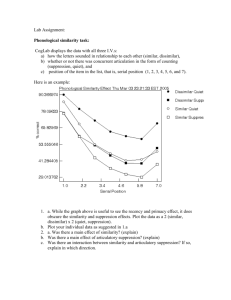
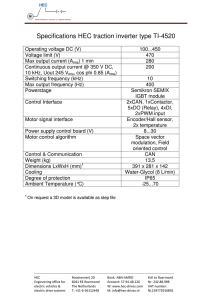
Document 11263827
advertisement

Investigating the Suppressive Effects of a Low-frequency Bias-tone on Auditory-nerve Responses to Tail-frequency Tones SACHU E INSTITUTE IT OF TECOLO~y By SEP 27 2011 Hui S. Nam B.S. Electrical Science and Engineering Massachusetts Institute of Technology, 1994 L R I SUBMITTED TO THE DEPARTMENT OF ELECTRICAL ENGINEERING AND COMPUTER SCIENCE IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE IN ELECTRICAL ENGINEERING AND COMPUTER SCIENCE AT THE MASSACHUSETTS INSTITUTE OF TECHOLOGY ARCHIVES SEPTEMBER, 2011 @ 2011 Massachusetts Institute of Technology. All rights reserved Signature of Author Harvard-MIT Division of Healt Sciences and Technology September 2, 2011 Certified by John J. Guinan, Jr., Ph.D Professor of Otology and Laryngology, Harvard Medical School Thesis Supervisor Accepted by Leslie A.Kolodziejski, Ph.D Professor of Electrical Engineering and Computer Science Chair, Committee for Graduate Students Investigating the Suppressive Effects of a Low-frequency Bias-tone on Auditory-nerve Responses to Tail-frequency Tones By Hui S. Nam Submitted to the Department of Electrical Engineering and Computer Science on September 2, 2011 in Partial Fulfillment of the Requirements for the Degree of Master of Science in Electrical Engineering and Computer Science ABSTRACT Sounds are amplified and frequency analyzed in the cochlea and the resulting signals are sent to the brain through the auditory nerve (AN). AN fiber threshold vs. frequency-response characteristics are band-pass shaped, with a sensitive, sharply-tuned tip centered at the "characteristic frequency" (CF) of the fiber. It is now well established that the sharp frequency tuning and high sensitivity of the tip is produced by mechanical feedback amplification from cochlear outer hair cells (OHCs). Specifically, sound vibrates the cochlear basilar membrane and this motion is transmitted to the OHC mechano-electric transduction function which produces a voltage change in the OHCs. This voltage change, through OHC electromotility, feeds back energy that amplifies basilar-membrane motion. For AN fibers with CF > 5 kHz (high-CF fibers), the sharp tuning-curve tip region is flanked on its low-frequency side with a broadly-tuned, less-sensitive "tailfrequency" region where the role of OHC motility has been thought as not significant. Contrary to this prevailing view, it has been experimentally found that the electromotility of OHCs is involved in generating AN responses in a narrow band of frequencies near 2.5 kHz in the tail. The main objective of this thesis was to investigate whether and how the mechano-electric transduction function of OHCs is involved in generating AN responses to tail-frequency tone at 2.5 kHz, and to determine any differences in the details of the mechano-electric transduction function in the tail-frequency responses versus in low-level CF-tone responses. The experimental strategy was to use the suppressive effects of a low-frequency bias-tone on AN responses driven by the active motility of OHCs. AN responses are affected by the mechano-electric transduction function of OHCs such that AN responses to low-level CF-tones are suppressed twice per bias-tone period by high-level bias-tones. This suppression is due to the saturation of the mechano-electric transduction as the bias-tone moves the operating point of OHC mechano-electric transduction into the nonlinear saturation regions twice per period of the bias-tone. Specifically, a bias-tone at 50 Hz was applied together with a second probe-tone, which was either a low-level CF-tone or a nearthreshold tail-frequency tone at 2.5 kHz, in order to characterize and compare the suppressive effects on these two response types. Single AN fiber recordings were made from 27 fibers from 6 cats. As expected, the characteristic pattern of twice-per-bias-tone-period suppression was found from the low-level CF- tone responses from all of the recorded fibers. As for the tail-frequency tone responses, significant suppression was found on 10 of the 27 fibers recorded. Among those 10 fibers, the twice-per-biastone-period suppression pattern associated with the non-linearity of the mechano-electric transduction function of OHCs was found for fibers with CF < 15 kHz. The lack of suppression for fibers with CF > 15 kHz may be due to the bias-tone sound levels not being high enough. These results directly show that the mechano-electric transduction function of OHCs is involved in generation of AN responses to 2.5 kHz tones for fibers with CF < 15 kHz in a similar way to how it is involved in producing cochlear amplification for low-level CF-tones. Further, comparisons of the details of the suppression pattern, e.g., most importantly the phase of major suppression, did not reveal any significant differences between the two response types. Overall these results indicate the detailed mechanisms of the OHC mechano-electric transduction function that are involved in producing the two responses types are similar. Additionally, the effects of a low-frequency bias-tone on the phase of AN responses to tailfrequency tones were compared with previous work on the effects of medial olivocochlear (MOC) efferent stimulation on the phase of AN responses to tail-frequency tones. Suppression of the gain of the OHC mechano-electric transduction function by a low-frequency bias-tone may affect the phase as well as the rate of AN responses driven by OHCs; however, effects of a low-frequency bias-tone on the phase of AN responses have not been reported in the literature. Bias-tone induced phase shifts were quantified as the difference in the phase of excitation to 2.5 kHz tones between bias-tone levels below and above the suppression threshold. Results were collected from five fibers. The shift in the phase of AN responses to 2.5 kHz tail-frequency tones induced by a bias-tone ranged from -45* (phase lag) to +5* (phase lead). A phase lag was found from 4 of the 5 fibers. These results are in general agreement with the MOC efferent effects on the phase of AN responses which ranged from -80* to +60* with an average of -15*, a phase lag. These results suggest that a lowfrequency bias-tone and MOC efferent stimulation affect the phase of AN responses to a tail-frequency tone through a similar mechanism by lowering the gain of the OHCs. Thesis Supervisor: John J. Guinan, Jr., Ph.D Title: Professor of Otology and Laryngology, Harvard Medical School Table of Contents List o f Figu re s ................................................................................................................................................ Introduction and Background .......................................................................................................-----... . 6 8 A. AN response characteristics of high-CF fibers: tip VS tail frequencies ........................................ 8 B. OHC involvement in the generation of AN responses to tail frequencies .................................... 9 C. Non-linearity in the OHC transduction function and suppressive effects on AN responses induced 11 by a low-frequency bias-tone ................................................................................................................. Thesis Objectives............................................................................................................................. D. Experimental M ethods ....................................................................................................................... 