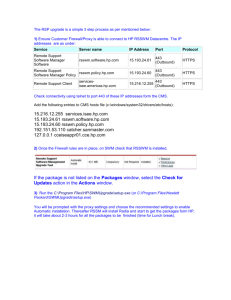
Does the human odometer use an extrinsic or intrinsic metric?
advertisement

Atten Percept Psychophys (2014) 76:230–246 DOI 10.3758/s13414-013-0549-3 Does the human odometer use an extrinsic or intrinsic metric? Elizabeth R. Chrastil & William H. Warren Published online: 3 October 2013 # Psychonomic Society, Inc. 2013 Abstract It is commonly assumed that path integration is based on an extrinsic measure of the objective distance traversed during locomotion. In contrast, biological odometers may rely on embodied intrinsic measures, such as idiothetic information specific to an action mode. We investigated this question using a distance reproduction task in which participants traveled an outbound distance and then reproduced that distance using the same or a different action mode. The extrinsic model predicted that distance reproduction should be invariant across action modes, whereas the intrinsic model predicted invariance only within an action mode. In Experiment 1, we held the outbound mode constant while varying the response mode (walk–walk, walk–throw) and corrected for response production error (view–walk, view–throw). In Experiment 2, we crossed different gaits in the outbound and response modes (walk, gallop). In both cases, we found that distance reproduction was significantly more accurate when the outbound and response modes matched, consistent with the intrinsic model. The results indicate that the human odometer preferentially relies on an intrinsic, rather than an extrinsic, metric. This solution is sufficient to support successful path integration within an action mode (but not across action modes), without the difficulties of objective distance estimation. Keywords Perception and action . Navigation . Locomotion Path integration, also known as dead reckoning, refers to the ability to keep track of one’s position and orientation in the environment by temporally integrating information about selfmotion. Accurate path integration implies that an animal could walk a circuitous outbound path from a home location and then take a direct path back to its starting point, a behavior known as homing. Animals such as desert ants are indeed capable of remarkably accurate and precise homing (Müller & Wehner, 1988; Sommer & Wehner, 2004; Wehner & Wehner, 1986), although homing performance in humans indicates that path integration is biased and more variable (Chance, Gaunet, Beall, & Loomis, 1998; Kearns, Warren, Duchon, & Tarr, 2002; Klatzky et al., 1990; Loomis et al., 1993; Peruch, May, & Wartenberg, 1997). Although error patterns in human path integration have been well studied, the underlying processes remain unclear. Fujita, Klatzky, Loomis, and Golledge (1993) identified three component processes in homing tasks: (1) encoding the distances and angles traversed on the outbound path, (2) integrating those distances and angles to compute a homebound trajectory or homing vector, and (3) executing that homebound path. The authors argued that error may be associated with each component. This analysis raised a number of questions that remain outstanding. First, what exactly is encoded on the outbound path; that is, what is the metric of path integration? Second, what is the source of the path integration errors observed in human homing, and can they be parsed into encoding, integration, and execution errors? Third, what is the nature of the integration process; for example, is the homing vector derived via a trigonometric computation or a heuristic process? Before the latter questions can be answered, the first needs to be addressed; in this article, we will focus on the problem of distance measurement, or odometry. The distance metric of the human odometer E. R. Chrastil : W. H. Warren Cognitive, Linguistic & Psychological Sciences, Brown University, Providence, RI, USA E. R. Chrastil (*) Psychology Department, Boston University, 2 Cummington Mall, Boston, MA 02215, USA e-mail: chrastil@bu.edu First, consider the metric of path integration. It is often assumed that path integration encodes the objective distances and angles that an animal traverses through the environment, using an extrinsic metric—that is, an absolute measure, defined in objectively scaled units. But there is no reason to believe a priori that biological systems evolved to extract Atten Percept Psychophys (2014) 76:230–246 environmental properties as measured by pre-19th-century geometers, such as linear distances in Euclidean space. Rather, biological systems may measure behaviorally relevant properties using an embodied intrinsic metric—that is, a relational measure in action-scaled units that are specific to the action involved (Gibson, 1979; Warren, 1984, 2007). As was first suggested by Bishop Berkeley (1709/2010), distance may be “measured by motion of the body” as a tangible locomotor interval. Subsequent research has shown that simple locomotor units such as the number of steps, step length, elapsed time, or expended energy are inadequate to characterize the odometer, and it is likely that the system depends on a higher-order complex of idiothetic variables (Durgin, Akagi, Gallistel, & Haiken, 2009; Mittelstaedt & Mittelstaedt, 2001; Schwartz, 1999; Turvey, et al., 2009; Wittlinger, Wehner, & Wolf, 2006, 2007). To investigate this question, we operationalized two conceptual models of distance encoding by the human odometer and tested them in a distance reproduction task. In a standard distance reproduction task, the participant walks a specified distance on an outbound path and is then asked to walk an equivalent distance on the response path. The extrinsic model proposes that objective distance (physical distance in the world) is encoded on the outbound path and then produced on the response path. This model predicts that distance reproduction should be invariant over changes in action mode, once execution error is controlled for. The intrinsic model proposes that an action-scaled measure based on idiothetic (motor commands, proprioceptive input, and vestibular input) information is registered on the outbound path and then reproduced on the response path. On this view, distance reproduction would be invariant only over actions that possess the same intrinsic metric, so that performance should be best when the outbound and response modes match. The intrinsic model still permits accurate distance reproduction and homing, provided that the same action mode is maintained during travel, without a strong assumption of objective distance measurement. We tested these predictions using a distance reproduction task in which the action mode changed between the outbound and response paths. Encoding and execution error Next, consider the three possible sources of path integration error identified by Fujita et al. (1993): encoding error, integration error, and execution error. The authors reasoned that execution error makes only a small contribution to the overall error, on the basis of the finding that blind walking to a target is quite accurate (Elliott, 1986, 1987; Loomis, da Silva, Fujita, & Fukusima, 1992; Ooi, Wu, & He, 2001; Philbeck & Loomis, 1997; Thomson, 1983). In the standard blind-walking task, also called visually 231 directed walking , a participant views a target, closes his or her eyes, and then walks to the target location. Since the target distance is given visually, the task eliminates locomotor encoding error, so the authors argued that blindwalking performance provides a measure of execution error. Furthermore, they assumed that integration error in computing the homebound path is negligible. Thus, Fujita et al. (1993) concluded that the primary source of error in path integration is encoding error, as summarized in their encoding-error model. Following this logic, researchers have estimated encoding error using distance and angle reproduction tasks (Jürgens, Boß, & Becker, 1999; Klatzky et al., 1990; Loomis et al., 1993; Marlinsky, 1999; Schwartz, 1999). Assuming that the traversed distance is encoded on the outbound path, and that little execution error occurs on the response path, then the overall reproduction error—the error in traveling a distance and then reproducing that distance—can be interpreted as a measure of encoding error. For example, an overshoot on the response path would indicate an overestimate of the distance during encoding, and vice versa. However, blind walking does not necessarily provide a valid estimate of execution error, and hence distance reproduction may not offer a valid estimate of encoding error. In blind walking, it is possible that both visual distance perception and locomotor execution are inaccurate, but compensatory. Suppose that the visual–motor system has established a calibration between visually perceived distance and locomotor displacement (Rieser, Pick, Ashmead, & Garing, 1995). Thanks to this calibration, blind-walking performance could be quite accurate, despite large perceptual and execution errors. For example, if distance were visually underestimated and locomotor execution produced overshooting, these errors would cancel out to yield accurate blind walking. Thus, performance in the standard blind-walking task cannot be taken as a measure of either perceptual accuracy or execution error. Recognizing this problem, researchers have developed other tasks, such as indirect walking or continuous pointing, in which the ability to walk to a perceived location cannot be explained by a simple calibration (Campos, Siegle, Mohler, Bülthoff, & Loomis, 2009; Loomis, Klatzky, Philbeck, & Golledge, 1998; Loomis & Philbeck, 2008; Philbeck, Loomis, & Beall, 1997). However, errors on indirect paths tend to differ from those on direct paths (Philbeck et al., 1997), and it remains possible that the standard blind-walking task is based on a visual–locomotor calibration. Thus, we question whether the execution error of walking has been firmly established. Moreover, the notion that blind walking provides an estimate of pure execution error presumes that the locomotor response is preplanned and ballistic. That is, it presumes that the response is executed without simultaneously encoding the traversed distance on the response path. But it seems highly 232 likely that the locomotor system is continuously registering idiothetic information for travel distance, and unlikely that the system would ignore such information on the response path when it could be used to confirm that the intended distance is actually being reproduced. Thus, performance in a blindwalking task cannot be taken as a measure of pure execution error, because it may also reflect encoding error during the response. We will refer to the combined source of encoding and execution error during a response as production error. Consequently, the standard distance reproduction task may not provide a valid estimate of encoding error. First, if execution error cannot be