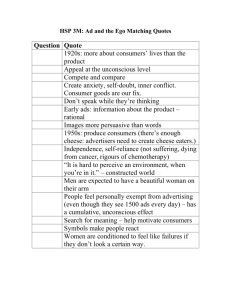
Document 11612149
advertisement

AN ABSTRACT OF THE THESIS OF Freddy Mauricio Lemus for the degree of Master of Science in Food Science and Technology presented on September 19, 2012 Title: Flavor Development of Cheddar Cheese Under Different Manufacturing Practices Abstract approved: Michael C. Qian Cheddar Cheese samples (good cheese, weak cheese, cheese made with pasteurized milk, cheese made with heat-shocked milk, cheese from production plant A, cheese from production plant B, cheese made with adjunct culture, and cheese made without adjunct culture), were evaluated during the ripening stage. Proteolysis was studied by a fractionation scheme, resulting in an insoluble fraction analyzed by urea polyacrylamide gel electrophoresis (UreaPAGE), and a soluble fraction which was further investigated through water soluble nitrogen (WSN), trichloroacetic acid soluble nitrogen (TCA-SN) and phosphotungstic acid soluble nitrogen (PTA-SN) analyzed by total Kjeldahl nitrogen content (TKN). Reversed phase high performance liquid chromatography (RP-HPLC) was used to study the peptide profile of the water soluble fraction. Lipolyisis was studied by levels of individual free fatty acids determined through gas chromatography-flame ionization detection (GC-FID) after isolation employing solid phase extraction (SPE). Volatile sulfur compounds were studied using head space solid phase micro-extraction (SPME) coupled with gas chromatography-pulsed flame photometric detection (PFPD). It was found that Urea-PAGE is capable to differentiate samples according their age, but cannot discriminate samples regarding the treatment assessed, quality or origin of the samples. However, measurements of total Kjeldahl Nitrogen (TKN) of the WSN, TCA-SN, and PTA- SN fractions, and the principal component analysis of the RP-HPLC peptide profile of the WSN fraction, revealed differences in the rate and pattern of proteolysis for each one of the manufacturing cases. Good cheese, cheese produce in plant TCCA, cheese made in plant CRP with adjunct culture isolated from plant TCCA cheese, and cheese made with heat-shocked milk developed higher level of total nitrogen for the WSN, TCA-SN and PTA-SN fractions, indicating that primary and secondary proteolysis were faster for these samples. This is supported by a PCA model with three principal components that account for the 80-83% of the variability of the data from the RP-HPLC peptide profile analysis, which discriminates the samples according to age and manufacturing practice. In addition, FFA profiles demonstrated higher levels of low and medium chain free fatty acids for good cheese, cheese produce in plant TCCA, cheese made in plant CRP with adjunct culture, and cheese made with heatshocked milk samples, which suggest faster lipolysis during ripening. The Volatile Sulfur Compounds (VSC) analysis showed higher levels of DMS and MeSH and lower levels of H2S, suggesting faster catabolism of sulfur containing amino acids in good cheese, cheese produce in plant TCCA, cheese made in plant CRP with adjunct culture, and cheese made with heatshocked milk. © Copyright by Freddy Mauricio Lemus September 19, 2012 All Rights Reserved Flavor Development of Cheddar Cheese Under Different Manufacturing Practices by Freddy Mauricio Lemus A THESIS submitted to Oregon State University in partial fulfillment of the requirements for the degree of Master of Science Presented September 19, 2012 Commencement June 2013 Master of Science thesis of Freddy Mauricio Lemus presented on September 19, 2012 APPROVED: Major Professor, representing Food Science and Technology Head of the Department of Food Science and Technology Dean of the Graduate School I understand that my thesis will become part of the permanent collection of Oregon State University libraries. My signature below authorizes the release of my thesis to any reader request Freddy Mauricio Lemus Munoz, Author ACKNOWLEDGEMENTS I sincerely thank Dr. Michael C. Qian, for his invaluable support, guidance, intellectual stimulation and especially for his patience and trust during the most challenging and difficult period of my life and my days at Oregon State University. I thank Dr. Robert Mc Gorrin, Head of the department of Food Science and Technology, Dr. Lisbeth Goddik, Dr. Thomas Shellhammer and Dr. Manoj Pastey, members of my Graduate Committee for providing an opportunity to pursue the M.S in Food Science and Technology at Oregon State University and for their continuous willing to help. My special thanks go to Cody Osborn and Family, Daniel Sharp and Family, Phillip Widstock, Helena Sanchez, Barb Kralj, Salaheddine Ziadeh, Nancy and Faith for their sincere and unconditional friendship that gave me happiness, keep me a float and was light during the darkest moments, and without whom this would not be possible. I would like to thank Dr. Yan Ping Qian, for her constant support, significant inputs, and absolute and unreserved collaboration during the various stages of this research project. I certainly would like to thank Linda Dunn and Linda Hoyser for their assistance to come here and for helping me to make a dream reality. I would like to thank the Office Staff for their wonderful work I appreciate the enthusiasm, strength and commitment of Hui Fang, Qin Zou and Grace, whose assistance and inputs were fundamental for my research work. Finally and most importantly, I thank my family... mi viejita la luna rockera, el Gordis, Sarah, Jan… and the best and only one … mi calvo, mi parce, mi padre y mi hermano, mi Heverth… for all of you there are no words enough!! CONTRIBUTION OF AUTHORS Dr. Michael C Qian, my advisor and pillar of this project, arranging the required funding, supervision and assistance at every stage of the project. Dr Will Austin provided some of the facilities of the Central analytical Laboratory for analysis of plant tissue, soil and water for nutrients at Oregon State University. Dr Yan Ping Qian, assisted the development of methodology for the Urea PAGE electrophoresis and TKN Kjeldahl analyses. Hui Fang assisted in the collection of some FFA analysis data. And Helen Burbank assisted in the collection of some VSC analysis data. TABLE OF CONTENTS Page CHAPTER 1----------------------------------------------------------------------------------------------------- 1 INTRODUCTION --------------------------------------------------------------------------------------------- 1 BACKGROUND ........................................................................................................................ 2 LITERATURE REVIEW ........................................................................................................... 3 CHEDDAR CHEESE ............................................................................................................. 3 FLAVOR FORMATION OF CHEESE ................................................................................. 3 Glycolysis ........................................................................................................................... 4 Lipolysis ............................................................................................................................. 4 Proteolysis .......................................................................................................................... 7 FACTORS AFFECTING THE DEVELOPMENT OF VOLATILE COMPOUNDS ......... 11 Milk preparation: selection, pasteurization & standardization ......................................... 12 Acidification ..................................................................................................................... 13 Coagulation....................................................................................................................... 13 Syneresis ........................................................................................................................... 14 Cheddaring........................................................................................................................ 15 Salting ............................................................................................................................... 15 Pressing and molding........................................................................................................ 16 CHEESE FLAVOR AND FLAVOR ANALYSIS ............................................................... 17 Instrumental analysis of cheddar cheese........................................................................... 19 Gas Chromatography .................................................................................................... 20 Extraction-Distillation .............................................................................................. 20 Headspace methods .................................................................................................. 21 Separation ................................................................................................................. 23 Reverse Phase-High Performance Liquid Chromatography (RP-HPLC) ..................... 24 Electrophoresis ............................................................................................................. 26 Aroma of Cheddar cheese................................................................................................. 28 REFERENCES ......................................................................................................................... 31 CHAPTER 2--------------------------------------------------------------------------------------------------- 44 TABLE OF CONTENTS (CONTINUED) EVALUATION OF THE EFFECT OF HEAT-SHOCK AND PASTEURIZATION TREATMENTS OF CHEESE MILK ON RIPENING RELATED CHANGES IN CHEDDAR CHEESE ------------------------------------------------------------------------------------------------------- 44 ABSTRACT ............................................................................................................................. 45 INTRODUCTION .................................................................................................................... 46 MATERIALS AND METHODS ............................................................................................. 48 CHEESE SAMPLES ............................................................................................................ 48 FREE FATTY ACIDS ANALYSIS ..................................................................................... 48 Chemicals ......................................................................................................................... 48 Extraction.......................................................................................................................... 49 Chromatography ............................................................................................................... 49 Quantitative analysis......................................................................................................... 49 VOLATILE SULFUR COMPOUNDS (VSC’S) ................................................................. 50 Chemicals ......................................................................................................................... 50 Extraction.......................................................................................................................... 50 Chromatography ............................................................................................................... 50 Quantitative analysis......................................................................................................... 51 Matrix effect ................................................................................................................. 51 Sulfur standards and internal standard preparation ....................................................... 51 PROTEOLYSIS .................................................................................................................... 52 Chemicals ......................................................................................................................... 52 Sample preparation and fractionation ............................................................................... 52 Macro blog digestion (Kjeldahl Digestion) ...................................................................... 53 Reversed phase High performance liquid chromatography analysis ................................ 53 Electrophoresis ................................................................................................................. 54 STATISTICAL ANALYSIS ................................................................................................ 54 RESULTS AND DICUSSION ................................................................................................. 54 FREE FATTY ACIDS FFA ................................................................................................. 54 Volatile Sulfur Compounds VSC’s....................................................................................... 58 Effect of treatment on Proteolysis ........................................................................................ 63 Soluble Nitrogen Fractions and TKN ............................................................................... 63 Peptide analysis by RP-HPLC .......................................................................................... 68 Electrophoresis ................................................................................................................. 75 CONCLUSION ........................................................................................................................ 77 REFERENCES ......................................................................................................................... 77 TABLE OF CONTENTS (CONTINUED) CHAPTER 3--------------------------------------------------------------------------------------------------- 90 COMPARISON OF RIPENING CHARACTERISTICS OF CHEDDAR CHEESE FROM THE SAME COMPANY MANUFACTURED IN DIFFERENT PRODUCTION PLANTS 90 ABSTRACT ............................................................................................................................. 91 INTRODUCTION .................................................................................................................... 92 MATERIALS AND METHODS ............................................................................................. 94 CHEESE SAMPLES ............................................................................................................ 94 FREE FATTY ACIDS ANALYSIS ..................................................................................... 94 Chemicals ......................................................................................................................... 94 Extraction.......................................................................................................................... 94 Chromatography ............................................................................................................... 95 Quantitative analysis......................................................................................................... 95 VOLATILE SULFUR COMPOUNDS (VSC’S) ................................................................. 95 Chemicals ......................................................................................................................... 95 Extraction.......................................................................................................................... 96 Chromatography ............................................................................................................... 96 Quantitative analysis......................................................................................................... 96 Matrix effect ................................................................................................................. 96 Sulfur standards and internal standard preparation ....................................................... 97 PROTEOLYSIS .................................................................................................................... 98 Chemicals ......................................................................................................................... 98 Sample preparation and fractionation ............................................................................... 98 Macro blog digestion (Kjeldahl Digestion) ...................................................................... 98 Reversed phase High performance liquid chromatography analysis ................................ 99 Electrophoresis ................................................................................................................. 99 STATISTICAL ANALYSIS .............................................................................................. 100 RESULTS AND DICUSSION ............................................................................................... 100 FREE FATTY ACIDS FFA ............................................................................................... 100 Volatile Sulfur Compounds VSC’s..................................................................................... 103 Effect of treatment on Proteolysis ...................................................................................... 108 Soluble Nitrogen Fractions and TKN ............................................................................. 108 Peptide analysis by RP-HPLC ........................................................................................ 112 Electrophoresis ............................................................................................................... 118 CONCLUSION ...................................................................................................................... 120 TABLE OF CONTENTS (CONTINUED) REFERENCES ....................................................................................................................... 121 CHAPTER 4-------------------------------------------------------------------------------------------------- 126 COMPARISION OF CHEDDAR CHEESE RIPENING MANUFACTURED WITH AND WITHOUT ADJUNCT CULTURE --------------------------------------------------------------------- 126 ABSTRACT ........................................................................................................................... 127 INTRODUCTION .................................................................................................................. 128 MATERIALS AND METHODS ........................................................................................... 130 CHEESE SAMPLES .......................................................................................................... 130 FREE FATTY ACIDS ANALYSIS ................................................................................... 130 Chemicals ....................................................................................................................... 130 Extraction........................................................................................................................ 130 Chromatography ............................................................................................................. 131 Quantitative analysis....................................................................................................... 131 VOLATILE SULFUR COMPOUNDS (VSC’S) ............................................................... 131 Chemicals ....................................................................................................................... 131 Extraction........................................................................................................................ 132 Chromatography ............................................................................................................. 132 Quantitative analysis....................................................................................................... 133 Matrix effect ............................................................................................................... 133 Sulfur standards and internal standard preparation ..................................................... 133 PROTEOLYSIS .................................................................................................................. 134 Chemicals ....................................................................................................................... 134 Sample preparation and fractionation ............................................................................. 134 Macro blog digestion (Kjeldahl Digestion) .................................................................... 134 Reversed phase High performance liquid chromatography analysis .............................. 135 Electrophoresis ............................................................................................................... 135 STATISTICAL ANALYSIS .............................................................................................. 136 RESULTS AND DICUSSION ............................................................................................... 136 FREE FATTY ACIDS FFA ............................................................................................... 136 Volatile Sulfur Compounds VSC’s..................................................................................... 139 Effect of treatment on Proteolysis ...................................................................................... 142 Soluble Nitrogen Fractions and TKN ............................................................................. 142 Peptide analysis by RP-HPLC ........................................................................................ 146 Electrophoresis ............................................................................................................... 152 TABLE OF CONTENTS (CONTINUED) CONCLUSION ...................................................................................................................... 154 REFERENCES ....................................................................................................................... 155 CHAPTER 5-------------------------------------------------------------------------------------------------- 159 COMPARATIVE CHEMICAL ANALYSIS OF GOOD AND WEAK CHEDDAR CHEESE DURING RIPENING -------------------------------------------------------------------------------------- 159 ABSTRACT ........................................................................................................................... 160 INTRODUCTION .................................................................................................................. 161 MATERIALS AND METHODS ........................................................................................... 162 CHEESE SAMPLES .......................................................................................................... 162 FREE FATTY ACIDS ANALYSIS ................................................................................... 162 Chemicals ....................................................................................................................... 162 Extraction........................................................................................................................ 162 Chromatography ............................................................................................................. 163 Quantitative analysis....................................................................................................... 163 VOLATILE SULFUR COMPOUNDS (VSC’S) ............................................................... 163 Chemicals ....................................................................................................................... 163 Extraction........................................................................................................................ 164 Chromatography ............................................................................................................. 164 Quantitative analysis....................................................................................................... 165 Matrix effect ............................................................................................................... 165 Sulfur standards and internal standard preparation ..................................................... 165 PROTEOLYSIS .................................................................................................................. 166 Chemicals ....................................................................................................................... 166 Sample preparation and fractionation ............................................................................. 166 Macro blog digestion (Kjeldahl Digestion) .................................................................... 167 Reversed phase High performance liquid chromatography analysis .............................. 167 Electrophoresis ............................................................................................................... 167 STATISTICAL ANALYSIS .............................................................................................. 168 RESULTS AND DICUSSION ............................................................................................... 168 FREE FATTY ACIDS FFA ............................................................................................... 168 Volatile Sulfur Compounds VSC’s..................................................................................... 172 Effect of treatment on Proteolysis ...................................................................................... 177 Soluble Nitrogen Fractions and TKN ............................................................................. 178 TABLE OF CONTENTS (CONTINUED) Peptide analysis by RP-HPLC ........................................................................................ 182 Electrophoresis ............................................................................................................... 188 CONCLUSION ...................................................................................................................... 189 REFERENCES ....................................................................................................................... 190 CONCLUSION ---------------------------------------------------------------------------------------------- 195 CONCLUSION ---------------------------------------------------------------------------------------------- 195 BIBLIOGRAFY --------------------------------------------------------------------------------------------- 197 APPENDICES ----------------------------------------------------------------------------------------------- 209 APENDIX A ........................................................................................................................... 209 ANOVAS ............................................................................................................................... 209 FFA ..................................................................................................................................... 209 Heat-shocked Vs Pasteurized ......................................................................................... 209 TCCA Vs CRP................................................................................................................ 211 Adjunct culture Vs Not ................................................................................................... 213 Good Vs Week................................................................................................................ 216 VSC’s.................................................................................................................................. 218 Heat-shocked Vs Pasteurized ......................................................................................... 218 TCCA Vs CRP................................................................................................................ 219 Adjunct culture Vs Not ................................................................................................... 219 Good Vs Weak................................................................................................................ 220 TKN .................................................................................................................................... 221 Heat-shocked Vs Pasteurized ......................................................................................... 221 TCCA Vs CRP................................................................................................................ 222 Adjunct culture Vs Not ................................................................................................... 222 Good Vs Weak................................................................................................................ 223 APENDIX B ........................................................................................................................... 225 PCA CORRELATION MATRIX .......................................................................................... 225 HEAT SHCOKED VS PASTEURIZATION ..................................................................... 225 TCCA Vs CRP.................................................................................................................... 225 ADJUCNT CULTURE Vs NOT ........................................................................................ 226 GOOD VS WEAK............................................................................................................. 227 LIST OF FIGURES Figure 1 FFA chromatogram Heat –schokecd Vs Pasteurized milk ......................................... 56 Figure 2 Development of individual FFA in Heat-shocked and Pasteurized milk cheese ....... 56 Figure 3 VSC's chromatogram Heat –schocked Vs Pasteurized milk ...................................... 62 Figure 4 Calibration curve DMS .............................................................................................. 62 Figure 5 Calibration Curve H2S ................................................................................................ 62 Figure 6 Development of MeSH in Heat-schocked and Pasteurized milk ............................... 62 Figure 7 Development of H2S in Heat-schocked and Pasteurized milk ................................... 62 Figure 8 Development of DMS in Heat-schocked and Pasteurized milk ................................. 63 Figure 9 TKN fractions heat-schocked cheese milk ................................................................. 67 Figure 10 TKN fractions pasteurized cheese milk.................................................................... 67 Figure 11 WSN heat-shocked and pasteurized cheese milk ..................................................... 67 Figure 12 TCA heat-schocked and pasteurized cheese milk .................................................... 68 Figure 13 PTA-SN heat-schocked and pasteurized cheese milk .............................................. 68 Figure 14 Scree plot of heat-shocked and pasteurized cheese milk.......................................... 73 Figure 15 Score plot of heat-shocked and pasteurized cheese milk (age) ................................ 73 Figure 16 Score plot of heat-shocked and pasteurized cheese milk (temperature) ................... 74 Figure 17 Loading plot for heat-shocked and pasteurized cheese milk .................................... 74 Figure 18 Peptide profile pasteurized cheese (A and B) Vs Heat-shocked (C and D) milk, for 2 and 12 months ........................................................................................................................... 75 Figure 19 Urea PAGE for ripening of cheese made from heat-shocked milk .......................... 76 Figure 20 Urea PAGE for heat-schocked Vs pasteurized cheeses (12 months) ....................... 76 Figure 21 FFA chromatogram TCCA Vs CRP....................................................................... 101 Figure 22 Development of individual FFA for TCCA and CRP cheese ................................ 102 Figure 23 VSC's chromatogram for TCCA cheese ................................................................. 107 LIST OF FIGURES (CONTINUED) Figure 24 Development of H2S for TCCA Vs CRP cheese .................................................... 107 Figure 25Development of MeSH for TCCA Vs CRP cheese ................................................. 107 Figure 26 Development of DMS for TCCA Vs CRP cheese ................................................. 108 Figure 27 TKN fractions TCCA cheese ................................................................................. 110 Figure 28 TKN fractions CRP cheese..................................................................................... 111 Figure 29 WSN TCCA Vs CRP cheese .................................................................................. 111 Figure 30 TCA-SN TCCA Vs CRP cheese ............................................................................ 111 Figure 31 PTA-SN TCCA Vs CRP cheese ............................................................................. 112 Figure 32 Scree plot of TCCA Vs CRP cheese ...................................................................... 116 Figure 33 Score plot of TCCA Vs CRP cheese (age) ............................................................. 116 Figure 34 Score plot of TCCA Vs CRP cheese (Origin) ........................................................ 117 Figure 35 Loading plot of TCCA Vs CRP cheese .................................................................. 117 Figure 36 Peptide profile CRP cheese (A and B) Vs TCCA (C and D) cheese, for 2 and 14 months .................................................................................................................................... 118 Figure 37 Urea PAGE for ripening of TCCA cheese ............................................................. 119 Figure 38 Urea PAGE for TCCA Vs CRP cheeses (12 m)..................................................... 120 Figure 39 Urea PAGE for TCCA Vs CRP cheeses (2 months) .............................................. 137 Figure 40 Development of individual FFA for cheese with and without adjunct culture....... 137 Figure 41 VSC chromatogram for Cheese with adjunct culture ............................................. 141 Figure 42 Development of H2S in Cheese made with and without adjunct culture................ 141 Figure 43 Development of MeSH in Cheese made with and without adjunct culture............ 141 Figure 44 Development of DMS in Cheese made with and without adjunct culture ............. 142 Figure 45 TKN fractions cheese with adjunct culture ........................................................... 144 Figure 46 TKN fractions cheese without adjunct culture ...................................................... 145 LIST OF FIGURES (CONTINUED) Figure 47 WSN Cheese with Adj culture Vs Cheese without ................................................ 145 Figure 48 TCA-SN Cheese with Adj culture Vs Cheese without ........................................... 145 Figure 49 PTA-SN Cheese with Adj culture Vs Cheese without ........................................... 146 Figure 50 Scree plot of cheese with and without adjunct culture ........................................... 150 Figure 51 Score plot of cheese with and without adjunct culture (age).................................. 150 Figure 52 Score plot of cheese with and without adjunct culture (culture) ............................ 151 Figure 53 Loading plot of cheese with and without adjunct culture....................................... 151 Figure 54 Peptide profile cheese with adjunct culture (A and B) Vs Not (C and D) cheese, for 2 and 10 months ...................................................................................................................... 152 Figure 55 Urea PAGE for ripening of cheese made with and without adjunct culture .......... 153 Figure 56 Urea PAGE for ripening of cheese made with and without adjunct culture .......... 154 Figure 57 FFA chromatogram week cheese ........................................................................... 169 Figure 58 Development of individual FFA in good cheese vs weak cheese .......................... 170 Figure 59 VSC's chromatogram Good Cheese ....................................................................... 175 Figure 60 Calibration Curve H2S ............................................................................................ 176 Figure 61 Calibration Curve DMS ......................................................................................... 176 Figure 62 Development of MeSH in Good and weak cheese ................................................. 176 Figure 63 Development of H2S in Good and weak cheese .................................................... 177 Figure 64 Development of DMS in Good and weak cheese.................................................. 177 Figure 65 TKN fractions Good cheese ................................................................................... 180 Figure 66 TKN fractions Good cheese ................................................................................... 180 Figure 67 WSN Good Vs Weak cheese .................................................................................. 181 Figure 68 WSN Good Vs Weak cheese .................................................................................. 181 Figure 69 PTA-SN Good Vs Weak cheese............................................................................. 182 LIST OF FIGURES (CONTINUED) Figure 70 Scree plot of Good Vs Weak cheese ...................................................................... 185 Figure 71 Score plot of Good Vs Weak cheese (age) ............................................................ 186 Figure 72 Score plot of Good Vs Weak cheese (quality) ....................................................... 186 Figure 73 Peptide profile for heat-shocked and pasteurized cheese milk ............................... 186 Figure 74 Peptide profile Weak Cheese (A and B) Vs Good Cheese (C and D) .................... 187 Figure 75 Urea PAGE for ripening of Good cheese ............................................................... 188 Figure 76 77 Urea PAGE for Good Vs Weak cheese (12 months) ........................................ 189 LIST OF TABLES Table 1 Aroma compounds of Cheddar cheese ........................................................................ 28 Table 2 Anova FFA of Heat-shocked Vs Pasteurized ............................................................ 209 Table 3 Anova FFA of TCCA Vs CRP ................................................................................. 211 Table 4 Anova FFA of Adjunct culture Vs Not...................................................................... 213 Table 5 Anova FFA of Good Vs Weak .................................................................................. 216 Table 6 Anova VSC of Heat-Shocked Vs Pasteurized ........................................................... 218 Table 7 Anova VSC TCCA Vs CRP ...................................................................................... 219 Table 8 Anova of VSC Adjunct Vs Not ................................................................................. 219 Table 9 Anova of VSC Good Vs Weak .................................................................................. 220 Table 10 Anova of TKN Heat-Shocked Vs Pasteurized......................................................... 221 Table 11 TKN TCCA Vs CRP ............................................................................................... 222 Table 12 TKN Adjunct culture Vs Not ................................................................................... 222 Table 13 TKN Good Vs Weak ............................................................................................... 223 Table 14 Correlation matrix for HEAT SHCOKED VS PASTEURIZATION...................... 225 Table 15 Coreelation matrix of TCCA Vs CRP ..................................................................... 226 Table 16 Correlation matrix of Adj Vs Not ............................................................................ 226 Table 17 Correlation matrix Good Vs Weak .......................................................................... 227 DEDICATED TO MY BROTHER FLAVOR DEVELOPMENT OF CHEDDAR CHEESE UNDER DIFFERENT MANUFACTURING PRACTICES CHAPTER 1 INTRODUCTION 2 BACKGROUND The quality of food products is often associated to their flavor. It is essentially determined by sensations and perceptions in our body resulting from metabolic responses to flavor compounds in those foods we eat. These impressions are collected by the set of complex detectors in our senses and scanned by our brains. Whereby, only through the presence of certain sapid and volatile compounds, at specific levels and ratios, it is possible to determine the characteristic odor and flavor of a product. Therefore, those compounds with greater impact on the perceived aroma and taste, known as character impact compounds, are matter of devoted research. Cheddar cheese is a low temperature hard rennet-coagulated cheese, traditionally made from cow’s milk, high in fat and solids, resulting in a cheese with firm consistency, with no holes and a flavor described as mild or pungent depending on age. The main operations during cheese manufacturing that induce flavor formation include milk selection, standardization and heat treatment, pre-acidification, addition of starter and adjunct cultures, coagulation, cooking, washing of curds, determination of size and shape of cheese, pressing, resting, salting, and ripening of curds. Other factors affecting flavor of Cheddar cheese include salt in moisture level and fat content (P. Walstra et al. 1999) The aim of this work was to create a model to assess quality variables during the maturation of the cheddar cheese system, which could be used to construct a set of data to propose a finger print of quality parameter to reproduce through the proper handling of processing variables. 3 LITERATURE REVIEW CHEDDAR CHEESE Cheddar cheese is a fermented milk base food product, resulting from a two stages dehydration process: 1) curds preparation and 2) ripening of curds; where fats and caseins are concentrated 6 to 12 fold. It is as well a low temperature, hard, and unwashed variety, traditionally made using mesophilic starter culture. The US Food and Drug Administration (FDA) states that cheddar cheese may be classified as a food product prepared by any procedure resulting in a cheese with a minimum milk fat content of 50% by weight of solids, and a maximum moisture content of 39% by weight (FDA 2011). Its moisture content and water activity (aw) vary from 30 to 50% and from 0.87 to 0.98 respectively, and has a pH between 5.0 and 5.3 (P. Walstra et al. 1999). Other typical composition by weight percentage for cheddar cheese is: protein (24.9%), fat (33%), total CHO (1.3%), Ash (3.9%), Ca (0.72%), P (0.51%), salt (1.8%), and salt in moisture (4.9%). In addition, about 10.6 billion pounds of cheese were produced in the United States in 2011, and around the 33% of the total production was Cheddar cheese (NASS, 2011). FLAVOR FORMATION OF CHEESE Flavor compounds in cheese arise from biochemical reactions happening during the ripening stage. These are mainly the degradation of proteins (caseins), lipids, lactose and citrate in milk, and subsequent catabolic reactions. They are grouped into: Glycolysis: metabolism of lactose and citrate Lipolysis: liberation of free fatty acids (FFA) from triacylglycerols, and subsequent metabolism to volatile compounds Proteolysis: degradation of casein matrix into peptides, and ultimately free amino acids (FAA), followed by the catabolism of FAA to produce flavor compounds, such as carboxylic acids and sulfur compounds. 4 Glycolysis Although lactose metabolism does not contribute directly to cheese flavor, it is significant at determining the texture of the cheese and therefore the rate of liberation of sapid compound. During the syneresis stage most of the lactose is lost in the whey drainage. However, the 0.8 1.5% of the total lactose remains in the cheese curd as substrate for the starter lactic acid bacteria (LAB) (Huffman and Kristoffersen 1984). In the case of Cheddar cheese, it is mostly fermented to lactic acid L+ before salting and molding (P. F. Fox, Lucey, and Cogan 1990), because LAB cultures are stimulated by low levels of NaCl and are strongly inhibited over 2.5% (Turner and Thomas 1980). Whereby, salting controls the metabolism of lactose by decreasing the starter activity and lactic acid production. When LAB activity has declined, shortly after the addition of salt, nonstarter lactic acid bacteria (NLAB), metabolizes the remaining lactose to DL-lactate and then racemizes it to Llactate (P. F. Fox, Lucey, and Cogan 1990). This last reaction is not significant for the flavor of cheese, but is determining for its structure and texture, since racemization support the development of calcium-D Lactate crystals, which are incorrectly interpreted by consumers as spoilage, resulting in product rejection. In addition lactate can be metabolized by LAB and NLAB to CO2 and acetate, which at high concentration may be perceive as an off-flavor (Aston and Dulley 1982). Alternatively, lactate can undergo anaerobic fermentation to butyrate, H2 and CO2 (P. F. Fox and McSweeney 2004) On the other hand, citrate is an important precursor of flavor compounds, and even though the 90% of the citrate in milk is lost in the whey, Cheddar cheese contains 0.2 to 0.5% that is metabolized by LAB and NLSB to diacetyl, acetate, acetoin, 2,3 butandiol and CO2 (P. F. Fox, Lucey, and Cogan 1990) Lipolysis Fat content has an important role in cheese flavor, and in addition to proportionate precursors for the development of flavor compounds, and to be solvent for hydrophobic compounds, it provides a fat-water-protein interface for catabolic reactions (P. Walstra et al. 1999). Indeed, 5 hydrolysis of lipids could be thought as the second most important metabolic reaction toward the generation of flavor compounds in cheese. Lipolytic activity is specific of the outer ester linkage of tri and/or diacylglycerides (Deeth and Touch 2000a). Esterases and lipases hydrolyze acyl ester chains between 2 and 8 carbons, and chains of 10 or more carbons respectively. The relative proportion of FFA in cheese from C6:0 to C18:3 is similar to that in milk fat, but it is higher for free C4:0, evidence that it is selectively release or synthesized by cheese microflora (Bills and Day 1964; P. F. Fox and McSweeney 1998; Paul L.H. McSweeney and Sousa 2000). Lipolytic agents are indigenous, endogenous and/or exogenous enzymes from: 1) milk; 2) rennet; 3) starter culture; 4) nonstarter culture; and 5) adjunct cultures. Milk contains a potent lipoprotein lipase (LPL) that in presence of an apolipoprotein activator (C-II apolipoprotein-glutamic acid), releases sufficient free fatty acids (FFA) to gives a rancid flavor to milk. However, LPL rarely reaches full activity due to its compartmentalization along with fat and casein micelles, being surrounded by a lipoprotein membrane, which once is damaged it may later promote off-flavors in cheese. LPL is relatively not specific for fatty acids, but is for the sn-1 and sn-3 sites of mono, di and triacylglycerides, being selective for short and medium chain triglycerides (Olivecrona et al. 1992). Due to LPL sensitivity to temperature, its contribution to flavor generation is more important in raw-milk cheeses. Indeed, the 73-95% of LPL activity is inactivated after pasteurization of milk (Yvonne F. Collins, McSweeney, and Wilkinson 2003). Usually commercial Rennets are free from lipase activity. However, Greek and Italian varieties prepared with rennet paste, contains a potent lipase, pregastric esterase, which is highly specific for short chain acids at the sn-3 position (P. F. Fox et al. 2000). Although lipolysis occurs in most cheeses, it is more noticeable in varieties made using rennet paste (longer ripening and/or developing of secondary flora). In Cheddar cheese estereolityc/lipolytic enzymes of LAB are the main lipolytic agents during cheese ripening. These enzymes are intracellular and can hydrolyze esters of fatty acids, tri, di and monoglycerides (Holland and Coolbear 1996; Chich, Marchesseau, and Gripon 1997; P. F. Fox et al. 1999; Liu, Holland, and Crow 2001). Although lactococcus and lactobacillus spp 6 are weakly lipolytic, due to their high concentration during long ripening period, they become significant in the final levels of FFA. Moreover, it has been reported that esterase activity is higher than lipase activity for many lactobacilli and lactococcus strains, such as Lb. helveticus, Lb. delbrueckii subps bulgaricus, Lb. delbrueckii subps lactis, Lb. acidophilus, Lc. lactis subps lactis, and Lc. lactis subps cremoris (Tsakalidou and Kalantzopoulos 1992). Due to LAB enzymes are intracellular, they are release by cell autolysis. And it has been reported that in Cheddar cheese the use of autolytic strains such as Lc. lactis subps cremoris AM2, result in higher levels of FFA as well as higher levels of secondary proteolysis than those for the strain Lc. lactis subps cremoris HP (Wilkinson et al. 1994; Yvonne F. Collins, McSweeney, and Wilkinson 2003). FFA are precursors molecules for many catabolic reactions resulting in flavor and aroma compounds such as methyl ketones, lactones, esters, alcanes and secondary alcohols (Gripon et al. 1991; P. F. Fox et al. 1999; Paul L.H. McSweeney and Sousa 2000). Methyl ketones are important flavor compounds generated from β-oxidations of fatty acids followed by decarboxilation supported by a low redox potential and micro-aerophilic conditions. They are very important and key compounds for the blue cheese flavor, yet they are identified as impact compounds in other cheeses (Sablé and Cottenceau 1999; Qian and Reineccius 2002). However, in full fat Cheddar cheese it has been reported that levels of heptan-2-one, non-2one, and undecan-2-one increase for 14 weeks and then decrease. In low fat milk Cheddar cheese, methyl ketones levels correspond to 25% of those in full-fat cheeses (Dimos 1992; Gerda Urbach 1993). Esters are other product of FFA catabolism. They are highly flavored and arise from the reaction between short to medium fatty acids and alcohols derived from the fermentation of lactose. In Cheddar cheese some LAB cultures hydrolyze milk fat and esterifies certain short fatty acids with ethanol, resulting in esters with fruity flavor notes such as ethyl butanoate, (Molimard and Spinnler 1996a), which is considered as defect in Cheddar cheese (Paul L.H. McSweeney and Sousa 2000). It has been reported that in Cheddar cheese esters of FFA are ethyl derivates (Arora, Cormier, and Lee 1995). 7 Secondary alcohols are product from lipolysis, and result specifically from enzymatic reduction of methyl ketones (Engels et al. 1997). In 1993 Urbach reported that in Cheddar cheese, 2-propanol is product from acetone and 2-butanol from butanone. The odors can be described as fruity, green, fuel oil like, and earthy, but the real contribution to the overall cheese flavor is limited because they have high odor thresholds Lactones are cyclic esters formed by esterification of hydroxy fatty acids. They confer a nutty, coconut and buttery-type character to cheese (Wallace J.M. and Fox P.F. 1997; Dirinck and De Winne 1999; Paul L.H. McSweeney and Sousa 2000). In addition, -lactones and lactones are present in milk and consequently in all types of cheese. Their concentrations are correlated to the extension of lipolysis, and in Cheddar cheese it has been reported that levels of lactones reach concentrations above their threshold during early ripening (Yvonne F. Collins, McSweeney, and Wilkinson 2003b). However, high levels of lactones with high molecular mass have been associated with rancid Cheddar cheese (Wong, Ellis, and LaCroix; Jolly and Kosikowski 1975). -Lactones have higher detection threshold than -lactones, and contribute to fruity notes such as coconut, peach and apricot (Dutossé et al., 1994; O´Keefe et al., 1969). It is proposed that cheese flavor is the balance of the contribution of many compounds present at certain level. Even though, some varieties have specific major contributors, like methyl ketones in mould ripening cheeses or FFA in hard Italian cheeses. However, for Cheddar cheese, the exact role of individual compounds and the right balance among them is still subject of research. Furthermore, in order to relate flavor of Cheddar cheese to levels of FFA, research has been based on: 1) determination of individual levels of FFA; 2) addition of plain bases to determine if they can be produced or improved; 3) selective removal of FFA to detect any alteration in perceptible flavor; 4) manufacture of reduce fat cheese or cheese with vegetable fat as substitute to milk fat (Wijesundera and Drury; Aston and Dulley 1982; Qian and Reineccius 2002) Proteolysis Casein metabolism is the consequence of bacteria using proteins as substrate once lactose is exhausted. This process contributes directly and indirectly to the development of flavor and off-flavor compounds in Cheddar cheese. After protein breakdown into peptides and free 8 amino acids, catabolic reactions such as transamination, deamination, decarboxylation, desulphuration and catabolism of aromatic amino acids, result in aroma compounds such as volatile sulfur compounds, aldehydes, ketones, esters, and thioesters. Proteolysis is the biochemical process that dominates the later phase of ripening (J. E. Christensen et al. 1999; Tammam et al. 2000). The decomposition of the casein network occurs due to the action of enzymes from the coagulant, the milk, the starter bacteria, nonstarter bacteria, and secondary cultures. Traditionally, cheese is manufactured by using an enzymatic coagulant extracted from abomasa of milk-fed calves known as rennet, and only up to 15% of the coagulant activity remains after whey drainage (Upadhyay et al. 2004). Chymosin (EC 3.4.23.4) and bovine pepsin (EC 3.4.23.1) are the major proteolytic enzymes in the coagulant and have a clotting activity of 88-94%, and 6-12% respectively. Specifically Chymosin coagulates milk by the rupture of the bond Phe105-Met106 in k-casein (Mulvihill and Fox 1979; P. F. Fox et al. 2000). In addition, Chymosin acts mostly on αs1-casein, by hydrolyzing the bonds Phe23Phe24 to form the peptides αs1-CN f24-199, αs1-CN f 1-23 and αs1 CN f102-109 (Richardson et al. 1974; T.K. Singh et al. 1994). Hydrolysis of αs1-casein makes the texture of curd smoother and homogeneous, and it has been acknowledged that increasing the salt in moisture level does inhibits the subsequent hydrolysis of the peptide αs1-CN f24-199 (Exterkate, Alting, and Slangen 1995). However, Chymosin activity on β-casein is extremely inhibited due to the presence of NaCl, though the presence of the peptides β-CN-f1-192 and β-CN-f193-209 is evidence of some enzymatic activity (S. ;Hup Visser 1983; Paul L. H. McSweeney et al. 1994). No major activity has been reported in αs2-casein and/or para-κ-casein (T.K. Singh et al. 1994). Therefore with high ionic strength and low water activity breakdown of α s1-casein is faster than that of β-casein (S. Visser 1993). Among the most representative indigenous milk proteinase are Plasmin (the principal and most studied one), Cathepsin D, Cathepsin B, and other proteolytic enzymes in lysosomes of somatic cells such as the serine proteinase, Elastase. 9 Plasmin acts mainly on β-casein, and due to the complex system of activators and inhibitors of the precursor Plasminogen, its enzymatic activity differs between cheese varieties. Plasmin hydrolyses β-casein at the bonds Lys28-lys29, Lys 105-His106 and Lys107-Glu108, resulting in the formation of γ1-[β-CN f29-209], γ2-[β-CN f106-209], γ3-[β-CN f108-209] caseins, along with proteose peptones. The γ-caseins accumulate during ripening while proteose peptones are hydrolyzed by starter bacteria peptidases which yield small peptides and free amino acids (Tanoj K. Singh, Fox, and Healy 1995). Cathepsin D, is an acid protease, with an optimum pH of 4.0, that produces the glycomacropeptide κ-CN(f106-169) (P. F. Fox and McSweeney 1996). However, in spite of Cathepsin D specificity is similar to that of Chymosin, Cathepsin D is not significant in milk coagulation due to its milk clotting potential is fairly low (P. F. Fox and McSweeney 1996; Larsen et al. 1996). Cathepsin D cleavage sites on αs1-casein and β-casein are similar to those of Chymosin, but significantly different from those on αs2-casein (Larsen et al. 1996). In addition, due to Cathepsin D is a heat labile enzyme, it is important in proteolysis of dairy products made with pasteurized milk and in proteolysis of rennet free cheeses. Reason why Cathepsin D activity during proteolysis of cheddar cheese is difficult to quantify (Hurley et al. 1999; V. Crow, Curry, and Hayes 2001) Cathepsin B is a lysosomal cysteine proteinase that works in the degradation of proteins by the cell. It is activated by dithiothreitol (DTT), has a pH optimum of 6.0, partially survives to the pasteurization processes, and is capable of degrading αs1-casein and β-casein extensively (Knecht 1999; Considine et al. 2004). Cathepsin B proteolytic activity is of great interest due to the activity of lysosomal enzymes has been related to the poor quality of dairy products (Grandison and Ford 1986; Verdi and Barbano 1991) Elastase is a neutral serine proteinase, and its essential physiological function is the degradation of elastin. However, it has a broad specificity on αs1-casein and β-casein, with a preferred specificity for bonds involving uncharged, non-aromatic amino acids (Naughton and Sanger 1961), cleaving 25 and 19 sited respectively (Considine et al. 2000). On β-casein, some of the Elastase cleavage sites are identical to, or near to those cleaved by Plasmin, 10 Chymosin or cell envelope-associated proteinase of several strains of Lactococcus (Considine et al. 1999). Regarding to the starter lactic acid bacteria (LAB), the principal starter cultures used in cheese manufacturing are the mesophilic Lactococcus and Leuconostoc species, and the thermophilic Lactobacillus and streptococcus thermophiles species. Their main role is to decrease the pH by producing lactic acid from lactose. Lactic acid bacteria (LAB) possess a complex proteinase/peptidase system, very important during the secondary proteolysis, which includes: 1) a cell envelope proteinase, lactocepine; 2) the intracellular oligoendopeptidases PepO and PepF; 3) the general aminopeptidases PepN, PepC, PepG, along with the glutamyl aminopeptidase (PepA), the pyrolidone carboxylyl peptidase (PCP), the leucyl aminopeptidase (PepL), the X-prolyldipeptidyl aminopeptidase (PepX), the proline inmunopeptidase, the aminopeptidase P (PepP), the prolinase (PepR), and the prolidase (PepQ); 4) the general dipeptidase PepV, PepD, PepDA and the general tripeptidase (PepT) (Sousa, Ardö, and McSweeney 2001). These peptidases can be classified into: 1) endopeptidases, relevant for the degradation of oligopeptides to shorter peptides; and 2) exopeptidases such as carboxypeptidases or aminopeptidases, which release free amino acids from short peptides. The most important enzyme is lactocepine, a serine proteinase that degrades intermediate size peptides produced from Chymosin and Plasmin activity (Upadhyay et al. 2004). However, since caseins are rich in proline and because of its particular structure, proline specific enzymes have significant contribution to the proteolysis of cheese, by making released peptides accessible to other peptidases. Although the role of non-starter lactic acid bacteria (NSLAB) in the development of the flavor of cheddar cheese is not totally understood, it has been used to manipulate the final sensory characteristics of the product. Indeed, when LAB population declines, NSLAB population becomes the dominant microflora in the maturation of cheese (Peterson and Marshall 1990; V. L. Crow et al. 1995; P. Fox, McSweeney, and Lynch 1998). NSLB are essentially constituted by homo and hetero-fermentative species of lactobacilli and are determinant during the secondary proteolysis. However, hetero-fermentative and certain Lactobacillus strains have been associated with off-flavors in cheddar cheese (Puchades, Lemieux, and Simard 1989), and it has been reported that thermophilic lactobacilli do not influence the development of 11 cheddar flavor (Lloyd, Horwood, and Barlow 1980). Actually, it has been suggested that the addition of selected adjunct strains of Lactobacillus spp, positively influence the quality of cheese (Drake et al. 1996; C.N. Lane and Fox 1996a; C. M. Lynch et al. 1996; Muir, Banks, and Hunter 1996), and accelerate the ripening of standard and reduced-fat Cheddar cheese (M. A. El Soda 1993; Christensen J.E., Johnson M.E., and Steele J.L. 1995) Thus, due to there are not criteria for the selection of adjuncts, research has been done to clarify the proteolytic and lipolytic systems of NSLAB which contribute to the maturation FACTORS AFFECTING THE DEVELOPMENT OF VOLATILE COMPOUNDS Cheese flavor development and most of its chemical and physical properties relates to microbiological and biochemical events that depend on physicochemical parameters, established during the early stages of cheese manufacture. Factors such as pH, water activity (aw), buffer capacity, oxidation-reduction potential (redox), physical history of milk and size and geometry of curds are set during the manufacturing of curd, which as mentioned above it is a dehydration process in which fat and caseins are concentrated 6 to 12 fold through the following operations: 1) Preparation of milk (pasteurization); 2) Acidification (LAB); 3) Rennet coagulation; 4) Syneresis; 5) Pressing and shaping the curd; 6) Salting; 7) Other operations. However, other physical factor such as time and temperature are relevant dynamic factors during the ripening stage that are of great interest in the discussion of cheese flavor. 