WINTER COLD HARDINESS OF SEVEN
WILDLAND SHRUBS
R. H. Walser
D.J. Weber
E. D. McArthur
S. C. Sanderson
ABSTRACT
supercooling of water (ice formation avoidance) is involved
(George and others 1974). Death occurs when ice crystals
form within the cells and rupture cellular membranes in
this manner; ice, not low temperatures, causes death.
Plants can also deharden following warm spells during
the winter period. Howell and Weiser (1970) found that
short-term changes in cold resistance of apple stems were
closely related to the air temperatures of the preceding
day. Hardy plants dehardened during the winter as much
as 15 oc in 1 day in a warm greenhouse, and rehardened
15 oc in 3 days when they were held at -15 °C. The maximum cold hardiness achieved by 'Haralson' apple bark
was approximately -50 °C in midwinter and -7 °C in the
early fall and at bloom time in the spring. Thus, a cold
spell in early fall or spring or severe cold in midwinter
following a warm spell could cause considerable damage.
Various methods have been used in the laboratory to
determine shoot and bud cold hardiness. Stergios and
Howell (1973) evaluated several viability tests for coldstressed plants. They found that the tissue browning test
was the most reliable, but required considerable time and
was qualitative. In this test, 15-cm-long stem sections
were placed in a chamber where the temperature was
lowered 10 °Cih. Samples were removed at 5 oc intervals
and placed in a humid chamber at room temperature for
5 days to incubate, they then were examined for damage.
Undamaged bark remained green, while damaged bark
first became water soaked, then darkened, and ultimately
became soft. We used this technique to evaluate seven
wildland shrubs in relation to the temperatures that first
cause damage and temperatures that would most likely
kill the plant.
Winter kill of wildland shrubs can be caused by anumber of factors including freezing. Freezing damage and
lethal temperatures were determined and are as follows for
rubber rabbit brush (Chrysothamnus nauseosus) (-30 °C),
Stansbury cliffrose (Cowania stansburiana) (-55 °C), antelope bitterbrush (Purshia tridentata) (-50 °C), big sagebrush (Artemisia tridentata) (-80 °C), fourwing saltbush
(Atriplex canescens) (-80 °C), shadscale (Atriplex confertifolia) (-80 °C) and winterfat (Ceratoides lanata) (-80 °C).
In April as the buds began to swell, shoots could be killed
at temperatures around -5 °C for rubber rabbitbrush,
-15 °C for antelope bitterbrush and big sagebrush, -25 °C
for shadscale and Stansbury cliffrose, and -35
for
fourwing saltbush and winterfat.
oc
INTRODUCTION
Winter kill of perennial agriculture crops is well documented. On the other hand, wildland shrubs are assumed
to be very hardy and few studies have been done to determine their cold hardiness (VanEpps 1975; Jensen and
Urness 1979; Hanson and others 1982; Nelson and
Tiernan 1983). Wildland shrubs represent a major forage
source for livestock and wildlife during the harsh winter
period (McArthur 1988). Reports oflarge areas of die-off
have increased the interest in the possibility of winter
damage as a factor in shrub die-off. Temperate zone
woody plants have the ability to acclimate during the fall
season and thus withstand extremely cold winter temperatures. Research has shown that cold acclimation is a
2-stage sequence. Photoperiod is the initial stimulus that
triggers the various metabolic events leading to cold acclimation, and cold temperature triggers the second state of
the process (Howell and Weiser 1970). The mechanisms
utilized by plants to avoid freezing injury are not
co~pletely understood, but it is generally accepted that
MATERIALS AND METHODS
One-year-old shoots from rubber rabbitbrush,
Stansbury cliffrose, antelope bitterbrush, big sagebrush,
fourwing saltbush, shadscale, and winterfat were collected monthly from natural populations in or near Skull
Valley, UT, for two winters (1986 and 1987). The shoots
were cut into 15-cm-long sections and three sections from
each species were bundled together. Eight large bundles
were then formed with one bundle from each species and
treated as follows: seven bundles were placed in a freezing
chamber programmed to lower the temperature 10 °Cih.