neglected, the reproduction error might confound encoding error on the outbound path with execution error on the response path. Second, if distance encoding occurs on both the outbound and response paths, reproduction would include both types of encoding error. In sum, it may not be possible to disentangle encoding error from execution error in a standard reproduction task. However, we can estimate relative encoding error by manipulating the action modes on the outbound and response paths. Extrinsic and intrinsic models This brings us back to the issue of the path integration metric. Models of path integration generally assume that the human odometer encodes objective distance during locomotion through the environment (e.g., Fujita et al., 1993; Mittelstaedt & Mittelstaedt, 2001). Alternatively, the odometer might register intrinsic action-scaled information during locomotion and reproduce it on the response path. Here we detail these alternatives and explain their predictions for distance reproduction. Although it is theoretically possible that both extrinsic and intrinsic models might have ballistic versions, we think it unlikely for the reasons described above, so we will focus on matching models in which information is monitored during both the outbound and response paths. Extrinsic models In an extrinsic model, the objective distance traveled on the outbound path is encoded and stored; during the response, objective distance is also encoded, and locomotion continues until this value matches the stored value (Fig. 1a). On this model, errors might occur due to (1) distance encoding on the outbound path, (2) decay of the stored distance value, (3) distance encoding on the response path, and (4) comparison of the stored distance value with the response value. The extrinsic model predicts that once the outbound distance is encoded, performance should not depend on the action mode used to produce the same distance during the response. Although distance is presumably not measured in feet or meters, the extrinsic model proposes that objective distance (the physical distance in the world) is mapped into Atten Percept Psychophys (2014) 76:230–246 some internal quantity that provides a common “yardstick” across different action modes. For example, in gaits with a double-support phase such as walking, if leg length is known and the angle between the two legs during double-support is given by proprioception, then step length can be determined, and the traversed distance could be registered by a “stride integrator,” independent of speed. A different mechanism might apply in single-support gaits such as skipping (e.g., a mechanism based on leg angle and impulse at kickoff), but it likewise maps objective distance into the same common metric, which could also be used to control other actions such as throwing. Thus, different action modes are calibrated to each other because they encode the same objective distance. Consequently, the extrinsic model predicts that the constant error in distance reproduction should be unaffected by the match between the outbound and response modes. Obviously, different types of responses may have different accuracies, so when testing the prediction we correct for this by measuring the production error of the response alone and subtracting it from the overall reproduction error. This computation yields an estimate of the encoding error in the outbound mode. The extrinsic model predicts that this estimated encoding error should be independent of the response mode, because objective distance is encoded and reproduced regardless of the particular action that carries it out. Mittelstaedt and Mittelstaedt (2001) proposed such a distance-matching model, in which an idiothetic transfer function encodes a representation of distance traversed on the outbound path, and a similar independent function encodes distance traversed on the response path, until the difference between them is brought to zero. Note that this model presupposes that the purpose of the transfer function is to convert idiothetic information into a measure of objective distance. If such a transfer function is linear with a constant gain, then distance reproduction will be invariant over a variety of outbound and response conditions. Contrary to this expectation, Mittelstaedt and Mittelstaedt reported systematic reproduction errors when walking speed was changed between the outbound and response path. Specifically, constant errors were lowest when the outbound and response speeds were the same, but significantly higher when the walking speeds differed. Nevertheless, they explained this effect within the extrinsic framework, proposing that the idiothetic transfer function is nonlinear (a leaky integrator), with parameters that depend on walking speed. In our view, to claim that different actions have different transfer functions—in essence, that each action mode possesses its own “yardstick” for measuring distance—implies that objective distance is not internally represented. The claim that distance is encoded differently in each action mode reduces to the claim that what is encoded is intrinsic (modespecific) information, not extrinsic distance. If distance reproduction is not invariant across action modes, this implies that Atten Percept Psychophys (2014) 76:230–246 233 Fig. 1 Comparison of the two models. a In the extrinsic model, as an observer walks forward, the objective distance is encoded as an internal quantity in common units; the observer then walks back until the same distance is produced. Note that the production error on the response path may include both execution error and encoding error during the response, but these two errors cannot be dissociated. b In the intrinsic model, an observer walks forward, registering some action-scaled idiothetic quantity, and then walks back until the idiothetic quantity is dissipated or matched there is no common metric for objective distance. We propose to take the distance-matching model one step farther, by suggesting that no representation of extrinsic distance may exist at all, but merely the matching of intrinsic idiothetic information. outbound path and idiothetic information associated with running on the response path, nothing would guarantee that the same objective distance would be traversed. Moreover, this quantity would not readily used to control other actions such as throwing. It is possible that different action modes become mutually calibrated through experience, such as repeated travel between two locations using different gaits, or throwing and walking to retrieve the projectile, but that this is contingent on specific experience. Thus, the intrinsic model predicts that the estimated encoding error should depend on the response mode, for reproduction error would be smaller when the outbound and response modes matched. Previous research has indicated that mismatches between the outbound and response modes tend to yield larger constant reproduction errors, as expected by the intrinsic model. Schwartz (1999) manipulated the step length and step frequency of walking on the outbound path and asked participants to return to the starting point at a normal walk, over a distance range of 10 to 50 m. When the step lengths of the outbound and response modes differed, the response range was compressed and absolute errors increased, relative to the normal walk–walk condition.1 Interestingly, Schwartz reported that the walking speed on the response path was correlated with the prescribed speed on the outbound path, even though participants were instructed to return at a normal walk. This observation suggests that they were attempting to match the outbound and response gaits, perhaps in order to match the idiothetic information. Intrinsic models In an intrinsic model, action-scaled information is registered on the outbound path as a measure of locomotor displacement, and action-scaled information is also registered on the response path until the outbound value is matched (Fig. 1b). For example, suppose that some idiothetic quantity is accumulated during the outbound path, depending on the action mode. During the response, another idiothetic quantity is accumulated by a similar process until the outbound value is matched (or a reciprocal process dissipates the accumulated value until it goes to zero). To the extent that the mappings between distance and idiothetic information were the same in the outbound and response modes, the objective distance would be reproduced. This model suggests that errors may occur in (1) the accumulation of idiothetic information on the outbound path, (2) decay of the stored idiothetic value, (3) the accumulation (or dissipation) process on the response path, and (4) the comparison of two accumulated values. The predictions of the intrinsic model contrast with those of the extrinsic model. Specifically, according to this model one would expect that constant errors in distance reproduction would be smaller when a participant used the same action mode on the outbound and response paths, once response accuracy was controlled for. If the navigator were to register idiothetic information associated with walking on the 1 Schwartz (1999) concluded that an effect of step frequency was mediated by its influence on step length. 