12 Milk preparation: selection, pasteurization & standardization After refrigeration of raw milk, microflora is dominated by psychrothrops which produces heat stable lipases and proteinases, and in concentrations over 106 CFU/ml, they reduce yield or can cause development of off-flavors during ripening (Payne and Kroll 1991) Mainly, because of heat-induce changes in cheese micro-flora, cheese made from raw milk ripens faster and develops stronger flavors than those made from pasteurized milk (P. F. Fox and McSweeney 1998). In addition to the changes in microflora, heat treatments of milk, such as pasteurization or heat shock, causes inactivation of indigenous enzymes (mainly lipoprotein lipase inactivation), resulting in cheeses with weaker flavor and slower ripening (P. F. Fox and McSweeney 1998; Y.F. Collins, McSweeney, and Wilkinson 2004). On the other hand, standardization is a process through which is possible to control the ratio of casein to fat, influencing in this way parameters such as moisture and moisture in non fat substances, improving cheese yield. Some of the different methods to pre-determine the composition of milk to assure a homogenous production during the year involve the use of membrane concentration techniques or vacuum concentration techniques. In the case of cheddar cheese (full fat and reduce fat cheeses), most of the research work is related to vacuum condensation, from where industrial and academic results show that concentration cannot exceed 1.8:1, because at higher concentration sweetness and saltiness problems occur due to excessive lactose and minerals (Anderson et al. 1993). Contrary, membrane concentration techniques (reverse osmosis, nanofiltration, microfiltration and ultrafiltration) are common in cheddar cheese production. Indeed, low concentration or protein standardization by ultrafiltration is a popular method to secure the uniformity of milk composition, lower casein loss through a firmer curd, more efficiency and better yield (Mistry and Maubois 2004). Additionally, different dairy products such as milk powder, milk protein, milk permeate can be used as means to standardize components in cheese milk (Mistry, Metzger, and Maubois 1996). 13 Acidification In addition to initiate the fermentation of lactose to lactate, during the acidification, starter cultures provide enzymes for the ripening stage, promoting as well the prevention of spoilage because of the reduction of redox potential of the system. Due to Lactic acid bacteria (LAB’s) are auxothropic for many amino acids, they have a proteinase and peptidase systems to liberate amino acids from caseins. Also, they poses intracellular metabolic enzymes which catalyze the catabolism of amino acids and contribute to the formation of volatile flavor compounds (Yvon and Rijnen 2001; Y.F. Collins, McSweeney, and Wilkinson 2004; P. F. Fox and McSweeney 2004). Consequently the reduction of pH due to the fermentation of lactose to lactate and lactic acid determines: 1) The retention of coagulant activity in the curd; 2) The rate of Syneresis; 3) The growth of added and native microorganism; 4) The enzymatic activity during ripening. LAB for cheddar cheese is a mesophilic culture with optimum temperature for growth at 2030 C, so when the cooking temperature is higher than the optimum the rate of syneresis is increased but the acid development decreases. Coagulation As it was mentioned above, it occurs via limited proteolysis of k-caseins at or near Phe105Met106 bond followed by the aggregation of the Ca2+ rennet altered micelles at temperature over 18 C (Home and Banks 2004). The traditional rennet is a brine of the abomasa of milk fed calves (or other young dairy animal), which contains mainly Chymosin and low levels of pepsin. The retention of the rennet activity added to milk varies from 0-15% depending on factors such as type of enzyme, pH at whey drainage, cook temperature, and moisture content of the curd (Upadhyay et al. 14 2004). Thus, later on during the ripening stage, parameters for coagulation will influence the degree of hydrolysis of αS1-casein, the generation of flavor compounds, texture of curds and cheeses, and subsequently the liberation of sapid compounds. Ph determines the casein micelle matrix, and along with other factors such as heating, protein concentration in milk, and addition of Ca2+, it establishes the equilibrium between colloidal and dissolved calcium phosphate and between dissolved calcium phosphate and other ions. For example acidity reduces the negative charge on the micelles (caseins isoelectric point is 4.6) and increases solubility of citrates (pH 5.5 completely soluble) and colloidal calcium phosphate (pH 5 completely soluble). Heating reduces dissolved calcium and phosphates, and promote coagulation by association of colloidal phosphates with the casein micelles. Milk concentration increases buffer capacity and colloidal calcium phosphate and its association to the micelles. Therefore, it is possible to observe how coagulation depends on dynamic properties like pH and the solubility of calcium salts, and the cleavage of κ-caseins by rennet enzymes. Thus, defects such as sour and/or bitter flavor and soft and pasty body are associated with excessive acidity and a pH under 5.0. Syneresis The gel formed due to the rennet induced coagulation is stable, but once it is broken, most of the liquid entrapped in the gel is expelled readily as whey. Syneresis and consequently moisture content of cheese are controlled by the milk composition, size of the curd particles, cooking temperature, time, rate of acidification and rate of stirring the curd-whey mixture (P. F. Fox and McSweeney 2004). Therefore, the mentioned parameters and the operations involved with Syneresis have an impact on ripening. Reason why high moisture cheese ripens faster than low moisture cheese. In addition to promote syneresis, high cooking temperature inactivates the remaining Chymosin and increases the levels of Plasmin due to denaturation of Plasmin inhibitors and inhibitors of Plasminogen activators (Farkye and Fox 1990). High temperature during rennet gelation increases the initial porosity of the gel and consequently the initial rate of syneresis after the curd is cut. The rate of syneresis is also 15 increased by smaller curd size at cutting and a vigorous agitation (Pieter Walstra, Wouters, and Geurts 2006). Cheddaring During cheddaring the pH of curds decrease to 5.4, causing dissolution of colloidal calcium phosphate, which modifies the texture of the curd by altering Ca:protein ratio. Some of the objectives of the cheddaring process are to remove small amount of whey to allow a better acid development, to have moisture control, to develop a proper texture, and curiously to repress the growth of gas forming spoilage organisms. Salting Salting can be achieved by immersion in brine (most chesses), by mixing dry salt with the milled curd, which is the case of Cheddar cheese, or by application of salt on the surface after molding. The brine option is intended for cheeses with high level of salt and the rate at which salt in moisture increases is slow since NaCl should diffuse from the surface. Contrary in the dry salted varieties NaCl uptake is very fast (Guinee 2004). This process influence the flavor profiles of cheese by controlling microbial growth, determine enzyme activity and impact on water activity. Salting affects bacterial and enzymatic activity. Indeed, inhibition of acid production occurs at NaCl concentrations over 1.5%, which has been used in some cheese varieties to stop acidification and to fix the pH (Guinee 2004). In addition, it is well known that microbial enzymes can be inhibited or stimulated by moderately high NaCl levels, particularly at low pH, depending on the type of enzyme. As matter of fact, approximately 2 kg of H2O are lost by absorption of 1 Kg of NaCl , which means a reduction on the water activity of the system. For example, Plasmin (principal indigenous proteinase in milk) is stimulated by 2% NaCl but inhibited at high concentration (Farkye and Fox 1990), but for Chymosin (principal enzyme in the rennet), the addition of NaCl increases the ionic strength, promoting interactions between hydrophobic C-terminals of β-caseins, inhibiting the access of the enzyme to cleavage sites. Thus decreasing NaCl levels facilitates Chymosin action on β-caseins and production of hydrophobic peptides. 16 In the case of cheddar cheese most of the acid is developed before salting and molding, therefore cooking time and temperature along with culture activity determine curd pH, buffer capacity and rennet retention at draining. Pressing and molding Curds for brine salted are molded prior to salting while those for dry-salted are molded and pressed after addition of salt. Cheese size is not determined only due to esthetic reasons, contrary because of many biochemical events occur at the surface of the cheese during the ripening stage, thus controlling the ratio of surface area to volume is key to control and assure an homogenous ripen, and depending on the variety, it allows the development of certain characteristics such as eyes formation due to sufficient partial pressure of CO2. In the case of cheddar the time and temperature of pressing and the temperature during the first days of ripening influence the extent of acid development and the minimum pH. Other important factor involved with flavor development is the oxidation/reduction system of milk, which include Fe2+/Fe3+, Cu+/Cu2+, dehydro ascorbate, rivoflavin, and lactate/pyruvate (P. Walstra et al. 1999). By decreasing the pH and increasing temperature free sulfydryl groups are produces in proteins and amino acids lowering the redox potential. Usually the redox potential decreases during setting, rises during, cutting, cooking and draining, decreases during cheddaring, increases during milling, and decreases during pressing and ripening. Maintaining a low Eh (-150 to -300 mV) promotes the production and stability of sulfur volatile compounds (G. Urbach 1995; Beresford et al. 2001) Therefore, after the early stages of cheese making, the main factors that determine the structure and flavor of most varieties are: 1) the extent of acid production in the vat; 2) amount of starter culture 3) the residual plasmin and rennet: 4) the pH of curds at draining; 5) the residual lactose; 6) and the mineral content of the curd. 17 CHEESE FLAVOR AND FLAVOR ANALYSIS As a response to the need for exact and predictable results of the ripening stage, monitoring the ripening process has been of interest for cheese makers and food scientist, especially when large productions are involved and “standardization of flavor” is associated to high quality products by the consumer. Thus, different methods to monitor the extension of ripening have been developed, and this work will be based on a metabolic approach. From this perspective, ripening can be followed by determining the metabolite products from primary carbon metabolism paths such as 1) the conversion of lactose and citrate; 2) lipolysis; 3) and proteolysis. Since the degradation and conversion of caseins is the most important biochemical step for flavor formation in hard and semi-hard cheeses, tracking the progress of proteolysis is an excellent indicator of the ripening process. There are different analytical techniques to monitor patterns of proteolysis; one of them is a fractionation scheme to extract nitrogen compounds based on pH. Nonetheless, choosing the scheme and methodology depends on 1) the availability of equipment and resources; 2) the cheese variety and its characteristics; and 3) the objective of the study. (Ardö and Frederiksberg 1999; Sousa, Ardö, and McSweeney 2001). These fractions can be used to obtain information about one or more proteolysis agents, such as action of certain enzyme, comparison of coagulants, and analysis of different starters or adjunct culture. The insoluble fraction can be used to study effects on primary proteolysis by means of Urea-polyacrylamide gel electrophoresis or by capillary electrophoresis. As well insoluble and soluble fractions can be used to determine peptide profiles by reverse phase high performance liquid chromatography (RP-HPLC), and to perform an analysis of individual amino acids. However, it is important to remember that proteolysis is too complex to be described by a single index, and in addition other chromatographic techniques involving mass spectrometry such as HPLC MS-MS or GC-MS are a more powerful approach for identifying and monitoring specific key flavor and taste compounds in a fast and accurate way (Cserháti 2002; Careri and Mangia 2003). Regarding the cheddar cheese making process, other metabolites could be tracked too; however, not all of them can be use because of certain limitations related to the metabolic 18 path. For example, the remaining lactose in cheddar cheese cannot be considered as a relevant parameter to monitor the ripening progress due to the remaining lactose is metabolized to Llactic acid, ethanol and CO2 within the first few weeks. And in spite that ethanol can esterified with free fatty acids to produce ethyl-esters, compounds associated to fruity notes, they are not good descriptors either. On the other hand, despite lipolysis is very limited for this variety, it is an important metabolic path to track, where the low but steady increased in FFA’s, especially those levels for C4 to C8 ones, can be use as an indicator of the progress of the ripening (Yvonne F. Collins, McSweeney, and Wilkinson 2003b; Y.F. Collins, McSweeney, and Wilkinson 2004). However, it is important to keep in mind that this last one as well as other indicators of proteolysis does not correlate directly to the production of relevant volatile compounds. Furthermore, the aroma analysis highly relies on the technique for extraction and concentration use to isolate the different aromas compounds. And due to there is not a single universal method to extract all kind of aroma compounds at once, many techniques has been used to study the aroma compounds of cheddar cheese, such as: solvent extraction, simultaneous distillation extraction, static and dynamic head space, purge and trap, ion exchange chromatography and solid phase microextraction. Additionally, for works intended for tracking the progress of ripening, the methodology selected should focus on the expected results as consequence of the assessed agent or parameter. By using the methods mentioned above, a wide variety of compounds such as acids, esters, ketones, aldehydes, alcohols, lactones, phenols and volatile sulfur compounds have been identified in cheddar cheese. Most volatile aroma compounds are found in the lipid content because they are hydrophobic. Thus, in the case of fatty acids it is relevant to know the individual concentration because each one has different sensory threshold and aroma attributes and contribution. They can be determined by gas chromatography (GC) without derivatization, but they require separation from triglycerides and others lipids such as cholesterol and phospholipids, prior to GC analysis. This is done by the use of aminopropyl weak anion exchange columns, in which the 100% of the recoveries have been achieved and a good repeatability can be expected (De Jong and Badings 1990; Chavarri et al. 1997; Qian and Reineccius 2002). 19 Based on the research done on amino acids catabolism (D. J. Manning, Chapman, and Hosking 1976; B. Weimer, Seefeldt, and Dias 1999; Stuart, Chou, and Weimer 1999), particularly by LAB enzymes, breakdown of key compounds such as phenylalanine (aromatic), leucine (branched chain) and sulphurous (methionine and cysteine), have been characterized. Thus, volatile sulphur compounds (VSC’s) such as methanethiol, dimethyl sulphide, dimethyl disulphide and dimethyl trisulphide, most of them with low odor threshold, are important flavor compounds that can be used as tracking metabolite for monitoring the progress of the ripening stage. VSC’s cannot been studied by conventional techniques including static head space and purge and trap due to the loss of analyte during concentration stage and the potential formation of thermal artifacts, particularly in these case where the studied compounds are highly volatile and chemically reactive. A better approach is the solid phase microextraction technique (SPME) along with the a sulphur pulsed photometric detector (PFPD), which makes possible the analysis of VSC’s in food faster and without tedious sample preparation involving the use of solvents (Fang and Qian 2005; Vazquez-Landaverde, Torres, and Qian 2006; H. M. Burbank and Qian 2005) . Nonetheless, there are many kind of fibers, but the most appropriate type for the study of dairy products is Carboxen-polydemethylsiloxane (CAR-PDMS), which can readily extract high volatile and low molecular compounds including VSC’s Instrumental analysis of cheddar cheese Chromatography methods are essential in understanding the aroma chemistry of cheddar cheese, and consequently they offer a thorough picture of the impact of manufacturing variables during the production process on the ready to sell product. Some of the preferred methods are gas chromatography (GC), including the use of detectors such as flame ionization detector (FID), pulsed flame photometric detector (PFPD) and mass spectrometry (MS) among others, and high performance liquid chromatography (HPLC), operating in the reverse phase separation mode. However, cheese matrix is complex and samples cannot be injected directly into the injection port of the devices, and as consequence isolation of volatile, nonvolatile and/or sapid compounds is required in order to protect the instruments and to avoid degradation of samples and formation of artifacts due to the high temperatures in the GC 20 injection ports. Thus, to achieve meaningful and reproducible results, isolation and concentration of aroma compounds is mandatory, and depending on the chemical composition of the target analytes, different approaches can be use to perform their separation from the matrix. Therefore, procedures involving pH, solubility, polarity and volatility are often used in the aroma analysis of cheddar cheese. Gas Chromatography The separation, identification, quantification and analysis of volatiles is generally complicated due to their concentration levels, which could be as low as parts per billion (ppb) in the nonvolatile matrix. The most common isolation methods used for volatiles are solvent extraction and distillation, and headspace techniques, including static headspace analysis, dynamic headspace analysis and headspace-solid phase microextraction (SPME). Extraction-Distillation Cheese is usually low moisture, high fat and high protein product, and in spite of that difficult matrix, solvent extraction became a common procedure to separate volatiles. It involves the use of organic solvents such as acetonitrile, dichloromethane and diethyl ether, where the last one is considered as the most suitable for cheese analysis due to its low density, low boiling point and high selectivity for aroma compounds. Moreover, purity of solvents is critical, which implies an obligatory pre-distillation/purification process before its use. In the case of cheddar cheese and other hard and semi-hard cheeses, the samples are usually frozen employing liquid nitrogen, then grated or grinded, and finally the analytes are extracted with the selected solvent (Preininger and Grosch 1994; Milo and Reineccius 1997; Suriyaphan et al. 2001; Zehentbauer and Reineccius 2002; Qian and Reineccius 2002; Avsar et al. 2004; Mary E. Carunchia Whetstine, Cadwallader, and Drake 2005), or alternatively, an aqueous extract is prepared followed by solvent extraction (Moio et al. 1993). However, because cheese is not a free-fat product, an additional extraction is required and usually done by dialysis (Benkler and Reineccius 1980) or by a low-temperature high vacuum distillation (Suriyaphan et al., 2001) to separate non-polar and non-volatile lipids, and other non-volatile constituents, followed by the separation of analytes into neutral, basic and acidic fractions if it is necessary. 21 The approach through direct solvent extraction is effective to separate semi-volatiles such as lactones, free fatty acids and phenolics, but has some disadvantages such as evaporation of large quantities of solvent, being a slow process, and adsorption of components of the membrane surface and catalysis of acetone condensation in the case of the dialysis (Benkler and Reineccius 1980). Regarding distillation as method to purify the solvent extracts, high vacuum distillation and steam distillation techniques are suitable when the sample will be analyzed by GC on capillary column. The most popular techniques are simultaneous steam distillation extraction (SDE) (Chaintreau 2001) and Solvent assisted flavor evaporation distillation (SAFE) (Engel, Bahr, and Schieberle 1999). The first one has been used for the analysis of aqueous slurry of Cheddar, Gouda, Edam, Swiss and Parmesan cheese, with the Likens-Nickerson extractor using diethyl ether, and following a procedure described by Parliament (1998). However, this technique can lead to artifact formation due to it is performed at high temperatures. Regarding SAFE, it has been reported that the method resulted in better recovery than the traditional high vacuum transfer, and it was effective for most volatiles from solvent extracts of cheese (Werkhoff et al., 2002; Carunchia-Whetsine et al., 2005, 2006; Cadwallader et al., 2006; Schlichtherle-Cerny et al., 2006) Headspace methods As it was mentioned above, headspace methods include static headspace analysis (SHA), dynamic head space analysis (DHA) and headspace-solid phase microextraction (SPME), which are limited to the equilibrium of volatile compounds into the gas phase, reason why they are considered non destructive and require minimal sample preparation. SHA is used when analysis of major component is satisfactory. In this technique, the sample is contained in a closed vessel and volatiles are allowed to reach the equilibrium by partition into the head space and the matrix, which is a process influenced by temperature, vessel size, ratio of sample to head space volumes, addition of salt, and agitation. Then an aliquot of the headspace is taken and injected into the GC. SHA has been used for the analysis of highly volatile sulfur compounds such as hydrogen sulphide, methanethiol and dimethyl sulphide (Lin and Jeon 1985), acetaldehyde and other low molecular weight Strecker aldehydes such as 22 methyl propanal, 2-methyl butanal, and 3-methyl butanal (Fernández-García 1996), which are important contributors to the cheddar cheese flavor. However, this technique lacks of sensitivity and is really difficult to standardize. By using a flow of carrier gas and an intermediate adsorption or cryogenic step before the injection into the GC, larger amounts of volatiles can be collected and the efficiency of the headspace analysis is improved, turning it into Dynamic headspace analysis (DHA) or purgeand-trap analysis. Usually the carrier gas could be helium or nitrogen, while the adsorbent might be a polymeric material such as Tenax (poly-2, 6-diphenyl-p-phenylene oxide) or active charcoal. Adsorbent trapping is more common than cryogenic focusing, because it prevents the damage of the GC column. Once the volatiles are isolated, they are desorbed by heating, they could be concentrated by cryo-focusing, and then they are transferred to the GC column by thermal desorption. Some of the advantage is the low risk of artifact formation, the minimal sample preparation and the fact that it is solvent-free. However the principal limitation is that the method is not efficient for semi-volatiles. Extracts by this technique have been analyzed by multidimensional CG-O/FID/MS and in total 5 aldehydes, 6 ketones, 8 alcohols, 3 esters, 11 hydrocarbons, 3 halides and 3 sulfur compounds were identified (Arora et al., 1995/ paper; Dunn and Lindsay, 1985; Barbieri et al, 1994; Thierry et al, 1999, 2004; Larrayoz et al, 2001; Rychlik and Bosset, 2001a; Valero et al., 2001; Qian and Reineccius, 2002; Boscaini et al., 2003; Avasar et al., 2004). Another solvent-free isolation method is SPME, which can concentrate volatiles from various matrices in a single step (Kataoka and others, 2000). It reduced the time required for sampling and the cost of analysis, and it became the most common method for analysis of volatiles in the last decade. It is based on the partition of the analytes in the headspace and the polymer coated fiber. The method counts with great selectivity and specificity due to the several adsorbents phases and film thicknesses available. The adsorption depends on temperature, vessel size, ratio of sample to head space volumes, addition of salt, agitation, nature of the coating and exposure time, for which is recommended short times (1-5 min) for highly volatiles and (5-30 min) for semi-volatiles (Roberts et al., 2000). Volatiles are transfer to the GC by thermal desorption in the splitless injection mode. Cheddar cheese was initially analyzed using fibers coated with non-polar polydimethylsiloxane (PDMS) and polar 23 Polyacrylate (PA), showing better results for the polar coating (Chin et al., 1996). Volatile components such as fatty acids and δ-lactones were found, and differences in profiles were observed between varieties. Later, the technique was consolidated for the analysis of cheddar cheese aroma, by a comparison polydimethylsiloxane/divinylbenzen of five types (PDMS-DVB), of PA, fiber coatings, carboxen/PDMS PDMS, and carbowax/DVB. The study revealed that bipolar coatings PDMS/DVB and carboxen/PDMS showed better selectivity, measured by the amount of peaks absorbed, besides confirming the relevance of the adsorption temperature and exposition time (Dufour et al., 2001). Initially it was suggested that unlike solvent extraction techniques or DHA, quantification of volatiles was not possible, however it was demonstrated the fact that it was possible to perform a quantification of volatiles extracted by SPME. Separation The injection technique and the analytical column stationary phase could be considered as the most critical parameters influencing the result of the GC analysis. Regarding the first one, programmable temperature vaporizer (PTV) and on-column injectors are the best options for analysis of aroma extracts. They provide versatility, which make them even suitable for works with cryogenic focusing from DHA, and ramped heated injections in the splitless or split modes. Besides, they proportionate the possibility of avoiding thermal degradation in the cool on-column injection for extracts from direct solvent extraction coupled with high vacuum distillation. On the other hand, polarity of the stationary phase determines the type of analytes that could be identified. Thus, DBWAX and FFAP phases constitute the adequate options for analysis of polar compounds, while DB-5 phase is the right choice for non-polar compounds. To study aroma compounds that make a contribution to the odor of a food, there are different approaches including gas chromatography-olfactometry (GCO), odor activity value (OAV) calculation, and sensory analysis. In GCO, the most often used techniques are the dilution analysis, where the most popular ones are the aroma extract dilution analysis (AEDA) (Grosch, 1993), its variation called aroma extract concentration analysis (AECA) (Kerscher and Grosh, 1997) and GCO-headspace dilution analysis. In AEDA a dilution series, or concentration series in the case of AECA, of an aroma extract is evaluated by GCO, where compounds are ranked according to its potency based on the highest dilution or concentration that can be perceived, which is defined as flavor dilution factor (FD). However, FD factors do 24 not account for highly volatile compounds lost during extraction and concentration, underestimating important compounds. GCO-headspace dilution analysis provides a complementary evaluation of the aroma composition. And in these methods, dilutions are achieved by decreasing the headspace volume, or the purge gas volume in the case of the DHA. In addition, retention indices are used for confirming a proper identification of compounds contributing to the aroma of a food. These are based on the retention times of target compounds compared to those of certain standards (usually hydrocarbons). In addition, OAV relates analytical data to sensory characteristics of a sample, and essentially it establishes a relation between the concentration of a target compound in the food to its odor threshold; allowing to discriminate accurately the compounds that really contribute to the specific aroma of a food. To the date, something to keep in mind is that the methodologies mentioned above coupled with the separation efficiency of gas chromatography and the excellent identification potential of mass spectrometry, offer to scientists and engineers the possibility to understand, describe and improve production processes towards the satisfaction of customers, by maintaining constancy in the quality of a product, in this case the traditional aroma of Cheddar cheese. Reverse Phase-High Performance Liquid Chromatography (RP-HPLC) The assessment of proteolysis through peptide profiling of cheese extracts by RP-HPLC, is widely used to study quality and authenticity of samples. It is based on the principle that caseins might not be soluble in certain solvents, but peptides resolved from their degradation can be. What is evident when the amount of intermediate and small peptides increase as the proteolysis of caseins takes place during ripening cheese (Bansal, Piraino, and McSweeney 2009; Piraino, Parente, and McSweeney 2004). Most of the HPLC separations are done in the reverse-phase mode, recognized by the use of a non-polar stationary phase and a polar mobile phase, where solutes are mainly retained due to hydrophobic interaction with the non polar one, and are eluted in order of decreasing polarity. Indeed, solutes retention decreases by increasing the organic solvent content of the mobile 25 phase. And as in the case of GC, samples cannot be directly injected in to the instrument, meaning that isolation of analytes is previously required. The most common way to perform the extraction of peptides from cheese, is following a fractionation scheme that involve the use of solvents such as water, buffers at pH 4.6, Trichloroacetic acid (TCA), phosphotungstic acid (PTA), and ethanol (Ardö and Frederiksberg 1999). This, in addition to be a sort of sample preparation, allows measuring of proteolytic activity through the analysis of nitrogen content of resolved fractions, and RPHPLC analysis of water and pH 4.6 soluble extracts. As a matter of fact, fractionation with water is a method use for mature cheddar cheese due to its low and relatively constant pH (Ardö and Frederiksberg 1999). Furthermore, water soluble nitrogen extracts (WSN) have been used as index of ripening, which is only employed when there is no variation of pH during ripening or between samples. Whereby, Bansal and others (2009), have suggested that pH 4.6 soluble nitrogen should be used instead of WSN as index of primary proteolysis. However, it is important to keep in mind that results from this method are somewhat smaller, it is more difficult to perform, but is easier to standardize. On the other hand, because cheese is a dynamic system, meaning that results of analysis depend on the age of samples (Sousa, Ardö, and McSweeney 2001), RP-HPLC analysis of water and/or pH 4.6 soluble extracts can be used as a discriminant technique for the characterization of secondary proteolysis. Regarding TCA, it has been used to precipitate peptides from water and pH 4.6 soluble extracts, at concentration ranging from 2 to 12%, which depends on the degree of fractionation required (Bansal, Piraino, and McSweeney 2009). It has been reported that during the fractionation with TCA, parameters such as extraction time and extraction temperature have little or none effect on the amount of nitrogen obtained. In contrast, the amount and type of peptides that can be extracted vary according with parameters such as cheese to water ratio, pH, NaCl content of cheese and type of previous fraction (Polyachroniadou et al., 1999 chapter). Currently, ethanol, ranging from 30 to 80%, has been used as an alternative to TCA because it offers similar precipitation levels and it can be easily evaporated for further analysis of peptides in this fraction. Phosphotunstic acid (PTA) is a very discriminant protein precipitant that is employed in a range from 1 to 6.5%, where only free amino acids (expect lysine and argentine), and peptides under 600 Da are soluble. However, there are other 26 techniques to perform the fractionation based on peptides molecular mass rather than on solubility such as dialysis, ultra filtration (UF), and size exclusion chromatography, which are really useful for taste panel work due to they are solvent free (Fox, 1989 chapter). Separation and characterization of peptides has been done by RP-HPLC analysis of WSN extract, pH 4.6 soluble and insoluble extracts, 10 kDa UF permeate, 70% soluble and insoluble extracts, and fractions from gel permeating chromatography. The elution of analytes takes place in a Nucleosil RP-8 (250 × 4.6 mm, 5 μm particle size, 300 Å pore size) analytical and guard columns (4.6 × 10 mm), and is usually done in the gradient mode using system such as water/acetonitrile or water/methanol (Allan J. Cliffe, Marks, and Mulholland 1993); however it is possible to perform it by isocratic elution using a phosphate buffer as mobil phase (Pham and Nakai, 1984). The detection of analytes is done by monitoring the carbonyl group in the peptides bonds, using a UV detector at wavelength ranging from 200 to 230 nm, for which the most used ion-pair reagent is trifluroacetic acid (TFA). Due to peptide profiles are multivariate in nature, the identification of analytes from raw data has been done by visual matching (A.H Pripp et al. 1999; Are Hugo Pripp, Stepaniak, and Sørhaug 2000), or by division of chromatograms in classes of retention time and peaks in each class (Barile, 2006) followed by mass spectrometry. Whereas, data analysis has been done by descriptive techniques such as Principal Component Analysis (PCA) (A.H Pripp et al. 1999; Are Hugo Pripp, Stepaniak, and Sørhaug 2000; Piraino, Parente, and McSweeney 2004), or by descriptive and inferential techniques such as Linear Discriminant Analysis on Principal Component Scores (Hynes et al., 2003) and (O’Shea et al., 1996), Partial Least Squares Regression (PLSR) and/or Partial Least Squares Discriminant Analysis (PLSDA). Electrophoresis Besides chromatographic methods such as GC and RP-HPLC, another analytical method for assessment of proteolysis is electrophoresis, which is a specific technique that in addition to give information about the extent of proteolysis and the general contribution of proteolytic agents, also resolves, isolates and identifies peptides (Bansal, Piraino, and McSweeney 2009). Indeed, it is a technique applied to the study of cheese ripening, and is particularly useful for 27 the comprehension of primary proteolysis, since it is limited to monitoring hydrolysis of parent caseins, where only protein and large peptides can be visualized. Among the different electrophoretic methods used in the study of cheese ripening, it is possible to find works with paper electrophoresis, free boundary electrophoresis, high voltage paper electrophoresis, starch gel electrophoresis, isoelectric focusing, capillary electrophoresis (CE), and polyacrilamide gel electrophoresis (Fox 1989; Mcsweeney and Fox 1993; Fox et al, 1995; Bican and Spahni, 1993; Trieu-Cuot and Gripon, 1982; Addeo et al 1983, 1990; Moio, Luccia and Addeo 1989, 1992; Creamer, 1992; Amigo et al, 1992; Strange et al 1992; Ledford et al., 1966; Shalabi and Fox 1987; Andrews 1983; Blakesley and B, 1977; Lindeber 1996; Goulds Worthy et al 1999; Bansal, Piraino, & McSweeney, 2009). The last one is the most common method applied, it usually works with discontinuous buffer systems, utilizing urea or sodium dodecyl sulfate (SDS) as dissociating agents. However, in the case of cheese analysis, due to caseins have similar molecular weights, it has been reported that buffer systems using urea are more sensitive and adequate than those using SDS, facilitating in this way to resolve proteins or peptides of comparable size. Nonetheless, other works using SDS as dissociating agent suggest that valuable information can be obtained. The most recommended method for the study of cheese involves the stacking system of Andrews () in alkaline gels containing 6M urea and the direct staining procedure of Blakesley and Boezi, with Coomassie blue G250. Even so, it is reported that low molecular peptides can be visualized using a silver staining technique involving glutaraldehyde as fixing agent, which has not been applied yet to cheese analysis. On the other hand, the arrangement between electrophoretic methods, or the application of a separation method followed by electrophoresis, which is known as two dimensional electrophoresis, has been widely used in the study of cheese ripening. Examples of this are works where SDS-PAGE is one dimension and isoelectric focusing is the other, or where thin layer chromatography (TLC) is followed by electrophoresis. Usually, peptides resolved on PAGE are isolated and identified by excision of the bands or by electroblotting. This last one is the most utilized, and peptides are identified by N-terminal 28 amino sequencing rather than mass spectroscopy (MS), due to they are stained. Nonetheless, most of the major degradation products are known in most PAGE systems. An alternative and potentially strong methodology to PAGE systems is capillary electrophoresis (CE), which to date, is a technique not well implemented in the study of cheese proteolysis, and counts with the capacity to resolve complicated mixtures of peptides, using buffer filled capillary and an electric field that promotes separation based on the net charge, molecular mass and Strokes’ radius. Aroma of Cheddar cheese Volatile flavor compounds identified in cheddar cheese includes a wide variety of acids, alcohols, esters, aldehydes, ketones, phenolics and sulfur compounds summarized in the following table. Table 1 Aroma compounds of Cheddar cheese COMPOUND Dimethyl sulphide Dimethyl disulphide Dimethyl trisulphide Hexanethiol TYPE OF COMPOUND Volatile sulfur compound Volatile sulfur compound Volatile sulfur compound Volatile sulfur compound Hydrogen sulfide Methanethiol Acetic acid n-butanoic acid Volatile sulfur compound Volatile sulfur compound Organic acid Organic acid n-decanoic acid Isovaleric acid butanoic acid Hexanoic acid Butyric acid 3-methyl n-octanoic acid n-pentanoic acid Phenyl acetic acid Propionic acid β - angelicalactone γ - decalactone δ - decalactone δ - dodecalactone 6-(Z)-dodecenyl- γ - decalactone Organic acid Organic acid Organic acid Organic acid Organic acid Organic acid Organic acid Organic acid Lactones Lactones Lactones Lactones Lactones ATRIBUTE Boiled cabbage Cabbage, strong onion Ripe cheese, garlic burnt fat, sulfury meaty, fatty garlic roasted burnt Rotten egg Rotten cabbage, fecal Vinegar, sour, pungent Sweaty, cheesy, fecal, rancid, sharp Rancid, waxy, soapy Swiss cheese, waxy, sweaty, old socks, fecal Goat like Sharp, dairy-like, cheesy, buttery with a fruity nuance Body odor, sweaty Swiss cheese Flowery pungent Coconut Peachy, coconut Cheesy, coconut Soapy 29 sotolon furaneol Lactones Lactones Homofuraneol or ethyl furaneol δ - octalactone n-butanol 2-butanol 2,3 butanediol p-cresol ethanol 2 ethyl butanol n-hexanol Isobutanol 2-methyl-1-butanol 3-methyl-1-butanol 2 octanol 2,4 pentanediol 2 pentanol 2 phenyl ethanol n-propanol acetaldehyde benzaldehyde butanal decanal (E,E)-2,4-decadienal Lactones Lactones Alcohol Alcohol Alcohol Alcohol Alcohol Alcohol Alcohol Alcohol Alcohol Alcohol Alcohol Alcohol Alcohol Alcohol Alcohol Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes (E,Z)-2,4-decadienal Aldehydes Trans-4,5-epoxy-2-(E)-decenal heptanal (Z)-4-heptenal n-hexanal 2-hexenal isohexanal 2-methyl butanal 3-methyl butanal 2-methyl propanal nonanal (E)-2-nonenal (Z)-2-nonenal (E,Z)-2,6-nonadienal (E,E)-2,4-nonadienal octanal Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes pentanal propanal Phenyl aldehyde propenal Thiophen-2-aldehyde acetone acetophenone Aldehydes Aldehydes Aldehydes Aldehydes Aldehydes Ketones Ketones Curry, seasoning Sweet, caramel,burnt sugar, strawberry Caramel Fruity, peachy, sweet Floral, fruity, sweet Alcoholic Fruity Unclean, medical, cowy, barny Aalcohol Fatty, green, floral Wine Fruity, alcohol, solvent, grainy Mushroom, coconut, oil, rancid Sweet, alcoholic, fruity, nutty Rosy Pungent Sweet, pungent Almond Pungent Soapy, flowery Mayonnaise, bread, fatty, tallow, fruity Mayonnaise, bread, fatty, tallow, fruity Metallic Fatty, oily, green Creamy, biscuit Green Almond bitter, green, fatty Dark chocolate, malt Dark chocolate, malt Malt Green Green, fatty Green Melon, cucumber soapy Green, fatty, soapy, orange peel Pungent, almond like Solvent Rosy Solvent-like Almond, musty, glue fruity, 30 2-butanone 2,3 butanedione (E)- β damascenone 2- heptanone Ketones Ketones Ketones Ketones 2-hexanone 1-hexen-3-one 3-hydroxy-2-butanone (acetoin) 3-methyl-2butanone 3-methyl-2-pentanone 2-nonanone Ketones Ketones Ketones Ketones Ketones Ketones (Z)-1,5-octadien-3-one 2-octanone Ketones Ketones 1-octen-3one Pentan-2one 2-tridecanone 2-undecanone Ketones Ketones Ketones Ketones 2-butyl acetate n-butyl butyrate n-butyl acetate Ethyl acetate Ethyl propionate Ethyl butyrate Ethyl hexanoate Ethyl octanonoate Methyl acetate Methyl propionate Methyl hexanoate Propyl acetate n-propyl butyrate Geosmin Guaicol indole limonene linalool α- pinene Pyrazine, 2 acetyl Pyrazine, 2-isobutyl-3-methoxy Pyrazine, 2-isopropiyl-3methoxy Pyrroline, 2 –acetyl-1 skatole Thiazoline, 2-acetyl-2 Esters Esters Esters Esters Esters Esters Esters Esters Esters Esters Esters Esters Esters Terpene Terpene Pyrazine Pyrazine Pyrazine pyrrol (B. C. Weimer 2007; T. K Singh, Drake, and Cadwallader 2003) Etheric Buttery Apple sauce Blue cheese, fruity, soapy Fruity, ketone Cooked vegetable, Buttery Camphor musty, Green, earthy, blue cheese, fatty, musty, varnish Green metallic Floral, fruity, soapy, ketone, musty Mushroom Acetone, sweet, fruity, ketone Floral, fruity, green, musty, tallow Pear Fruity, solvent, sweet Fruity Bubble gum, fruity Fruity Fruity pineapple Pineapple, Earthy, moistened soil Smoky, spicy Mothball Citrus Sweet, floral, honey Pine Popcorn Bell pepper Earthy, soil, green, beany Roasted Unclean, mothball, fecal roasted 31 REFERENCES Alichanidis, Efstathios, Julia H. M. Wrathall, and Anthony T. Andrews. 1986. “Heat Stability of Plasmin (milk Proteinase) and Plasminogen.” Journal of Dairy Research V53 (02): pp. 259–269. Alting, A. C., W. Engels, S. van Schalkwijk, and F. A. Exterkate. 1995. “Purification and Characterization of Cystathionine (beta)-Lyase from Lactococcus Lactis Subsp. Cremoris B78 and Its Possible Role in Flavor Development in Cheese.” Applied and Environmental Microbiology V61 (11) (November 1): pp. 4037–4042. Amarita, F., T. Requena, G. Taborda, L. Amigo, and C. Pelaez. 2001. “Lactobacillus Casei and Lactobacillus Plantarum Initiate Catabolism of Methionine by Transamination.” Journal of Applied Microbiology V90 (6): pp. 971–978. Andersen, Lene T., Ylva Ardö, and Wender L.P. Bredie. 2010. “Study of Taste-active Compounds in the Water-soluble Extract of Mature Cheddar Cheese.” International Dairy Journal V20 (8) (August): pp. 528–536. Anderson, D.L., V.V. Mistry, R.L. Brandsma, and K.A. Baldwin. 1993. “Reduced Fat Cheddar Cheese from Condensed Milk. 1. Manufacture, Composition, and Ripening.” Journal of Dairy Science V76 (10) (October): pp. 2832–2844. Ardö, Y., and Frederiksberg. 1999. “Evaluating Proteolysis by Analysing the N Content of Cheese Fractions.” Bulletin - International Dairy Federation V337: pp. 4–9. Arora, Gulshan, Francois Cormier, and Byong Lee. 1995. “Analysis of Odor-Active Volatiles in Cheddar Cheese Headspace by Multidimensional GC/MS/Sniffing.” J. Agric. Food Chem. V43 (3): pp. 748–752. Aston, J. W., and L. K. Creamer. 1986. “Contribution of the components of the water-soluble fraction to the flavour of Cheddar cheese.” New Zealand Journal of Dairy Science and Technology V21 (3): pp. 229–248. Aston, J. W., and John R. Dulley. 1982. “Cheddar cheese flavor review.” Australian Journal of Dairy Technology V37 (2) (June): pp. 59–64. Avsar, Y.K., Y. Karagul-Yuceer, M.A. Drake, T.K. Singh, Y. Yoon, and K.R. Cadwallader. 2004. “Characterization of Nutty Flavor in Cheddar Cheese.” Journal of Dairy Science V87 (7) (July): pp. 1999–2010. Ayad, Eman H.E, Annette Verheul, Catrienus de Jong, Jan T.M Wouters, and Gerrit Smit. 1999. “Flavour Forming Abilities and Amino Acid Requirements of Lactococcus Lactis Strains Isolated from Artisanal and Non-dairy Origin.” International Dairy Journal V9 (10) (October): pp. 725–735. Baer, A., and I. Ryba. 1992. “Serological Identification of Propionibacteria in Milk and Cheese Samples.” International Dairy Journal V2 (5): pp. 299–310. Balcão, Victor M., and F.Xavier Malcata. 1998. “Lipase Catalyzed Modification of Milkfat.” Biotechnology Advances V16 (2) (March): pp. 309–341. Bansal, N, Paolo Piraino, and P. L. H. McSweeney. 2009. “Determination of Proteolysis in Cheese.” In Handbook of Dairy Foods Analysis, pp. 405–417. CRC Press. Bassett, Harold J., and W.L. Slatter. 1953. “The Heat Resistance of Some Lactobacilli Found in Swiss Cheese.” Journal of Dairy Science V36 (4) (April): pp. 409–413. Bastian, E.D., C.G. Lo, and K.M.M. David. 1997. “Plasminogen Activation in Cheese Milk: Influence on Swiss Cheese Ripening.” Journal of Dairy Science 80 (2) (February): 245–251. 32 Bastian, Eric D., and Rodney J. Brown. 1996. “Plasmin in Milk and Dairy Products: An Update.” International Dairy Journal V6 (5) (May): pp. 435–457. Benfeldt, Connie, and John Sørensen. 2001. “Heat Treatment of Cheese Milk: Effect on Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4–7) (July 11): pp. 567–574. Benfeldt, Connie, John Sørensen, Katrine H. Ellegård, and Torben E. Petersen. 1997. “Heat Treatment of Cheese Milk: Effect on Plasmin Activity and Proteolysis During Cheese Ripening.” International Dairy Journal 7 (11) (October 24): 723–731. Benkler, K. F., and G. A. Reineccius. 1980. “FLAVOR ISOLATION FROM FATTY FOODS VIA SOLVENT EXTRACTION AND MEMBRANE DIALYSIS.” Journal of Food Science V45 (4): pp. 1084–1085. Bentley, Ronald, and Thomas G. Chasteen. 2004. “Environmental VOSCs––formation and Degradation of Dimethyl Sulfide, Methanethiol and Related Materials.” Chemosphere V55 (3) (April): pp. 291–317. Beresford, Tom P, Nora A Fitzsimons, Noelle L Brennan, and Tim M Cogan. 2001. “Recent Advances in Cheese Microbiology.” International Dairy Journal V11 (4–7) (July 11): pp. 259–274. Beuvier, E., and S. Buchin. 2004. “Raw Milk Cheeses.” In Cheese Chemistry, Physics and Microbiology, Volume 1:319–345. Academic Press. Bills, D.D., and E.A. Day. 1964. “Determination of the Major Free Fatty Acids of Cheddar Cheese.” Journal of Dairy Science V47 (7) (July): pp. 733–738 Burbank, Helen M., and Michael C. Qian. 2005. “Volatile Sulfur Compounds in Cheddar Cheese Determined by Headspace Solid-phase Microextraction and Gas Chromatograph-pulsed Flame Photometric Detection.” Journal of Chromatography A V1066 (1–2) (February 25): pp. 149–157. Burbank, Helen, and Michael C. Qian. 2008. “Development of Volatile Sulfur Compounds in Heat-shocked and Pasteurized Milk Cheese.” International Dairy Journal V18 (8) (August): pp. 811–818. Burton, H. 1986. “Microbiological aspects.” (200): 9–14. Careri, M, and A Mangia. 2003. “Analysis of Food Proteins and Peptides by Chromatography and Mass Spectrometry.” Journal of Chromatography A V1000 (1–2) (June 6): pp. 609–635. Carunchia Whetstine, M E, P J Luck, M A Drake, E A Foegeding, P D Gerard, and D M Barbano. 2007. “Characterization of Flavor and Texture Development Within Large (291 Kg) Blocks of Cheddar Cheese.” Journal of Dairy Science V90 (7) (July): pp. 3091–3109. Carunchia Whetstine, Mary E., Keith R. Cadwallader, and MaryAnne Drake. 2005. “Characterization of Aroma Compounds Responsible for the Rosy/Floral Flavor in Cheddar Cheese.” J. Agric. Food Chem. V53 (8): pp. 3126–3132. del Castillo-Lozano, M. López, S. Mansour, R. Tâche, P. Bonnarme, and S. Landaud. 2008. “The Effect of Cysteine on Production of Volatile Sulphur Compounds by Cheeseripening Bacteria.” International Journal of Food Microbiology V 122 (3) (March 20): pp. 321–327. Chaintreau, Alain. 2001. “Simultaneous Distillation–extraction: From Birth to Maturity— review.” Flavour and Fragrance Journal V16 (2): pp. 136–148. Chatfield, Chris, and A. Collins. 1981. “Finding New Underlying Variables.” In Introduction to Multivariate Analysis, pp. 57–78. CRC Press. 33 Chavarri, Felisa, Mailo Virto, Celia Martin, Ana I. Nájera, Arantza Santisteban, Luis J. R. Barrón, and Mertxe De Renobales. 1997. “Determination of Free Fatty Acids in Cheese: Comparison of Two Analytical Methods.” Journal of Dairy Research V64 (3): pp. 445–452. Chich, Jean-François, Karine Marchesseau, and Jean-Claude Gripon. 1997. “Intracellular Esterase from Lactococcus Lactis Subsp. Lactis NCDO 763: Purification and Characterization.” International Dairy Journal V7 (2-3) (February): pp. 169–174. Chopin, Alain. 1993. “Organization and Regulation of Genes for Amino Acid Biosynthesis in Lactic Acid Bacteria.” FEMS Microbiology Reviews V12 (1–3) (September): pp. 21– 37. Christensen J.E., Johnson M.E., and Steele J.L. 1995. “Production of Cheddar Cheese Using a Lactococcus Lactis Ssp. Cremoris SK11 Derivative with Enhanced Aminopeptidase Activity.” International Dairy Journal V5 (4): pp. 367–379. Christensen, Jeffrey E., Edward G. Dudley, Jeffrey A. Pederson, and James L. Steele. 1999. “Peptidases and Amino Acid Catabolism in Lactic Acid Bacteria.” Antonie Van Leeuwenhoek V76 (1): pp. 217–246. Christensen, K.R., and G.A. Reineccius. 1992. “Gas Chromatographic Analysis of Volatile Sulfur Compounds from Heated Milk Using Static Headspace Sampling.” Journal of Dairy Science V75 (8) (August): pp. 2098–2104 Christensen, Søren, Tove Wiegers, John Hermansen, and Lars Sottrup-Jensen. 1995. “Plasmaderived Protease Inhibitors in Bovine Milk.” International Dairy Journal V5 (5): pp. 439–449. Christensen, T. M. I. E., A.-M. Bech, and H. Werner. 1991. “Methods for crude fractionation (extraction and precipitation) of nitrogen components in cheese.” Bulletin International Dairy Federation (261): pp. 4–9. Christensen, Tove M. I. E., Kristian R. Kristiansen, and Jesper S. Madsen. 1989. “Proteolysis in Cheese Investigated by High Performance Liquid Chromatography.” Journal of Dairy Research V56 (5): pp. 823–828. Cliffe, A.J., D. Revell, and B.A. Law. 1989. “A Method for the Reverse Phase High Performance Liquid Chromatography of Peptides from Cheddar Cheese.” Food Chemistry 34 (2): 147–160. Cliffe, Allan J., Jonathan D. Marks, and Francis Mulholland. 1993. “Isolation and Characterization of Non-volatile Flavours from Cheese: Peptide Profile of Flavour Fractions from Cheddar Cheese, Determined by Reverse-phase High-performance Liquid Chromatography.” International Dairy Journal V3 (4–6): pp 379–387. Collin, J. C., D. le Bars, I. Ryba, and A. Bear. 1990. “Comparative measurements of the plasminogen-plasmin system of milk.”International Dairy Federation. Collins, Y.F., P.L.H. McSweeney, and M.G. Wilkinson. 2004. “Lipolysis and Catabolism of Fatty Acids in Cheese.” In Cheese Chemistry, Physics and Microbiology, Volume 1:373–389. Academic Press. Collins, Yvonne F., Paul L.H. McSweeney, and Martin G. Wilkinson. 2003. “Lipolysis and Free Fatty Acid Catabolism in Cheese: a Review of Current Knowledge.” International Dairy Journal V13 (11): pp. 841–866. Considine, T., A. Healy, A.L. Kelly, and P.L.H. McSweeney. 1999. “Proteolytic Specificity of Elastase on Bovine Β-casein.” Food Chemistry V66 (4) (September): pp. 463–470. Considine, T., Á. Healy, A.L. Kelly, and P.L.H. McSweeney. 2000. “Proteolytic Specificity of Elastase on Bovine Αs1-casein.” Food Chemistry V69 (1) (April): pp. 19–26. 34 Considine, T., Á. Healy, A.L. Kelly, and P.L.H. McSweeney. 2004. “Hydrolysis of Bovine Caseins by Cathepsin B, a Cysteine Proteinase Indigenous to Milk.” International Dairy Journal V14 (2) (February): pp. 117–124. Crow, V.L., T. Coolbear, P.K. Gopal, F.G. Martley, L.L. McKay, and H. Riepe. 1995. “The Role of Autolysis of Lactic Acid Bacteria in the Ripening of Cheese.” International Dairy Journal V5 (8): pp. 855–875. Crow, Vaughan, Brian Curry, and Michelle Hayes. 2001. “The Ecology of Non-starter Lactic Acid Bacteria (NSLAB) and Their Use as Adjuncts in New Zealand Cheddar.” International Dairy Journal V11 (4–7) (July 11): pp 275–283. Cserháti, Tibor. 2002. “Mass Spectrometric Detection in Chromatography. Trends and Perspectives.” Biomedical Chromatography V16 (5): pp. 303–310. Curioni, P.M.G., and J.O. Bosset. 2002. “Key Odorants in Various Cheese Types as Determined by Gas Chromatography-olfactometry.” International Dairy Journal v12 (12): pp. 959–984. doi:10.1016/S0958-6946(02)00124-3. Datta, N., A. J. Elliott, M. L. Perkins, and H. C. Deeth. 2002. “Ultra-high-temperature (UHT) Treatment of Milk: Comparison of Direct and Indirect Modes of Heating” V57 (3): pp. 211–227. Deeth, H. C, and V. Touch. 2000. “Methods for detecting lipase activity in milk and milk products.” Australian Journal of Dairy Technology V55 (3): pp. 153–168. Dias, Benjamin, and Bart Weimer. 1998. “Conversion of Methionine to Thiols by Lactococci, Lactobacilli, and Brevibacteria.” Applied and Environmental Microbiology V64 (9) (September 1): pp. 3320–3326. Dimos, Tony. 1992. “A comparative study by GC/MS of the flavour volatiles produced during the maturation of full-fat and low-fat Cheddar cheese.” Dirinck, P, and A De Winne. 1999. “Flavour Characterisation and Classification of Cheeses by Gas Chromatographic–mass Spectrometric Profiling.” Journal of Chromatography A V847 (1-2) (June 25): pp. 203–208. Dobric, Nada, Gäetan K.Y. Limsowtin, Alan J. Hillier, Nicholas P.B. Dudman, and Barrie E. Davidson. 2000. “Identification and Characterization of a Cystathionine Β/γ-lyase from Lactococcus Lactis Ssp. Cremoris MG13631.” FEMS Microbiology Letters V182 (2): pp. 249–254. Drake, M.A., T.D. Boylston, K.D. Spence, and B.G. Swanson. 1996. “Chemical and Sensory Effects of a Lactobacillus Adjunct in Cheddar Cheese.” Food Research International V29 (3–4) (April): pp. 381–387. El Soda, M., S.A. Madkor, and P.S. Tong. 2000. “Adjunct Cultures: Recent Developments and Potential Significance to the Cheese Industry.” Journal of Dairy Science V83 (4) (April): pp. 609–619. El Soda, Morsi A. 1993. “The Role of Lactic Acid Bacteria in Accelerated Cheese Ripening.” FEMS Microbiology Reviews V12 (1–3) (September): pp. 239–251. El-Soda, M., S. A. Madkor, and P. S. Tong. 2000. “Evaluation of commercial adjuncts for use in cheese ripening: 4. Comparison between attenuated and not attenuated lactobacilli.” Milchwissenschaft V55 (5): pp. 260–263. Engel, Wolfgang, Wolfgang Bahr, and P. Schieberle. 1999. “Solvent Assisted Flavour Evaporation – a New and Versatile Technique for the Careful and Direct Isolation of Aroma Compounds from Complex Food Matrices.” European Food Research and Technology V209 (3): pp. 237–241. 35 Engels, W.J.M., R. Dekker, C. de Jong, R. Neeter, and S. Visser. 1997. “A Comparative Study of Volatile Compounds in the Water-soluble Fraction of Various Types of Ripened Cheese.” International Dairy Journal V7 (4) (April): pp. 255–263. Exterkate, Fred A.., Arno C.. Alting, and Charles J. Slangen. 1995. “Conversion of αs1Casein-(24-199)-Fragment and β-Casein Under Cheese Conditions by Chymosin and Starter Peptidases.” Systematic and Applied Microbiology V18 (1): pp. 7–12. Fang, Yu, and Michael C. Qian. 2005. “Sensitive Quantification of Sulfur Compounds in Wine by Headspace Solid-phase Microextraction Technique.” Journal of Chromatography A V1080 (2) (July 8): pp. 177–185. Farkye, Nana Y., and Patrick F. Fox. 1990. “Observations on Plasmin Activity in Cheese.” Journal of Dairy Research V57 (3): pp. 413–418. FDA. 2011. “Code of Federal Regulations Title 21.” http://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr=133.111. Fennema, Owen R., and Damodaran. 1996. “Amino Acids, Peptides, and Proteins.” In Food Chemistry, pp. 322–425. CRC Press. Fernández, Manuela, Tanoj K. Singh, and Patrick F. Fox. 1998. “Isolation and Identification of Peptides from the Diafiltration Permeate of the Water-Soluble Fraction of Cheddar Cheese.” Journal of Agricultural and Food Chemistry V46 (11) (November 1): pp. 4512–4517. Fernández, María, Wim van Doesburg, Ger A. M. Rutten, Joey D. Marugg, Arno C. Alting, Richard van Kranenburg, and Oscar P. Kuipers. 2000. “Molecular and Functional Analyses of themetC Gene of Lactococcus Lactis, Encoding Cystathionine β-Lyase.” Applied and Environmental Microbiology V66 (1) (January 1): pp. 42–48. Fernández-García, Estrella. 1996. “Use of Headspace Sampling in the Quantitative Analysis of Artisanal Spanish Cheese Aroma.” Journal of Agricultural and Food Chemistry V44 (7) (January 1): pp. 1833–1839. Ferreira, A.D., and B.C. Viljoen. 2003. “Yeasts as Adjunct Starters in Matured Cheddar Cheese.” International Journal of Food Microbiology V86 (1–2) (September 1): pp. 131–140. Forde, Amanda, and Gerald F Fitzgerald. 2000. “Biotechnological Approaches to the Understanding and Improvement of Mature Cheese Flavour.” Current Opinion in Biotechnology V11 (5) (October 1): pp. 484–489. Fox, P. F., T.P. Guinee, T. M. Cogan, and P. L. H. McSweeney. 2000. “Enzymatic Coagulation of Milk.” In Fundamentals of Cheese Science, pp. 98135. Springer. Fox, P. F., Law J, P. L. H. McSweeney, and Wallace J.M. 1999. “Biochemistry of Cheese Ripening.” In Cheese: Chemistry, Physics and Microbiology, pp. 382–450. Springer. Fox, P. F., J. A. Lucey, and T. M. Cogan. 1990. “Glycolysis and Related Reactions During Cheese Manufacture and Ripening.” Critical Reviews in Food Science and Nutrition V29 (4) (January): pp. 237–253. Fox, P. F., and P. L. H. McSweeney. 1996. “Proteolysis in Cheese During Ripening.” Food Reviews International V12 (4): pp. 457–509. Fox, P. F., and P. L. H. McSweeney. 1998. “Milk Lipids.” In Dairy Chemistry and Biochemistry, pp. 67–141. Springer. Fox, P. F., J. M. Wallace, S. Morgan, C. M. Lynch, E. J. Niland, and J. Tobin. 1996. “Acceleration of Cheese Ripening.” Antonie Van Leeuwenhoek 70 (2): 271–297. Fox, P.F. 1989. “Proteolysis During Cheese Manufacture and Ripening.” Journal of Dairy Science V72 (6) (June): pp. 1379–1400. 36 Fox, P.F., and P.L.H. McSweeney. 2004. “Cheese: An Overview.” In Cheese Chemistry, Physics and Microbiology, Volume 1:1–18. Academic Press. Fox, P.F., and L. Stepaniak. 