PROCEEDINGS THE ROYAL S O C I E T Y
advertisement

Downloaded from rspb.royalsocietypublishing.org on January 28, 2014
PROCEEDINGS THE ROYAL
O F
SO C IE T Y
B IO L O G IC A L
SCIENCES
How superdiffusion gets arrested: ecological encounters
explain shift from Levy to Brownian movement
Monique de Jager, Frederic Bartumeus, Andrea Kölzsch, Franz J. Weissing, Geerten M. Flengeveld,
Bart A. Nolet, Peter M. J. Flerman and Johan van de Koppei
Proc. R. Soc. S 2014 281 , 20132605, published 13 November 2013
Supplementary data
"Data Supplement"
http://rspb.royalsocietypublishing.org/content/suppl/2013/11/12/rspb.2013.2605.DC1 .h
tml
References
T his artic le cites 29 artic les, 8 o f w h ich can be a c ce ss ed free
Qopen access
Subject Collections
http://rspb.royalsocietypubiishing.Org/content/281/1774/20132605.full.html#ref-list-1
This article is free to access
Articles on similar topics can be found in the following collections
ecology (1536 articles)
R eceive free em ail alerts w hen new articles cite this article - sign up in the box at the top
rig ht-ha nd corn er or the article or click here
To subscribe to Proc. R. Soc. B g o to : http://rspb.royalsocietypublishing.org/subscriptions
Downloaded from rspb.royalsocietypublishing.org on January 28, 2014
PROCEEDINGS
O F ----------
TH E ROYAL
SOCIETY
rspb.royalsocietypublishing.org
Research
3 ®
CrossMark
Cite this article: d e Jager M, Bartum eus F,
Kölzsch A, W eissing FJ, H engeveld GM, N olet
BA, Herman PMJ, van d e Koppel J. 2 0 1 4 How
superdiffusion g e ts arrested: ecological
encou nters explain shift from Lévy to Brownian
m ovem en t. Proc. R. Soc. B 281: 20132 605.
http ://d x.doi.O rg/10.1098/rsp b .2 0 1 3.2605
Received: 4 O ctober 2013
Accepted: 17 October 2013
Subject Areas:
ecology
Keywords:
Brownian m otion , Lévy w alk, anim al
m o vem en t, M ytilus edulis, search efficiency,
resource density
How superdiffusion gets arrested:
ecological encounters explain shift from
Lévy to Brownian movement
M onique de Jager1-2-4, Frederic Bartum eus5, Andrea Kölzsch6-7, Franz
J. W eissing2, G eerten M. H engeveld6-7, Bart A. N olet6-7, Peter M. J. H erm an1
and Johan van de K oppei1-3-7
1S p atial Ecology D e p a rtm e n t, Royal N e th e rla n d s In s titu te fo r Sea R esearch (NIOZ), PO Box 1 4 0 , 4 4 0 0 AC V erseke,
T he N e th e rla n d s
^T heo retical B iology G roup, a n d C o m m u n ity a n d C o nserv ation Ecology G roup, C entre fo r Ecological a n d
Ev olutio nary S tu d ie s, U niversity o f G ro n in g en , N ijen b o rg h 7,
9 7 4 7 AG G ro n in g e n , T he N e th e rla n d s
i n s t i t u t e o f In te g ra tiv e Biology, ETH Zürich, U n iv e rsita e tstra sse 16, 8 0 9 2 Zürich, S w itz erla n d
5 C e n te r fo r A d van ced S tu d ie s o f B lanes (CEAB-CSIC), A ccès Cala S a n t Francesc, 1 4 ,1 7 3 0 0 B lanes, G irona, Spain
d e p a r t m e n t o f A nim al Ecology, a n d 7 P ro ject G roup M o v e m e n t Ecology, N e th e rla n d s In s titu te o f Ecology
, PO Box 5 0, 6 7 0 0 AB W a g e n in g e n , T he N e th e rla n d s
Ecological theory uses Brownian m otion as a default tem plate for describing
ecological m ovem ent, despite lim ited m echanistic underpinning. The general­
ity of Brownian m otion has recently been challenged b y em pirical studies that
highlight alternative m ovem ent patterns of anim als, especially w h en foraging
in resource-poor environm ents. Yet, em pirical studies reveal anim als m oving
in a Brownian fashion w h en resources are abundant. We dem onstrate that
Einstein's original theory of collision-induced Brownian m otion in physics
provides a parsim onious, mechanistic explanation for these observations.
Here, Brownian m otion results from frequent encounters betw een organism s
in dense environm ents. In density-controlled experiments, m ovem ent patterns
of m ussels shifted from Levy tow ards Brownian m otion w ith increasing den ­
sity. W hen the analysis w as restricted to moves n o t truncated b y encounters,
this shift d id n o t occur. U sing a theoretical argum ent, w e explain that any
m ovem ent pattern approxim ates B rownian m otion at high-resource densities,
provided that m ovem ent is in terru p ted u p o n encounters. Hence, the observed
shift to Brownian m otion does n o t indicate a density-dependent change in
m ovem ent strategy b u t rather results from frequent collisions. O u r results
em phasize the need for a m ore mechanistic use of Brownian m otion in ecology,
highlighting that especially in rich environm ents, Brownian m otion em erges
from ecological interactions, rather th an being a default m ovem ent pattern.
