zn65 Seaward content of euphausiids decreased markedly. close to shore there was a gradual decrease in radioactivity to Again, our salinity data, as well as those of Anderson et al. indicate that the Columbia River plume extended south of Coos Bay. Figure 4 shows that in January Zn 65 was still present in appreciable quantities in euphausiids taken off Newport, but values at stations off Coos Bay, particularly near shore, were lower than in November. The low I Cr51 4000 COLUMBIA RIVER ( Corbett - Jan. 10, 1963) 3000 Zn65 C e 141 1 2000 1 Z r95 Mn54-56 A 1000 C S 137 Ru103 l ) K 40 SC 46 i I F- i WILLAMETTE RIVER Ce 141 ( Buena Vista Ferry 3000 - Jan. 10, 1963) Zr 95 2000 Zn65 1000 1 0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 CHANNELS Figure 1. Gamma-ray spectra of particulate material in two Oregon rivers. Note chromium-51 and zinc-65 peaks in the Columbia River spectrum. BROOKINGS Figure 2. Zinc-65 in euphausiids (bars) and salinity distribution (contours) during summer, 1961 (see text for dates). Numbers above bars indicate picocuries per gram dry weight of euphausiids. All stations where euphausiids were collected are indicated by bars or dots. Dots show stations where euphausiids were insufficient for radioanalyses. In no case where analysis was possible was Zn65 absent. 47 X05 \45 125 \65 DEPTH 50 IN 100 METERS 150 COOS BAY SALT N ITY 28 30%. 32.5%° Figure 3. Same as Figure 2 for November 1961. 13 ASTORIA 17 0 DEPTH N 50 100 METERS 200 COOS BAY SALINITY < Figure 4. 28 %o Same as Figure 2 for January 1962. 113 salinity of this inshore water was undoubtedly due to increased winter runoff from the nearby Umpqua and Coos Rivers, rather than to the Columbia plume. Zinc-65 values at the four stations nearest shore off Coos Bay may also have been reduced by admixture of water from the south (Davidson Current) which occurs frequently in this area at this season, whereas, water further offshore continues to flow from the north (H. 0. Publ. 570 1947; Sverdrup et al. 1942; Anderson et al. 1962). Anderson et al. (1962) point out that prevailing winds in winter probably carry the Columbia plume northward. However, low salinity and relatively high Zn65 values off Newport during January indicated the continued influence of plume water in this area. euphausiids from 125 miles On the other hand, off,Astoria contained little Zn65, although low salinity water was present there, We believe that this low salinity water off Astoria was not from the Columbia River, but instead was part of the general low-salinity belt (modified Sub-Arctic Water) extending to the Gulf of Alaska (Anderson et al. 1961). This conclusion is based both on the low Zn65 value and on the fact that winds that would drive the plume westward are absent. Exceptionally high values of Zn65 were found during very near the mouth of the Columbia River (Fig. 5). March-April 1962 These high Zn 65 values occurred at a time when the plume was "pooled" close to the mouth of the river by several months of strong winds from the south-west (Anderson et al. 1962). High radioactivity off Newport seems to be assoc- iated with water fresher than 32.5 0/0o in spite of the fact that the Columbia River plume is not thought to extend this far south during the spring (Anderson et al. 1962). 114 Figure Zn65 (July-August 6 1962) represents the same season as Figure was again lower off Astoria than off Newport or, during this year, even Coos Bay. Relatively high values of Zn65 in euphausiids were found off Brookings in the tongue of fresher water offshore, which again seems to be part of the Columbia River plume. Much lower Zn65 values were found in the four stations nearest the coast where the salinity was almost 34 0/oo (undoubtedly due to upwelling). In summary, the maximum value of Zn65 in E. pacifica, was found off the mouth of the Columbia River in spring, 1962, (Fig. 5). However, large amounts were found off the central coast of Oregon throughout the year. In summer, these high values were from euphausiids taken in the area of the Columbia River plume, as indicated by salinity measurements. It is surprising that values of Zn 65 were sometimes. higher off central Oregon than off the mouth of the Columbia River (Fig. 2, 4, and 6) Moreover, relatively high concentrations were found off southern Oregon as much as 250 miles from the Columbia (Fig. 6). find that