1993. “Enzymes in Cheese Technology.” International Dairy Journal V3 (4–6): pp. 509–530. Fox, PF, PLH McSweeney, and CM Lynch. 1998. “Significance of Non-starter Lactic Acid Bacteria in Cheddar Cheese.” Australian Journal of Dairy Technology V53 (2): pp. 7– 8. Gao, Song, Ed S. Mooberry, and James L. Steele. 1998. “Use of 13C Nuclear Magnetic Resonance and Gas Chromatography To Examine Methionine Catabolism by Lactococci.” Applied and Environmental Microbiology V64 (12) (December 1): pp. 4670–4675. Gomez, Maria J., Sonia Garde, Pilar Gaya, Margarita Medina, and Manuel Nuñez. 1997. “Relationship Between Level of Hydrophobic Peptides and Bitterness in Cheese Made from Pasteurized and Raw Milk.” Journal of Dairy Research V64 (2): pp. 289–297. Grandison, Alistair S., and Graeme D. Ford. 1986. “Effects of Variations in Somatic Cell Count on the Rennet Coagulation Properties of Milk and on the Yield, Composition and Quality of Cheddar Cheese.” Journal of Dairy Research V53 (4): pp. 645–655. Grappin, Rémy, and Eric Beuvier. 1997. “Possible Implications of Milk Pasteurization on the Manufacture and Sensory Quality of Ripened Cheese.” International Dairy Journal V7 (12) (November 21): pp. 751–761. Green, Margaret L., and Donald J. Manning. 1982. “Development of Texture and Flavour in Cheese and Other Fermented Products.” Journal of Dairy Research V49 (4): pp. 737– 748. Grieve, Paul A., and Barry J. Kitchen. 1985. “Proteolysis in Milk: The Significance of Proteinases Originating from Milk Leucocytes and a Comparison of the Action of Leucocyte, Bacterial and Natural Milk Proteinases on Casein.” Journal of Dairy Research 52 (1): 101–112. Griffiths, M. W. 1986. “Use of milk enzymes as indices of heat treatment.” Journal of Food Protection V49 (9): pp. 696–705. Gripon, Jean-Claude, V Monnet, G Lambert, and M. J. Desmazeaud. 1991. “Microbial enzymes in cheese ripening.” In Food enzymology, vol 1. Guinee, T P. 2004. “Salting and the Role of Salt in Cheese.” International Journal of Dairy Technology V57 (2-3): pp. 99–109. Habibi‐Najafi, Mohammad B., Byong H. Lee, and Barry Law. 1996. “Bitterness in Cheese: A Review.” Critical Reviews in Food Science and Nutrition V36 (5): pp. 397–411. Heegaard, Christian W., Lone K. Rasmussen, and Peter A. Andreasen. 1994. “The Plasminogen Activation System in Bovine Milk: Differential Localization of Tissuetype Plasminogen Activator and Urokinase in Milk Fractions Is Caused by Binding to Casein and Urokinase Receptor.” Biochimica Et Biophysica Acta (BBA) - Molecular Cell Research V1222 (1) (May 26): pp. 45–55. Hickey, D.K., K.N. Kilcawley, T.P. Beresford, and M.G. Wilkinson. 2007. “Lipolysis in Cheddar Cheese Made from Raw, Thermized, and Pasteurized Milks.” Journal of Dairy Science V90 (1) (January): pp. 47–56. Holland, Ross, and Tim Coolbear. 1996. “Purification of Tributyrin Esterase from Lactococcus Lactis Subsp. Cremoris E8.” Journal of Dairy Research V63 (1): pp. 131–140. Home, D.S., and J.M. Banks. 2004. “Rennet-induced Coagulation of Milk.” In Cheese Chemistry, Physics and Microbiology, Volume 1:pp. 47–70. Academic Press. 37 Huffman, L. M.;, and Kristoffersen. 1984. “Role of lactose in Cheddar cheese manufacturing and ripening.” New Zealand Journal of Dairy Science and Technology V19 (2): pp. 151–162. Hurley, M.J, B.M O’Driscoll, A.L Kelly, and P.L.H McSweeney. 1999. “Novel Assay for the Determination of Residual Coagulant Activity in Cheese.” International Dairy Journal V9 (8) (August): pp 553–558. Hutton, J.T., and S. Patton. 1952. “The Origin of Sulfhydryl Groups in Milk Proteins and Their Contributions to ‘Cooked’ Flavor.” Journal of Dairy Science V35 (8) (August): pp. 699–705. Jolly, Ramesh C., and Frank V. Kosikowski. 1975. “Quantification of Lactones in Ripening Pasteurized Milk Blue Cheese Containing Added Microbial Lipases.” J. Agric. Food Chem. V23 (6): pp. 1175–1176. De Jong, Catrienus, and Herman T. Badings. 1990. “Determination of Free Fatty Acids in Milk and Cheese Procedures for Extraction, Clean up, and Capillary Gas Chromatographic Analysis.” Journal of High Resolution Chromatography V13 (2): pp. 94–98. K. Arfi, H.Spinnler, R. Tache, and P. bonnarme. 2002. “Production of Volatile Compounds by Cheese-ripening Yeasts: Requirement for a Methanethiol Donor for S-methyl Thioacetate Synthesis by Kluyveromyces Lactis.” Applied Microbiology and Biotechnology V58 (4): pp. 503–510. Knecht, E. 1999. “Lysomomal Cysteine Proteinases (2nd Edition).” Cell Biology International V23 (8) (8) (August): pp 590–591. Laan, Harry, Saw Eng Tan, Paul Bruinenberg, Gaëtan Limsowtin, and Malcolm Broome. 1998. “Aminopeptidase Activities of Starter and Non-Starter Lactic Acid Bacteria Under Simulated Cheddar Cheese Ripening Conditions.” International Dairy Journal V8 (4) (April): pp. 267–274. Lane, C.N., and P.F. Fox. 1996 “Contribution of Starter and Adjunct Lactobacilli to Proteolysis in Cheddar Cheese During Ripening.” International Dairy Journal V6 (7) (July): pp. 715–728. Lane, Cáit N., and Patrick F. Fox. 1997. “Role of Starter Enzymes During Ripening of Cheddar Cheese Made from Pasteurized Milk Under Controlled Microbiological Conditions.” International Dairy Journal V7 (1) (January): pp. 55–63. Larsen, Lotte B., Connie Benfeldt, Lone K. Rasmussen, and Torben E. Petersen. 1996. “Bovine Milk Procathepsin D and Cathepsin D: Coagulation and Milk Protein Degradation.” Journal of Dairy Research V63 (1): pp 119–130. Lau, Kum Y., David M. Barbano, and Robert R. Rasmussen. 1990. “Influence of Pasteurization on Fat and Nitrogen Recoveries and Cheddar Cheese Yield.” Journal of Dairy Science V73 (3) (March): pp. 561–570. doi:10.3168/jds.S0022-0302(90)787036. Lau, Kum Y., David M. Barbano, and Robert R. Rasmussen. 1991. “Influence of Pasteurization of Milk on Protein Breakdown in Cheddar Cheese During Aging.” Journal of Dairy Science V74 (3) (March): pp. 727–740. Law, A. J. R., J. M. Banks, D. S. Horne, J. Leaver, and I. G. West. 1994. “Denaturation of the whey proteins in heated milk and their incorporation into Cheddar cheese.” Milchwissenschaft V49 (2): pp. 63–67. Lee, Won-Jae, Dattatreya S. Banavara, Joanne E. Hughes, Jason K. Christiansen, James L. Steele, Jeffery R. Broadbent, and Scott A. Rankin. 2007. “Role of Cystathionine βLyase in Catabolism of Amino Acids to Sulfur Volatiles by Genetic Variants of 38 Lactobacillus Helveticus CNRZ 32.” Applied and Environmental Microbiology V73 (9) (May 1): pp. 3034–3039. Lin, J. C. C., and I. J. Jeon. 1985. “Headspace Gas Sampling/GC Method for the Quantitative Analysis of Volatile Compounds in Cheese.” Journal of Food Science V50 (3): pp. 843–844. Liu, Shao-Quan, Ross Holland, and Vaughan L Crow. 2001. “Purification and Properties of Intracellular Esterases from Streptococcus Thermophilus.” International Dairy Journal V11 (1–2) (January): pp. 27–35. Lloyd, G. T., J. F. Horwood, and I. Barlow. 1980. “The effect of yoghurt culture YB on the flavour and maturation of Cheddar cheese.” Australian Journal of Dairy Technology V35 (4): pp. 137–139. Lynch, C.M., P.L.H. McSweeney, P.F. Fox, T.M. Cogan, and F.D. Drinan. 1996. “Manufacture of Cheddar Cheese with and Without Adjunct Lactobacilli Under Controlled Microbiological Conditions.” International Dairy Journal V6 (8–9) (August): pp. 851–867. Lynch, C.M., D.D. Muir, J.M. Banks, P.L.H. McSweeney, and P.F. Fox. 1999. “Influence of Adjunct Cultures of Lactobacillus Paracasei Ssp. Paracasei or Lactobacillus Plantarum on Cheddar Cheese Ripening.” Journal of Dairy Science V82 (8) (August): pp. 1618– 1628. Lynch, Joanna M., David M. Barbano, and J. Richard Fleming. 2002. “Determination of the Total Nitrogen Content of Hard, Semihard, and Processed Cheese by the Kjeldahl Method: Collaborative Study.” Journal of AOAC International V85 (2): pp. 445–455. Madkor, S.A., P.S. Tong, and M. El Soda. 2000. “Ripening of Cheddar Cheese with Added Attenuated Adjunct Cultures of Lactobacilli.” Journal of Dairy Science V83 (8) (August): pp. 1684–1691. Manning, D. J., Helen R. Chapman, and Zena D. Hosking. 1976. “The Production of Sulphur Compounds in Cheddar Cheese and Their Significance in Flavour Development.” Journal of Dairy Research V43 (2): pp. 313–320. Manning, Donald J., and Carolyn Moore. 1979. “Headspace Analysis of Hard Cheeses.” Journal of Dairy Research V46 (3): pp. 539–545. McSweeney, P.L.H., P.F. Fox, J.A. Lucey, K.N. Jordan, and T.M. Cogan. 1993. “Contribution of the Indigenous Microflora to the Maturation of Cheddar Cheese.” International Dairy Journal V3 (7): pp. 613–634. McSweeney, P.L.H., P.F. Fox, and N.F. Olson. 1995. “Proteolysis of Bovine Caseins by Cathepsin D: Preliminary Observations and Comparison with Chymosin.” International Dairy Journal V5 (4): pp. 321–336. McSweeney, Paul L. H., Sylvie Pochet, Patrick F. Fox, and Aine Healy. 1994. “Partial Identification of Peptides from the Water-Insoluble Fraction of Cheddar Cheese.” Journal of Dairy Research V61 (4): pp. 587–590. McSweeney, Paul L.H., and Maria José Sousa. 2000. “Biochemical Pathways for the Production of Flavour Compounds in Cheeses During Ripening: A Review.” Le Lait V80 (3): pp. 293–324. Metwalli, Ali A.M, Harmen H.J de Jongh, and Martinus A.J.S van Boekel. 1998. “Heat Inactivation of Bovine Plasmin.” International Dairy Journal V8 (1) (January 1): pp. 47–56. Milo, C., and G. A. Reineccius. 1997. “Identification and Quantification of Potent Odorants in Regular-Fat and Low-Fat Mild Cheddar Cheese.” Journal of Agricultural and Food Chemistry V45 (9) (September 1): pp. 3590–3594. 39 Mistry, V.V., and J.-L. Maubois. 2004. “Application of Membrane Separation Technology to Cheese Production.” In Cheese Chemistry, Physics and Microbiology, Volume 1:pp. 261–VII. Academic Mistry, V.V., L.E. Metzger, and J.L. Maubois. 1996. “Use of Ultrafiltered Sweet Buttermilk in the Manufacture of Reduced Fat Cheddar Cheese.” Journal of Dairy Science V79 (7) (July): pp. 1137–1145 Moio, Luigi, Dominique Langlois, Patrick Etievant, and Francesco Addeo. 1993. “Powerful Odorants in Bovine, Ovine, Caprine and Water Buffalo Milk Determined by Means of Gas Chromatography–olfactometry.” Journal of Dairy Research V60 (02): pp. 215– 222. Molimard, P., and H.E. Spinnler. 1996 “Review: Compounds Involved in the Flavor of Surface Mold-Ripened Cheeses: Origins and Properties.” Journal of Dairy Science V79 (2) (February): pp. 169–184. Mooney, J.S, P.F Fox, A Healy, and J Leaver. 1998. “Identification of the Principal Waterinsoluble Peptides in Cheddar Cheese.” International Dairy Journal V8 (9) (September): pp. 813–818. Muir, D.D., J.M. Banks, and E.A. Hunter. 1996. “Sensory Properties of Cheddar Cheese: Effect of Starter Type and Adjunct.” International Dairy Journal V6 (4) (April): pp. 407–423. Mulvihill, Donald M., and Patrick F. Fox. 1979. “Proteolytic Specificity of Chymosin on Bovine Αs1-Casein.” Journal of Dairy Research V46 (4): pp. 641–651. Naughton, M. A., and F. Sanger. 1961. “Purification and Specificity of Pancreatic Elastase.” Biochemical Journal V78 (1): pp. 156–163. Niven, C. F., L. G. Buettner, and James B. Evans. 1954. “Thermal Tolerance Studies on the Heterofermentative Lactobacilli That Cause Greening of Cured Meat Products.” Applied Microbiology V2 (1) (January): pp. 26–29. O’Keeffe, Arthur M., Patrick F. Fox, and Charles Daly. 1978. “Proteolysis in Cheddar Cheese: Role of Coagulant and Starter Bacteria.” Journal of Dairy Research V45 (3): pp. 465– 477. O’Keeffe, R. B., P. F. Fox, and C. Daly. 1976. “Contribution of Rennet and Starter Proteases to Proteolysis in Cheddar Cheese.” Journal of Dairy Research 43 (01): 97–107. O’Shea, B.A., T. Uniacke-Lowe, and P.F. Fox. 1996. “Objective Assessment of Cheddar Cheese Quality.” International Dairy Journal V6 (11-12) (November): pp. 1135– 1147. Olivecrona, T., S. Vilaró, G. Bengtsson-Olivecrona, and P. F Fox. 1992. “Indigenous enzyme in milk. II. Lipases in milk.” In Advanced Dairy Chemistry 1, 292–310. Payne, Martin J., and Rohan G. Kroll. 1991. “Methods for the Separation and Concentration of Bacteria from Foods.” Trends in Food Science & Technology V2 (0): pp. 315–319. Peterson, S.D., and R.T. Marshall. 1990. “Nonstarter Lactobacilli in Cheddar Cheese: A Review.” Journal of Dairy Science V73 (6) (June): pp. 1395–1410. Piraino, Paolo, Eugenio Parente, and Paul L. H. McSweeney. 2004. “Processing of Chromatographic Data for Chemometric Analysis of Peptide Profiles from Cheese Extracts: A Novel Approach.” Journal of Agricultural and Food Chemistry 52 (23) (November 1): 6904–6911. Precetti, A.S., M.P. Oria, and S.S. Nielsen. 1997. “Presence in Bovine Milk of Two Protease Inhibitors of the Plasmin System.” Journal of Dairy Science V80 (8) (August): pp. 1490–1496. 40 Preininger, Martin, and Werner Grosch. 1994. “Evaluation of Key Odorants of the Neutral Volatiles of Emmentaler Cheese by the Calculation of Odour Activity Values.” LWT Food Science and Technology V27 (3) (June): pp. 237–244. Pripp, A.H, Shakeel-Ur-Rehman, P.L.H McSweeney, and P.F Fox. 1999. “Multivariate Statistical Analysis of Peptide Profiles and Free Amino Acids to Evaluate Effects of Single-strain Starters on Proteolysis in Miniature Cheddar-type Cheeses.” International Dairy Journal V9 (7) (July): pp. 473–479. Pripp, Are Hugo, Leszek Stepaniak, and Terje Sørhaug. 2000. “Chemometrical Analysis of Proteolytic Profiles During Cheese Ripening.” International Dairy Journal V10 (4): pp. 249–253. Puchades, R., L. Lemieux, and R.e. Simard. 1989. “Evolution of Free Amino Acids During the Ripening of Cheddar Cheese Containing Added Lactobacilli Strains.” Journal of Food Science V54 (4): pp. 885–888. Qian, M., and G. Reineccius. 2002. “Identification of Aroma Compounds in ParmigianoReggiano Cheese by Gas Chromatography/Olfactometry.” Journal of Dairy Science V85 (6) (June): pp. 1362–1369. Rehman, Shakeel-Ur, J.M Banks, E.Y Brechany, D.D Muir, P.L.H McSweeney, and P.F Fox. 2000. “Influence of Ripening Temperature on the Volatiles Profile and Flavour of Cheddar Cheese Made from Raw or Pasteurised Milk.” International Dairy Journal V10 (1–2) (January): pp. 55–65. Richardson, B. C., L. K. Creamer, K. N. Pearce, and R. E. Munford. 1974. “Comparative Micelle Structure: II. Structure and Composition of Casein Micelles in Ovine and Caprine Milk as Compared with Those in Bovine Milk.” Journal of Dairy Research V41 (2): pp. 239–247. Roy, Denis, Isabelle Mainville, and Francine Mondou. 1997. “Selective Enumeration and Survival of Bifidobacteria in Fresh Cheese.” International Dairy Journal V7 (12) (November 21): pp. 785–793. Sablé, Sophie, and Gilles Cottenceau. 1999. “Current Knowledge of Soft Cheeses Flavor and Related Compounds.” J. Agric. Food Chem. V47 (12): pp. 4825–4836. Seefeldt, K.E., and B.C. Weimer. 2000. “Diversity of Sulfur Compound Production in Lactic Acid Bacteria.” Journal of Dairy Science V83 (12) (December): pp. 2740–2746. Shakeel-Ur-Rehman, J. M Banks, P. L. H Mcsweeney, and P. F Fox. 2000. “Effect of ripening temperature on the growth and significance of non-starter lactic acid bacteria in Cheddar cheese made from raw or pasteurised milk.” International dairy journal V10 (1-2): pp. 45–53. Singh, T. K, M. A Drake, and K. R Cadwallader. 2003. “Flavor of Cheddar Cheese: A Chemical and Sensory Perspective.” Comprehensive Reviews in Food Science and Food Safety V2 (4) (October 1): pp. 166–189. Singh, T.K., P.F. Fox, P. Højrup, and A. Healy. 1994. “A Scheme for the Fractionation of Cheese Nitrogen and Identification of Principal Peptides.” International Dairy Journal V4 (2): pp. 111–122. Singh, Tanoj K., Patrick F. Fox, and Áine Healy. 1995. “Water-Soluble Peptides in Cheddar Cheese: Isolation and Identification of Peptides in the Diafiltration Retentate of the Water-Soluble Fraction.” Journal of Dairy Research V62 (4): pp. 629–640. Singh, Tanoj K., Patrick F. Fox, and Áine Healy 1997. “Isolation and Identification of Further Peptides in the Diafiltration Retentate of the Water-soluble Fraction of Cheddar Cheese.” Journal of Dairy Research V64 (03): pp. 433–443. 41 Smacchi, Emanuele, and Marco Gobbetti. 1998. “Purification and Characterization of Cystathionine Γ-lyase from Lactobacillus Fermentum DT41.” FEMS Microbiology Letters V166 (2): pp. 197–202. Smit, Gerrit, Bart A. Smit, and Wim J.M. Engels. 2005. “Flavour Formation by Lactic Acid Bacteria and Biochemical Flavour Profiling of Cheese Products.” FEMS Microbiology Reviews V29 (3): pp. 591–610. Smith, A. M., and S. Nakai. 1990. “Classification of cheese varieties by multivariate analysis of HPLC profiles.” Canadian Institute of Food Science and Technology Journal V23 (1): pp. 53–58. Sousa, M.J, Y Ardö, and P.L.H McSweeney. 2001. “Advances in the Study of Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4–7) (July 11): pp. 327– 345. Stuart, Mark R., Lan Szu Chou, and Bart C. Weimer. 1999. “Influence of Carbohydrate Starvation and Arginine on Culturability and Amino Acid Utilization of Lactococcus Lactis Subsp. Lactis.” Applied and Environmental Microbiology V65 (2) (February 1): pp. 665–673. Subramanian, A., W.J. Harper, and L.E. Rodriguez-Saona. 2009. “Cheddar Cheese Classification Based on Flavor Quality Using a Novel Extraction Method and Fourier Transform Infrared Spectroscopy.” Journal of Dairy Science V92 (1) (January): pp. 87–94. Suriyaphan, Orasa, MaryAnne Drake, X. Q. Chen, and Keith R. Cadwallader. 2001. “Characteristic Aroma Components of British Farmhouse Cheddar Cheese.” Journal of Agricultural and Food Chemistry V49 (3) (March 1): pp. 1382–1387. Tammam, J. D, A. G Williams, J. Noble, D. Lloyd, J. D Tammam, A. G Williams, J. Noble, and D. Lloyd. 2000. “Amino acid fermentation in non‐starter Lactobacillus spp. isolated from Cheddar cheese, Amino acid fermentation in non‐starter Lactobacillus spp. isolated from Cheddar cheese.” Letters in Applied Microbiology, Letters in Applied Microbiology V30 (5) (May 1): pp. 370–374. Tanaka, Hidehiko, Nobuyoshi Esaki, and Kenji Soda. 1985. “A Versatile Bacterial Enzyme: L-methionine Γ-lyase.” Enzyme and Microbial Technology V7 (11) (November): pp. 530–537. Thomsen, D., and H. Stapelfeldt. 1990. “Yield and quality of Danbo 45+ cheese from milk heat treated at temperatures between 72 and 95°C.” (44): 39. Tsakalidou, E., and G. Kalantzopoulos. 1992. “Purification and Partial Characterization of an Esterase from Lactococcus Lactis Ssp Lactis Strain ACA-DC 127.” Le Lait V72 (6): pp. 533–543. Turner, K. W, and T. D Thomas. 1980. “Lactose fermentation in Cheddar cheese and the effect of salt.” New Zealand Journal of Dairy Science and Technology V15 (3): pp. 265–276. Upadhyay, V.K., P.L.H. McSweeney, A.A.A. Magboul, and P.F. Fox. 2004. “Proteolysis in Cheese During Ripening.” In Cheese Chemistry, Physics and Microbiology, Volume 1:pp. 391–433. Urbach, G. 1995. “Contribution of Lactic Acid Bacteria to Flavour Compound Formation in Dairy Products.” International Dairy Journal V5 (8): pp. 877–903. Urbach, Gerda. 1993. “Relations Between Cheese Flavour and Chemical Composition.” International Dairy Journal V3 (4–6): pp. 389–422. USDA. 2011. Dairy Products Annual Sumary. 42 Vazquez-Landaverde, P.A., J.A. Torres, and M.C. Qian. 2006. “Quantification of Trace Volatile Sulfur Compounds in Milk by Solid-Phase Microextraction and Gas Chromatography–Pulsed Flame Photometric Detection.” Journal of Dairy Science V89 (8) (August): pp. 2919–2927. Verdi, Robert J., and David M. Barbano. 1991. “Properties of Proteases from Milk Somatic Cells and Blood Leukocytes.” Journal of Dairy Science V74 (7) (July): pp. 2077– 2081. Visser, S. ;Hup. 1983. “Bitter flavour in cheese, 2: model studies on the formation and degradation of bitter peptides by proteolytic enzymes from calf rennet, starter cells and starter cell fractions.” Netherlands Milk and Dairy Journal V37 (3): pp. 169–180. Visser, Servaas. 1993. “Proteolytic Enzymes and Their Relation to Cheese Ripening and Flavor: An Overview.” Journal of Dairy Science V76 (1) (January): pp. 329–350. Voigt, Daniela D., François Chevalier, John A. Donaghy, Margaret F. Patterson, Michael C. Qian, and Alan L. Kelly. 2012. “Effect of High-pressure Treatment of Milk for Cheese Manufacture on Proteolysis, Lipolysis, Texture and Functionality of Cheddar Cheese During Ripening.” Innovative Food Science & Emerging Technologies V13 (0) (January): pp. 23–30. Wallace J.M., and Fox P.F. 1997. “Effect of Adding Free Amino Acids to Cheddar Cheese Curd on Proteolysis, Flavour and Texture Development.” International Dairy Journal V7 (2): pp. 157–167. Walstra, P., T. J Geurts, A. Noomen, A. Jellema, and M. A. J. S. van Boekel. 1999. “Cheese Varieties.” In Dairy technology: principles of milk properties and processes, pp 651– 708. Walstra, Pieter, Jan T. M. Wouters, and Tom J. Geurts. 2006. Dairy Science And Technology. CRC Press. Weimer, B. C. 2007. Improving the flavour of cheese. CRC Press. Weimer, Bart, Kimberly Seefeldt, and Benjamin Dias. 1999. “Sulfur Metabolism in Bacteria Associated with Cheese.” Antonie Van Leeuwenhoek 76 (1): 247–261. Wijesundera, C., and L. Drury. “Role of milk fat in production of cheddar cheese flavour using a fat-substituted cheese model.” Australian journal of dairy technology V54 (1): pp. 28–35. Wilkinson, Martin G., Timothy P. Guinee, Daniel M. O’Callaghan, and Patrick F. Fox. 1994. “Autolysis and Proteolysis in Different Strains of Starter Bacteria During Cheddar Cheese Ripening.” Journal of Dairy Research V61 (2): pp. 249–262. De Wit, M., G. Osthoff, B.C. Viljoen, and A. Hugo. 2005. “A Comparative Study of Lipolysis and Proteolysis in Cheddar Cheese and Yeast-inoculated Cheddar Cheeses During Ripening.” Enzyme and Microbial Technology V37 (6) (November 1): pp. 606–616. de Wit, Rudy, and Hans Nieuwenhuijse. 2008. “Kinetic Modelling of the Formation of Sulphur-containing Flavour Components During Heat-treatment of Milk.” International Dairy Journal V18 (5) (May): pp. 539–547. Wong, N. P, R. Ellis, and D. E LaCroix. “Quantitative determination of lactones in Cheddar cheese.” Journal of Dairy Science V58 (10): pp. 1437–1441. Yvon, Mireille, Claire Chabanet, and Jean-Pierre Pélissier. 1989. “Solubility of Peptides in Trichloroacetic Acid (TCA) Solutions Hypothesis on the Precipitation Mechanism.” International Journal of Peptide and Protein Research V34 (3): pp. 166–176. Yvon, Mireille, and Liesbeth Rijnen. 2001. “Cheese Flavour Formation by Amino Acid Catabolism.” International Dairy Journal V11 (4–7) (July 11): pp. 185–201. 43 Zehentbauer, Gerhard, and G. A. Reineccius. 2002. “Determination of Key Aroma Components of Cheddar Cheese Using Dynamic Headspace Dilution Assay.” Flavour and Fragrance Journal V17 (4): pp. 300–305 44 CHAPTER 2 EVALUATION OF THE EFFECT OF HEAT-SHOCK AND PASTEURIZATION TREATMENTS OF CHEESE MILK ON RIPENING RELATED CHANGES IN CHEDDAR CHEESE Lemus Muñoz, Freddy Mauricio, Qian Michael Department of Food Science and Technology, Oregon State University Corvallis Oregon 97331 45 ABSTRACT The effect of heat shock (66C for 30 sec) and pasteurization (72C for 15C) treatments of cheese milk was studied during the ripening stage of Cheddar cheese. Proteolysis was investigated by a fractionation scheme, resulting in an insoluble fraction analyzed by urea polyacrylamide gel electrophoresis (Urea-PAGE), and a soluble fraction which was further investigated through water soluble nitrogen (WSN), trichloroacetic acid soluble nitrogen (TCA-SN) and phosphotungstic acid soluble nitrogen (PTA-SN) analyzed by total Kjeldahl nitrogen content (TKN). Reversed phase high performance liquid chromatography (RPHPLC) was used to study the peptide profile of the water soluble fraction. Lipolysis was studied by levels of individual free fatty acids determined through gas chromatography-flame ionization detection (GC-FID) after isolation employing solid phase extraction (SPE). Volatile sulfur compounds were studied using head space solid phase micro-extraction (SPME) coupled with gas chromatography-pulsed flame photometric detection (PFPD). The Urea-PAGE method was able to differentiate samples according their age, but it could not discriminate samples regarding their treatment. Nonetheless, measurements of total Kjeldahl Nitrogen (TKN) of the WSN, TCA-SN, and PTA-SN fractions, and the principal component analysis of the RP-HPLC peptide profile of the WSN fraction, revealed differences in the rate and pattern of proteolysis. Levels of total nitrogen for the WSN, TCA-SN and PTA-SN fractions increased as cheese aged and were lower for samples made from pasteurized milk, indicating that primary and secondary proteolysis were faster for samples made with heatshocked milk. It was obtained a PCA model with 3 principal components that accounted for the 82.6% of the variability from data collected. This model discriminate the samples according age and quality, suggesting the samples undergo more or faster proteolysis. FFA profiles reveal minor but not significant difference in the extension of the inactivation of the lipoprotein lipase and its role during ripening, which is related to a higher lipolytic activity for heat-shocked samples. The Volatile Sulfur Compounds (VSC) analysis showed that cheeses made from heat-shocked milk developed higher concentrations of H2S, DMS and MeSH, suggesting slower catabolism of sulfur containing amino acids in cheese made with pasteurized milk. 46 INTRODUCTION Although cheeses made from raw milk develop stronger flavors and ripen faster (P. F. Fox et al. 1999), because of the commercial interest in maintaining a microbiological safe product during longer periods of time in the market, thermal treatments of cheese milk have become the first unit operation that determines the characteristic flavor of commercial cheddar cheese during the cheese making process. However, differences in manufacturing procedures, such as type of heat treatment applied to the cheese milk, result in different flavor profiles, corresponding to its evident influence on the development of attributes. Whereby, it is of industrial interest to find a well adjusted way to reduce microbial number without significant effects on organoleptic characteristics and/or nutritional components, while emerging technologies can be totally implemented. Comparisons between the difference in the development of attributes of cheddar cheese made from raw milk and pasteurized milk have been rigorously studied (Lau, Barbano, and Rasmussen 1990; Lau, Barbano, and Rasmussen 1991; Grappin and Beuvier 1997; Hickey et al. 2007; Cáit N. Lane and Fox 1997). However, complementary analysis of temperature effects on cheese milk during ripening are still required to fulfill the gap of knowledge in differences of standard heat treatments. Thus, its is worthy to focus again in common approaches such as heat-shock (66 °C for 30 sec) and pasteurization (72 °C for 15 sec), which are slightly different. Furthermore, it is key to maintain the scope in low processing temperatures, based on that previous studies show that defects in body and flavor, and a reduced proteolytic activity arise from severe heat treatments due to inactivation of Plasmin by thiol-disulphide reactions with denatured-lactoglobulin and formation of complex between caseins and b-lactoglobulin, that result in the interference of whey protein in the maturation of cheese (Thomsen and Stapelfeldt 1990; Law et al. 1994) Essentially, heat affects the indigenous enzyme activity and thermo-labile non-starter lactic acid bacteria (NSLAB) of milk. On the one hand, the enzyme lipoprotein lipase (LPL) and its activator (C-II apolipoprotein-glutamic acid) are practically inactivated due to their sensitivity to temperature. Indeed, the 73-95% of LPL activity is lost after pasteurization of milk. On the other hand, because of Plasmin and its pro-enzyme Plasminogen depend on a thermo-labile heterogeneous system of inhibitors (Heegaard, Rasmussen, and Andreasen 1994; Metwalli, de 47 Jongh, and van Boekel 1998; E.D. Bastian, Lo, and David 1997; Precetti, Oria, and Nielsen 1997), heat treatment of cheese milk leads to higher plasmin acitvity during the ripening stage (Grappin and Beuvier 1997). In addition, heat induces the destruction of some beneficial NSLAB (Lau, Barbano, and Rasmussen 1991; Rehman et al. 2000), or results in a significant reduction of NSLAB species depending on the severity of the treatment (P.L.H. McSweeney et al. 1993; Roy, Mainville, and Mondou 1997; Beuvier and Buchin 2004), which has a direct impact on the texture and quality of the product as consequence of the lactose metabolism, in which a reduced racemization of L(+)-lactate and formation of calcium lactate crystals lactate take place. In the case of cheddar cheese and other hard and semi-hard varieties, standard heat treatments of cheese milk do not influence secondary proteolysis, which is mainly depend on proteinases and peptidases from LAB and other adjunct cultures, but affect primary proteolysis where the main enzymatic activity is provided by the rennet and the indigenous milk proteinase plasmin (P. F. Fox 1989; S. Visser 1993; P.L.H. McSweeney et al. 1993). However, rennet enzymatic activity is not directly affected by heat treatments of cheese milk, but it is reported that the accessibility to αs1 caseins is altered. On the contrary, proteolytic activity of plasmin is affected directly by temperature (P. F. Fox 1989; Benfeldt et al. 1997). Previous studies showed that cheese manufactured from pasteurized milk displayed higher proteolytic digestion of β-casein and an analogous increase in the amount of γ-casein compared to cheeses manufactured from raw milk (Grappin and Beuvier 1997). However, it has been reported as well that in spite of the stability of Plasmin at elevated temperatures, its activity decrease as the temperature and the holding time increased (Benfeldt et al. 1997; Benfeldt and Sørensen 2001). Regarding the degradation of milk fat, it has been documented that the level of lipolysis in cheese made from pasteurized milk was lower than that attained in cheese made from raw milk (P.L.H. McSweeney et al. 1993; Shakeel-Ur-Rehman et al. 2000), which can be explained by the inactivation of the indigenous enzyme LPL and differences in growth rates of NSLAB (P.L.H. McSweeney et al. 1993; Roy, Mainville, and Mondou 1997; Beuvier and Buchin 2004), resulting in final product with lower levels of individual free fatty acids and 48 other compounds like methyl ketones and lactones in comparison to those found in cheese made from raw milk (Olivecrona et al. 1992). Thus, in order to understand the influence of heat treatments of cheese milk on the biochemical changes of cheddar cheese during ripening; Calculating levels of individual FFA (Qian and Reineccius 2002), monitoring the development of volatile sulfur compounds (H. M. Burbank and Qian 2005; H. Burbank and Qian 2008), and tracking proteolysis through the nitrogen content of certain fractions (J. M. Lynch, Barbano, and Fleming 2002), and the analysis of peptide profiles by chromatographic and electrophoretic techniques (Dirinck and De Winne 1999; Benfeldt and Sørensen 2001; Sousa, Ardö, and McSweeney 2001) are important and appropriate approaches to be used. The objective of this study was to track the proteolysis and lipolysis during the ripening of cheddar cheese samples to investigate agerelated changes resulting from different heat treatment practices of cheese milk. MATERIALS AND METHODS CHEESE SAMPLES Cheeses were manufactured in a local dairy plant, using pasteurized milk and heat – shocked milk. For the pasteurization treatment, milk was heated at 72 C for 16 seconds and cheese was made according with standard protocols. Milk for the heat-shocked treatment was heated at 60 C for 30 seconds and the cheese was made according with standard protocols. Three blocks of cheese made from each milk treatment were selected randomly from three consecutive manufacturing days. All cheeses were aged using the same conditions at manufacturer’s facility, Every month a 2 lb portion was sampled from each block and sent to the lab, where samples are stored at (-37C) to stop ageing process until analysis is completed. FREE FATTY ACIDS ANALYSIS Chemicals Pentanoic acid, heptanoic acid, nonanoic acid, undecanoic acid, and heptadecanoic acid were used as internal standards, they were purchased from Eastman (Rochester, N.Y., U.S.A). Butanoic acid, hexanoic acid, octanoic acid, decanoic acid, dodecanoic acid, tetradecanoic acid, 9-tetradecanoic acid, hexadecanoic acid, 9-hexadecanoic acid, octadecanoic acid, 9- 49 octadecanoic acid, 9,12-octadecanoic acid and 6,9,12 octadecanoic acid were used for the standard stock solution, and were obtained from Aldrich Chemical Co. Inc (Milwaukee, Wisconsin, U.S.A). Heptane, Isopropanol, Sulfuric acid, anhydrous sodium sulfate, chloroform, formic acid and diethyl ether were obtained from Fisher. Extraction From each 2lb block of cheese, 100 grams were wrapped in alumina foil, frozen with liquid nitrogen during 6 minutes, and then grinded for 30 seconds to obtain a fine powder. Six grams of this previously freeze-ground cheese, 1 ml of 2N sulphuric acid and 1 ml of internal standard solution (C5:0, C7:0, C9:0, C11:0 and C17:0 in 1:1 hetpane-isopropanol) were mixed with 7 grams of anhydrous sodium sulfate and 20 ml of 1:1 diethyl ether- heptane in a 40 ml amber vial using a sonicator and manual agitation. During sonication, the salt-slurry solution is initially exposed for 15 minutes, after which each vial is shake vigorously to continue with a second sonication period of 20 minutes. With a glass-Pasteur pipette, the sample extract (solvent) is transferred to an AccuBOND amino cartridge (Agilent Technologies) conditioned previously with 10 ml of heptane. After the addition of the sample, the column is washed with 5 ml of 2:1 Chloroform-Isopropanol to remove non volatile triglycerides and phospholipids using a manifold vacuum chamber. Once the washing step is complete, free fatty acids are eluted with 5ml of 2% formic acid in diethyl ether, collected in a 20 ml vial and stored in the freezer until GC analysis. Chromatography The analysis was performed using a Hewlett-Packard 5890 gas chromatograph equipped with a flame ionization detector (FID). Samples were analyzed on a DB-FFAP column (15m x 0.53mm ID, 1 m film thickness; Supelco Wax10, Supelco U.S.A). Injector and detector temperatures were 250C. Nitrogen was used as carrier gas at a flow rate of 15 ml per minute at a split ratio of 1 to 1. The oven temperature was programmed for a 2 minutes hold at 60C, raised to 230C at a rate of 8C per minute with a hold of 20 minutes at 230C. Quantitative analysis The levels of free fatty acids concentrations were calculated based on individual peak area from GC-FID response in comparison to the internal standard peak area, by using standard 50 calibration curve of individual free fatty acid using Peak Simple software (SRI instruments, Torrance, CA). Each experimental value corresponds to the average of the 3 extraction replicates. VOLATILE SULFUR COMPOUNDS (VSC’S) Chemicals Dimethyl sulfide (DMS) was purchased from TCI America (Portland, OR, U.S.A.); gaseous methanethiol (MeSH) was obtained from Aldrich Chemical Co. Inc (Milwaukee, Wisconsin, U.S.A), and a solution was prepared by bubbling the gas into cold methanol; a H2S solution was prepared by dissolving Na2S.9 H2O (Sigma Co) in acidic water stabilized with citric acid (pH 3). Extraction From each 2lb block of cheese, 100 grams were wrapped in alumina foil, frozen with liquid nitrogen during 6 minutes, and then grinded for 30 seconds to obtain a fine powder. Then one gram of this freshly prepared powder is added to a 20ml vial (formerly flushed with argon), followed by the addition of 4 ml of 1M citric acid and 20 l of the internal standard solution. After addition of sample vials were immediately sealed with screw caps with teflon-lined silicone septa. The vials used in this study were previously deactivated with DMTCS 5% solution in toluene, toluene, methanol and distillate water. The volatile sulfur compounds were extracted with an 85 m Carbox-PDMS fiber (Supelco, Bellefonte, PA, U.S.A.). Prior to use, the fiber was conditioned at 300 C for 90 minutes. The fiber was then placed into a SPME adapter of a CombiPAL autosampler (CTC analytics AG, Zwingen, Switzerland) Fitted with a vial heater/agitator. Samples were pre-equilibrated at 500 RPM at 40C for 5 minutes, and the extraction of VSC’s was done at 250 RPM at 40C for 25 minutes. The desorption time was 5 minutes and 30 seconds. Chromatography The analysis was performed using a Varian CP-3800 gas chromatograph (Varian, Walnut Creek, CA, U.S.A.) equipped with a pulsed flame photometric detector (PFPD). The separation of analytes was made using a DB-FFAP fused silica capillary column (30m, 0.32 51 mm ID and 1 m film thickness; Agilent, Palo Alto, CA, USA) and nitrogen as carrier gas at constant flow at 2 ml per minute. The injector temperature was 300 C and it was in the splitless mode. The oven temperature was programmed for a 3 minutes hold at 35C, raised to 150C at a rate of 10C per minute, held for 5 minute, and then heated to 220C at a rate of 20C per minute with a final hold of 3 minutes. The PFPD was held at 300 C and 450 V with the following flow rates: Air 1 at 17 ml per min, H2 at 14 ml per min, and Air 2 at 10 ml/min. The detector response signal was integrated using the software Star Workstation 6.2, Varian) Quantitative analysis Matrix effect In order to retain the matrix effect during the construction of the calibration curves, cheese powder from the “youngest sample” is used. It is de-volatilized by exposure to room conditions in a hood for 2 hours. Then 1 gram of powder is added to 4 ml of 1M citric acid in a 20 ml vial and exposed to a 50C water bath for 30 mins, prior to the addition of standards and internal standard solutions. Sulfur standards and internal standard preparation Two internal standards were used for the quantification of VSC’s: ethyl methyl sulfide (EMS) for H2S, MeSH and DMS, and isopropyl disulfide (IsoProDS) for DMDS and DMTS. The concentration of the internal standard solution was 500 ppm EMS and 500 ppm IsoProDS in methanol. Calibration curves were constructed by spiking cheese samples with a range of known concentrations of H2S, MeSH and DMS. Hydrogen sulfide (H2S) was prepared by dissolving Na2S.9 H2O in acidic water (pH = 3). Different concentrations of sodium sulfide solutions were made, and the concentrations of H2S were calculated based on the amounts of salt added to the matrix. A standard solution of 100 ppm of DMS was individually prepared in cooled methanol (-15C), and dilutions were made with cooled methanol at the same temperature. The mesh standard was prepared as following: 1) newly deactivated, recently flushed with argon, and cooled vials were used; 2) The original standard solution was made by bubbling pure MeSH into cooled methanol; 3) Dilutions were prepared by taking aliquots from the original solution contained in a sealed vial, through the teflon-lined silicone septa by using a syringe. And then injecting the aliquots into new sealed vials containing proportional amount of cooled methanol through the septa; 4) 1 gr of devolatilized cheese is added to a 52 recently flushed vial (argon was used), which is immediately flushed again; 5) simultaneous argon flushing and addition of 4ml of “free” dissolved oxygen-1M citric acid solution and quick sealing of the vials; 6) Addition of 20 l of internal standard and MeSH standard through septa. The identification of target compounds was made by comparing retention times with those of pure standards. Ratios of the square root of the standard area to the corresponding square root of the internal standard area were plotted Vs concentration ratios to determine the relation between the response and concentration for the unknowns. Triplicate analysis was performed for all samples PROTEOLYSIS Chemicals Sulfuric acid was purchased from Fisher Scientific International Inc. (Pittsburgh, PA, U.S.A.); Trichloroacetic acid was purchased from Alfa Aesar (Royston, UK); and phosphotungstic acid was obtained from Aldrich Chemical Co. Inc (Milwaukee, Wis, U.S.A) Sample preparation and fractionation From each 2lb block of cheese, 60 grams are blended with 120 ml of distilled water pre-heated to 55C. The mixture is blended for 5 minutes and the homogenate is incubated at 55C for 1 hour. Then the pH is adjusted to 4.6 with 1M HCl and the mixture is centrifuged at 3000g for 30 minutes at 4C. Suspension and supernatant were filtered thoroughly 3 times through glass wool. The filtrate was safe at -20C for macro blog digestion method analysis, and RP-HPLC analysis. The insoluble pellet was frozen at -20C for further Urea-PAGE gel electrophoresis analysis. The trichloroacetic acid soluble nitrogen fraction (TCA-SN) was prepared by the addition of 25 ml of pH 4.6 soluble fraction (WSN) to 25 ml of 24% trichloroacetic acid solution. Then the mixture is equilibrated for 2 hours at room temperature and filtered through filter paper Whatman No 40 before macro blog digestion method analysis. For the phosphotungstic acid soluble nitrogen fraction (PTA-SN), 10 ml of WSN are added to 7 ml of 3.95 M H2SO4 and 3 ml of 33% phosphotungstic acid solution. Then the mixture is 53 equilibrated overnight at 4C and filtered through filter paper Whatman No 40 before macro blog digestion method analysis. Duplicate analysis was performed for all samples. Macro blog digestion (Kjeldahl Digestion) From the fractions collected an aliquot (2 ml for the Water soluble fraction, 1ml for TCA-SN and 1 ml for PTA-SN) is added into a 70 ml Kjeldahl Digestion flask with 10 ml of H2SO4 and the catalyst pellet containing 0,075 and 1,5 grams of mercuric oxide and potassium sulfate respectively. The mixture is warmed to 150 C and hold for 1 hour, then heated to 250 C and hold for 1 hour, and finally heated to 350 C and hold for 2 hours. After digestion the sample is cooled down overnight to room temperature, and diluted with distillate water to 70 ml, followed by a gentile agitation. Then a 5 ml aliquot is used to determine the nitrogen content by a rapid flow analyzer FOSS II. Reversed phase High performance liquid chromatography analysis The RP-HPLC analysis was performed using a Shimadzu 6 series liquid chromatograph (Shimadzu scientific instruments, Kyoto Japan), consisting of an autosampler, 2 pumps, a multi-wavelength spectrophotometer and a controller unit. It was used a nucleosil RP-8 analytical column (250x 4mm, 5 m particle size, 300 A pore size) and a guard column (4.6 x10 mm) from waters (Milford, MA, U.S.A.). The mobile phase consists of solvent A (0.1% TFA in deionized and vacuum filtered water) and solvent B (0.1% TFA in acetonitrile). The elution was monitored at 214nm. The following gradient elution was performed: 1) 100% solvent A for 5 minutes followed by a linear gradient to 55% solvent B (v/v); 2) elution at 55% solvent B for 6 minutes followed by a linear gradient to 60%; 3) elution at 60% solvent B for 3 minutes; 4) The column is washed using 95% solvent B during 5 minutes; 5) the column is equilibrated using 100% solvent A during 10 minutes. The sample (WSN fraction) was dissolved in solvent A (10 mg per ml) and then micro-centrifuged at 14000 RPM for 10 minutes. An aliquot of 40 l from the extract was injected to a flow rate of 0.75 ml per min. 54 Electrophoresis Samples of the water-insoluble nitrogen fraction were dry frozen prior to analysis. Samples were dissolved in a buffer (0.75 g tris, (hydroxymethyl) methylamine, 49 gr urea and 0.4 ml concentrated HCl, 0.7 ml 2-mercaptoethanol and 0.15 gr bromophenol blue, dissolved to 100ml) and hold at 50C for 40 min. Urea-polyacrylamide gel electrophoresis (urea-PAGE) was carried out using a Protean II xi cell vertical slab unit (Bio-Rad Laboratories ltd., Hemel Hempstead, Herts, UK). Urea-PAGE gels (12.5%) were prepared and run according to the method or Ardö (1999). Reagents used were obtained by Sigma-Aldrich, Inc and Fisher Scientific. STATISTICAL ANALYSIS A two-way analysis of variance (ANOVA) on data was carried out using a general linear model procedure with Turkey’s pair wise comparison at 95% confidence level, using the package Minitab 16 (minitab Ltda., Coventry, UK). RESULTS AND DICUSSION FREE FATTY ACIDS FFA The levels of lipolysis obtained in this work, measured as the amount of individual FFA, showed subtle but interesting difference between the cheeses prepared with pasteurized milk and those made with heat-shocked milk. In spite of the initial loss of some short and medium chain FFA during whey draining, it can be seen the tendency of FFA to increase during maturation of samples. Indeed, it is evident in Figures (2) that short chain FFA tend to increase faster, reaching concentrations about 4 times the initial one by the end of the observation period for both type of treatments. On the one hand, this can be related to the mobility and better access of enzymes to these substrates, which are essentially located at the positions sn-1 and sn-3 (Balcão and Malcata 1998). On the other hand, this behavior indicates that enzymatic activity is most likely dominated by lipases since they are specific for the outer ester bonds of tri- or diacylglycerides (Deeth and Touch 2000). In addition, this last observation is support by the fact that in spite of the low change, long chain FFA increased their concentration during ripening, which is directly related to lipolytic activity rather than activity promoted by esterases. 55 Despite the most dominant peaks or FFA in the chromatograms correspond to C14:0, C16:0, C18:0 and C18:1, due to their significantly lower odor thresholds (Molimard and Spinnler 1996), they are not considered as important contributors to the overall aroma of Cheddar cheese. Contrary, in the case of FFA such as C4:0, C6:0, C8:0, C10:0 and C12:0, it is known that in spite of their lower concentration, they contribute directly and indirectly to the characteristic aroma of Cheddar cheese, and it was seen that their rate of generation depend on the type of heating treatment of cheese milk. Nonetheless, no significant difference was found, but it is evident the trend of cheeses prepared with heat shocked milk to develop slightly higher levels of short chain FFA. This observation can be related to higher LPL activity in samples made with heat-shocked milk, since it is well known that 78 C for 10 seconds are required for the complete inactivation of this enzyme (Driessen, 1989), and it has been accepted that despite 72 C for 15 seconds inactivate the enzyme extensively, it still contributes to lipolysis in pasteurized milk cheese. Therefore, a lower temperature treatment is expected to affect the activity of this enzyme to a lesser extent. Indeed, this can be supported by the fact that LPL is specific for the sn-1 and sn-3 positions of mono, di and triacylglicerides (Olivecrona et al. 1992), which are the positions where C4:0 and C6:0 are predominately located along with other unsaturated FFA (Balcão and Malcata 1998). However, since the findings are subtle differences rather than significant, it is important to keep in mind that this kind of ripening behavior might not be reproducible. Alternatively, based on the fact that the principal lipolytic activity is provided by LAB enzymes, which are mostly intracellular and have nothing to do with the heat treatment of milk, it could be thought that the differences found in this work can be attributed to physical alteration of milk as consequence of the heating treatment of cheese milk (Dalgleish and Banks., 1991), resulting in a more difficult access for lipases and esterases of LAB in pasteurized milk cheeses to their substrates (Hickey et al. 2007), and probably higher LPL activity in Heat-shocked milk cheeses. 56 Figure 1 FFA chromatogram Heat –schokecd Vs Pasteurized milk Figure 2 Development of individual FFA in Heat-shocked and Pasteurized milk cheese 57 58 Volatile Sulfur Compounds VSC’s The decomposition of sulfur containing amino acids (cysteine and methionine) is another biochemical event occurring during Cheddar cheese ripening resulting in the characteristic aroma of this variety. In addition to free fatty acids (FFA), Volatile sulfur compounds (VSC’s) correlate with good Cheddar cheese flavor (B. Weimer, Seefeldt, and Dias 1999). When smelled alone they smell like garlic, onion, cabbage and skunk, but when they are mixed, they contribute to pleasant Cheddar cheese flavor notes. It has been reported that high concentrations of H2S, MeSH, and DMS are found in Cheddar Cheese, while DMDS, DMTS and 3-methylthiopropionaldehyde have low concentrations. Other compounds such as Carbonyl sulfide, carbon disulfide and dimethyl sulfone are not important contributors (H. M. Burbank and Qian 2005). 59 The results from this work suggest that after assuring the stability of samples through the use of an organic acid buffer solution (citric acid 1M), and the proper de-activation of injection liner and vials to prevent methanethiol (MeSH) oxidation, only hydrogen sulphide (H2S), carbon disulphide (CS2), MeSH, and dimethyl sulphide (DMS) were the volatile sulfur compounds developed during ripening. However, in figure (3) it can be seen that only small and negligible amounts of dimethyl disulphide (DMDS) and dimethyl trisulphide (DMTS) were rarely found in the chromatograms for the samples analyzed. Whereby, only the development of MeSH, DMS and H2S will be discussed. Because of the wide range of volatility and concentration of the VSC’s in the samples, and the different selectivity of the CAR-PDMS SPME fiber, standard calibration curves had to be calculated for each compound. It can be seen in figures 4 and 5 that good linear correlation coefficients were obtained for H2S and DMS, but unfortunately not for MeSH. Whence, the interpretation of results for this last compound was done based on the area ratio, between MeSH and the internal standard EMS, instead of using its concentration during the ageing process. The results indicate that the heating treatment of cheese milk had a significant effect on the VSC’s development during ripening. In both cases, pasteurization and heat-shock treatments, temperature influenced the VSC’s concentration during ripening, suggesting that NSLAB population and denaturation of serum proteins of milk may have contributed to their formation. In figure 6, hydrogen sulfide did not show a steady development for the heat-shock treatment whereas it increased as cheese aged for the pasteurization treatment. Indeed, during the initial stage of the ageing process the difference between the treatments was not significant, however after 6 months it became evident, and the samples made with pasteurized cheese milk displayed a higher concentration of H2S than those made with heat-shocked milk. It is also possible to see that in spite of the unclear difference between treatments during the first 6-7 months the concentration of H2S increased over time, reaching a plateau after 9 months in the case of pasteurized milk samples. Moreover, in the case of the heat-shock samples, it was difficult to establish any trend during the ripening. 60 Since H2S sensory threshold is 10 ppb in water (Rychlik et al. 1998) and its concentration along the ageing process varied from 12 to 50 ppm in the case of pasteurized samples, and from 20 to 35 ppm for the heat-shocked samples; it was possible to confirm its role as key contributor to the cheddar cheese aroma. The appreciable higher concentration for the pasteurized samples can be attributed to higher denaturation and incorporation of βlactoglobulins to the casein micelles. Indeed, Cysteine, a sulfur containing amino acid present in limited amounts in caseins, is the main precursor of H2S and it can be generated from the sulfydryl groups once thermal breakdown of cysteine takes place (Fennema and Damodaran 1996). Thus, the results obtained suggest that the higher temperature used in the pasteurization treatment promotes more coagulation of whey protein, increasing the availability of cysteine and consequently the development of H2S during the cheese ageing. Indeed, Hutton and Patton 1952; K. R. Christensen and Reineccius 1992 reported that the concentration of H2S in milk increases linearly with heating temperature. In the case of MeSH, because of the limited sulfur content of amino acids in caseins, it was initially expected a higher concentration of this compound for the pasteurization treatment during the cheese ripening due to the possible higher inclusion of whey protein into the cheese curd. However, the results in figure 7 indicate a clear difference between treatments and surprisingly higher amounts of MeSH for the samples made with heat-shocked cheese milk. In addition, this trend suggests that the difference between treatments can arise from the enzymatic activity of LAB and survival NSLAB coupled with the effect of the treatments on the chemical structure of milk. The higher development of MeSH for the heat shock treatment can be related to a bigger population of indigenous bacteria as consequence of the lower temperature employed, resulting in a possible increase of activity of the enzymes L-methionine γ-lyase (Tanaka, Esaki, and Soda 1985) and/or cystathionine β-lyase and γ-lyase (Alting et al. 1995) during ripening. As a matter of fact, some of the potent odorants in the cheddar cheese profile result from leucine and methionine degradation, and it has been proposed that MeSH can be produce from L-methionine via: 1) lysis of the C-S bond by L-methionine γ-lyase and/or cystathionine β-lyase or γ-lyase), or 2) through a two-step pathway that involves the transamination of L- 61 methionine in the presence of a-keto glutarate to form a-keto-g-methyl thiobutyrate (KMTB) (Yvon, et al 1997; Gao et al. 1998; Amarita et al. 2001), followed by its enzymatic breakdown to form 3-methylthiopropionaldehyde and MeSH. In addition, when methanethiol is produce from cysteine through the β-elimination reaction hydrogen sulfide is produced (Dobric et al. 2000; María Fernández et al. 2000) Regarding the general absence in the results of this study of the sub-products DMDS and DMTS resulting from the oxidation of MeSH, it might be possible to say based on the thorough sample preparation work and the fact that cheese has a low redox potential, -150 to 200 mV, (Donald J. Manning and Moore 1979; Green and Manning 1982), that these compounds are not original odorants produce during the ripening of Cheddar cheese, resulting in “negligible” amounts. On the other hand DMS had high concentrations. The results in figure 8 show a significant difference between treatments, and it can be seen a steady increase of DMS for both type of samples during the ripening stage, and higher amounts of DMS for the cheese prepared with heat-shocked milk. The DMS concentration for most of the samples evaluated was higher than its sensory threshold, 2ppm in water (Rychlik et al. 1998), demonstrating that this compound is an important odorant in the Cheddar cheese matrix. The concentrations in pasteurized milk varied from 1 to 14 ppm, while those for the heat-shock samples changed from 8 to 45 ppm, which are amounts comparable to the ones obtained by Burbank and Qian (2008). As far as we know there is no a proved and clear mechanism for the generation of DMS during cheese ripening. However, it is well known that DMS concentration in raw milk is significant and it is influenced by the diet of the cows (Manning et al. 1976; Forss 1979). Furthermore, it is known that DMS can be generated from sulfhydryl group of milk proteins, mainly β-lactoglobulin and if present the milk fat globule membrane proteins, where methionine is most likely the precursor for its generation (Datta et al. 2002). Therefore it is expected that DMS is a natural component of the cheese milk, either raw or heated. Moreover, lately in the work of (R. de Wit and Nieuwenhuijse 2008), it has been reported even the possibility of a oxidation of MeSH into DMS and H2S (discussed later in the next chapter). 62 Figure 3 VSC's chromatogram Heat –schocked Vs Pasteurized milk Figure 5 Calibration Curve H2S Figure 7 Development of MeSH in Heat-schocked and Pasteurized milk Figure 4 Calibration curve DMS Figure 6 Development of H2S in Heat-schocked and Pasteurized milk 63 Figure 8 Development of DMS in Heat-schocked and Pasteurized milk Effect of treatment on Proteolysis Cheddar cheese samples manufactured from pasteurized milk (72C for 15 sec) and heatshocked milk (66C for 30 sec) were examined during an 18 month ripening period by measurements of Total Kjeldahl Nitrogen (TKN) of the WSN, TCA-SN and PTA-SN fractions, RP-HPLC peptide profile of the WSN fraction and Urea-PAGE peptide profile of the water insoluble fraction. The results revealed differences in the rate and pattern of proteolysis. Indeed, from the three methods employed to evaluate the extend of proteolysis, the TKN measurements and the peptide profile analysis by RP-HPLC of the WSN fraction, were more effective than the Urea-PAGE analysis for discriminating the impact of the heat treatment of cheese milk between samples. As a matter of fact, it was possible to observe considerable differences in the primary and secondary proteolysis. Soluble Nitrogen Fractions and TKN Alteration to the ongoing proteolysis due to the heat treatment of cheese milk was obvious by making use of the described fractionation scheme, which is based on the fact that extractability of nitrogen compounds depend on pH (Ardö and Frederiksberg 1999; Sousa, Ardö, and McSweeney 2001). Thus, it can be seen in figures 9 to 13, that the nitrogen level in the recovery fractions increased during time and was different regarding the heating treatment of cheese milk. Among the nitrogen indices proposed in this part of the study, the WSN fraction includes all casein breakdown products, but native caseins and high molecular weight peptides; the 12% trichloracetic TCA-SN fraction contains small peptides and FAA; and PTA- 64 SN fraction, which contains the smallest peptides (600 Da) and FAA (T. M. I. E. Christensen, Bech, and Werner 1991). In the figures 9 and 10, the levels of WSN were the highest for both types of cheeses, and had values about 4 times higher than those of the TCA-SN fraction and 10 times higher in comparison to those of the PTA-SN fraction, except for those points corresponding to the first 4 months of maturation, which were much higher, close to 6 and 12 times respectively, which might be related to the primary proteolysis. As a matter of fact after 6 months of aging, the values of TCA-SN and PTA-SN increased substantially for the all the samples. On the other hand, it is possible to appreciate in figures 11, 12 and 13, how the values were significantly affected by the treatment, and the last nitrogen contents in every fraction were 1.5 to 2 fold higher than those at the beginning of the observation. Regarding the levels of the WSN fraction, figure 11 shows how the values increased over time, and although the amounts were similar at the beginning of the ageing process for both types of samples, they were higher for those samples prepared with heat-shocked cheese milk. In addition, the result in this figure also confirm the use of this fraction as an effective index of primary proteolysis (Bansal, Piraino, and McSweeney 2009), and showed how the heating treatment of the cheese milk, regardless the intensity and resemblance of it, can cause an appreciable difference in the maturation of the cheese samples.This can be related to the fact that the main enzymatic activity during primary proteolysis is provided by the remaining rennet and the indigenous milk proteinase (S. Visser 1993; Paul L. H. McSweeney et al. 1994). Also, this is believed to do with the altered accessibility of the retained Chymosin to the substrates, αs1 and κ caseins (Mulvihill and Fox 1979; P. F. Fox 1989), and with the impact of the treatment on the Plasmin activity, which in spite of being stable at elevated temperature, is definitively influenced by the heating protocol (Alichanidis, Wrathall, and Andrews 1986; Metwalli, de Jongh, and van Boekel 1998). Indeed, it is possible to suggest that the difference between treatments could be attributed to better accessibility of Chymosin to the bonds Leu101-Lys102, Phe32-Gly33 and Leu109-Glu110 of the large peptides. The levels of TCA-SN in Figure 12 also illustrate a proteolysis development affected by the heating treatment of cheese milk. The increment in the soluble nitrogen in this fraction for the 65 experimental cheeses revealed a trend similar to that of the WSN fraction. The pasteurization of milk decreased the concentration of nitrogen in the fraction with respect to the results of the heat-shocked milk samples. However, it is possible to observe that the evolution of this fraction for both types of samples was more regular and sustained throughout ripening, resulting in final values that are approximately 3 times higher than those at the beginning of the observation. Also, despite the results were slightly lower than those values found by (Voigt et al. 2012) for the control sample, they are still comparable. Based on the fact that TCA is a fraction rich in small peptides of low and medium hydrophobicity (Kuchroo & Fox 1982), which contain peptides or traces of peptides mostly derived from the N-terminal half of β-casein and from the N-terminal half of αs1-casein (Tanoj K. Singh, Fox, and Healy 1995; T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998) resulting from the starter and NSLAB enzymatic activity (O’Keefe et al. 1978). It might be possible to suggest that the difference between treatments in this fraction can be attributed to intracellular and extracellular peptidases and amino peptidase of the LAB and NSLAB. Indeed, it has been reported that in spite of the destruction by pasteurization of pathogens, coliforms, psychotrophs and the reduction of most of the LAB strain by 6 log (Burton 1986), it has been found that one strain of Lactobacillus casei var. casei was reduced by only 3.5 log and that thermophilic LAB were definitively more resistant to this treatment, in addition to other facts such as that in Swiss cheese only 4 of 60 lactobacilli isolated were eliminated at 71C for 18 s (Bassett and Slatter 1953; Niven, Buettner, and Evans 1954), and that Propionibacterium freudenreichii, found in raw milk (Baer and Ryba 1992), survived heating at 62.8 C for 30 min. Therefore, it is evident that in the case of a milder treatment of the cheese milk such as Heat-shock, it can be expected to find a more significant growth of NSLAB in Cheddar Cheese, resulting in a TCA fraction with higher content of soluble nitrogen. On the other hand, Lau, Barbano, and Rasmussen 1990; Lau, Barbano, and Rasmussen 1991 found slightly higher level of moisture in cheddar cheese made from pasteurized milk in comparison to cheese made from raw milk, which indicates that syneresis could be affected by the treatment and can impact on the proteolysis pattern because of the levels of bound water that can result in less protein hydrolysis. It was found in the work of Whetstine et al. 2007, 66 that Cheddar cheese blocks corresponding to the inner side of a 291 Kg block had smaller moisture content and had faster proteolysis in comparison to those of the outer side, characterized by higher moisture content and less protein hydrolysis, which might be reflected in the mobility of small peptides that contribute to the TCA-SN fraction. Which is another reason why it can be expected that the heat-shock treatment, which is milder, shows slightly higher levels of soluble nitrogen in TCA fraction. The PTA-SN fraction is an index of secondary proteolysis because it is mainly constituted by very small peptides (<15 kDa) and amino acids of approximately 600 Da (Aston and Dulley 1982). The results for this fraction showed in Fig 13, indicate a similar trend to the other fractions, where the values increased over time and showed faster proteolysis for the heatshocked samples. Moreover, it is possible to appreciate that in comparison to the other fractions, the increment of nitrogen content was more regular and sturdy, displaying final values that are 4 times higher than those at the begin of the observation. Additionally, it can be seen that the values for both treatments were really tight during the first 4 months, which coincides with the period of time corresponding to the primary proteolysis. In a similar way to the TCA fraction, these results could be explain by the low efficiency of the milder heating treatments on NSLAB, elucidating the role of milk flora in the rate of proteolysis during the ripening stage. In general, it is possible to appreciate that as cheese ages the total amount of water soluble peptides increases. In addition, it is feasible to affirm that the fractionation using water is sufficient to extract the majority of water soluble peptides to establish a fair comparison between treatments through the recovered nitrogen. Some possible explanations to the appreciable difference in the proteolysis between treatments has to do with 1) The increase in plasmin activity, regardless if it was due to denaturation of the inhibitors of the plasminogen activators, or because of the inactivation of plasmin inhibitors (Baer et al. Collin et al. 1990; Bastian & Brown 1996); 2) The denaturation of whey protein and subsequent alteration of the rennetability of milk, affecting the moisture content the and LAB enzymatic activity; 3) And as it has been mentioned above, a low elimination of NSLAB activity. 