Author for correspondence:
M onigue d e Jager
e-m ail: m onigue.deJager@ usys.ethz.ch
Electronic supp lem en tary material is available
at h ttp ://d x.d oi.org /10.1098/rsp b .2013.260 5 or
1. Introduction
Traditionally, ecologists ap ply Brow nian m otion an d diffusive dispersal as
default m odels for anim al m ovem ent [1,2], b o th at in dividual a n d at population
levels [3-5]. Recently, how ever, em pirical studies have show n th at anim al m ove­
m ent can strongly deviate from Brow nian m otion [6], revealing superdiffusive,
Lévy-like m ovem ent in resource-poor environm ents, b u t stan d ard Brownian
m otion w h en resource availability is h igh [7-11]. A nim al ecologists have
explained this change from Lévy to Brow nian m otion by an active shift in indi­
v idu al m ovem ent strategy, reflecting the assum ption that different m ovem ent
strategies are optim al u n d er different environm ental conditions [10-13]. In h et­
erogeneous, resource-poor environm ents, Lévy m ovem ent w ill typically b e m ore
efficient th an a Brownian w alk because it provides faster dispersal an d prevents
revisiting the sam e sites [14]. In resource-rich environm ents, a Brownian w alk
m ay be equally or even m ore efficient th an a Lévy w alk, because large steps
via http://rspb.royalsocietypublishing.org.
© 2 01 3 The A u th o rs. P u b lish ed by th e Royal S ociety u n d e r th e te r m s o f th e C reative C o m m o n s A ttrib u tio n
Royal S ociety P u b lis h in g
License h ttp ://c re a tiv e c o m m o n s .O rg /lic e n s e s/b y /3 .0 /, w h ich p e rm its u n re stric te d u se , p ro vided th e o riginal
a u th o r a n d so u rce a re cre d ite d .
Downloaded from rspb.royalsocietypublishing.org on January 28, 2014
2ae
(2 . 1)
w here a is the length betw een position i an d i + 1, b is the
length betw een position i — 1 an d i + 1 an d c is the length
betw een positions i — 1 an d i. W henever a w as larger than
a threshold angle aT, a new step is considered to start. Follow­
ing Turchin's approach [24], w e used aT = tt/ 5 for our step
length calculations, as this value m inim ized autocorrelation
betw een subsequent turns. Using other threshold angles did
not change our conclusions.
We studied the changes in the statistical properties of the
observed m ovem ent p attern b y recording 10 individual
m ovem ent trajectories for five different density treatm ents
each (0, 1.3, 2.0, 3.3 an d 5.2 kg m -2 , approx. 1, 950, 1550,
2500 an d 3850 m ussels m -2 ) d u rin g the initial 300 m in of p at­
tern form ation [23]. W hen a m ussel encountered an obstacle,
for exam ple a conspecific, it w as forced to truncate its step,
w hich w ill probably alter the properties of the m ovem ent
pattern. We u sed the com plem entary cum ulative distribution
function (CCDF) of the observed step lengths of each individ­
ual m ussel in the five density treatm ents to illustrate the
observed m ovem ent patterns. This CCDF is a preferred
m ethod for fitting pow er distributions as it provides a
m ore reliable representation of m ovem ent pattern s th an
other portraying m ethods [3]. For each step length /, the
CCDF(/ ) of the observed step lengths in each density treat­
m ent indicates the fraction of step lengths that w ere at least
as long as I. U sing m axim um -likelihood m ethods, w e esti­
m ated the scaling exponent /i of a pow er-law step length
distribution,
(2 .2 )
2. Experiments
(a) M eth o d s
U sing mesocosm experiments, w e investigated how mussel
m ovem ent patterns are affected b y m ussel density. Young
blue mussels (M. edulis) of approxim ately 1.5 cm in length
w ere obtained from w ooden wave-breaker poles on the beaches
near Vlissingen, The N etherlands (51°46' N , 3°53' E). After care­
ful separation an d cleaning, the mussels w ere kept in containers
an d fed live cultures of diatom s (Phaeodactylum tricornutum)
w here I is the step length an d /nlin is the m inim al
step length of young m ussels (/nlin < I) [3,18,25,26]. The
step length distribution corresponds to a Lévy w alk for
1 < |x < 3 an d it approxim ates a Brownian w alk w hen
> 3 [27]. We apply a sim ple pow er-law m odel rather
th an a m ore com plex com posite m odel because w e are inter­
ested in the change of general statistical properties w ith
m ussel density rather th an in a detailed statistical description
of m ussel m ovem ent [18-20]. First, w e kept the m inim al step
length constant at the fixed value /min = 3 m m . G iven /min,
Proc. /?. Soc. B 281: 20132605
daily. Fresh, unfiltered seaw ater w as supplied to the container
at a rate of approxim ately 11 m in -1; a constant w ater tem pera­
ture of 16°C w as m aintained during the experiments. A t the
start of each experiment, m ussels w ere spread hom ogeneously
over an 80 x 60 cm red PVC sheet in a 120 x 80 x 30 cm con­
tainer. We used a red PVC sheet to provide a contrast-rich
surface for later analysis an d considered only the m ovem ents
of the m ussels w ithin this 80 x 60 cm arena. The container
w as illum inated using fluorescent lam ps. M ussel m ovem ent
w as recorded b y photographing the m ussels at 1 m in intervals
for a duration of 300 min; w e used a Logitech Q uickCam 9000
Pro w ebcam (w w w.logitech.com ), w hich w as positioned
about 60 cm above the w ater surface.
We derived the step lengths b y calculating the distance
betw een tw o reorientation events (e.g. w here a m ussel clearly
changes its direction of movement) using Turchin's angle
m ethod [18,24]. First, the observed m ovem ent path is discre­
tized into steps on basis of changes in the angle (a) of the
m ovem ent p ath at observed position i using the prior (i — 1)
an d the subsequent (i + 1) observed locations as follows:
rspb.royalsocietypublishing.org
(which are the hallm ark of Lévy m ovem ent) provide little
benefit u n d er these circum stances [11].