seasonal variations in the It was particularly unexpected to Zn65 in euphausiids were not great, and that comparatively high values were found off Oregon in the winter, when the plume presumably extends northward and is absent off the Oregon Coast. Zn65 as an Indicator of Columbia River Water In the absence of fallout Zn65, the Zn65 which we measured in euphau- siids must be derived from Hanford water. activities, i.e., the Columbia River In interpreting the distributions of euphausiids containing Zn65 and the disparity between this and the location of the Columbia River 136 COOS BAY E 30% 32.5 % Figure 5. Same as Figure 2 for March-April 1962. BROOKINGE M, Figure 6. Same as Figure 2 for July-August 1962. 115 plume, we consider the following physical and biological factors: (1) variations in the amount of Zn65 introduced into the Columbia at Hanford, (2) amounts of Zn65 reaching the ocean, decay, (3) losses at sea due to nuclear (4) currents affecting the movement,of the plume, currents and vertical migrations on transport Zn65 in the mixed layer, and (7) f Zn 65, uptake and retention (5) effect of (6) half-life of of Zn65 by euphausiids. (1) Variations in the Amount of Zn65 Introduced into the Columbia at Hanford The quantity of but data are by the Zn65 entering the, Pacific Ocean undoubtedly varies, not available on the variations in the amount of Zn65 released reactors. Recent reports indicate that. improved techniques have reduced Zn65 output by about 25 percent (Schneller 1962), but the date of this change was Pacific Ocean not'given. for 1961 However, equilibrium values of Zn65 in the (Nelson, 1962) are unchanged from the previous year (Nelson, 1961), which indicates that this reduction occurred after 1961. Perhaps the lower values off Newport in the summer, of 1962 (Fig. 6), com- pared with the summer of 1961 (Fig. 2), reflect this reduction. (2) Amounts of Zn65 Reaching the Ocean Because of the transitory nature of,Zn65, the amount which reaches the ocean will be affected by the time it takes water to flow from Hanford to the sea. Dams on the Columbia, by increasing this time, allow more decay and thus tend to reduce the output of Zn65 into the ocean. Another factor is uptake by the biota of the river (Davis and Foster, no date). We have no quantitative estimates of these complex factors, but believe their bearing on the problem of tracing the Columbia River is negligible 116 compared with other uncertainties. (3) Losses at Sea Due to Nuclear Decay If only physical 245 days. decay is considered, half of the Zn65 is lost in However, steady state conditions for Zn65 exist in the ocean off Oregon in that decay is approximately balanced by replenishment of Zn65 from the Hanford operations (Nelson 1961). the river flows southward in the ocean (as it appears summer), decay will result in time and distance. But as the plume of a gradual decrease to do in the in radioactivity with If this were the only loss, the transit time of the water could be computed by measuring the change in Zn65 concentration in the water. Let us assume for the moment that measurement of Zn65 in euphausiids is equivalent to measurement of Zn65 in water, The spatial and temporal changes in Zn65 concentration in euphausiids off Astoria are very large. Therefore, interpretation of this series of samples would be difficult. However, Zn65 values for the Newport and Coos Bay series are less variable and more confidence can be placed in averages. The average of the Coos Bay series was always lower than that of the Newport series. If the lower amounts of Zn65 at Coos Bay are attributed entirely to nuclear decay as the water flows from north to south, the travel time ranged from 34 days in November 1961 to 309 days in summer 1961. Anderson et al. (1961) state that "the current drift (of the southerlysetting California Current) as computed from the dynamic topography of the sea surface is small, with average values of the order of 5 cm/sec or less and maxima up to 20 cm/sec (10 miles per day) in the spring and 117 summer." at these rates Zn65 carried by the southward. drift would cover the distance between Newport and Coos Bay in about 8 to 40 days;'this is considerably less than the times computed from the rate of physical decay of Zn65 This leads us to conclude that, although radioactive decay is undoubtedly a factor effecting a decrease