67 Figure 9 TKN fractions heat-schocked cheese milk Figure 10 TKN fractions pasteurized cheese milk Figure 11 WSN heat-shocked and pasteurized cheese milk 68 Figure 12 TCA heat-schocked and pasteurized cheese milk Figure 13 PTA-SN heat-schocked and pasteurized cheese milk Peptide analysis by RP-HPLC The peptide analysis by RP-HPLC is an alternative index to express the degree of secondary proteolysis. In addition, it can be used in authenticity studies and optimization process (Upadhyay et al. 2004). In this case, in order to evaluate the impact of the heating treatments of the cheese milk on the peptide profiles in the chromatograms, it was used a principal component analysis (PCA), which is essentially a multivariate analysis tool use in descriptive statistics, to estimate the 69 linear relationship between variables when their number is very large (Chatfield and Collins 1981). The raw data from chromatograms were processed based on Piraino, Parente, and McSweeney 2004 work, where the complexity of the profiles containing more than 70 peaks, is initially reduced by establishing a set of time intervals according the elution time; for which the area of each one is expressed as percentage of total area of the chromatogram. Then the variability due to treatments, biological factors (ripening, cheese making process, milk quality, etc) and technical factors (sampling, extraction steps, and measurement of peak and intervals area), is measured and analyzed through the variation of the contributing eigenvalues in the correlation matrix for the principal components of the resulting model. Two treatments were compared through a randomized block design with three replicates. The peptide profiles revealed differences that were supported by the PCA results. Indeed, the PCA analysis leads to a model with three principal components (PCs) that explain the 82.6% of the variability of the data as can be seen in figure 14. However, after comparing in pairs the score plots for any combination of the principal components, only the score plot for the PC1 Vs PC2 revealed a correlation. This is in agreement with the published results of (Benfeldt and Sørensen 2001). The first and second principal components explained the 73.1% of the variance. In the score plot for this two PC’s (Fig. 15 and 16), it is possible to see how the PC1 differentiates the samples according to their age, while PC2 represent mainly the contribution of the heating treatment to the variation. Thus, the figures display higher scores for those samples with longer ripening that correspond to the heat-shock treatment, which suggest that proteolysis is slower for cheeses made from pasteurized milk. The loading plot in figure 17, presents the projection of the eluting intervals on the PC1 and PC2, and allows determining correlations between variables and their effect on the amount of peptides eluting within certain retention times. Regarding the segments from 12-20 and 20-25 minutes, it can be seen that they have high scores for the PC1 and values close to 0 for PC2, which suggest that the amount of peptides eluting in this zones increases during time and is relatively unaffected by the type of heat treatment of the cheese milk. This interpretation could be explain by the fact that the segments from 12-20 and 20-25 are mainly composed by 70 peptides products from the action of Chymosin on αs1-CN and k-caseins. In addition, there are other peptides resulting from the action of cell envelope proteinase (CEP) from LAB such as αs1-CN(f1-9) and αs1-CN(f1-13), which accumulate during ripening (Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998). This indicates that the formation of these hydrophilic peptides is mainly dependent on proteolytic systems that are not affected by the heat treatment of the cheese milk, and it is probable that the subtle differences between samples can be attributed to the amount of NSLA that survive after the treatment. In contrast, the segments 25-30 and 30-35 got negative loadings for the PC1, meaning that the relative amount of peptides eluting in this intervals decrease over time. This is believed to do with the breakdown of the αs1-CN, αs2-CN, αs1-CN (f24-199) and β-CN peptides, which is related to enzyme activity proportionate by the rennet and indigenous milk enzymes. However, it is important to keep in mind that the concentration of αs2-casein appears to decrease during ripening, but no large peptides derived from αs2-casein have been reported yet (Mooney et al. 1998), and only a few small peptides have been identified in the WSE (T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998). Indeed, these observations are in agreement with previous works in Danbo cheese (Benfeldt et al. 1997; Benfeldt and Sørensen 2001), and make sense since the breakdown of caseins by chymosin and plasmin takes place along the ripening (especially during the primary proteolysis). Nonetheless, the PC2 loading scores for these 2 intervals reveal a mild reduction in the amount of peptides eluting within these retention times as the temperature of the heating treatment increases. Certainly, it is expected that the Plasminogen activation system and the plasma-derived proteinase inhibitors respond to any thermal change. As a matter of fact, while Plasmin and the complex Plasminogen activation system is related to the casein micelles and are stable at high temperatures (S. Christensen et al. 1995), the proteinase inhibitory activity in milk is associated to the serum phase (Heegaard, Rasmussen, and Andreasen 1994), which is susceptible to heat denaturation due to its secondary and tertiary structures. Thus, it can be thought that a lower treatment temperature causes less denaturation and association of serum proteins to the casein micelles, and consequently less integration of inhibitors of Plasminogen activators into the cheese. Additionally, less thermally induced thiol disulphide exchange 71 between Plasmin and β-lactoglobulin can be expected, (P. F. Fox and Stepaniak 1993), resulting in higher Plasmin activity, reflected in the loading scores for these intervals that contain the αs2-CN and β-CN peaks. Also, a possible reduced activity of Chymosin on the αs1-CN might be expected and can be related to the association of β-lactoglobulin to the casein micelle, which makes more difficult the accessibility of Chymosin to its active sites, leading to a subtle difference between the treatments, which is appreciable as well in the score for the 25-30 that is slightly off from zero regarding to the PC2. On the other hand, the intervals 35-40 and 40-45 exhibit slightly positive loading scores in relation to the PC1, and fairly negative ones regarding to the PC2. This suggests that the relative amount of peptides in this elution fractions raise with the cheese ageing and diminishes as the treatment temperature is increased. The reasons for this behavior can be found in the fact that these fractions contain mainly hydrophobic peptides such as: 1) the fragments β-CN(f29–209), β-CN(f106–209) and β-CN(f108–209) (γ1, γ 2, and γ 3, respectively), whose concentrations increase during ripening (Farkye and Fox 1990) and are the result from the hydrolysis of β-Casein by Plasmin at Lys28-Lys29, Lys105-Gln106 and Lys107-Glu108 bonds; 2) the peptides αs1-CN(f93–?), αs1-CN(f24–30), αs1-CN(f26–32), αs1-CN(f26–34) resulting from the hydrolysis of the peptide αs1-CN(f24–199) by Chymosin, CEP and aminopeptidase (T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1995; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998); 3) peptides αs2CN(f204–207), which is a C-terminal residue and product of lactococcal CEP (T.K. Singh et al. 1994). Furthermore, the slightly difference between heating treatments of cheese milk regarding the amount of peptides eluting during these intervals can be related as well to a higher or total inactivation of the acid proteinase Cathepsin D and the leucocytes proteinase by pasteurization. Cathepsin D is more active on αs1-CN than on β-CN, and results in similar breakdown products than Chymosin does, specially on the peptide αs1-CN(f24–199) (P. F. Fox and McSweeney 1996; Larsen et al. 1996). This enzyme is completely inactivated at pH 7.0 and temperatures higher than 60C for 10 min, whereby it can be expected that the pasteurization treatment had a higher impact in its final proteolytic activity during ripening (Ducastaing et al.,1976). On the other hand, proteolytic activity of leucocyte proteinases is similar to that of cathepsin D, and it may be a source of cathepsin D in milk by itself (P.L.H. McSweeney, Fox, and Olson 1995). 72 Thus, because of the complete loss of leucocyte vitality by pasteurization (Grieve and Kitchen 1985), chemical alteration of the milk structure, and lower efficiency of the treatments in the elimination of native milk enzymes (Lau, Barbano, and Rasmussen 1991), it could be suggested that a “milder treatment” will be reflected on the scores for the loading plot and the scores plot. However, it is important to keep in mind that only 50% of the of β-casein in Cheddar cheese is hydrolyzed (T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1995; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998), and a still unknown role of the alkaline phosphatase enzyme, which is inactivated at 70C for 15 sec, may have a contribution to the slight difference between treatments (Griffiths 1986). Regarding the scores for the intervals between 45-65 minutes, in spite they are composed by hydrophobic compounds as well, they display a different trend to that shown in the intervals between 35 to 45 min, which suggest that a considerable amount of amino acids might elute in this zone, resulting from enzyme activity that is not severely affected by the heating treatment. However, it is possible to see in the loading plot that their amount decreases as cheese aged, which might has to do with the fact that as cheese ages more caseins, long, medium and small peptides are broken into smaller pieces that may or may not be water soluble and finally can undergo catabolic reactions (Lau, Barbano, and Rasmussen 1991). In addition, it has been thought that the material eluting in this region could correspond to high molecular mass molecules or molecules that contain aromatic amino acids, which are characterized for being very hydrophobic. Lau, Barbano, and Rasmussen 1990, proposed that the products of peptides containing aromatic amino acids can be highly hydrophobic and no longer water soluble, which may indicate smaller amounts of hydrophobic peptides as cheese aged (an observation that fits to the results obtain in this work). A complement to this last explanation is that after Aston and Creamer, 1986, demonstrate that the water-soluble nitrogen fraction is an important contributor to the nonvolatile flavor of cheese, Allan J. Cliffe, Marks, and Mulholland 1993, reported that fractions isolated by gel filtration of the water soluble extract of well matured Cheddar cheese, may vary from bitterness in the higher molecular mass components to savory for those of lower molecular mass. Therefore, bitter fractions can be related to material that was eluting late on the RP-HPLC column since they can have higher hydrophobic interactions. In addition, it has been said that bitterness in cheese is associated to the presence aromatic 73 amino acids in free form or as part of a peptide. Furthermore, based on earlier works that describe these segments as fractions mainly composed by amino acids, it can be expected that once they are in its free form they become precursors of flavor compounds through catabolic reactions, resulting in decreasing amounts of components for these zones of the chromatogram. Figure 14 Scree plot of heat-shocked and pasteurized cheese milk Figure 15 Score plot of heat-shocked and pasteurized cheese milk (age) 74 Figure 16 Score plot of heat-shocked and pasteurized cheese milk (temperature) Figure 17 Loading plot for heat-shocked and pasteurized cheese milk 75 Figure 18 Peptide profile pasteurized cheese (A and B) Vs Heat-shocked (C and D) milk, for 2 and 12 months A C B D Electrophoresis The results of the urea-PAGE of the pH4.6 insoluble nitrogen fraction of experimental Cheddar cheese in Figs 19 and 20, clearly exhibit the progressive change of αS1-CN into the peptides αS1-CN (f24-199), αS1-CN (f121-199), αS1-CN (f99-199), and the β-CN into peptides β-CN (f29-202), β-CN (f108-209) and β-CN (f106-209) during ripening. In addition bands of γ-CNs became more noticeable after 4 months. Also it can be seen that apparently the development of peptides from αS1-CN is faster than those from β-CN, which can be related to primary proteolysis and the actual amount of β-CN that is hydrolyzed (only the 50%). However, the representative urea-PAGE gel do not show any appreciable difference between treatment, and barely minor differences are perceived between treatments after 14 months in comparison to those of the early stages of ripening. 76 Figure 19 Urea PAGE for ripening of cheese made from heat-shocked milk β -CN(f106-209) β -CN(f29-209) β -CN(f108-209) β -CN αs1-CN αs1CN(f121-199) αs1 -CN(f24-199) 2M 4M 8M 10 M 12 M 14 M Figure 20 Urea PAGE for heat-schocked Vs pasteurized cheeses (12 months) Heat-shock Pasteurized 77 CONCLUSION The present study demonstrates that the use of FFA profile, VSC’s profile, measurement of the levels of the Total Kjeldahl nitrogen for the WSN, TCA-SN and PTA-SN fractions, and the analysis of the RP-HPLC peptide profile of the WSN fraction by using a PCA, are effective tools and indices of ripening to differentiate Cheddar cheese samples regarding to age and type of heat treatment of the cheese milk. In the case of Urea-PAGE, it was demonstrated that its use as index of primary proteolysis can be effective to differentiate samples by their age; nonetheless it is clear that it is a method not adecuate to detect differences between samples made from heat-shocked or pasteurized cheese milk unless it is coupled to other systems to obtain electrophotograms. In addition, it was demonstrated that proteolysis is faster for cheeses made with heat-shocked cheese milk since the results of nitrogen level for all the 3 fractions analyzed were higher than those found for the pasteurized cheese milk samples. This was supported by the PCA model obtained, which suggest alterations to the structure of milk and to the amount of remaining indigenous bacteria. It was found slightly higher levels of short and medium FFA; however, the difference FFA is not significant. The amounts found for DMS, H2S and MeSH showed appreciable differences between samples, and it can be seen that cheeses made from heat-shocked milk undergo faster catabolism of sulfur containing amino acids such as methionine and cysteine. Another interesting observation is that it seems like the DMDS and DMTS, reported in previos works as important contributors to cheese flavor, can be artifacts from extraction and separation procedures, consecuence of the oxidation of MeSH, rather than metabolites from the ripening of Cheddar cheese. REFERENCES Alichanidis, Efstathios, Julia H. M. Wrathall, and Anthony T. Andrews. 1986. “Heat Stability of Plasmin (milk Proteinase) and Plasminogen.” Journal of Dairy Research V53 (2): pp. 259–269. Alting, A. C., W. Engels, S. van Schalkwijk, and F. A. Exterkate. 1995. “Purification and Characterization of Cystathionine (beta)-Lyase from Lactococcus Lactis Subsp. Cremoris B78 and Its Possible Role in Flavor Development in Cheese.” Applied and Environmental Microbiology V61 (11) (November 1): pp. 4037–4042. Amarita, F., T. Requena, G. Taborda, L. Amigo, and C. Pelaez. 2001. “Lactobacillus Casei and Lactobacillus Plantarum Initiate Catabolism of Methionine by Transamination.” Journal of Applied Microbiology V90 (6): pp. 971–978. 78 Andersen, Lene T., Ylva Ardö, and Wender L.P. Bredie. 2010. “Study of Taste-active Compounds in the Water-soluble Extract of Mature Cheddar Cheese.” International Dairy Journal V20 (8) (August): pp. 528–536. Anderson, D.L., V.V. Mistry, R.L. Brandsma, and K.A. Baldwin. 1993. “Reduced Fat Cheddar Cheese from Condensed Milk. 1. Manufacture, Composition, and Ripening.” Journal of Dairy Science V76 (10) (October): pp. 2832–2844. Ardö, Y., and Frederiksberg. 1999. “Evaluating Proteolysis by Analysing the N Content of Cheese Fractions.” Bulletin - International Dairy Federation V337: pp. 4–9. Arora, Gulshan, Francois Cormier, and Byong Lee. 1995. “Analysis of Odor-Active Volatiles in Cheddar Cheese Headspace by Multidimensional GC/MS/Sniffing.” J. Agric. Food Chem. V43 (3): pp. 748–752. Aston, J. W., and L. K. Creamer. 1986. “Contribution of the components of the water-soluble fraction to the flavour of Cheddar cheese.” New Zealand Journal of Dairy Science and Technology V21 (3): pp. 229–248. Aston, J. W., and John R. Dulley. 1982. “Cheddar cheese flavor review.” Australian Journal of Dairy Technology V37 (2) (June): pp. 59–64. Avsar, Y.K., Y. Karagul-Yuceer, M.A. Drake, T.K. Singh, Y. Yoon, and K.R. Cadwallader. 2004. “Characterization of Nutty Flavor in Cheddar Cheese.” Journal of Dairy Science V87 (7) (July): pp. 1999–2010. Ayad, Eman H.E, Annette Verheul, Catrienus de Jong, Jan T.M Wouters, and Gerrit Smit. 1999. “Flavour Forming Abilities and Amino Acid Requirements of Lactococcus Lactis Strains Isolated from Artisanal and Non-dairy Origin.” International Dairy Journal V9 (10) (October): pp. 725–735. Baer, A., and I. Ryba. 1992. “Serological Identification of Propionibacteria in Milk and Cheese Samples.” International Dairy Journal V2 (5): pp. 299–310. Balcão, Victor M., and F.Xavier Malcata. 1998. “Lipase Catalyzed Modification of Milkfat.” Biotechnology Advances V16 (2) (March): pp. 309–341. Bansal, N, Paolo Piraino, and P. L. H. McSweeney. 2009. “Determination of Proteolysis in Cheese.” In Handbook of Dairy Foods Analysis, pp. 405–417. CRC Press. Bassett, Harold J., and W.L. Slatter. 1953. “The Heat Resistance of Some Lactobacilli Found in Swiss Cheese.” Journal of Dairy Science V36 (4) (April): pp. 409–413. Bastian, E.D., C.G. Lo, and K.M.M. David. 1997. “Plasminogen Activation in Cheese Milk: Influence on Swiss Cheese Ripening.” Journal of Dairy Science 80 (2) (February): 245–251. Bastian, Eric D., and Rodney J. Brown. 1996. “Plasmin in Milk and Dairy Products: An Update.” International Dairy Journal V6 (5) (May): pp. 435–457. Benfeldt, Connie, and John Sørensen. 2001. “Heat Treatment of Cheese Milk: Effect on Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4–7) (July 11): pp. 567–574. Benfeldt, Connie, John Sørensen, Katrine H. Ellegård, and Torben E. Petersen. 1997. “Heat Treatment of Cheese Milk: Effect on Plasmin Activity and Proteolysis During Cheese Ripening.” International Dairy Journal 7 (11) (October 24): 723–731. Benkler, K. F., and G. A. Reineccius. 1980. “FLAVOR ISOLATION FROM FATTY FOODS VIA SOLVENT EXTRACTION AND MEMBRANE DIALYSIS.” Journal of Food Science V45 (4): pp. 1084–1085. Bentley, Ronald, and Thomas G. Chasteen. 2004. “Environmental VOSCs––formation and Degradation of Dimethyl Sulfide, Methanethiol and Related Materials.” Chemosphere V55 (3) (April): pp. 291–317. 79 Beresford, Tom P, Nora A Fitzsimons, Noelle L Brennan, and Tim M Cogan. 2001. “Recent Advances in Cheese Microbiology.” International Dairy Journal V11 (4–7) (July 11): pp. 259–274. Beuvier, E., and S. Buchin. 2004. “Raw Milk Cheeses.” In Cheese Chemistry, Physics and Microbiology, Volume 1:319–345. Academic Press. Bills, D.D., and E.A. Day. 1964. “Determination of the Major Free Fatty Acids of Cheddar Cheese.” Journal of Dairy Science V47 (7) (July): pp. 733–738. Burbank, Helen M., and Michael C. Qian. 2005. “Volatile Sulfur Compounds in Cheddar Cheese Determined by Headspace Solid-phase Microextraction and Gas Chromatograph-pulsed Flame Photometric Detection.” Journal of Chromatography A V1066 (1–2) (February 25): pp. 149–157. Burbank, Helen, and Michael C. Qian. 2008. “Development of Volatile Sulfur Compounds in Heat-shocked and Pasteurized Milk Cheese.” International Dairy Journal V18 (8) (August): pp. 811–818. Burton, H. 1986. “Microbiological aspects.” (200): 9–14. Careri, M, and A Mangia. 2003. “Analysis of Food Proteins and Peptides by Chromatography and Mass Spectrometry.” Journal of Chromatography A V1000 (1-2) (June 6): pp. 609–635. Carunchia Whetstine, M E, P J Luck, M A Drake, E A Foegeding, P D Gerard, and D M Barbano. 2007. “Characterization of Flavor and Texture Development Within Large (291 Kg) Blocks of Cheddar Cheese.” Journal of Dairy Science V90 (7) (July): pp. 3091–3109. Carunchia Whetstine, Mary E., Keith R. Cadwallader, and MaryAnne Drake. 2005. “Characterization of Aroma Compounds Responsible for the Rosy/Floral Flavor in Cheddar Cheese.” J. Agric. Food Chem. V53 (8): pp. 3126–3132. del Castillo-Lozano, M. López, S. Mansour, R. Tâche, P. Bonnarme, and S. Landaud. 2008. “The Effect of Cysteine on Production of Volatile Sulphur Compounds by Cheeseripening Bacteria.” International Journal of Food Microbiology V 122 (3) (March 20): pp. 321–327. Chaintreau, Alain. 2001. “Simultaneous Distillation–extraction: From Birth to Maturity— review.” Flavour and Fragrance Journal V16 (2): pp. 136–148. Chatfield, Chris, and A. Collins. 1981. “Finding New Underlying Variables.” In Introduction to Multivariate Analysis, pp. 57–78. CRC Press. Chavarri, Felisa, Mailo Virto, Celia Martin, Ana I. Nájera, Arantza Santisteban, Luis J. R. Barrón, and Mertxe De Renobales. 1997. “Determination of Free Fatty Acids in Cheese: Comparison of Two Analytical Methods.” Journal of Dairy Research V64 (3): pp. 445–452. Chich, Jean-François, Karine Marchesseau, and Jean-Claude Gripon. 1997. “Intracellular Esterase from Lactococcus Lactis Subsp. Lactis NCDO 763: Purification and Characterization.” International Dairy Journal V7 (2-3) (February): pp. 169–174. Chopin, Alain. 1993. “Organization and Regulation of Genes for Amino Acid Biosynthesis in Lactic Acid Bacteria.” FEMS Microbiology Reviews V12 (1–3) (September): pp. 21– 37. Christensen J.E., Johnson M.E., and Steele J.L. 1995. “Production of Cheddar Cheese Using a Lactococcus Lactis Ssp. Cremoris SK11 Derivative with Enhanced Aminopeptidase Activity.” International Dairy Journal V5 (4): pp. 367–379. 80 Christensen, Jeffrey E., Edward G. Dudley, Jeffrey A. Pederson, and James L. Steele. 1999. “Peptidases and Amino Acid Catabolism in Lactic Acid Bacteria.” Antonie Van Leeuwenhoek V76 (1) (1): pp. 217–246. Christensen, K.R., and G.A. Reineccius. 1992. “Gas Chromatographic Analysis of Volatile Sulfur Compounds from Heated Milk Using Static Headspace Sampling.” Journal of Dairy Science V75 (8) (August): pp. 2098–2104. Christensen, Søren, Tove Wiegers, John Hermansen, and Lars Sottrup-Jensen. 1995. “Plasmaderived Protease Inhibitors in Bovine Milk.” International Dairy Journal V5 (5): pp. 439–449. Christensen, T. M. I. E., A.-M. Bech, and H. Werner. 1991. “Methods for crude fractionation (extraction and precipitation) of nitrogen components in cheese.” Bulletin International Dairy Federation (261): pp. 4–9. Christensen, Tove M. I. E., Kristian R. Kristiansen, and Jesper S. Madsen. 1989. “Proteolysis in Cheese Investigated by High Performance Liquid Chromatography.” Journal of Dairy Research V56 (5): pp. 823–828. Cliffe, A.J., D. Revell, and B.A. Law. 1989. “A Method for the Reverse Phase High Performance Liquid Chromatography of Peptides from Cheddar Cheese.” Food Chemistry 34 (2): 147–160. Cliffe, Allan J., Jonathan D. Marks, and Francis Mulholland. 1993. “Isolation and Characterization of Non-volatile Flavours from Cheese: Peptide Profile of Flavour Fractions from Cheddar Cheese, Determined by Reverse-phase High-performance Liquid Chromatography.” International Dairy Journal V3 (4–6): pp 379–387. Collin, J. C., D. le Bars, I. Ryba, and A. Bear. 1990. “Comparative measurements of the plasminogen-plasmin system of milk.” International Dairy Federation. Collins, Y.F., P.L.H. McSweeney, and M.G. Wilkinson. 2004. “Lipolysis and Catabolism of Fatty Acids in Cheese.” In Cheese Chemistry, Physics and Microbiology, Volume 1:373–389. Academic Press. Collins, Yvonne F., Paul L.H. McSweeney, and Martin G. Wilkinson. 2003 “Lipolysis and Free Fatty Acid Catabolism in Cheese: a Review of Current Knowledge.” International Dairy Journal V13 (11): pp. 841–866. Considine, T., A. Healy, A.L. Kelly, and P.L.H. McSweeney. 1999. “Proteolytic Specificity of Elastase on Bovine Β-casein.” Food Chemistry V66 (4) (September): pp. 463–470. Considine, T., Á. Healy, A.L. Kelly, and P.L.H. McSweeney. 2000. “Proteolytic Specificity of Elastase on Bovine Αs1-casein.” Food Chemistry V69 (1) (April): pp. 19–26. doi:10.1016/S0308-8146(99)00217-4. Considine, T., Á. Healy, A.L. Kelly, and P.L.H. McSweeney. 2004. “Hydrolysis of Bovine Caseins by Cathepsin B, a Cysteine Proteinase Indigenous to Milk.” International Dairy Journal V14 (2) (February): pp. 117–124. Crow, V.L., T. Coolbear, P.K. Gopal, F.G. Martley, L.L. McKay, and H. Riepe. 1995. “The Role of Autolysis of Lactic Acid Bacteria in the Ripening of Cheese.” International Dairy Journal V5 (8): pp. 855–875. Crow, Vaughan, Brian Curry, and Michelle Hayes. 2001. “The Ecology of Non-starter Lactic Acid Bacteria (NSLAB) and Their Use as Adjuncts in New Zealand Cheddar.” International Dairy Journal V11 (4-7) (4–7) (July 11): pp 275–283. Cserháti, Tibor. 2002. “Mass Spectrometric Detection in Chromatography. Trends and Perspectives.” Biomedical Chromatography V16 (5): pp. 303–310. 81 Curioni, P.M.G., and J.O. Bosset. 2002. “Key Odorants in Various Cheese Types as Determined by Gas Chromatography-olfactometry.” International Dairy Journal v12 (12): pp. 959–984. Datta, N., A. J. Elliott, M. L. Perkins, and H. C. Deeth. 2002. “Ultra-high-temperature (UHT) Treatment of Milk: Comparison of Direct and Indirect Modes of Heating” V57 (3): pp. 211–227. Deeth, H. C, and V. Touch. 2000a. “Methods for detecting lipase activity in milk and milk products.” Australian Journal of Dairy Technology V55 (3): pp. 153–168. Deeth, H. C., and V. Touch. 2000b. “Methods for detecting lipase activity in milk and milk products.” Australian Journal of Dairy Technology V55 (3): pp. 153–168. Dias, Benjamin, and Bart Weimer. 1998. “Conversion of Methionine to Thiols by Lactococci, Lactobacilli, and Brevibacteria.” Applied and Environmental Microbiology V64 (9) (September 1): pp. 3320–3326. Dimos, Tony. 1992. “A comparative study by GC/MS of the flavour volatiles produced during the maturation of full-fat and low-fat Cheddar cheese.” Dirinck, P, and A De Winne. 1999. “Flavour Characterisation and Classification of Cheeses by Gas Chromatographic–mass Spectrometric Profiling.” Journal of Chromatography A V847 (1-2) (June 25): pp. 203–208. Dobric, Nada, Gäetan K.Y. Limsowtin, Alan J. Hillier, Nicholas P.B. Dudman, and Barrie E. Davidson. 2000. “Identification and Characterization of a Cystathionine Β/γ-lyase from Lactococcus Lactis Ssp. Cremoris MG13631.” FEMS Microbiology Letters V182 (2): pp. 249–254. Drake, M.A., T.D. Boylston, K.D. Spence, and B.G. Swanson. 1996. “Chemical and Sensory Effects of a Lactobacillus Adjunct in Cheddar Cheese.” Food Research International V29 (3–4) (April): pp. 381–387. El Soda, M., S.A. Madkor, and P.S. Tong. 2000. “Adjunct Cultures: Recent Developments and Potential Significance to the Cheese Industry.” Journal of Dairy Science V83 (4) (April): pp. 609–619. El Soda, Morsi A. 1993. “The Role of Lactic Acid Bacteria in Accelerated Cheese Ripening.” FEMS Microbiology Reviews V12 (1–3) (September): pp. 239–251. El-Soda, M., S. A. Madkor, and P. S. Tong. 2000. “Evaluation of commercial adjuncts for use in cheese ripening: 4. Comparison between attenuated and not attenuated lactobacilli.” Milchwissenschaft V55 (5): pp. 260–263. Engel, Wolfgang, Wolfgang Bahr, and P. Schieberle. 1999. “Solvent Assisted Flavour Evaporation – a New and Versatile Technique for the Careful and Direct Isolation of Aroma Compounds from Complex Food Matrices.” European Food Research and Technology V209 (3): pp. 237–241. Engels, W.J.M., R. Dekker, C. de Jong, R. Neeter, and S. Visser. 1997. “A Comparative Study of Volatile Compounds in the Water-soluble Fraction of Various Types of Ripened Cheese.” International Dairy Journal V7 (4) (4) (April): pp. 255–263. Exterkate, Fred A.., Arno C.. Alting, and Charles J. Slangen. 1995. “Conversion of αs1Casein-(24-199)-Fragment and β-Casein Under Cheese Conditions by Chymosin and Starter Peptidases.” Systematic and Applied Microbiology V18 (1) (1): pp. 7–12. Fang, Yu, and Michael C. Qian. 2005. “Sensitive Quantification of Sulfur Compounds in Wine by Headspace Solid-phase Microextraction Technique.” Journal of Chromatography A V1080 (2) (July 8): pp. 177–185. Farkye, Nana Y., and Patrick F. Fox. 1990. “Observations on Plasmin Activity in Cheese.” Journal of Dairy Research V57 (03): pp. 413–418. 82 FDA. 2011. “Code of Federal Regulations Title 21.” http://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr. Fennema, Owen R., and Damodaran. 1996. “Amino Acids, Peptides, and Proteins.” In Food Chemistry, pp. 322–425. CRC Press. Fernández, Manuela, Tanoj K. Singh, and Patrick F. Fox. 1998. “Isolation and Identification of Peptides from the Diafiltration Permeate of the Water-Soluble Fraction of Cheddar Cheese.” Journal of Agricultural and Food Chemistry V46 (11) (November 1): pp. 4512–4517. Fernández, María, Wim van Doesburg, Ger A. M. Rutten, Joey D. Marugg, Arno C. Alting, Richard van Kranenburg, and Oscar P. Kuipers. 2000. “Molecular and Functional Analyses of themetC Gene of Lactococcus Lactis, Encoding Cystathionine β-Lyase.” Applied and Environmental Microbiology V66 (1) (January 1): pp. 42–48. Fernández-García, Estrella. 1996. “Use of Headspace Sampling in the Quantitative Analysis of Artisanal Spanish Cheese Aroma.” Journal of Agricultural and Food Chemistry V44 (7) (January 1): pp. 1833–1839. Ferreira, A.D., and B.C. Viljoen. 2003. “Yeasts as Adjunct Starters in Matured Cheddar Cheese.” International Journal of Food Microbiology V86 (1-2) (1–2) (September 1): pp. 131–140. Forde, Amanda, and Gerald F Fitzgerald. 2000. “Biotechnological Approaches to the Understanding and Improvement of Mature Cheese Flavour.” Current Opinion in Biotechnology V11 (5) (October 1): pp. 484–489. Fox, P. F., T.P. Guinee, T. M. Cogan, and P. L. H. McSweeney. 2000. “Enzymatic Coagulation of Milk.” In Fundamentals of Cheese Science, pp. 98135. Springer. Fox, P. F., Law J, P. L. H. McSweeney, and Wallace J.M. 1999. “Biochemistry of Cheese Ripening.” In Cheese: Chemistry, Physics and Microbiology, pp. 382–450. Springer. Fox, P. F., J. A. Lucey, and T. M. Cogan. 1990. “Glycolysis and Related Reactions During Cheese Manufacture and Ripening.” Critical Reviews in Food Science and Nutrition V29 (4) (January): pp. 237–253. Fox, P. F., and P. L. H. McSweeney. 1996. “Proteolysis in Cheese During Ripening.” Food Reviews International V12 (4): pp. 457–509. Fox, P. F., and P. L. H. McSweeney. 1998. “Milk Lipids.” In Dairy Chemistry and Biochemistry, pp. 67–141. Springer. Fox, P. F., J. M. Wallace, S. Morgan, C. M. Lynch, E. J. Niland, and J. Tobin. 1996. “Acceleration of Cheese Ripening.” Antonie Van Leeuwenhoek 70 (2): 271–297. Fox, P.F. 1989. “Proteolysis During Cheese Manufacture and Ripening.” Journal of Dairy Science V72 (6) (6) (June): pp. 1379–1400. Fox, P.F., and P.L.H. McSweeney. 2004. “Cheese: An Overview.” In Cheese Chemistry, Physics and Microbiology, Volume 1:1–18. Academic Press. Fox, P.F., and L. Stepaniak. 1993. “Enzymes in Cheese Technology.” International Dairy Journal V3 (4–6): pp. 509–530. Fox, PF, PLH McSweeney, and CM Lynch. 1998. “Significance of Non-starter Lactic Acid Bacteria in Cheddar Cheese.” Australian Journal of Dairy Technology V53 (2): pp. 7– 8. Gao, Song, Ed S. Mooberry, and James L. Steele. 1998. “Use of 13C Nuclear Magnetic Resonance and Gas Chromatography To Examine Methionine Catabolism by Lactococci.” Applied and Environmental Microbiology V64 (12) (December 1): pp. 4670–4675. 83 Gomez, Maria J., Sonia Garde, Pilar Gaya, Margarita Medina, and Manuel Nuñez. 1997. “Relationship Between Level of Hydrophobic Peptides and Bitterness in Cheese Made from Pasteurized and Raw Milk.” Journal of Dairy Research V64 (2): pp. 289–297. Grandison, Alistair S., and Graeme D. Ford. 1986. “Effects of Variations in Somatic Cell Count on the Rennet Coagulation Properties of Milk and on the Yield, Composition and Quality of Cheddar Cheese.” Journal of Dairy Research V53 (4): pp. 645–655. Grappin, Rémy, and Eric Beuvier. 1997. “Possible Implications of Milk Pasteurization on the Manufacture and Sensory Quality of Ripened Cheese.” International Dairy Journal V7 (12) (November 21): pp. 751–761. Green, Margaret L., and Donald J. Manning. 1982. “Development of Texture and Flavour in Cheese and Other Fermented Products.” Journal of Dairy Research V49 (04): pp. 737–748. Grieve, Paul A., and Barry J. Kitchen. 1985. “Proteolysis in Milk: The Significance of Proteinases Originating from Milk Leucocytes and a Comparison of the Action of Leucocyte, Bacterial and Natural Milk Proteinases on Casein.” Journal of Dairy Research 52 (1): 101–112. Griffiths, M. W. 1986. “Use of milk enzymes as indices of heat treatment.” Journal of Food Protection V49 (9): pp. 696–705. Gripon, Jean-Claude, V Monnet, G Lambert, and M. J. Desmazeaud. 1991. “Microbial enzymes in cheese ripening.” In Food enzymology, vol 1. Guinee, T P. 2004. “Salting and the Role of Salt in Cheese.” International Journal of Dairy Technology V57 (2-3): pp. 99–109. Habibi‐Najafi, Mohammad B., Byong H. Lee, and Barry Law. 1996. “Bitterness in Cheese: A Review.” Critical Reviews in Food Science and Nutrition V36 (5): pp. 397–411. Heegaard, Christian W., Lone K. Rasmussen, and Peter A. Andreasen. 1994. “The Plasminogen Activation System in Bovine Milk: Differential Localization of Tissuetype Plasminogen Activator and Urokinase in Milk Fractions Is Caused by Binding to Casein and Urokinase Receptor.” Biochimica Et Biophysica Acta (BBA) - Molecular Cell Research V1222 (1) (May 26): pp. 45–55. Hickey, D.K., K.N. Kilcawley, T.P. Beresford, and M.G. Wilkinson. 2007. “Lipolysis in Cheddar Cheese Made from Raw, Thermized, and Pasteurized Milks.” Journal of Dairy Science V90 (1) (January): pp. 47–56. Holland, Ross, and Tim Coolbear. 1996. “Purification of Tributyrin Esterase from Lactococcus Lactis Subsp. Cremoris E8.” Journal of Dairy Research V63 (1): pp. 131–140. Home, D.S., and J.M. Banks. 2004. “Rennet-induced Coagulation of Milk.” In Cheese Chemistry, Physics and Microbiology, Volume 1:pp. 47–70. Academic Press. Huffman, L. M.;, and Kristoffersen. 1984. “Role of lactose in Cheddar cheese manufacturing and ripening.” New Zealand Journal of Dairy Science and Technology V19 (2): pp. 151–162. Hurley, M.J, B.M O’Driscoll, A.L Kelly, and P.L.H McSweeney. 1999. “Novel Assay for the Determination of Residual Coagulant Activity in Cheese.” International Dairy Journal V9 (8) (August): pp 553–558. Hutton, J.T., and S. Patton. 1952. “The Origin of Sulfhydryl Groups in Milk Proteins and Their Contributions to ‘Cooked’ Flavor.” Journal of Dairy Science V35 (8) (August): pp. 699–705 84 Jolly, Ramesh C., and Frank V. Kosikowski. 1975. “Quantification of Lactones in Ripening Pasteurized Milk Blue Cheese Containing Added Microbial Lipases.” J. Agric. Food Chem. V23 (6): pp. 1175–1176. De Jong, Catrienus, and Herman T. Badings. 1990. “Determination of Free Fatty Acids in Milk and Cheese Procedures for Extraction, Clean up, and Capillary Gas Chromatographic Analysis.” Journal of High Resolution Chromatography V13 (2): pp. 94–98. K. Arfi, H.Spinnler, R. Tache, and P. bonnarme. 2002. “Production of Volatile Compounds by Cheese-ripening Yeasts: Requirement for a Methanethiol Donor for S-methyl Thioacetate Synthesis by Kluyveromyces Lactis.” Applied Microbiology and Biotechnology V58 (4): pp. 503–510. Knecht, E. 1999. “Lysomomal Cysteine Proteinases (2nd Edition).” Cell Biology International V23 (8) (August): pp 590–591. Laan, Harry, Saw Eng Tan, Paul Bruinenberg, Gaëtan Limsowtin, and Malcolm Broome. 1998. “Aminopeptidase Activities of Starter and Non-Starter Lactic Acid Bacteria Under Simulated Cheddar Cheese Ripening Conditions.” International Dairy Journal V8 (4) (April): pp. 267–274. Lane, C.N., and P.F. Fox. 1996. “Contribution of Starter and Adjunct Lactobacilli to Proteolysis in Cheddar Cheese During Ripening.” International Dairy Journal V6 (7) (July): pp. 715–728. Lane, Cáit N., and Patrick F. Fox. 1997. “Role of Starter Enzymes During Ripening of Cheddar Cheese Made from Pasteurized Milk Under Controlled Microbiological Conditions.” International Dairy Journal V7 (1) (January): pp. 55–63. Larsen, Lotte B., Connie Benfeldt, Lone K. Rasmussen, and Torben E. Petersen. 1996. “Bovine Milk Procathepsin D and Cathepsin D: Coagulation and Milk Protein Degradation.” Journal of Dairy Research V63 (1): pp 119–130. Lau, Kum Y., David M. Barbano, and Robert R. Rasmussen. 1990. “Influence of Pasteurization on Fat and Nitrogen Recoveries and Cheddar Cheese Yield.” Journal of Dairy Science V73 (3) (March): pp. 561–570. Lau, Kum Y., David M. Barbano, and Robert R. Rasmussen.1991. “Influence of Pasteurization of Milk on Protein Breakdown in Cheddar Cheese During Aging.” Journal of Dairy Science V74 (3) (March): pp. 727–740. Law, A. J. R., J. M. Banks, D. S. Horne, J. Leaver, and I. G. West. 1994. “Denaturation of the whey proteins in heated milk and their incorporation into Cheddar cheese.” Milchwissenschaft V49 (2): pp. 63–67. Lee, Won-Jae, Dattatreya S. Banavara, Joanne E. Hughes, Jason K. Christiansen, James L. Steele, Jeffery R. Broadbent, and Scott A. Rankin. 2007. “Role of Cystathionine βLyase in Catabolism of Amino Acids to Sulfur Volatiles by Genetic Variants of Lactobacillus Helveticus CNRZ 32.” Applied and Environmental Microbiology V73 (9) (May 1): pp. 3034–3039. Lin, J. C. C., and I. J. Jeon. 1985. “Headspace Gas Sampling/GC Method for the Quantitative Analysis of Volatile Compounds in Cheese.” Journal of Food Science V50 (3): pp. 843–844. Liu, Shao-Quan, Ross Holland, and Vaughan L Crow. 2001. “Purification and Properties of Intracellular Esterases from Streptococcus Thermophilus.” International Dairy Journal V11 (1–2) (January): pp. 27–35. 85 Lloyd, G. T., J. F. Horwood, and I. Barlow. 1980. “The effect of yoghurt culture YB on the flavour and maturation of Cheddar cheese.” Australian Journal of Dairy Technology V35 (4): pp. 137–139. Lynch, C.M., P.L.H. McSweeney, P.F. Fox, T.M. Cogan, and F.D. Drinan. 1996. “Manufacture of Cheddar Cheese with and Without Adjunct Lactobacilli Under Controlled Microbiological Conditions.” International Dairy Journal V6 (8–9) (August): pp. 851–867. Lynch, C.M., D.D. Muir, J.M. Banks, P.L.H. McSweeney, and P.F. Fox. 1999. “Influence of Adjunct Cultures of Lactobacillus Paracasei Ssp. Paracasei or Lactobacillus Plantarum on Cheddar Cheese Ripening.” Journal of Dairy Science V82 (8) (August): pp. 1618– 1628. Lynch, Joanna M., David M. Barbano, and J. Richard Fleming. 2002. “Determination of the Total Nitrogen Content of Hard, Semihard, and Processed Cheese by the Kjeldahl Method: Collaborative Study.” Journal of AOAC International V85 (2): pp. 445–455. Madkor, S.A., P.S. Tong, and M. El Soda. 2000. “Ripening of Cheddar Cheese with Added Attenuated Adjunct Cultures of Lactobacilli.” Journal of Dairy Science V83 (8) (August): pp. 1684–1691. Manning, D. J., Helen R. Chapman, and Zena D. Hosking. 1976. “The Production of Sulphur Compounds in Cheddar Cheese and Their Significance in Flavour Development.” Journal of Dairy Research V43 (02): pp. 313–320. Manning, Donald J., and Carolyn Moore. 1979. “Headspace Analysis of Hard Cheeses.” Journal of Dairy Research V46 (03): pp. 539–545. McSweeney, P.L.H., P.F. Fox, J.A. Lucey, K.N. Jordan, and T.M. Cogan. 1993. “Contribution of the Indigenous Microflora to the Maturation of Cheddar Cheese.” International Dairy Journal V3 (7): pp. 613–634. McSweeney, P.L.H., P.F. Fox, and N.F. Olson. 1995. “Proteolysis of Bovine Caseins by Cathepsin D: Preliminary Observations and Comparison with Chymosin.” International Dairy Journal V5 (4): pp. 321–336. McSweeney, Paul L. H., Sylvie Pochet, Patrick F. Fox, and Aine Healy. 1994. “Partial Identification of Peptides from the Water-Insoluble Fraction of Cheddar Cheese.” Journal of Dairy Research V61 (4): pp. 587–590. McSweeney, Paul L.H., and Maria José Sousa. 2000. “Biochemical Pathways for the Production of Flavour Compounds in Cheeses During Ripening: A Review.” Le Lait V80 (3): pp. 293–324. Metwalli, Ali A.M, Harmen H.J de Jongh, and Martinus A.J.S van Boekel. 1998. “Heat Inactivation of Bovine Plasmin.” International Dairy Journal V8 (1) (January 1): pp. 47–56. Milo, C., and G. A. Reineccius. 1997. “Identification and Quantification of Potent Odorants in Regular-Fat and Low-Fat Mild Cheddar Cheese.” Journal of Agricultural and Food Chemistry V45 (9) (September 1): pp. 3590–3594. Mistry, V.V., and J.-L. Maubois. 2004. “Application of Membrane Separation Technology to Cheese Production.” In Cheese Chemistry, Physics and Microbiology, Volume 1:pp. 261–VII. Academic Press. Mistry, V.V., L.E. Metzger, and J.L. Maubois. 1996. “Use of Ultrafiltered Sweet Buttermilk in the Manufacture of Reduced Fat Cheddar Cheese.” Journal of Dairy Science V79 (7) (July): pp. 1137–1145. Moio, Luigi, Dominique Langlois, Patrick Etievant, and Francesco Addeo. 1993. “Powerful Odorants in Bovine, Ovine, Caprine and Water Buffalo Milk Determined by Means of 86 Gas Chromatography–olfactometry.” Journal of Dairy Research V60 (2): pp. 215– 222. Molimard, P., and H.E. Spinnler. 1996. “Review: Compounds Involved in the Flavor of Surface Mold-Ripened Cheeses: Origins and Properties.” Journal of Dairy Science V79 (2) (February): pp. 169–184. Mooney, J.S, P.F Fox, A Healy, and J Leaver. 1998. “Identification of the Principal Waterinsoluble Peptides in Cheddar Cheese.” International Dairy Journal V8 (9) (September): pp. 813–818. Muir, D.D., J.M. Banks, and E.A. Hunter. 1996. “Sensory Properties of Cheddar Cheese: Effect of Starter Type and Adjunct.” International Dairy Journal V6 (4) (April): pp. 407–423. Mulvihill, Donald M., and Patrick F. Fox. 1979. “Proteolytic Specificity of Chymosin on Bovine Αs1-Casein.” Journal of Dairy Research V46 (4): pp. 641–651. Naughton, M. A., and F. Sanger. 1961. “Purification and Specificity of Pancreatic Elastase.” Biochemical Journal V78 (1): pp. 156–163. Niven, C. F., L. G. Buettner, and James B. Evans. 1954. “Thermal Tolerance Studies on the Heterofermentative Lactobacilli That Cause Greening of Cured Meat Products.” Applied Microbiology V2 (1) (January): pp. 26–29. O’Keeffe, Arthur M., Patrick F. Fox, and Charles Daly. 1978. “Proteolysis in Cheddar Cheese: Role of Coagulant and Starter Bacteria.” Journal of Dairy Research V45 (3): pp. 465– 477. O’Keeffe, R. B., P. F. Fox, and C. Daly. 1976. “Contribution of Rennet and Starter Proteases to Proteolysis in Cheddar Cheese.” Journal of Dairy Research 43 (01): 97–107. O’Shea, B.A., T. Uniacke-Lowe, and P.F. Fox. 1996. “Objective Assessment of Cheddar Cheese Quality.” International Dairy Journal V6 (11-12) (November): pp. 1135– 1147. Olivecrona, T., S. Vilaró, G. Bengtsson-Olivecrona, and P. F Fox. 1992. “Indigenous enzyme in milk. II. Lipases in milk.” In Advanced Dairy Chemistry 1, 292–310. Payne, Martin J., and Rohan G. Kroll. 1991. “Methods for the Separation and Concentration of Bacteria from Foods.” Trends in Food Science & Technology V2 (0): pp. 315–319. Peterson, S.D., and R.T. Marshall. 1990. “Nonstarter Lactobacilli in Cheddar Cheese: A Review.” Journal of Dairy Science V73 (6) (June): pp. 1395–1410. Piraino, Paolo, Eugenio Parente, and Paul L. H. McSweeney. 2004. “Processing of Chromatographic Data for Chemometric Analysis of Peptide Profiles from Cheese Extracts: A Novel Approach.” Journal of Agricultural and Food Chemistry 52 (23) (November 1): 6904–6911. Precetti, A.S., M.P. Oria, and S.S. Nielsen. 1997. “Presence in Bovine Milk of Two Protease Inhibitors of the Plasmin System.” Journal of Dairy Science V80 (8) (August): pp. 1490–1496. Preininger, Martin, and Werner Grosch. 1994. “Evaluation of Key Odorants of the Neutral Volatiles of Emmentaler Cheese by the Calculation of Odour Activity Values.” LWT Food Science and Technology V27 (3) (June): pp. 237–244. Pripp, A.H, Shakeel-Ur-Rehman, P.L.H McSweeney, and P.F Fox. 1999. “Multivariate Statistical Analysis of Peptide Profiles and Free Amino Acids to Evaluate Effects of Single-strain Starters on Proteolysis in Miniature Cheddar-type Cheeses.” International Dairy Journal V9 (7) (July): pp. 473–479. 87 Pripp, Are Hugo, Leszek Stepaniak, and Terje Sørhaug. 2000. “Chemometrical Analysis of Proteolytic Profiles During Cheese Ripening.” International Dairy Journal V10 (4): pp. 249–253.. Puchades, R., L. Lemieux, and R.e. Simard. 1989. “Evolution of Free Amino Acids During the Ripening of Cheddar Cheese Containing Added Lactobacilli Strains.” Journal of Food Science V54 (4): pp. 885–888. Qian, M., and G. Reineccius. 2002. “Identification of Aroma Compounds in ParmigianoReggiano Cheese by Gas Chromatography/Olfactometry.” Journal of Dairy Science V85 (6) (June): pp. 1362–1369. Rehman, Shakeel-Ur, J.M Banks, E.Y Brechany, D.D Muir, P.L.H McSweeney, and P.F Fox. 2000. “Influence of Ripening Temperature on the Volatiles Profile and Flavour of Cheddar Cheese Made from Raw or Pasteurised Milk.” International Dairy Journal V10 (1–2) (January): pp. 55–65. Richardson, B. C., L. K. Creamer, K. N. Pearce, and R. E. Munford. 1974. “Comparative Micelle Structure: II. Structure and Composition of Casein Micelles in Ovine and Caprine Milk as Compared with Those in Bovine Milk.” Journal of Dairy Research V41 (2): pp. 239–247. Roy, Denis, Isabelle Mainville, and Francine Mondou. 1997. “Selective Enumeration and Survival of Bifidobacteria in Fresh Cheese.” International Dairy Journal V7 (12) (November 21): pp. 785–793. Sablé, Sophie, and Gilles Cottenceau. 1999. “Current Knowledge of Soft Cheeses Flavor and Related Compounds.” J. Agric. Food Chem. V47 (12): pp. 4825–4836. Seefeldt, K.E., and B.C. Weimer. 2000. “Diversity of Sulfur Compound Production in Lactic Acid Bacteria.” Journal of Dairy Science V83 (12) (December): pp. 2740–2746. Shakeel-Ur-Rehman, J. M Banks, P. L. H Mcsweeney, and P. F Fox. 2000. “Effect of ripening temperature on the growth and significance of non-starter lactic acid bacteria in Cheddar cheese made from raw or pasteurised milk.” International dairy journal V10 (1-2): pp. 45–53. Singh, T. K, M. A Drake, and K. R Cadwallader. 2003. “Flavor of Cheddar Cheese: A Chemical and Sensory Perspective.” Comprehensive Reviews in Food Science and Food Safety V2 (4) (October 1): pp. 166–189. Singh, T.K., P.F. Fox, P. Højrup, and A. Healy. 1994. “A Scheme for the Fractionation of Cheese Nitrogen and Identification of Principal Peptides.” International Dairy Journal V4 (2): pp. 111–122. Singh, Tanoj K., Patrick F. Fox, and Áine Healy. 1995. “Water-Soluble Peptides in Cheddar Cheese: Isolation and Identification of Peptides in the Diafiltration Retentate of the Water-Soluble Fraction.” Journal of Dairy Research V62 (4): pp. 629–640. Singh, Tanoj K., Patrick F. Fox, and Áine Healy 1997. “Isolation and Identification of Further Peptides in the Diafiltration Retentate of the Water-soluble Fraction of Cheddar Cheese.” Journal of Dairy Research V64 (3): pp. 433–443. Smacchi, Emanuele, and Marco Gobbetti. 1998. “Purification and Characterization of Cystathionine Γ-lyase from Lactobacillus Fermentum DT41.” FEMS Microbiology Letters V166 (2): pp. 197–202. Smit, Gerrit, Bart A. Smit, and Wim J.M. Engels. 2005. “Flavour Formation by Lactic Acid Bacteria and Biochemical Flavour Profiling of Cheese Products.” FEMS Microbiology Reviews V29 (3): pp. 591–610. 88 Smith, A. M., and S. Nakai. 1990. “Classification of cheese varieties by multivariate analysis of HPLC profiles.” Canadian Institute of Food Science and Technology Journal V23 (1): pp. 53–58. Sousa, M.J, Y Ardö, and P.L.H McSweeney. 2001. “Advances in the Study of Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4-7) (July 11): pp. 327– 345. Stuart, Mark R., Lan Szu Chou, and Bart C. Weimer. 1999. “Influence of Carbohydrate Starvation and Arginine on Culturability and Amino Acid Utilization of Lactococcus Lactis Subsp. Lactis.” Applied and Environmental Microbiology V65 (2) (February 1): pp. 665–673. Subramanian, A., W.J. Harper, and L.E. Rodriguez-Saona. 2009. “Cheddar Cheese Classification Based on Flavor Quality Using a Novel Extraction Method and Fourier Transform Infrared Spectroscopy.” Journal of Dairy Science V92 (1) (1) (January): pp. 87–94. Suriyaphan, Orasa, MaryAnne Drake, X. Q. Chen, and Keith R. Cadwallader. 2001. “Characteristic Aroma Components of British Farmhouse Cheddar Cheese.” Journal of Agricultural and Food Chemistry V49 (3) (March 1): pp. 1382–1387. Tammam, J. D, A. G Williams, J. Noble, D. Lloyd, J. D Tammam, A. G Williams, J. Noble, and D. Lloyd. 2000. “Amino acid fermentation in non‐starter Lactobacillus spp. isolated from Cheddar cheese, Amino acid fermentation in non‐starter Lactobacillus spp. isolated from Cheddar cheese.” Letters in Applied Microbiology, Letters in Applied Microbiology V30 (5) (May 1): pp. 370–374. Tanaka, Hidehiko, Nobuyoshi Esaki, and Kenji Soda. 1985. “A Versatile Bacterial Enzyme: L-methionine Γ-lyase.” Enzyme and Microbial Technology V7 (11) (November): pp. 530–537. Thomsen, D., and H. Stapelfeldt. 1990. “Yield and quality of Danbo 45+ cheese from milk heat treated at temperatures between 72 and 95°C.” (44): 39. Tsakalidou, E., and G. Kalantzopoulos. 1992. “Purification and Partial Characterization of an Esterase from Lactococcus Lactis Ssp Lactis Strain ACA-DC 127.” Le Lait V72 (6): pp. 533–543. Turner, K. W, and T. D Thomas. 1980. “Lactose fermentation in Cheddar cheese and the effect of salt.” New Zealand Journal of Dairy Science and Technology V15 (3): pp. 265–276. Upadhyay, V.K., P.L.H. McSweeney, A.A.A. Magboul, and P.F. Fox. 2004. “Proteolysis in Cheese During Ripening.” In Cheese Chemistry, Physics and Microbiology, Volume 1:pp. 391–433. Academic Press. Urbach, G. 1995. “Contribution of Lactic Acid Bacteria to Flavour Compound Formation in Dairy Products.” International Dairy Journal V5 (8): pp. 877–903. Urbach, Gerda. 1993. “Relations Between Cheese Flavour and Chemical Composition.” International Dairy Journal V3 (4–6): pp. 389–422. USDA. 2011. Dairy Products Annual Sumary. Vazquez-Landaverde, P.A., J.A. Torres, and M.C. Qian. 2006. “Quantification of Trace Volatile Sulfur Compounds in Milk by Solid-Phase Microextraction and Gas Chromatography–Pulsed Flame Photometric Detection.” Journal of Dairy Science V89 (8) (August): pp. 2919–2927. Verdi, Robert J., and David M. Barbano. 1991. “Properties of Proteases from Milk Somatic Cells and Blood Leukocytes.” Journal of Dairy Science V74 (7) (July): pp. 2077– 2081. 89 Visser, S. ;Hup. 1983. “Bitter flavour in cheese, 2: model studies on the formation and degradation of bitter peptides by proteolytic enzymes from calf rennet, starter cells and starter cell fractions.” Netherlands Milk and Dairy Journal V37 (3): pp. 169–180. Visser, Servaas. 1993. “Proteolytic Enzymes and Their Relation to Cheese Ripening and Flavor: An Overview.” Journal of Dairy Science V76 (1) (January): pp. 329–350. Voigt, Daniela D., François Chevalier, John A. Donaghy, Margaret F. Patterson, Michael C. Qian, and Alan L. Kelly. 2012. “Effect of High-pressure Treatment of Milk for Cheese Manufacture on Proteolysis, Lipolysis, Texture and Functionality of Cheddar Cheese During Ripening.” Innovative Food Science & Emerging Technologies V13 (0) (January): pp. 23–30. Wallace J.M., and Fox P.F. 1997. “Effect of Adding Free Amino Acids to Cheddar Cheese Curd on Proteolysis, Flavour and Texture Development.” International Dairy Journal V7 (2): pp. 157–167. Walstra, P., T. J Geurts, A. Noomen, A. Jellema, and M. A. J. S. van Boekel. 1999. “Cheese Varieties.” In Dairy technology: principles of milk properties and processes, pp 651– 708. Marcel Dekker. Walstra, Pieter, Jan T. M. Wouters, and Tom J. Geurts. 2006. Dairy Science And Technology. CRC Press. Weimer, B. C. 2007. Improving the flavour of cheese. CRC Press. http://www.cabdirect.org/abstracts/20083142333.html. Weimer, Bart, Kimberly Seefeldt, and Benjamin Dias. 1999. “Sulfur Metabolism in Bacteria Associated with Cheese.” Antonie Van Leeuwenhoek 76 (1): 247–261. Wijesundera, C., and L. Drury. “Role of milk fat in production of cheddar cheese flavour using a fat-substituted cheese model.” Australian journal of dairy technology V54 (1): pp. 28–35. Wilkinson, Martin G., Timothy P. Guinee, Daniel M. O’Callaghan, and Patrick F. Fox. 1994. “Autolysis and Proteolysis in Different Strains of Starter Bacteria During Cheddar Cheese Ripening.” Journal of Dairy Research V61 (2): pp. 249–262. De Wit, M., G. Osthoff, B.C. Viljoen, and A. Hugo. 2005. “A Comparative Study of Lipolysis and Proteolysis in Cheddar Cheese and Yeast-inoculated Cheddar Cheeses During Ripening.” Enzyme and Microbial Technology V37 (6) (November 1): pp. 606–616. de Wit, Rudy, and Hans Nieuwenhuijse. 2008. “Kinetic Modelling of the Formation of Sulphur-containing Flavour Components During Heat-treatment of Milk.” International Dairy Journal V18 (5) (May): pp. 539–547. Wong, N. P, R. Ellis, and D. E LaCroix. “Quantitative determination of lactones in Cheddar cheese.” Journal of Dairy Science V58 (10): pp. 1437–1441. Yvon, Mireille, Claire Chabanet, and Jean-Pierre Pélissier. 1989. “Solubility of Peptides in Trichloroacetic Acid (TCA) Solutions Hypothesis on the Precipitation Mechanism.” International Journal of Peptide and Protein Research V34 (3): pp. 166–176. Yvon, Mireille, and Liesbeth Rijnen. 2001. “Cheese Flavour Formation by Amino Acid Catabolism.” International Dairy Journal V11 (4–7) (July 11): pp. 185–201. Zehentbauer, Gerhard, and G. A. Reineccius. 2002. “Determination of Key Aroma Components of Cheddar Cheese Using Dynamic Headspace Dilution Assay.” Flavour and Fragrance Journal V17 (4): pp. 300–305. 