Physical theory offers an alternative, m ore parsim onious
explanation for the occurrence of Brownian m otion in
resource-rich environm ents. Einstein, followed b y Langevin,
theorized that Brownian m otion in solutes results from col­
lisions betw een particles [15,16]. Likewise, Brownian m otion
in ecology m ight result from frequent 'collisions' of anim als
w ith the resources they are searching for (food, shelter or
conspecifics), or w ith item s that they are trying to avoid (e.g. terri­
tory boundaries [17]). Untangling w hether the observed
m ovem ent patterns in searching animals reflect adaptation of
intrinsic m ovem ent strategies, or are the consequence of chan­
ging encounter (collision) rates w ith resources, is crucial both
for sound mechanistic understanding of Brownian m otion an d
for predicting anim al m ovem ent patterns in ecosystems w here
resource availability varies in space or time.
Here, w e provide evidence that, as in physics, Brownian
w alks in anim al m ovem ents can be caused by frequent encoun­
ters, rather than being the result of adaptation to high-density
conditions. In density-controlled experim ents w ith young
m ussels (Mytilus edulis), w e w ere able to distinguish betw een
intrinsic m ovem ent strategy an d the effects of resource density
b y separating the m ovem ent steps that w ere truncated by
encounters from those that w ere term inated spontaneously.
Recently, it w as show n that the individual m ovem ent of
young m ussels can be approxim ated b y a sim ple Lévy w alk
[18] (or a m ore complex multi-scale w alk, w hich provides an
even better fit [19,20]). The m ovem ent of individual m ussels
results in a self-organized m ussel bed w ith a regular labyrinth-like pattern w here local aggregation yields protection
against w ave stress an d predation w hile it reduces com petition
for algal food resources [21-23]. As the m ovem ent of individ­
ual m ussels can be experim entally studied in considerable
detail, this system provides a unique opportunity to investigate
how anim al m ovem ent patterns are affected by truncation of
m oves ow ing to encounters.
This pap er is structured as follows. First, w e describe m ove­
m ent of young m ussels observed in density-controlled
experiments, revealing that m ovem ent patterns are affected by
changes in the density of mussels. By distinguishing betw een
obstructed an d unobstructed m ovem ent steps, w e investigate
the relationship betw een intended an d realized m ovem ent p at­
terns. Second, w e create an individual-based m odel of self­
organized pattern form ation in m ussel beds to examine w hether
m ussel density could cause a change in the efficiency of Brow­
nian and Lévy walks, explaining a possible active shift in
m ussel m ovem ent strategy. Third, w e use a general argum ent
to dem onstrate that the interplay betw een any intrinsic m ove­
m ent strategy an d frequent ecological encounters will often
result in Brownian motion.
Downloaded from rspb.royalsocietypublishing.org on January 28, 2014
( a)
50 n
m ussel 2
5 .2kgnW
rspb.royalsocietypublishing.org
mussel 15
O k g rrr2
2 5 0 -,
40-
200-
3020-
50-
10 -
0-1
0J
20
30
40
50
U
15
(d)
(c)
&
Q
U
10
60
0 .5 0 -
0 .5 0 -
0 .2 0 -
0.2 0 -
0 .0 5 -
0.05 -
LW
— BW
0.01 J
0 . 01-1
0.05
0.20
1.00
5.00
20.00
step length (mm)
8
10
12
14
step length (mm)
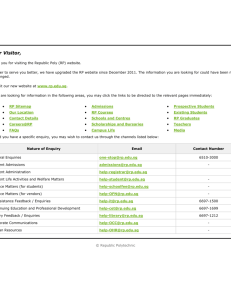
Figure 1. Step length distributions and m odel fits for m o vem en t trajectories at tw o m ussel den sities. Step len gth freguency distributions o f m ussel 15 in th e
0 kg m - 2 treatm ent (a) and m ussel 2 in th e 5.2 kg m - 2 treatm ent (6), tog eth er w ith an illustration o f th e m o vem en t paths (red dots indicate encounters
w ith conspecifics). The fitted lines to th e CCDFs o f th e step len gth s o f m ussel 15 (c) and m ussel 2 ( d ) indicate h o w w ell th e m o vem en t trajectories are represented
by a Lévy w alk (LW) and a Brownian w alk (BW).
the exponent p, can b e estim ated from the likelihood function
[25,26,28,29]
l (m
, . . . , z„ ) =
Um
i
(2.3)
w here {/,-... /„} are the observed step lengths. Taking the
natural logarithm of L an d m axim izing w ith respect to |x
yields the m axim um -likelihood estim ate
(i = 1 + U ■
ln(/;) - In (/min))
.
(2.4)
To check for the robustness of our results, w e also fitted the
observed step length distribution to a pow er law w here the
value of /min w as estim ated separately for each individual tra­
jectory (by equating /min w ith the m inim al observed step
length). O ur conclusions w ere n o t affected in any way.
By labelling steps as truncated w henever the step end ed
directly in front of another m ussel, w e w ere able to dis­
tinguish pure, non-truncated steps from those truncated by
collisions w ith conspecifics. For the sam e 10 individuals in
the five density treatm ents (50 m ussels in total), w e split
the steps into truncated a n d non-truncated steps, exam ining
the distributions separately.
(b) R esults
O ur mesocosm experiments illustrate that the observed move­
m ent patterns are strongly affected by mussel density (figures
1 an d 2; see the electronic supplem entary material, table SI).