in Zn65 concentration as the water flows from the latitude of Newport to that of Coos Bay, it cannot be the only or even the most important factor. (4) Currents Affecting the Movement of the Plume The plume itself is a very shallow phenomenon and may be thought of as a thin anticyclonic gyre embedded surface currents. Currents generally southward in in the prevailing or "steering" within 100 miles of, the Oregon coast are summer, and, less consistently, northward in winter (H. 0. Publ. 5,0 1947). The outer edge of the plume generally Most of these currents are weak lies in the prevailing southward drift., with speeds of less than half a knot. In addition to the general coastal drift, offshore motion of the upper layers is present during summer when upwelling occurs along the coast (Pattullo et al. in prep.). Consider the behavior of a hypothetical plume that leaves the mouth of the Columbia as a discrete discharge during the month of June. It would drift southward (at about 5 cm/sec at the core), and seaward, undoubtedly spreading and becoming saltier with time. Zinc-!65 concentration in the plume would be diluted by mixing with salt water, but,, on'the whole, would tend to. remain near would disappear as a result the surface.. of turbulent Eventually, the plume mixing. 118 Although grossly oversimplified, this general pattern agrees with the behavior of the plume as described by Anderson et al. (1961, 1962). The principal difference between the simplified "model" and the real river plume is that the flow is not a single burst in June but has some persistence during the summer months. simply detach itself from the river Therefore, the plume body does not as a fixed quantity of fresh water, but increases in freshwater content, as well as in area, during the summer. Winter mixing is such, however, that the plume completely loses identity each year and can be considered an annually recurring event. Occasionally a secondary maximum in Columbia River flow is observed during winter. This does not reinforce the old plume body off the Oregon coast, normally, but is driven northward along the Washington coast instead. Surface drift is directly influenced, apparently, by winds. summer the North Pacific high pressure area is well developed. During The Oregon coast lies under the influence of the northwesterly winds at the northeastern edge of the high. During winter, this high retreats to the southwest and the southeastern sector of the Aleutian low frequently dominates the area; this leads to southerly winds. However, at any particular time the meteorological conditions may not conform to this generalized pattern, and deviations from prevailing flow can be expected to result. During winter southwesterly winds result in a "pooling" of river water along the shore adjacent to the mouth of the Columbia River (Anderson et al. 1962). is the'area At this season, river discharge is low and so of the "pool" compared with the summertime pattern. However, the Zn65 content is high because of the high concentration of river water in a restricted area. 119 65 (5) Effect of Currents and Vertical Migrations on the Transport of Zn If all the Zn65 released by the Columbia River were retained entirely within the plume, its study would involve only surface flow and transport. Since euphausiids and other vertical migrants are found to concentrate Zn65, however, we must consider the motions of the euphausiids and the currents that affect them, as well as the currents which affect only the plume. Euphausia pacifica is known to make daily vertical migrations (Esterly 1914, Tucker 1951, Barham 1956, Brinton 1962). Off Oregon, E. It is pacifica is abundant in catches from the upper 200 m at night. uncommon during the day, presumably descending into deeper water. This migration takes euphausiids well below the Columbia River plume water, and, in fact below the temperature-salinity gradients that separate the low density waters of the surface mixed layer from denser waters below. Thus, biological transport of zinc across the pycnocline by such animals as E. pacifica is undoubtedly more important than transport through this layer by physical processes (see Ketchum and Bowen 1958). Direct measurements of currents indicate that the pycnocline (at 100 to 200 m) is a region of velocity shear. Recent data show that throughout the year surface waters