90 CHAPTER 3 COMPARISON OF RIPENING CHARACTERISTICS OF CHEDDAR CHEESE FROM THE SAME COMPANY MANUFACTURED IN DIFFERENT PRODUCTION PLANTS Lemus Muñoz, Freddy Mauricio, Qian Michael Department of Food Science and Technology, Oregon State University Corvallis Oregon 97331 91 ABSTRACT The difference in ripening patterns between samples from the same manufacturer but produced in different plants (location) was studied. Proteolysis was investigated by a fractionation scheme, resulting in an insoluble fraction analyzed by urea polyacrylamide gel electrophoresis (Urea-PAGE), and a soluble fraction which was further investigated through water soluble nitrogen (WSN), trichloroacetic acid soluble nitrogen (TCA-SN) and phosphotungstic acid soluble nitrogen (PTA-SN) analyzed by total Kjeldahl nitrogen content (TKN). Reversed phase high performance liquid chromatography (RP-HPLC) was used to study the peptide profile of the water soluble fraction. Lipolysis was studied by levels of individual free fatty acids determined through gas chromatography-flame ionization detection (GC-FID) after isolation employing solid phase extraction (SPE). Volatile sulfur compounds were studied using head space solid phase micro-extraction (SPME) coupled with gas chromatography-pulsed flame photometric detection (PFPD). The Urea-PAGE method was able to differentiate samples according their age, but it could not discriminate samples regarding their origin. Nonetheless, measurements of total Kjeldahl Nitrogen (TKN) of the WSN, TCA-SN, and PTA-SN fractions, and the principal component analysis of the RP-HPLC peptide profile of the WSN fraction, revealed differences in the rate and pattern of proteolysis for the samples. Levels of total nitrogen for the WSN, TCA and PTA fractions increased as cheese aged and were lower for cheeses made in the production plant CRP (B). From the RP-HPLC analysis data it was developed a PCA model with 3 principal components that accounted for the 80.6% of the variability. This model discriminates the samples according age and quality, and suggests that the cheese samples from TCCA (A) plant undergo more or faster proteolysis. FFA profiles reveal significant difference in the extension of lipolysis, which can be mostly related to variations in manufacturing practices and indicates that good cheese samples had faster lipolysis. The Volatile Sulfur Compounds (VSC) analysis showed that cheeses made in the production plant A developed higher concentrations of H2S, DMS and MeSH, suggesting slower catabolism of sulfur containing amino acids in cheese made in plant CRP; however H2S did not exhibit a continuous development as the cheese aged 92 INTRODUCTION Essentially, cheddar cheese is a casein matrix that contains a balanced mixture of moisture, peptides, amino acids, free fatty acids, lipids, minerals, microflora and other compounds; whose complex flavor profile is determined by: 1) variation in the composition and quality of milk (especially in seasonal dairying countries, where variations of protein and fat levels, and lactose concentration occurs during the year) and other raw materials; 2) manufacturing practices; and 3) the extent of biochemical events such as proteolysis, lipolysis, and glycolysis occurring during ripening (P. F. Fox et al. 1999). This makes cheese manufacturing with consistent quality and uniform sensory properties, a really challenging labor. However, it should be kept in mind that it is expectable that cheeses produced in different regions, or in different production plants belonging to the same company that follow “identical” manufacturing procedure, might have unique and distinctive flavor attributes. The quality of Cheddar cheese is associated to maturity, flavor intensity and texture, and it is usually assessed by expensive panels of trained people, following a cheese-grader set of criteria based and dependent on the presence or absence of defects, which besides of being a time consuming practice, it can result in ambiguous and subjective assessments, consequence of the different customer and manufacturers preferences from region to region. Thus, in order to be competitive in an industry producing about 3.3 billion pounds of cheddar cheese per year (USDA 2011), cheese makers require more reliable standards for classifying and grading cheese, such as quantitative measurements of compositional parameters involving instrumental methods and chemical analysis. Hence, an accurate evaluation of flavor quality will improve the relationship between the final sensory character of the product and the factors to control it during curd manufacture and ripening. Because of the capacity to simultaneously monitor many key compounds, the use of instrumental methods such as gas chromatography (GC), high performance liquid chromatography (HPLC), and electrophoresis, have allowed the identification of hundreds of compounds contributing to the characteristic flavor of cheese (Paul L.H. McSweeney and Sousa 2000), which are mainly separated between sapid and aromatic compounds (T. K Singh, Drake, and Cadwallader 2003). Nonetheless, an adequate correlation of existing sensory 93 criteria and the chosen measurements is required for predicting, classifying and reproducing products with equivalent quality. Thus, based on the fact that the volatile fraction contributes to the aroma while the water soluble fraction play a role in the taste (Aston and Creamer 1986), the discrimination of samples regarding their origin can be done using parameters such as the profile and abundance of headspace volatiles (Subramanian, Harper, and RodriguezSaona 2009), and degree of proteolysis and lipolysis as indices of ripening (P. F. Fox 1989; Y.F. Collins, McSweeney, and Wilkinson 2004). The use of proteolysis and lipolysis implies identification and quantification of fatty acids, amino acids, peptides and soluble nitrogen among other measurements. Indeed, the analysis by HPLC have made possible the separation of bitter peptides and the estimation of the ratio of hydrophobic to hydrophilic peptides, the differentiation between varieties, between young and mature cheese, and between cheese made from raw and thermally treated milk (Smith and Nakai 1990; Lau, Barbano, and Rasmussen 1991), which is complemented by the characterization of low molecular mass pepetides through electrophoretic methods (Sousa, Ardö, and McSweeney 2001) and the amino acid composition analysis by the N-terminals, resulting in the characterization of degradation products (Allan J. Cliffe, Marks, and Mulholland 1993; A.J. Cliffe, Revell, and Law 1989). In a similar way, gas chromatography (GC) has allowed the characterization and quantification of more than a hundred volatile flavor compounds in cheese, which coupled with olfactometry analysis and headspace extraction techniques such as aroma extract dilution analysis (AEDA ) and solid phase micro extraction (SPME), have made possible the detection and discrimination of the potent odorants in different varieties (Preininger and Grosch 1994; Fernández-García 1996; Qian and Reineccius 2002). Nonetheless, in order to validate the degree of ripening estimated through chromatography; experimental controls and/or statistical analysis are required to interpret data, and to establish a accurate correlation between objective methods, sensory analysis and the classification according to a particular variable. This makes possible documenting differences in attributes as consequence of cheese origin, resulting in a better understanding of the process variables for standardizing the product between different locations of production regions. 94 The objective of this study was to apply objective measurements to characterize the differences found in cheddar cheese made in different production plants MATERIALS AND METHODS CHEESE SAMPLES Cheeses were manufactured by Tillamok county creamery. Cheeses were made according with standard protocols in Boardman cheese factory and Tillamok cheese factory. Three blocks of cheese made from each processing plant were selected randomly from three consecutive manufacturing days. All cheeses were aged using the same conditions at manufacturer’s facility, Every 2 months a 2 lb portion was sampled from each block and sent to the lab, where samples are stored at (-37C) to stop ageing process until analysis is completed FREE FATTY ACIDS ANALYSIS Chemicals Pentanoic acid, heptanoic acid, nonanoic acid, undecanoic acid, and heptadecanoic acid were used as internal standards, they were purchased from Eastman (Rochester, N.Y., U.S.A). Butanoic acid, hexanoic acid, octanoic acid, decanoic acid, dodecanoic acid, tetradecanoic acid, 9-tetradecanoic acid, hexadecanoic acid, 9-hexadecanoic acid, octadecanoic acid, 9octadecanoic acid, 9,12-octadecanoic acid and 6,9,12 octadecanoic acid were used for the standard stock solution, and were obtained from Aldrich Chemical Co. Inc (Milwaukee, Wisconsin, U.S.A). Heptane, Isopropanol, Sulfuric acid, anhydrous sodium sulfate, chloroform, formic acid and diethyl ether were obtained from Fisher. Extraction From each 2lb block of cheese, 100 grams were wrapped in alumina foil, frozen with liquid nitrogen during 6 minutes, and then grinded for 30 seconds to obtain a fine powder. Six grams of this previously freeze-ground cheese, 1 ml of 2N sulphuric acid and 1 ml of internal standard solution (C5:0, C7:0, C9:0, C11:0 and C17:0 in 1:1 hetpane-isopropanol) were mixed with 7 grams of anhydrous sodium sulfate and 20 ml of 1:1 diethyl ether- heptane in a 40 ml amber vial using a sonicator and manual agitation. During sonication, the salt-slurry solution is initially exposed for 15 minutes, after which each vial is shake vigorously to continue with a 95 second sonication period of 20 minutes. With a glass-Pasteur pipette, the sample extract (solvent) is transferred to an AccuBOND amino cartridge (Agilent Technologies) conditioned previously with 10 ml of heptane. After the addition of the sample, the column is washed with 5 ml of 2:1 Chloroform-Isopropanol to remove non volatile triglycerides and phospholipids using a manifold vacuum chamber. Once the washing step is complete, free fatty acids are eluted with 5ml of 2% formic acid in diethyl ether, collected in a 20 ml vial and stored in the freezer until GC analysis. Chromatography The analysis was performed using a Hewlett-Packard 5890 gas chromatograph equipped with a flame ionization detector (FID). Samples were analyzed on a DB-FFAP column (15m x 0.53mm ID, 1 m film thickness; Supelco Wax10, Supelco U.S.A). Injector and detector temperatures were 250C. Nitrogen was used as carrier gas at a flow rate of 15 ml per minute at a split ratio of 1 to 1. The oven temperature was programmed for a 2 minutes hold at 60C, raised to 230C at a rate of 8C per minute with a hold of 20 minutes at 230C. Quantitative analysis The levels of free fatty acids concentrations were calculated based on individual peak area from GC-FID response in comparison to the internal standard peak area, and by using standard calibration curve of individual free fatty acid using Peak Simple software (SRI instruments, Torrance, CA). Each experimental value corresponds to the average of the 3 extraction replicates. VOLATILE SULFUR COMPOUNDS (VSC’S) Chemicals Dimethyl sulfide (DMS) was purchased from TCI America (Portland, OR, U.S.A.); gaseous methanethiol (MeSH) was obtained from Aldrich Chemical Co. Inc (Milwaukee, Wisconsin, U.S.A), and a solution was prepared by bubbling the gas into cold methanol; a H2S solution was prepared by dissolving Na2S.9 H2O (Sigma Co) in acidic water stabilized with citric acid (pH 3). 96 Extraction From each 2lb block of cheese, 100 grams were wrapped in alumina foil, frozen with liquid nitrogen during 6 minutes, and then grinded for 30 seconds to obtain a fine powder. Then one gram of this freshly prepared powder is added to a 20ml vial (formerly flushed with argon), followed by the addition of 4 ml of 1M citric acid and 20 l of the internal standard solution. After addition of sample vials were immediately sealed with screw caps with teflon-lined silicone septa. The vials used in this study were previously deactivated with DMTCS 5% solution in toluene, toluene, methanol and distillate water. The volatile sulfur compounds were extracted with an 85 m Carbox-PDMS fiber (Supelco, Bellefonte, PA, U.S.A.). Prior to use, the fiber was conditioned at 300 C for 90 minutes. The fiber was then placed into a SPME adapter of a CombiPAL autosampler (CTC analytics AG, Zwingen, Switzerland) Fitted with a vial heater/agitator. Samples were pre-equilibrated at 500 RPM at 40C for 5 minutes, and the extraction of VSC’s was done at 250 RPM at 40C for 25 minutes. The desorption time was 5 minutes and 30 seconds. Chromatography The analysis was performed using a Varian CP-3800 gas chromatograph (Varian, Walnut Creek, CA, U.S.A.) equipped with a pulsed flame photometric detector (PFPD). The separation of analytes was made using a DB-FFAP fused silica capillary column (30m, 0.32 mm ID and 1 m film thickness; Agilent, Palo Alto, CA, USA) and nitrogen as carrier gas at constant flow at 2 ml per minute. The injector temperature was 300 C and it was in the splitless mode. The oven temperature was programmed for a 3 minutes hold at 35C, raised to 150C at a rate of 10C per minute, held for 5 minute, and then heated to 220C at a rate of 20C per minute with a final hold of 3 minutes. The PFPD was held at 300 C and 450 V with the following flow rates: Air 1 at 17 ml per min, H2 at 14 ml per min, and Air 2 at 10 ml/min. The detector response signal was integrated using the software Star Workstation 6.2, Varian) Quantitative analysis Matrix effect 97 In order to retain the matrix effect during the construction of the calibration curves, cheese powder from the “youngest sample” is used. It is de-volatilized by exposure to room conditions in a hood for 2 hours. Then 1 gram of powder is added to 4 ml of 1M citric acid in a 20 ml vial and exposed to a 50C water bath for 30 mins, prior to the addition of standards and internal standard solutions. Sulfur standards and internal standard preparation Two internal standards were used for the quantification of VSC’s: ethyl methyl sulfide (EMS) for H2S, MeSH and DMS, and isopropyl disulfide (IsoProDS) for DMDS and DMTS. The concentration of the internal standard solution was 500 ppm EMS and 500 ppm IsoProDS in methanol. Calibration curves were constructed by spiking cheese samples with a range of known concentrations of H2S, MeSH and DMS. Hydrogen sulfide (H2S) was prepared by dissolving Na2S.9 H2O in acidic water (pH = 3). Different concentrations of sodium sulfide solutions were made, and the concentrations of H2S were calculated based on the amounts of salt added to the matrix. A standard solution of 100 ppm of DMS was individually prepared in cooled methanol (-15C), and dilutions were made with cooled methanol at the same temperature. The mesh standard was prepared as following: 1) newly deactivated, recently flushed with argon, and cooled vials were used; 2) The original standard solution was made by bubbling pure MeSH into cooled methanol; 3) Dilutions were prepared by taking aliquots from the original solution contained in a sealed vial, through the teflon-lined silicone septa by using a syringe. And then injecting the aliquots into new sealed vials containing proportional amount of cooled methanol through the septa; 4) 1 gr of devolatilized cheese is added to a recently flushed vial (argon was used), which is immediately flushed again; 5) simultaneous argon flushing and addition of 4ml of “free” dissolved oxygen-1M citric acid solution and quick sealing of the vials; 6) Addition of 20 l of internal standard and MeSH standard through septa. The identification of target compounds was made by comparing retention times with those of pure standards. Ratios of the square root of the standard area to the corresponding square root of the internal standard area were plotted Vs concentration ratios to determine the relation between the response and concentration for the unknowns. Triplicate analysis was performed for all samples 98 PROTEOLYSIS Chemicals Sulfuric acid was purchased from Fisher Scientific International Inc. (Pittsburgh, PA, U.S.A.); Trichloroacetic acid was purchased from Alfa Aesar (Royston, UK); And phosphotungstic acid was obtained from Aldrich Chemical Co. Inc (Milwaukee, Wis, U.S.A) Sample preparation and fractionation From each 2lb block of cheese, 60 grams are blended with 120 ml of distilled water pre-heated to 55C. The mixture is blended for 5 minutes and the homogenate is incubated at 55C for 1 hour. Then the pH is adjusted to 4.6 with 1M HCl and the mixture is centrifuged at 3000g for 30 minutes at 4C. Suspension and supernatant were filtered thoroughly 3 times through glass wool. The filtrate was safe at -20C for macro blog digestion method analysis, and RP-HPLC analysis. The insoluble pellet was frozen at -20C for further Urea-PAGE gel electrophoresis analysis. The trichloroacetic acid soluble nitrogen fraction (TCA-SN) was prepared by the addition of 25 ml of pH 4.6 soluble fraction (WSN) to 25 ml of 24% trichloroacetic acid solution. Then the mixture is equilibrated for 2 hours at room temperature and filtered through filter paper Whatman No 40 before macro blog digestion method analysis. For the phosphotungstic acid soluble nitrogen fraction (PTA-SN), 10 ml of WSN are added to 7 ml of 3.95 M H2SO4 and 3 ml of 33% phosphotungstic acid solution. Then the mixture is equilibrated overnight at 4C and filtered through filter paper Whatman No 40 before macro blog digestion method analysis. Duplicate analysis was performed for all samples. Macro blog digestion (Kjeldahl Digestion) From the fractions collected an aliquot (2 ml for the Water soluble fraction, 1ml for TCA-SN and 1 ml for PTA-SN) is added into a 70 ml Kjeldahl Digestion flask with 10 ml of H2SO4 and the catalyst pellet containing 0,075 and 1,5 grams of mercuric oxide and potassium sulfate respectively. The mixture is warmed to 150 C and hold for 1 hour, then heated to 250 C and 99 hold for 1 hour, and finally heated to 350 C and hold for 2 hours. After digestion the sample is cooled down overnight to room temperature, and diluted with distillate water to 70 ml, followed by a gentile agitation. Then a 5 ml aliquot is used to determine the nitrogen content by a rapid flow analyzer FOSS II. Reversed phase High performance liquid chromatography analysis The RP-HPLC analysis was performed using a Shimadzu 6 series liquid chromatograph (Shimadzu scientific instruments, Kyoto Japan), consisting of an autosampler, 2 pumps, a multi-wavelength spectrophotometer and a controller unit. It was used a nucleosil RP-8 analytical column (250x 4mm, 5 m particle size, 300 A pore size) and a guard column (4.6 x10 mm) from waters (Milford, MA, U.S.A.). The mobile phase consists of solvent A (0.1% TFA in deionized and vacuum filtered water) and solvent B (0.1% TFA in acetonitrile). The elution was monitored at 214nm. The following gradient elution was performed: 1) 100% solvent A for 5 minutes followed by a linear gradient to 55% solvent B (v/v); 2) elution at 55% solvent B for 6 minutes followed by a linear gradient to 60%; 3) elution at 60% solvent B for 3 minutes; 4) The column is washed using 95% solvent B during 5 minutes; 5) the column is equilibrated using 100% solvent A during 10 minutes. The sample (WSN fraction) was dissolved in solvent A (10 mg per ml) and then micro-centrifuged at 14000 RPM for 10 minutes. An aliquot of 40 l from the extract was injected to a flow rate of 0.75 ml per min. Electrophoresis Samples of the water-insoluble nitrogen fraction were dry frozen prior to analysis. Samples were dissolved in a buffer (0.75 g tris, (hydroxymethyl) methylamine, 49 gr urea and 0.4 ml concentrated HCl, 0.7 ml 2-mercaptoethanol and 0.15 gr bromophenol blue, dissolved to 100ml) and hold at 50C for 40 min. Urea-polyacrylamide gel electrophoresis (urea-PAGE) was carried out using a Protean II xi cell vertical slab unit (Bio-Rad Laboratories ltd., Hemel Hempstead, Herts, UK). Urea-PAGE gels (12.5%) were prepared and run according to the method or Ardö (1999). Reagents used were obtained by Sigma-Aldrich, Inc and Fisher Scientific. 100 STATISTICAL ANALYSIS A two-way analysis of variance (ANOVA) on data was carried out using a general linear model procedure with Turkey’s pair wise comparison at 95% confidence level, using the package Minitab 15 (minitab Ltda., Coventry, UK). RESULTS AND DICUSSION FREE FATTY ACIDS FFA It is important to keep in mind that levels of lipolysis measured as FFA released are considered to be moderate for Cheddar cheese (P.L.H. McSweeney et al. 1993; P. F. Fox et al. 1999; Paul L.H. McSweeney and Sousa 2000), which is the reason why excessive lipolysis is undesirable and may be considered as rancid by some consumers (Yvonne F. Collins, McSweeney, and Wilkinson 2003). Due to their considerably lower perception thresholds (Molimard and Spinnler 1996), the most important FFA contributing direct and indirectly to the background ofCheddar cheese flavor are those of short (C4:0 to c8:0) and medium (C10:0 to C12:0) chain.Thus, it is relevant to point out that evident differences were found between the two types of samples assessed. Also, noticeable differences were found for long chain fatty acids, and in spite of their low contribution to the overall flavor of Cheddar cheese, they are still a good index of the degree of lipolysis. In addition it can be seen in the figures 22 that the slope of the lines corresponding to short chain fatty acids in these graphs is steeper, which might indicates that lipolysis of triglycerides containing short chain fatty acids could be faster. This makes sense since enzymes have better access to these substrates that are usually located at the sn-1 and sn3 position (Balcão and Malcata 1998). Nonetheless, in this case it is difficult to suggests what could be the most important enzymatic activity between lipases and esterases, since lipolytic enzymes are specific for the outer ester bonds of tri- or diacylglycerides (Deeth and Touch 2000; Metwalli, de Jongh, and van Boekel 1998) and it has been reported that butanoic, and other short and medium chain fatty acids are preferentially released by lipolytic activity (Bills and Day 1964; Chavarri et al. 1997; Yvonne F. Collins, McSweeney, and Wilkinson 2003). However, based on the fact that the most important lipolytic activity is provided by LAB 101 enzymes, it is not possible to tell whether lipases or esterases influence more the lipolysis in these samples without a study of specificity which takes into account the bacteria strain. As it is mentioned above C4:0 and C6:0 were the FFA with highest increment whereas C18:0, C14:1, C16:1 and C18:3 were the FFA with the lowest raise during ripening. This is explain by the accessibility of lipases to this substrates, since all of them are located at the sn-1 and sn3 position (Balcão and Malcata 1998). In addition, it can be seen in figure 21, that the most dominant FFA were C14:0, C16:0, C18:0 and C18:1; however, in spite of this quantitative importance, this is related to the fact that these are the most abundant FFA in milk (Yvonne F. Collins, McSweeney, and Wilkinson 2003). And finally the most important observation is that lipolysis is faster for cheeses made in the TCCA plant, which is not easy to explain due to it might be related to many factors, and it is evident that these two facilities are not following the same procedures or standardization of raw materials. Some of these factors are: 1) differences in cell viability and autolysis of the starter strain; 2) higher activity of the lipoprotein lipase LPL as consequence of subtle differences in the preparation of cheese milk such as heating or homogenization protocols; 3) Differences in the control of relative humidity and temperature during ripening of curds, which could proportionate better conditions for NSLAB growth; and 4) Differences during salting, which for sure could have inhibitory effect in different zones of the original blocks, because LAB enzymes are really sensitive to the salt in moisture content (Gripon et al. 1991; P. F. Fox and Stepaniak 1993). Figure 21 FFA chromatogram TCCA Vs CRP 102 Figure 22 Development of individual FFA for TCCA and CRP cheese 103 Volatile Sulfur Compounds VSC’s One type of the distinctive aromas of cheddar cheese results from the decomposition of sulfur containing amino acids such as cysteine and methionine. Indeed it has been reported that volatile sulfur compounds (VSC’s) correlate with good Cheddar cheese flavor in spite of their individual attributes described as garlic, onion, cabbage and skunk (D. J. Manning, Chapman, and Hosking 1976; B. Weimer, Seefeldt, and Dias 1999). Furthermore, it has been stated that the most important contributors are H2S, MeSH, and DMS, and contrary compounds such as 104 DMDS, DMTS, 3-methylthiopropionaldehyde, Carbonyl sulfide, carbon disulfide and dimethyl sulfone do not have a significant odor activity (H. M. Burbank and Qian 2005). By preventing the development of artifacts through a thorough de-activation work on vials and injection ports (liner), it is possible to suggest that the results of this work confirm that hydrogen sulphide (H2S), carbon disulphide (CS2), methanethiol (MeSH), and dimethyl sulphide (DMS) are the only volatile sulfur compounds formed during cheese ripening, and that compounds such as dimethyl disulphide (DMDS) and dimethyl trisulphide (DMTS) are decomposition or oxidation products from MeSH once the samples are exposed to environments where considerable amounts of oxygen are present. Accounting with the facts mentioned above, only the development of MeSH, DMS and H2S will be addressed in this work, thus only calibration curves for these compounds were calculated. However, good linear correlation coefficients were attained for H2S and DMS. Unfortunately, in the case of MeSH it was not possible to achieve a satisfying calibration curve free from DMDS and DMTS, whereby the interpretation of results for this compound was done based on the area ratio with respect to the internal standard EMS. The figures 25 and 26, revealed that only MeSH and DMS had a steady development during ripening, and that its concentration increased as cheese aged in comparison to H2S, which did not exhibit a regular development pattern in figure 23. Nonetheless, these figures also demonstrate that there are not significant and noticeable differences of the sulfur attributes related to the origin of the samples. Although it was not found in this work, due to the initial increment of the H2S principal precursor, cysteine, product from the denaturation and incorporation of β-lactoglobulins to casein micelles (which have a limited amount of this amino acid) as consequence of the heat treatment of cheese milk, it was expected that H2S showed a rising tendency along the maturation of samples. However, nothing was visible despite of the possible degradation and conversion of sulfydryl groups (Fennema and Damodaran 1996) through α- and/or βelimination reaction of cysteine by enzymes such as Cystathionine β- and γ-lyase (found in brevibacteria and bacilli, potentially NSLAB) resulting in hydrogen sulfide formation (B. 105 Weimer, Seefeldt, and Dias 1999; Seefeldt and Weimer 2000), which is a mechanism still unclear and not well understood. Moreover, it can be seen that H2S is an important contributor to the attributes of cheddar cheese based on the concentration found in this work and its odor threshold, 10 ppb in water (Rychlik et al. 1998), resulting in a relevant odor activity value. In addition it is possible to notice that in spite of the lack of a trend of generation, the samples from the TCCA plant had a slightly higher H2S concentrations than those for the CRP plant samples, which could be mainly explained by 1) subtle differences in the heat treatment of cheese milk between facilities that promote incorporation of β-lactoglobulins to casein micelles, 2) differences in the NSLAB microflora or 3) definitively lack of standardization for the raw milk Unfortunately, in this work methanethiol could not being confirm as a potent odorant in cheese due to the lack of effectiveness in constructing a calibration curve free of its oxidation products DMDS and DMTS. Nonetheless, it can be seen that in both cases the amount of MeSH increased during ripening time, which is in agreement with Urbach 1995. This might be attributed to enzymatic reaction provided by secondary microflora rather than by chemical reaction. And as a matter of fact, regardless it has been reported that MeSH can be obtained in chemical reduced cheese slurries made without starter cultures from the chemical decomposition of methathione (Donald J. Manning and Moore 1979; Green and Manning 1982), it is more likely that MeSH can be formed by a enzymatic process, which may require less activation energy, based on the fact that it is a catalytic reaction by nature (Alting et al. 1995; Smacchi and Gobbetti 1998; Dias and Weimer 1998). The enzymatic formation of MeSH could be via a single pathway catalysed by the catabolism of L-methionine by Lmethionine γ-lyase (Tanaka, Esaki, and Soda 1985) or cystathionine β or γ-lyase (Alting et al. 1995), or via two-step pathways during transamination of L-methionine in the presence of aketo glutarate to form a-keto-γ-methylthiobutyrate (KMTB) (Gao, Mooberry, and Steele 1998; Yvon and Rijnen 2001; Amarita et al. 2001), which can then be further broken down enzymatically to form 3-methylthiopropionaldehyde and MeSH. However, these results show no significant difference between samples from these two plants regarding this attribute, which it is not clear or easy to explain. In addition, because of the limited content of sulfur containing amino acids in caseins, it seems possible that the preparation of the cheese milk could be very similar or at least not drastically different since higher concentration could be 106 expected from more inclusion of whey protein into the cheese curd as consecuence of the heat treatment of milk, which is not what happened Something similar to what is described above occurred in the case of DMS. The results in figure 26 show that although this compound is usually present in raw and heat treated milk (Datta et al. 2002; Lee et al. 2007), its concentration increased during the ripening stage, and slightly higher amounts were found for the TCCA samples. Indeed, the results confirm that this is an important contributor to the aroma of cheddar cheese (Milo and Reineccius 1997) since its concentration varied from 5ppm to 30 ppm and its odor threshold in water is 2ppm (Rychlik 1998), which are amounts comparable to the ones obtained by Burbank and Qian, 2008. On the other hand, there is no proved and/or clear mechanism for the generation of DMS during cheese ripening; nonetheless, it has been stated that DMS is product of the metabolism of propioni-bacteria, present in milk microflora (Baer and Ryba 1992), and formed from methionine (Curioni and Bosset 2002). Additionally it is known that methionine is mainly present in β-lactoglobulins and integrated to caseins after thermal denaturation (Fennema and Damodaran 1996; Datta et al. 2002), which provides the sulfhydryl group required for DMS generation. Moreover, it can be seen in the figure 26 that there is no significant difference between samples from these two production plants, however, the concentration of samples from the TCCA plant are slightly higher, which is really hard to explain and is most likely attributed to difference in activity of NSLAB. On the one hand, due to the heat treatment of cheese milk, DMS is most likely formed via protein-bound methionine and the formation of DMS from methionine required more energy than the formation of MeSH. This means that the actual reaction is probably way more complicated than that proposed by R. de Wit and Nieuwenhuijse 2008, which involves the possibility of oxidation of MeSH into DMS and H2S. On the other hand, the production of DMS by catabolic reaction of methionine during ripening involves both non-enzymatic and enzymatic decomposition of Sulfonium salts resulting from the catabolism of methionine or cysteine such as a-keto-γ-methyl thiobutyrate (KMTB) (Gao, Mooberry, and Steele 1998; Yvon and Rijnen 2001; Amarita et al. 2001), Smethyl thioacetate, S- methyl thiopropionate, S- methyl thiobutyrate, and maybe Smethylmethionine, which can be used as substrate by a wide range of enzymes resulting in 107 DMS formation (Bentley and Chasteen 2004). In addition, there is a possibility that MeSH can be further transformed to DMS by a methyl transfer reactions involving thiol transferases and lyases for sulfur-containing amino acids provided by secondary microflora such as Brevibacterium linens (Dias and Weimer 1998), different strains of Lactococcus lactis subsp. cremoris (Alting et al. 1995), Lactococcus lactis subsp. lactis, Lacto bacillus sp., Propionibacterium shermanii and/or the yeast Geotrichum candidum and Kluyveromyces lactis (K. Arfi et al. 2002) Figure 23 VSC's chromatogram for TCCA cheese Figure 24 Development of H2S for TCCA Vs CRP cheese Figure 25Development of MeSH for TCCA Vs CRP cheese 108 Figure 26 Development of DMS for TCCA Vs CRP cheese Effect of treatment on Proteolysis Based on the results of this work, it is possible to state that measurements of Total Kjeldahl Nitrogen (TKN) and RP-HPLC revealed clear differences in the rate and pattern of proteolysis. Unfortunately, the results of the Urea-PAGE test are not conclusive to discriminate the origin or quality of the samples. Soluble Nitrogen Fractions and TKN Following a fractionation scheme in which the extractability of nitrogen compounds depend on pH (Ardö et al. 1999; Sousa, Ardö, and McSweeney 2001;Voigt et al. 2012), it was possible to appreciate how nitrogen concentrations increased during time, and was different regarding to the origin of the samples. In figures 27 and 28, it can be seen that the nitrogen levels for the WSN fraction were the highest in comparison to the other fraction, displaying values 4 to 5 times higher than the TCA-SN fraction and 8 to 18 times higher in comparison to those of the PTA-SN fraction. In addition, by the end of the observation period the nitrogen content values were at least twice those at the beginning. With regard to the levels of WSN, it can be seen that during the first 6 months of maturation, the nitrogen content is similar for both types of samples (TCCA and CRP); however, after this period the difference between samples became visible, showing higher values for those 109 corresponding to the TCCA plant. Thus, as it is suggested in the work of Bansal, Piraino, and McSweeney 2009, using this fraction as a reliable index of primary proteolysis, which is related to the remaining rennet activity and indigenous milk proteinases (S. Visser 1993; Paul L. H. McSweeney et al. 1994), it may be possible to focus the attention on variables in the process that are capable to alter the performance of this enzymes such as, acid development, temperature profiles, salt uptake and diffuse, and syneresis, The levels of TCA-SN in figure 30 also illustrate differences in the proteolysis development between samples from these two plants. In a similar way to the WSN fraction, this one results in final values that are approximately 3 times higher than those at the beginning of the observation, showing higher values for the samples of the TCCA plant during the ripening stage, which are slightly lower values but still comparable to those found by (Voigt et al. 2012). It is well known that in this fraction the higher the concentration of TCA, the lower the number of soluble peptides. Therefore, it is probable that a 12%TCA solution can be rich in medium sized and small peptides, and amino acids with low and medium hydrophobicity (Kuchroo & Fox 1982), from the N-terminal half of αs1-casein (Tanoj K. Singh, Fox, and Healy 1997), the N-terminal half of β-casein; most of them products of the action of starter enzymes (R. B. O’Keeffe, Fox, and Daly 1976; A. M. O’Keeffe, Fox, and Daly 1978), and endopeptidases from products of Chymosin or Plasmin. Whereby, it would be possible to attribute this difference to the activity of the starter enzymes. Indeed, the ability to degrade peptides from the action of Chymosin and Plasmin in cheese by LAB and NSLAB is tightly determined by the right combination of growth conditions, enzymes activity, ability to lyse and ripening conditions. Thus water activity and pH determine the survival and growth of microorganisms in cheese and indirectly their enzyme activity. However, the difference in substrate availability related to the activity of Plasmin and rennet is still a possibility.As a matter of fact, much of the work in lab is focused on maximizing cell mass starter activity and stability during storage rather than in proteolytic activity. Another factor that can be taking into account could be the lack of reproducible starter performance, related to undefined mixed strain cultures selected from original natural cultures rather than defined mixtures of pure characterized strains. 110 The PTA-SN fraction is mainly composed by very small peptides (<15 kDa) and amino acids of approximately 600 Da (Aston and Dulley 1982), and had a similar trend to that observed in the results for last fractions, where the values indicate a faster secondary proteolysis for the TCCA samples. Moreover, it is possible to appreciate that in comparison to the other fractions, the increment of nitrogen content was more steadfast, displaying final values for the TCCA samples that were 2.5 higher than those at the beginning. Alike the TCA fraction, these results could be related to non-reproducible processing variables between the two plants. In addition, taking in to account that the PTA-SN fraction is a index of secondary proteolysis, the results for this fraction can be pointed to dissimilar conditions of humidity and temperature in the maturation rooms for these two plants, or simply a different starter culture.and cheese microflora From the results of these fractions, it is reasonable to affirm that the water based fractionation scheme used is sufficient to extract the majority of water soluble peptides to establish a fair comparison between products of these two plants. And the most probable explanation to the difference in the proteolysis is found in dissimilar ripening conditions, and non-reproducible manufacturing practices. Figure 27 TKN fractions TCCA cheese 111 Figure 28 TKN fractions CRP cheese Figure 29 WSN TCCA Vs CRP cheese Figure 30 TCA-SN TCCA Vs CRP cheese 112 Figure 31 PTA-SN TCCA Vs CRP cheese Peptide analysis by RP-HPLC Another index of proteolysis to evaluate the difference between the ripening of samples from different origin is their peptide profile estimated by RP-HPLC. In this work, it was used a principal component analysis (PCA) to interpret the raw data from chromatograms based on Piraino, Parente, and McSweeney, 2004; and Benfeldt and Sørensen, 2001 works; where the complexity of the profiles containing more than 70 peaks, is initially reduced by establishing a set of elution time intervals (for which the area of each one is expressed as percentage of the total area of the chromatogram), and then the variability is measured and analyzed through the variation of the contributing eigenvalues in the correlation matrix for the principal components of the resulting model. A comparison of the ripening characteristics of these Cheddar cheese samples was set up by randomized block design with three replicates. Visible differences were supported by the PCA analysis, which resulted in a model with three principal components (PCs) that explain the 80.6% of the variability of the data, and after comparing in pairs the score plots for any combination of the principal components, only the score plot for the PC1 Vs PC2 revealed a correlation. These two principal components explain the 70.8% of the variability of the data. In figures 33 and 34 it can be seen that PC1 differentiate the samples according to their age, while PC2 represent mainly the contribution from the origin to the variation. Therefore, the points in these figures displaying higher scores indicate samples with longer ripening that 113 correspond to the TCCA plant, which suggest that proteolysis is slower for cheeses made in the CRP plant. The projection of the eluting intervals on the PC1 and PC2 is shown in figure 35, and it presents the correlation between variables and their effect on the amount of peptides eluting within certain retention times. The segments 12-20, 20-25, and 35-40 have high scores for the PC1, which suggest that the amount of peptides eluting in this zones increase during ripening. In contrast the segments 25-30, 30-35 40-45, 45-50, 50-55, 55-60 and 60-65 have lower scores, indicating that the amount of peptides in these segments reduces over time. On the other hand, the segments, 30-35, 55-60 and 60-65 have values close to 0 in the PC2, which suggest that the peptides eluting in this intervals have no variability as consequence of their origin. In a different way, the segments 12-20, 20-25, 25-30, 45-50 and 50-55 show low scores for the PC2, which indicates that the amount of peptides in these zones is smaller for the samples that correspond to the CRP plant, while the segments 35-40 and 40-45 have high scores for PC2, displaying a higher concentration of peptides in these intervals for the chesses made in the TCCA plant. Regarding the intervals 12-20 and 20-25, based on the works of Tove M. I. E. Christensen, Kristiansen, and Madsen 1989; T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1995; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998, it is possible to say that these sections of the chromatogram are mainly composed by peptides, which are products from the hydrolysis of the αs1-casein. Indeed, besides to para-κ-caseins, these parts of the chromatogram are dominated by the peptides αs1-CN(f1-9) and αs1-CN(f1-13), which are accumulated during cheese ripening and are originated from the hydrolysis of the peptide αs1-CN(f1-23) by the starter cell-envelope proteinase (CEP). This last peptide is result of the cleavage of the bond Phe23-Phe24 of αs1-caseins by Chymosin during the early stages of proteolysis. In addition, other peptides that constitute these segments and are product of the action of action of CEP and amino peptidase on the peptide αs1-CN (f1-23) are αs1-CN (f18), αs1-CN(f8-23), αs1-CN(f9-23), αs1-CN(f14-23), and the N terminal residues αs1-CN(f10?), αs1-CN(f17-?), αs1-CN(f18-?) and αs1-CN(f11-?). Other peptides in these segments with different origin are αs1-CN(f25-31), product of the hydrolysis of αs1-CN(f24-199) by aminopeptidase and/or Chymosin; αs1-CN(f92/93-?), which may involve activity of starter 114 endopeptidases (Pep O, Pep F); And finally αs2-CN(f1-?) and αs2-CN(191-197) that might involve lactococcal CEP and aminopeptidase activity due to the C terminus of the last peptide. Therefore, from the kind of peptides in this retention times and the proteolytic systems involved, it is feasible to attribute the difference between the ripening of the samples from both plants to the rennet, starter LAB, or to processing variables affecting these systems such as NaCl (salting), temperature and relative humidity of ripening rooms, and pH at draining, which in the case of the CRP plant seems like to contribute to the depletion of the enzymatic activity. The segments 25-30 and 30-35 got negative loadings for the PC1, which indicates that the relative amount of peptides eluting in this intervals decrease over time. Additionally it can be seen that these segment obtained negative loading scores for PC2, which are related to a slower proteolysis in the samples from the CRP plant. The fact that the amounts of peptides eluting in the first segment decrease over time has to do with the type of peptides dominating this zone, which are αs1-CN, αs1-CN (f24-199) and αs2-CN that tend to decrease as proteolysis takes place by the activity of Chymosin and CEP, specially over the first two. In the case of the segment 30-35 min, in spite of the possible accumulation of the peptides αs1CN (f85-91), αs1-CN (f11-?), αs2-CN (f170-?) and αs1-CN (f175-182), the presence of β-CN seems to strongly influence the amount of relative peptides eluting in this zone, which of course is expected to decrease over time (in spite of the fact that it is a water insoluble component and only the 50% of the β-CN are hydrolyzed during ripening in Cheddar cheese) (Tove M. I. E. Christensen, Kristiansen, and Madsen 1989; T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1995; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998; Sousa, Ardö, and McSweeney 2001). Regarding the scores in PC2, it might be explained by the fact that the water-soluble peptide profiles are directly related to the variety, and reflect the specificity of the LAB and NSLAB enzymes (P. F. Fox and McSweeney 1996). In addition it is obvious the influence of Chymosin on αs1-CN, αs1-CN (f24-199), whereby it is possible once again to focus the attention on possible distinct renneting or starter systems activity, consequence of smaller retention of coagulant activity due to denaturation by cooking temperature of curds, or because of low moisture level in the final cheese (P. F. Fox et al. 1999), or because a non optimum pH at draining of curds, which in addition can cause dissociation of plasmin and plasminogen from micelles. Also the 115 conditions for the LAB and NSLAB growing and survival might be different, probably due to calibration of instruments. With respect to the segments 35-40 and 40-45, it will be possible to say that the peptides eluting in this zone have a hydrophobic character due to the type of mobile phase mixture they are eluting in. In addition, as it can be seen in the loading plot the relative amount of peptides in the section 35-40 slightly increased, whereas relative amounts in the section 40-45 decrease during the ripening. However, both sections had positive scores respect to PC2, specially for the segment 35-40 min, which suggest that proteolysis related to this type of peptides is faster for samples from the TCCA plant. From the works of Tove M. I. E. Christensen, Kristiansen, and Madsen 1989; T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1995; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998, it might be possible to locate the peptides αs1-CN(f93–?), αs1-CN(f24–30), αs1-CN(f26–32), αs1-CN(f26–34) resulting from the hydrolysis of the peptide αs1-CN(f24–199) by Chymosin, CEP and aminopeptidase in the segment 35-40. Also peptides αs2-CN(f204–207), which is a C-terminal residue and product of lactooccal CEP (Paul L. H. McSweeney et al. 1994), β-CN(f45-52), as well reported product of hydrolysis by CEP and aminopeptidase, and traces of γ-caseins, which accumulate during ripening and are mainly present in the retentate of the filtration of the WSE, correspond to this segment. This again is evidence of the differences mentioned above. Regarding the components eluting in the segments from 40 to 65 mins, it can be seen that they have negative scores respect to PC1, which indicates that their amount decrease during ripening, and in a similar way these segments, excepting 40-45, have negative scores for the PC2, suggesting that elements in segment 40-45 undergo a faster proteolysis in the TCCA plant whereas segments from 45 to 65 have slower proteolysis in the CRP plant. This can be explain based on the fact that these segments, and fractions studied in earlier works correspond to the last part of the gradient elution and are mainly composed by amino acids. Therefore it can be expected that once they are in its free form they become precursors of flavor compounds, and catabolic reactions, resulting in decreasing amounts of components for these zones. However, it is difficult to explain the reason why proteolysis goes faster in the segment 40-45 for the samples from TCCA and slower for the rest of the chromatogram for 116 the samples from CRP since there are no real measurements of what type of amino acids are present in these regions and the subsequent catabolic pathway for their degradation into flavor compounds such as α-keto acids, aldehydes, carboxylic acids, alcohols, esters and thioesters Figure 32 Scree plot of TCCA Vs CRP cheese Figure 33 Score plot of TCCA Vs CRP cheese (age) 117 Figure 34 Score plot of TCCA Vs CRP cheese (Origin) Figure 35 Loading plot of TCCA Vs CRP cheese 118 Figure 36 Peptide profile CRP cheese (A and B) Vs TCCA (C and D) cheese, for 2 and 14 months A B C D Electrophoresis From the results of the results of the urea-PAGE of the insoluble nitrogen fraction in figures 38, 39 and 40, it can be seen the progressive degradation of αS1-CN into the peptides αS1-CN (f24-199), αS1-CN (f121-199), αS1-CN (f99-199), and the β-CN into γ-CNs during ripening. Also it can be seen that apparently the development of peptides from αS1-CN is faster than that for peptides from β-CN, which can be related to the fact that Chymosin is the starter of hydrolysis of caseins and only the 50% of β-CN is hydrolyzed. However the primary proteolysis pattern for both types of samples is similar during progress of ripening, which is the reason why it is possible to attribute the difference in quality of these cheeses according to its origin to the secondary proteolysis rather than the primary proteolysis 119 Figure 37 Urea PAGE for ripening of TCCA cheese β -CN(f106-209) β -CN(f29-209) β -CN(f108-209) β -CN αs1 -CN αz1-CN(f24-199) αs1-CN(f121-199) 2M 4M 6M 10 M 12 M 15 M 120 Figure 38 Urea PAGE for TCCA Vs CRP cheeses (12 m) TCCA CRP CONCLUSION The present study demonstrates that the use of FFA profile, VSC’s profile, measurement of the level of the Total Kjeldahl nitrogen for the WSN, TCA-SN and PTA-SN fractions, and the analysis of the RP-HPLC peptide profile of the WSN fraction by using a PCA, are effective tools and ripening indices to differentiate Cheddar cheese samples regarding their age and origin. The urea-PAGE was effective to differentiate samples by their age; nonetheless it is clear that it is not sensitive enough to detect differences related to the origin of the sample. In addition, the results of levels of nitrogen for all the 3 fractions analyzed demonstrated that proteolysis is faster for cheeses made in the TCCA plant. This was supported by the PCA model obtained which suggest evident differenced in manufacturing practices between the evaluated facilities. In a similar way, it was proved that lipolysis is slower for cheese produce 121 in the CRP plant, whose samples showed lower levels of individual FFA. The amounts of DMS, H2S and MeSH showed differences between samples. However, despite difference is not significant, it is possible to see a trend indicating that the catabolism of sulfur containing amino acids such as methionine and cysteine can be faster for the cheeses made in TCCA. Another interesting outcome from this analysis was to point out the possibility that DMDS and DMTS are artifacts from extraction and separation procedures rather than metabolites from the ripening of Cheddar cheese. REFERENCES Alting, A. C., W. Engels, S. van Schalkwijk, and F. A. Exterkate. 1995. “Purification and Characterization of Cystathionine (beta)-Lyase from Lactococcus Lactis Subsp. Cremoris B78 and Its Possible Role in Flavor Development in Cheese.” Applied and Environmental Microbiology V61 (11) (November 1): pp. 4037–4042. Amarita, F., T. Requena, G. Taborda, L. Amigo, and C. Pelaez. 2001. “Lactobacillus Casei and Lactobacillus Plantarum Initiate Catabolism of Methionine by Transamination.” Journal of Applied Microbiology V90 (6): pp. 971–978. Aston, J. W., and L. K. Creamer. 1986. “Contribution of the components of the water-soluble fraction to the flavour of Cheddar cheese.” New Zealand Journal of Dairy Science and Technology V21 (3): pp. 229–248. Aston, J. W., and John R. Dulley. 1982. “Cheddar cheese flavor review.” Australian Journal of Dairy Technology V37 (2): pp. 59–64. Baer, A., and I. Ryba. 1992. “Serological Identification of Propionibacteria in Milk and Cheese Samples.” International Dairy Journal V2 (5): pp. 299–310. Balcão, Victor M., and F.Xavier Malcata. 1998. “Lipase Catalyzed Modification of Milkfat.” Biotechnology Advances V16 (2) (March): pp. 309–341. Bansal, N, Paolo Piraino, and P. L. H. McSweeney. 2009. “Determination of Proteolysis in Cheese.” In Handbook of Dairy Foods Analysis, pp. 405–417. CRC Press. Benfeldt, Connie, and John Sørensen. 2001. “Heat Treatment of Cheese Milk: Effect on Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4–7): pp. 567–574. Bentley, Ronald, and Thomas G. Chasteen. 2004. “Environmental VOSCs––formation and Degradation of Dimethyl Sulfide, Methanethiol and Related Materials.” Chemosphere V55 (3): pp. 291–317. Bills, D.D., and E.A. Day. 1964. “Determination of the Major Free Fatty Acids of Cheddar Cheese.” Journal of Dairy Science V47 (7): pp. 733–738. Burbank, Helen M., and Michael C. Qian. 2005. “Volatile Sulfur Compounds in Cheddar Cheese Determined by Headspace Solid-phase Microextraction and Gas Chromatograph-pulsed Flame Photometric Detection.” Journal of Chromatography A V1066 (1–2): pp. 149–157. Burbank, Helen, and Michael C. Qian. 2008. “Development of Volatile Sulfur Compounds in Heat-shocked and Pasteurized Milk Cheese.” International Dairy Journal V18 (8) (August): pp. 811–818. 122 Chavarri, Felisa, Mailo Virto, Celia Martin, Ana I. Nájera, Arantza Santisteban, Luis J. R. Barrón, and Mertxe De Renobales. 1997. “Determination of Free Fatty Acids in Cheese: Comparison of Two Analytical Methods.” Journal of Dairy Research V64 (03): pp. 445–452. Christensen, Tove M. I. E., Kristian R. Kristiansen, and Jesper S. Madsen. 1989. “Proteolysis in Cheese Investigated by High Performance Liquid Chromatography.” Journal of Dairy Research V56 (05): pp. 823–828. Cliffe, A.J., D. Revell, and B.A. Law. 1989. “A Method for the Reverse Phase High Performance Liquid Chromatography of Peptides from Cheddar Cheese.” Food Chemistry 34 (2): 147–160. Cliffe, Allan J., Jonathan D. Marks, and Francis Mulholland. 1993. “Isolation and Characterization of Non-volatile Flavours from Cheese: Peptide Profile of Flavour Fractions from Cheddar Cheese, Determined by Reverse-phase High-performance Liquid Chromatography.” International Dairy Journal V3 (4–6): pp 379–387. Collins, Yvonne F., Paul L.H. McSweeney, and Martin G. Wilkinson. 2003. “Lipolysis and Free Fatty Acid Catabolism in Cheese: a Review of Current Knowledge.” International Dairy Journal V13 (11): pp. 841–866. Collins, Y.F., P.L.H. McSweeney, and M.G. Wilkinson. 2004. “Lipolysis and Catabolism of Fatty Acids in Cheese.” In Cheese Chemistry, Physics and Microbiology, Volume 1:373–389. Academic Press. Curioni, P.M.G., and J.O. Bosset. 2002. “Key Odorants in Various Cheese Types as Determined by Gas Chromatography-olfactometry.” International Dairy Journal v12 (12): pp. 959–984. Datta, N., A. J. Elliott, M. L. Perkins, and H. C. Deeth. 2002. “Ultra-high-temperature (UHT) Treatment of Milk: Comparison of Direct and Indirect Modes of Heating” V57 (3): pp. 211–227. Deeth, H. C., and V. Touch. 2000. “Methods for detecting lipase activity in milk and milk products.” Australian Journal of Dairy Technology V55 (3): pp. 153–168. Dias, Benjamin, and Bart Weimer. 1998. “Conversion of Methionine to Thiols by Lactococci, Lactobacilli, and Brevibacteria.” Applied and Environmental Microbiology V64 (9): pp. 3320–3326. Farkye, Nana Y., and Patrick F. Fox. 1990. “Observations on Plasmin Activity in Cheese.” Journal of Dairy Research V57 (03): pp. 413–418. Fennema, Owen R., and Damodaran. 1996. “Amino Acids, Peptides, and Proteins.” In Food Chemistry, pp. 322–425. CRC Press. Fernández, Manuela, Tanoj K. Singh, and Patrick F. Fox. 1998. “Isolation and Identification of Peptides from the Diafiltration Permeate of the Water-Soluble Fraction of Cheddar Cheese.” Journal of Agricultural and Food Chemistry V46 (11): pp. 4512–4517. Fernández-García, Estrella. 1996. “Use of Headspace Sampling in the Quantitative Analysis of Artisanal Spanish Cheese Aroma.” Journal of Agricultural and Food Chemistry V44 (7): pp. 1833–1839. Fox, P. F., Law J, P. L. H. McSweeney, and Wallace J.M. 1999. “Biochemistry of Cheese Ripening.” In Cheese: Chemistry, Physics and Microbiology, pp. 382–450. Springer. Fox, P.F. 1989. “Proteolysis During Cheese Manufacture and Ripening.” Journal of Dairy Science V72 (6): pp. 1379–1400. Fox, P.F., and L. Stepaniak. 1993. “Enzymes in Cheese Technology.” International Dairy Journal V3 (4–6): pp. 509–530.. 123 Fox, P. F., and P. L. H. McSweeney. 1996. “Proteolysis in Cheese During Ripening.” Food Reviews International V12 (4): pp. 457–509. Fox, P. F., Law J, P. L. H. McSweeney, and Wallace J.M. 1999. “Biochemistry of Cheese Ripening.” In Cheese: Chemistry, Physics and Microbiology, pp. 382–450. Springer. Gao, Song, Ed S. Mooberry, and James L. Steele. 1998. “Use of 13C Nuclear Magnetic Resonance and Gas Chromatography To Examine Methionine Catabolism by Lactococci.” Applied and Environmental Microbiology V64 (12): pp. 4670–4675. Green, Margaret L., and Donald J. Manning. 1982. “Development of Texture and Flavour in Cheese and Other Fermented Products.” Journal of Dairy Research V49 (04): pp. 737–748. Gripon, Jean-Claude, V Monnet, G Lambert, and M. J. Desmazeaud. 1991. “Microbial enzymes in cheese ripening.” In Food enzymology, vol 1. K. Arfi, H.Spinnler, R. Tache, and P. bonnarme. 2002. “Production of Volatile Compounds by Cheese-ripening Yeasts: Requirement for a Methanethiol Donor for S-methyl Thioacetate Synthesis by Kluyveromyces Lactis.” Applied Microbiology and Biotechnology V58 (4): pp. 503–510. Lau, Kum Y., David M. Barbano, and Robert R. Rasmussen. 1991. “Influence of Pasteurization of Milk on Protein Breakdown in Cheddar Cheese During Aging.” Journal of Dairy Science V74 (3): pp. 727–740. Lee, Won-Jae, Dattatreya S. Banavara, Joanne E. Hughes, Jason K. Christiansen, James L. Steele, Jeffery R. Broadbent, and Scott A. Rankin. 2007. “Role of Cystathionine βLyase in Catabolism of Amino Acids to Sulfur Volatiles by Genetic Variants of Lactobacillus Helveticus CNRZ 32.” Applied and Environmental Microbiology V73 (9): pp. 3034–3039. Manning, D. J., Helen R. Chapman, and Zena D. Hosking. 1976. “The Production of Sulphur Compounds in Cheddar Cheese and Their Significance in Flavour Development.” Journal of Dairy Research V43 (2): pp. 313–320. Manning, Donald J., and Carolyn Moore. 1979. “Headspace Analysis of Hard Cheeses.” Journal of Dairy Research V46 (3): pp. 539–545. McSweeney, P.L.H., P.F. Fox, J.A. Lucey, K.N. Jordan, and T.M. Cogan. 1993. “Contribution of the Indigenous Microflora to the Maturation of Cheddar Cheese.” International Dairy Journal V3 (7): pp. 613–634. McSweeney, Paul L. H., Sylvie Pochet, Patrick F. Fox, and Aine Healy. 1994. “Partial Identification of Peptides from the Water-Insoluble Fraction of Cheddar Cheese.” Journal of Dairy Research V61 (4) (04): pp. 587–590. McSweeney, Paul L.H., and Maria José Sousa. 2000. “Biochemical Pathways for the Production of Flavour Compounds in Cheeses During Ripening: A Review.” Le Lait V80 (3): pp. 293–324. Metwalli, Ali A.M, Harmen H.J de Jongh, and Martinus A.J.S van Boekel. 1998. “Heat Inactivation of Bovine Plasmin.” International Dairy Journal V8 (1): pp. 47–56. Milo, C., and G. A. Reineccius. 1997. “Identification and Quantification of Potent Odorants in Regular-Fat and Low-Fat Mild Cheddar Cheese.” Journal of Agricultural and Food Chemistry V45 (9) (September 1): pp. 3590–3594. Molimard, P., and H.E. Spinnler. 1996. “Review: Compounds Involved in the Flavor of Surface Mold-Ripened Cheeses: Origins and Properties.” Journal of Dairy Science V79 (2) (February): pp. 169–184. 124 O’Keeffe, Arthur M., Patrick F. Fox, and Charles Daly. 1978. “Proteolysis in Cheddar Cheese: Role of Coagulant and Starter Bacteria.” Journal of Dairy Research V45 (3): pp. 465– 477.. O’Keeffe, R. B., P. F. Fox, and C. Daly. 1976. “Contribution of Rennet and Starter Proteases to Proteolysis in Cheddar Cheese.” Journal of Dairy Research 43 (1): 97–107. Piraino, Paolo, Eugenio Parente, and Paul L. H. McSweeney. 2004. “Processing of Chromatographic Data for Chemometric Analysis of Peptide Profiles from Cheese Extracts: A Novel Approach.” Journal of Agricultural and Food Chemistry 52 (23): 6904–6911. Preininger, Martin, and Werner Grosch. 1994. “Evaluation of Key Odorants of the Neutral Volatiles of Emmentaler Cheese by the Calculation of Odour Activity Values.” LWT Food Science and Technology V27 (3): pp. 237–244. Qian, M., and G. Reineccius. 2002. “Identification of Aroma Compounds in ParmigianoReggiano Cheese by Gas Chromatography/Olfactometry.” Journal of Dairy Science V85 (6) pp. 1362–1369. Seefeldt, K.E., and B.C. Weimer. 2000. “Diversity of Sulfur Compound Production in Lactic Acid Bacteria.” Journal of Dairy Science V83 (12): pp. 2740–2746. Singh, T. K, M. A Drake, and K. R Cadwallader. 2003. “Flavor of Cheddar Cheese: A Chemical and Sensory Perspective.” Comprehensive Reviews in Food Science and Food Safety V2 (4): pp. 166–189. Singh, T.K., P.F. Fox, P. Højrup, and A. Healy. 1994. “A Scheme for the Fractionation of Cheese Nitrogen and Identification of Principal Peptides.” International Dairy Journal V4 (2): pp. 111–122. Singh, Tanoj K., Patrick F. Fox, and Áine Healy. 1995. “Water-Soluble Peptides in Cheddar Cheese: Isolation and Identification of Peptides in the Diafiltration Retentate of the Water-Soluble Fraction.” Journal of Dairy Research V62 (4): pp. 629–640. Singh, Tanoj K., Patrick F. Fox, and Áine Healy. 1997. “Isolation and Identification of Further Peptides in the Diafiltration Retentate of the Water-soluble Fraction of Cheddar Cheese.” Journal of Dairy Research V64 (3): pp. 433–443. Smacchi, Emanuele, and Marco Gobbetti. 1998. “Purification and Characterization of Cystathionine Γ-lyase from Lactobacillus Fermentum DT41.” FEMS Microbiology Letters V166 (2): pp. 197–202. Smith, A. M., and S. Nakai. 1990. “Classification of cheese varieties by multivariate analysis of HPLC profiles.” Canadian Institute of Food Science and Technology Journal V23 (1): pp. 53–58. Sousa, M.J, Y Ardö, and P.L.H McSweeney. 2001. “Advances in the Study of Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4-7): pp. 327–345. Subramanian, A., W.J. Harper, and L.E. Rodriguez-Saona. 2009. “Cheddar Cheese Classification Based on Flavor Quality Using a Novel Extraction Method and Fourier Transform Infrared Spectroscopy.” Journal of Dairy Science V92 (1): pp. 87–94. Tanaka, Hidehiko, Nobuyoshi Esaki, and Kenji Soda. 1985. “A Versatile Bacterial Enzyme: L-methionine Γ-lyase.” Enzyme and Microbial Technology V7 (11) (November): pp. 530–537. Urbach, G. 1995. “Contribution of Lactic Acid Bacteria to Flavour Compound Formation in Dairy Products.” International Dairy Journal V5 (8): pp. 877–903. USDA. 2011. Dairy Products Annual Sumary. Visser, Servaas. 1993. “Proteolytic Enzymes and Their Relation to Cheese Ripening and Flavor: An Overview.” Journal of Dairy Science V76 (1): pp. 329–350. 125 Voigt, Daniela D., François Chevalier, John A. Donaghy, Margaret F. Patterson, Michael C. Qian, and Alan L. Kelly. 