Long steps occur less frequently w ith increasing mussel density
(figure 2a). The scaling exponent p, increases w ith mussel density
from a value below 2.5 at low densities to values above 3.5 at high
densities (figure 2b). As a second test of our hypothesis that
observed m ovement trajectories become more Brownian-like
w ith increased resource density, w e used the Akaike information
criterion for deciding w hether the individual trajectories in each
density class were better fitted by a pow er law or by an exponen­
tial distribution (corresponding to a Brownian walk). In 83% of
the m ovement trajectories in the lowest density treatment, a
Lévy w alk provided a better fit to the step length data than a
Brownian walk. By contrast, 75% of the tracks in the high-density
treatm ent were better approxim ated by a Brownian w alk than by
a Lévy walk. Again, w e conclude that m ovement trajectories
become more Brownian-like w ith increasing mussel density.
Closer exam ination of the m ovem ent data indicates that the
change of step length distribution w ith m ussel density results
from the frequent truncation of step lengths at high densities
(figure 2c,d). The fraction of truncated steps increases w ith
m ussel density (figure 2c), presum ably because the num ber
of encounters leading to an interruption of the m ovem ent
increases w ith density. W hen only considering non-truncated
steps, m ussel m ovem ent does n ot significantly differ b et­
w een density treatm ents (figure 2d). We conclude that the
intrinsic m ovem ent strategy of the m ussels does n o t change
w ith density an d that the observed change from Lévy-like to
Brownian-like m ovem ent results solely from the increased
m ussel encounter rates at high density.
Proc. R. Soc. B 281: 20132605
10
Downloaded from rspb.royalsocietypublishing.org on January 28, 2014
(b) 1.0
-,
rspb.royalsocietypublishing.org
È 0.8
0. 1 g
0.6 -1
G
G
Ü
0 .0 1 -
0.4 -
G
O k g rrr2
2.0 kg n r
5.2 kg n r
0.001 -I
1
*
10
0.2
-
0
-
100
30
0
step length (mm)
1.3
2.0
3.3
5.2
mussel density (kgm -2)
(c)
( d )
5.0
5.0
all steps
4.5 H
4 .5 -
4.0
4 .0 -
3.5
3.5
3.0
3 .0 -
2.5 H
2 .5 -
2.0
2 .0
1.5
T
0
1- - - - - - - 1- -----
r
1
2
3
4
5
mussel density (kgm -2 )
non-truncated steps only
-
1 .5 -
i-----------1----------1----------1----------1-------- 1
0
1
2
3
4
mussel density (kgm -2)
5
Figure 2. Effect o f m ussel d en sity on individual m o vem en t trajectories, (a) CCDF o f th e pooled step len gth s o f m oving m u ssels m easured for three d en sity treat­
m en ts. W ith increasing m ussel density, th e fraction o f lon g step s decreases. (6) Estim ated scaling exp on en t ¡a as a function o f m ussel density; ¡x increases w ith
m ussel den sity (linear regression, /3 n = 0.73, r = 0 .4 6 , d.f. = 46, p < 0.001; bars indicate average ¡x per den sity group ± s . e . ) and takes on valu es beyond 3 at
high den sities, (c) The fraction o f step s th at are truncated by collisions increases w ith m ussel d en sity (bars indicate m ean s + s.e.). (rf) W hen considering th e nontruncated steps only, th e scaling exp on en t ¡x rem ains approxim ately constan t (linear regression, /3 n = 0.18, r = 0, d.f. = 26, p = 0.593; bars indicate average ¡x
per d en sity group + s.e.).
3. A model of mussel movement
(a) M eth o d s
U sing a well-established m odel for m ussel m ovem ent [18],
w e investigated w hether an active sw itch from Lévy to Brow­
nian m ovem ent at high densities is m ore efficient th an the
persistent use of Lévy m ovem ent. We ran individual-based
com puter sim ulations for a range of values of the scaling
exponent p, an d at various densities, w here w e repeated
each sim ulation 10 tim es to account for stochasticity. W hen­
ever a displacem ent w as restricted b y the presence of a
conspecific, the step w as truncated. In each sim ulation, w e
determ ined the sum D of all displacem ents required before
the m ussels settled in a stable pattern. The inverse of D can
b e view ed as a m easure of the patterning efficiency of the
m ovem ent strategy u n d er consideration [18,30].
(b) R esults
Brow nian m ovem ent is often assum ed to be m ore efficient in
dense environm ents; som e researchers thus argue th at ani­
m als sw itch from Lévy to Brownian m ovem ent w h en
encountering areas of higher resource density. H ow ever,
sim ulations w ith ou r individual-based m odel [18] of m ussel
m ovem ent dem onstrate that Lévy m ovem ent is at least as
efficient as Brownian m otion at all densities. A t low densities,
a Lévy w alk w ith exponent p, ~ 2 is the m ost efficient
m ovem ent strategy (figure 3). A t higher densities, all m ove­
m ent strategies w ith 2 < p, < 3 lead to Brownian-like
m ovem ent patterns a n d therefore have a sim ilar patterning
efficiency; hence, the sim ulations do n o t su p p o rt the h y p o th ­
esis that Brow nian m ovem ent strategies lead to m ore efficient
aggregation th an Lévy m ovem ent strategies. This im plies that
there is no necessity to sw itch to a Brow nian strategy w ith
increasing density an d the m ussels in o u r experim ents do
n o t behave suboptim ally w h en using a Lévy w alk at high
densities (figure 2d).