have had southward velocity relative to the deeper waters (Pattullo et al. in prep.). animals such as Therefore, for vertically migrating euphausiids, which reside in deep waters during the day, the net drift to the south would be less than that of the plume. an atom of Zn65 in a euphausiid would move less rapidly out of the local area than one floating freely in the plume, and concentration of euphausiids Hence could be expected to lag behind the plume body. Zn65 in 120 Seasonal variations in subsurface currents also may affect euphausiid distribution. During wintertime, both subsurface (at least to 200 m) and surface flow are probably slightly northward (Pattullo et al. in prep.). This would tend to return to Oregon coastal areas euphausiids and Zn65 that had drifted southward during the summer. Consequently, the patterns of currents and vertical distributions of E. pacifica, both on a daily and seasonal basis, appear to reduce the dispersion of Zn65 from the local area off Oregon and may contribute to the absence of large seasonal vari- ations of this radionuclide in euphausiids. The effects of vertical mi- grations on the horizontal distributions of Antarctic euphausiids have been described by Hardy and Gunther (1935) and Mackintosh (1937). They also show that both daily and seasonal vertical migrations may affect the retention of a population within a geographic area. (6) Half-life of Zn 65 in the Mixed Layer Budinger et al.(unpublished) report that mixing of the plume with salt water occurs in such a way that radionuclides in the plume tend to remain in the surface layers. That is, salt water mixes vertically upward into the overlying plume, while little fresh water is lost through the resulting halocline. Diffusion occurs along the outer edges of the plume, and often "cells" of fresher water become separated from the plume (Anderson et al. 1962). Thus, radioactivity per unit volume of surface water in the plume decreases with distance from the mouth of the river even though little physical loss occurs vertically. Zn65 Thus most of the from the Columbia River is retained in the euphonic zone where it readily enters into food chains. 121 Dilution of river water at sea makes measurement of Zn65 extremely difficult, except in marine organisms, of our non-biological measurements which concentrate the zinc. Most therefore, been made in near have, the Columbia estuary, where levels are much higher. or, There, Zn65 is found in several conditions: (1) as ionic zinc in the filtrate (concentrated on cation and chelating resins), filtering surface water (2) attached to particles (collected. by through membrane filters), and (3) in organisms. The concentration of ionic zinc is low and diminishes rapidly seaward, so that it is not generally detectable by our techniques beyond 25 miles from Zn65 the river's mouth (Osterberg and Cutshall lab notes)... in or on particulate material can be detected farther at.sea.= -A small;.. but distinct Zn65 peak appeared in the spectrum of a filter through which had been passed 109 liters of surface sea water from 45 miles off Astoria. Zn65 in marine organisms remains easily detectable over the entire area of our with observations, organisms. This indicates that much of the Zn65 is associated Major loss from the mixed layer therefore may be caused by excretion and predation below the thermocline by. animals undertaking diurnal migrations. (7) Uptake and Retention of Zinc-65 The biological it is Euphausiids half-life of Zn65 in long, as it is in some marine E. pacifica is not known.. If fish (Chipman et al. 1958), radio zinc may be accumulated and retained for several months. This would help explain the small difference in Zn65 content between winter (when the plume is not evident) and summer present). (when the plume is obviously 122 Zinc is a constituent of several enzyme systems (Vallee 1957) and marine organisms probably have a physiological need for it (Joyner 1962). Metabolic demand for this element, which was the most common artificial gamma emitter in marine animals off the Oregon coast (Osterberg et al, in press), may be greater during certain seasons of the year, or for certain stages in the life history of euphausiids than at other times. Davis and Foster (no date) show that fluctuations in the radioactivity of freshwater minnows (Richardsonius dependent. balteatus), for were strongly temperature- example, They attribute the 75-fold increase in concentration of radio- isotopes in the fish between winter and late summer to different metabolic requirements, due largely to temperature changes. High Zn65 values near the mouth of the Columbia River in spring and somewhat increased values in the fall correspond fall plankton E. pacifica grazes on "blooms." 