2012. “Effect of High-pressure Treatment of Milk for Cheese Manufacture on Proteolysis, Lipolysis, Texture and Functionality of Cheddar Cheese During Ripening.” Innovative Food Science & Emerging Technologies V13 (0): pp. 23–30. Weimer, Bart, Kimberly Seefeldt, and Benjamin Dias. 1999. “Sulfur Metabolism in Bacteria Associated with Cheese.” Antonie Van Leeuwenhoek 76 (1): 247–261. de Wit, Rudy, and Hans Nieuwenhuijse. 2008. “Kinetic Modelling of the Formation of Sulphur-containing Flavour Components During Heat-treatment of Milk.” International Dairy Journal V18 (5): pp. 539–547. Yvon, Mireille, and Liesbeth Rijnen. 2001. “Cheese Flavour Formation by Amino Acid Catabolism.” International Dairy Journal V11 (4–7): pp. 185–201. 126 CHAPTER 4 COMPARISION OF CHEDDAR CHEESE RIPENING MANUFACTURED WITH AND WITHOUT ADJUNCT CULTURE Lemus Muñoz, Freddy Mauricio, Qian Michael Department of Food Science and Technology, Oregon State University Corvallis Oregon 97331 127 ABSTRACT The effect of addition of adjunct culture isolated from cheese manufactured at the TCCA plant to cheese produce in the CRP plant was studied during the ripening stage of samples. Proteolysis was investigated by a fractionation scheme, resulting in an insoluble fraction analyzed by urea polyacrylamide gel electrophoresis (Urea-PAGE), and a soluble fraction which was further studied through water soluble nitrogen (WSN), trichloroacetic acid soluble nitrogen (TCA-SN) and phosphotungstic acid soluble nitrogen (PTA-SN) analyzed by total Kjeldahl nitrogen content (TKN). Reversed phase high performance liquid chromatography (RP-HPLC) was used to study the peptide profile of the water soluble fraction. Lipolysis was studied by levels of individual free fatty acids determined through gas chromatography-flame ionization detection (GC-FID) after isolation employing solid phase extraction (SPE). Volatile sulfur compounds were studied using head space solid phase micro-extraction (SPME) coupled with gas chromatography-pulsed flame photometric detection (PFPD). The Urea-PAGE method was able to differentiate samples according their age, but it could not discriminate samples regarding their treatment. Nonetheless, measurements of total Kjeldahl Nitrogen (TKN) of the WSN, TCA-SN, and PTA-SN fractions, and the principal component analysis of the RP-HPLC peptide profile of the WSN fraction, revealed differences in the rate and pattern of proteolysis for the samples. Levels of total nitrogen for the WSN, TCA and PTA fractions increased as cheese aged and were lower for cheeses made without adjunct culture. The principal component analysis of the RP-HPLC data resulted in PCA model with 3 principal components that accounted for the 83.4% of the variability. This model discriminates the samples according age and treatment, suggesting that samples made with adjunct culture undergoes more or faster proteolysis. FFA profiles reveal significant difference in the extension of lipolysis, which was higher or faster for cheese made with adjunct culture. The Volatile Sulfur Compounds (VSC) analysis showed that cheeses made with adjunct culture developed higher concentrations of H2S, DMS and MeSH, suggesting slower catabolism of sulfur containing amino acids in cheese made without adjunct culture. 128 INTRODUCTION Cheddar cheese is the product of a dehydration process involving 2 stages: 1) preparation of curd, which is usually done during the first 24 hours, and 2) ripening of curds, which can take up to 12 months. The development of flavor and texture is the result of three complex biochemical and microbiological process occurring during ripening such as glycolysis, lipolysis and proteolysis, whose early products constitute the precursors for the formation of volatile and non-volatile flavor compounds (Wallace J.M. and Fox P.F. 1997; Paul L.H. McSweeney and Sousa 2000). During ripening the major sources of enzymatic activity are the rennet enzymes (pepsin and Chymosin), indigenous milk enzymes (Plasmin and Cathepsin B and D), and starter lactic acid bacteria (LAB), non-starter lactic acid bacteria (NSLAB), and adjunct cultures enzymes (P. F. Fox et al. 1999). This stage is determined mainly by the extent of proteolysis (P. F. Fox 1989), which is the biochemical event where proteolytic enzymes hydrolyze αs1, αs2, β and κ caseins, originating large and intermediate peptides that are further hydrolyzed by LAB, NSLAB and adjunct culture intracellular enzymes into free amino acids (FAA) and other smaller peptides, which are important contributors to the background of cheese flavor and essential precursors for deamination, decarboxylation and/or transamination reactions, among others, to produce volatile and sapid compounds (Allan J. Cliffe, Marks, and Mulholland 1993; P. F. Fox et al. 1999; Paul L.H. McSweeney and Sousa 2000). In a similar way fat plays an important role during ripening, and besides of being solvent for aromatic and non-volatile compounds, its degradation through lipolysis results in the release of free fatty acids (FFA), which are flavor compounds that also can be catabolized into other compounds such as methyl ketones, alcohols and lactones (Gerda Urbach 1993; P. F. Fox et al. 1999). Regarding glycolysis and ripening, residual lactose is mainly metabolized to l-lactate, and further transformed to acetate through oxidation by LAB, NSLA or adjunct culture enzymes; this compound has been found significant in cheddar cheese flavor (P. F. Fox et al. 1996), and can be a positive attribute or an off-flavor depending on its concentration. However, ripening is an expensive and long process that has received significant attention due to the interest of manufacturers in accelerating this stage in order to reduce cost of production, and to improve the quality and consistency of attributes of the ready to sell product. This fact 129 has promoted research related to the addition of adjunct cultures to accelerate ripening and improve flavor (P. F. Fox and Stepaniak 1993; C.N. Lane and Fox 1996b). Co-starting or adjunct cultures can be nonstarter lactic acid bacteria, consisting mainly of Lactobacillus sp, or certain yeast species. The dominant NSLAB strains are mesophilic lactobacilli such as Lactobacillus casei ssp. casei, Lactobacillus paracasei ssp. paracasei, Lactobacillus plantarum, Lactobacillus curvatus, and Lactobacillus helveticus) (P.L.H. McSweeney et al. 1993; C.N. Lane and Fox 1996b; M. El Soda, Madkor, and Tong 2000) as well as other probiotic strains such as Lactobacillusacidophilus 4962, Lb. casei 279, Bifidobacterium longum 1941, Lb. acidophilus LAFTI® L10 (Shah 2006), that have been recently studied. While in the case of yeast, the preferred strains to work with are Debaryomyces hansenii and Yarrowia lipolytica (Ferreira and Viljoen 2003; M. De Wit et al. 2005). In order to take full advantage of adjunct NSLAB cultures, studies about cell autolysis have been conducted through the use of attenuated cultures, which has been done by means of sublethal treatments such as freeze shocking (FS), heat shocking (HS), and spray drying (SD), to provide more control on the release of intracellular enzymes in to the cheese matrix (M. A. El Soda 1993; M. El Soda, Madkor, and Tong 2000; Madkor, Tong, and El Soda 2000), reporting acceleration of the breakdown of peptides and consequently increase in the amount of amino nitrogen and decrease in bitterness, a considerably enhanced flavor intensity without affecting cheese composition, and also a reduction in off-flavors (Madkor, Tong, and El Soda 2000; M. El Soda, Madkor, and Tong 2000). Nevertheless, because of acidification is one of the crucial operations in the manufacture of cheddar cheese, adjunct cultures should meet certain criteria in order to be considered as agents for accelerating the ripening process. Therefore, qualities such as: 1) a potent lipolytic and proteolytic system with high autolytic activity, 2) assimilation of lactose and organic acids, 3) resistance to high salt concentration, low water activity and low pH, 4) ability to grow at low temperatures, and 5) in the case of yeast cultures, compatibility with the starter culture; are desire and should be take into account at the moment of selecting any culture. 130 The objective of this study is to establish a comparison between samples of Cheddar cheese manufactured in the CRP plant that were made with and without adjunct culture isolated from cheese produced in the TCCA plant, by monitoring lipolysis and proteolysis using: 1) free fatty acids profiles, 2) peptides profiles by RP-HPLC, 3) Urea-PAGE, 4) development of sulfur volatile compounds, and 5) nitrogen content of cheese fraction MATERIALS AND METHODS CHEESE SAMPLES Cheeses were manufactured by Tillamok county creamery. Cheeses were made according with standard protocols in the CRP plant The adjunct culture included is unknown and was extracted from good quality cheese produced at the TCCA plant. One blocks of cheese made from each treatment were selected randomly. All cheeses were aged using the same conditions at manufacturer’s facility, Every 2 months a 2 lb portion was sampled from each block and sent to the lab, where samples are stored at (-37C) to stop ageing process until analysis is completed. FREE FATTY ACIDS ANALYSIS Chemicals Pentanoic acid, heptanoic acid, nonanoic acid, undecanoic acid, and heptadecanoic acid were used as internal standards, they were purchased from Eastman (Rochester, N.Y., U.S.A). Butanoic acid, hexanoic acid, octanoic acid, decanoic acid, dodecanoic acid, tetradecanoic acid, 9-tetradecanoic acid, hexadecanoic acid, 9-hexadecanoic acid, octadecanoic acid, 9octadecanoic acid, 9,12-octadecanoic acid and 6,9,12 octadecanoic acid were used for the standard stock solution, and were obtained from Aldrich Chemical Co. Inc (Milwaukee, Wisconsin, U.S.A). Heptane, Isopropanol, Sulfuric acid, anhydrous sodium sulfate, chloroform, formic acid and diethyl ether were obtained from Fisher. Extraction From each 2lb block of cheese, 100 grams were wrapped in alumina foil, frozen with liquid nitrogen during 6 minutes, and then grinded for 30 seconds to obtain a fine powder. Six 131 grams of this previously freeze-ground cheese, 1 ml of 2N sulphuric acid and 1 ml of internal standard solution (C5:0, C7:0, C9:0, C11:0 and C17:0 in 1:1 hetpane-isopropanol) were mixed with 7 grams of anhydrous sodium sulfate and 20 ml of 1:1 diethyl ether- heptane in a 40 ml amber vial using a sonicator and manual agitation. During sonication, the salt-slurry solution is initially exposed for 15 minutes, after which each vial is shake vigorously to continue with a second sonication period of 20 minutes. With a glass-Pasteur pipette, the sample extract (solvent) is transferred to an AccuBOND amino cartridge (Agilent Technologies) conditioned previously with 10 ml of heptane. After the addition of the sample, the column is washed with 5 ml of 2:1 Chloroform-Isopropanol to remove non volatile triglycerides and phospholipids using a manifold vacuum chamber. Once the washing step is complete, free fatty acids are eluted with 5ml of 2% formic acid in diethyl ether, collected in a 20 ml vial and stored in the freezer until GC analysis. Chromatography The analysis was performed using a Hewlett-Packard 5890 gas chromatograph equipped with a flame ionization detector (FID). Samples were analyzed on a DB-FFAP column (15m x 0.53mm ID, 1 m film thickness; Supelco Wax10, Supelco U.S.A). Injector and detector temperatures were 250C. Nitrogen was used as carrier gas at a flow rate of 15 ml per minute at a split ratio of 1 to 1. The oven temperature was programmed for a 2 minutes hold at 60C, raised to 230C at a rate of 8C per minute with a hold of 20 minutes at 230C. Quantitative analysis The levels of free fatty acids concentrations were calculated based on individual peaks areas from GC-FID response in comparison to the internal standard peaks areas, using standard calibration curve of individual free fatty acid using Peak Simple software (SRI instruments, Torrance, CA). Each experimental value corresponds to the average of the 3 extraction replicates. VOLATILE SULFUR COMPOUNDS (VSC’S) Chemicals Dimethyl sulfide (DMS) was purchased from TCI America (Portland, OR, U.S.A.); gaseous methanethiol (MeSH) was obtained from Aldrich Chemical Co. Inc (Milwaukee, Wisconsin, 132 U.S.A), and a solution was prepared by bubbling the gas into cold methanol; a H2S solution was prepared by dissolving Na2S.9 H2O (Sigma Co) in acidic water stabilized with citric acid (pH 3). Extraction From each 2lb block of cheese, 100 grams were wrapped in alumina foil, frozen with liquid nitrogen during 6 minutes, and then grinded for 30 seconds to obtain a fine powder. Then one gram of this freshly prepared powder is added to a 20ml vial (formerly flushed with argon), followed by the addition of 4 ml of 1M citric acid and 20 l of the internal standard solution. After addition of sample vials were immediately sealed with screw caps with teflon-lined silicone septa. The vials used in this study were previously deactivated to its use with DMTCS 5% solution in toluene, toluene, methanol and distillate water. The volatile sulfur compounds were extracted with a 85 m Carbox-PDMS fiber (Supelco, Bellefonte, PA, U.S.A.). Prior to use, the fiber was conditioned at 300 C for 90 minutes. The fiber was then placed into a SPME adapter of a CombiPAL autosampler (CTC analytics AG, Zwingen, Switzerland) Fitted with a vial heater/agitator. Samples were pre-equilibrated at 500 RPM at 40C for 5 minutes, and the extraction of VSC’s was done at 250 RPM at 40C for 25 minutes. The desorption time was 5 minutes and 30 seconds. Chromatography The analysis was performed using a Varian CP-3800 gas chromatograph (Varian, Walnut Creek, CA, U.S.A.) equipped with a pulsed flame photometric detector (PFPD). The separation of analytes was made using a DB-FFAP fused silica capillary column (30m, 0.32 mm ID and 1 m film thickness; Agilent, Palo Alto, CA, USA) and nitrogen as carrier gas at constant flow at 2 ml per minute. The injector temperature was 300 C and it was in the splitless mode. The oven temperature was programmed for a 3 minutes hold at 35C, raised to 150C at a rate of 10C per minute, held for 5 minute, and then heated to 220C at a rate of 20C per minute with a final hold of 3 minutes. The PFPD was held at 300 C and 450 V with the following flow rates: Air 1 at 17 ml per min, H2 at 14 ml per min, and Air 2 at 10 ml/min. The detector response signal was integrated using the software Star Workstation 6.2, Varian) 133 Quantitative analysis Matrix effect In order to retain the matrix effect during the construction of the calibration curves, cheese powder from the “youngest sample” is used. It is de-volatilized by exposure to room conditions in a hood for 2 hours. Then 1 gram of powder is added to 4 ml of 1M citric acid in a 20 ml vial and exposed to a 50C water bath for 30 mins, prior to the addition of standards and internal standard solutions. Sulfur standards and internal standard preparation Two internal standards were used for the quantification of VSC’s: ethyl methyl sulfide (EMS) for H2S, MeSH and DMS, and isopropyl disulfide (IsoProDS) for DMDS and DMTS. The concentration of the internal standard solution was 500 ppm EMS and 500 ppm IsoProDS in methanol. Calibration curves were constructed by spiking cheese samples with a range of known concentrations of H2S, MeSH and DMS. Hydrogen sulfide (H2S) was prepared by dissolving Na2S.9 H2O in acidic water (pH = 3). Different concentrations of sodium sulfide solutions were made, and the concentrations of H2S were calculated based on the amounts of salt added to the matrix. A standard solution of 100 ppm of DMS was individually prepared in cooled methanol (-15C), and dilutions were made with cooled methanol at the same temperature. The mesh standard was prepared as following: 1) newly deactivated, recently flushed with argon, and cooled vials were used; 2) The original standard solution was made by bubbling pure MeSH into cooled methanol; 3) Dilutions were prepared by taking aliquots from the original solution contained in a sealed vial, through the teflon-lined silicone septa by using a syringe. And then injecting the aliquots into new sealed vials containing proportional amount of cooled methanol through the septa; 4) 1 gr of devolatilized cheese is added to a recently flushed vial (argon was used), which is immediately flushed again; 5) simultaneous argon flushing and addition of 4ml of “free” dissolved oxygen-1M citric acid solution and quick sealing of the vials; 6) Addition of 20 l of internal standard and MeSH standard through septa. The identification of target compounds was made by comparing retention times with those of pure standards. Ratios of the square root of the standard area to the corresponding square root of the internal standard area were plotted Vs concentration ratios to determine the relation between the response and concentration for the unknowns. Triplicate analysis was performed for all samples 134 PROTEOLYSIS Chemicals Sulfuric acid was purchased from Fisher Scientific International Inc. (Pittsburgh, PA, U.S.A.); Trichloroacetic acid was purchased from Alfa Aesar (Royston, UK); And phosphotungstic acid was obtained from Aldrich Chemical Co. Inc (Milwaukee, Wis, U.S.A) Sample preparation and fractionation From each 2lb block of cheese, 60 grams are blended with 120 ml of distilled water pre-heated to 55C. The mixture is blended for 5 minutes and the homogenate is incubated at 55C for 1 hour. Then the pH is adjusted to 4.6 with 1M HCl and the mixture is centrifuged at 3000g for 30 minutes at 4C. Suspension and supernatant were filtered thoroughly 3 times through glass wool. The filtrate was safe at -20C for macro blog digestion method analysis, and RP-HPLC analysis. The insoluble pellet was frozen at -20C for further Urea-PAGE gel electrophoresis analysis. The trichloroacetic acid soluble nitrogen fraction (TCA-SN) was prepared by the addition of 25 ml of pH 4.6 soluble fraction (WSN) to 25 ml of 24% trichloroacetic acid solution. Then the mixture is equilibrated for 2 hours at room temperature and filtered through filter paper Whatman No 40 before macro blog digestion method analysis. For the phosphotungstic acid soluble nitrogen fraction (PTA-SN), 10 ml of WSN are added to 7 ml of 3.95 M H2SO4 and 3 ml of 33% phosphotungstic acid solution. Then the mixture is equilibrated overnight at 4C and filtered through filter paper Whatman No 40 before macro blog digestion method analysis. Duplicate analysis was performed for all samples. Macro blog digestion (Kjeldahl Digestion) From the fractions collected an aliquot (2 ml for the Water soluble fraction, 1ml for TCA-SN and 1 ml for PTA-SN) is added into a 70 ml Kjeldahl Digestion flask with 10 ml of H2SO4 and the catalyst pellet containing 0,075 and 1,5 grams of mercuric oxide and potassium sulfate 135 respectively. The mixture is warmed to 150 C and hold for 1 hour, then heated to 250 C and hold for 1 hour, and finally heated to 350 C and hold for 2 hours. After digestion the sample is cooled down overnight to room temperature, and diluted with distillate water to 70 ml, followed by a gentile agitation. Then a 5 ml aliquot is used to determine the nitrogen content by a rapid flow analyzer FOSS II. Reversed phase High performance liquid chromatography analysis The RP-HPLC analysis was performed using a Shimadzu 6 series liquid chromatograph (Shimadzu scientific instruments, Kyoto Japan), consisting of an autosampler, 2 pumps, a multi-wavelength spectrophotometer and a controller unit. It was used a nucleosil RP-8 analytical column (250x 4mm, 5 m particle size, 300 A pore size) and a guard column (4.6 x10 mm) from waters (Milford, MA, U.S.A.). The mobile phase consists of solvent A (0.1% TFA in deionized and vacuum filtered water) and solvent B (0.1% TFA in acetonitrile). The elution was monitored at 214nm. The following gradient elution was performed: 1) 100% solvent A for 5 minutes followed by a linear gradient to 55% solvent B (v/v); 2) elution at 55% solvent B for 6 minutes followed by a linear gradient to 60%; 3) elution at 60% solvent B for 3 minutes; 4) The column is washed using 95% solvent B during 5 minutes; 5) the column is equilibrated using 100% solvent A during 10 minutes. The sample (WSN fraction) was dissolved in solvent A (10 mg per ml) and then micro-centrifuged at 14000 RPM for 10 minutes. An aliquot of 40 l from the extract was injected to a flow rate of 0.75 ml per min. Electrophoresis Samples of the water-insoluble nitrogen fraction were dry frozen prior to analysis. Samples were dissolved in a buffer (0.75 g tris, (hydroxymethyl) methylamine, 49 gr urea and 0.4 ml concentrated HCl, 0.7 ml 2-mercaptoethanol and 0.15 gr bromophenol blue, dissolved to 100ml) and hold at 50C for 40 min. Urea-polyacrylamide gel electrophoresis (urea-PAGE) was carried out using a Protean II xi cell vertical slab unit (Bio-Rad Laboratories ltd., Hemel Hempstead, Herts, UK). Urea-PAGE gels (12.5%) were prepared and run according to the method or Ardö (1999). Reagents used were obtained by Sigma-Aldrich, Inc and Fisher Scientific. 136 STATISTICAL ANALYSIS A two-way analysis of variance (ANOVA) on data was carried out using a general linear model procedure with Turkey’s pair wise comparison at 95% confidence level, using the package Minitab 15 (minitab Ltda., Coventry, UK). RESULTS AND DICUSSION FREE FATTY ACIDS FFA Levels of individual free fatty acids are shown in figures 40. It can be seen in figure 39 that the most dominant peaks were C14:0, C16:0, C18:0 and C18:1, however, despite these apparent relevant quantities, they are not important contributors to the Cheddar cheese aroma because of their high odor threshold. In addition, their high concentration is related to the fact that these are the most abundant FFA in milk (Banks 1991; Gunstone, Harwood, & Padley 1994), and also it is possible to see in the results that these FFA and C14:1, C16:1, C18:1 and C18:3 are the ones with the lowest increase over time, which is related to the harder access of lipases to their active sites. Indeed, most of them are located in the same position than the short chain fatty acids within the tri and diacylglycerides. On the other hand, the increment for the short chain fatty acids C4:0 and C6:0 was proportionally the highest and fastest, reaching at least twice their concentration at the beginning of the observation period, which makes sense since lipases and esterases have better access to these substrates located at the sn-1 and sn-3 position of the triacylglicerides. Also it is possible to appreciate that cheeses made with adjunct culture in general exhibited higher levels of FFA liberation during ripening in comparison to the control samples. These results indicate that adjunct cultures (possibly other strains of lactococci or lactobacilli) contribute to lipolysis, which is in agreement with Madkor, Tong, and El Soda 2000, and in addition, they have high activity of intracellular lipase upon autolysis. However it is important to keep in mind that Lactococcus and Lactobacillus spp are considered to be weakly lipolytic in comparison to species such as Pseudomonas, Acinetobacter and Flavobacterium, and it should be expected that the degree and pace of lipolysis is related to the autolysis capacity of the strains selected, which in this case are unknown. Therefore it is important to take into account that the adjunct culture used could have high rates of autolysis with high level of 137 enzymatic activity, or high autolytic activity and low level of enzymatic activity, or low rates of autolysis with high level of enzymatic activity, or low autolysis and enzymatic activity (ElSoda, Madkor, and Tong 2000). Figure 39 Urea PAGE for TCCA Vs CRP cheeses (2 months) Figure 40 Development of individual FFA for cheese with and without adjunct culture 138 139 Volatile Sulfur Compounds VSC’s Volatile sulfur compounds correlate with good Cheddar cheese flavor, and the most important contributors are H2S, MeSH, and DMS. These are product from the decomposition of amino acids such as cysteine and methionine (B. Weimer, Seefeldt, and Dias 1999; D. J. Manning, Chapman, and Hosking 1976; H. M. Burbank and Qian 2005). As it was mentioned earlier in the last two chapters, Hydrogen sulphide (H2S), carbon disulphide (CS2), methanethiol (MeSH), and dimethyl sulphide (DMS) are the only volatile sulfur compounds originally found in this work as consequence of biochemical reactions during ripening. Therefore only their development was discussed; however, because CS2 is not considered as a relevant contributor to cheese aroma it is not going to be part of the analysis. Calibration curves were calculated for H2S, DMS and MeSH; nonetheless only for the first two compounds acceptable linear correlation was achieved, reason why the interpretation of results for MeSH was done based on the area ratio respect the internal standard EMS. The development of DMS and MeSH was similar to the last two studies; it was steady during the observation period, resulting in increasing concentration of these compounds over time. In the case of H2S there was not an evident development pattern, but a clear higher concentration for those samples made with adjunct culture was observed. Most of adjunct cultures for Cheddar cheese are NSLAB consisting mainly of lactobacillus sp.; however, other strains of lactoccus lactis sp. and Brevibacterium linens are included in this group (C. M. Lynch et al. 1996; C. M. Lynch et al. 1999; P. Fox, McSweeney, and Lynch 140 1998; B. Weimer, Seefeldt, and Dias 1999). Indeed, in comparison to the industrial strains, the wild strain varieties of lactoccus and lactobacillus are more dependent on their own enzymatic amino acid activity to survive, consequently their capacity to synthesize their own amino acids is reflected on the amount of flavor compounds (such as VSC’s) that can be found in those cheeses manufactured with them in parallel with the starter culture. Therefore, it is possible to base this analysis on the fact that starter culture, adjunct bacteria and NSLAB all contribute to the formation of methanethiol (Forde and Fitzgerald 2000; M. El Soda, Madkor, and Tong 2000; Seefeldt and Weimer 2000). Actually, the results of this work confirm that, and it can be seen in figure 45 that the samples made with adjunct culture have significant higher concentration of methanethiol. This can be explained by the many metabolic pathways that adjunct culture can use to metabolize methionine, from which the cystathionine is the principal one for most of the strains (Ayad et al. 1999; Seefeldt and Weimer 2000). Nonetheless, the synthesis of MeSH via methionine γ-lyase is more efficient and is characteristic of B. Linens. As a matter of fact it has been reported that lactococci can grow in absence of cysteine but not methionine, and lactobacilli could not grow in the absence of either, which indicate that both types of bacteria are auxothropic for both amino acids, but their growth requirements are strain specific (Chopin 1993; Seefeldt and Weimer 2000). Another fact that reinforces the last hypothesis is that these adjunct bacteria have potent proteolytic systems composed by extracellular proteinase, endopeptidases, exopeptidases and amino peptidases that increase proteolysis and remove amino acids from the amino terminal ends of various peptides, which result in the increment of free amino acids such as methionine (that was showed and explained in last section), which in high concentrations inhibits cystathionine-lyase activity of Lc lactis spp. cremoris because the enzymes responsible for this activity are biosynthetic and methionine inhibit their expression (Dias and Weimer 1998). Therefore, it is possible to state that in cheese made with adjunct culture, methanethiol is mainly provided by other strains of Lactococci, less susceptible to methionine concentration in the growth medium, or by Lactobacilli, and/or B. Linens, which methionine presence had little or no effect on cystathionine-lyase or methionine-γ-lyase activity. Essentially the same principle describe above can be used to explain the higher levels of H2S for the samples containing adjunct culture in figure 44. In addition to the H2S produced from 141 the sulfhydryl groups from thermal breakdown of cysteine, consequence of the denaturation of whey protein that coagulate with caseins after thermal treatment of cheese milk, more H 2S is generated when methionine is produce from cysteine through the β-elimination reaction (Dobric et al. 2000; María Fernández et al. 2000), which is a reaction reproducible by other lactococcus strains and certain genetic variants of lactobacillus helveticus, one of the possible adjunct cultures (Smit, Smit, and Engels 2005; Lee et al. 2007). Regarding DMS, it is way more difficult to explain the reason why there is and evident increment in its concentration for the samples containing adjunct culture since its generation is still poorly understood and unclear even for the starter culture strains. Figure 41 VSC chromatogram for Cheese with adjunct culture Figure 43 Development of MeSH in Cheese made with and without adjunct culture Figure 42 Development of H2S in Cheese made with and without adjunct culture 142 Figure 44 Development of DMS in Cheese made with and without adjunct culture Effect of treatment on Proteolysis The results of this work indicate that the methodology for monitoring proteolysis and classifying Cheddar cheeses according to maturity and treatment can be based on measurements of Total Kjeldahl Nitrogen (TKN) and a principal component analysis of the RP-HPLC peptide profile of the WSN fraction. Contrary the Urea PAGE results do not present evidence of differences in order to discriminate sample. Soluble Nitrogen Fractions and TKN Based on the same fractionation scheme describe in the last two chapters (Sousa, Ardö, and McSweeney 2001; Voigt et al. 2012), it can be seen that nitrogen concentrations increased during ripening time, and was different regarding the content of adjunct culture. Figures 45 and 46 show that for all samples the nitrogen levels for the WSN fraction was the highest in comparison to the TCA-SN and PTA-SN fractions. Similar to the cases of the results in the last 2 chapters, the values collected were about 4 and 8 times higher respectively. And after an observation period of 10 months, nitrogen concentrations were at least the double of those by the beginning of the assessment Regarding the TKN levels of WSN, in the figure 47 is possible to appreciate that during the second and fourth month, the levels are higher for the samples made with adjunct culture, but by the sixth month the difference became smaller and was sustained during the next 4 months. 143 However, it is clear that the samples containing adjunct culture have the tendency of concentrating more nitrogen, and despite this fraction has been recognized as an index of primary proteolysis (Bansal, Piraino, and McSweeney 2009), it is evident the influence of NSLAB or adjunct culture enzymes in primary proteolysis, and although it is expected to be expressed later in the ripening stage when numbers of LAB decrease and the adjunct culture become dominant, its role in early proteolysis can be seen. However it is important to keep in mind the TKN of the WSN fraction is a percentage of total Nitrogen and has no specific information about the composition of the WSN fraction; therefore, these result could be a synergic effect from different breakdown products and proteolytic agents. Nonetheless, the results are in agreement with the works of Habibi-Najafi, Lee, and Law 1996; Laan et al. 1998; El-Soda, Madkor, and Tong 2000; Madkor, Tong, and El Soda 2000; that report higher intracellular activity after de addition of lactobacilli strains and higher concentration of water soluble nitrogen. In figure 48, it can be seen that the levels of TCA-SN indicate differences in the proteolysis development between samples containing adjunct cultures and the ones that do not. Additionally, this figure show as well that the nitrogen levels in this fraction after 10 months are about twice those by 2 months. So it is possible to appreciate how the difference in the nitrogen extracted was larger as the extent of proteolysis was higher. This fraction is a selective precipitation by TCA to fractionate peptides in the WSN. The peptide solubility is related to hydrophobicity (Yvon, Chabanet, and Pélissier 1989), therefore, it is expected that this fraction is rich in medium sized and small peptides, amino acids, which can have low and medium hydrophobicity (T.K. Singh et al. 1994). Thus, it is possible that many peptides derived from the N-terminal half of αs1-casein and the N-terminal half of βcasein might be extracted in this fraction; nonetheless, due to it is not known what was the nature of the adjunct culture used, it is difficult to suggest a relation between the role of the bacteria with the extra amount of nitrogen extracted, because it can be attributed to specificity of the adjunct culture or to the rate of cell autolysis. In addition it might be possible that the adjunct culture can be a combination of two NSLAB cultures that for sure will increase the rate of proteolysis. Another interesting observation is that during the first 4 months the 144 amounts of total nitrogen are similar, which can be evidence of competition for substrate between the starter and adjunct culture enzymes. Regarding the PTA-SN fraction, it is possible to see that the amount of nitrogen extracted is directly related to the extension of the ripening of samples, which increases along with time. Also it can be seen an increment of 3 and 4 times the concentration at 2 months by the end of the observation period (10 months) for the samples without and with adjunct culture respectively. This observation is in agreement with the work of (C. M. Lynch et al. 1996), which reports that the presence of lactobacilli led to increase the levels of small peptides and amino acids, which is basically the composition of the PTA fraction, very small peptides (<15 kDa) and amino acids of approximately 600 Da (Aston and Dulley 1982). However, as in the case of the WSN fraction, this fraction is related to the extent of secondary proteolysis and has no specific information about the composition. Nonetheless, by focusing the attention on the first 4 months, it might be supported the observation in the TCA fraction about the competition for the available substrate between starter and adjunct culture enzymes. Alike, the TCA and WSN fraction it is really difficult to establish any relation between the results obtained and the specific role of the adjunct culture without knowing its nature, because as it is mentioned above, difference can be attributed to viable or to non-viable cells, or to the combination of both. From the results of these fractions, it is reasonable to affirm that it is possible to establish a fair comparison between samples made with and without adjunct culture by using the proposed fractionation scheme. Figure 45 TKN fractions cheese with adjunct culture 145 Figure 46 TKN fractions cheese without adjunct culture Figure 47 WSN Cheese with Adj culture Vs Cheese without Figure 48 TCA-SN Cheese with Adj culture Vs Cheese without 146 Figure 49 PTA-SN Cheese with Adj culture Vs Cheese without Peptide analysis by RP-HPLC Based on the same procedure mentioned in the last 2 chapters for analyzing the raw data from the peptide profile obtained by RP-HPLC by employing a PCA (Smith and Nakai 1990; Benfeldt and Sørensen 2001; Piraino, Parente, and McSweeney 2004), it was possible to establish a fair assessment of the proteolysis of Cheddar cheese samples prepared with and without the addition of adjunct culture with a randomize design based in duplicate observations. The PCA results lead to a model with three principal components (PCs) that explain the 83.4% of the variability of the data. The only combination that exposed correlation is the one for PC1 Vs PC2, which explain the 74.9% of the variability of the data and are related to the discrimination of samples according age and adjunct culture treatment respectively as it can be seen in figures 53 and 54. This indicate that the higher scores in PC1 correspond to samples with longer ripening time while the higher scores in PC2 correspond to samples made with addition of secondary culture, which means that proteolysis is faster for this cheeses. In addition, the loading plot in the figure 55 shows high score for the segments 12-20, 20-25, 3540, 40-45 and 60-65 with regarding to PC1, which suggest that the relative amount of peptides and amino acids eluting in this zones increase during ripening, whereas the scores for the segments 25-30, 30-35, 45-50,50-55 and 55-60 indicate that the amount of peptides and amino acids in these segments reduces over time. Additionally the scores for PC2 point out a increase 147 in the material eluting in the segments 12-20, 20-25, 35-40, 40-45, 45-50 and 50-55, and reveal a reduction for the segments 25-30, 30-35, 55-60 and 60-65 as consequence of the adjunct culture treatment. Regarding intervals 12-20 and 20-25, the results in the loading plot indicate that the amount of peptides and amino acids eluting in these segments increased as consequence of the maturation and due to the addition of adjunct culture. This interpretation can be explain based on the fact that proteolytic activity of secondary cultures complement the activity of the starter, producing peptides with similar molecular weight and free amino acids (C.N. Lane and Fox 1996a; C. M. Lynch et al. 1996; C. M. Lynch et al. 1999). Normally, these segments are mostly composed by amino acids and small hydrophilic peptides product from the hydrolysis of αs1-CN and κ-caseins followed by the action of the cell envelope proteinase (CEP) and amino peptidase from LAB, resulting in peptides such as αs1-CN(f1-9) and αs1-CN(f1-13) that accumulate during ripening, and others like αs1-CN (f1-8), αs1-CN(f8-23), αs1-CN(f923), αs1-CN(f14-23), αs1-CN(f10-?), αs1-CN(f17-?), αs1-CN(f18-?) and αs1-CN(f11-?) that are N-terminal fragments from αs1-CN. Also, amino acids such as glutamic acid, valine, isoleucine, leucine, lysine and proline can be found in this zone, which reveal the activity of amino peptidases (Pep A), (Pep N) and proline iminopeptidase (T.K. Singh et al. 1994; Manuela Fernández, Singh, and Fox 1998; Andersen, Ardö, and Bredie 2010). Therefore, it is possible to suggest that the increment could be related to adjunct culture of lactoballi, which have 5 to 100 times higher intracellular enzyme activity (Habibi-Najafi, Lee, and Law 1996; Laan et al. 1998). In addition if the strain used has high peptidolytic potential with low acidification ability, high levels of proteolysis can be expected (El-Soda, Madkor, and Tong 2000). Contrary, the segments 25-30 and 30-35 got negative loadings for the PC1 and PC2, which means that peptides eluting in these zones decrease over time and that the addition of the secondary culture, apparently decreases their rate of proteolysis. This observation has to do with the breakdown of αs1-CN, αs2-CN, αs1-CN (f24-199) and β-CN peptides (Tove M. I. E. Christensen, Kristiansen, and Madsen 1989; Tanoj K. Singh, Fox, and Healy 1995; Tanoj K. Singh, Fox, and Healy 1997; Benfeldt et al. 1997), which are related to enzymatic activity that is essentially provided by Chymosin and Plasmin and has not so much relation to the 148 intracellular activity of adjunct culture. Therefore, it might be possible to say that the relative decrease in the amount of eluents in these sections due to the adjunct culture can be related to a general increase in the total amount of peptides. Additionally, in spite of the feasible early accumulation of peptides such as αs1-CN (f85-91), αs1-CN (f11-?), αs2-CN (f170-?) and αs1CN (f175-182) resulting from the action of CEP and activity of endopeptidases (Pep O, Pep F), it might be possible that the decreased amount of peptides in the 30-35 segment is related to faster depletion of available substrate as consequence of the competition between the proteolytic systems of LAB and the adjunct culture . The intervals 35-40 and 40-45 exhibit fairly positive loading scores in relation to the PC1, and slightly positive scores regarding PC2. This suggests that the relative amount of peptides in these fractions rise with the cheese age and slightly increased due to the addition of adjunct culture. Once again based on previous studies, these hydrophobic peptides could be: 1) αs1CN(f93–?), αs1-CN(f24–30), αs1-CN(f26–32), αs1-CN(f26–34) resulting from the hydrolysis of the peptide αs1-CN(f24–199) by Chymosin, CEP and aminopeptidase in the segment; 2) αs2-CN(f204–207), which is a C-terminal residue and product of lactooccal CEP (Fox et al., 1994); and 3) β-CN(f45-52), as well reported as a product of hydrolysis by CEP and aminopeptidase, and traces of γ-caseins (Tove M. I. E. Christensen, Kristiansen, and Madsen 1989; T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1995; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998). In addition, some amounts of amino acids related to bitterness such as phenylalanine and histidine might be present in the last part of the last segment (Manuela Fernández, Singh, and Fox 1998; Andersen, Ardö, and Bredie 2010). This makes sense since it has been reported that peptides eluting in this region of the chromatogram correspond to high molecular mass molecules or molecules that contain aromatic amino acids, which are characterized for being very hydrophobic (Gripon et al. 1991; Gomez et al. 1997), and for eluting late in the peptide profiling by RP-HPLC and gel filtration of the water soluble extract (Allan J. Cliffe, Marks, and Mulholland 1993). Therefore, a minor increase in levels of proteolysis for these fractions has to do probably with the complementary and supplementary enzymatic activity of adjunct culture, which might provide homofermentative mesophilic lactobacilli that cause more bitterness (Habibi-Najafi, Lee, and Law 1996) and consequently increased these values. Also some adjunct strains have high 149 peptidolytic activity (El-Soda, Madkor, and Tong 2000). However, it is important to keep in mind that the role of these organisms in amino acid production requires more study. Moreover, something really interesting is the reinforced reducing effect of material eluting in the intervals 45-50 and 50-55 visible as negative scores for PC1 and positive for PC2, which indicates that the relative amount of peptides and amino acids in this part of the chromatogram reduces over time and is intensified by the addition of adjunct culture. As it was mentioned above, material eluting in the last section of the chromatogram is related to bitter fractions (Gripon et al. 1991; Allan J. Cliffe, Marks, and Mulholland 1993; Gomez et al. 1997), and in addition it was reported by Singh et al. 1994; Manuela Fernández, Singh, and Fox 1998, that the last fraction of the chromatogram mainly contained tryptophan, another aromatic amino acid related to bitterness in cheese. Therefore this behavior can by basically explained based on the findings of Habibi-Najafi, Lee, and Law 1996 and El-Soda, Madkor, and Tong 2000, which points out the debitterase action and high potential to degrade hydrophobic aminoacids and reducing bitter off-flavor of adjunct cultures, specially strains of L Helveticus, and mixed strains of lactococci and Br. linens. Finally the low scores in the remaining segments respect to PC1 and practically close to 0 for PC 2, indicate that enzymatic activity associated to the reduction of the material eluting in this zone, is apparently unaffected or less affected by the addition of adjunct culture. Indeed, the enzymatic activity in this zone might be related to the catabolism of free amino acids. Nonetheless because of the uncertainty associated to the type of adjunct culture employed in the manufacture of the samples, it is really difficult to establish any accurate association. However, it is important to recall that it is a common industrial practice to use isolated cultures from good quality cheese where the criteria for selection is still vague, resulting in potentially non reproducible results, which reminds the importance and need to identify the proteolytic and lipolytic systems to be used. . 150 Figure 50 Scree plot of cheese with and without adjunct culture Figure 51 Score plot of cheese with and without adjunct culture (age) 151 Figure 52 Score plot of cheese with and without adjunct culture (culture) Figure 53 Loading plot of cheese with and without adjunct culture 152 Figure 54 Peptide profile cheese with adjunct culture (A and B) Vs Not (C and D) cheese, for 2 and 10 months A B C D Electrophoresis From the results of the Urea-PAGE it is possible to see that αs1-casein and β-casein are breaking down during ripening. Indeed it can be seen that the degradation of αs1-casein is faster and stronger than that of β-casein. However the primary proteolysis pattern for the different observation is similar for αs1-casein and β-casein for the cheeses made with and without adjunct culture. In addition, in the figures 57 to 59 it is possible to see a parallel increase of those bands corresponding to γ-caseins over time. Therefore it is possible to say that proteolysis patterns found are related to the activity of Chymosin and Plasmin rather than associated to the activity of the starter or the adjunct culture 153 Figure 55 Urea PAGE for ripening of cheese made with and without adjunct culture β -CN(f106-209) β -CN(f29-209) β -CN(f108-209) β -CN αs1 -CN αz1-CN(f24-199) αs1-CN(f121-199) 2M-A 2M-NA 4M-A 4M-NA 6M-A 6M-NA 154 Figure 56 Urea PAGE for ripening of cheese made with and without adjunct culture 6M-A 6M-NA 8M-A 8M-NA 10M-A 10M-NA CONCLUSION The present study demonstrates that the use of FFA profile, VSC’s profile, measurement of the level of the Total Kjeldahl nitrogen for the WSN, TCA-SN and PTA-SN fractions, and the PCA of the RP-HPLC peptide profile of the WSN fraction are effective tools and ripening indices to differentiate Cheddar cheese samples regarding to their age and the adjunct culture treatment. The urea-PAGE was effective to differentiate samples by their age; nonetheless it is clear that it is not sensitive enough to detect differences related to the addition of adjunct culture. On the other hand, , the results of levels of nitrogen for all the 3 fractions analyzed demonstrated that proteolysis is faster for cheeses made with adjunct culture. This was supported by the PCA model obtained which suggests differences caused by the role of the secondary culture as supplement to the starter culture during ripening. Lipolysis was slower for cheese produced without adjunct culture, which showed lower levels of individual FFA. 155 The amounts of DMS, H2S and MeSH showed differences between treatments and the tendency to accelerate the catabolism of sulfur containing amino acids by including adjunct culture. Once again, the results for DMDS and DMTS suggest that they are artifacts from extraction and separation procedures rather than metabolites from the ripening of Cheddar cheese. REFERENCES Andersen, Lene T., Ylva Ardö, and Wender L.P. Bredie. 2010. “Study of Taste-active Compounds in the Water-soluble Extract of Mature Cheddar Cheese.” International Dairy Journal V20 (8): pp. 528–536. Aston, J. W., and John R. Dulley. 1982. “Cheddar cheese flavor review.” Australian Journal of Dairy Technology V37 (2): pp. 59–64. Ayad, Eman H.E, Annette Verheul, Catrienus de Jong, Jan T.M Wouters, and Gerrit Smit. 1999. “Flavour Forming Abilities and Amino Acid Requirements of Lactococcus Lactis Strains Isolated from Artisanal and Non-dairy Origin.” International Dairy Journal V9 (10): pp. 725–735. Bansal, N, Paolo Piraino, and P. L. H. McSweeney. 2009. “Determination of Proteolysis in Cheese.” In Handbook of Dairy Foods Analysis, pp. 405–417. CRC Press. Benfeldt, Connie, John Sørensen, Katrine H. Ellegård, and Torben E. Petersen. 1997. “Heat Treatment of Cheese Milk: Effect on Plasmin Activity and Proteolysis During Cheese Ripening.” International Dairy Journal 7 (11): 723–731.. Benfeldt, Connie, and John Sørensen. 2001. “Heat Treatment of Cheese Milk: Effect on Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4–7): pp. 567–574. Burbank, Helen M., and Michael C. Qian. 2005. “Volatile Sulfur Compounds in Cheddar Cheese Determined by Headspace Solid-phase Microextraction and Gas Chromatograph-pulsed Flame Photometric Detection.” Journal of Chromatography A V1066 (1–2): pp. 149–157. Chopin, Alain. 1993. “Organization and Regulation of Genes for Amino Acid Biosynthesis in Lactic Acid Bacteria.” FEMS Microbiology Reviews V12 (1–3): pp. 21–37. Christensen, Tove M. I. E., Kristian R. Kristiansen, and Jesper S. Madsen. 1989. “Proteolysis in Cheese Investigated by High Performance Liquid Chromatography.” Journal of Dairy Research V56 (5): pp. 823–828. Cliffe, Allan J., Jonathan D. Marks, and Francis Mulholland. 1993. “Isolation and Characterization of Non-volatile Flavours from Cheese: Peptide Profile of Flavour Fractions from Cheddar Cheese, Determined by Reverse-phase High-performance Liquid Chromatography.” International Dairy Journal V3 (4–6): pp 379–387. Dias, Benjamin, and Bart Weimer. 1998. “Conversion of Methionine to Thiols by Lactococci, Lactobacilli, and Brevibacteria.” Applied and Environmental Microbiology V64 (9): pp. 3320–3326. Dobric, Nada, Gäetan K.Y. Limsowtin, Alan J. Hillier, Nicholas P.B. Dudman, and Barrie E. Davidson. 2000. “Identification and Characterization of a Cystathionine Β/γ-lyase from Lactococcus Lactis Ssp. Cremoris MG13631.” FEMS Microbiology Letters V182 (2): pp. 249–254. 156 El Soda, Morsi A. 1993. “The Role of Lactic Acid Bacteria in Accelerated Cheese Ripening.” FEMS Microbiology Reviews V12 (1–3) (September): pp. 239–251. El Soda, M., S.A. Madkor, and P.S. Tong. 2000. “Adjunct Cultures: Recent Developments and Potential Significance to the Cheese Industry.” Journal of Dairy Science V83 (4): pp. 609–619. El-Soda, M., S. A. Madkor, and P. S. Tong. 2000. “Evaluation of commercial adjuncts for use in cheese ripening: 4. Comparison between attenuated and not attenuated lactobacilli.” Milchwissenschaft V55 (5): pp. 260–263. Fernández, Manuela, Tanoj K. Singh, and Patrick F. Fox. 1998. “Isolation and Identification of Peptides from the Diafiltration Permeate of the Water-Soluble Fraction of Cheddar Cheese.” Journal of Agricultural and Food Chemistry V46 (11): pp. 4512–4517. Fernández, María, Wim van Doesburg, Ger A. M. Rutten, Joey D. Marugg, Arno C. Alting, Richard van Kranenburg, and Oscar P. Kuipers. 2000. “Molecular and Functional Analyses of themetC Gene of Lactococcus Lactis, Encoding Cystathionine β-Lyase.” Applied and Environmental Microbiology V66 (1): pp. 42–48. Ferreira, A.D., and B.C. Viljoen. 2003. “Yeasts as Adjunct Starters in Matured Cheddar Cheese.” International Journal of Food Microbiology V86 (1-2): pp. 131–140. Forde, Amanda, and Gerald F Fitzgerald. 2000. “Biotechnological Approaches to the Understanding and Improvement of Mature Cheese Flavour.” Current Opinion in Biotechnology V11 (5): pp. 484–489. Fox, P.F. 1989. “Proteolysis During Cheese Manufacture and Ripening.” Journal of Dairy Science V72 (6): pp. 1379–1400. Fox, P.F., and L. Stepaniak. 1993. “Enzymes in Cheese Technology.” International Dairy Journal V3 (4–6): pp. 509–530. Fox, P. F., J. M. Wallace, S. Morgan, C. M. Lynch, E. J. Niland, and J. Tobin. 1996. “Acceleration of Cheese Ripening.” Antonie Van Leeuwenhoek 70 (2): 271–297. Fox, PF, PLH McSweeney, and CM Lynch. 1998. “Significance of Non-starter Lactic Acid Bacteria in Cheddar Cheese.” Australian Journal of Dairy Technology V53 (2): pp. 7– 8. Fox, P. F., Law J, P. L. H. McSweeney, and Wallace J.M. 1999. “Biochemistry of Cheese Ripening.” In Cheese: Chemistry, Physics and Microbiology, pp. 382–450. Springer. Gomez, Maria J., Sonia Garde, Pilar Gaya, Margarita Medina, and Manuel Nuñez. 1997. “Relationship Between Level of Hydrophobic Peptides and Bitterness in Cheese Made from Pasteurized and Raw Milk.” Journal of Dairy Research V64 (02): pp. 289–297. Gripon, Jean-Claude, V Monnet, G Lambert, and M. J. Desmazeaud. 1991. “Microbial enzymes in cheese ripening.” In Food enzymology, vol 1. Habibi‐Najafi, Mohammad B., Byong H. Lee, and Barry Law. 1996. “Bitterness in Cheese: A Review.” Critical Reviews in Food Science and Nutrition V36 (5): pp. 397–411. Laan, Harry, Saw Eng Tan, Paul Bruinenberg, Gaëtan Limsowtin, and Malcolm Broome. 1998. “Aminopeptidase Activities of Starter and Non-Starter Lactic Acid Bacteria Under Simulated Cheddar Cheese Ripening Conditions.” International Dairy Journal V8 (4): pp. 267–274. Lane, C.N., and P.F. Fox. 1996a. “Contribution of Starter and Adjunct Lactobacilli to Proteolysis in Cheddar Cheese During Ripening.” International Dairy Journal V6 (7): pp. 715–728. Lee, Won-Jae, Dattatreya S. Banavara, Joanne E. Hughes, Jason K. Christiansen, James L. Steele, Jeffery R. Broadbent, and Scott A. Rankin. 2007. “Role of Cystathionine βLyase in Catabolism of Amino Acids to Sulfur Volatiles by Genetic Variants of 157 Lactobacillus Helveticus CNRZ 32.” Applied and Environmental Microbiology V73 (9): pp. 3034–3039. Lynch, C.M., P.L.H. McSweeney, P.F. Fox, T.M. Cogan, and F.D. Drinan. 1996. “Manufacture of Cheddar Cheese with and Without Adjunct Lactobacilli Under Controlled Microbiological Conditions.” International Dairy Journal V6 (8–9): pp. 851–867. Lynch, C.M., D.D. Muir, J.M. Banks, P.L.H. McSweeney, and P.F. Fox. 1999. “Influence of Adjunct Cultures of Lactobacillus Paracasei Ssp. Paracasei or Lactobacillus Plantarum on Cheddar Cheese Ripening.” Journal of Dairy Science V82 (8): pp. 1618–1628. Madkor, S.A., P.S. Tong, and M. El Soda. 2000. “Ripening of Cheddar Cheese with Added Attenuated Adjunct Cultures of Lactobacilli.” Journal of Dairy Science V83 (8): pp. 1684–1691. Manning, D. J., Helen R. Chapman, and Zena D. Hosking. 1976. “The Production of Sulphur Compounds in Cheddar Cheese and Their Significance in Flavour Development.” Journal of Dairy Research V43 (2): pp. 313–320. McSweeney, P.L.H., P.F. Fox, J.A. Lucey, K.N. Jordan, and T.M. Cogan. 1993. “Contribution of the Indigenous Microflora to the Maturation of Cheddar Cheese.” International Dairy Journal V3 (7): pp. 613–634. McSweeney, Paul L.H., and Maria José Sousa. 2000. “Biochemical Pathways for the Production of Flavour Compounds in Cheeses During Ripening: A Review.” Le Lait V80 (3): pp. 293–324. Piraino, Paolo, Eugenio Parente, and Paul L. H. McSweeney. 2004. “Processing of Chromatographic Data for Chemometric Analysis of Peptide Profiles from Cheese Extracts: A Novel Approach.” Journal of Agricultural and Food Chemistry 52 (23): 6904–6911. Seefeldt, K.E., and B.C. Weimer. 2000. “Diversity of Sulfur Compound Production in Lactic Acid Bacteria.” Journal of Dairy Science V83 (12): pp. 2740–2746. Singh, T.K., P.F. Fox, P. Højrup, and A. Healy. 1994. “A Scheme for the Fractionation of Cheese Nitrogen and Identification of Principal Peptides.” International Dairy Journal V4 (2): pp. 111–122. Singh, Tanoj K., Patrick F. Fox, and Áine Healy. 1995. “Water-Soluble Peptides in Cheddar Cheese: Isolation and Identification of Peptides in the Diafiltration Retentate of the Water-Soluble Fraction.” Journal of Dairy Research V62 (4): pp. 629–640. Singh, Tanoj K., Patrick F. Fox, and Áine Healy. 1997. “Isolation and Identification of Further Peptides in the Diafiltration Retentate of the Water-soluble Fraction of Cheddar Cheese.” Journal of Dairy Research V64 (3): pp. 433–443. Smit, Gerrit, Bart A. Smit, and Wim J.M. Engels. 2005. “Flavour Formation by Lactic Acid Bacteria and Biochemical Flavour Profiling of Cheese Products.” FEMS Microbiology Reviews V29 (3): pp. 591–610. Smith, A. M., and S. Nakai. 1990. “Classification of cheese varieties by multivariate analysis of HPLC profiles.” Canadian Institute of Food Science and Technology Journal V23 (1): pp. 53–58. Sousa, M.J, Y Ardö, and P.L.H McSweeney. 2001. “Advances in the Study of Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4-7): pp. 327–345. Urbach, Gerda. 1993. “Relations Between Cheese Flavour and Chemical Composition.” International Dairy Journal V3 (4–6): pp. 389–422. Voigt, Daniela D., François Chevalier, John A. Donaghy, Margaret F. Patterson, Michael C. Qian, and Alan L. Kelly. 2012. “Effect of High-pressure Treatment of Milk for 158 Cheese Manufacture on Proteolysis, Lipolysis, Texture and Functionality of Cheddar Cheese During Ripening.” Innovative Food Science & Emerging Technologies V13 (0): pp. 23–30. Wallace J.M., and Fox P.F. 1997. “Effect of Adding Free Amino Acids to Cheddar Cheese Curd on Proteolysis, Flavour and Texture Development.” International Dairy Journal V7 (2): pp. 157–167. Weimer, Bart, Kimberly Seefeldt, and Benjamin Dias. 1999. “Sulfur Metabolism in Bacteria Associated with Cheese.” Antonie Van Leeuwenhoek 76 (1): 247–261. De Wit, M., G. Osthoff, B.C. Viljoen, and A. Hugo. 2005. “A Comparative Study of Lipolysis and Proteolysis in Cheddar Cheese and Yeast-inoculated Cheddar Cheeses During Ripening.” Enzyme and Microbial Technology V37 (6) (November 1): pp. 606–616. Yvon, Mireille, Claire Chabanet, and Jean-Pierre Pélissier. 1989. “Solubility of Peptides in Trichloroacetic Acid (TCA) Solutions Hypothesis on the Precipitation Mechanism.” International Journal of Peptide and Protein Research V34 (3): pp. 166–176. 159 CHAPTER 5 COMPARATIVE CHEMICAL ANALYSIS OF GOOD AND WEAK CHEDDAR CHEESE DURING RIPENING Lemus Muñoz, Freddy Mauricio, Qian Michael Department of Food Science and Technology, Oregon State University Corvallis Oregon 97331 160 ABSTRACT Cheddar cheese samples graded as good and/or weak by a trained panel were analyzed during ripening. Proteolysis was studied by a fractionation scheme, resulting in an insoluble fraction analyzed by urea polyacrylamide gel electrophoresis (Urea-PAGE), and a soluble fraction which was further investigated through water soluble nitrogen (WSN), trichloroacetic acid soluble nitrogen (TCA-SN) and phosphotungstic acid soluble nitrogen (PTA-SN) analyzed by total Kjeldahl nitrogen content (TKN). Reversed phase high performance liquid chromatography (RP-HPLC) was used to study the peptide profile of the water soluble fraction. Lipolysis was studied by levels of individual free fatty acids determined through gas chromatography-flame ionization detection (GC-FID) after isolation employing solid phase extraction (SPE). Volatile sulfur compounds were studied using head space solid phase microextraction (SPME) coupled with gas chromatography-pulsed flame photometric detection (PFPD). It was found that Urea-PAGE is capable to differentiate samples according their age, but it could not be used to discriminate samples regarding their quality. Nonetheless, measurements of total Kjeldahl Nitrogen (TKN) of the WSN, TCA-SN, and PTA-SN fractions, and the principal component analysis of the RP-HPLC peptide profile of the WSN fraction, revealed differences in the rate and pattern of proteolysis for the samples. Good cheese, developed higher level of total nitrogen for the WSN, TCA-SN and PTA-SN fractions, indicating that primary and secondary proteolysis were faster for these samples during ripening. It was obtained a PCA model with 3 principal components that accounted for the 80.7% of the variability from data collected. This model discriminate the samples according age and quality, suggesting the samples undergo more or faster proteolysis. In addition, FFA profiles demonstrated higher levels of low and medium chain free fatty acids for good cheese, which suggest faster lipolysis during ripening. The Volatile Sulfur Compounds (VSC) analysis showed higher levels of DMS and MeSH and lower levels of H2S, suggesting slower catabolism of sulfur containing amino acids in weak cheese. 