1. 13 15 A. Surgical preparation and acoustic setup..................................................................................... 15 B. Data Collection ................................................................................................................................ 15 C. Bias-tone only Paradigm: Excitation Threshold and Phase.......................................................... 16 1. Acoustic Stimulus and Data Collection .................................................................................... 16 2. Data Analysis............................................................................................................................... 16 Experimental Paradigm for Bias-tone Effects on Low-level CF-tone Responses......................... 18 D. 1. Acoustic Stimulus and Data Collection .................................................................................... 18 2. Analysis of the Bias-tone Period Histograms for Suppression Effects .................................... 21 E. Experimental Paradigm for Bias-tone Effects on AN Responses to Near-threshold-level Tailfrequency Tones...................................................................................................................................... 24 1. Acoustic Stimulus and Data Collection .................................................................................... 24 2. Analysis of the Bias-tone Period Histograms for Suppression Effects .................................... 24 3. Analysis of the Bias-tone Effects on the Phase of AN Responses to the Tail Frequency Tone at 26 2 .5 kHz................................................................................................................................................. Re su lts ............................................................................................................................................. 29 A. Exam ple Result from CT023_U066: CF=7.16 kHz, 80 sps............................................................. 30 B. Exam ple Result from CT031_U078: CF=7.37 kHz, SR=13.2 sps.................................................... 35 C. Exam ple Result from CT026_U009: CF=12.8 kHz, SR=1.7 sps...................................................... 41 D. Exam ple Result from CT026_UO41: CF=13.43 kHz, SR=78.7 sps.................................................. 44 E. Exam ple Result from CT028_UO44: CF=19.37 kHz, SR=0.6 sps.................................................... 48 F. Phase of Excitation by 50 Hz Bias-tone Alone and Phase of Suppression on Low-level CF-tone Re sp o n se s ............................................................................................................................................... 54 G. Comparison of Bias-tone Effects on AN Responses to Low-level CF-tone Responses VS Nearthreshold-level Tail-frequency Tone at 2.5 kHz .................................................................................. 55 Ill. H. IV. V. Bias-tone Effects on the Phase of AN Responses to Tail-frequency tone at 2.5 kHz..................58 Discu ssio n........................................................................................................................................ 60 A. Concerns for Potential Artifacts Caused by Direct Stimulation of IHC Stereocilia by the Bias-tone 60 B. Bias-tone Induced Suppression Patterns on Tail-frequency Tone Responses............................60 C. Bias-tone Induced Phase Shift in Tail-frequency Tone Responses ............................................. Re fe re nce s .......................................................................................................................................... 61 63 List of Figures Fig. 1 The effects of MOC stimulation on rate-level curves of a high-CF (CF=17.78 kHz) fiber in response to tones at tail frequencies and CF, with and without MOC efferent stimulation.....................10 Fig. 2 Effects of MOC efferent stimulation on AN responses to tail frequency tone at 1 kHz. ............. 11 12 Fig. 3 A model of the mechano-electric transduction function of OHCs. .............................................. of a fiber a high-CF from Fig. 4 Suppressive effects of 200 Hz bias-tone on low-level CF tone responses 13 ca t .................................................................................................................................................. Fig. 5 Results of the bias-tone alone paradigm from the example fiber, CT026_U050, CF=7.19 kHz, 17 SR=3 .6 sps. ..................................................................................................................................... Paradigm...........................................19 Burst on Tone Bias-tone Fig. 6 Acoustic Stimulus Setup for the Fig. 7 Threshold tuning Curve of the example fiber is plotted together with the CF-tone and tailfrequency tones applied on this fiber......................................................................................... 20 Fig. 8 Bias-tone level series of the period histograms from the bias on low-level CF-tone paradigm on 21 the exam ple fiber........................................................................................................................... Fig. 9 The detailed data analysis on the bias-tone level functions on the suppression effects on CF-tone 23 responses from the exam ple fiber............................................................................................. Fig. 10 Bias-tone effects on a level series of 2.5 kHz tail-frequency tone from the example fiber........... 25 Fig. 11 Effects of a bias-tone level series on AN responses to the tail-frequency tone at 70 dB SPL........26 Fig. 12 Bias-tone effects on the synchrony of AN responses to the tail-frequency tone at 2.5 kHz for the 27 exam ple fiber. ................................................................................................................................ 29 Fig. 13 AN fiber count over the three octave bands of CF from 5 kHz to 20 kHz .................................. 30 the fiber............................................. Fig. 14 CT023_U066: CF=7.16 kHz, SR=80 sps. Threshold TC of 31 Fig. 15 CT023_U066: Bias-tone level series of the bias-tone period histograms .................................. Fig. 16 CT023_U066: The detailed data analysis on the bias-tone level functions on the suppression 32 effects on CF-tone responses ........................................................................................................ Fig. 17 CT023_U066: Suppression effects of 50 Hz bias-tone at the fixed level of 90 dB SPL on a level 33 series of tail-frequency tone at 2.5 kHz ...................................................................................... AN responses of level series on probe-tone bias-tone of 50 Hz Fig. 18 CT023 U066: Suppressive effects 34 to 1 kH z to ne.................................................................................................................................. Fig. 19 CTO31 U078: CF=7.37 kHz, SR=13.2 sps. Threshold TC of the fiber........................................... 36 Fig. 20 CT031_U078: Bias-tone level series of period histograms.......................................................... 37 Fig. 21 CT031_U078: The detailed data analysis on the bias-tone level functions on the suppression 38 effects on CF-tone responses ........................................................................................................ 