1954), from which it appears to obtain Zn65 roughly to spring and phytoplankton (Ponamereva (Osterberg et al. 1963a). Thus, a relationship is suggested between phytoplankton abundance and increased Zn65 content in either to increased growth of the This relationship may be due content per unit of phytoplankton during rapid Zn65 population, Foster and Davis euphausiids. or to increased grazing rates (1955) state that by the euphausiids. freshwater diatoms reach equilib- rium with reactor effluent water in about an hour. Marine diatoms also appear to concentrate radionuclides rapidly, since maximum Zn65 values are found in filter samples from nearest the mouth of the Columbia River (Osterberg and Curl lab notes). Nevertheless, the highest in euphausiids was generally found off Newport, Zn65 content well to the south. This suggests that radioactivity of euphausiids is dependent on the time that 123 the animal has spent in water containing Zn65. That is, a long period is required for maximum concentration to be reached in the euphausiid. In contrast, near the mouth of the Columbia River, Zn65 content of mussels (Mytilus spp.) was found to be generally higher than that of euphausiids, but a similar comparison at Coos Bay showed less in mussels (Watson et al. 1961). The smaller gradient of Zn65 concentration in euphausiids with distance is probably typical of plankton which drift with the moving water. On the other hand, the water surrounding sessile organisms is continually renewed with changes of wind and tide.. CONCLUSIONS Surface salinity patterns off the Oregon.coast change in,response to the position of the Columbia River plume, which in turn is influenced by a seasonal change in prevailing winds. While Zn65 is shown to be associated with the Columbia River, seasonal variations of euphausiids are not Zn65 in marked, and the amount of Zn65 does not diminish greatly in the absence of the plume in winter. Vertical stability in the ocean in this area (which generally precludes mixing-to great depths), seasonal current reversal and the migratory habits of euphau- siids appear to maintain high levels of the year. Zn65 in euphausiids throughout A long biological half-life of Zn65 would also help maintain high Zn65 levels in the winter. Maximum concentration of Zn65 in euphausiids occurs close to the mouth of the river in spring when the river's effluent is driven inshore and "pooled" by winds from the higher off Newport, to the southwest. south, Otherwise levels. are generally perhaps due to a longer residence time 124 in plume waters and time lag in Zn65 incorporation. Levels are lower.off Coos Bay due to dilution and spreading of the plume and to decay of . Two prominent local phenomena, the inshore wintertime current from the south (Davidson Current) and summertime upwelling off Brookings, are both reflected by lower Zn65 concentrations in euphausiids from those areas. The great affinity of marine organisms for zinc and sensitivity of modern gamma-ray spectrographic techniques make Zn65 in euphausiids easy to measure. Unfortunately, however, use of biological organisms as monitors introduces many uncertainties. The most important is that we do not know how accurately the radioactivity of the euphausiid reflects the radioactivity of his immediate environment. This difficulty is com- pounded if variations in stable zinc occur; local variations seem likely because of the affinity of marine organisms for zinc. However, no com- parable data exist for stable zinc. Another approach to the problem might be through the study of a radioisotope that is not utilized extensively by marine organisms and thus would be a more conservative indicator of plume waters. Chromium-51 meets this requirement and furthermore is even more abundant in Columbia River water than is Zn65 (Fig. 1) (Nelson 1961). However, this lack of biological concentration of Cr51 makes it especially difficult to measure, except near the mouth of the river before much