4. A general argument
By m eans of a general argum ent, it can b e seen th at the tran ­
sition from non-Brow nian to Brow nian m otion at high
densities is a general phenom enon an d n ot restricted to
m ussel m ovem ent. C onsider a pop u latio n of anim als w here
the individuals have a certain intrinsic m ovem ent strategy,
for exam ple a Lévy w alk w ith a given exponent p,. If all indi­
v iduals could com plete their m ovem ent steps u ninterrupted,
this m ovem ent strategy w o u ld result in a step length dis­
tribution w ith a com plem entary cum ulative distribution
function CCDFintended^ ) (as in figure 2a, CCDF(/) corre­
sponds to the probability th at a step is longer th an or equal
to /). Suppose now that an anim al term inates its m ovem ent
w henever it encounters its desired target, such as food or
Proc. R. Soc. B 281; 20132605
3
.2
Downloaded from rspb.royalsocietypublishing.org on January 28, 2014
5. Discussion
77= 500
77= 750
77= 1000
77= 1250
„ = 1500
>
o>
c
»
o
0-1
1.5
2.0
2.5
3.0
A*
Figure 3. Patterning efficiency as a function o f th e scaling exp on en t /a in
m odel sim u lations for five different m ussel den sities. At low m ussel d en sity
{n = 50 0 ), a LW w ith /a « 2 has th e highest patterning efficiency, i.e. this
m o vem en t strategy creates a spatial pattern w ith a m inim um o f displace­
m en ts. At higher d en sities, a LW w ith ¡x « 2 still appears op tim al, but
m ost other m o vem en t strategies (including a BW) perform equally w ell.
Bars indicate m ean s o f 10 sim ulations + s.d.; lines illustrate cubic sm oothin g
splin es through th e m odel results. Patterning efficiency, m easured as th e
inverse o f th e distance D m oved per m ussel until a pattern w as form ed,
w as norm alized by dividing by th e largest efficiency found in all sim ulations.
shelter. (The sam e argum ents apply w hen m oves are term i­
nated ow ing to encounters w ith obstacles or the presence of
a potential danger, such as a predator or a rival.) If the
encounters of the m oving anim als w ith the target objects
are random , the probability that an intended step of length
I w ill not be term inated is given b y the zero term of a Poisson
distribution: e~kAl, w here A is the density of target objects
an d k is a constant of proportionality th at reflects aspects
such as the search w indow of the anim al or the size an d visi­
bility of the target objects. As a consequence, the CCDF of the
realized (and observed) step length distribution is given by
CCDFrealized(/) = CCDFintendedj/) ■e^M'.
(4.5)
As step lengths w ill becom e shorter ow ing to the term in­
ation of steps b y encounters, the realized step length
distribution w ill have a different signature than the intended
step length distribution. In particular, intended longer steps
w ill be term inated m ore often than intended shorter steps,
an d the probability that a step is term inated w ill d epen d on
the density of target objects. For large densities of the target
object, the exponential term becom es dom inant an d forces
the tail of the CCDF tow ards the exponential distribution
th at is characteristic of Brow nian w alks (figure 4). For
example, the CCDF of an intended Lévy w alk w ith exponent
/¿intended = 2 results in a realized CCDF that, ow ing to the ter­
m ination of steps b y encounters w ith the target object,
resem bles the CCDF of a Lévy w alk w ith a larger exponent
/¿realized (figure 4fl). In m ore general term s, an intended m ove­
m ent strategy that is n o t Brow nian at all takes on the
signature of Brow nian m otion w hen intended m ovem ent
steps are frequently term inated because of a high density of
target objects (figure 4b).
Proc. /?. Soc. B 281; 20132605
0 .2 -
Einstein dem onstrated that Brownian m otion of dissolved
particles can b e explained b y heat-driven collisions of these
particles w ith the molecules of the liquid [15,16]. Despite
obvious differences betw een m ovem ent in particles an d organ­
isms, o u r study show s that in analogy to physics, encounters
betw een organism s result in Brownian m otion, in particular
w h en found in encounter-rich environm ents. We observed
that u n d er controlled, experim ental conditions, m ussel m ove­
m ent patterns shifted from Lévy to Brownian m otion w ith
increasing m ussel density. By separating truncated from
non-truncated steps, w e w ere able to show that this change
in m ovem ent pattern is entirely the consequence of increased
encounter rate, as w e d id n o t observe a shift in intrinsic
m ovem ent strategy. We furtherm ore dem onstrated the u n i­
versality of this principle w ith a sim ple argum ent, show ing
that in general, encounters lead to Brownian m otion in
anim al m ovem ent patterns.
The shift from Lévy-like to Brownian m ovem ent w ith
increasing density has so far been explained as an adaptation
to increased resource availability. Anim als are considered to
ad a p t to increased encounters w ith food item s b y refraining
from large-scale m ovem ent steps, hence leading to adaptive
Brownian w alks [12,31]. H ow ever, o ur study provides a differ­
ent perspective on the observed shift from Lévy-like to
Brownian movement. W hen encounter rates are low, the
observed m ovem ent p attern reflects the intrinsic search strat­
egy, w hich can strongly deviate from Brownian movement.
W hen encounter rates are high, the signature of the intrinsic
search strategy is lost; large m ovem ent steps are frequently
truncated b y encounters an d the m ovem ent p attern resembles
Brownian m otion irrespective of the underlying intrinsic strat­
egy. This has im portant im plications for ecological theory, as
here Brownian m otion is n o t a default, intrinsic m ovem ent
m ode that underlies anim al dispersal b u t em erges from eco­
logical encounters betw een organism s, such as encounters
w ith food item s or interference w ith conspecifics, sim ilar to
the physical obstruction of m ussel m ovem ent observed in
o u r study.