39 SPL .......... at 70 dB tone kHz tail-frequency Fig. 22 CT031_U078: Bias-tone effects on responses to 2.5 Fig. 23 CT031_U078: Bias-tone effects on the phase of AN responses to near-threshold tail-frequency 40 to ne ................................................................................................................................................ 41 Fig. 24 CT026_U009: CF=12.8 kHz, SR=1.7 sps. Threshold TC of the fiber ............................................ 42 Fig. 25 CT026_U009, Bias on CF-tone Responses .................................................................................. Fig. 26 CT026_U009, Bias on 2.5 kHz Tail-frequency Responses............................................................ 43 Fig. 27 CT026_U041: CF=13.43 kHz, SR=78.7 sps. Threshold TC of the fiber is plotted together with the 44 CF-tone and tail-frequency tones applied on this fiber ....................................................... Fig. 28 CT026_UO41, CF=13.43 kHz: Bias-tone effects on low-level CF-tone responses. ..................... 45 Fig. 29 CT026_UO41, CF=13.43 kHz: Bias-tone effects on AN responses to 2.5 kHz tone at 80 dB SPL.46 Fig. 30 CT026_UO41, CF=13.43 kHz: Bias-tone effects on the phase of AN responses to 2.5 kHz tone at 8 0 d B SPL........................................................................................................................................ 47 Fig. 31 CT028_UO44, CF=19.37 kHz, SR=0.6 sps: Threshold TC of the fiber ........................................... 48 Fig. 32 CT028, U044: CF=19.37 kHz. Bias-tone level series of bias-tone period histograms ................ 49 Fig. 33 CT028_U44: The detailed data analysis on the bias-tone level functions of the suppression effects on CF-tone responses ........................................................................................................ 50 Fig. 34 CT028_U44: Bias-tone effects on AN responses to 2.5 kHz tail-frequency tone at 90 dB SPL ...... 51 Fig. 35 CT028_UO44: Bias-tone effects on the phase of AN responses to 2.5 kHz tail-frequency tone at 90 d B S PL............................................................................................................................................. 52 Fig. 36 Phase of excitation by the "bias-tone alone" and the phase of major suppression on low-level CFto ne re spo nses...............................................................................................................................5 5 Fig. 37 Major suppression phase of AN responses to low-level CF-tones and near-threshold level tailfrequency to ne at 2.5 kH z ............................................................................................................. 56 Fig. 38 Measure of symmetry between the major and minor suppression phase for bias-tone effects on AN responses to low-level CF-tones and tail-frequency tone at 2.5 kHz .................................. 57 Fig. 39 Threshold of suppression of AN responses to low-level CF-tones and tail-frequency tones at 2.5 kHz..................................................................................................................................................5 8 Fig. 40 Shift in the phase of AN responses to tail-frequency tone at 2.5 kHz induced by the bias-tone.. 59 I. Introduction and Background Key aspects of the operation of the mammalian cochlea are still poorly understood. With advances in basilar membrane (BM) measurement techniques [Ruggero et al, 1996], it has been found that the tuning and sensitivity in the tip region of auditory-nerve (AN) fibers with high characteristic frequencies (CFs) matches well with the tuning of BM motion [Narayan et al, 1998]. These results have led to the currently prevailing view on the mechanism behind sound signal transmission to inner hair cells (IHCs), and through the IHCs, to AN fibers: (1) sound travels through a single transmission pathway, the BM traveling wave, to drive IHC stereocilia and thereby to excite AN fibers [Ruggero et al, 1996], (2) the drive to the IHCs can be described as a sensitive tip enhanced by an OHC-based feedback amplifier, called the "cochlear amplifier", that has compressive non-linearity, and outside of this sensitive CF region, the drive to the IHCs is mainly passive [Ruggero et al, 1997], and (3) BM motion measurements at the base of the cochlea can explain the auditory nerve firing pattern throughout the cochlea [Narayan et al, 1998]. Alongside this view, however, there is an abundance of experimental results on AN firing patterns both at near threshold and at high sound levels, and also for both low-CF and high-CF fibers, that cannot be adequately explained by this view. The objective of this thesis is further investigation on one such unexpected finding in responses of high-CF AN fibers. Specifically, in conflict with the prevailing view on cochlear mechanics, OHCs have been found to be actively involved in generating AN responses to tone stimuli from one frequency region of the high-threshold, low-frequency "tail region" [Stankovic and Guinan, 1999]. A. AN response characteristics of high-CF fibers: tip VS tail frequencies The neural-threshold tuning curve of a cat high-CF AN fiber with CF= 17.78 kHz is shown in the upper-left panel of Fig. 1. In this figure the sensitive, sharply-tuned tip region around CF can be readily distinguished from the broadly-tuned, low-frequency "tail region" which has a much higher threshold than the tip by 60 - 70 dB. Studies have shown that damage to OHCs produced by acoustic trauma [Liberman and Kiang, 1978] affect AN responses in tip vs. tail regions drastically differently. Specifically, damage to the OHCs has been found to raise neural response thresholds from the sensitive tip region significantly, while the sensitivity of neural responses in the tail region remain relatively unaffected or even increased [Liberman and Dodds, 1984]. It is now widely accepted that active motility in OHCs is the underlying mechanism behind the highsensitivity and fine-tuning of AN responses to tip-region, CF-tone stimuli and their vulnerability to damage, particularly damage to OHCs [Robles and Ruggero, 2001]. OHC active motility is thought to enhance the sensitivity and frequency tuning of BM motion and AN firing in response to tone stimuli near CF through a mechanism termed the "cochlear amplifier". The critical part of the "cochlear amplifier" lies in the mechanical feedback between the BM and OHCs whereby the stereocilia of OHCs transduce the BM vibration to OHC transmembrane receptor voltage which, through OHC electromotility, returns as changes to BM motion [Ruggero, 2001]. B. OHC involvement in the generation of AN responses to tail frequencies Measurements of BM motion from the basal, high-CF region of chinchilla and guinea pig cochleae show that for low-frequency tones that are an octave or more below the CF of the measurement site, the BM motion response grows linearly with stimulus level and is largely unaffected by acoustic trauma [Ruggero et al, 1997]. These results are distinctly different from the compressive growth and sensitivity to acoustic trauma found in BM responses to CF tones [Ruggero et al, 1997]. Under the prevailing view of cochlear mechanics, these distinct characteristics of CF versus tail-frequency responses are explained by there being a large contribution from the OHC cochlear amplifier in CF-tone responses and a negligible contribution from the OHC cochlear amplifier in tail-frequency-tone responses. Therefore, if BM motion with these characteristics is the only drive to the IHCs produced by a tail-frequency tone, as assumed in the prevailing view of cochlear mechanics, electrical stimulation of medial