The samples were removed from the chamber when the
predetermined temperatures were reached. The samples
were then placed under a mist system in a greenhouse
(20 °C) and evaluated for damage (browning of phloem,
Paper presented at the Symposium on Cheatgrass Invasion, Shrub DieOff and Other Aspects of Shrub Biology and Management, Las Vegas, NV,
April 5-7, 1989.
R. H. Walser is Associate Professor, Department of Agronomy and
Horticulture and D. J. Weber is Professor, Department of Botany and
Range Science, Brigham Young University, Provo, UT 84602; E. D.
McArthur is Project Leader and Chief Research Geneticist and S. C.
Sanderson is Research Geneticist, Intermountain Research Station, Forest
Service, U.S. Department of Agriculture, Shrub Sciences Laboratory,
Provo, UT 84606.
115
This file was created by scanning the printed publication.
Errors identified by the software have been corrected;
however, some errors may remain.
buds, or xylem) following 1, 4, and 10 days in the greenhouse. The eighth bundle (control) was placed directly in
the greenhouse and evaluated with the treated samples.
Shoots were rated for the temperature when the first
visible tissue damage was evident and when no visible
live tissue was evident and no bud growth occurred following 10 days in the greenhouse.
In January of each winter, a double amount of shoots
was harvested from the field. One set of shoots was processed as previously described, while the second set of
shoots was placed in the greenhouse (20 °C) for 6 days,
after which they were exposed to low temperatures as
previously described. The purpose of this experiment was
to determine the amount of dehardening each of the species was capable of achieving during midwinter.
RESULTS AND DISCUSSION
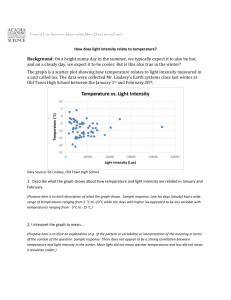
The results illustrate the great amount of cold tolerance
that is developed by the tested shrubs. In all cases, the
lowest temperature to kill the shrub shoots occurred in
the coldest part of the winter (January-February) as
shown in figures 1 through 4. However, there was a very
wide temperature range between the point where first
visible damage occurred and where no visible live tissue
was evident (for example, -27 oc and -80 oc in big sagebrush). Both temperatures were considerably higher in
December and March. Based on our experience with fruit
trees and ornamental shrubs, and our observations of the
progression of damage and regrowth of the treated shoots,
we suggest that the temperature which would cause
0
0
[]
•
Damaged
Killed
Rubber Rabbitbrush
Stansbury Cliffrose
-10
6
!!_..
-20
0
-20
!!_..
Q)
~
~-40
Cll
ia.
.a
e
e -30
Q)
E
Q)
Q)
1-
1-
-60
-40
••
--80
-50
Dec
Jan
Feb
Dec
Mar
Jan
Feb
Mar
Time (months)
Time (months)
0
Damaged
Killed
0
Big Sagebrush
Antelope Bitterbrush
-20
-20
E -40
0
!!_..
~
.a
Q)
~ -40
('(I
;
~a.
a.
-60
E
Q)
E
1-
Q)
1-
-60
-80
•
••
-100
Dec
Jan
Feb
Damaged
Killed
-80
Mar
Dec
Time (months)
Jan
Feb
Time (months)
Figure 1-Winter shoot damage (dark square)
and winter plant death (dark triangle) in rubber
rabbitbrush (top graph) and big sagebrush (bottom graph).
Figure 2-Winter shoot damage (dark square) and
winter plant death (dark triangle) in Stansbury
cliffrose (top graph) and antelope bitterbrush (bottom graph).
116
Mar
-20
-30
••
-20
Damaged
Killed
Damaged
Killed
Fourwing Saltbush
-30
Wlnterfat
-40
-40
0
0
t.. -50
t..
!:::J
~
:::J
; -50
;
i_
Q;
0..
-SO
E
~
E
~ -SO
-70
-70
•
-80
•
-80
Dec
Jan
Feb
-90
Dec
Mar
Feb
Jan
Mar
Time (months)
Time (months)
Figure 4-Winter shoot damage (dark square)
and winter plant death (dark triangle) in winterfat.
-20
Damaged
Killed
Shad scale
-30
Winter! at
-40
E
Shadscale
Cll
.aas
-50
Sattbush
Q;
0..
l!