The explanation of encounters driving Brownian m otion can
clarify observations from a num ber of terrestrial an d marine
studies. For instance, studies b y Bartum eus et al. [8], De Knegt
et al. [9] an d H um phries et al. [10-11] illustrate that microzooplankton, goats, m arine predators an d albatrosses all exhibit
Brownian m otion in areas w ith high food density an d Lévylike m ovem ent in resource-poor environm ents. These studies
highlight that an increased prevalence of Brownian m otion in
resource-rich environm ents is a general trend in ecological
systems. O ur explanation that encounters obscure innate m ove­
m ent strategy into an observed m ovem ent pattern that closely
resembles a Brownian w alk rationalizes this universal trend.
As a variety of ecological encounters, such as p red a to r-p rey
interactions, m ating or aggregation, are prone to occur in real
ecosystems, observed anim al m ovem ent patterns w ill always
deviate from the em ployed intrinsic m ovem ent strategy.
Especially in rich environm ents, resource encounters m ay
alter the m ovem ent p attern extensively. Hence, o u r study n ot
only illustrates the generality of this principle, b u t also high­
lights the im portance of ecological interactions in shaping
m ovem ent patterns of organism s throughout nature.
W hile density-dependence of dem ographic processes,
such as grow th an d predation, form s the cornerstone of
B
rspb.royalsocietypublishing.org
l.O -i*
.
«
*
0 -8 - .
Downloaded from rspb.royalsocietypublishing.org on January 28, 2014
(b)
5.0 n
4.5 Brownian walk
4 .0 •o 3 .5 -
s 10-2 "
&
Q
Ü 1 0 -3 Ü
_N
3.0 — 2.5 2.0
5
10
50
500
Levy walk
-
1.5
2.0
step length
2.5
3.0
3.5
4.0
¿'intended
Figure 4. Difference b etw een in tend ed and realized step len gth distribution for various den sities o f th e target object, (a) CCDFs o f th e realized step len gth s o f
organism s using a LW w ith scaling exp on en t /¿¡„tended = 2 as their intrinsic m o vem en t strategy. Only at zero d en sity, th e realized CCDF corresponds to th e in tend ed
CCDF, w h ile th e fatn ess o f th e tail o f th e distribution strongly decreases at higher den sities. The realized CCDF approxim ately correspond to th e CCDF o f a pow er law
w ith scaling exp on en t /¿realized = 2.5 , 2.9, 3.0 and 3.5 for th e increasing den sities, respectively. (6) Relationship b etw een intrinsic scaling exp on en t /¿¡„tended and
realized scaling exp on en t /¿reanzed for various ob ject den sities. M ovem ent patterns are often classified as a LW w h en th e estim a ted value o f /¿ is b etw een 1 and 3
and as BW w h en /¿ > 3.
ecological theory, anim al m ovem ent an d dispersal are ty p i­
cally approxim ated by density-independent linear diffusion,
b ased on the assum ption of Brow nian m otion. This study,
in com bination w ith previous w ork [7-11,18,23] show s that
for m any organism s, this assum ption is n ot valid; b o th m ove­
m ent rates an d m ovem ent characteristics m ay change as a
function of the local density of food item s or conspecifics,
being either through ecological encounters as advocated in
this paper, or through adaptation of m ovem ent [10]. As a
consequence, m ovem ent characteristics at the population
level m ay change w ith density, for instance from superdiffusive dispersal at low encounter rates, to m ore conservative
linear diffusion at h igh encounter rates. This can have im por­
tan t consequences for, for instance, the rate of spread of
infectious diseases an d invasive species or the form ation of
self-organized patterns. As the underlying m ovem ent strat­
egy w ill often be m asked u n d er high-density conditions
an d organism s thus m ight behave differently u n d er low-
density conditions, one m ust b e careful n o t to draw too farreaching conclusions from m ovem ent p attern s observed in
dense environm ents. A m ore m echanistic und erstan d in g of
ecological m ovem ent, facilitated by current im provem ents
in techniques to m onitor m oving anim als, w ill greatly
expand our ability to examine, m odel an d com prehend
anim al m ovem ent pattern s an d their influence on other
ecological processes.
A c k n o w le d g e m e n ts. W e th a n k V. Jan sen , F. M e y sm an , B. R. S illim an,
G. V isw a n a th a n a n d th e a n o n y m o u s referees for th e ir co m m en ts.
F or assista n ce, w e th a n k A . V a d e r, J. v a n S oelen a n d H . d e Paoli.
F.B. a c k n o w le d g e s th e S p a n ish m in istry of Science a n d In n o v a tio n
(REF. 2009-04133, B FU 2010-22337). T his is p u b lic a tio n 5527 of th e
N e th e rla n d s In stitu te of Ecology.
F u nd in g S ta te m e n t. M .D .J. is s u p p o r te d b y a g ra n t fro m th e N e th e rla n d s
O rg a n isa tio n for Scientific R esearch— E a rth a n d Life Sciences
(N W O -A LW ), a n d Project G ro u p M o v e m e n t E cology b y th e K N A W
S trategic F und.
References
1.
S k ellam JG. 1951 R an d o m d isp e rsa l in th e o re tic a l
6.
10 9 3 /b io m e t/3 8 .1 -2.1 96 )
2.
3.
N o let BA, M ooij W M . 2 0 0 2 S earch p a th s o f sw a n s
m a rin e p re d a to rs. Nature 465,1 0 6 6 - 1 0 6 9 . (do i:10 .