olivocochlear (MOC) efferents, which is known to inhibit AN firings through the inhibitive synapses on OHCs [Guinan, 1996], should have a negligible impact on AN responses to tail-frequency tones. This topic was extensively investigated in [Stankovic & Guinan, 1999] which measured the effects of MOC stimulation on rate-level functions of cat high-CF AN fibers in response to tail-frequency tones and to low-level CF tones. As expected, MOC stimulation strongly inhibited AN responses to low-level CF tones. However, in contrast to the predictions from the prevailing view of the cochlear mechanics, Stankovic and Guinan found a MOC-inhibited region between 1.5 and 4 kHz in the tail of high-CF fibers. Fig. 1 shows the effects of MOC stimulation on the rate-level curves in response to a CF tone and to the tail-frequency tones at 1 kHz, 2 kHz, 2.5 kHz and 3 kHz. An average of the MOC effects over the tail-frequency region from 500 Hz to 5 kHz for high-CF fibers with CFs between 10 and 30 kHz has shown that the MOC inhibitory effects are most prominent at a tail frequency of 2.5 kHz [Stankovic & Guinan, 1999]. These results strongly suggest that AN responses to tones from this MOC-sensitive region are generated by an OHC-based active drive. Further, since damage to OHCs does not appreciably affect BM responses to tail-frequency tones [Ruggero et al, 1997], it has been hypothesized that the IHC drive that produces the AN responses to tail-frequency tones is likely to involve a mechanism that must be substantially different from the BM-based cochlear amplifier [Stankovic & Guinan, 1999]. In addition to the MOC efferent effects on the rate-level function of AN responses to tail-frequency tones, the effects of MOC efferents on the synchrony and phase of AN responses to tail frequency tones were investigated in [Stankovic and Guinan, 2000]. This study has found that in the absence of MOC stimulation, the phase of AN responses to tail frequency tones exhibit increasing delay with increase in tone stimulus intensity by as much as 90* over the tone stimulus range of 30-40 dB from the threshold level. Further, it has been found that stimulation of MOC efferents introduces on average 15* of phase lag with the largest phase shift at tone levels near threshold. A typical example for this finding is shown in the left panel of Fig. 2 where the period histograms of AN responses to a level series of tail-tone at 1 kHz with and without MOC efferent stimulation are plotted together [Stankovic and Guinan, 2000]. In the right panel of Fig. 2, the effects of MOC efferent stimulation on phase response of AN fibers to 1 kHz tone level series are compared with the changes in the phase of 1 kHz component of the cochlear microphonic (CM) [Stankovic and Guinan, 2000]. Since In the base of the cochlea, the phase of CM in response to a low frequency tone such as 1 kHz follows the phase of BM displacement [Dallos et al., 1974], comparison of the MOC efferent effects on the phase of AN responses to tail frequency tones VS the phase of CM responses allows a test to determine whether the mechanism of changes in the phase of AN responses by MOC efferent stimulation involves changes in the motion of the BM [Stankovic and Guinan, 2000]. As shown in the right panel of Fig. 2, the changes in the phase of CM responses were substantially less than those of AN responses [Stankovic and Guinan, 20001. These results were interpreted as further support for the hypothesis that the OHC motility acts on generating AN responses of high-CF fibers to tail frequency through a mechanism which does not involve BM motion thereby substantially different from the cochlear amplifier [Stankovic and Guinan, 20001. * with efferent stimulation o without efferent stimulation 0- 75 300- 300 50-. f 300- 1 kHz f 2.0 kHz C, 25 4 0 " 10 1 Frequency (kHz) o ) 0) i 0 10 40 __ C 300 300 f= 2.5 kHz 0 100 f =3.0 kHz 100 ' 40 100 300- C/) CU f= CF (17.78 kHz) 0 40 100 0 40 100 0 0 80 Sound Level (dB SPL) Fig. 1 The effects of MOC stimulation on rate-level curves of a high-CF (CF=17.78 kHz) fiber in response to tones at tail frequencies and CF, with and without MOC efferent stimulation. Note the prominent MOC effects on CF responses and at the narrow region around 2.5 kHz. Taken from [Stankovic & Guinan, 1999] O-o No efferent stimulation ---- -o With efferent stimulation 500A 500 S z 82 dB B 76 dB C 0 0 W 500 0 U)- CM 60 30 .0 Cn 0 500 ANF 7 w -30 -60 50 0 6-90 0.5 . o no efferent stimulation 9 efferent stimulation 1 1.5 TIME (ms) (Left) 2 -120 50 60 70 80 90 SOUND LEVEL (dB SPL) 100 (Right) Fig. 2 Effects of MOC efferent stimulation on AN responses to tail frequency tone at 1 kHz. Taken from FIG 2 and FIG 13 of [Stankovic and Guinan, 2000]. (Left) Level series of period histograms of AN responses to 1 kHz tone over the range of 64 dB SPL to 88 dB SPL with and without MOC efferent stimulation. Near threshold, MOC efferent stimulation introduced significant phase delay which diminished at higher levels of the tone stimulus; (Right) Comparison of the bias-tone effects on the phase response of high-CF AN fibers VS cochlear microphonic (CM) to level series of 1 kHz tail-frequency tone. Note that the effects on AN response phase were substantially larger than on CM. C. Non-linearity in the OHC transduction function and suppressive effects on AN responses induced by a low-frequency bias-tone Stimulation of MOC efferents results in a reduction of the gain of OHC somatic electromotility, i.e., the electromechanical or "reverse" transduction of OHCs, thereby inhibiting AN responses. Alternatively, AN responses that are actively amplified by OHCs can also be suppressed by affecting OHC forward transduction, i.e., the mechanoelectric transduction process of OHCs. This suppression involves the saturating non-linearity of the OHC mechanoelectric transduction function [Robles and Ruggero, 20011. In vitro and in vivo measurements of OHC mechanoelectric transduction functions show saturating plateaus at both ends of the output range, as shown in Fig. 3 which displays a schematized version of a typical shape of the mechanoelectric transduction function of OHCs from the basal part of the cochlea. It is thought that the gain of cochlear amplification depends on the slope of the mechanoelectric transduction function. At rest, the operating point of the transduction function lies at a high-gain region with a high slope in the transduction curve and well away from the saturation ends. A low frequency bias-tone tone in the range of 30 - 100 Hz with high enough amplitude, but at an amplitude still under the neural response threshold, can (at certain phases of the bias-tone) deflect the OHC stereocilia wide enough to move the OHC mechanoelectric operating point into the low gain region. Consequently, AN responses to a low-level CF probe tone stimulus presented together with such a bias-tone would be suppressed in synchrony with bias-tone movement of the OHC stereocilia into their saturation regions [Ruggero et al, 1992]. Specifically, as the level of the bias-tone is increased, the suppression pattern on AN firing in response to a low-level, cochlear-amplified, CF-tone typically progresses from a single suppression phase (called the "major suppression phase"), to two suppression phases that are separated by half a period of the bias-tone [Cai and Geisler, 1996a]. An example of the suppressive effects of a low-frequency bias-tone on an AN response is shown in Fig. 4 in which a bias-tone of 200 Hz and a low-level CF-tone were presented together and responses were measured from a cat high-CF fiber with a CF of 