234 Schwartz concluded that distance reproduction is based on a complex of proprioceptive variables that does not simply correspond to a stride integrator. Turvey et al. (2009) extended these findings to different gaits. In contrast to Schwartz, they found that differing speeds on the outbound path did not affect reproduction distance on the response path. They did find, however, that galloping or performing a hesitation step on the outbound path reduced accuracy when walking on the response path. Turvey et al. (2009; Turvey, Harrison, Frank, & Carello, 2012) proposed that different gaits fall into specific symmetry classes (Golubitsky, Stewart, Buono, & Collins, 1999; Pinto & Golubitsky, 2006), and only gaits within the same class allow accurate distance reproduction. Sun, Campos, Young, Chan, and Ellard (2004) crossed visual information and idiothetic information on the outbound and response phases, yielding four conditions: view–view (visual matching), view–walk (blind walking), walk–walk (standard distance reproduction), and walk–view. Conditions with matching outbound and response modes (view–view and walk–walk) tended to have the lowest constant errors, whereas mismatching conditions (view–walk and walk–view) yielded large errors, consistent with the intrinsic model. However, an exception occurred when they added visual information during walking (Sun et al., 2004). Walking with vision on both outbound and response paths (walk– walk with vision) led to significant errors. Walking with vision on the response path only (view–walk with vision) led to overshooting the target distance, whereas vision on the outbound path only (walk with vision–view) led to undershooting. These findings suggest that visual information such as optic flow may result in an underestimation of traversed distance and interfere with matching idiothetic information on the outbound and response paths (see also Bremmer & Lappe, 1999; Campos, Byrne, & Sun, 2010; Frenz, Bremmer, & Lappe, 2003; Harris, Jenkin, & Zikovitz, 2000; Lappe, Jenkin, & Harris, 2007; Mossio, Vidal, & Berthoz, 2008; Redlick, Jenkin, & Harris, 2001). The potential interaction of visual and idiothetic information in the human odometer needs further exploration. Finally, research on visual and vestibular information also implies that objective distance may not be encoded. Rather than rely on distance, participants may rely on the velocity profile of the outbound and return paths, matching them whenever possible (Berthoz, Israël, George-Francois, Grasso, & Tsuzuku, 1995; Bremmer & Lappe, 1999; Glasauer, Schneider, Grasso, & Ivanenko, 2007; Grasso, Glasauer, Georges-François, & Israël, 1999; Israel & Berthoz, 1989; Israel, Chapuis, Glasauer, Charade, & Berthoz, 1993; Israel, Grasso, Georges-François, Tsuzuku, & Berthoz, 1997). These findings are consistent with the intrinsic model, in that participants reproduce the spatiotemporal dynamics of the outbound path rather than using a distance metric. Atten Percept Psychophys (2014) 76:230–246 In sum, the previous literature suggests that matching the outbound and response modes facilitates distance reproduction, consistent with an intrinsic model of distance reproduction. These experiments have tested numerous mismatches in speed, step length, gait, and velocity profile. The findings suggest that the idiothetic information on the outbound and response paths must match fairly closely for accurate distance reproduction to be observed. In the present study, we tested very different action modes, such as throwing. Although the previous literature has shown that matched outbound and response modes tend to yield more accurate distance reproduction, they have not successfully estimated the potential sources of error. In particular, prior attempts to measure and directly compare encoding error have proven inconclusive. Thus, there has yet to be a strong test of alternative models of path integration. The aims of the present study were (1) to establish whether the human odometer encodes extrinsic distance or intrinsic actionscaled information and (2) to derive a useful estimate of encoding error. Logic of the experiments The two experiments presented here were designed to test whether the metric of the human odometer is extrinsic or intrinsic by manipulating the match between outbound and response modes in a distance reproduction task. In the first experiment, we held the outbound action mode constant while varying the response mode, whereas in the second we crossed the outbound mode and the response mode. Previous experiments have typically manipulated the outbound mode while keeping the response measure constant. Although this makes good sense, it also has a drawback: Encoding accuracy in the outbound mode is not controlled, so a difference in performance might be due to a difference in encoding error rather than to the match between the outbound and response modes. An alternative would be to hold the outbound mode constant and manipulate the response mode. The obvious drawback here is that response accuracy is not controlled, and may differ between response modes; thus, one must estimate and correct for production error during the response. In the present study, we pursued both of these approaches in order to obtain converging measures of performance with matching and mismatching action modes. In Experiment 1, we held the outbound mode constant and varies the response mode while controlling for differences in production. In Experiment 2, we fully crossed the outbound mode and the response mode. These two experiments would enable us to determine whether distance reproduction improves when the outbound and response modes match, as is predicted by the intrinsic model. Atten Percept Psychophys (2014) 76:230–246 Experiment 1 The extrinsic model predicts that objective distance is encoded and should transfer to other responses, whereas the intrinsic model predicts that performance should be more accurate when the outbound and response modes match. In Experiment 1, we held the outbound mode constant and varied the response mode, yielding two distance reproduction conditions: walk–walk and walk–throw. However, we also had to control for the possibility that people are worse at throwing a given distance than walking it. We thus measured the difference in production error between walking and throwing in two additional conditions—view–walk and view–throw—and used the result to correct the reproduced distances. The extrinsic model predicts that this correction should completely account for the difference between walk–walk and walk–throw: If the outbound distance is encoded in the same way, the only source of error would be the production error. In contrast, the intrinsic model predicts that reproduction errors in the walk–walk condition will still be smaller than those in the walk–throw condition, even after the correction: When the action modes match, the outbound idiothetic information can be matched during the response, but this information cannot be used to control throwing accurately, because it does not encode distance. The correction for walking and throwing accuracy amounts to subtracting production error from total reproduction error. This subtraction is equivalent to estimating the encoding error on the outbound path (see below). We performed two such estimates, one in which the outbound and response modes matched (walk–walk) and one in which they did not (walk– throw). The extrinsic model predicts that these two estimates of encoding error would be the same, because objective distance would be encoded and reproduced, whereas the intrinsic model predicts that they would differ, because no common distance metric would be present. In particular, the transfer of idiothetic information from walking to throwing might result in increased error as compared to a walking response, in which the idiothetic information could be matched. The extrinsic model thus predicts that the estimates of encoding error should be the same, regardless of the response mode, whereas the intrinsic model predicts that the estimated encoding errors would depend on the response mode. Experiment 1 provided a test of these predictions. Model predictions The computation of reproduction error, corrected for differences in production, makes the following assumptions: (1) Constant errors are additive; that is, errors from different sources, such as encoding or production error, can be summed linearly to yield total reproduction error. (2) Encoding error 235 when viewing a target is the same, regardless of the response mode. The extrinsic model predicts that distance reproduction should be accurate (after correction for production error) whether or not the action modes match, whereas the intrinsic model predicts more accurate distance reproduction when the outbound and response modes are the same than when they differ. In order to determine whether distance reproduction depends on the match between the outbound and response modes, in Experiment 1 we held the outbound mode constant and varied the response mode (walk–walk and walk–throw), while controlling for differences in response accuracy. Specifically, we corrected the overall reproduction error by subtracting a measure of production error (view–walk and view–throw), as follows. As is illustrated in Fig. 1a, the constant error in distance reproduction (REPRO) is the sum of the encoding error on the outbound path (ENCout) and the production error on the response path (PROD): REPRO ¼ ENC out þ PROD: ð1Þ Note that production error includes both execution error (EX) and encoding error during the response (PROD = EX + ENCresp).2 Although we cannot measure execution or encoding errors directly, we can measure reproduction and production errors. Once reproduction errors are corrected for any difference in production, they can be used to test the extrinsic and intrinsic models. In particular, these corrected reproduction errors yield estimates of the encoding error (ÊNCout), as can be seen by rearranging Eq. 1: b out ¼ REPRO−PROD: ENC ð2Þ We will compare two such estimates of encoding error, one in which the outbound and response modes match, and one in which they differ (refer to Fig. 2b). First, we measure overall reproduction error when the action modes match (walk–walk) and when they do not (walk–throw). Second, because throwing ability might simply be poorer than walking ability, we measure the production errors for both walking (view–walk) and throwing (view–throw). A difference between these conditions would be due to a difference between walking and throwing accuracy. Finally, we correct for any difference in production accuracy by computing two 2 Substituting for PROD, we see that REPRO = ENCout + (EX + ENCresp). Interestingly, if the outbound and response encoding processes are indeed the same, the encoding errors would cancel, such that ENCresp = −ENCout, and the reproduction error would simply reflect residual execution error, REPRO = EX. 236 Atten Percept Psychophys (2014) 76:230–246 Fig. 2 a View of the experimental setup and materials. b Conditions in Experiment 1, illustrating how reproduction errors (left) were corrected for production errors (center), yielding two estimates of encoding error (right) estimates of the outbound encoding error.3 The first subtracts the production error for walking from the overall reproduction error [(walk–walk) – (view–walk)]: b ww ¼ REPROww −PRODw : ENC ð3Þ The second estimate subtracts the production error for throwing from the reproduction error [(walk–throw) – (view–throw)]: b wt ¼ REPROwt −PRODt : ENC 3 ð4Þ Our purpose here was to compare these two estimates of encoding error; we are not claiming that the estimates are accurate. The computations assume that the encoding error when viewing the target is the same whether the response is walking or throwing (although they do not assume that the encoding error is zero). Thus, production errors after viewing a target are somewhat different from those in distance reproduction when walking: PRODv = ENCv + ENCresp + EX. Because we assume that the encoding error for viewing is the same regardless of the response mode, we can safely subtract production errors in order to compare the encoding errors. However, this estimate of encoding error may include a component ENCv that does not reflect the actual encoding error for walking. A difference between these two estimates (ÊNCww < ÊNCwt) would indicate that the overall reproduction error is smaller when the outbound and response modes match, even after