10 3 8 /n a tu re 0 9 1 16)
m o v e m e n t a s a c o rre la te d ra n d o m w alk. Oecologia
Eco!. 71, 4 5 1 - 4 6 2 . (d o i:1 0 .1 0 4 6 /j.1 3 6 5 -2 6 5 6 .2 0 0 2 .
56, 2 3 4 - 2 3 8 . (d o i:1 0 .1 0 0 7 /B F 0 03 79 69 5)
006 10 .x )
B e n h a m o u S. 2 0 0 7 H ow m a n y a n im a ls really d o th e
7.
H u m p h rie s NE et al. 2 0 1 0 E n v iro n m en tal c o n te x t
ex p lain s Levy a n d B row nian m o v e m e n t p a tte rn s o f
fo ra g in g on sp atially a u to c o rre la te d tu b e rs . J. Anim.
8.
Lévy flig h ts re co rd ed in situ. Proc. Natl Acad.
Sei. USA 109, 7 1 6 9 - 7 1 7 4 . (d o i:1 0 .1 0 7 3 /p n a s.
to reso urce availab ility in m icro zo o p lan k to n . Proc. Natl
Acad. Sei. USA 100, 12 7 7 1 - 1 2 7 75 . (d o i:1 0 .1 0 7 3 /
1121201109)
1 2.
p n a s.2 1 3 72 43 10 0)
9.
H u m p h rie s NE, W eim erskirch H, Q ueiroz N, S o u th all
EJ, Sim s DW. 2 0 1 2 F o raging success o f biological
B a rtu m e u s F, P e ters F, Pueyo S, M arrase C, C atalan J.
Sim s DW et al. 2 0 0 8 Scaling law s o f m a rin e
p re d a to r search b eh a v io u r. Nature 451,
1 1.
200 3 Helical Levy w alks: a d ju stin g search in g s ta tis tic
0 6 -1 7 6 9 .1 )
1 09 8 - 1 1 0 2 . (d o i:1 0 .1 0 3 8 /n a tu re 0 6 5 1 8)
5.
1 0.
K areiva PM , S h ig e sa d a N. 1 98 3 A nalyzing insect
Levy w alk? Ecology 88, 1 9 6 2 - 1 9 6 9 . (d o i:1 0 .1 8 9 0 /
4.
K lafter J, Sokolov IM. 2 00 5 A n o m alo u s diffusion
s p re a d s its w in g s. Phys. World 18, 29.
p o p u latio n s. Biometrika 38, 1 9 6 - 2 1 8 . (do i:10 .
Bell W J. 1991 Searching behaviour, the behavioural
ecology o f finding resources. L o nd on , UK: C h ap m an
De K negt HJ, H en gev eld GM, v an L a n g ev eld e F,
a n d Haii.
E d w ard s AM , F reem an MP, B reed GA, Jo n se n ID.
d e B oer WF, K irkm an KP. 2 0 0 7 Patch
1 3.
2 0 1 2 Incorrect lik elih oo d m e th o d s w e re u se d to
d e n sity d e te rm in e s m o v e m e n t p a tte rn s
PJ, S o uza TA, V isw a n a th a n GM . 2011 H ow
R aposo EP, B u rtu m eu s F, d a Luz MGE, R ibeiro-N eto
infer scalin g law s o f m a rin e p re d a to r search
a n d fo ra g in g efficiency o f larg e h erb iv ores.
la n d sc a p e h e te ro g e n e ity fra m e s o p tim a l diffusivity
b eh a v io u r. PLoS ONE 7, e 4 5 1 7 4 . (d o i:1 0 .1 3 7 1 /
Behav. Eco!. 80, 1 0 6 5 - 1 0 7 2 . (d o i:1 0 .1 0 9 3 /
in search in g p ro cesses. Comput. Biol. 7, e1 0 0 2 2 3 3 .
jo u rn a l.p o n e .0 0 4 5 1 7 4 )
b e h e c o /a rm 0 8 0 )
(d o i:1 0 .1 3 7 1 /jo u rn a l.p c b i.1 0 0 2 2 3 3 )
Proc. R. Soc. B 281: 20132605
1
rspb.royalsocietypublishing.org
0 kg m — 1.3 kg nr
2.0 kg nr
3.3 kg nr
5.2 kg nr
Downloaded from rspb.royalsocietypublishing.org on January 28, 2014
14.
ra te in biological in tera c tio n s: Lévy v ersu s B row nian
c o m m e n t o n 'Lévy w alks evo lv e th ro u g h
s tra te g ie s . Phys. Rev. Lett. 88, 0 9 7 9 0 1 . (d o i:1 0 .1 1 0 3 /
in tera c tio n b e tw e e n m o v e m e n t a n d e n v iro n m e n ta l
P h y sR evL ett.8 8.09 79 01 )
c o m p lex ity '. Science 335, 9 1 8 . (d o i:1 0 .1 1 2 6 /sc ie n c e .
Einstein A. 1 90 5 On th e m o v e m e n t o f sm all
1 21 59 03 )
21.
C lau set A, Shalizi CR, N ew m an ME. 2 0 0 9 P o w e r-la w
d istrib u tio n s in em p irical d a ta . SIAM Rev. 51,
6 6 1 - 7 0 3 . (d o i:1 0 .1 1 3 7 /0 7 0 7 1 0 1 1 1 )
2 7.
B a rtu m e u s F, d a Luz MGE, V isw a n a th a n GM, C atalan
th e m o le c u la r-k in e tic th e o ry o f h e a t. Ann. Phys. 17,
m u sse ls o n rocky sho res: in te g ra tin g p ro cess to
ra n d o m -w a lk analysis. Ecology 86, 3 0 7 8 - 3 0 8 7 .