161 INTRODUCTION The production of Cheddar cheese with constant quality during the year is limited due to natural variation of milk composition or the extent of the enzymatic activity during ripening. However, in terms of homogeneity in order to satisfy customer expectation and to assure a reliable production, it is necessary to document and understand the chemical composition differences related to samples characterized as good or weak according to a experienced sensory panel. As a matter of fact, the traditional scope to assess quality based on off-flavor and defects (O’Shea, Uniacke-Lowe, and Fox 1996), could be complemented by monitoring biochemical changes occurring during ripening, which also allow to identify variations in manufacturing practices (M E Carunchia Whetstine et al. 2007; J. M. Lynch, Barbano, and Fleming 2002) resulting in perceptible disparity of flavor and texture in the ready to sale product. Indeed, currently most of the assessment of cheddar cheese quality is done by trained sensory panels that expensive, time consuming and essentially depend on the presence or absence of defects, leading to results that are subjective rather than objective. Thus, better grading implies the use of accurate measurements and reliable instrumental methods to predict and determine the flavor quality of cheese. Cheddar cheese flavor is a balance of several volatile and non-volatile sapid compounds (Engels et al. 1997; Curioni and Bosset 2002). The volatile fraction contributes to its aroma and the water soluble fraction is responsible for its taste (Aston and Dulley 1982; Aston and Creamer 1986). Thus sample differentiation based on proteolysis and lipolysis implies the separation, characterization and quantification of peptides, amino acids, free fatty acids and another key volatile compounds using chromatographic methods such as RP-HPLC, GC-MS, GC-PFPD and GC-FID, electrophoretic methods, and other emerging technologies such as Fourier transform infrared (FT-IR) spectroscopy (Smith and Nakai 1990; Dimos 1992; Subramanian, Harper, and Rodriguez-Saona 2009). It has been reported significant correlations between levels of pH 4.6-soluble nitrogen, phosphotungstic acid (PTA)-soluble nitrogen and trichloroacetic acid (TCA)-soluble nitrogen and the age, flavor intensity, and flavor development in Cheddar cheese (O’Shea, UniackeLowe, and Fox 1996; Are Hugo Pripp, Stepaniak, and Sørhaug 2000; Upadhyay et al. 2004). In addition, RP-HPLC has been effectively used to identify cheese variety and to determine its 162 age because of its high resolution power, reproducibility and low time consumption (Bican & Spahni, 1991). Thus, in order to contribute to the understanding of the flavor development of Cheddar cheese during ripening, the aim of this study was to use complementary approaches to objectively evaluate and correlate samples of different quality and age by using analytical methods such as GC-PFPD, GC-FID and HPLC, Urea-polyacrilamide gel electrophoresis and determination of nitrogen content of different soluble fractions. MATERIALS AND METHODS CHEESE SAMPLES Cheeses samples were manufactured by Tillamok county creamery according with standard protocols. Blocks of cheese of different quality were randomly selected from three consecutive manufacturing days. All samples are stored at (-37C) to stop ageing process until analysis is completed. FREE FATTY ACIDS ANALYSIS Chemicals Pentanoic acid, heptanoic acid, nonanoic acid, undecanoic acid, and heptadecanoic acid were used as internal standards, they were purchased from Eastman (Rochester, N.Y., U.S.A). Butanoic acid, hexanoic acid, octanoic acid, decanoic acid, dodecanoic acid, tetradecanoic acid, 9-tetradecanoic acid, hexadecanoic acid, 9-hexadecanoic acid, octadecanoic acid, 9octadecanoic acid, 9,12-octadecanoic acid and 6,9,12 octadecanoic acid were used for the standard stock solution, and were obtained from Aldrich Chemical Co. Inc (Milwaukee, Wisconsin, U.S.A). Heptane, Isopropanol, Sulfuric acid, anhydrous sodium sulfate, chloroform, formic acid and diethyl ether were obtained from Fisher. Extraction From each 2lb block of cheese, 100 grams were wrapped in alumina foil, frozen with liquid nitrogen during 6 minutes, and then grinded for 30 seconds to obtain a fine powder. Six grams of this previously freeze-ground cheese, 1 ml of 2N sulphuric acid and 1 ml of internal 163 standard solution (C5:0, C7:0, C9:0, C11:0 and C17:0 in 1:1 hetpane-isopropanol) were mixed with 7 grams of anhydrous sodium sulfate and 20 ml of 1:1 diethyl ether- heptane in a 40 ml amber vial using a sonicator and manual agitation. During sonication, the salt-slurry solution is initially exposed for 15 minutes, after which each vial is shake vigorously to continue with a second sonication period of 20 minutes. With a glass-Pasteur pipette, the sample extract (solvent) is transferred to an AccuBOND amino cartridge (Agilent Technologies) conditioned previously with 10 ml of heptane. After the addition of the sample, the column is washed with 5 ml of 2:1 Chloroform-Isopropanol to remove non volatile triglycerides and phospholipids using a manifold vacuum chamber. Once the washing step is complete, free fatty acids are eluted with 5ml of 2% formic acid in diethyl ether, collected in a 20 ml vial and stored in the freezer until GC analysis. Chromatography The analysis was performed using a Hewlett-Packard 5890 gas chromatograph equipped with a flame ionization detector (FID). Samples were analyzed on a DB-FFAP column (15m x 0.53mm ID, 1 m film thickness; Supelco Wax10, Supelco U.S.A). Injector and detector temperatures were 250C. Nitrogen was used as carrier gas at a flow rate of 15 ml per minute at a split ratio of 1 to 1. The oven temperature was programmed for a 2 minutes hold at 60C, raised to 230C at a rate of 8C per minute with a hold of 20 minutes at 230C. Quantitative analysis The levels of free fatty acids concentrations were calculated based on individual peak area from GC-FID response in comparison to the internal standard peak area, by using standard calibration curve of individual free fatty acid using Peak Simple software (SRI instruments, Torrance, CA). Each experimental value corresponds to the average of the 3 extraction replicates. VOLATILE SULFUR COMPOUNDS (VSC’S) Chemicals Dimethyl sulfide (DMS) was purchased from TCI America (Portland, OR, U.S.A.); gaseous methanethiol (MeSH) was obtained from Aldrich Chemical Co. Inc (Milwaukee, Wisconsin, U.S.A), and a solution was prepared by bubbling the gas into cold methanol; a H2S solution 164 was prepared by dissolving Na2S.9 H2O (Sigma Co) in acidic water stabilized with citric acid (pH 3). Extraction From each 2lb block of cheese, 100 grams were wrapped in alumina foil, frozen with liquid nitrogen during 6 minutes, and then grinded for 30 seconds to obtain a fine powder. Then one gram of this freshly prepared powder is added to a 20ml vial (formerly flushed with argon), followed by the addition of 4 ml of 1M citric acid and 20 l of the internal standard solution. After addition of sample vials were immediately sealed with screw caps with teflon-lined silicone septa. The vials used in this study were previously deactivated with DMTCS 5% solution in toluene, toluene, methanol and distillate water. The volatile sulfur compounds were extracted with an 85 m Carbox-PDMS fiber (Supelco, Bellefonte, PA, U.S.A.). Prior to use, the fiber was conditioned at 300 C for 90 minutes. The fiber was then placed into a SPME adapter of a CombiPAL autosampler (CTC analytics AG, Zwingen, Switzerland) Fitted with a vial heater/agitator. Samples were pre-equilibrated at 500 RPM at 40C for 5 minutes, and the extraction of VSC’s was done at 250 RPM at 40C for 25 minutes. The desorption time was 5 minutes and 30 seconds. Chromatography The analysis was performed using a Varian CP-3800 gas chromatograph (Varian, Walnut Creek, CA, U.S.A.) equipped with a pulsed flame photometric detector (PFPD). The separation of analytes was made using a DB-FFAP fused silica capillary column (30m, 0.32 mm ID and 1 m film thickness; Agilent, Palo Alto, CA, USA) and nitrogen as carrier gas at constant flow at 2 ml per minute. The injector temperature was 300 C and it was in the splitless mode. The oven temperature was programmed for a 3 minutes hold at 35C, raised to 150C at a rate of 10C per minute, held for 5 minute, and then heated to 220C at a rate of 20C per minute with a final hold of 3 minutes. The PFPD was held at 300 C and 450 V with the following flow rates: Air 1 at 17 ml per min, H2 at 14 ml per min, and Air 2 at 10 ml/min. The detector response signal was integrated using the software Star Workstation 6.2, Varian) 165 Quantitative analysis Matrix effect In order to retain the matrix effect during the construction of the calibration curves, cheese powder from the “youngest sample” is used. It is de-volatilized by exposure to room conditions in a hood for 2 hours. Then 1 gram of powder is added to 4 ml of 1M citric acid in a 20 ml vial and exposed to a 50C water bath for 30 mins, prior to the addition of standards and internal standard solutions. Sulfur standards and internal standard preparation Two internal standards were used for the quantification of VSC’s: ethyl methyl sulfide (EMS) for H2S, MeSH and DMS, and isopropyl disulfide (IsoProDS) for DMDS and DMTS. The concentration of the internal standard solution was 500 ppm EMS and 500 ppm IsoProDS in methanol. Calibration curves were constructed by spiking cheese samples with a range of known concentrations of H2S, MeSH and DMS. Hydrogen sulfide (H2S) was prepared by dissolving Na2S.9 H2O in acidic water (pH = 3). Different concentrations of sodium sulfide solutions were made, and the concentrations of H2S were calculated based on the amounts of salt added to the matrix. A standard solution of 100 ppm of DMS was individually prepared in cooled methanol (-15C), and dilutions were made with cooled methanol at the same temperature. The mesh standard was prepared as following: 1) newly deactivated, recently flushed with argon, and cooled vials were used; 2) The original standard solution was made by bubbling pure MeSH into cooled methanol; 3) Dilutions were prepared by taking aliquots from the original solution contained in a sealed vial, through the teflon-lined silicone septa by using a syringe. And then injecting the aliquots into new sealed vials containing proportional amount of cooled methanol through the septa; 4) 1 gr of devolatilized cheese is added to a recently flushed vial (argon was used), which is immediately flushed again; 5) simultaneous argon flushing and addition of 4ml of “free” dissolved oxygen-1M citric acid solution and quick sealing of the vials; 6) Addition of 20 l of internal standard and MeSH standard through septa. The identification of target compounds was made by comparing retention times with those of pure standards. Ratios of the square root of the standard area to the corresponding square root of the internal standard area were plotted Vs concentration ratios to 166 determine the relation between the response and concentration for the unknowns. Triplicate analysis was performed for all samples PROTEOLYSIS Chemicals Sulfuric acid was purchased from Fisher Scientific International Inc. (Pittsburgh, PA, U.S.A.); Trichloroacetic acid was purchased from Alfa Aesar (Royston, UK); and phosphotungstic acid was obtained from Aldrich Chemical Co. Inc (Milwaukee, Wis, U.S.A) Sample preparation and fractionation From each 2lb block of cheese, 60 grams are blended with 120 ml of distilled water pre-heated to 55C. The mixture is blended for 5 minutes and the homogenate is incubated at 55C for 1 hour. Then the pH is adjusted to 4.6 with 1M HCl and the mixture is centrifuged at 3000g for 30 minutes at 4C. Suspension and supernatant were filtered thoroughly 3 times through glass wool. The filtrate was safe at -20C for macro blog digestion method analysis, and RP-HPLC analysis. The insoluble pellet was frozen at -20C for further Urea-PAGE gel electrophoresis analysis. The trichloroacetic acid soluble nitrogen fraction (TCA-SN) was prepared by the addition of 25 ml of pH 4.6 soluble fraction (WSN) to 25 ml of 24% trichloroacetic acid solution. Then the mixture is equilibrated for 2 hours at room temperature and filtered through filter paper Whatman No 40 before macro blog digestion method analysis. For the phosphotungstic acid soluble nitrogen fraction (PTA-SN), 10 ml of WSN are added to 7 ml of 3.95 M H2SO4 and 3 ml of 33% phosphotungstic acid solution. Then the mixture is equilibrated overnight at 4C and filtered through filter paper Whatman No 40 before macro blog digestion method analysis. Duplicate analysis was performed for all samples. 167 Macro blog digestion (Kjeldahl Digestion) From the fractions collected an aliquot (2 ml for the Water soluble fraction, 1ml for TCA-SN and 1 ml for PTA-SN) is added into a 70 ml Kjeldahl Digestion flask with 10 ml of H2SO4 and the catalyst pellet containing 0,075 and 1,5 grams of mercuric oxide and potassium sulfate respectively. The mixture is warmed to 150 C and hold for 1 hour, then heated to 250 C and hold for 1 hour, and finally heated to 350 C and hold for 2 hours. After digestion the sample is cooled down overnight to room temperature, and diluted with distillate water to 70 ml, followed by a gentile agitation. Then a 5 ml aliquot is used to determine the nitrogen content by a rapid flow analyzer FOSS II. Reversed phase High performance liquid chromatography analysis The RP-HPLC analysis was performed using a Shimadzu 6 series liquid chromatograph (Shimadzu scientific instruments, Kyoto Japan), consisting of an autosampler, 2 pumps, a multi-wavelength spectrophotometer and a controller unit. It was used a nucleosil RP-8 analytical column (250x 4mm, 5 m particle size, 300 A pore size) and a guard column (4.6 x10 mm) from waters (Milford, MA, U.S.A.). The mobile phase consists of solvent A (0.1% TFA in deionized and vacuum filtered water) and solvent B (0.1% TFA in acetonitrile). The elution was monitored at 214nm. The following gradient elution was performed: 1) 100% solvent A for 5 minutes followed by a linear gradient to 55% solvent B (v/v); 2) elution at 55% solvent B for 6 minutes followed by a linear gradient to 60%; 3) elution at 60% solvent B for 3 minutes; 4) The column is washed using 95% solvent B during 5 minutes; 5) the column is equilibrated using 100% solvent A during 10 minutes. The sample (WSN fraction) was dissolved in solvent A (10 mg per ml) and then micro-centrifuged at 14000 RPM for 10 minutes. An aliquot of 40 l from the extract was injected to a flow rate of 0.75 ml per min. Electrophoresis Samples of the water-insoluble nitrogen fraction were dry frozen prior to analysis. Samples were dissolved in a buffer (0.75 g tris, (hydroxymethyl) methylamine, 49 gr urea and 0.4 ml concentrated HCl, 0.7 ml 2-mercaptoethanol and 0.15 gr bromophenol blue, dissolved to 100ml) and hold at 50C for 40 min. Urea-polyacrylamide gel electrophoresis (urea-PAGE) was carried out using a Protean II xi cell vertical slab unit (Bio-Rad Laboratories ltd., Hemel Hempstead, Herts, UK). Urea-PAGE gels (12.5%) were prepared and run according to the 168 method or Ardö (1999). Reagents used were obtained by Sigma-Aldrich, Inc and Fisher Scientific. STATISTICAL ANALYSIS A two-way analysis of variance (ANOVA) on data was carried out using a general linear model procedure with Turkey’s pair wise comparison at 95% confidence level, using the package Minitab 15 (minitab Ltda., Coventry, UK). RESULTS AND DICUSSION FREE FATTY ACIDS FFA The levels of lipolysis tracked by the amount of individual FFA showed differences between the cheeses graded as good and weak. However, in both cases it can be seen the tendency of FFA to increase during maturation of samples. Indeed, it is evident in figures 58, that short chain FFA tend to increase faster, reaching concentrations about 4 times the initial one during the observation period. This can be related to more mobility and better access of enzymes to these substrates, which are essentially located at the positions sn-1 and sn-3 of the triacylglicerides (Balcão and Malcata 1998). On the other hand, this behavior might suggestes that enzymatic activity is most likely dominated by lipases since they are specific for the outer ester bonds of tri or diacylglycerides (Deeth and Touch 2000a). In addition, in spite of the low change, long chain FFA increased their concentration during ripening too, displaying as well higher levels for good cheese. which seems related to lipolytic activity rather than estereolityc activity. Although the most dominant peaks in figure 57 correspond to C14:0, C16:0, C18:0 and C18:1, due to their significant lower odor thresholds (Molimard and Spinnler 1996a), they are not considered as important contributors to the overall aroma of Cheddar cheese. As a matter of fact, this relative quantitative relevance has to do with the fact that these FFA are the most abundant ones in raw milk (Yvonne F. Collins, McSweeney, and Wilkinson 2003). Contrary, in the case of short chain FFA such as C4:0 and C6:0, or FFA such as C8:0, C10:0 and C12:0, despite of their lower concentration, they contribute directly and indirectly to the characteristic aroma of Cheddar cheese, and it was evident the trend of good cheeses to developed higher 169 levels, specially for the 6, 9 and 12 month of maturation. Their rate of generation mainly depend on enzymatic activity; however, based on the fact that the most important lipolytic activity is provided by LAB enzymes, composed by esterases and lipases, it is not possible to tell which one had higher influence in the lipolysis of these samples without a study of specificity. Nonetheless, in order to explain the difference between these 2 types of samples, it might be a better approach to focus on variables that contribute to the decriment of the LAB enzymatic activity such as temperature and relative humidity of ripening rooms, and differences during salting (which for sure could have inhibitory effect since LAB enzymes are really sensitive to the salt in moisture content (Gripon et al. 1991; P. F. Fox and Stepaniak 1993)), rather than look at other manufacturing operation such as the heat treatment or standardization of milk which do not have a direct impact on intracellular enzymes. Another factor related to the lower lipolysis of weak cheese can be differences in the cell viability and autolysis of the starter strain, which indeed might suggests the use of different starter during the acidification in the manufacturing of these samples. Figure 57 FFA chromatogram week cheese 170 Figure 58 Development of individual FFA in good cheese vs weak cheese 171 172 Volatile Sulfur Compounds VSC’s In addition to free fatty acids (FFA), Volatile sulfur compounds (VSC’s) correlate with good Cheddar cheese flavor (B. Weimer, Seefeldt, and Dias 1999). When smelled alone they smell like garlic, onion, cabbage and skunk, but when they are mixed, they contribute to pleasant Cheddar cheese flavor notes. They result from decomposition of sulfur containing amino acids such as cysteine and methionine. Therefore this is another biochemical event occurring during Cheddar cheese ripening that can be use to track the extension of the maturation of samples. Indeed, It has been reported that high concentrations of H2S, MeSH, and DMS are found in Cheddar Cheese, while DMDS, DMTS and 3-methylthiopropionaldehyde have low concentrations. Other compounds such as Carbonyl sulfide, carbon disulfide and dimethyl sulfone are not important contributors (H. M. Burbank and Qian 2005). Subsequent to a thorough de-activation of injection liner and vials to prevent methanethiol (MeSH) oxidation, and an adecuate stabilization of analytes by using an organic acid buffer solution (citric acid 1M), the results from this work suggest that only hydrogen sulphide (H2S), carbon disulphide (CS2), MeSH, and dimethyl sulphide (DMS) were developed during ripening, and only small and negligible amounts of dimethyl disulphide (DMDS) and dimethyl trisulphide (DMTS) were scarcely found in the chromatograms for the samples analyzed. Which is the reason why only the development of MeSH, DMS and H2S will be discussed. 173 Standard calibration curves were calculates for each compound, and it is possible to observe in figures 60 and 61 that good linear correlation coefficients were obtained for H2S and DMS. Nonetheless it was not possible to achieve a descent calibration curve for MeSH due to its oxidation to DMDS and DMTS, therefore the interpretation of results for this last one was based on the area ratio between MeSH and the internal standard EMS, instead of using its concentration The results in figures 62, 63 and 64 show a steady development for all the compounds and samples excepting H2S good cheese, which apparently did not increase so much during the observation period. Also, this figures demonstrate that there are considerable differences of the sulfur attributes related to the quality of the samples graded by trained panel. In figure 63, hydrogen sulfide did not show a steady development for the good samples. Moreover , weak samples displayed a higher concentration of H2S. However, the difference between these types of samples during the initial stage of the ageing process was absolutely not significant, but after 9 months it became noticible and evident. Therefore, in the case of good cheese it was difficult to establish any trend during the ripening in contrast to weak cheese, which increases H2S concentration along the maturation process. In addition, based on the fact that the H2S sensory threshold is 10 ppb in water (Rychlik et al. 1998) and the concenntration for weak samples varied from 20 to 30 ppm it was possible to confirm its role as key contributor to the cheddar cheese aroma. The higher concentration for weak samples could be related to 1) differences in the cheese milk, either because of the standardization process or due to the heat treatment, which potentially can incorporate β-lactoglobulins to the casein micelles and consequently increase the amounts of cysteine, which along with methionine are the main precursors of H2S (B. Weimer, Seefeldt, and Dias 1999; Lee et al. 2007; del Castillo-Lozano et al. 2008). Indeed, (Hutton and Patton 1952; K. R. Christensen and Reineccius 1992) reported that the concentration of H2S in milk increases linearly with heating temperature; and 2) changes related to the LAB and NSLAB microflora, which supply enzymes such as methionine-γ-lyase, cystathionine-β-lyase and cystathionine-γ-lyase that produce methanethiol and H2S. 174 Regarding MeSH, figure 62 indicates a significant difference between samples, showing higher concentration for the good samples. Unfortunately, it was not possible to confirm MeSH as a potent odorant in cheese due to the lack of effectiveness in constructing a calibration curve free of its oxidation products DMDS and DMTS. Nonetheless, the analysis based on area ratios suggests a steady concentration increase for both types of samples in agreement with Urbach 1995. As in the case of H2S, the higher concentration for the good samples can be related to more enzymatic activity of LAB and NSLAB, and to a possible higher availabilty of substrate. This last possibility can be consequence of incorporation of whey protein to the casein micelles during the heating treatment of cheese milk, or due to adjustments to the protein content of the cheese milk during standardization, which from the results seems like it is different for the manufacturing process of these to types of samples. On the other hand, because it has been proposed that MeSH is enzymatically produce rather than by chemical reactions (Alting et al. 1995; Smacchi and Gobbetti 1998; Dias and Weimer 1998), it might be pertinent and more likely to attribute the different response to a raise in the L-methionine γ-lyase and/or cystathionine β-lyase and γ-lyase activity (Alting et al. 1995) (Tanaka, Esaki, and Soda 1985) resulting from, 1) a milder heating treatment of the cheese milk, which results in a higher population of indigenous bacteria, 2) addition of adjuct cultures, or 3) the use of LAB starters with different specificity and autolysis ability. In a similar way to MeSH, figure 64 shows a steady increment of the concentration of DMS for both types of samples during maturation. Aslo, it can be observed a noticeable difference between the good and weak samples, with higher concentration for the first type. Additionally, it can be seen that this sulfur compound is a key contributor to the overall flavor of Cheddar cheese since the concentrations found range between 10 and 45 ppm, amounts comparable to the ones obtained by Burbank and Qian (2008), while the sensory threshold is 2ppm in water (Rychlik et al. 1998). The higher DMS concentration for good samples is difficult to explain since its generation mechanism has not been well understood yet. And in spite that it is known that DMS concentration in raw milk is significant and is influenced by the diet of the cows (Manning et al. 1976; Forss 1979), and it can be generated from sulfhydryl group of milk proteins, mainly β-lactoglobulin and if present the milk fat globule membrane proteins, where methionine is 175 most likely the precursor for its generation after protein thermal denaturation (Datta et al. 2002), it is not possible to suggest other reasons why good cheese has higher concentration more than: 1) a completely different type of cheese milk, which could be supported by the fact that the initial observation point displays a obvios difference in comparision to those from the H2S and MeSH graphs; and 2) elevated numbers of secondary micro flora such as propionibacteria, present in the milk microflora (Baer and Ryba 1992), resulting from different approaches to the heating treatment of milk, leading to more formation of DMS from methionine (Curioni and Bosset 2002); or other bacteria sucha s Brevibacterium linens (Dias and Weimer 1998), different strains of Lactococcus lactis subsp. cremoris (Alting et al. 1995), Lactococcus lactis subsp. lactis, Lacto bacillus sp., Propionibacterium shermanii and/or the yeast Geotrichum candidum and Kluyveromyces lactis (K. Arfi et al. 2002) that could as well to provide enzymes such as thiol transferases to promote the conversion of some MeSH into DMS (Bentley and Chasteen 2004). Regarding the general absence of the sub-products DMDS and DMTS in the results of this study, it might be possible to say that based on the thorough sample preparation work and the fact that cheese has a low redox potential, -150 to -200 mV, (Donald J. Manning and Moore 1979; Green and Manning 1982), these compounds might not be generated during the ripening of curds, and instead their formation is the simple consequence of oxidation of MeSH once the cheese is exposed to a non controlled environment. Figure 59 VSC's chromatogram Good Cheese 176 Figure 60 Calibration Curve H2S Figure 61 Calibration Curve DMS Figure 62 Development of MeSH in Good and weak cheese 177 Figure 63 Development of H2S in Good and weak cheese Figure 64 Development of DMS in Good and weak cheese Effect of treatment on Proteolysis Cheddar cheese graded as “Good” and “Weak” by a trained panel was investigated during a 15 month maturation period through: 1) measurements of Total Kjeldahl Nitrogen (TKN) of the water soluble nitrogen (WSN), trichloroacetic acid soluble nitrogen (TCA-SN) and phosphotungstic acid soluble nitrogen (PTA-SN) fractions; 2) RP-HPLC peptide profiles of the WSN fraction; and 3) Urea-PAGE peptide profiles of the water insoluble nitrogen fraction. The results reveal clear differences in the rate and pattern of proteolysis. Nonetheless, from the three methods employed to evaluate the extend of proteolysis, the TKN of fractions 178 WSN, TCA-SN and PTA-SN, and the peptide profile analysis by RP-HPLC of the WSN fraction, were more effective than the Urea-PAGE electrophoresis analysis for discriminating the samples according to its quality. And as a matter of fact, it was possible to observe considerable differences in the primary and secondary proteolysis by means of these methods. Soluble Nitrogen Fractions and TKN The results from this analysis show differences in the primary and secondary protelolysis related to the quality of the samples assessed. Thus from the fractionation scheme proposed and employed by Ardö and Frederiksberg 1999, and Sousa, Ardö, and McSweeney 2001, it can be seen in figures 65 to 69, that the nitrogen concentration increased during time and it was higher for the good samples. The WSN includes all casein breakdown products, but native caseins and high molecular weight peptides; the 12% trichloracetic TCA-SN contains small peptides and FAA; and PTA-SN, which contains the smallest peptides (600 Da) and FAA (T. M. I. E. Christensen, Bech, and Werner 1991). In figures 65 and 66, it was confirmed that the nitrogen levels for the WSN fraction were the highest in comparison to TCA-SN and PTA-SN fractions. Indeed, the WSN fraction had values about 3 times larger than those of the TCA-SN fraction and 7 to 8 times higher in comparison to those of the PTA-SN fraction. On th other hand, it is possible to see in figures 67, 68 and 69 that nitrogen content by the end of the observation for the WSN and TCA-SN fractions was 1.5 to 2 times higher than those at the beginning of the observation, while in the case of the PTA-SN fraction, the nitrogen values at the end were 2 to 2.5 times bigger than those starting with. In relation to the nitrogen levels of the WSN fraction in figure 67, it can be seen that the values increased during ripening and were clearly different between samples from the very beginning of the maturation process. Also, it is possible to appreciate that the results for the “good” samples were higher than those for the “weak samples”. Therefore, based on the fact that this fraction represents the primary proteolysis (Bansal, Piraino, and McSweeney 2009), where the main enzymatic activity is proportionate by the rennet and the indigenous milk proteinases (Allan J. Cliffe, Marks, and Mulholland 1993; A.J. Cliffe, Revell, and Law 1989), it is possible to relate the difference in patterns of proteolysis to a marked disparity in 179 manufacturing operations that are capable to alterate the performance of these enzymes such as: 1) standardization of milk (adjustments of milk composition, and pasteurization of cheese milk), 2) pH achieved during acidification and at whey drainage, which determines the retention of coagulant activity, 3) and moisture content of the curd; resulting in defects such as sour and/or bitter flavor and soft and pasty body. Concerning the nitrogen levels of the TCA-SN fraction, the figure 68 shows the same behavior of the WSN fraction, where nitrogen levels increased during time and the “good” samples had higher values. Also it can be seen that the evolution of this fraction results in final values that are 1.5 bigger than the ones at the beginng of the observation. This fraction is rich in small peptides of low and medium hydrophobicity (Kuchroo & Fox 1982), mostly resulting from LAB, NSLAB and rennet enzymatic activity (Tanoj K. Singh, Fox, and Healy 1995; T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998), which is the reason why it is possible to attribute the difference found to inconsistency in processing variables such as temperature and relative humidity in ageing rooms, heat treatment of cheese milk, amount of salt added during salting and pH at salting, which affect the intracellular and extracellular proteolityc activity proportionated by LAB and NSLAB. As it is well known the PTA-SN fraction is an index of secondary proteolysis because it is mainly constituted by very small peptides (<15 kDa) and amino acids of approximately 600 Da (Aston and Dulley 1982). The results in figure 69, showed a similar trend to the other fractions; the values increased over time and were higher for the “good” samples. Even so, the proportional increase in nitrogen for this fraction was more steadfast, and the final values were about 2 to 3 times higher than those at the begin of the observation. Alike the WSN and TCA-SN fractions, the difference between samples was evident since the beginning of the observation period and as in the case of the TCA-SN fraction it could be explain through the difference in processing variable that affect LAB and NSLAB In general, it is possible to state that the fractionation scheme based on extraction of peptides with water can establish a fair comparison between samples of different quality in order to track and evaluate the extend of proteolysis. 180 Figure 65 TKN fractions Good cheese Figure 66 TKN fractions Good cheese 181 Figure 67 WSN Good Vs Weak cheese Figure 68 WSN Good Vs Weak cheese 182 Figure 69 PTA-SN Good Vs Weak cheese Peptide analysis by RP-HPLC The peptide analysis by RP-HPLC is another index of secondary proteolysis. In addition, it can be used in authenticity studies and optimization process (Upadhyay et al. 2004). In this work to evaluate the difference in the peptide profile for the different samples a principal component analysis (PCA) was used, which is a multivariate analysis tool use in descriptive statistics, to estimate the linear relationship between variables when their number is very large (Chatfield and Collins 1981). As matter of fact, the data from chromatograms was processed based on Piraino, Parente, and McSweeney 2004 work, where the complexity of profiles containing more than 70 peaks is initially reduced using time intervals, whose area is expressed as a percentage of total area of the chromatogram. Then the variability due the characteristics of the samples in terms of treatments, biological factors (ripening, cheese making process, milk quality, etc) and technical factors (sampling, extraction steps, and measurement of peak and intervals area), is measured and analyzed through the contributing eigenvalues in the correlation matrix for the principal components of the resulting model. The peptide profiles revealed visible differences that were supported by the PCA results. Indeed, the PCA analysis leads to a model with three principal components (PCs) that explain 183 the 80.7% of the variability of the data. However, after comparing in pairs the score plots for any combination of the principal components, only the score plot for the PC1 Vs PC2 revealed a correlation. This is in agreement with the published results of (Benfeldt and Sørensen 2001). The first and second principal components explained the 72% of the variance, and the score plots in figures 71 and 72 show how the model can differentiate the samples according to their age in the PC1 and according their quality in PC2. Therefore, the figures display higher scores for those samples with longer ripening that correspond to the good samples, which indicate that proteolysis is faster for cheeses corresponding to the “good” samples. The loading plot in figure 73, shows the projection of the eluting intervals on the PC1 and PC2, and it allows to establish a correlations between the type of sample, its age and the amount of peptides eluting within certain retention times. Thus, it can be seen that the segments from 12-20 and 20-25 minutes have high scores for the PC1 and positive values close to 0 for PC2. This trend suggests that the amount of peptides eluting in this zones rise over time and higher amounts could be associate to good quality cheese. This interpretation could be explain by the fact that the segments from 12-20 and 20-25 are mainly composed by hydroplhilic peptides and free amino acids (FAA) resulting from the action of Chymosin on αs1-CN and k-caseins, and the cell envelope proteinase (CEP) on the peptide αs1-CN(f1-23), which accumulate during ripening (Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998). Thus, based on the works of Singh and Fox 1998, which describes the WSN fraction as the one that contains many compounds associated to the characteristic savory flavor in cheese. It could be possible to suggest that higher amounts of these hydrophilic peptides and free aminoacids might be related to “good” quality cheese. Which can associated to processing variables such as …………. that direct and indirectly affect proteolytic systems such as Chymosin and CEP. Contrary, the segments 25-30 and 30-35 got negative loadings for PC1 and PC2, which means that the relative amount of peptides eluting in this intervals decrease over time and therefore higher amounts might be related to “weak” cheese. This is believed to do with the breakdown of the αs1-CN, αs2-CN, αs1-CN (f24-199) and β-CN peptides, corresponding to 184 enzymatic activity proportionate by the rennet, indigenous milk enzymes and LAB enzymes (T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998); making sense since the breakdown of the main caseins is progressive and a higher concentration of them could be related to the original character of curd or to a mild and definitevily “non”-sharp Cheddar cheese. Nonetheless, in spite of the clear discrimination of samples according to quality, it is not easy to explain the reason why, which is consequence of the proteolytic systems involved in these segments that basically are all of them. This indicates the obvious dissimilarity between the production process of these two types of samples. Regarding the intervals 35-40 and 40-45, they are mainly constituted by hydrophobic peptides such as: 1) the fragments β-CN(f29–209), β-CN(f106–209) and β-CN(f108–209) (γ1, γ 2, and γ 3, respectively), whose concentrations increase during ripening (Farkye and Fox 1990) and are the result from the hydrolysis of β-Casein by Plasmin at Lys28-Lys29, Lys105-Gln106 and Lys107-Glu108 bonds; 2) the peptides αs1-CN(f93–?), αs1-CN(f24–30), αs1-CN(f26–32), αs1-CN(f26–34) resulting from the hydrolysis of the peptide αs1-CN(f24–199) by Chymosin, CEP and aminopeptidase (T.K. Singh et al. 1994; Tanoj K. Singh, Fox, and Healy 1995; Tanoj K. Singh, Fox, and Healy 1997; Manuela Fernández, Singh, and Fox 1998); 3) peptides αs2CN(f204–207), which is a C-terminal residue and product of lactococcal CEP (T.K. Singh et al. 1994). Therefore, the positive scores for PC1 and the negative ones for PC2 means that the amount of peptides eluting in these intervals increased during time and higher amounts are related to “good” cheese. As in the case of the last two intervals, the development of these peptides involved all proteolytic systems, thus it is difficult to point out a specific part of the manufacturing process that causes the difference detected. The intervals between 45-65 minutes are mainly constituted by hydrophobic compounds; however, they displayed a different trend to that of the intervals between 35 to 45 min. The scores in figure 73 showed negative values for the intervals 45-50, 50-55 and 55-60 in the PC1 while the interval 60-65 got a small and positive score, which is close to zero. This means that those the amount of peptides eluting in the first two intervals decreased during time whereas the constituents of last interval increased. On the other hand the interval 45-50 and 50-55 got scores fairly close to zero for the PC2, which suggests that there is not a big 185 difference between samples for this part of the chromatograms. Contrary the interval 55-60 got a positive value in PC2, which indicates that the smaller is the amount of material eluting in this interval the better the quality of the sample. The interval 60-65 got a negative score and this suggests that the bigger is the amount of the material eluting in this zone the lower is the quality of the samples. A possible explanation for the the way the samples were discriminated by the model might has to do with the bitter character of some of the hydrophobic peptides and free amino acids of this zone, which essentially are those that contain aromatic amino acid (Allan J. Cliffe, Marks, and Mulholland 1993). Therefore the last two observations respect the PC2 makes sense since bitterness is one of the defects to avoid in Cheddar cheese. Figure 70 Scree plot of Good Vs Weak cheese 186 Figure 71 Score plot of Good Vs Weak cheese (age) Figure 72 Score plot of Good Vs Weak cheese (quality) 187 Figure 74 Peptide profile Weak Cheese (A and B) Vs Good Cheese (C and D) A C B D 188 Electrophoresis The results of the urea-PAGE of the water insoluble nitrogen fraction of experimental Cheddar cheese in figures 75 and 76, clearly exhibit the progressive change of αS1-CN into the peptides αS1-CN (f24-199), αS1-CN (f121-199), αS1-CN (f99-199), and the β-CN into peptides β-CN (f29-202), β-CN (f108-209) and β-CN (f106-209) during ripening. In addition bands of γ-CNs became more noticeable after 4 months. Also it can be seen that apparently the development of peptides from αS1-CN is faster than those from β-CN, which can be related to primary proteolysis and the actual amount of β-CN that is hydrolyzed (only the 50%). However, the representative urea-PAGE gel do not show any appreciable difference between samples of different quality. Figure 75 Urea PAGE for ripening of Good cheese β -CN(f106-209) β -CN(f29-209) β -CN(f108-209) β -CN αs1 -CN αs1 -CN(f24-199) αs1-CN(f121-199) 2M 6M 9M 12M 15MG 15MW 189 Figure 76 77 Urea PAGE for Good Vs Weak cheese (12 months) Good Weak CONCLUSION The present study demonstrates that the use of FFA profile, VSC’s profile, measurement of the levels of the Total Kjeldahl nitrogen for the WSN, TCA-SN and PTA-SN fractions, and the PCA of the RP-HPLC peptide profile of the WSN fraction are effective tools and ripening indices to differentiate Cheddar cheese samples regarding their age and quality. The urea-PAGE was effective to differentiate samples by their age; nonetheless it was not sensitive enough to detect differences related to quality. On the other hand, , the results of levels of nitrogen for all the 3 fractions analyzed demonstrated that proteolysis is faster for good cheeses This was supported by the PCA model obtained which suggests many possible causes. Lipolysis was slower for weak cheese, which showed lower levels of individual FFA. The amounts of DMS, H2S and MeSH showed noticeable differences between samples and it can be seen that good cheese undergoes a faster catabolism of sulfur containing. Once again, the results for DMDS and DMTS suggest that they are artifacts from extraction and separation procedures rather than metabolites from the ripening of Cheddar cheese. 190 REFERENCES Alting, A. C., W. Engels, S. van Schalkwijk, and F. A. Exterkate. 1995. “Purification and Characterization of Cystathionine (beta)-Lyase from Lactococcus Lactis Subsp. Cremoris B78 and Its Possible Role in Flavor Development in Cheese.” Applied and Environmental Microbiology V61 (11) (November 1): pp. 4037–4042. Amarita, F., T. Requena, G. Taborda, L. Amigo, and C. Pelaez. 2001. “Lactobacillus Casei and Lactobacillus Plantarum Initiate Catabolism of Methionine by Transamination.” Journal of Applied Microbiology V90 (6): pp. 971–978. Aston, J. W., and L. K. Creamer. 1986. “Contribution of the components of the water-soluble fraction to the flavour of Cheddar cheese.” New Zealand Journal of Dairy Science and Technology V21 (3): pp. 229–248. Aston, J. W., and John R. Dulley. 1982. “Cheddar cheese flavor review.” Australian Journal of Dairy Technology V37 (2): pp. 59–64. Baer, A., and I. Ryba. 1992. “Serological Identification of Propionibacteria in Milk and Cheese Samples.” International Dairy Journal V2 (5): pp. 299–310. Balcão, Victor M., and F.Xavier Malcata. 1998. “Lipase Catalyzed Modification of Milkfat.” Biotechnology Advances V16 (2) (March): pp. 309–341. Bansal, N, Paolo Piraino, and P. L. H. McSweeney. 2009. “Determination of Proteolysis in Cheese.” In Handbook of Dairy Foods Analysis, pp. 405–417. CRC Press. Benfeldt, Connie, and John Sørensen. 2001. “Heat Treatment of Cheese Milk: Effect on Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4–7): pp. 567–574. Bentley, Ronald, and Thomas G. Chasteen. 2004. “Environmental VOSCs––formation and Degradation of Dimethyl Sulfide, Methanethiol and Related Materials.” Chemosphere V55 (3): pp. 291–317. Bills, D.D., and E.A. Day. 1964. “Determination of the Major Free Fatty Acids of Cheddar Cheese.” Journal of Dairy Science V47 (7): pp. 733–738. Burbank, Helen M., and Michael C. Qian. 2005. “Volatile Sulfur Compounds in Cheddar Cheese Determined by Headspace Solid-phase Microextraction and Gas Chromatograph-pulsed Flame Photometric Detection.” Journal of Chromatography A V1066 (1–2): pp. 149–157. Burbank, Helen, and Michael C. Qian. 2008. “Development of Volatile Sulfur Compounds in Heat-shocked and Pasteurized Milk Cheese.” International Dairy Journal V18 (8) (August): pp. 811–818. Chavarri, Felisa, Mailo Virto, Celia Martin, Ana I. Nájera, Arantza Santisteban, Luis J. R. Barrón, and Mertxe De Renobales. 1997. “Determination of Free Fatty Acids in Cheese: Comparison of Two Analytical Methods.” Journal of Dairy Research V64 (03): pp. 445–452. Christensen, Tove M. I. E., Kristian R. Kristiansen, and Jesper S. Madsen. 1989. “Proteolysis in Cheese Investigated by High Performance Liquid Chromatography.” Journal of Dairy Research V56 (05): pp. 823–828. Cliffe, A.J., D. Revell, and B.A. Law. 1989. “A Method for the Reverse Phase High Performance Liquid Chromatography of Peptides from Cheddar Cheese.” Food Chemistry 34 (2): 147–160. 191 Cliffe, Allan J., Jonathan D. Marks, and Francis Mulholland. 1993. “Isolation and Characterization of Non-volatile Flavours from Cheese: Peptide Profile of Flavour Fractions from Cheddar Cheese, Determined by Reverse-phase High-performance Liquid Chromatography.” International Dairy Journal V3 (4–6): pp 379–387. Collins, Yvonne F., Paul L.H. McSweeney, and Martin G. Wilkinson. 2003. “Lipolysis and Free Fatty Acid Catabolism in Cheese: a Review of Current Knowledge.” International Dairy Journal V13 (11): pp. 841–866. Collins, Y.F., P.L.H. McSweeney, and M.G. Wilkinson. 2004. “Lipolysis and Catabolism of Fatty Acids in Cheese.” In Cheese Chemistry, Physics and Microbiology, Volume 1:373–389. Academic Press. Curioni, P.M.G., and J.O. Bosset. 2002. “Key Odorants in Various Cheese Types as Determined by Gas Chromatography-olfactometry.” International Dairy Journal v12 (12): pp. 959–984. Datta, N., A. J. Elliott, M. L. Perkins, and H. C. Deeth. 2002. “Ultra-high-temperature (UHT) Treatment of Milk: Comparison of Direct and Indirect Modes of Heating” V57 (3): pp. 211–227. Deeth, H. C., and V. Touch. 2000. “Methods for detecting lipase activity in milk and milk products.” Australian Journal of Dairy Technology V55 (3): pp. 153–168. Dias, Benjamin, and Bart Weimer. 1998. “Conversion of Methionine to Thiols by Lactococci, Lactobacilli, and Brevibacteria.” Applied and Environmental Microbiology V64 (9): pp. 3320–3326. Farkye, Nana Y., and Patrick F. Fox. 1990. “Observations on Plasmin Activity in Cheese.” Journal of Dairy Research V57 (03): pp. 413–418. Fennema, Owen R., and Damodaran. 1996. “Amino Acids, Peptides, and Proteins.” In Food Chemistry, pp. 322–425. CRC Press. Fernández, Manuela, Tanoj K. Singh, and Patrick F. Fox. 1998. “Isolation and Identification of Peptides from the Diafiltration Permeate of the Water-Soluble Fraction of Cheddar Cheese.” Journal of Agricultural and Food Chemistry V46 (11): pp. 4512–4517. Fernández-García, Estrella. 1996. “Use of Headspace Sampling in the Quantitative Analysis of Artisanal Spanish Cheese Aroma.” Journal of Agricultural and Food Chemistry V44 (7): pp. 1833–1839. Fox, P. F., Law J, P. L. H. McSweeney, and Wallace J.M. 1999. “Biochemistry of Cheese Ripening.” In Cheese: Chemistry, Physics and Microbiology, pp. 382–450. Springer. Fox, P.F. 1989. “Proteolysis During Cheese Manufacture and Ripening.” Journal of Dairy Science V72 (6): pp. 1379–1400. Fox, P.F., and L. Stepaniak. 1993. “Enzymes in Cheese Technology.” International Dairy Journal V3 (4–6): pp. 509–530.. Fox, P. F., and P. L. H. McSweeney. 1996. “Proteolysis in Cheese During Ripening.” Food Reviews International V12 (4): pp. 457–509. Fox, P. F., Law J, P. L. H. McSweeney, and Wallace J.M. 1999. “Biochemistry of Cheese Ripening.” In Cheese: Chemistry, Physics and Microbiology, pp. 382–450. Springer. Gao, Song, Ed S. Mooberry, and James L. Steele. 1998. “Use of 13C Nuclear Magnetic Resonance and Gas Chromatography To Examine Methionine Catabolism by Lactococci.” Applied and Environmental Microbiology V64 (12): pp. 4670–4675. Green, Margaret L., and Donald J. Manning. 1982. “Development of Texture and Flavour in Cheese and Other Fermented Products.” Journal of Dairy Research V49 (04): pp. 737–748. 192 Gripon, Jean-Claude, V Monnet, G Lambert, and M. J. Desmazeaud. 1991. “Microbial enzymes in cheese ripening.” In Food enzymology, vol 1. K. Arfi, H.Spinnler, R. Tache, and P. bonnarme. 2002. “Production of Volatile Compounds by Cheese-ripening Yeasts: Requirement for a Methanethiol Donor for S-methyl Thioacetate Synthesis by Kluyveromyces Lactis.” Applied Microbiology and Biotechnology V58 (4): pp. 503–510. Lau, Kum Y., David M. Barbano, and Robert R. Rasmussen. 1991. “Influence of Pasteurization of Milk on Protein Breakdown in Cheddar Cheese During Aging.” Journal of Dairy Science V74 (3): pp. 727–740. Lee, Won-Jae, Dattatreya S. Banavara, Joanne E. Hughes, Jason K. Christiansen, James L. Steele, Jeffery R. Broadbent, and Scott A. Rankin. 2007. “Role of Cystathionine βLyase in Catabolism of Amino Acids to Sulfur Volatiles by Genetic Variants of Lactobacillus Helveticus CNRZ 32.” Applied and Environmental Microbiology V73 (9): pp. 3034–3039. Manning, D. J., Helen R. Chapman, and Zena D. Hosking. 1976. “The Production of Sulphur Compounds in Cheddar Cheese and Their Significance in Flavour Development.” Journal of Dairy Research V43 (2): pp. 313–320. Manning, Donald J., and Carolyn Moore. 1979. “Headspace Analysis of Hard Cheeses.” Journal of Dairy Research V46 (3): pp. 539–545. McSweeney, P.L.H., P.F. Fox, J.A. Lucey, K.N. Jordan, and T.M. Cogan. 1993. “Contribution of the Indigenous Microflora to the Maturation of Cheddar Cheese.” International Dairy Journal V3 (7): pp. 613–634. McSweeney, Paul L. H., Sylvie Pochet, Patrick F. Fox, and Aine Healy. 1994. “Partial Identification of Peptides from the Water-Insoluble Fraction of Cheddar Cheese.” Journal of Dairy Research V61 (4) (04): pp. 587–590. McSweeney, Paul L.H., and Maria José Sousa. 2000. “Biochemical Pathways for the Production of Flavour Compounds in Cheeses During Ripening: A Review.” Le Lait V80 (3): pp. 293–324. Metwalli, Ali A.M, Harmen H.J de Jongh, and Martinus A.J.S van Boekel. 1998. “Heat Inactivation of Bovine Plasmin.” International Dairy Journal V8 (1): pp. 47–56. Milo, C., and G. A. Reineccius. 1997. “Identification and Quantification of Potent Odorants in Regular-Fat and Low-Fat Mild Cheddar Cheese.” Journal of Agricultural and Food Chemistry V45 (9) (September 1): pp. 3590–3594. Molimard, P., and H.E. Spinnler. 1996. “Review: Compounds Involved in the Flavor of Surface Mold-Ripened Cheeses: Origins and Properties.” Journal of Dairy Science V79 (2) (February): pp. 169–184. O’Keeffe, Arthur M., Patrick F. Fox, and Charles Daly. 1978. “Proteolysis in Cheddar Cheese: Role of Coagulant and Starter Bacteria.” Journal of Dairy Research V45 (3): pp. 465– 477.. O’Keeffe, R. B., P. F. Fox, and C. Daly. 1976. “Contribution of Rennet and Starter Proteases to Proteolysis in Cheddar Cheese.” Journal of Dairy Research 43 (1): 97–107. Piraino, Paolo, Eugenio Parente, and Paul L. H. McSweeney. 2004. “Processing of Chromatographic Data for Chemometric Analysis of Peptide Profiles from Cheese Extracts: A Novel Approach.” Journal of Agricultural and Food Chemistry 52 (23): 6904–6911. Preininger, Martin, and Werner Grosch. 1994. “Evaluation of Key Odorants of the Neutral Volatiles of Emmentaler Cheese by the Calculation of Odour Activity Values.” LWT Food Science and Technology V27 (3): pp. 237–244. 193 Qian, M., and G. Reineccius. 2002. “Identification of Aroma Compounds in ParmigianoReggiano Cheese by Gas Chromatography/Olfactometry.” Journal of Dairy Science V85 (6) pp. 1362–1369. Seefeldt, K.E., and B.C. Weimer. 2000. “Diversity of Sulfur Compound Production in Lactic Acid Bacteria.” Journal of Dairy Science V83 (12): pp. 2740–2746. Singh, T. K, M. A Drake, and K. R Cadwallader. 2003. “Flavor of Cheddar Cheese: A Chemical and Sensory Perspective.” Comprehensive Reviews in Food Science and Food Safety V2 (4): pp. 166–189. Singh, T.K., P.F. Fox, P. Højrup, and A. Healy. 1994. “A Scheme for the Fractionation of Cheese Nitrogen and Identification of Principal Peptides.” International Dairy Journal V4 (2): pp. 111–122. Singh, Tanoj K., Patrick F. Fox, and Áine Healy. 1995. “Water-Soluble Peptides in Cheddar Cheese: Isolation and Identification of Peptides in the Diafiltration Retentate of the Water-Soluble Fraction.” Journal of Dairy Research V62 (4): pp. 629–640. Singh, Tanoj K., Patrick F. Fox, and Áine Healy. 1997. “Isolation and Identification of Further Peptides in the Diafiltration Retentate of the Water-soluble Fraction of Cheddar Cheese.” Journal of Dairy Research V64 (3): pp. 433–443. Smacchi, Emanuele, and Marco Gobbetti. 1998. “Purification and Characterization of Cystathionine Γ-lyase from Lactobacillus Fermentum DT41.” FEMS Microbiology Letters V166 (2): pp. 197–202. Smith, A. M., and S. Nakai. 1990. “Classification of cheese varieties by multivariate analysis of HPLC profiles.” Canadian Institute of Food Science and Technology Journal V23 (1): pp. 53–58. Sousa, M.J, Y Ardö, and P.L.H McSweeney. 2001. “Advances in the Study of Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4-7): pp. 327–345. Subramanian, A., W.J. Harper, and L.E. Rodriguez-Saona. 2009. “Cheddar Cheese Classification Based on Flavor Quality Using a Novel Extraction Method and Fourier Transform Infrared Spectroscopy.” Journal of Dairy Science V92 (1): pp. 87–94. Tanaka, Hidehiko, Nobuyoshi Esaki, and Kenji Soda. 1985. “A Versatile Bacterial Enzyme: L-methionine Γ-lyase.” Enzyme and Microbial Technology V7 (11) (November): pp. 530–537. Urbach, G. 1995. “Contribution of Lactic Acid Bacteria to Flavour Compound Formation in Dairy Products.” International Dairy Journal V5 (8): pp. 877–903. USDA. 2011. Dairy Products Annual Sumary. Visser, Servaas. 1993. “Proteolytic Enzymes and Their Relation to Cheese Ripening and Flavor: An Overview.” Journal of Dairy Science V76 (1): pp. 329–350. Voigt, Daniela D., François Chevalier, John A. Donaghy, Margaret F. Patterson, Michael C. Qian, and Alan L. Kelly. 2012. “Effect of High-pressure Treatment of Milk for Cheese Manufacture on Proteolysis, Lipolysis, Texture and Functionality of Cheddar Cheese During Ripening.” Innovative Food Science & Emerging Technologies V13 (0): pp. 23–30. Weimer, Bart, Kimberly Seefeldt, and Benjamin Dias. 1999. “Sulfur Metabolism in Bacteria Associated with Cheese.” Antonie Van Leeuwenhoek 76 (1): 247–261. de Wit, Rudy, and Hans Nieuwenhuijse. 2008. “Kinetic Modelling of the Formation of Sulphur-containing Flavour Components During Heat-treatment of Milk.” International Dairy Journal V18 (5): pp. 539–547. Yvon, Mireille, and Liesbeth Rijnen. 2001. “Cheese Flavour Formation by Amino Acid Catabolism.” International Dairy Journal V11 (4–7): pp. 185–201. 194 195 CONCLUSION The present study demonstrates that the use of FFA profile, VSC’s profile, measurement of the level of the Total Kjeldahl nitrogen for the WSN, TCA-SN and PTA-SN fractions, and the PCA analysis of the RP-HPLC peptide profile of the WSN fraction are effective tools and ripening indices to differentiate Cheddar cheese samples regarding variables such as quality, age, heat treatment of milk, origin, and addition of adjunct culture. In the case of Urea-PAGE, it was demonstrated that its use as index of primary proteolysis can be effective to differentiate samples by their age; nonetheless it is not sensitive enough to detect differences related to the changes caused by heat treatment of cheese milk, origin of samples and addition of adjunct culture. It was demonstrated that proteolysis is faster for good cheese and cheeses made with adjunct culture, made in the TCCA plant and made from heat-shocked milk. In general the levels of nitrogen for the 3 fractions analyzed were the highest for these treatments. The results were supported by the PCA models obtained, which suggest differences caused by: 1) the role of the adjunct culture as supplement to the starter culture during ripening, 2) distinct manufacturing practices between prodcutction plants, and 3) physical changes to milk structure and population of NSLAB as consequence of heat treatments of cheese milk. Llipolysis is slower for weak samples, cheese produce without adjunct culture, cheese made with pasteurized milk and for samples made in the CRP plant, which showed lower levels of individual FFA, speacially for short chain fatty acids. Finally the amounts of DMS, H2S and MeSH, revealed differences between the treatments, and it is possible to appreciate the tendency to accelerate the catabolism of sulfur containing amino acids, such as methionine and cysteine, for the samples with adjunct culture, made from heat-shocked milk and from the TCCA plant. An important outcome from this research work was to point out the possibility that DMDS and DMTS are artifacts from extraction and separation procedures rather than metabolites from the ripening of Cheddar cheese. Other tests such as analysis of amino acids or measurement of the levels of other potent volatile compounds through GC-MS should be performed in order to complement the proposed chemical analysis and to obtain more information to keep explaining the causes of 196 the difference in rates and patterns of lipolysis and proteolysis. In addition, it would be usuful to totally assure that the same manufacturing procedures and raw materials are used during the manufacture of samples, which might give a better idea about the influence of the investigated parameters in the final results by eliminating the noise of other manufacturing variables. Other areas that deserve more research and are not well understood yet, are related to the specific role of important strains of NSLAB in lipolysis and proteolysis, and the generation of DMS and H2S. 197 BIBLIOGRAFY Alichanidis, Efstathios, Julia H. M. Wrathall, and Anthony T. Andrews. 1986. “Heat Stability of Plasmin (milk Proteinase) and Plasminogen.” Journal of Dairy Research V53 (2): pp. 259–269. Alting, A. C., W. Engels, S. van Schalkwijk, and F. A. Exterkate. 1995. “Purification and Characterization of Cystathionine (beta)-Lyase from Lactococcus Lactis Subsp. Cremoris B78 and Its Possible Role in Flavor Development in Cheese.” Applied and Environmental Microbiology V61 (11): pp. 4037–4042. Amarita, F., T. Requena, G. Taborda, L. Amigo, and C. Pelaez. 2001. “Lactobacillus Casei and Lactobacillus Plantarum Initiate Catabolism of Methionine by Transamination.” Journal of Applied Microbiology V90 (6): pp. 971–978. Andersen, Lene T., Ylva Ardö, and Wender L.P. Bredie. 2010. “Study of Taste-active Compounds in the Water-soluble Extract of Mature Cheddar Cheese.” International Dairy Journal V20 (8): pp. 528–536. Anderson, D.L., V.V. Mistry, R.L. Brandsma, and K.A. Baldwin. 1993. “Reduced Fat Cheddar Cheese from Condensed Milk. 1. Manufacture, Composition, and Ripening.” Journal of Dairy Science V76 (10): pp. 2832–2844. Ardö, Y., and Frederiksberg. 1999. “Evaluating Proteolysis by Analysing the N Content of Cheese Fractions.” Bulletin - International Dairy Federation V337: pp. 4–9. Arora, Gulshan, Francois Cormier, and Byong Lee. 1995. “Analysis of Odor-Active Volatiles in Cheddar Cheese Headspace by Multidimensional GC/MS/Sniffing.” J. Agric. Food Chem. V43 (3): pp. 748–752. Aston, J. W., and L. K. Creamer. 1986. “Contribution of the components of the water-soluble fraction to the flavour of Cheddar cheese.” New Zealand Journal of Dairy Science and Technology V21 (3): pp. 229–248. Aston, J. W., and John R. Dulley. 1982. “Cheddar cheese flavor review.” Australian Journal of Dairy Technology V37 (2): pp. 59–64. Avsar, Y.K., Y. Karagul-Yuceer, M.A. Drake, T.K. Singh, Y. Yoon, and K.R. Cadwallader. 2004. “Characterization of Nutty Flavor in Cheddar Cheese.” Journal of Dairy Science V87 (7): pp. 1999–2010. Ayad, Eman H.E, Annette Verheul, Catrienus de Jong, Jan T.M Wouters, and Gerrit Smit. 1999. “Flavour Forming Abilities and Amino Acid Requirements of Lactococcus Lactis Strains Isolated from Artisanal and Non-dairy Origin.” International Dairy Journal V9 (10): pp. 725–735. Baer, A., and I. Ryba. 1992. “Serological Identification of Propionibacteria in Milk and Cheese Samples.” International Dairy Journal V2 (5): pp. 299–310. Balcão, Victor M., and F.Xavier Malcata. 1998. “Lipase Catalyzed Modification of Milkfat.” Biotechnology Advances V16 (2) (March): pp. 309–341. Bansal, N, Paolo Piraino, and P. L. H. McSweeney. 2009. “Determination of Proteolysis in Cheese.” In Handbook of Dairy Foods Analysis, pp. 405–417. CRC Press. Bassett, Harold J., and W.L. Slatter. 1953. “The Heat Resistance of Some Lactobacilli Found in Swiss Cheese.” Journal of Dairy Science V36 (4): pp. 409–413. Bastian, Eric D., and Rodney J. Brown. 1996. “Plasmin in Milk and Dairy Products: An Update.” International Dairy Journal V6 (5): pp. 435–457. 198 Bastian, E.D., C.G. Lo, and K.M.M. David. 1997. “Plasminogen Activation in Cheese Milk: Influence on Swiss Cheese Ripening.” Journal of Dairy Science 80 (2): pp. 245–251. Benfeldt, Connie, John Sørensen, Katrine H. Ellegård, and Torben E. Petersen. 1997. “Heat Treatment of Cheese Milk: Effect on Plasmin Activity and Proteolysis During Cheese Ripening.” International Dairy Journal 7 (11): 723–731. Benfeldt, Connie, and John Sørensen. 2001. “Heat Treatment of Cheese Milk: Effect on Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4–7): pp. 567–574. Benkler, K. F., and G. A. Reineccius. 