correcting for differences in production error, consistent with the intrinsic model. The actual encoding error in walk–walk is presumably the same as in walk–throw. However, the models make different predictions regarding the two estimates of encoding error. The extrinsic model predicts that distance reproduction should be independent of the response mode; hence, the estimated encoding error should be as well. In contrast, the intrinsic model predicts that distance reproduction should be most accurate, and the estimated encoding error smaller, when the outbound and response modes match. Method Participants A group of 20 participants took part in Experiment 1 (ten male, ten female; age 24.05 ± 5.987 years, range 19–43); three additional participants failed to complete all Atten Percept Psychophys (2014) 76:230–246 experiment sessions and were not included in the analysis. They all had normal or corrected-to-normal vision. All participants gave informed consent to participate in the study in accordance with the Brown University Institutional Review Board. Stimuli and setup The experiment was composed of a total of four conditions, with 50 trials in each: walk–walk, walk– throw, view–walk, and view–throw. Trials were blocked by condition and presented within subjects during three experimental sessions, along with the conditions for Experiment 2b and one other condition not presented here. In the walking condition, strips of foam (approximately 4.5 cm wide and 1.5 cm high) were attached to the carpeted floor with Velcro and used to guide participants on a straight path (Fig. 2a). Participants held a cane made of plastic tubing (4-cm diameter) with a wiffle ball attached to the bottom to create a smooth foot. They stood approximately 36 cm to the left of the foam guide, grasped the cane with their right hand, and held it against the right side of the strip to guide their walking direction. To indicate where to stop when walking, another foam strip crossed over the first; when participants felt the cane touch this junction, it signaled them to stop. Audio cues also instructed participants when to start and stop walking. For the throwing condition, a beanbag (approximately 50 g) was covered with patches of Velcro to minimize sliding after it hit the carpeted floor and to keep it in its landing location. Two participants preferred to throw the beanbag with their left hand, and the rest used their right hand. In the view condition, the visual target was a vertical section of white PVC pipe (approximately 8.5 cm high and 10.5 cm in diameter), which was chosen to avoid a familiar size cue. Other distance information was preserved, including declination angle, motion parallax, binocular stereopsis, vergence, accommodation, and relative size across trials. For this condition, only a single foam strip in the orthogonal direction remained on the floor to guide responses, while the single “outbound” foam track in the target direction remained in place. The participant’s head position was tracked using an IS900 hybrid sonic/inertial tracking system (InterSense, Billerica, MA), hosted by a Dell XPS 730X computer. The experimenter’s head position was also tracked, as a means of recording the landing location of the beanbag. A wireless mouse was used to mark the beginning and end of each response in the position data file. Audio cues were presented through two loudspeakers driven by the computer’s sound system, and the participant wore headphones to reduce external noise. Procedure After giving informed consent, participants received practice walking with their eyes open and closed along the foam guides and practice throwing a beanbag to targets with their eyes open. The practice targets were colored rolls of 237 gaffer tape placed at different distances from those in the test phase, over a similar range; the experimenter called out a target color in a random order until each had been practiced five times. Participants were instructed to throw the beanbag while standing on two feet, and if they needed to throw farther, they should bend their knees more and arc the throw higher. Finally, participants were given two practice trials at the beginning of each experimental condition to familiarize themselves with the procedure, again at nontest distances but over a similar range. Distance encoding by counting steps or intervals was minimized by presenting a simple arithmetic problem during the “outbound” phase of each test trial, generated by randomly adding, subtracting, or multiplying the digits 2 through 9. In the walk conditions, an experimenter gave one arithmetic problem just after the participant started walking. In the view conditions, an experimenter gave one problem after the participant had viewed the target and put on a blindfold. In addition, participants were instructed that they might be tempted at times to count their steps, but they were asked to refrain from counting as the experimenters were not interested in how well they could count steps. Five outbound distances were used in each of the four conditions: 2, 4, 5, 6, and 8 m. Participants completed ten trials at each distance in a random order, yielding 50 trials per condition, for a total of 200 trials. Trials were blocked by condition, and the order of the blocks was randomized for each participant. Participants completed all four blocks as part of three 1.5-h sessions, two blocks per session, for a within-subjects design. At the end of each session, they filled out a brief questionnaire, in which they described any strategies they might have used, and rated the difficulty of each of the tasks on a 1–7 scale. Four experimental conditions were compared in this experiment: walk–walk, walk–throw, view–walk, and view–throw (Fig. 2b). These conditions were created by crossing two outbound modes (view and walk) with two response modes (walk and throw). Walk–walk This condition measured the reproduction error for walking (REPROww). Participants wore a blindfold during the entire block of trials. An audio cue (“Go”) directed participants to start walking. They walked forward while blindfolded until the cane touched one of the crossing foam strips and they heard a loud beep, indicating that they should stop walking and turn 90º to the left. After a 7-s pause, in which they turned and the cane was set on the next guide strip, another audio cue (“Go”) directed them to start walking again. Participants were instructed to walk the same distance along the new guide as they had walked on the first one. When they judged that they had travelled the same distance, participants stopped and clicked the mouse, at which point their position was recorded and a chime sounded. An experimenter guided the participant back to the starting location. 238 Walk–throw This condition measured the reproduction error when walking is reproduced by throwing (REPROwt). The procedure was identical to that of walk–walk except that after the 7-s pause, during which they turned 90º, participants were instructed to throw the beanbag the same distance they had walked on the first track. An experimenter recorded the position of the beanbag. A chime sounded when the position was recorded, and another experimenter walked the participant back to the start location. Again, participants kept the blindfold on during the entire block. View–walk This condition measured the production error for walking (PRODw). The procedure was identical to that of the walk–walk condition, except that the outbound distance was perceived by viewing a target. An audio cue (“Blindfold off”) directed participants to move the blindfold so that they could see the target. Participants were allowed to view the target for 5 s, and then an audio cue (“Blindfold on”) played. During the ensuing 7-s pause, participants were to replace the blindfold and turn 90º to the left. A final audio cue (“Go”) directed participants to walk an equivalent distance along a guide strip in that direction. Participants kept the blindfold on during the block except when viewing the target. View–throw This condition measured the production errors for throwing (PRODt). The procedure was identical to that of the view–walk condition, except that after viewing the target, participants were instructed to replace the blindfold, turn 90º to the left, and, after the 7-s pause, throw the beanbag an equivalent distance in that direction. Again, they kept the blindfold on during the block except when viewing the target. Occasional trials were excluded from the analysis when the tracker lost the participant’s position, the mouse did not record the data properly, the participant lost his or her place, or the beanbag was thrown into a wall or the ceiling. These errors occurred on fewer than 2 % of the trials. The analysis was performed on both the response distance (to derive constant error) and the within-subjects standard deviation of response distance (variable error) in each condition. Results and discussion Initial examination of the absolute (unsigned) errors offers an overall picture of accuracy in distance reproduction. The mean absolute errors were walk–walk (M = 0.611 m, SD = 0.206), walk–throw (M = 1.105 m, SD = 0.419), view–walk (M = 0.727 m, SD = 0.229), and view–throw (M = 0.801 m, SD = 0.427). A two-way repeated measures analysis of variance (ANOVA) on absolute error found a significant main effect of outbound distance [F (4, 76) = 16.144, p < .001], a significant effect of condition [F (3, 57) = 4.671, p < .01], and a significant Condition × Distance interaction [F(12, 228) = Atten Percept Psychophys (2014) 76:230–246 1.926, p < .05]. A post-hoc test comparing the walk–walk condition and the walk–throw condition found a significant difference between the mean absolute errors for these conditions (p < .01). These results suggest that conditions in which the outbound and response modes match (walk–walk) are more accurate than those in which they do not match (walk– throw). But the result of interest in Experiment 1 was the estimated encoding error. To obtain this value, we stepped through the analysis of reproduction error and production error, starting with the raw distance reproduction data (see Figs. 3 and 4); errors corresponded to overshooting or undershooting the diagonal in these figures. The corrected reproduction errors—equivalent to the estimated encoding errors—are represented in Fig. 5. First, we plotted the mean response distance as a function of outbound distance in the walk–walk and walk–throw conditions (Fig. 3a). Responses were more accurate (closer to the diagonal) when the action modes matched (walk–walk; REPROww) than when they differed (walk–throw; REPROwt), in which case participants tended to overshoot short distances and undershoot long distances. A planned two-way repeated measures ANOVA on response distance revealed a significant main effect of outbound distance [F(4, 76) = 462.956, p < .001] and a significant