12.82 kHz [Cai and Geisler, 1996a]. Note that a single suppression phase, the major suppression phase, appeared at the bias-tone level of 85 dB SPL in the left plot of Fig. 4. Then, at 90 dB SPL of the bias-tone, the suppression pattern developed into twoper-cycle suppression with the suppression phases separated by a half a cycle. Typical Shape of OHC Mechanoelectric Transduction Function from Base of the Cochlea C: CU cu CU 0.0 0 Sound Pressure, arbitrary unit Fig. 3 A model of the mechano-electric transduction function of OHCs with a saturating non-linearity at both ends of the voltage output. Positive sound pressure indicates rarefaction. The gain of the cochlear amplifier has been linked to the slope of the transduction function at a particular operating point. The operating point "at rest" is indicated by a red dot in the above plot. The operating point has been found to lie at varying degrees of asymmetry between the two saturating ends. This particular plot shows the typical symmetric shape found in the basal region of the cochlea. SUP Intensity: 85 dB 100 90 dB W7W 12820Hz 30 so e -UdS li ilhk1m 0.5 1 0.5 1 Bias Tone Period Fig. 4 Suppressive effects of 200 Hz bias-tone on low-level CF tone responses from a high-CF fiber of a cat. The CF of the fiber was 12.82 kHz, and the bias-tone frequency was 200 Hz. The level of the bias-tone corresponding to each of the two period histograms isnoted as 'SUP Intensity' in each plot. Taken from Figure 10 of [Cai and Geisler, 1996a] There are two objectives of measuring the suppression of AN responses produced by a lowfrequency bias-tone: (1) the suppression pattern can reveal whether the mechanoelectric transduction function of OHCs is involved in AN response generation, and (2) the suppression pattern can provide useful data on the micromechanics of AN response generation within the Organ of Corti. Studies have shown that the phase of AN response suppression due to a low-frequency bias-tone can be related to the phase of BM displacement. Specifically, in guinea pigs the major suppression phase has been found to correspond to the phase of peak BM displacement toward scala tympani (ST) [Patuzzi et al, 1984a]. Further, in the apical part of the cochlea, the asymmetry of the depth of the two suppression phases has been used to show that the operating point of the OHC mechanoelectric transduction function is located asymmetrically, i.e. the operating point is closer to the saturation plateau in hyperpolarizing direction than to the plateau in the depolarizing direction [Cai and Geisler, 1996c]. The aforementioned properties of suppressive effects of a low-frequency bias-tone on the mechano-electric function of OHCs and the AN responses driven by it can serve as an investigative tool to determine whether and how the mechano-electric transduction function of OHCs are involved in generation of AN responses to tail frequency tones. Note that the inhibitory effects of MOC stimulation on AN responses to tail-frequency tones in [Stankovic and Guinan, 1999] demonstrated that the OHC reverse transduction function, i.e., OHC electromotility, is involved in producing AN responses to tailfrequency tones while the involvement of the forward transduction, i.e., the mechanoelectric transduction function, of OHCs has not been directly shown. D. Thesis Objectives The main objective of this thesis is to determine whether and how the mechano-electric transduction function of OHCs is involved in generating AN responses to tail-frequency tones from highCF fibers of cats. Specifically, if two oppositely located suppression phases were to be found from the bias-tone effects on AN responses to tail frequency tones, it would indicate involvement of the mechano-electric transduction function of OHCs in the mechanism producing AN responses to tail frequency tones. Further, if the mechano-electric transduction function of OHCs is indeed involved in generation of AN responses to tail frequency tones, comparison of the major suppression phase and the symmetry of depth of suppression phases of the AN responses to tail frequency tones VS low-level CFtones can reveal any differences in the shape or the operating point at rest of the OHC mechano-electric transduction function producing these AN response types. Investigation is focused on one particular tail-frequency of 2.5 kHz where the inhibitive effects of MOC efferent stimulation were found to be the strongest [Stankovic and Guinan, 1999]. Additionally, the effects of a low-frequency bias-tone on the phase of AN responses to tailfrequency tones are compared with the effects of MOC efferent stimulation on the phase of AN responses to tail-frequency tones reported in [Stankovic and Guinan, 2000]. The objective of this comparison is largely exploratory. Suppression of the gain of the mechano-electric transduction function of OHCs by a low-frequency bias-tone is expected to affect the phase as well as the rate of AN responses driven by OHCs; however, the effects of a low-frequency bias-tone on the phase of AN responses to tones at any frequency from either high or low-CF fibers have not been reported in the literature. II. Experimental Methods A. Surgical preparation and acoustic setup The experiments were carried out on cats which typically survive through 48 - 60 hours of experimentation. The experimental methods for animal handling, surgical approach to the AN and methods for acoustic setup and recording from AN fibers of cats were as described in [Kiang et al, 1965] and [Stankovic & Guinan, 1999], and the methods for monitoring the health of hearing during experiments are as described in [Stankovic & Guinan, 1999]. Briefly, all experiments were done inside a double-walled, reduced-reflection chamber. Sound stimuli were delivered with an acoustic assembly consisting of two earphones and a microphone with a probe tube which was calibrated for the magnitude and phase of the frequency response of the entire assembly including the probe tube. A DT48 dynamic earphone, capable of high output at low frequency, was used to generate the lowfrequency bias-tone, and all other stimulus types were generated by a 1-inch condenser earphone [Bruel & Kjaer (B&K) 4145]. Throughout all experiments, the frequency of the bias-tone was fixed at 50 Hz since a low frequency tone below 200-600 Hz can more selectively deflect the OHC stereocilia compared to the IHC stereocilia [Russell and Sellick, 1983] B. Data Collection A silver electrode was inserted through the bulla opening and placed near the round window to measure the cochlear compound action potentials (CAP). An automated tone-pip audiogram was measured over the frequency range from 0.5 kHz to 32 kHz at an octave interval with the criterion of 10 pV pp of the CAP. Tone pip audiograms were run periodically every hour or so, and also after a series of good data in order to ensure good health of the cochlea associated with the collected data. A glass micropipette electrode filled with 3 M KCL electrolyte with 10 -20 MO impedance was driven through the exposed view of the auditory nerve by a remotely controlled microdrive until isolating an AN fiber using a wideband noise burst search stimulus. Spike timings were detected with the resolution of 10 plsec. Upon isolating a unit using a wideband, noise-burst search stimulus, a threshold tuning curve and the spontaneous firing rate of the fiber were measured. Upon identifying a high-CF unit with its CF in the range 5 to 30 kHz (the range- used in [Stankovic & Guinan, 1999]), the following 50 Hz bias-tone paradigms were run: e e e Bias-tone alone paradigm Bias-tone