1980. “Flavor isolation from fatty foods via solvent extraction and membrane dialysis.” Journal of Food Science V45 (4): pp. 1084–1085. Bentley, Ronald, and Thomas G. Chasteen. 2004. “Environmental VOSCs––formation and Degradation of Dimethyl Sulfide, Methanethiol and Related Materials.” Chemosphere V5 (3): pp. 291–317. Beresford, Tom P, Nora A Fitzsimons, Noelle L Brennan, and Tim M Cogan. 2001. “Recent Advances in Cheese Microbiology.” International Dairy Journal V11 (4–7): pp. 259– 274. Beuvier, E., and S. Buchin. 2004. “Raw Milk Cheeses.” In Cheese Chemistry, Physics and Microbiology, Volume 1:319–345. Academic Press. Bills, D.D., and E.A. Day. 1964. “Determination of the Major Free Fatty Acids of Cheddar Cheese.” Journal of Dairy Science V47 (7): pp. 733–738. Burbank, Helen M., and Michael C. Qian. 2005. “Volatile Sulfur Compounds in Cheddar Cheese Determined by Headspace Solid-phase Microextraction and Gas Chromatograph-pulsed Flame Photometric Detection.” Journal of Chromatography A V1066 (1–2): pp. 149–157. Burbank, Helen, and Michael C. Qian. 2008. “Development of Volatile Sulfur Compounds in Heat-shocked and Pasteurized Milk Cheese.” International Dairy Journal V18 (8): pp. 811–818. Burton, H. 1986. “Microbiological aspects.” V200: pp. 9–14. Careri, M, and A Mangia. 2003. “Analysis of Food Proteins and Peptides by Chromatography and Mass Spectrometry.” Journal of Chromatography A V1000 (1–2): pp. 609–635. Carunchia Whetstine, Mary E., Keith R. Cadwallader, and MaryAnne Drake. 2005. “Characterization of Aroma Compounds Responsible for the Rosy/Floral Flavor in Cheddar Cheese.” J. Agric. Food Chem. V53 (8): pp. 3126–3132. Carunchia Whetstine, M E, P J Luck, M A Drake, E A Foegeding, P D Gerard, and D M Barbano. 2007. “Characterization of Flavor and Texture Development Within Large (291 Kg) Blocks of Cheddar Cheese.” Journal of Dairy Science V90 (7): pp. 3091– 3109. Chaintreau, Alain. 2001. “Simultaneous Distillation–extraction: From Birth to Maturity— review.” Flavour and Fragrance Journal V16 (2): pp. 136–148. Chatfield, Chris, and A. Collins. 1981. “Finding New Underlying Variables.” In Introduction to Multivariate Analysis, pp. 57–78. CRC Press. Chavarri, Felisa, Mailo Virto, Celia Martin, Ana I. Nájera, Arantza Santisteban, Luis J. R. Barrón, and Mertxe De Renobales. 1997. “Determination of Free Fatty Acids in Cheese: Comparison of Two Analytical Methods.” Journal of Dairy Research V64 (3): pp. 445–452. Chich, Jean-François, Karine Marchesseau, and Jean-Claude Gripon. 1997. “Intracellular Esterase from Lactococcus Lactis Subsp. Lactis NCDO 763: Purification and 199 Characterization.” International Dairy Journal V7 (2-3) (2–3) (February): pp. 169– 174. Chopin, Alain. 1993. “Organization and Regulation of Genes for Amino Acid Biosynthesis in Lactic Acid Bacteria.” FEMS Microbiology Reviews V12 (1–3): pp. 21–37. Christensen J.E., Johnson M.E., and Steele J.L. 1995. “Production of Cheddar Cheese Using a Lactococcus Lactis Ssp. Cremoris SK11 Derivative with Enhanced Aminopeptidase Activity.” International Dairy Journal V5 (4): pp. 367–379. Christensen, Jeffrey E., Edward G. Dudley, Jeffrey A. Pederson, and James L. Steele. 1999. “Peptidases and Amino Acid Catabolism in Lactic Acid Bacteria.” Antonie Van Leeuwenhoek V76 (1): pp. 217–246. Christensen, K.R., and G.A. Reineccius. 1992. “Gas Chromatographic Analysis of Volatile Sulfur Compounds from Heated Milk Using Static Headspace Sampling.” Journal of Dairy Science V75 (8): pp. 2098–2104. Christensen, Søren, Tove Wiegers, John Hermansen, and Lars Sottrup-Jensen. 1995. “Plasmaderived Protease Inhibitors in Bovine Milk.” International Dairy Journal V5 (5): pp. 439–449. Christensen, Tove M. I. E., Kristian R. Kristiansen, and Jesper S. Madsen. 1989. “Proteolysis in Cheese Investigated by High Performance Liquid Chromatography.” Journal of Dairy Research V56 (5): pp. 823–828. Christensen, T. M. I. E., A.-M. Bech, and H. Werner. 1991. “Methods for crude fractionation (extraction and precipitation) of nitrogen components in cheese.” Bulletin International Dairy Federation (261): pp. 4–9. Cliffe, A.J., D. Revell, and B.A. Law. 1989. “A Method for the Reverse Phase High Performance Liquid Chromatography of Peptides from Cheddar Cheese.” Food Chemistry 34 (2): 147–160. Cliffe, Allan J., Jonathan D. Marks, and Francis Mulholland. 1993. “Isolation and Characterization of Non-volatile Flavours from Cheese: Peptide Profile of Flavour Fractions from Cheddar Cheese, Determined by Reverse-phase High-performance Liquid Chromatography.” International Dairy Journal V3 (4–6): pp 379–387. Collin, J. C., D. le Bars, I. Ryba, and A. Bear. 1990. “Comparative measurements of the plasminogen-plasmin system of milk.” In International Dairy Federation. Collins, Yvonne F., Paul L.H. McSweeney, and Martin G. Wilkinson. 2003. “Lipolysis and Free Fatty Acid Catabolism in Cheese: a Review of Current Knowledge.” International Dairy Journal V13 (11): pp. 841–866. Collins, Y.F., P.L.H. McSweeney, and M.G. Wilkinson. 2004. “Lipolysis and Catabolism of Fatty Acids in Cheese.” In Cheese Chemistry, Physics and Microbiology, Volume 1:373–389. Academic Press. Considine, T., A. Healy, A.L. Kelly, and P.L.H. McSweeney. 1999. “Proteolytic Specificity of Elastase on Bovine Β-casein.” Food Chemistry V66 (4): pp. 463–470.. Considine, T., Á. Healy, A.L. Kelly, and P.L.H. McSweeney. 2000. “Proteolytic Specificity of Elastase on Bovine Αs1-casein.” Food Chemistry V69 (1): pp. 19–26. Considine, T., A. Healy, A.L. Kelly. 2004. “Hydrolysis of Bovine Caseins by Cathepsin B, a Cysteine Proteinase Indigenous to Milk.” International Dairy Journal V14 (2): pp. 117–124. Crow, V.L., T. Coolbear, P.K. Gopal, F.G. Martley, L.L. McKay, and H. Riepe. 1995. “The Role of Autolysis of Lactic Acid Bacteria in the Ripening of Cheese.” International Dairy Journal V5 (8): pp. 855–875. 200 Crow, Vaughan, Brian Curry, and Michelle Hayes. 2001. “The Ecology of Non-starter Lactic Acid Bacteria (NSLAB) and Their Use as Adjuncts in New Zealand Cheddar.” International Dairy Journal V11 (4-7): pp 275–283. Cserháti, Tibor. 2002. “Mass Spectrometric Detection in Chromatography. Trends and Perspectives.” Biomedical Chromatography V16 (5): pp. 303–310. Curioni, P.M.G., and J.O. Bosset. 2002. “Key Odorants in Various Cheese Types as Determined by Gas Chromatography-olfactometry.” International Dairy Journal v12: pp. 959–984. Datta, N., A. J. Elliott, M. L. Perkins, and H. C. Deeth. 2002. “Ultra-high-temperature (UHT) Treatment of Milk: Comparison of Direct and Indirect Modes of Heating” V57 (3): pp. 211–227. Deeth, H. C, and V. Touch. 2000. “Methods for detecting lipase activity in milk and milk products.” Australian Journal of Dairy Technology V5 (3): pp. 153–168. Dias, Benjamin, and Bart Weimer. 1998. “Conversion of Methionine to Thiols by Lactococci, Lactobacilli, and Brevibacteria.” Applied and Environmental Microbiology V64 (9): pp. 3320–3326. Dimos, Tony. 1992. “A comparative study by GC/MS of the flavour volatiles produced during the maturation of full-fat and low-fat Cheddar cheese.” Dirinck, P, and A De Winne. 1999. “Flavour Characterisation and Classification of Cheeses by Gas Chromatographic–mass Spectrometric Profiling.” Journal of Chromatography A V847 (1–2): pp. 203–208. Dobric, Nada, Gäetan K.Y. Limsowtin, Alan J. Hillier, Nicholas P.B. Dudman, and Barrie E. Davidson. 2000. “Identification and Characterization of a Cystathionine Β/γ-lyase from Lactococcus Lactis Ssp. Cremoris MG13631.” FEMS Microbiology Letters V182 (2): pp. 249–254. Drake, M.A., T.D. Boylston, K.D. Spence, and B.G. Swanson. 1996. “Chemical and Sensory Effects of a Lactobacillus Adjunct in Cheddar Cheese.” Food Research International V29 (3–4): pp. 381–387. El Soda, Morsi A. 1993. “The Role of Lactic Acid Bacteria in Accelerated Cheese Ripening.” FEMS Microbiology Reviews V12 (1–3): pp. 239–251. El Soda, M., S.A. Madkor, and P.S. Tong. 2000. “Adjunct Cultures: Recent Developments and Potential Significance to the Cheese Industry.” Journal of Dairy Science V83 (4): pp. 609–619. El-Soda, M., S. A. Madkor, and P. S. Tong. 2000. “Evaluation of commercial adjuncts for use in cheese ripening: 4. Comparison between attenuated and not attenuated lactobacilli.” Milchwissenschaft V55 (5): pp. 260–263. Engel, Wolfgang, Wolfgang Bahr, and P. Schieberle. 1999. “Solvent Assisted Flavour Evaporation – a New and Versatile Technique for the Careful and Direct Isolation of Aroma Compounds from Complex Food Matrices.” European Food Research and Technology V209 (3): pp. 237–241. Engels, W.J.M., R. Dekker, C. de Jong, R. Neeter, and S. Visser. 1997. “A Comparative Study of Volatile Compounds in the Water-soluble Fraction of Various Types of Ripened Cheese.” International Dairy Journal V7 (4): pp. 255–263. Exterkate, Fred, Arno C. Alting, and Charles J. Slangen. 1995. “Conversion of αs1-Casein(24-199)-Fragment and β-Casein Under Cheese Conditions by Chymosin and Starter Peptidases.” Systematic and Applied Microbiology V18 (1): pp. 7–12. 201 Fang, Yu, and Michael C. Qian. 2005. “Sensitive Quantification of Sulfur Compounds in Wine by Headspace Solid-phase Microextraction Technique.” Journal of Chromatography A V1080 (2): pp. 177–185. Farkye, Nana Y., and Patrick F. Fox. 1990. “Observations on Plasmin Activity in Cheese.” Journal of Dairy Research V57 (3): pp. 413–418. FDA. 2011. “Code of Federal Regulations Title 21.” http://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr=133.111. Fennema, Owen R., and Damodaran. 1996. “Amino Acids, Peptides, and Proteins.” In Food Chemistry, pp. 322–425. CRC Press. Fernández, Manuela, Tanoj K. Singh, and Patrick F. Fox. 1998. “Isolation and Identification of Peptides from the Diafiltration Permeate of the Water-Soluble Fraction of Cheddar Cheese.” Journal of Agricultural and Food Chemistry V46 (11): pp. 4512–4517. Fernández, María, Wim van Doesburg, Ger A. M. Rutten, Joey D. Marugg, Arno C. Alting, Richard van Kranenburg, and Oscar P. Kuipers. 2000. “Molecular and Functional Analyses of themetC Gene of Lactococcus Lactis, Encoding Cystathionine β-Lyase.” Applied and Environmental Microbiology V66 (1): pp. 42–48.. Fernández-García, Estrella. 1996. “Use of Headspace Sampling in the Quantitative Analysis of Artisanal Spanish Cheese Aroma.” Journal of Agricultural and Food Chemistry V44 (7): pp. 1833–1839. Ferreira, A.D., and B.C. Viljoen. 2003. “Yeasts as Adjunct Starters in Matured Cheddar Cheese.” International Journal of Food Microbiology V86 (1-2): pp. 131–140. Forde, Amanda, and Gerald F Fitzgerald. 2000. “Biotechnological Approaches to the Understanding and Improvement of Mature Cheese Flavour.” Current Opinion in Biotechnology V11 (5): pp. 484–489. Fox, P.F. 1989. “Proteolysis During Cheese Manufacture and Ripening.” Journal of Dairy Science V72 (6): pp. 1379–1400. Fox, P. F., J. A. Lucey, and T. M. Cogan. 1990. “Glycolysis and Related Reactions During Cheese Manufacture and Ripening.” Critical Reviews in Food Science and Nutrition V29 (4): pp. 237–253. Fox, P.F., and L. Stepaniak. 1993. “Enzymes in Cheese Technology.” International Dairy Journal V3 (4–6): pp. 509–530. Fox, P. F., and P. L. H. McSweeney. 1996. “Proteolysis in Cheese During Ripening.” Food Reviews International V12 (4): pp. 457–509. Fox, P. F., J. M. Wallace, S. Morgan, C. M. Lynch, E. J. Niland, and J. Tobin. 1996. “Acceleration of Cheese Ripening.” Antonie Van Leeuwenhoek 70 (2): 271–297. Fox, PF, PLH McSweeney, and CM Lynch. 1998. “Significance of Non-starter Lactic Acid Bacteria in Cheddar Cheese.” Australian Journal of Dairy Technology V53 (2): pp. 7– 8. Fox, P.F., and P.L.H. McSweeney 1998. “Milk Lipids.” In Dairy Chemistry and Biochemistry, pp. 67–141. Springer. Fox, P. F., Law J, P. L. H. McSweeney, and Wallace J.M. 1999. “Biochemistry of Cheese Ripening.” In Cheese: Chemistry, Physics and Microbiology, pp. 382–450. Springer. Fox, P. F., T.P. Guinee, T. M. Cogan, and P. L. H. McSweeney. 2000. “Enzymatic Coagulation of Milk.” In Fundamentals of Cheese Science, pp. 98135. Springer. Fox, P.F., and P.L.H. McSweeney. 2004. “Cheese: An Overview.” In Cheese Chemistry, Physics and Microbiology, Volume 1:1–18. Academic Press. 202 Gao, Song, Ed S. Mooberry, and James L. Steele. 1998. “Use of 13C Nuclear Magnetic Resonance and Gas Chromatography To Examine Methionine Catabolism by Lactococci.” Applied and Environmental Microbiology V64 (12): pp. 4670–4675. Gomez, Maria J., Sonia Garde, Pilar Gaya, Margarita Medina, and Manuel Nuñez. 1997. “Relationship Between Level of Hydrophobic Peptides and Bitterness in Cheese Made from Pasteurized and Raw Milk.” Journal of Dairy Research V64 (2): pp. 289–297. Grandison, Alistair S., and Graeme D. Ford. 1986. “Effects of Variations in Somatic Cell Count on the Rennet Coagulation Properties of Milk and on the Yield, Composition and Quality of Cheddar Cheese.” Journal of Dairy Research V53 (4): pp. 645–655. Grappin, Rémy, and Eric Beuvier. 1997. “Possible Implications of Milk Pasteurization on the Manufacture and Sensory Quality of Ripened Cheese.” International Dairy Journal V7 (12): pp. 751–761. Green, Margaret L., and Donald J. Manning. 1982. “Development of Texture and Flavour in Cheese and Other Fermented Products.” Journal of Dairy Research V49 (4): pp. 737– 748. Grieve, Paul A., and Barry J. Kitchen. 1985. “Proteolysis in Milk: The Significance of Proteinases Originating from Milk Leucocytes and a Comparison of the Action of Leucocyte, Bacterial and Natural Milk Proteinases on Casein.” Journal of Dairy Research 52 (1): 101–112. Griffiths, M. W. 1986. “Use of milk enzymes as indices of heat treatment.” Journal of Food Protection V49 (9): pp. 696–705. Gripon, Jean-Claude, V Monnet, G Lambert, and M. J. Desmazeaud. 1991. “Microbial enzymes in cheese ripening.” In Food enzymology, vol 1. Guinee, T P. 2004. “Salting and the Role of Salt in Cheese.” International Journal of Dairy Technology V57 (2-3): pp. 99–109. Habibi‐Najafi, Mohammad B., Byong H. Lee, and Barry Law. 1996. “Bitterness in Cheese: A Review.” Critical Reviews in Food Science and Nutrition V36 (5): pp. 397–411. Heegaard, Christian W., Lone K. Rasmussen, and Peter A. Andreasen. 1994. “The Plasminogen Activation System in Bovine Milk: Differential Localization of Tissuetype Plasminogen Activator and Urokinase in Milk Fractions Is Caused by Binding to Casein and Urokinase Receptor.” Biochimica Et Biophysica Acta (BBA) - Molecular Cell Research V1222 (1): pp. 45–55. Hickey, D.K., K.N. Kilcawley, T.P. Beresford, and M.G. Wilkinson. 2007. “Lipolysis in Cheddar Cheese Made from Raw, Thermized, and Pasteurized Milks.” Journal of Dairy Science V90 (1): pp. 47–56. Holland, Ross, and Tim Coolbear. 1996. “Purification of Tributyrin Esterase from Lactococcus Lactis Subsp. Cremoris E8.” Journal of Dairy Research V63 (1): pp. 131–140. Home, D.S., and J.M. Banks. 2004. “Rennet-induced Coagulation of Milk.” In Cheese Chemistry, Physics and Microbiology, Volume 1:pp. 47–70. Academic Press. Huffman, L. M.;, and Kristoffersen. 1984. “Role of lactose in Cheddar cheese manufacturing and ripening.” New Zealand Journal of Dairy Science and Technology V19 (2): pp. 151–162. Hurley, M.J, B.M O’Driscoll, A.L Kelly, and P.L.H McSweeney. 1999. “Novel Assay for the Determination of Residual Coagulant Activity in Cheese.” International Dairy Journal V9(8): pp 553–558. 203 Hutton, J.T., and S. Patton. 1952. “The Origin of Sulfhydryl Groups in Milk Proteins and Their Contributions to ‘Cooked’ Flavor.” Journal of Dairy Science V35 (8): pp. 699– 705. Jolly, Ramesh C., and Frank V. Kosikowski. 1975. “Quantification of Lactones in Ripening Pasteurized Milk Blue Cheese Containing Added Microbial Lipases.” J. Agric. Food Chem. V23 (6): pp. 1175–1176. De Jong, Catrienus, and Herman T. Badings. 1990. “Determination of Free Fatty Acids in Milk and Cheese Procedures for Extraction, Clean up, and Capillary Gas Chromatographic Analysis.” Journal of High Resolution Chromatography V13 (2): pp. 94–98. K. Arfi, H.Spinnler, R. Tache, and P. bonnarme. 2002. “Production of Volatile Compounds by Cheese-ripening Yeasts: Requirement for a Methanethiol Donor for S-methyl Thioacetate Synthesis by Kluyveromyces Lactis.” Applied Microbiology and Biotechnology V58 (4): pp. 503–510. Knecht, E. 1999. “Lysomomal Cysteine Proteinases (2nd Edition).” Cell Biology International V23 (8): pp 590–591. Laan, Harry, Saw Eng Tan, Paul Bruinenberg, Gaëtan Limsowtin, and Malcolm Broome. 1998. “Aminopeptidase Activities of Starter and Non-Starter Lactic Acid Bacteria Under Simulated Cheddar Cheese Ripening Conditions.” International Dairy Journal V8 (4): pp. 267–274. Lane, C.N., and P.F. Fox. 1996a. “Contribution of Starter and Adjunct Lactobacilli to Proteolysis in Cheddar Cheese During Ripening.” International Dairy Journal 6 (7): 715–728. Lane, Cáit N., and Patrick F. Fox. 1997. “Role of Starter Enzymes During Ripening of Cheddar Cheese Made from Pasteurized Milk Under Controlled Microbiological Conditions.” International Dairy Journal V7 (1): pp. 55–63. Larsen, Lotte B., Connie Benfeldt, Lone K. Rasmussen, and Torben E. Petersen. 1996. “Bovine Milk Procathepsin D and Cathepsin D: Coagulation and Milk Protein Degradation.” Journal of Dairy Research V63 (1): pp 119–130. Lau, Kum Y., David M. Barbano, and Robert R. Rasmussen. 1990. “Influence of Pasteurization on Fat and Nitrogen Recoveries and Cheddar Cheese Yield.” Journal of Dairy Science V73 (3): pp. 561–570. Lau, Kum Y. Lau, David M. Barbano, and Robert R. Rasmussen. 1991. “Influence of Pasteurization of Milk on Protein Breakdown in Cheddar Cheese During Aging.” Journal of Dairy Science V74 (3): pp. 727–740. Law, A. J. R., J. M. Banks, D. S. Horne, J. Leaver, and I. G. West. 1994. “Denaturation of the whey proteins in heated milk and their incorporation into Cheddar cheese.” Milchwissenschaft V49 (2): pp. 63–67. Lee, Won-Jae, Dattatreya S. Banavara, Joanne E. Hughes, Jason K. Christiansen, James L. Steele, Jeffery R. Broadbent, and Scott A. Rankin. 2007. “Role of Cystathionine βLyase in Catabolism of Amino Acids to Sulfur Volatiles by Genetic Variants of Lactobacillus Helveticus CNRZ 32.” Applied and Environmental Microbiology V73 (9): pp. 3034–3039. Lin, J. C. C., and I. J. Jeon. 1985. “Headspace Gas Sampling/GC Method for the Quantitative Analysis of Volatile Compounds in Cheese.” Journal of Food Science V50 (3): pp. 843–844. 204 Liu, Shao-Quan, Ross Holland, and Vaughan L Crow. 2001. “Purification and Properties of Intracellular Esterases from Streptococcus Thermophilus.” International Dairy Journal V11 (1–2): pp. 27–35. Lloyd, G. T., J. F. Horwood, and I. Barlow. 1980. “The effect of yoghurt culture YB on the flavour and maturation of Cheddar cheese.” Australian Journal of Dairy Technology V35 (4): pp. 137–139. Lynch, C.M., P.L.H. McSweeney, P.F. Fox, T.M. Cogan, and F.D. Drinan. 1996. “Manufacture of Cheddar Cheese with and Without Adjunct Lactobacilli Under Controlled Microbiological Conditions.” International Dairy Journal V6 (8–9): pp. 851–867. Lynch, C.M., D.D. Muir, J.M. Banks, P.L.H. McSweeney, and P.F. Fox. 1999. “Influence of Adjunct Cultures of Lactobacillus Paracasei Ssp. Paracasei or Lactobacillus Plantarum on Cheddar Cheese Ripening.” Journal of Dairy Science V82 (8): pp. 1618–1628. Lynch, Joanna M., David M. Barbano, and J. Richard Fleming. 2002. “Determination of the Total Nitrogen Content of Hard, Semihard, and Processed Cheese by the Kjeldahl Method: Collaborative Study.” Journal of AOAC International V85 (2): pp. 445–455. Madkor, S.A., P.S. Tong, and M. El Soda. 2000. “Ripening of Cheddar Cheese with Added Attenuated Adjunct Cultures of Lactobacilli.” Journal of Dairy Science V83 (8): pp. 1684–1691. Manning, D. J., Helen R. Chapman, and Zena D. Hosking. 1976. “The Production of Sulphur Compounds in Cheddar Cheese and Their Significance in Flavour Development.” Journal of Dairy Research V43 (2): pp. 313–320. Manning, Donald J., and Carolyn Moore. 1979. “Headspace Analysis of Hard Cheeses.” Journal of Dairy Research V46 (3): pp. 539–545. McSweeney, P.L.H., P.F. Fox, J.A. Lucey, K.N. Jordan, and T.M. Cogan. 1993. “Contribution of the Indigenous Microflora to the Maturation of Cheddar Cheese.” International Dairy Journal V3 (7): pp. 613–634. McSweeney, Paul L. H., Sylvie Pochet, Patrick F. Fox, and Aine Healy. 1994. “Partial Identification of Peptides from the Water-Insoluble Fraction of Cheddar Cheese.” Journal of Dairy Research V61 (4): pp. 587–590. McSweeney, P.L.H., P.F. Fox, and N.F. Olson. 1995. “Proteolysis of Bovine Caseins by Cathepsin D: Preliminary Observations and Comparison with Chymosin.” International Dairy Journal V5 (4): pp. 321–336. McSweeney, Paul L.H., and Maria José Sousa. 2000. “Biochemical Pathways for the Production of Flavour Compounds in Cheeses During Ripening: A Review.” Le Lait V80 (3): pp. 293–324.. Metwalli, Ali A.M, Harmen H.J de Jongh, and Martinus A.J.S van Boekel. 1998. “Heat Inactivation of Bovine Plasmin.” International Dairy Journal V8 (1): pp. 47–56. Milo, C., and G. A. Reineccius. 1997. “Identification and Quantification of Potent Odorants in Regular-Fat and Low-Fat Mild Cheddar Cheese.” Journal of Agricultural and Food Chemistry V45 (9): pp. 3590–3594. Mistry, V.V., L.E. Metzger, and J.L. Maubois. 1996. “Use of Ultrafiltered Sweet Buttermilk in the Manufacture of Reduced Fat Cheddar Cheese.” Journal of Dairy Science V79 (7): pp. 1137–1145. Mistry, V.V., and J.-L. Maubois. 2004. “Application of Membrane Separation Technology to Cheese Production.” In Cheese Chemistry, Physics and Microbiology, Volume 1:pp. 261–VII. Academic Press. 205 Moio, Luigi, Dominique Langlois, Patrick Etievant, and Francesco Addeo. 1993. “Powerful Odorants in Bovine, Ovine, Caprine and Water Buffalo Milk Determined by Means of Gas Chromatography–olfactometry.” Journal of Dairy Research V60 (2): pp. 215– 222. Molimard, P., and H.E. Spinnler. 1996. “Review: Compounds Involved in the Flavor of Surface Mold-Ripened Cheeses: Origins and Properties.” Journal of Dairy Science V79 (2): pp. 169–184. Mooney, J.S, P.F Fox, A Healy, and J Leaver. 1998. “Identification of the Principal Waterinsoluble Peptides in Cheddar Cheese.” International Dairy Journal V8 (9): pp. 813– 818. Muir, D.D., J.M. Banks, and E.A. Hunter. 1996. “Sensory Properties of Cheddar Cheese: Effect of Starter Type and Adjunct.” International Dairy Journal V6 (4): pp. 407– 423. Mulvihill, Donald M., and Patrick F. Fox. 1979. “Proteolytic Specificity of Chymosin on Bovine Αs1-Casein.” Journal of Dairy Research V46 (4): pp. 641–651. Naughton, M. A., and F. Sanger. 1961. “Purification and Specificity of Pancreatic Elastase.” Biochemical Journal V78 (1): pp. 156–163. Niven, C. F., L. G. Buettner, and James B. Evans. 1954. “Thermal Tolerance Studies on the Heterofermentative Lactobacilli That Cause Greening of Cured Meat Products.” Applied Microbiology V2 (1): pp. 26–29. O’Keeffe, R. B., P. F. Fox, and C. Daly. 1976. “Contribution of Rennet and Starter Proteases to Proteolysis in Cheddar Cheese.” Journal of Dairy Research 43 (1): 97–107. O’Keeffe, Arthur M., Patrick F. Fox, and Charles Daly. 1978. “Proteolysis in Cheddar Cheese: Role of Coagulant and Starter Bacteria.” Journal of Dairy Research V45 (3): pp. 465– 477. Olivecrona, T., S. Vilaró, G. Bengtsson-Olivecrona, and P. F Fox. 1992. “Indigenous enzyme in milk. II. Lipases in milk.” In Advanced Dairy Chemistry 1,pp 292–310.. Payne, Martin J., and Rohan G. Kroll. 1991. “Methods for the Separation and Concentration of Bacteria from Foods.” Trends in Food Science & Technology V2 (0): pp. 315–319. Peterson, S.D., and R.T. Marshall. 1990. “Nonstarter Lactobacilli in Cheddar Cheese: A Review.” Journal of Dairy Science V73 (6): pp. 1395–1410. Piraino, Paolo, Eugenio Parente, and Paul L. H. McSweeney. 2004. “Processing of Chromatographic Data for Chemometric Analysis of Peptide Profiles from Cheese Extracts: A Novel Approach.” Journal of Agricultural and Food Chemistry 52 (23): 6904–691. Precetti, A.S., M.P. Oria, and S.S. Nielsen. 1997. “Presence in Bovine Milk of Two Protease Inhibitors of the Plasmin System.” Journal of Dairy Science V80 (8): pp. 1490–1496. Preininger, Martin, and Werner Grosch. 1994. “Evaluation of Key Odorants of the Neutral Volatiles of Emmentaler Cheese by the Calculation of Odour Activity Values.” LWT Food Science and Technology V27 (3): pp. 237–244. Pripp, A.H, Shakeel-Ur-Rehman, P.L.H McSweeney, and P.F Fox. 1999. “Multivariate Statistical Analysis of Peptide Profiles and Free Amino Acids to Evaluate Effects of Single-strain Starters on Proteolysis in Miniature Cheddar-type Cheeses.” International Dairy Journal V9 (7): pp. 473–479. Pripp, Are Hugo, Leszek Stepaniak, and Terje Sørhaug. 2000. “Chemometrical Analysis of Proteolytic Profiles During Cheese Ripening.” International Dairy Journal V10 (4): pp. 249–253. 206 Puchades, R., L. Lemieux, and R.e. Simard. 1989. “Evolution of Free Amino Acids During the Ripening of Cheddar Cheese Containing Added Lactobacilli Strains.” Journal of Food Science V54 (4): pp. 885–888. Qian, M., and G. Reineccius. 2002. “Identification of Aroma Compounds in ParmigianoReggiano Cheese by Gas Chromatography/Olfactometry.” Journal of Dairy Science V85 (6): pp. 1362–1369. Rehman, Shakeel-Ur, J.M Banks, E.Y Brechany, D.D Muir, P.L.H McSweeney, and P.F Fox. 2000. “Influence of Ripening Temperature on the Volatiles Profile and Flavour of Cheddar Cheese Made from Raw or Pasteurised Milk.” International Dairy Journal V10 (1-2): pp. 55–65. Richardson, B. C., L. K. Creamer, K. N. Pearce, and R. E. Munford. 1974. “Comparative Micelle Structure: II. Structure and Composition of Casein Micelles in Ovine and Caprine Milk as Compared with Those in Bovine Milk.” Journal of Dairy Research V41 (2): pp. 239–247. Roy, Denis, Isabelle Mainville, and Francine Mondou. 1997. “Selective Enumeration and Survival of Bifidobacteria in Fresh Cheese.” International Dairy Journal V7 (12): pp. 785–793. Sablé, Sophie, and Gilles Cottenceau. 1999. “Current Knowledge of Soft Cheeses Flavor and Related Compounds.” J. Agric. Food Chem. V47 (12): pp. 4825–4836. Seefeldt, K.E., and B.C. Weimer. 2000. “Diversity of Sulfur Compound Production in Lactic Acid Bacteria.” Journal of Dairy Science V83 (12): pp. 2740–2746. Shakeel-Ur-Rehman, J. M Banks, P. L. H Mcsweeney, and P. F Fox. 2000. “Effect of ripening temperature on the growth and significance of non-starter lactic acid bacteria in Cheddar cheese made from raw or pasteurised milk.” International dairy journal V10 (1-2): pp. 45–53. Singh, T.K., P.F. Fox, P. Højrup, and A. Healy. 1994. “A Scheme for the Fractionation of Cheese Nitrogen and Identification of Principal Peptides.” International Dairy Journal V4 (2): pp. 111–122. Singh, Tanoj K., Patrick F. Fox, and Áine Healy. 1995. “Water-Soluble Peptides in Cheddar Cheese: Isolation and Identification of Peptides in the Diafiltration Retentate of the Water-Soluble Fraction.” Journal of Dairy Research V62 (4). 629–640. Singh, Tanoj K., Patrick F. Fox, and Áine Healy. 1997. “Isolation and Identification of Further Peptides in the Diafiltration Retentate of the Water-soluble Fraction of Cheddar Cheese.” Journal of Dairy Research V64 (3): pp. 433–443. Singh, T. K, M. A Drake, and K. R Cadwallader. 2003. “Flavor of Cheddar Cheese: A Chemical and Sensory Perspective.” Comprehensive Reviews in Food Science and Food Safety V2 (4): pp. 166–189. Smacchi, Emanuele, and Marco Gobbetti. 1998. “Purification and Characterization of Cystathionine Γ-lyase from Lactobacillus Fermentum DT41.” FEMS Microbiology Letters V166 (2): pp. 197–202. Smit, Gerrit, Bart A. Smit, and Wim J.M. Engels. 2005. “Flavour Formation by Lactic Acid Bacteria and Biochemical Flavour Profiling of Cheese Products.” FEMS Microbiology Reviews V29 (3): pp. 591–610. Smith, A. M., and S. Nakai. 1990. “Classification of cheese varieties by multivariate analysis of HPLC profiles.” Canadian Institute of Food Science and Technology Journal V23 (1): pp. 53–58. Sousa, M.J, Y Ardö, and P.L.H McSweeney. 2001. “Advances in the Study of Proteolysis During Cheese Ripening.” International Dairy Journal V11 (4-7): pp. 327–345. 207 Stuart, Mark R., Lan Szu Chou, and Bart C. Weimer. 1999. “Influence of Carbohydrate Starvation and Arginine on Culturability and Amino Acid Utilization of Lactococcus Lactis Subsp. Lactis.” Applied and Environmental Microbiology V65 (2): pp. 665– 673. Subramanian, A., W.J. Harper, and L.E. Rodriguez-Saona. 2009. “Cheddar Cheese Classification Based on Flavor Quality Using a Novel Extraction Method and Fourier Transform Infrared Spectroscopy.” Journal of Dairy Science V92 (1): pp. 87–94. Suriyaphan, Orasa, MaryAnne Drake, X. Q. Chen, and Keith R. Cadwallader. 2001. “Characteristic Aroma Components of British Farmhouse Cheddar Cheese.” Journal of Agricultural and Food Chemistry V49 (3): pp. 1382–1387. Tammam, J. D, A. G Williams, J. Noble, D. Lloyd, J. D Tammam, A. G Williams, J. Noble, and D. Lloyd. 2000. “Amino acid fermentation in non‐starter Lactobacillus spp. isolated from Cheddar cheese, Amino acid fermentation in non‐starter Lactobacillus spp. isolated from Cheddar cheese.” Letters in Applied Microbiology, Letters in Applied Microbiology V30 (5): pp. 370–374. Tanaka, Hidehiko, Nobuyoshi Esaki, and Kenji Soda. 1985. “A Versatile Bacterial Enzyme: L-methionine Γ-lyase.” Enzyme and Microbial Technology V7 (11): pp. 530–537. Thomsen, D., and H. Stapelfeldt. 1990. “Yield and quality of Danbo 45+ cheese from milk heat treated at temperatures between 72 and 95°C.” V44: pp. 39. Tsakalidou, E., and G. Kalantzopoulos. 1992. “Purification and Partial Characterization of an Esterase from Lactococcus Lactis Ssp Lactis Strain ACA-DC 127.” Le Lait V72 (6): pp. 533–543. Turner, K. W, and T. D Thomas. 1980. “Lactose fermentation in Cheddar cheese and the effect of salt.” New Zealand Journal of Dairy Science and Technology V15 (3): pp. 265–276. Upadhyay, V.K., P.L.H. McSweeney, A.A.A. Magboul, and P.F. Fox. 2004. “Proteolysis in Cheese During Ripening.” In Cheese Chemistry, Physics and Microbiology, Volume 1:pp. 391–433. Academic Press. Urbach, Gerda. 1993. “Relations Between Cheese Flavour and Chemical Composition.” International Dairy Journal V3 (4–6): pp. 389–422. Urbach, Gerda. 1995. “Contribution of Lactic Acid Bacteria to Flavour Compound Formation in Dairy Products.” International Dairy Journal V5 (8): pp. 877–903. USDA. 2011. Dairy Products Annual Sumary. Vazquez-Landaverde, P.A., J.A. Torres, and M.C. Qian. 2006. “Quantification of Trace Volatile Sulfur Compounds in Milk by Solid-Phase Microextraction and Gas Chromatography–Pulsed Flame Photometric Detection.” Journal of Dairy Science V89 (8): pp. 2919–2927. Verdi, Robert J., and David M. Barbano. 1991. “Properties of Proteases from Milk Somatic Cells and Blood Leukocytes.” Journal of Dairy Science V74 (7): pp. 2077–2081. Visser, Servas 1983. “Bitter flavour in cheese, 2: model studies on the formation and degradation of bitter peptides by proteolytic enzymes from calf rennet, starter cells and starter cell fractions.” Netherlands Milk and Dairy Journal V37 (3): pp. 169–180. Visser, Servaas. 1993. “Proteolytic Enzymes and Their Relation to Cheese Ripening and Flavor: An Overview.” Journal of Dairy Science V76 (1): pp. 329–350 Voigt, Daniela D., François Chevalier, John A. Donaghy, Margaret F. Patterson, Michael C. Qian, and Alan L. Kelly. 2012. “Effect of High-pressure Treatment of Milk for Cheese Manufacture on Proteolysis, Lipolysis, Texture and Functionality of Cheddar 208 Cheese During Ripening.” Innovative Food Science & Emerging Technologies V13 (0): pp. 23–30. Wallace J.M., and Fox P.F. 1997. “Effect of Adding Free Amino Acids to Cheddar Cheese Curd on Proteolysis, Flavour and Texture Development.” International Dairy Journal V7 (2): pp. 157–167. Walstra, Pieter, T. J Geurts, A. Noomen, A. Jellema, and M. A. J. S. van Boekel. 1999. “Cheese Varieties.” In Dairy technology: principles of milk properties and processes, pp 651–708. Marcel Dekker. Walstra, Pieter, Jan T. M. Wouters, and Tom J. Geurts. 2006. Dairy Science And Technology. CRC Press. Weimer, B. C. 2007. Improving the flavour of cheese. CRC Press. Weimer, Bart, Kimberly Seefeldt, and Benjamin Dias. 1999. “Sulfur Metabolism in Bacteria Associated with Cheese.” Antonie Van Leeuwenhoek 76 (1): 247–261. Wijesundera, C., and L. Drury. “Role of milk fat in production of cheddar cheese flavour using a fat-substituted cheese model.” Australian journal of dairy technology V54 (1): pp. 28–35. Wilkinson, Martin G., Timothy P. Guinee, Daniel M. O’Callaghan, and Patrick F. Fox. 1994. “Autolysis and Proteolysis in Different Strains of Starter Bacteria During Cheddar Cheese Ripening.” Journal of Dairy Research V61 (2): pp. 249–262. De Wit, M., G. Osthoff, B.C. Viljoen, and A. Hugo. 2005. “A Comparative Study of Lipolysis and Proteolysis in Cheddar Cheese and Yeast-inoculated Cheddar Cheeses During Ripening.” Enzyme and Microbial Technology V37 (6): pp. 606–616. de Wit, Rudy, and Hans Nieuwenhuijse. 2008. “Kinetic Modelling of the Formation of Sulphur-containing Flavour Components During Heat-treatment of Milk.” International Dairy Journal V18 (5): pp. 539–547. Wong, N. P, R. Ellis, and D. E LaCroix. “Quantitative determination of lactones in Cheddar cheese.” Journal of Dairy Science V58: pp. 1437–1441. Yvon, Mireille, Claire Chabanet, and Jean-Pierre Pélissier. 1989. “Solubility of Peptides in Trichloroacetic Acid (TCA) Solutions Hypothesis on the Precipitation Mechanism.” International Journal of Peptide and Protein Research V34 (3): pp. 166–176. Yvon, Mireille, and Liesbeth Rijnen. 2001. “Cheese Flavour Formation by Amino Acid Catabolism.” International Dairy Journal V11 (4–7): pp. 185–201. Zehentbauer, Gerhard, and G. A. Reineccius. 2002. “Determination of Key Aroma Components of Cheddar Cheese Using Dynamic Headspace Dilution Assay.” Flavour and Fragrance Journal V17 (4): pp. 300–305. 209 APPENDICES APENDIX A ANOVAS FFA Heat-shocked Vs Pasteurized Table 2 Anova FFA of Heat-shocked Vs Pasteurized General Linear Model: C4:0, C6:0, ... versus treatment, time Factor treatment time Type random fixed Levels 2 7 Values HS, P 10M, 12M, 14M, 2M, 4M, 6M, 8M Analysis of Variance for C4:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 8.026 605.649 3.354 * 617.029 Adj SS 8.026 605.649 3.354 * Adj MS 8.026 100.941 0.559 * F 14.36 180.56 ** P 0.009 0.000 Analysis of Variance for C6:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 2.9257 122.0326 1.5343 * 126.4926 Adj SS 2.9257 122.0326 1.5343 * Adj MS 2.9257 20.3388 0.2557 * F 11.44 79.54 ** P 0.015 0.000 Analysis of Variance for C8:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 0.6487 19.9431 0.3181 * 20.9099 Adj SS 0.6487 19.9431 0.3181 * Adj MS 0.6487 3.3239 0.0530 * F 12.24 62.70 ** P 0.013 0.000 Analysis of Variance for C10:0, using Adjusted SS for Tests Source treatment time treatment*time DF 1 6 6 Seq SS 3.1114 71.3699 0.8686 Adj SS 3.1114 71.3699 0.8686 Adj MS 3.1114 11.8950 0.1448 F 21.49 82.17 ** P 0.004 0.000 210 Error Total 0 13 * 75.3499 * * Analysis of Variance for C12:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 3.2335 102.7589 1.8224 * 107.8147 Adj SS 3.2335 102.7589 1.8224 * Adj MS 3.2335 17.1265 0.3037 * F 10.65 56.39 ** P 0.017 0.000 Analysis of Variance for C14:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 96.68 1213.26 23.50 * 1333.44 Adj SS 96.68 1213.26 23.50 * Adj MS 96.68 202.21 3.92 * F 24.68 51.63 ** P 0.003 0.000 Analysis of Variance for C14:1, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 0.33913 8.01428 0.45428 * 8.80769 Adj SS 0.33913 8.01428 0.45428 * Adj MS 0.33913 1.33571 0.07571 * F 4.48 17.64 ** P 0.079 0.001 Analysis of Variance for C16:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 503.88 3988.00 150.99 * 4642.87 Adj SS 503.88 3988.00 150.99 * Adj MS 503.88 664.67 25.17 * F 20.02 26.41 ** P 0.004 0.000 Analysis of Variance for C16:1, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 0.6079 20.0450 0.7335 * 21.3863 Adj SS 0.6079 20.0450 0.7335 * Adj MS 0.6079 3.3408 0.1223 * F 4.97 27.33 ** P 0.067 0.000 Analysis of Variance for C18:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 25.42 1361.62 42.00 * 1429.04 Adj SS 25.42 1361.62 42.00 * Adj MS 25.42 226.94 7.00 * F 3.63 32.42 ** P 0.105 0.000 Analysis of Variance for C18:1, using Adjusted SS for Tests 211 Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 92.14 6277.51 247.49 * 6617.13 Adj SS 92.14 6277.51 247.49 * Adj MS 92.14 1046.25 41.25 * F 2.23 25.36 ** P 0.186 0.001 Analysis of Variance for C18:2, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 0.024 1057.288 50.452 * 1107.764 Adj SS 0.024 1057.288 50.452 * Adj MS 0.024 176.215 8.409 * F 0.00 20.96 ** P 0.959 0.001 Analysis of Variance for C18:3, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 0.00072 2.59199 0.09388 * 2.68659 Adj SS 0.00072 2.59199 0.09388 * Adj MS 0.00072 0.43200 0.01565 * F 0.05 27.61 ** P 0.837 0.000 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. TCCA Vs CRP Table 3 Anova FFA of TCCA Vs CRP General Linear Model: C4:0, C6:0, ... versus treatment, time Factor treatment time Type random fixed Levels 2 7 Values CRP, TCCA 10M, 12M, 14M, 2M, 4M, 6M, 8M Analysis of Variance for C4:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 26.612 444.280 40.857 * 511.749 Adj SS 26.612 444.280 40.857 * Adj MS 26.612 74.047 6.810 * F 3.91 10.87 ** P 0.095 0.005 Analysis of Variance for C6:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 5.9080 73.1742 5.4894 * 84.5717 Adj SS 5.9080 73.1742 5.4894 * Adj MS 5.9080 12.1957 0.9149 * F 6.46 13.33 ** P 0.044 0.003 212 Analysis of Variance for C8:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 7.9018 11.0297 0.7586 * 19.6901 Adj SS 7.9018 11.0297 0.7586 * Adj MS 7.9018 1.8383 0.1264 * F 62.50 14.54 ** P 0.000 0.002 Analysis of Variance for C10:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 47.0056 51.2884 10.9329 * 109.2269 Adj SS 47.0056 51.2884 10.9329 * Adj MS 47.0056 8.5481 1.8221 * F 25.80 4.69 ** P 0.002 0.041 Analysis of Variance for C12:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 80.376 65.265 9.070 * 154.711 Adj SS 80.376 65.265 9.070 * Adj MS 80.376 10.878 1.512 * F 53.17 7.20 ** P 0.000 0.015 Analysis of Variance for C14:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 817.27 498.97 167.39 * 1483.62 Adj SS 817.27 498.97 167.39 * Adj MS 817.27 83.16 27.90 * F 29.29 2.98 ** P 0.002 0.105 Analysis of Variance for C14:1, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 10.2390 5.6458 1.7850 * 17.6698 Adj SS 10.2390 5.6458 1.7850 * Adj MS 10.2390 0.9410 0.2975 * F 34.42 3.16 ** P 0.001 0.093 Analysis of Variance for C16:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 1152.33 1788.07 273.61 * 3214.01 Adj SS 1152.33 1788.07 273.61 * Adj MS 1152.33 298.01 45.60 * F 25.27 6.54 ** P 0.002 0.019 Analysis of Variance for C16:1, using Adjusted SS for Tests Source treatment time DF 1 6 Seq SS 14.9829 18.9973 Adj SS 14.9829 18.9973 Adj MS 14.9829 3.1662 F 22.34 4.72 P 0.003 0.040 213 treatment*time Error Total 6 0 13 4.0239 * 38.0041 4.0239 * 0.6707 * ** Analysis of Variance for C18:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 296.820 537.598 34.560 * 868.978 Adj SS 296.820 537.598 34.560 * Adj MS 296.820 89.600 5.760 * F 51.53 15.56 ** P 0.000 0.002 Analysis of Variance for C18:1, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 3159.42 2227.26 613.90 * 6000.58 Adj SS 3159.42 2227.26 613.90 * Adj MS 3159.42 371.21 102.32 * F 30.88 3.63 ** P 0.001 0.071 Analysis of Variance for C18:2, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 3.4844 93.9442 7.2245 * 104.6531 Adj SS 3.4844 93.9442 7.2245 * Adj MS 3.4844 15.6574 1.2041 * F 2.89 13.00 ** P 0.140 0.003 Analysis of Variance for C18:3, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 6 6 0 13 Seq SS 12.2826 6.4034 3.5308 * 22.2169 Adj SS 12.2826 6.4034 3.5308 * Adj MS 12.2826 1.0672 0.5885 * F 20.87 1.81 ** P 0.004 0.244 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. Adjunct culture Vs Not Table 4 Anova FFA of Adjunct culture Vs Not General Linear Model: C4:0, C6:0, ... versus treatment, time Factor treatment time Type random fixed Levels 2 5 Values A, N 2, 4, 6, 8, 10 214 Analysis of Variance for C4:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 12.877 352.397 1.314 * 366.588 Adj SS 12.877 352.397 1.314 * Adj MS 12.877 88.099 0.329 * F 39.19 268.16 ** P 0.003 0.000 Analysis of Variance for C6:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 3.9297 44.7948 0.3472 * 49.0717 Adj SS 3.9297 44.7948 0.3472 * Adj MS 3.9297 11.1987 0.0868 * F 45.28 129.03 ** P 0.003 0.000 Analysis of Variance for C8:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 0.90706 6.38731 0.09790 * 7.39227 Adj SS 0.90706 6.38731 0.09790 * Adj MS 0.90706 1.59683 0.02448 * F 37.06 65.24 ** P 0.004 0.001 Analysis of Variance for C10:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 14.2226 28.8278 0.5657 * 43.6162 Adj SS 14.2226 28.8278 0.5657 * Adj MS 14.2226 7.2069 0.1414 * F 100.56 50.96 ** P 0.001 0.001 Analysis of Variance for C12:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 21.8046 37.0475 1.7017 * 60.5538 Adj SS 21.8046 37.0475 1.7017 * Adj MS 21.8046 9.2619 0.4254 * F 51.25 21.77 ** P 0.002 0.006 Analysis of Variance for C14:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 64.010 441.849 5.952 * 511.811 Adj SS 64.010 441.849 5.952 * Adj MS 64.010 110.462 1.488 * F 43.02 74.24 ** P 0.003 0.001 Analysis of Variance for C14:1, using Adjusted SS for Tests Source treatment DF 1 Seq SS 0.01894 Adj SS 0.01894 Adj MS 0.01894 F 0.95 P 0.386 215 time treatment*time Error Total 4 4 0 9 3.44150 0.08004 * 3.54048 3.44150 0.08004 * 0.86038 0.02001 * 43.00 ** 0.002 Analysis of Variance for C16:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 131.04 2371.44 28.37 * 2530.86 Adj SS 131.04 2371.44 28.37 * Adj MS 131.04 592.86 7.09 * F 18.48 83.59 ** P 0.013 0.000 Analysis of Variance for C16:1, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 3.8062 18.6749 0.1284 * 22.6095 Adj SS 3.8062 18.6749 0.1284 * Adj MS 3.8062 4.6687 0.0321 * F 118.54 145.41 ** P 0.000 0.000 Analysis of Variance for C18:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 45.274 299.232 6.749 * 351.256 Adj SS 45.274 299.232 6.749 * Adj MS 45.274 74.808 1.687 * F 26.83 44.33 ** P 0.007 0.001 Analysis of Variance for C18:1, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 183.67 2647.99 51.89 * 2883.55 Adj SS 183.67 2647.99 51.89 * Adj MS 183.67 662.00 12.97 * F 14.16 51.03 ** P 0.020 0.001 Analysis of Variance for C18:2, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 7.060 82.232 7.758 * 97.049 Adj SS 7.060 82.232 7.758 * Adj MS 7.060 20.558 1.939 * F 3.64 10.60 ** P 0.129 0.021 Analysis of Variance for C18:3, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 0.28568 1.18875 0.15069 * 1.62512 Adj SS 0.28568 1.18875 0.15069 * Adj MS 0.28568 0.29719 0.03767 * ** Denominator of F-test is zero or undefined. F 7.58 7.89 ** P 0.051 0.035 216 S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. Good Vs Week Table 5 Anova FFA of Good Vs Weak General Linear Model: C4:0, C6:0, ... versus treatment, time Factor treatment time Type random fixed Levels 2 5 Values Good, Weak 2, 6, 9, 12, 15 Analysis of Variance for C4:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 16.23 899.80 60.72 * 976.75 Adj SS 16.23 899.80 60.72 * Adj MS 16.23 224.95 15.18 * F 1.07 14.82 ** P 0.359 0.011 Analysis of Variance for C6:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 18.901 155.502 12.161 * 186.564 Adj SS 18.901 155.502 12.161 * Adj MS 18.901 38.876 3.040 * F 6.22 12.79 ** P 0.067 0.015 Analysis of Variance for C8:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 31.7506 26.5553 3.2097 * 61.5156 Adj SS 31.7506 26.5553 3.2097 * Adj MS 31.7506 6.6388 0.8024 * F 39.57 8.27 ** P 0.003 0.032 Analysis of Variance for C10:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 15.901 87.759 12.757 * 116.417 Adj SS 15.901 87.759 12.757 * Adj MS 15.901 21.940 3.189 * F 4.99 6.88 ** P 0.089 0.044 Analysis of Variance for C12:0, using Adjusted SS for Tests Source treatment time treatment*time Error DF 1 4 4 0 Seq SS 20.336 135.429 31.303 * Adj SS 20.336 135.429 31.303 * Adj MS 20.336 33.857 7.826 * F 2.60 4.33 ** P 0.182 0.093 217 Total 9 187.068 Analysis of Variance for C14:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 147.99 1152.15 107.94 * 1408.08 Adj SS 147.99 1152.15 107.94 * Adj MS 147.99 288.04 26.99 * F 5.48 10.67 ** P 0.079 0.021 Analysis of Variance for C14:1, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 0.7794 10.9725 1.5122 * 13.2641 Adj SS 0.7794 10.9725 1.5122 * Adj MS 0.7794 2.7431 0.3781 * F 2.06 7.26 ** P 0.224 0.040 Analysis of Variance for C16:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 2758.6 6210.3 347.2 * 9316.2 Adj SS 2758.6 6210.3 347.2 * Adj MS 2758.6 1552.6 86.8 * F 31.78 17.88 ** P 0.005 0.008 Analysis of Variance for C16:1, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 86.795 51.875 7.414 * 146.084 Adj SS 86.795 51.875 7.414 * Adj MS 86.795 12.969 1.853 * F 46.83 7.00 ** P 0.002 0.043 Analysis of Variance for C18:0, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 22.56 2848.10 460.17 * 3330.83 Adj SS 22.56 2848.10 460.17 * Adj MS 22.56 712.02 115.04 * F 0.20 6.19 ** P 0.681 0.053 Analysis of Variance for C18:1, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 2415.7 11093.8 871.1 * 14380.6 Adj SS 2415.7 11093.8 871.1 * Adj MS 2415.7 2773.4 217.8 * F 11.09 12.73 ** P 0.029 0.015 Analysis of Variance for C18:2, using Adjusted SS for Tests Source treatment DF 1 Seq SS 859.53 Adj SS 859.53 Adj MS 859.53 F 13.92 P 0.020 218 time treatment*time Error Total 4 4 0 9 346.00 246.92 * 1452.45 346.00 246.92 * 86.50 61.73 * 1.40 ** 0.376 Analysis of Variance for C18:3, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 48.3241 6.8976 6.6459 * 61.8677 Adj SS 48.3241 6.8976 6.6459 * Adj MS 48.3241 1.7244 1.6615 * F 29.09 1.04 ** P 0.006 0.486 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. VSC’s Heat-shocked Vs Pasteurized Table 6 Anova VSC of Heat-Shocked Vs Pasteurized General Linear Model: DMS, H2S, MeSH versus treatment, time Factor treatment time Type random fixed Levels 2 10 Values HS, P 2, 5, 6, 7, 8, 9, 10, 12, 14, 15 Analysis of Variance for DMS, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 9 9 0 19 Seq SS 2108.64 1316.67 295.56 * 3720.87 Adj SS 2108.64 1316.67 295.56 * Adj MS 2108.64 146.30 32.84 * F 64.21 4.45 ** P 0.000 0.018 Analysis of Variance for H2S, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 9 9 0 19 Seq SS 291.20 1245.71 1245.30 * 2782.21 Adj SS 291.20 1245.71 1245.30 * Adj MS 291.20 138.41 138.37 * F 2.10 1.00 ** P 0.181 0.500 Analysis of Variance for MeSH, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 9 9 0 19 Seq SS 0.189718 0.596347 0.137296 * 0.923361 Adj SS 0.189718 0.596347 0.137296 * Adj MS 0.189718 0.066261 0.015255 * F 12.44 4.34 ** P 0.006 0.020 219 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. TCCA Vs CRP Table 7 Anova VSC TCCA Vs CRP General Linear Model: DMS, H2S, MeSH versus treatment, time Factor treatment time Type random fixed Levels 2 8 Values CRP, TCCA 10M, 12M, 14M, 15M, 2M, 4M, 6M, 8M Analysis of Variance for DMS, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 7 7 0 15 Seq SS 10.461 743.070 16.942 * 770.472 Adj SS 10.461 743.070 16.942 * Adj MS 10.461 106.153 2.420 * F 4.32 43.86 ** P 0.076 0.000 Analysis of Variance for H2S, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 7 7 0 15 Seq SS 76.434 147.463 84.326 * 308.223 Adj SS 76.434 147.463 84.326 * Adj MS 76.434 21.066 12.047 * F 6.34 1.75 ** P 0.040 0.239 Analysis of Variance for MeSH, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 7 7 0 15 Seq SS 0.009245 0.170565 0.022100 * 0.201909 Adj SS 0.009245 0.170565 0.022100 * Adj MS 0.009245 0.024366 0.003157 * F 2.93 7.72 ** P 0.131 0.008 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. Adjunct culture Vs Not Table 8 Anova of VSC Adjunct Vs Not General Linear Model: DMS, H2S, MeSH versus treatment, time Factor treatment time Type random fixed Levels 2 5 Values adj, No 10M, 2M, 4M, 6M, 8M 220 Analysis of Variance for DMS, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 24.643 432.056 6.056 * 462.755 Adj SS 24.643 432.056 6.056 * Adj MS 24.643 108.014 1.514 * F 16.28 71.34 ** P 0.016 0.001 Analysis of Variance for H2S, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 116.266 87.452 43.852 * 247.570 Adj SS 116.266 87.452 43.852 * Adj MS 116.266 21.863 10.963 * F 10.61 1.99 ** P 0.031 0.260 Analysis of Variance for MeSH, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 0.021563 0.174265 0.005715 * 0.201542 Adj SS 0.021563 0.174265 0.005715 * Adj MS 0.021563 0.043566 0.001429 * F 15.09 30.49 ** P 0.018 0.003 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. Good Vs Weak Table 9 Anova of VSC Good Vs Weak General Linear Model: DMS, H2S, MeSH versus treatment, time Factor treatment time Type random fixed Levels 2 5 Values good, weak 12M, 15M, 2M, 6M, 9M Analysis of Variance for DMS, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 199.62 1332.04 65.53 * 1597.18 Adj SS 199.62 1332.04 65.53 * Adj MS 199.62 333.01 16.38 * F 12.18 20.33 ** P 0.025 0.006 Analysis of Variance for H2S, using Adjusted SS for Tests Source DF Seq SS Adj SS Adj MS F P 221 treatment time treatment*time Error Total 1 4 4 0 9 31.2594 25.5177 8.9782 * 65.7552 31.2594 25.5177 8.9782 * 31.2594 6.3794 2.2445 * 13.93 2.84 ** 0.020 0.168 Analysis of Variance for MeSH, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 0.046490 0.128501 0.026157 * 0.201148 Adj SS 0.046490 0.128501 0.026157 * Adj MS 0.046490 0.032125 0.006539 * F 7.11 4.91 ** P 0.056 0.076 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. TKN Heat-shocked Vs Pasteurized Table 10 Anova of TKN Heat-Shocked Vs Pasteurized General Linear Model: WSN, TCA, PTA versus treatment, time Factor treatment time Type random fixed Levels 2 10 Values HS, P 10m, 12m, 14m, 16m, 18m, 20m, 2m, 4m, 6m, 8m Analysis of Variance for WSN, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 9 9 0 19 Seq SS 22.4890 65.5854 1.5960 * 89.6704 Adj SS 22.4890 65.5854 1.5960 * Adj MS 22.4890 7.2873 0.1773 * F 126.82 41.09 ** P 0.000 0.000 Analysis of Variance for TCA, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 9 9 0 19 Seq SS 0.7169 23.8577 0.6346 * 25.2092 Adj SS 0.7169 23.8577 0.6346 * Adj MS 0.7169 2.6509 0.0705 * F 10.17 37.59 ** P 0.011 0.000 Analysis of Variance for PTA, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 9 9 0 19 Seq SS 0.04105 2.89366 0.01552 * 2.95023 Adj SS 0.04105 2.89366 0.01552 * Adj MS 0.04105 0.32152 0.00172 * F 23.80 186.44 ** P 0.001 0.000 222 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. TCCA Vs CRP Table 11 TKN TCCA Vs CRP General Linear Model: WSN, TCA, PTA versus treatment, time Factor treatment time Type random fixed Levels 2 8 Values CRP, TCCA 10m, 12m, 14m, 15m, 2m, 4m, 6m, 8m Analysis of Variance for WSN, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 7 7 0 15 Seq SS 4.0102 132.8055 1.1492 * 137.9648 Adj SS 4.0102 132.8055 1.1492 * Adj MS 4.0102 18.9722 0.1642 * F 24.43 115.57 ** P 0.002 0.000 Analysis of Variance for TCA, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 7 7 0 15 Seq SS 1.4495 16.4812 0.3755 * 18.3062 Adj SS 1.4495 16.4812 0.3755 * Adj MS 1.4495 2.3545 0.0536 * F 27.02 43.89 ** P 0.001 0.000 Analysis of Variance for PTA, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 7 7 0 15 Seq SS 0.72285 4.10058 0.12258 * 4.94600 Adj SS 0.72285 4.10058 0.12258 * Adj MS 0.72285 0.58580 0.01751 * F 41.28 33.45 ** P 0.000 0.000 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. Adjunct culture Vs Not Table 12 TKN Adjunct culture Vs Not General Linear Model: WSN, TCA, PTA versus treatment, time Factor treatment time Type random fixed Levels 2 5 Values Adj, Without 10m, 2m, 4m, 6m, 8m 223 Analysis of Variance for WSN, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 5.114 83.168 1.857 * 90.139 Adj SS 5.114 83.168 1.857 * Adj MS 5.114 20.792 0.464 * F 11.01 44.78 ** P 0.029 0.001 Analysis of Variance for TCA, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 1.19657 5.39730 0.36319 * 6.95706 Adj SS 1.19657 5.39730 0.36319 * Adj MS 1.19657 1.34933 0.09080 * F 13.18 14.86 ** P 0.022 0.011 Analysis of Variance for PTA, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 0.11281 1.25264 0.08796 * 1.45342 Adj SS 0.11281 1.25264 0.08796 * Adj MS 0.11281 0.31316 0.02199 * F 5.13 14.24 ** P 0.086 0.012 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. Good Vs Weak Table 13 TKN Good Vs Weak General Linear Model: WSN, TCA, PTA versus treatment, time Factor treatment time Type random fixed Levels 2 5 Values Good, Weak 12m, 15m, 2m, 6m, 9m Analysis of Variance for WSN, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 26.1002 46.5563 2.8000 * 75.4564 Adj SS 26.1002 46.5563 2.8000 * Adj MS 26.1002 11.6391 0.7000 * F 37.29 16.63 ** P 0.004 0.009 Analysis of Variance for TCA, using Adjusted SS for Tests Source treatment time DF 1 4 Seq SS 2.45150 5.36704 Adj SS 2.45150 5.36704 Adj MS 2.45150 1.34176 F 118.81 65.03 P 0.000 0.001 224 treatment*time Error Total 4 0 9 0.08253 * 7.90107 0.08253 * 0.02063 * ** Analysis of Variance for PTA, using Adjusted SS for Tests Source treatment time treatment*time Error Total DF 1 4 4 0 9 Seq SS 2.14584 5.83349 0.15910 * 8.13843 Adj SS 2.14584 5.83349 0.15910 * Adj MS 2.14584 1.45837 0.03978 * F 53.95 36.66 ** P 0.002 0.002 ** Denominator of F-test is zero or undefined. S = * * NOTE * Could not graph the specified residual type because MSE = 0 or the degrees of freedom for error = 0. 225 APENDIX B PCA CORRELATION MATRIX HEAT SHCOKED VS PASTEURIZATION Table 14 Correlation matrix for HEAT SHCOKED VS PASTEURIZATION Principal Component Analysis: Eigenanalysis of the Correlation Matrix Eigenvalue Proportion Cumulative 6.5409 0.503 0.503 2.9590 0.228 0.731 1.2323 0.095 0.826 0.7095 0.055 0.880 0.5463 0.042 0.922 Eigenvalue Proportion Cumulative 0.0950 0.007 0.988 0.0674 0.005 0.993 0.0522 0.004 0.997 0.0324 0.002 0.999 0.0100 0.001 1.000 Variable PC1 Temperature 0.360 Time 0.072 seg 12-20 0.325 seg 20-25 0.258 seg 25-30 -0.291 seg 30-35 -0.285 seg 35-40 0.331 seg 40-45 0.299 seg 45-50 -0.150 seg 50-55 -0.290 seg 55-60 -0.346 seg 60-65 -0.183 PC2 0.182 -0.440 0.074 0.144 -0.193 -0.292 -0.104 -0.225 0.498 0.397 0.123 -0.353 PC3 0.077 0.500 0.221 -0.467 0.003 0.182 -0.024 -0.310 0.198 -0.170 0.080 -0.471 PC4 -0.093 -0.161 0.264 0.234 0.320 0.176 -0.379 -0.260 -0.409 0.137 -0.237 -0.053 PC5 -0.364 0.280 0.455 0.208 -0.228 -0.404 -0.171 -0.298 -0.162 0.036 0.079 0.025 Variable PC9 Temperature 0.052 Time 0.076 seg 12-20 -0.058 seg 20-25 0.549 seg 25-30 -0.260 seg 30-35 -0.297 seg 35-40 0.315 seg 40-45 -0.260 seg 45-50 0.055 seg 50-55 -0.074 seg 55-60 0.591 seg 60-65 -0.071 PC10 0.322 -0.002 0.633 0.081 0.181 -0.085 -0.030 0.373 0.004 0.287 0.230 0.292 PC11 0.662 0.381 -0.272 0.117 -0.036 -0.117 -0.296 -0.005 0.291 0.033 -0.146 0.176 PC12 -0.304 0.271 0.095 0.038 0.629 0.112 -0.119 -0.212 0.530 -0.189 0.198 0.027 PC13 -0.164 0.432 -0.261 0.440 -0.075 0.396 -0.093 0.309 -0.141 0.040 -0.136 -0.019 TCCA Vs CRP 0.3552 0.027 0.949 PC6 0.149 -0.035 0.052 -0.198 -0.670 0.314 -0.259 -0.172 0.066 0.071 -0.322 0.419 0.2357 0.018 0.968 PC7 -0.034 -0.185 -0.064 0.196 -0.314 0.164 -0.397 0.168 -0.119 -0.505 0.547 -0.188 0.1641 0.013 0.980 PC8 -0.063 0.016 0.029 -0.040 -0.223 0.001 0.520 -0.217 -0.289 0.246 0.521 0.241 226 Table 15 Coreelation matrix of TCCA Vs CRP Principal Component Analysis: Eigenanalysis of the Correlation Matrix 35 cases used, 1 cases contain missing values Eigenvalue Proportion Cumulative 7.4190 0.571 0.571 1.7891 0.138 0.708 1.2763 0.098 0.806 0.8856 0.068 0.875 0.4454 0.034 0.909 Eigenvalue Proportion Cumulative 0.1580 0.012 0.977 0.1165 0.009 0.986 0.0801 0.006 0.992 0.0552 0.004 0.997 0.0426 0.003 1.000 Variable Location Time Batch seg 12-20 seg 20-25 seg 25-30 seg 30-35 seg 35-40 seg 40-45 seg 45-50 seg 50-55 seg 55-60 seg 60-65 PC1 0.060 0.334 0.269 0.255 0.317 -0.307 -0.335 0.225 -0.122 -0.289 -0.307 -0.326 -0.266 PC2 0.573 0.043 -0.213 -0.409 -0.196 -0.242 0.001 0.359 0.434 -0.183 -0.242 -0.062 0.061 PC3 0.524 -0.208 0.294 -0.268 0.091 -0.067 0.020 -0.323 -0.557 -0.219 -0.067 0.091 -0.071 PC4 0.006 -0.173 -0.352 0.258 0.019 0.182 -0.080 0.357 -0.363 -0.451 0.182 0.084 0.517 PC5 -0.034 -0.059 0.178 0.027 -0.404 0.385 -0.118 0.398 -0.070 -0.166 0.385 0.347 -0.571 Variable Location Time Batch seg 12-20 seg 20-25 seg 25-30 seg 30-35 seg 35-40 seg 40-45 seg 45-50 seg 50-55 seg 55-60 seg 60-65 PC9 0.288 0.178 0.245 0.174 0.484 0.047 -0.176 -0.006 -0.096 -0.228 0.527 0.427 -0.076 PC10 -0.198 0.131 -0.090 -0.052 0.187 -0.264 0.332 0.079 -0.227 -0.019 -0.351 0.563 0.469 PC11 -0.060 0.140 -0.081 -0.224 -0.365 -0.068 0.572 -0.377 -0.002 -0.288 0.316 0.170 -0.318 PC12 0.089 -0.369 0.441 0.590 -0.192 0.127 0.377 -0.042 0.018 0.302 -0.025 0.153 -0.002 PC13 0.392 -0.607 -0.293 -0.110 0.205 0.285 0.163 -0.091 0.017 -0.424 -0.144 -0.016 0.147 0.2948 0.023 0.932 PC6 -0.125 0.187 0.656 -0.085 -0.206 -0.070 0.417 0.340 -0.072 0.035 -0.070 -0.054 0.407 0.2420 0.019 0.950 PC7 0.148 0.344 0.191 -0.068 0.131 0.196 -0.371 -0.175 0.156 0.083 0.196 0.610 0.317 0.1953 0.015 0.965 PC8 0.000 0.290 -0.008 0.415 -0.384 -0.310 0.063 -0.355 0.183 -0.436 -0.310 0.175 0.019 ADJUCNT CULTURE Vs NOT Table 16 Correlation matrix of Adj Vs Not Principal Component Analysis: Eigenanalysis of the Correlation Matrix 35 cases used, 1 cases contain missing values Eigenvalue 7.6670 2.0734 1.1046 0.5703 0.4492 0.3360 0.2211 0.1501 227 Proportion Cumulative 0.590 0.590 0.159 0.749 0.085 0.834 0.044 0.878 0.035 0.913 Eigenvalue Proportion Cumulative 0.1360 0.010 0.978 0.1142 0.009 0.986 0.0863 0.007 0.993 0.0581 0.004 0.997 0.0335 0.003 1.000 Variable Time Culture Batch seg 12-20 seg 20-25 seg 25-30 seg 30-35 seg 35-40 seg 40-45 seg 45-50 seg 50-55 seg 55-60 seg 60-65 PC1 0.294 0.049 -0.146 0.220 0.256 -0.293 -0.328 0.329 0.320 -0.300 -0.302 -0.314 0.304 PC2 -0.294 0.254 0.570 0.358 0.274 -0.227 -0.055 0.153 0.166 0.235 0.218 -0.149 -0.299 PC3 -0.129 -0.860 0.223 0.050 0.171 0.230 -0.029 0.144 0.159 -0.151 0.186 -0.032 0.062 PC4 -0.270 0.041 -0.146 0.737 -0.028 0.369 -0.006 0.031 -0.213 -0.039 -0.323 0.244 0.108 PC5 0.254 0.002 0.358 0.224 -0.787 -0.063 0.081 0.073 0.236 -0.243 0.062 0.060 0.009 Variable Time Culture Batch seg 12-20 seg 20-25 seg 25-30 seg 30-35 seg 35-40 seg 40-45 seg 45-50 seg 50-55 seg 55-60 seg 60-65 PC9 -0.089 0.002 -0.182 -0.014 0.017 -0.331 -0.058 0.002 -0.225 -0.126 -0.713 0.140 0.501 PC10 0.342 -0.044 0.009 0.145 0.058 -0.074 -0.775 -0.017 -0.248 -0.171 0.007 -0.384 -0.111 PC11 0.034 -0.084 -0.268 -0.678 0.019 0.400 -0.045 0.059 0.201 -0.473 -0.067 -0.149 0.066 PC12 0.288 -0.085 0.154 -0.157 0.029 -0.301 0.284 -0.360 -0.445 -0.474 0.212 0.262 -0.152 PC13 0.434 -0.657 -0.034 0.375 0.112 0.330 0.234 -0.035 0.102 -0.017 -0.134 -0.027 0.177 0.026 0.939 PC6 0.084 0.191 -0.070 -0.051 0.108 0.171 -0.478 0.173 -0.113 -0.488 0.391 0.378 -0.321 0.017 0.956 PC7 -0.029 0.101 0.201 -0.261 0.034 0.217 -0.248 0.111 0.476 0.346 -0.258 0.540 0.233 0.012 0.967 PC8 0.437 0.120 0.106 0.148 0.425 0.129 0.578 -0.241 0.237 -0.230 0.069 0.243 -0.005 GOOD VS WEAK Table 17 Correlation matrix Good Vs Weak Principal Component Analysis: Eigenanalysis of the Correlation Matrix Eigenvalue Proportion Cumulative 5.2975 0.407 0.407 4.0625 0.313 0.720 1.1362 0.087 0.807 0.6540 0.050 0.858 0.5932 0.046 0.903 Eigenvalue Proportion Cumulative 0.0988 0.008 0.986 0.0925 0.007 0.993 0.0421 0.003 0.996 0.0287 0.002 0.999 0.0172 0.001 1.000 PC1 PC2 PC3 PC4 PC5 Variable 0.4589 0.035 0.939 0.3741 0.029 0.967 0.1443 0.011 0.979 PC6 PC7 PC8 228 Quality Time Batch seg 12-20 seg 20-25 seg 25-30 seg 30-35 seg 35-40 seg 40-45 seg 45-50 seg 50-55 seg 55-60 seg 60-65 0.000 0.383 0.238 0.339 0.390 -0.182 -0.374 0.295 0.293 -0.210 -0.312 -0.198 0.061 0.472 0.006 0.006 0.073 0.056 -0.466 -0.072 -0.386 -0.072 -0.080 -0.191 0.389 -0.445 0.339 -0.182 0.432 0.031 0.066 0.150 0.075 0.219 -0.389 -0.517 0.152 0.225 0.304 0.358 0.073 -0.313 0.289 -0.182 0.339 -0.052 0.062 0.192 0.247 -0.303 0.411 0.414 0.082 0.296 -0.103 -0.613 -0.049 0.224 -0.517 -0.052 -0.348 -0.045 -0.265 -0.051 0.010 Variable Treatment Time Batch seg 12-20 seg 20-25 seg 25-30 seg 30-35 seg 35-40 seg 40-45 seg 45-50 seg 50-55 seg 55-60 seg 60-65 PC9 -0.042 -0.417 0.181 0.383 0.047 0.271 0.251 -0.158 -0.436 0.370 0.250 0.267 -0.143 PC10 0.167 0.120 0.247 0.080 -0.139 -0.162 -0.576 0.153 -0.146 0.357 0.064 -0.306 -0.493 PC11 -0.046 -0.081 0.598 -0.039 0.168 -0.603 0.197 -0.099 -0.268 -0.295 0.030 -0.146 0.106 PC12 -0.339 0.668 0.144 0.007 -0.070 -0.165 0.064 -0.082 0.061 0.209 0.214 0.526 -0.071 PC13 0.552 0.267 -0.027 0.057 -0.036 -0.066 0.417 -0.028 0.155 0.326 0.384 -0.330 0.229 0.022 0.108 -0.443 0.217 -0.138 0.005 -0.441 0.053 0.060 -0.322 0.636 0.077 -0.100 -0.080 0.032 0.542 0.124 -0.213 0.501 -0.227 -0.427 0.298 0.000 0.088 0.070 -0.226 0.263 -0.320 -0.015 -0.539 0.166 -0.006 -0.028 0.175 0.672 -0.120 0.088 0.067 0.000