dilution occurs. Nevertheless, we are striving to improve our techniques so that radioactive elements in the Columbia River can become a more useful tool in physical and biological studies of the plume and its effects in the Oregon coastal region. It is this low-level radioactivity which dis- tinguishes the plume from other sources of fresh water in the area, and seems to offer the most promise as a tracer of the river water at sea. 125 ACKNOWLEDGMENTS We thank R. W. Perkins of General Electric's Hanford Laboratories for allowing us to use his equipment, and for his assistance with certain technical aspects of the gamma-ray spectrometry. to Harriet Van Arsdale, We, are indebted I. Lauren Larsen, Norman Cutshall, and the captain and crew of the R/V ACONA for their work in the laboratory and at sea. We gratefully acknowledge the use of salinity data made avail- able to us by Bruce Wyatt. This research was carried out under grants AT(45-;1)1750 and AT(45-1)1726 with the Atomic Energy Commission, Nonr 1286(02) with the Office of Naval Research, and grant GP-622 with the National Science Foundation, 126 REFERENCES ANDERSON, G. C., C. A. BARNES, T. F. BUDINGER, C. M. LOVE, AND D. A. McMANUS. 1961. The Columbia River discharge area of the northeast Pacific Ocean, a literature survey. pp. University of Washington, 99 (U.S.A.E.C. Ref. M61-25) Mimeographed. ANDERSON, G. C., K, BANSE, L. K. COACHMAN,'J. S. CREAGER, M. G. GROSS, AND D. A. McMANUS. Columbia River effects in the northeast 1962. University of Washington,17 pp. Pacific. (U.S.A.E.C. Ref. M62-5) Mimeographed. BARHAM, E. G. 1956. The ecology of sonic scattering layers in the Monterey Bay area, California. Doctoral Dissert. Ser. Publ. No. Univ. Microfilms, Inc., Ann Arbor, Mich., 182 p. 21,564. BARNES, C. A. AND R. G. PAQUETTE. 1957. Circulation near the Washington Proc. of the 8th Pacific Sci. Cong., 1953. coast. BRINTON, EDWARD. 1962. 3:585-608. The distribution of Pacific euphausiids. Bull. of the Scripps Institution of Oceanography of the University of California. pp. 51-269. BUDINGER, T. F., L. R. COACHMAN, AND C. A. BARNES. 1963. Description of the Columbia River plume and certain aspects of the mixing of river water in the sea (abstract). Trans. Am. Geophys. Union 44 (1) :208. BUDINGER, T. F. (unpublished). In paper presented at AGU Meeting, Seattle, Washington, November 2, 1962. CHHABRA, A. S. AND R. K. HUKKOO. activity in foodstuffs. 1962. Natural and fallout gamma Health Physics 7:151-155. 127 CHIPMAN, W. A., T. R. RICE, AND T. J. PRICE. 1958. Uptake and accum- ulation of radioactive zinc by marine plankton, fish, and shellfish. U. S. Fish and Wildlife Serv., Fish Bull. 135, 58:279-292. DAVIS, J. J. AND R. F. FOSTER. No date. Bioaccumulation of radioiso- Biology Operation, Hanford topes through aquatic food chains. Laboratories, General Electric Co., 15 pp. ESTERLY, C. 0. Mimeographed. The vertical distribution and movements of the 1915. Schizopoda of the San Diego region. Univ. Calif. Publs. Zool. 13:123-145. FOSTER, R. F. AND J. J. DAVIS. 1955. substances in aquatic forms. ful Uses of The accumulation of radioactive Proc. of the Intern.. Conf. on Peace- Atomic Energy, 13 (P/280):364-367. HARDY, A. C. AND E. R. GUNTHER. 1935. whaling grounds and adjacent The plankton of the South Georgia waters, 1926-27. Discovery Rep. 11: 1-146. Atlas of surface currents, Northeastern HYDROGRAPHIC OFFICE, U. S. Navy. Pacific Ocean. JOYNER, TIMOTHY. H. 0. Publ. 570. Washington, D. C., 1947. Effects of the biological specificity of zinc 1962. on the distribution of Zn65 in the fish fauna of a coral atoll Ph.D. Thesis. lagoon. Seattle, University of Washington. 111 pp. (Mimeographed). KETCHUM, B. H. AND V. T. BOWEN. in the sea. test site. Distribution of radioisotopes Second U. N. Intern. Conf. on the Peaceful Uses of Atomic Energy, 1958. LOWMAN, FRANK. 1958. 1960. Geneva. 18:429-433. Marine biological investigations at the Eniwetok Disposal of Radioactive Wastes, Vienna. 2:106-138. 128 MACKINTOSH, N. A. 1937. macroplankton. The seasonal circulation of Antarctic Discovery Rept, 16:367-412. MURTHY, G. K., A. S. GOLDIN, AND J. E. CAMPBELL. C. (Ed.). Evaluation of radiological conditions in 1961. Hanford Atomic Operations Rept. the vicinity of Hanford for 1960. No. HW-68435. ------- Zinc-65 in Science 130:1255-1256. foods. NELSON, I. 1959. 1962. 