Distance × Condition interaction [F (4, 76) = 38.927, p < .001], confirming that reproduction errors were significantly smaller when the outbound and response modes matched. Similarly, an ANOVA on within-subjects standard deviations also yielded a main effect of distance [F (4, 76) = 28.997, p < .001) and a significant Distance × Condition interaction [F (4, 76) = 3.392, p < .05]. Thus, variable errors were also lower when the outbound and response modes matched. A linear regression on the mean response distance yielded a slope of 0.892 (intercept = 0.411, r = .999) for the walk–walk condition, as compared to 0.589 (intercept = 1.611, r = .997) for the walk–throw condition (Fig. 3a), indicating that the range of distance responses was more compressed in the walk–throw condition. The former value is in agreement with prior research, which has generally found quite accurate distance reproduction, with a slight tendency to overshoot small distances and undershoot large distances (Klatzky et al., 1990; Loomis et al., 1993; Marlinsky, 1999; Schwartz, 1999; Turvey et al., 2009). The latter condition has not been previously studied. These results suggest that distance reproduction depends on whether the outbound and response modes match. But obviously, walking and throwing ability might differ, so the next step was to estimate production accuracy by plotting the mean response distance as a function of target distance in the view–walk (PRODw) and view–throw (PRODt) conditions (Fig. 3b). Walking responses were quite accurate (close to the diagonal), whereas throwing responses were compressed. Atten Percept Psychophys (2014) 76:230–246 Fig. 3 Mean response distance as a function of outbound distance in Experiment 1. a Overall reproduction performance for walking on the outbound path and either a walking or throwing response. b Production accuracy for walking and throwing responses, after viewing the target distance. The solid gray diagonal lines indicate accurate performance, whereas overshooting and undershooting correspond to constant error; error bars represent between-subjects standard errors A planned two-way repeated measures ANOVA on mean response distance revealed a significant main effect of target distance [F (4, 76) = 498.578, p < .001] and a significant Distance × Condition interaction [F(4, 76) = 8.595, p < .001], confirming that throwing had a significantly lower production accuracy than walking. In contrast, a similar ANOVA on the within-subjects standard deviation showed only a main effect of distance [F(4, 76) = 34.653, p < .001], indicating that the precision of these responses is comparable. Linear regressions on the mean response distance yielded slopes of 0.976 (intercept = −0.292, r = .999) for the view– walk condition and 0.793 (intercept = 0.470, r = .999) for the view–throw condition (Fig. 3b), confirming distance compression for throwing. These results are in line with previous 239 Fig. 4 Mean response distance as a function of outbound distance (data replotted from Fig. 3). a Performance for the walking response, when either walking or viewing the outbound distance. b Performance for the throwing response, when either walking or viewing the outbound distance. The difference between the curves is the estimated outbound encoding error. Error bars represent between-subjects standard errors research showing a greater underestimation of large distances for blind throwing than blind walking (Eby & Loomis, 1987). In contrast, a study by Sahm, Creem-Regehr, Thompson, and Willemsen (2005) showed no Distance × Response interaction; however, they only tested distances up to 6 m, whereas we tested out to 8 m, resulting in a stronger interaction. Thus, we observed significantly greater production errors for throwing than for walking. The final step was to use this result to correct the overall reproduction error by subtracting the production errors for walking and throwing (refer to Fig. 2b). This correction yielded two estimates of the outbound encoding error in walking. First, we subtracted the production error for walking from the overall reproduction error [(walk–walk) – (view–walk)]; that is, we computed the difference between the two curves in Fig. 4a (REPROww – 240 Fig. 5 Estimated encoding error (corrected reproduction error) as a function of outbound distance. One estimate is based on the walking response (ÊNCww), and the other is based on the throwing response (ÊNCwt); each has been corrected for response production error. Error bars represent between-subjects standard errors PRODw = ÊNCww). The resulting estimate of encoding error, based on the walking response, is plotted in Fig. 5 (filled symbols). Second, we subtracted the production error for throwing from the overall reproduction error [(walk–throw) – (view–throw)]; that is, we computed the difference between the two curves in Fig. 4b (REPROwt – PRODt = ÊNCwt). This estimate of encoding error, based on the throwing response, is also plotted in Fig. 5 (open symbols). Figure 5 thus represents the two estimates of encoding error (i.e., corrected reproduction error) as a function of outbound distance. It is apparent that the estimate from the walk–walk condition (ÊNCww), when the outbound and response modes match, is flatter than the estimate from the walk–throw condition (ÊNCwt), when they differ. A planned two-way repeated measures ANOVA on the two estimates yielded a main effect of distance [F(4, 76) = 9.817, p < .001] and a significant Distance × Condition interaction [F(4, 76) = 2.969, p < .05]. This result indicates that when the outbound and response modes match, the error range is smaller, whereas when the modes differ, short distances are more overestimated and long distances are more underestimated. A similar ANOVA on the within-subjects standard deviations yielded no effects, indicating that variable error does not depend on matching action modes. This finding suggests that random errors in encoding processes on the outbound and response paths are uncorrelated. Linear regressions of the two estimates yielded slopes of −0.084 (intercept = 0.703, r = .916) for the walk–walk estimate and −0.204 (intercept = 1.141, r = .991) for the walk– throw estimate, indicating that encoding error based on walking was flatter than encoding error based on throwing (Fig. 5). As we explained in the introduction to this experiment, the extrinsic model predicts that no difference should emerge between these two estimates of encoding error, whereas the Atten Percept Psychophys (2014) 76:230–246 intrinsic model predicts that the estimate based on walking should be smaller than the estimate based on throwing (ÊNCww < ÊNCwt). Our results are consistent with this prediction, indicating that the overall reproduction error is smaller with matching action modes. These findings suggest that additional error, such as error due to different idiothetic information in different modalities, may be associated with a mismatch between the outbound and response modes. Self-reported difficulty ratings for the four conditions had means of 2.63 (SD = 0.93) for walk–walk, 5.90 (SD = 1.25) for walk–throw, 3.58 (SD = 1.14) for view–walk, and 5.23 (SD = 1.88) for view–throw. These ratings are in ordinal agreement with the observed reproduction errors. Tukey tests on the self-reported difficulty ratings showed that walk–walk was judged as being significantly easier than walk–throw (p < .001), but not easier than view–walk (p = .188). View–walk was rated as being significantly easier than view–throw (p < .01). Finally, participants did not rate view–throw as being significantly easier than walk– throw (p = .560). Thus, distance reproduction is judged to be easiest when the outbound and response modes match. The results of Experiment 1 indicate that distance reproduction is more accurate with matching outbound and response modes, consistent with the intrinsic model. Yet performance did not completely fall apart when the action mode changed in the walk–throw condition, as one might expect for mismatched idiothetic information. This observation suggests that participants may have established a rough calibration between the idiothetic information associated with walking and that associated with throwing, perhaps on the basis of previous experience walking to retrieve thrown objects. However, such a calibration exhibits systematic error, for throwing overshoots short walked intervals and undershoots long walked intervals. The results of Experiment 1 are in agreement with previous research. Schwartz (1999) found that when step length and walking speed differed on the outbound and response paths, distance reproduction was compressed. Similarly, Mittelstaedt and Mittelstaedt (2001) asked participants to reproduce distances of 2–21 m, using slow, standard, or fast walking speeds on the outbound and response paths. When the outbound and response speeds matched, distance reproduction was highly accurate. However, when the walking speeds did not match, the length of the response path depended on the difference between the outbound and response speeds. Together, these findings indicate that mismatches in step length, step frequency, or walking speed can reduce reproduction accuracy. Sun et al. (2004) took these finding a step further, by comparing visual estimates and walking estimates. They found that when outbound and response modes matched, distance estimations were more accurate than when they mismatched. In sum, Experiment 1 contrasted the extrinsic and intrinsic models of distance reproduction. The results suggest that Atten Percept Psychophys (2014) 76:230–246 distance reproduction is more accurate when the outbound and response modes match, even when taking into account differing accuracies in production. These results are in line with previous literature finding that differences in gait type and speed affect distance reproduction. Although these previous findings hinted at the intrinsic model, none of them corrected for production error. The present results provide direct evidence for the intrinsic model and against the extrinsic model. In addition, our procedure offers a practical estimate of encoding error that can be used in future work on path integration. 