effects on low-level CF-tone responses Bias-tone effects on near-threshold tail-tone responses The methods for the above experimental paradigms are described in detail with actual data from the example fiber, CT023_U066, with its CF at 7.16 kHz and SR of 80 sps. Note that the experimental methods for the bias-tone alone and the bias-tone effects on CFtone responses are as described in [Nam, 2011], and briefly presented in this thesis for the convenience of the reader. The methods for the above experimental paradigms are described in detail with actual data from the example fiber, CT026_U050, with its CF at 7.19 kHz and SR of 3.6 sps. C. Bias-tone only Paradigm: Excitation Threshold and Phase The objective of this paradigm was to measure the neural excitation threshold, and phase of excitation, of the fiber produced by a level series of a 50 Hz bias-tone, presented alone. The thresholdexcitation level determined by this procedure was used during later data analysis of the bias-tone effects on CF-tone and tail-tones to select data that are not significantly distorted by the effects of the bias-tone acting directly on IHC stereocilia. Further, the phase of excitation by the bias-tone alone provides a phase reference that can be related to the phase of BM displacement [Patuzzi et al, 1984a] which, in turn, can provide insights into the details of the cochlea mechanics involved in generating the AN response to tail-frequency tones. 1. Acoustic Stimulus and Data Collection A randomized level series of the 50 Hz bias-tone over the range of 70 dB SPL to 120 dB SPL was presented. A single trial period was 500 ms with 50% stimulus ON duty cycle, and at each level of the bias-tone, 8 trials were repeated. The spike records during the stimulus ON periods from the 8 trials with the total spike collection time of 1600 ms were binned re. the bias-tone period in order to form a level series of bias-tone period histograms. 2. Data Analysis Synchrony analysis was done on the bias-tone period histograms through the vector phase analysis method in order to calculate the phase of excitation and the synchrony index [Goldberg and Brown, 1969; Johnson, 1980]. Briefly, each spike is represented as a unit vector with a phase corresponding to the phase of the spike within the bias-tone period. The magnitude and phase of the average vector representation of all the spikes are the synchrony index and vector phase, respectively. Mathematically the average vector corresponds to the first harmonic component of the Fourier series of the period histogram normalized by the total spike count [Goldberg and Brown, 1969]. Further, the statistical significance of the phase estimates were evaluated through the standard error of the vector phase estimates. The mathematical model for calculating the standard error of the phase estimate, S.E@, is described in [Mardia, 2000], and has been previously applied in AN spike data analysis [Stankovic & Guinan, 1999b]. The excitation threshold was determined as the bias-tone level at which the standard error on the phase estimate, S.E.@, dropped below a critical value, e.g., ± 30, and also where the firing rate exceeded the baseline rate by one standard deviation of the baseline rate. The baseline rate was defined as the spike rate at the minimum level of the bias-tone level series. The results of the bias-tone alone paradigm from the example fiber are shown in Fig. 5 where the level series of the period histograms are plotted in Fig. 5(a), and the detailed analysis is shown in Fig. 5(b). Note that both the rate and phase error criteria were met at 110 dB SPL. (b) Bias-tone Only: Data Analysis (a) Bias-tone Only: Period Histograms Spike Rate BIAS TONE LEVEL dBSPL 100 Rate Level Function Rate Threshold: +y Cl) 50 120 () Excitation Phase U -A 0 70 110 E 1- 80 90 100 110 Tone Level, (dB SPL) 120 Synchrony Index VS Bias Tone Level C 100 c 05 CaC 0 U) i 70 90 [ 80 - - 90 100 110 120 Tone Level, (dB SPL) Excitation Phase Estimate - - 1 F1 80 0 -7 Excitation Phase F8 a- 0.5 Above Threshold U) 0.5 -C 0~ 0 5 5 PZC time (ms) 20 70 90 100 110 80 Tone Lee (dB SPL) 120 Fig. 5 Results of the bias-tone alone paradigm from the example fiber, CT026_U050, CF=7.19 kHz, SR=3.6 sps. (a)The bias-tone level series of the histograms are shown as vertically stacked plots with the corresponding bias-tone level indicated to the right of each histogram. The scale of the firing rate indicated for the bottom histogram applies to all the plots. Note that the "PZC time" in the x-axis label refers to the time after the positive peak voltage of the 50 Hz earphone drive signal; (b)Detailed data analysis metrics to determine biastone alone response threshold & response phase. Top: Rate-level function from which the rate-threshold is determined when the spike rate exceeds one standard deviation; Middle: Level function of the synchrony index; Bottom: the excitation phase estimates with their standard error. The excitation phase & its standard error are determined by the vector phase method. The excitation threshold is met when both the rate threshold is met and also when the standard error on the phase estimate drops within 30*. For the example fiber, the excitation criteria were met at 110 dB SPIL as indicated on the period histogram where the excitation phase ismarked with a green upward triangle. D. Experimental Paradigm for Bias-tone Effects on Low-level Responses CF-tone The objective of this paradigm was to determine and characterize the suppressive effects of a lowfrequency bias-tone on AN responses to low-level CF-tones. Specifically, primary interest was on detection of the major suppression phase from the period histograms and whether there was two-perbias-tone-cycle suppression. Additionally, the bias-tone level of suppression threshold for AN responses to CF-tones determined by this paradigm provided a reference level of the bias-tone for the experimental paradigm for bias-tone effects on near-threshold tail-tone responses as described in the next section. 1. Acoustic Stimulus and Data Collection A randomized level series of a 50 Hz bias-tone over the range 70 to 120 dB SPL was presented together with low-level CF-tone at 10 - 20 dB (for CF tones) above its TC threshold. The bias-tone was presented during the 50% ON duty cycle within the 500 ms trial period as shown in Fig. 6(a). The spike records within the spike analysis window with the duration of 220 ms spanning 11 cycles of the 50 Hz bias tone as shown in Fig. 6(b) are binned re. the bias-tone period in order to form a level series of bias-tone period histograms. The spike records from 24 of such trials were binned re. the bias-tone-period of 20 ms to form the bias-tone-period histogram. Note that spike records were initially time-stamped with the resolution of 10 ps which would yield bias-tone-period histograms with the sample size of 200000 if left unprocessed. Since spike data analysis for bias-tone effects on period histograms are concerned mainly with low-frequency modulation on the spike records spanning first few harmonics of the bias-tone, the spike records were re-binned with the time bin width of 0.5 ms or the sample size of 40. Note that the sample of 40 allowed spectral analysis up to the first 19 harmonics of the bias-tone as discrete Fourier transform was applied on the period histograms. The zero phase reference of the period histogram is the positive peak of 50 Hz electrical drive signal to the earphone. The threshold tuning curve and the level of the CF-tone for the example fiber is shown in Fig. 7. Fig. 8(b) shows the bias-tone level series of the period