E
~ -SO
1::
as
a:
Sagebrush
Cliff rose
-70
Bitterbrush
-80
Dec
Jan
Feb
Rabbitbrush
Mar
Time (months)
l?LI
•
-100
Six days warming
No warming
-80
-60
-40
-20
0
20
Temperature (°C)
Figure 3-Winter shoot damage (dark square) and
winter plant death (dark triangle) in fourwing saltbush
(top graph) and shadscale (bottom graph).
Figure 5-Effect of January warming on the cold
tolerance of winterfat, shadscale, fourwing
saltbush, big sagebrush, Stansbury cliffrose, antelope bitterbrush, and rubber rabbitbrush.
enough damage to the plants so that they would not survive the stresses of the following summer would fall close
to the midpoint between the two temperatures ranges.
That midpoint temperature would be approximately
-35 °C in December and -20 oc in March for rubber
rabbitbrush, -50 oc and -40 oc for big sagebrush, -47 oc
and -36 oc for Stansbury cliffrose, -45 oc and -35 oc for
antelope bitterbrush, -55 oc and -47 oc for fourwing
saltbush, -55 oc and -47 oc for shadscale, and -57 °C and
-45 oc for winterfat.
All of the plants tested showed considerable de hardening following the January warm treatment. The first
damage and final killing damage temperatures were reduced by 50 percent in almost all of the shrubs (fig. 5).
The warming period had the least effect on rubber rabbitbrush. Temperatures in the area where the shoots were
collected occasionally reach lows of -25 °C to -30 oc
(Stevens and others 1983). The coldest temperature ever
recorded in Utah was -45 oc (Stevens and others 1983).
Under these extreme temperatures, it appears that considerable damage could occur to the shrubs investigated,
particularly if the cold spell was preceded by a few days
of temperatures considerably above freezing. Another fact
that needs to be considered is that roots and crown area
of the plants are less cold tolerant than the shoots (Howell
and Weiser 1970). Research performed in Minnesota
117
(Wildung and others 1973) indicated that apple stems
could tolerate temperatures of -40 °C, while roots were
killed at -14 °C. Thus, a cold spell occurring without a
protective snow cover would very likely cause much more
damage than with a snow cover.
Jensen, C. H.; Urness, P. J. 1979. Winter cold damage to
bitterbrush related to spring sheep grazing. Journal of
Range Management. 35: 142-145.
McArthur, E. D. 1988. New plant development in range
management. In: Tueller, P. T., ed. Vegetation science
applications for rangeland analysis and management.
The Netherlands: Dordrecht: 81-112.
Nelson, D. L.; Tieman, C. F. 1983. Winter injury of sagebrush and other wildland shrubs in the western United
States. Res. Pap. INT-314. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain
Forest and Range Experiment Station. 17 p.
Stergios, B. C.; Howell, C. S., Jr. 1973. Evaluation of viability tests for cold stressed plants. Journal of American
Society of Horticultural Science. 98: 325-330.
Stevens, D. J.; Brough, R. C.; Griffin, R. D.; Richardson,
E. A. 1983. Utah weather guide. West Jordan, UT: Center for Applied Climateology. 46 p.
VanEpps, G. A. 1975. Winter injury to fourwing saltbush.
Journal of Range Management. 28: 157-159.
Wildung, D. K.; Weiser, C. J.; Pellett, H. M. 1973. Temperature and moisture effects on hardening of apple
roots. HortScience. 8: 53-55.
ACKNOWLEDGMENT
This research was supported in part by National Science Foundation Grant PCM-8320462 and was facilitated
by a cooperative agreement between Intermountain Research Station (USDA Forest Service) and Utah Division
of Wildlife Resources Project W-82-R.
REFERENCES
George, M. F.; Burke, M. J.; Weiser, C. J. 1974. Supercooling in overwintering azalea flower buds. Plant
Physiology. 54: 29-35.
Hanson, C. L.; Johnson, C. W.; Wight, J. R. 1982. Foliage
mortality of mountain big sagebrush (Artemisia tridentata ssp. vaseyana) in southwestern Idaho during the
winter of 1976-77. Journal of Range Management. 35:
142-145.
Howell, G. S.; Weiser, C. J. 1970. Fluctuations in the cold
resistance of apple twigs during spring de hardening.
Journal of American Society of Horticultural Sciences.
95: 190-192.
118
 0
0