241 Turvey et al. (2012) over a shorter distance range (2–8 m, as opposed to 8–24 m). The extrinsic model predicts that participants should be able to encode objective distance whether walking or galloping, so distance reproduction should be equally accurate in all four conditions. In contrast, the intrinsic model predicts that participants should be less accurate in the gallop–walk and walk–gallop conditions, when the outbound and response modes differ, than in the walk–walk and gallop–gallop conditions, when the modes match. In Experiment 2a, we fully crossed walking and galloping, whereas in Experiment 2b we more closely examined the gallop–walk and walk–walk conditions. Experiment 2 Method Whereas in Experiment 1 we attempted to analyze distance reproduction into encoding error and production error, Experiment 2 directly tested whether distance reproduction depends on the match between the outbound and response modes. Specifically, we fully crossed the outbound and response modes, in this case a walking gait and a bipedal “galloping” gait, to ensure the proper control conditions. Walking and galloping are qualitatively more similar than the walking and throwing modes tested in Experiment 1. Both gaits generate idiothetic information for locomotor displacement that is produced by stepping with the lower limbs, with a double-support phase. However, in a gallop the participant leads every stride with one leg and follows with a catch-up step in the contralateral leg. Walking is a “primary” gait and galloping a “secondary” gait, as classified by symmetry group analysis of minimal neural pattern generators for bipedal and quadrupedal gaits (Golubitsky et al., 1999; Pinto & Golubitsky, 2006). Primary bipedal gaits are generated by similar signals to both legs (two-legged hop, two-legged jump) or by half-period phase shifts between them (walk, run), whereas secondary bipedal gaits (skip, gallop) require two types of signals. Turvey et al. (2009) hypothesized that distance measurement is invariant over gaits within a symmetry class, but not across classes. Turvey et al. (2009) found less accurate distance reproduction in a gallop–walk condition than in walk–walk and gallop–gallop conditions, consistent with the intrinsic model and the gait symmetry hypothesis. However, they did not fully cross the outbound and response modes with a walk–gallop condition, so it was not clear whether undershooting in the gallop–walk condition was due to a lack of transfer or reflected a calibration between galloping and walking. While the present article was in revision, Turvey et al. (2012) published a completely crossed experiment showing that participants in the walk–gallop condition reciprocally overshot relative to the gallop–gallop condition, suggesting a mutual calibration. Experiment 2 thus constitutes a replication of Participants A group of 19 participants took part in Experiment 2a (11 male, eight female; age 23.0 ±5.686 years, range 18–41); six additional participants failed to finish all experiment sessions and were not included in the analysis. The 20 participants in Experiment 1 also took part in Experiment 2b (ten male, ten female; age 24.05 ±5.987 years, range 19–43). They all had normal or corrected-to-normal vision and gave informed consent to participate in the study, in accordance with the Brown University Institutional Review Board. Procedure The experimental setup was the same as in Experiment 1. In addition, in Experiment 2a, audio cues were presented over a set of noise-canceling headphones, along with white noise to reduce external noise. In Experiment 2a, four conditions were presented: walk– walk, gallop–gallop, walk–gallop, and gallop–walk. Four outbound distances were used in each of the four conditions: 2, 4, 6, and 8 m. Participants completed eight trials at each distance in a random order, yielding 32 trials per condition, for a total of 128 trials. Trials were blocked by condition, and the order of the blocks was randomized for each participant. Participants completed all four blocks in two 1-h sessions, two blocks per session, for a within-subjects design. At the end of the second session, they filled out a brief questionnaire in which they described any strategies they might have used, and rated the difficulty of each of the tasks on a 1–7 scale. In Experiment 2b, two conditions were presented: walk– walk and gallop–walk. Five outbound distances were used in the two conditions: 2, 4, 5, 6, and 8 m. Participants completed ten trials at each distance in a random order, yielding 50 trials per condition, for a total of 100 trials. Trials were blocked by condition, and the order of the blocks was randomized for each participant. Participants completed both blocks together with Experiment 1, in three 1.5-h sessions, two blocks per session, in a within-subjects design. At the end of the last session, they filled out a brief questionnaire in which they 242 Atten Percept Psychophys (2014) 76:230–246 described any strategies that they might have used and rated the difficulty of each of the tasks on a 1–7 scale. The experimenter demonstrated the gallop step at the beginning of the session, instructing participants that they were to step forward with one foot and then bring the opposite foot in line with the first. Participants were allowed to lead with whichever foot they felt most comfortable and were allowed to pause between steps, as long as they were consistent throughout the block of trials. Thus, we did not distinguish gaits that Turvey et al. (2009) described as a walking gallop (with a pause in the double-support phase) and a running gallop (without a pause), although most participants used a running gallop. Otherwise, the procedure was the same as that in the walk–walk condition of Experiment 1. Occasional trials were excluded from the analysis when the tracker lost the participant’s position, the mouse did not record the data properly, or a participant lost his or her place. These errors occurred on fewer than 4 % of the trials. The analysis was performed on both the response distance (to derive constant error) and the within-subjects standard deviation of response distance (variable error) in each condition. Results and discussion The results of Experiment 2a are represented in Fig. 6, which plots mean response distance as a function of outbound distance for the four conditions. Distance reproduction appears to be more accurate (closer to the diagonal) when the outbound and response gaits match than when they differ. A planned two-way repeated measures ANOVA on response distance for the four conditions showed a main effect of outbound distance [F(3, 54) = 344.474, p < .001], a main effect of condition [F (3, 54) = 5.136, p < .01], and a significant interaction [F (9, 162) = 1.974, p < .05]. We also found an overall effect of outbound distance on the within-subjects standard deviation [F (3, 54) = 34.785, p < .001], but no effect of condition [F(3, 54) = 2.113, p = .109) or interaction [F (9, 162) = 0.329, p = .964]. To compare particular conditions, we performed planned comparisons using two-way repeated measures ANOVAs on response distance. The first comparison, between the walk– walk and gallop–gallop conditions, indicated that walking and galloping were equally accurate in distance reproduction (Fig. 6a): We observed a significant main effect of outbound distance [F(3, 54) = 728.169, p < .001], but no main effect of condition [F (1, 18) = 2.308, p = .146] and no interaction [F (3, 54) = 1.032, p = .386]. In addition, the within-subjects standard deviations for these conditions did not significantly differ [F (1, 18) = 0.005, p = .942]. In contrast, significant differences emerged between matching and mismatching gaits. A planned comparison of the walk–walk and walk–gallop conditions revealed a Fig. 6 Mean response distance as a function of outbound distance for the four conditions in Experiment 2a. The solid gray diagonal lines indicate accurate performance; error bars represent between-subjects standard errors. a Walk–walk and gallop–gallop. b Walk–gallop and gallop–walk significant effect of outbound distance [F(3, 54) = 497.390, p < .001] and a significant effect of condition [F(1, 18) = 13.587, p < .01], but no Distance × Condition interaction [F (3, 54) = 0.562, p = .642]. The planned comparison between the walk–walk and gallop–walk conditions revealed a main effect of distance [F (3, 54) = 396.282, p < .001], but no effect of condition [F (1, 18) = 1.024, p = .325] or interaction [F (3, 54) = 2.064, p = .116]. The planned comparison between the gallop–gallop and walk–gallop conditions yielded a main effect of distance [F(3, 54) = 682.563, p < .001], a main effect of condition [F (1, 18) = 4.633, p < .05], and no Distance × Condition interaction [F (3, 54) = 0.940, p = .428]. The comparison between the gallop–gallop and gallop–walk conditions yielded a main effect of distance [F (3, 54) = 564.724, p < .001], a marginal effect of condition [F (1, Atten Percept Psychophys (2014) 76:230–246 18) = 3.487, p = .078], and a significant interaction [F(3, 54) = 3.494, p < .05]. Taken together, these results indicate that performance with mismatched gaits differs from that with matched gaits, showing either a compressed response range or lower response distances. The final comparison, between the walk–gallop and gallop–walk conditions (Fig. 6b), yielded a main effect of distance [F (3, 54) = 500.018, p < .001], a main effect of condition [F(1, 18) = 7.416, p < .05], and a significant interaction [F (3, 54) = 3.605, p < .05]. This finding demonstrates that gallop–walk systematically undershot relative to walk–gallop. Linear regressions yielded slopes of 0.823 (intercept = 0.771, r = .999) for the walk–walk condition, 0.855 (intercept = 0.827, r = .998) for the gallop–gallop condition, 0.866 (intercept = 1.024, r = .998) for the walk–gallop condition, and 0.747 (intercept = 0.998, r = .999) for the gallop–walk condition. These results confirm that the gallop–walk condition exhibited the greatest response compression. The smallest response compression occurred in the walk–gallop condition, which also had the highest intercept, indicating a general overshoot in that condition. Considering the results in terms of the accuracy of distance reproduction, the mean absolute (unsigned) errors for the four conditions were walk–walk (M = 0.616 m, SD = 0.232), gallop–gallop (M = 0.638 m, SD = 0.105), gallop–walk (M = 0.854 m, SD = 0.334), and walk–gallop (M = 0.731 m, SD = 0.069). A two-way repeated measures ANOVA on absolute errors showed an effect of outbound distance [F (3, 54) = 4.550, p < .01], a marginal effect of condition [F(3, 54) = 2.552, p = .065], and a marginal Distance × Condition interaction [F(9, 162) = 1.905, p = .055]. Post-hoc comparisons between the conditions revealed that the walk–walk condition had significantly lower absolute errors than did the gallop– walk condition (p < .05). This finding suggests that conditions in which the outbound and response modes matched had lower errors than did conditions in which they did not match, supporting the intrinsic model. Finally, ratings of task difficulty showed an overall effect of condition [F(3, 54) = 30.150, p < .001]. Post-hoc Tukey tests revealed that the walk–walk condition (M = 2.45, SD = 1.07) was rated as being significantly easier than both the walk– gallop (M = 4.74, SD = 1.24, Tukey test, p < .001) and the gallop–walk (M = 4.84, SD = 1.21, Tukey test, p < .001) conditions, but was not different from the gallop–gallop task (M = 3.05, SD = 1.62, Tukey test, p = .482). Likewise, the gallop–gallop