histograms for the bias-tone effects on AN responses to low-level CF-tone for the example fiber. Qualitatively, the major suppression phase seems to emerge at the bias-tone level of 80 dB SPL, and the suppression pattern moved to the typical twiceper-bias-tone period pattern at 90 dB SPL. Note that the period histograms from the bias-tone alone paradigm is replotted in Fig. 8(a) for comparison. It can be seen that the excitation pattern by the biastone alone began to be dominant at the excitation threshold of 110 dB SPL and above. (a) Stimulus Paradigm for Bias-tone on Probe-tone 50 Hz Bias Tone 24 trials Probe Tone 0 25 400 200 255 300 PST time (ms) (b) Example PSTH and the Spike Analysis Window 100 500 Spike Analysis Window: 25 ms - 245 ms 220 ms duration or 11 periods of the 50 Hz bias-tone F' I 100 300 200 245 PST time (ms) _f 400 500 Fig. 6 Acoustic Stimulus Setup for the Bias-tone on Tone Burst Paradigm: Top: the probe tone & the 50 Hz biastone were presented together for 230 ms within the total trial period of 500 ms. Bottom: The post-stimulus time histogram (PSTH) of the spikes over the entire 500 ms trial period. Note the ON duty cycle of ~50% corresponding to the stimulus setup diagram above. The spike analysis window covered the duration of 220 ms or 11 periods of the 50 Hz bias-tone. Tuning Curve of CT26, Unit 50: CF=7.19 kHz, Th=-0.6 dB SPL 120 - - + 0 100 800 TC CF-tone Tail 2.5kHz 0 80 1 I I 1 I 1 I I I I 1I I I I 600 It1 I I I U) I1 I II I I I Hi I I I I|I I I I I I III I 20 40. 20 I I I + I I I I I -20 1 IIIIIII I I 1 i i 101 100 Tone Frequency (kHz) Fig. 7 Threshold tuning Curve of the example fiber is plotted together with the CF-tone and tail-frequency tones applied on this fiber. (b) Bias on CF-tone (a) Bias-tone Only BIAS TONE LEVEL dB SPL 120 120 110 110 E C,, - 100 100 O. C) 90 90 V_ 80 80 0.2 )- 0 III L 1 5 1 , 10 PZC time (ms) 15 20 0 0 5 10 15 PZC time (ms) Fig. 8 Bias-tone level series of the period histograms from the bias on low-level CF-tone paradigm on the example fiber. (a) period histograms from the bias-tone alone paradigm isreplotted here for comparison; (b) period histograms from the bias on low-level CF-tone paradigm. It can be seen qualitatively that the major suppression phase emerged at 80 dB SPL. At the excitation threshold of 110 dB SPL and above, the bias on CFtone histograms were dominated by the bias alone excitation pattern. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after the positive peak voltage of the 50 Hz earphone drive signal. 2. Analysis of the Bias-tone Period Histograms for Suppression Effects Synchrony analysis was performed on the bias-tone period histograms to calculate the phase of suppression and the synchrony index [Goldberg and Brown, 1969; Johnson, 1980] based on the framework of directional data analysis [Mardia, 2000]. For increasing bias-tone levels, the typical progression of suppression patterns seen in AN responses to low-level CF-tones moves from a single suppression phase to two suppression phases separated by a half a period [Patuzzi et al, 1984a; Cai & Geisler, 1996a]. From the period histograms with a single suppression phase, estimation of the major suppression phase was done through a vector phase analysis of the first harmonic component of the period histogram using a similar approach as described for estimation of the excitation phase. Specifically, significant suppression was detected when the standard error of the phase estimate dropped below a critical value of ± 30 as done in [Stankovic and Guinan, 2000]. For the analysis of period histograms with two oppositely-located suppression phases, vector phawe analysis of the second-harmonic component of the period histograms was applied to detect the presence of two-per-cycle suppression phases and also to determine the major suppression phase. Similarly to the first harmonic method described above, the presence of a significant two-per-cycle suppression pattern will be detected based on a phase-estimate error criterion. Since the second harmonic phase corresponds to two suppression phase locations of the bias-tone period without distinguishing which is the major suppression phase, an analysis procedure is needed to disambiguate which corresponds to the major suppression phase. The basic approach for this is to calculate the modulation depth around the two suppression phases and to select as the major suppression phase the one with the higher modulation index. The details of this method, which is termed as "Half-period Synchrony Analysis", are described in [Nanm, 2011]. For period histograms which meet the suppression threshold for both the first and second harmonic component, the major suppression phase was determined from the second harmonic phase estimate. Finally, the period histograms generated by a bias-tone level at or above the bias-tone alone excitation threshold were classified as "Excitation" and accordingly excluded from the suppression data pool. The analysis procedures described above are illustrated on the example fiber as shown in Fig. 9 where the level functions of the spike rate, synchrony index and vector phase data for the first and second harmonic are plotted in the top-down order. In this example, the suppression threshold for the first and second harmonic criterion was met at the bias-tone level of 80 and 90 dB SPL respectively. The major suppression phase would be determined by the first harmonic phase at the bias-tone level of 80 dB SPL as noted by a blue inverted triangle in Fig. 8(b). Spike Rate: BT-on-ProbeTone & BT-alone Bias on CF Bias alone 100 a) .50 CD .0CO) 70 80 0) C 90 100 110 Bias Tone level, dB SPL Synchrony Index 120 0 0.5 C) -C 1st Harmonic 2nd Harmonic C) 70 0 80 90 100 110 Bias Tone level, dB SPL Suppression Phase Estimate Significant 1st Har Significant 2nd Har (D 0 c) 120 0.5 C CU 70 80 90 100 110 Bias Tone level, dB SPL 120 Fig. 9 The detailed data analysis on the bias-tone level functions of the suppression effects for CF-tone responses from the example fiber. Top: the rate-level function for the run with the bias-tone alone and for the bias on the probe tone run. Middle: bias-tone level functions of the synchrony index for 1" & 2 harmonics. Note the sharp increase of the synchrony indices at the suppression thresholds. Bottom: level function of the suppression phase estimates for 1st & 2nd harmonic with their standard error. The data points meeting the error bound are noted with markers labeled as "Significant 1s Harmonic" and "Significant 2nd Harmonic". The 1s & 2" harmonic suppression thresholds are met at 80 & 90 dB SPIL respectively. E. Experimental Paradigm for Bias-tone Effects on AN Responses to Nearthreshold-level Tail-frequency Tones The objective of this paradigm is to determine and characterize the suppressive effects of a lowfrequency bias-tone at 50 Hz on AN responses to near-threshold level tail-frequency tones. Focus of the investigation was on one particular tail frequency of 2.5 kHz where the inhibitive effects of MOC efferent stimulation were found to be the strongest [Stankovic and Guinan, 1999]. As with the bias-tone effects on low-level CF-tone responses, primary interest was on detection of the major suppression phase from the period histograms and whether there was two-per-bias-tone-cycle suppression. Additionally, the effects of the bias-tone on the phase of AN responses to the tail frequency tone were examined. 