151 pp. Evaluation of radiological conditions in the vicinity of Hanford for 1961. Hanford Atomic Operations Rept. No. HW-71999. 250 pp. OSTERBERG, CHARLES. 1962. Zn65 content in salps and euphausiids. Limnol. and Oceanog. 7:478-479. ------- 1963. Radioactivity in oceanic Corvallis, Oregon State University. films organisms. Ph.D. Thesis. 134 pp. University Micro- (Order No. 63-3787). ------, LAWRENCE SMALL AND LYLE HUBBARD. 1963a. Radioactivity in large marine plankton as a function of surface area. ------, L. D. KULM AND JOHN V. BYRNE. sediments near the Columbia River. 1963b. Nature 197:883-884. Gamma emitters in marine Science 139:916-917. ------, WILLIAM G. PEARCY AND HERBERT CURL, JR. (In press). Radio- activity and its relationship to oceanic food chains... PATTULLO, JUNE, BRUCE WYATT AND PAUL MAUGHAN. (In prep.). Direct measurements of currents in the upper 200 m of water off the Oregon coast. 129 PERKINS, R. W., J. M. NIELSON, AND R. N. DIEBEL. 1960. Total absorp- tion gamma-ray spectrometers utilizing anticoincidence shielding. Rev. Sci. Instr. 31:1344-1349. PERKINS, R. W. 1961. A large well crystal for the direct measurement of trace amounts of radioisotopes in environmental samples. Health Physics 7:81-86. PONAMEREVA, L. A. 1954. Dokl. Akad. Nauk. SSSR 98:153 (Translated by V. 0. Pahn). SCHNELLER, MARVIN. sion 1962. Reactor Operations Branch - Production Divi- - U.S,A.E.C., Richland, Washington. (Personal Communication). SVERDRUP, H. U., M. W. JOHNSON, AND R. H. FLEMING. Prentice-Hall, N. J. TUCKER, G. H. 1951. 1957. The oceans. 1087 pp. Relation of fishes and other organisms to the scattering of underwater sound. VALLEE, B. L. 1942. J. Mar. Res. 10:215-238. Zinc and its biological significance. Archives of Industrial Health. 16:147-154. WATSON, D. G., J. J. DAVIS, AND W. C. HANSON. 1961. Zinc-65 in marine organisms along the Oregon and Washington coasts. 133:1826-1828. A.M.A. Science 131 PUBLICATIONS AND PAPERS Papers Published Osterberg, Charles. Fallout radionuclides in euphausiids. 1962. Science 138:529-530. (With AT (45 -1)1750) ----- -- 1962. Zn65 content of salps and euphausiids. 1:478-479. Limnol. Oceanogr. (With AT(45-1)1750) ------, Lawrence Small and Lyle Hubbard. 1963. Radioactivity in large marine plankton as a function of surface area. Nature 197:883-884. (With AT (45 -1)1750 Pearcy, W. G. 1962. Egg masses and early developmental stages of the Scorpaenid fish, Sabastolobus. ------, and Gilbert L. Voss. northeastern Pacific. ------, 1963. 1963. J. Fish. Res. Bd. Canada 19:1169-1173. A new species of gonatid squid from the Proc. Biol. Soc. Washington 76:105-111. Distribution of oceanic cephalopods off Oregon, U.S.A. Proc. XVI Int. Congress of Zool. 1:69. Papers Accepted for Publication Osterberg, Charles, W. G. Pearcy and Herbert Curl, Jr. Radioactivity and its relationship to oceanic food chains. J. Mar. Res. AT(45-1)1750) (With See Section 6 of this report. Papers Submitted for Publication Osterberg, Charles, June Pattullo and William Pearcy. Zinc-65 in euphausiids as related to Columbia River water off the Oregon coast. Limnol. Oceanogr. (With AT(45-1)1750) See Section 7 of this report. 132 Pearcy, W. G. Preliminary observations in the distribution of mesopelagic fishes off Oregon. J. Mar. Res. (See Section 1 of this report.) In Preparation Pearcy, William G. Species composition and distribution of pelagic cephalopods from the Pacific Ocean off Oregon. (See Section 2 of this report.) Pearcy, William G. and Charles Osterberg. tion of Zn65 and Zr95-Nb95 Notes on the vertical distribu- from oceanic animals. (With AT(45-1)1750) See Section 5 of this report. Pearcy, W. G. Vertical distribution of numbers and biomass of mesopelagic fishes with an improved Isaacs-Kidd midwater trawl. (See Section 4 of this report.) Papers Delivered at Scientific Meetings Pearcy, W. G. Intermediate forage animals. Northwest Pacific Oceanog- raphers, Vancouver, B. C., February 19, 1962. Pearcy, W. G. Distribution of bathypelagic fishes over the continental slope off Oregon. Amer. Soc. Limnol. Oceanogr. AIBS Meeting, Corvallis, Oregon, August 28, 1962. Pearcy, W. G. Deep-sea animals off the Oregon coast. Mar. Biol, Soc. of Oregon, Portland, Oregon, March 30, 1963. Pearcy, W. G. Preliminary observations on the vertical distribution of micronekton off Oregon, Pac. Div. Amer, Soc. Limnol. Oceanogr., Stanford, California, June 20, 1963.