condition was rated as being easier than both the walk–gallop (Tukey test, p = .001) and the gallop–walk (Tukey test, p < .001) conditions, which were not different from each other (Tukey test, p = .994). Experiment 2b pursued the contrast between the gallop– walk and walk–walk conditions in more detail (Fig. 7). Distance reproduction in the walk–walk condition appeared to be more accurate (closer to the diagonal), whereas gallop–walk 243 Fig. 7 Mean response distance as a function of outbound distance for the two conditions in Experiment 2b. The diagonal line indicates accurate performance; error bars represent between-subjects standard errors exhibited more undershooting at larger distances. A two-way repeated measures ANOVA on response distance showed a main effect of outbound distance [F(4, 76) = 619.409, p < .001] and a significant Distance × Condition interaction [F (4, 76) = 6.041, p < .001], indicating greater response compression in the gallop–walk condition. An analysis of the withinsubjects standard deviations revealed only a main effect of distance [F (4, 76) = 35.554, p < .001], with no interaction. Thus, distance reproduction appears to be more accurate when the outbound and response modes match. Linear regressions on the mean response distance yielded slopes of 0.892 (intercept = 0.411, r = .999) for the walk–walk condition and 0.777 (intercept = 0.734, r = .997) for the gallop–walk condition (Fig. 7), confirming greater response compression in gallop–walk. The mean absolute error for walk–walk was 0.611 m (SD = 0.206), whereas for gallop– walk the mean absolute error was 0.891 m (SD = 0.313). A two-way repeated measures ANOVA on the absolute errors yielded a significant effect of outbound distance [F(4, 76) = 12.029, p < .001] and a significant effect of condition [F (1, 19) = 6.732, p < .05]. This result indicates that the matched condition was more accurate than the mismatched condition. Similarly, the gallop–walk condition (M = 4.20, SD = 1.40) was rated as being significantly more difficult than the walk– walk condition (M = 2.63, SD = 0.93, Tukey test, p < .01), such that distance reproduction was found to be easier with matching outbound and response modes. The results of Experiment 2 are consistent with the intrinsic model for distance reproduction, as performance was more accurate when the action mode was the same on the outbound and response paths. In contrast to Experiment 1, both the outbound and response modes generated proprioceptive information from the legs and vestibular information for selfmotion, so the mismatch between walking and galloping 244 was limited to the details of these gait patterns. Nevertheless, the differences in idiothetic variables were sufficient to induce significant differences in distance reproduction. The consistent pattern of overshooting in the walk–gallop condition and undershooting in the gallop–walk condition offers an important clue to the path integration process. First, it shows that these two gaits are systematically related, but are not accurately calibrated to each other. The extrinsic model holds that different gaits encode objective distance, and hence they should be accurately calibrated, yet they are not. A possible explanation is that distance reproduction is simply poor in the unfamiliar galloping gait. But Experiment 2a showed that gallop–gallop is as accurate as walk–walk, indicating that galloping reproduces distance quite well. Thus, the finding that the gaits were not wellcalibrated to each other implies that they do not encode objective distance. Second, the over- and undershooting suggests that walking and galloping are mutually calibrated, but inaccurately so, presumably due to limited prior experience. Specifically, the slope ratio of gallop–walk (0.747) to walk–gallop (0.866) yields a scaling constant of 0.863 between walking and galloping, such that walking undershoots a previously galloped distance, and galloping overshoots a previously walked distance. The findings of Experiment 2 can thus be explained by the intrinsic model: Matched idiothetic information on the outbound and response paths allows for accurate distance reproduction, whereas mismatched information yields systematic overand undershooting. Our results replicate those of Turvey et al. (2012) over a shorter distance range. They likewise found that distance reproduction was significantly less accurate in the gallop– walk and walk–gallop conditions than in the walk–walk and gallop–gallop conditions, and that the latter conditions did not differ from each other. Moreover, their gallop–walk condition also undershot the corresponding matched-gait condition, whereas the walk–gallop condition overshot it. This pattern of results is consistent with both the intrinsic model and the gait symmetry hypothesis, which will be revisited in the General Discussion. We interpret the results of Experiment 2 to be contrary to the extrinsic model but supportive of the intrinsic model. Distance reproduction depends on the match between the outbound and response modes; specifically, performance is more accurate when the outbound and response gaits are the same, relative to when they differ. General discussion The present results offer converging evidence of an intrinsic, action-scaled metric for the human odometer. These two experiments showed that distance reproduction is significantly Atten Percept Psychophys (2014) 76:230–246 more accurate when the outbound action mode matches the response action mode. In Experiment 1, we held the outbound mode constant (walk) and varied the response mode (walk or throw). We found that reproduction error was significantly larger in the walk–throw than the walk–walk condition, even after correcting for production accuracy (view–walk and view–throw conditions). Equivalently, we found that the estimated encoding error of outbound walking was significantly smaller in the walk–walk than in the walk–throw condition. Thus, overall distance reproduction was more accurate when the outbound and response modes matched. This finding is inconsistent with the extrinsic model, which assumes that objective distance is encoded on the outbound path and reproduced during the response. On the other hand, it is consistent with the intrinsic model, which holds that actionscaled information on the outbound path is matched on the response path, within an action mode. When the outbound and response modes differ, distance reproduction is less accurate, but does not fail completely. We suggest that different action modes may become approximately calibrated to each other if they are used to traverse the same locations. In Experiment 2, we crossed the outbound mode with the response mode (walk or gallop). Distance reproduction was again more accurate when the action modes matched than when they did not. This effect is unexpected on the view that objective distance is encoded and reproduced in both gaits, but it is consistent with the matching of action-scaled information that is specific to each gait. In particular, the finding that the gallop–walk condition consistently undershot the walk–gallop condition cannot be explained by the notion that each gait encodes objective distance. Rather, it appears that walking and galloping are mutually calibrated, but with a proportional scaling. Turvey et al. (2009; Turvey et al., 2012) proposed that the measurement of distance by locomotion is specific to a gait symmetry class. Bipedal gaits have two symmetry classes: primary gaits, which are characterized by reflectional symmetry, and secondary gaits, which are only characterized by rotational symmetry. Turvey et al. (2009) reported that reproduction accuracy was high even with unmatched outbound and response gaits, as long as they both came from the same symmetry class, including the primary gaits run–walk and backward walk–walk, and the secondary gaits running gallop–walking gallop. Nonetheless, Turvey et al. (2009) maintained that the human odometer measures objective traversed distance. We believe that these results support the intrinsic model. Turvey et al.’s (2009) conclusion that distance measurement is gait-symmetry-specific implies that each gait class possesses its own yardstick, consistent with the intrinsic hypothesis. However, gait symmetry classes offer only a first approximation of the relevant action modes. On the one hand, accurate distance reproduction across different gaits within a symmetry Atten Percept Psychophys (2014) 76:230–246 class may be attributable to similarities in idiothetic variables. For example, within the secondary gaits, it seems likely that idiothetic information in a backward walk is quite similar to that in a walk, and a running gallop is quite similar to a walking gallop (the former merely eliminates a pause during double-support). On the other hand, action modes appear to be more fine-grained than symmetry classes. Contrary to the symmetry hypothesis, a change in step length or speed between the outbound and response paths interferes with distance reproduction in walking (Mittelstaedt & Mittelstaedt, 2001; Schwartz, 1999), despite the fact that the gait class remains the same. Action modes would seem to have a basis in common idiothetic information, rather than in gait symmetry classes per se. We contend that action modes are individuated by the family of actions over which distance reproduction is invariant, and that each family shares a common complex of idiothetic variables related to locomotor displacement. Two cases, in which distance reproduction is accurate despite different outbound and response gaits, appear to present a challenge to the intrinsic hypothesis. Schwartz (1999) reported that distance reproduction was equal in jog–walk and walk– walk conditions, and Turvey et al. (2009) reported the same in run–walk and walk–walk conditions. Walking, jogging, and running are all primary gaits, and they seem to generate different patterns of idiothetic information. However, the finding that distance reproduction is invariant over this family of gaits leads us to expect that they could share a common higher-order complex of idiothetic variables. Hence, both the intrinsic model and the gait symmetry hypothesis imply that the metric for human odometry is an action-scaled quantity based on idiothetic information, not an extrinsic measure of objective distance. The notion that each action mode or symmetry class has its own “yardstick” for distance would seem to be problematic for path integration, for any shift in action mode during the course of travel would yield homing errors. On the other hand, an intrinsic metric avoids the difficulties of converting idiothetic variables to objective distances: As long as the navigator remains in one action mode during the journey, homing performance will be reasonably accurate and robust. For example, in a triangle completion task, idiothetic variables that underestimate