5 4 9 - 5 6 0 . (d o i:1 0 .1 0 0 2 /a n d p .l9 0 5 3 2 2 0 8 0 6 )
u n d e rs ta n d p a tte rn . Ecology 82, 3 2 1 3 - 3 2 3 1 .
L an gev in P. 1 90 8 On the theory o f Brownian motion.
(do i:10 .1 8 9 0 /0 0 1 2 -9 6 5 8 (2 0 0 1 )0 8 2 [3 2 1 3 :P D O M O R ]2 .
G iuggioli L, P o tts JR, H arris S. 2 0 1 2 P redicting
22.
B ertran d S, B ertran d A, G uevara-C arrasco R, G erlotto
O.CO;2)
F. 2 0 0 7 S c a le -in v a ria n t m o v e m e n ts o f fish erm e n :
H u nt HL, S ch eiblin g RE. 2 0 0 2 M o v em e n t a n d w ave
th e s a m e fo rag in g stra te g y as n atu ra l p re d a to rs.
o scillatory d y n am ic s in t h e m o v e m e n t o f territo ria l
d is lo d g m e n t o f m u sse ls o n a w a v e -e x p o se d rocky
a n im a ls. J. R. Soc. Interface 9, 1 5 2 9 - 1 5 4 3 . (do i:10 .
sh o re . Veliger 45, 273 - 277 .
23.
(d o i:1 0 .1 8 9 0 /0 4 -1 8 0 6 )
2 8.
Eco!. Appl. 17, 3 3 1 - 3 3 7 . (d o i:1 0 .1 8 9 0 /0 6 -0 3 0 3 )
2 9.
E d w ard s AM. 2 0 0 8 U sing lik elih oo d to te s t fo r Lévy
Van d e K oppel J, G ascoign e JC, T h eraulaz G,
flig h t search p a tte rn s a n d fo r g e n e ra l p o w e r-la w
d istrib u tio n s in n a tu re . J. Anim. Eco!. 77, 1212 —
De J a g e r M, W eissing FJ, F lerm an PM J, N olet BA,
R ietkerk M, M ooij W M , F lerm an PM J. 2 0 0 8
v an d e n K oppel J. 2011 Lévy w a lk s evo lv e th ro u g h
E x p erim en tal e v id e n c e fo r sp a tia l self-o rg a n iz a tio n
in tera c tio n b e tw e e n m o v e m e n t a n d e n v iro n m e n ta l
a n d its e m e rg e n t effe c ts in m u ssel b ed eco sy stem s.
com p lex ity . Science 332,1 5 5 1 - 1 5 5 3 . (d o i:1 0 .1 1 2 6 /
Science 322, 7 3 9 - 7 4 2 . (d o i:1 0 .1 1 2 6 /sc ie n c e .
MGE, R aposo EP, S ta n ley FIE. 1 9 9 9 O p tim izing th e
scien ce.1 2 0 1 1 8 7 )
1 16 39 52 )
success o f ra n d o m s ea rc h e s. Nature 401, 9 1 1 - 9 1 4 .
J a n se n VAA, M a sh an o v a A, Petrovskii S. 2 01 2
24.
C o m m en t o n 'Lévy w alk s evo lv e th ro u g h in teractio n
b e tw e e n m o v e m e n t a n d e n v iro n m e n ta l c o m p lex ity '.
Science 335, 9 1 8 . (d o i:1 0 .1 1 2 6 /sc ie n c e .1 2 1 5 7 4 7 )
Turchin P. 1 99 8 Quantitative analysis o f movement.
S u n d e rla n d , MA: S in a u er A ssociates.
25.
1 22 2. (d o i:1 0 .1 1 1 1 /j .l 3 6 5 -2 6 5 6 .2 0 0 8 .0 1 428.x)
3 0.
V isw a n a th a n GM , B uldyrev SV, Havlin S, d a Luz
(d o i:1 0 .1 0 3 8 /4 4 8 3 1 )
3 1.
Frank SA. 2 0 0 9 T he c o m m o n p a tte rn s o f n atu re .
E d w ard s AM et al. 2 0 0 7 R evisiting Lévy flig h t search
J. Evol. Biol. 22, 1 5 6 3 - 1 5 8 5 . (d o i:1 0 .1 1 1 1 /j.1 4 2 0 -
p a tte rn s o f w a n d e rin g a lb a tro sse s, b u m b le b e e s a n d
9 1 0 1 .2 0 0 9 .0 1 7 7 5 .x )
Proc. /?. Soc. B 281: 20132605
19.
n atu re 0 6 1 9 9 )
2 6.
J. 2 00 5 A nim al search s tra te g ie s: a q u a n tita tiv e
1 0 9 8 /tsif.2 0 1 1.0 79 7)
18.
d e e r. Nature 449, 1 0 4 4 - 1 0 4 8 . (d o i:1 0 .1 0 3 8 /
H u nt HL, S ch eiblin g RE. 2001 Patch d y n am ic s o f
C. R. Acad. Sei. [Paris) 146, 5 3 0 - 5 3 3 .
17.
De J a g e r M, W eissing FJ, Flerm an PM J,
N o let BA, v an d e K oppel J. 2 0 1 2 R esp o n se to
p artic le s s u s p e n d e d in sta tio n a ry liq uids req u ired by
16.
20.
rspb.royalsocietypublishing.org
15.
B a rtu m e u s F, C atalan J, Fulco UL, Lyra ML,
V isw a n a th a n GM. 2 0 0 2 O p tim izing th e e n c o u n te r