objective distance would yield an “intrinsic path” that was larger than, but similar to, the triangle for the “extrinsic path,” and thus would successfully return to the home location. This similar-path argument also applies to more complex outbound routes. However, the argument does not hold for rotation: An intrinsic metric that misestimated objective turn angles would yield systematic homing errors. We will investigate this question in a subsequent study. In sum, we have found that distance reproduction does not depend on encoding objective distance, but on an actionscaled measure of locomotor displacement. The human 245 odometer thus appears to possess an intrinsic, rather than an extrinsic, metric: Distance is “measured by the body” using idiothetic information. This solution is sufficient to support successful path integration within an action mode, while avoiding the difficulties of measuring objective distance. Author note Preparation of this article was supported by National Science Foundation awards BCS-0214383 and BCS-0843940. The authors also thank Henry Harrison, Joost de Nijs, Mintao Zhao, Michael Fitzgerald, and the members of the VENLab for assistance with the research. References Berkeley, G. (2010). An essay towards a new theory of vision. Whitefish, MT: Kessinger. Original work published 1709. Berthoz, A., Israël, I., George-Francois, P., Grasso, R., & Tsuzuku, T. (1995). Spatial memory of body linear displacement: What is being stored? Science, 269, 95–98. Bremmer, F., & Lappe, M. (1999). The use of optical velocities for distance discrimination and reproduction during visually simulated self motion. Experimental Brain Research, 127, 33–42. Campos, J. L., Byrne, P., & Sun, H.-J. (2010). The brain weights bodybased cues higher than vision when estimating walked distances. European Journal of Neuroscience, 31, 1889–1898. doi:10.1111/j. 1460-9568.2010.07212.x Campos, J. L., Siegle, J. H., Mohler, B. J., Bülthoff, H. H., & Loomis, J. M. (2009). Imagined self-motion differs from perceived self-motion: Evidence from a novel continuous pointing method. PLoS ONE, 4, e7793. doi:10.1371/journal.pone.0007793 Chance, S. S., Gaunet, F., Beall, A. C., & Loomis, J. M. (1998). Locomotion mode affects the updating of objects encountered during travel: The contribution of vestibular and proprioceptive inputs to path integration. Presence: Teleoperators and Virtual Environments, 7, 168–178. Durgin, F. H., Akagi, M., Gallistel, C. R., & Haiken, W. (2009). The precision of locomotor odometry in humans. Experimental Brain Research, 193, 429–436. doi:10.1007/s00221-008-1640-1 Eby, D. W., & Loomis, J. M. (1987). A study of visually directed throwing in the presence of multiple distance cues. Perception & Psychophysics, 41, 308–312. doi:10.3758/BF03208231 Elliott, D. (1986). Continuous visual information may be important after all: A failure to replicate Thomson (1983). Journal of Experimental Psychology: Human Perception and Performance, 12, 388–391. doi:10.1037/0096-1523.12.3.388 Elliott, D. (1987). The influence of walking speed and prior practice on locomotor distance estimation. Journal of Motor Behavior, 19, 476–485. Frenz, H., Bremmer, F., & Lappe, M. (2003). Discrimination of travel distances from “situated” optic flow. Vision Research, 43, 2173– 2183. Fujita, N., Klatzky, R. L., Loomis, J. M., & Golledge, R. G. (1993). The encoding-error model of pathway completion without vision. Geographical Analysis, 25, 295–314. Gibson, J. J. (1979). The ecological approach to visual perception. Boston, MA: Houghton Mifflin. Glasauer, S., Schneider, E., Grasso, R., & Ivanenko, Y. P. (2007). Space– time relativity in self-motion reproduction. Journal of Neurophysiology, 97, 451–461. doi:10.1152/jn.01243.2005 Golubitsky, M., Stewart, I., Buono, P.-L., & Collins, J. J. (1999). Symmetry in locomotor central pattern generators and animal gaits. Nature, 401, 693–695. Grasso, R., Glasauer, S., Georges-François, P., & Israël, I. (1999). Replication of passive whole-body linear displacements from inertial 246 cues: Facts and mechanisms. Annals of the New York Academy of Sciences, 871, 345–366. Harris, L. R., Jenkin, M., & Zikovitz, D. C. (2000). Visual and non-visual cues in the perception of linear self motion. Experimental Brain Research, 135, 12–21. doi:10.1007/s002210000504 Israel, I., & Berthoz, A. (1989). Contribution of the otoliths to the calculation of linear displacement. Journal of Neurophysiology, 62, 247–263. Israel, I., Chapuis, N., Glasauer, S., Charade, O., & Berthoz, A. (1993). Estimation of passive horizontal linear whole-body displacement in humans. Journal of Neurophysiology, 70, 1270–1273. Israel, I., Grasso, R., Georges-François, P., Tsuzuku, T., & Berthoz, A. (1997). Spatial memory and path integration studied by self-driven passive linear displacement: I. Basic properties. Journal of Neurophysiology, 77, 3180–3192. Jürgens, R., Boß, T., & Becker, W. (1999). Estimation of self-turning in the dark: Comparison between active and passive rotation. Experimental Brain Research, 128, 491–504. doi:10.1007/s002210050872 Kearns, M. J., Warren, W. H., Duchon, A. P., & Tarr, M. J. (2002). Path integration from optic flow and body senses in a homing task. Perception, 31, 349–374. Klatzky, R. L., Loomis, J. M., Golledge, R. G., Cicinelli, J. G., Doherty, S., & Pellegrino, J. W. (1990). Acquisition of route and survey knowledge in the absence of vision. Journal of Motor Behavior, 22, 19–43. Lappe, M., Jenkin, M., & Harris, L. R. (2007). Travel distance estimation from visual motion by leaky path integration. Experimental Brain Research, 180, 35–48. doi:10.1007/s00221-006-0835-6 Loomis, J. M., da Silva, J. A., Fujita, N., & Fukusima, S. S. (1992). Visual space perception and visually directed action. Journal of Experimental Psychology: Human Perception and Performance, 18, 906– 921. doi:10.1037/0096-1523.18.4.906 Loomis, J. M., Klatzky, R. L., Golledge, R. G., Cicinelli, J. G., Pellegrino, J. W., & Fry, P. A. (1993). Nonvisual navigation by blind and sighted: Assessment of path integration ability. Journal of Experimental Psychology: General, 122, 73–91. doi:10.1037/0096-3445.122.1.73 Loomis, J. M., Klatzky, R. L., Philbeck, J. W., & Golledge, R. G. (1998). Assessing auditory distance perception using perceptually directed action. Perception & Psychophysics, 60, 966–980. doi:10.3758/ BF03211932 Loomis, J. M., & Philbeck, J. W. (2008). Measuring spatial perception with spatial updating and action. In R. L. Klatzky, B. MacWhinney, & M. Behrmann (Eds.), Embodiment, egospace, and action (pp. 1– 44). New York, NY: Taylor & Francis. Marlinsky, V. V. (1999). Vestibular and vestibulo-proprioceptive perception of motion in the horizontal plane in blindfolded man—I. Estimations of linear displacement. Neuroscience, 90, 389–394. Mittelstaedt, M.-L., & Mittelstaedt, H. (2001). Idiothetic navigation in humans: Estimation of path length. Experimental Brain Research, 139, 318–332. Mossio, M., Vidal, M., & Berthoz, A. (2008). Traveled distances: New insights into the role of optic flow. Vision Research, 48, 289–303. doi:10.1016/j.visres.2007.11.015 Müller, M., & Wehner, R. (1988). Path integration in desert ants, Cataglyphis fortis. Proceedings of the National Academy of Sciences, 85, 5287–5290. Ooi, T. L., Wu, B., & He, Z. J. (2001). Distance determined by the angular declination below the horizon. Nature, 414, 197–200. doi:10.1038/ 35102562 Atten Percept Psychophys (2014) 76:230–246 Peruch, P., May, M., & Wartenberg, F. (1997). Homing in virtual environments: Effects of field of view and path layout. Perception, 26, 301–311. Philbeck, J. W., & Loomis, J. M. (1997). Comparison of two indicators of perceived egocentric distance under full-cue and reduced-cue conditions. Journal of Experimental Psychology: Human Perception and Performance, 23, 72–85. doi:10.1037/0096-1523.23.1.72 Philbeck, J. W., Loomis, J. M., & Beall, A. C. (1997). Visually perceived location is an invariant in the control of action. Perception & Psychophysics, 59, 601–612. Pinto, C. M. A., & Golubitsky, M. (2006). Central pattern generators for bipedal locomotion. Journal of Mathematical Biology, 53, 474–489. Redlick, F. P., Jenkin, M., & Harris, L. R. (2001). Humans can use optic flow to estimate distance of travel. Vision Research, 41, 213–219. Rieser, J. J., Pick, H. L., Ashmead, D. H., & Garing, A. E. (1995). Calibration of human locomotion and models of perceptual–motor organization. Journal of Experimental Psychology: Human Perception and Performance, 21, 480–497. doi:10.1037/0096-1523.21.3.480 Sahm, C. S., Creem-Regehr, S. H., Thompson, W. B., & Willemsen, P. (2005). Throwing versus walking as indicators of distance perception in similar real and virtual environments. ACM Transactions on Applied Perceptions, 2, 35–45. Schwartz, M. (1999). Haptic perception of the distance walked when blindfolded. Journal of Experimental Psychology: Human Perception and Performance, 25, 852–865. doi:10.1037/0096-1523.25.3. 852 Sommer, S., & Wehner, R. (2004). The ant’s estimation of distance travelled: Experiments with desert ants, Cataglyphis fortis. Journal of Comparative Physiology A, 190, 1–6. Sun, H. J., Campos, J. L., Young, M., Chan, G. S. W., & Ellard, C. G. (2004). The contributions of static visual cues, nonvisual cues, and optic flow in distance estimation. Perception, 33, 49–65. Thomson, J. A. (1983). Is continuous visual monitoring necessary in visually guided locomotion? Journal of Experimental Psychology: Human Perception and Performance, 9, 427–443. doi:10.1037/ 0096-1523.9.3.427 Turvey, M. T., Harrison, S. J., Frank, T. D., & Carello, C. (2012). Human odometry verifies the symmetry perspective on bipedal gaits. Journal of Experimental Psychology: Human Perception and Performance, 38, 1014–1025. doi:10.1037/a0027853 Turvey, M. T., Romaniak-Gross, C., Isenhower, R. W., Arzamarski, R., Harrison, S., & Carello, C. (2009). Human odometer is gaitsymmetry specific. Proceedings of the Royal Society B, 276, 4309–4314. doi:10.1098/rspb.2009.1134 Warren, W. H. (1984). Perceiving affordances: Visual guidance of stair climbing. Journal of Experimental Psychology: Human Perception and Performance, 10, 683–703. doi:10.1037/ 0096-1523.10.5.683 Warren, W. H. (2007). Action-scaled information. In G. J. Pepping & M. L. Grealy (Eds.), Closing the gap: The scientific writings of David N. Lee (pp. 253–268). Mahwah, NJ: Erlbaum. Wehner, R., & Wehner, S. (1986). Path integration in desert ants: Approaching a long-standing puzzle in insect navigation. Monitore Zoologico Italiano, 20, 309–331. doi:10.1242/jeb.02657 Wittlinger, M., Wehner, R., & Wolf, H. (2006). The ant odometer: Stepping on stilts and stumps. Science, 312, 1965–1967. doi:10. 1126/science.1126912 Wittlinger, M., Wehner, R., & Wolf, H. (2007). The desert ant odometer: A stride integrator that accounts for stride length and walking speed. Journal of Experimental Biology, 210, 198–207.