1. Acoustic Stimulus and Data Collection As reported in [Stankovic and Guinan, 1999], the range of the tail frequency tone levels inhibited by MOC efferent stimulation was relatively narrow within 10 - 20 dB of the AN response threshold to the tail frequency tone. Also, it has been found during the course of the experiment that selecting the tail frequency tone level from the tuning curve of the fiber as done in the paradigm for bias-tone effects on low-level CF-tone was not adequate. Accordingly, it was necessary to identify the level of the tail frequency first by running a paradigm where a level series of the tail frequency tone was presented together with a bias-tone at the reference level determined from the bias-tone effects on low-level CFtone responses. The range of levels for the tail-frequency tone was selected so that it traversed through the threshold tuning curve, and the reference level of the bias-tone was set typically at 10 dB above the suppression threshold on the low-level CF-tone response of the fiber. For the example fiber, the reference level of the bias-tone was at 100 dB SPL, and the range of levels of the 2.5 kHz tail-frequency is noted on the tuning curve in Fig. 7 2. Analysis of the Bias-tone Period Histograms for Suppression Effects The tail-frequency-tone level series of the period histograms were analyzed to determine the range of levels of the tail-frequency tone with significant suppression using the same methods as described in the paradigm for bias-tone effects on low-level CF-tone response. The bias-tone period histograms and the details of the data analysis for the example fiber are shown in Fig. 10. Significant suppression was found at 60, 70 and 80 dB SPL of the tail-frequency tone while the neural threshold for 2.5 kHz tone from the tuning was ~67 dB SPL. Accordingly, for the subsequent runs with a bias-tone level series with a fixed level of the tail-frequency tone, the tail-frequency level of 70 dB SPL was selected as marked by a filled blue circle on the tuning curve in Fig. 7. With the level of the tail-frequency tone as described above, a randomized level series of 50 Hz bias-tone was presented together with a tail frequency tone at the fixed level using the same methods as described in the paradigm for bias-tone effects on low-level CF-tone responses. The results of the bias-tone level series run on the example fiber are shown in Fig. 11. The suppression threshold for the first and second harmonic was met at 90 and 100 dB SPL of the bias-tone as indicated by the blue and red labels of the bias-tone level. The major suppression phase at these suppression thresholds are indicated by blue and red inverted triangles. At the second harmonic threshold of 100 dB SPL, the typical pattern of two suppression dips at opposite phases can be observed. The location of the major suppression phase was quite similar to that of the bias-tone effects on CF-tone from Fig. 8. IL, 1 I 2.5 kHz Tail-ton e Level Series: Bias at 100dB SPL Spike Rate: BT-on-ProbeTone & BT-alone Q 200 Tai i-tone Level dB SPL D 100 1~I Ca -- 0 100 60 0 'IL I 90 80 70 Probe Tone leveldB SPL 100 Synchrony Index 90 _0 C 1st Harmonic 2nd Harmonic 0 c0. II -C ci, 0 60 70 80 90 Probe Tone level dB SPL 100 Suppression Phase Estimate 0 70 70 0- Signficant 1 Significant 2 CU CO, I; 0 0 5 15 10 PZC time (ms) 20 60 C') CU, 60 80 90 70 Probe Tone leve dB SPL 100 Fig. 10 Bias-tone effects on a level series of 2.5 kHz tail-frequency tone from the example fiber. The bias-tone was at the fixed level of 100 dB SPL which was 10 dB above the suppression threshold on low-level CF-tone responses of the example fiber. Data analysis showed significant suppression at the tail frequency tone level of 60, 70 and 80 dB SPL while the neural threshold from the tuning curve was at ~67 dB SPL. For the subsequent bias-tone level series runs, the tail frequency tone level of 70 dB SPL was selected for this fiber. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after the positive peak voltage of the 50 Hz earphone drive signal Bias on 2.5 kHz Tail Tone at 70 dB SPL Bias-tone Spike Rate: BT-on-ProbeTone & BT-alone Level 80 dB SPL 120 1200 a - - -60 40 1-0 (D Bias on tail 0 0B- a) 70 110 80 90 100 Bias alone 110 120 Bias Tone leveIdB SPL Synchrony Index E1 1st Harmonic - 2nd Harmonic -100 2 .5 CnC U) 0 90 70 90 100 110 120 80 Bias Tone level dB SPL Suppression Phase Estimate .0 80 Significant 1st Har Significant 2nd Har 1D CL C 0 0.5 0.2 0 0 5 15 10 PZC time (ms) 20 70 n 0 70 80 90 100 110 120 Bias Tone level dB SPL Fig. 11 Effects of a bias-tone level series on AN responses to a tail-frequency tone at 70 dB SPL. The 1s & 2" harmonic suppression thresholds were met at 90 & 100 dB SPL respectively. (Left) the bias-tone level series of bias-tone period histograms. The significant suppression phases for the first and second harmonic criteria are marked by blue and red inverted triangles. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after the positive peak voltage of the 50 Hz earphone drive signal; (Right) The detailed data analysis on the biastone level functions on the suppression effects on tail-tone responses from the example fiber. Top: the ratelevel function for the run with the bias-tone alone and for the bias on the probe tone run. Middle: bias-tone level function of the synchrony index for 1 & 2" harmonic. Note the sharp increase of the synchrony indices at the suppression thresholds. Bottom: level functions of the suppression phase estimates for 1S & 2nd harmonics with their standard errors. The data points meeting the error bound are noted with markers labeled as "Significant 1 StHarmonic" and "Significant 2nd Harmonic". Analysis of the Bias-tone Effects on the Phase of AN Responses to the 3. Tail Frequency Tone at 2.5 kHz The spike records from the bias-tone level series on a fixed level of tail-frequency tone were binned re. the period of the tail-frequency tone at 2.5 kHz in order to form a bias-tone level series of the tailfrequency tone period histograms. The effects of the increasing level of the bias-tone on the phase of AN responses to the tail-frequency tone were analyzed with the vector phase method by plotting the bias-tone level series of the synchrony index and excitation phase to the tail-frequency tone. For the example fiber, the tail-frequency tone at 2.5 kHz was presented at the fixed level of 70 dB SPL together with the bias-tone level series over the range of 70 dB - 120 dB SPL. The tail-frequency period histograms and the level function of the synchrony index and excitation phase for the example fiber are shown in the left and right panel of Fig. 12. The main objective of the data analysis was to quantify the change in the excitation phase due to suppressive effects of the bias-tone by calculating the phase delta from the minimum level of the bias-tone of the series to a bias-tone level at or above the suppression threshold which was detected by the analysis methods described earlier. Bias-tone Effects on 2.5 kHz Tail-tone Synchrony: Bias-tone level series of Tail-tone period histogram tail-tone level at 70 dB SPL I II Bias-tone Level dB SPL II I J 110 Synchrony Index Significant Excite Phase . 0 -A C II - -'-1 0.2 f ' -- I I 0- J I 0.1 0 1J: 111 1L 11 J J 1 00 11 I A 0.2 PZC time (ms) 0.4 70 70 80 90 100 110 Bias Tone level dB SPL Fig. 12 Bias-tone effects on the synchrony of AN responses to the tail-frequency tone at 2.5 kHz for the example fiber. The tail frequency-tone was at the fixed level of 70 dB SPL. (Left) Bias-tone level series of the tailfrequency period histograms. The suppression threshold for the first and second harmonic identified previously, i.e., 90 and 100 dB SPL, are noted in blue and red. The excitation phase at the sub- and supra-suppressionthreshold level are marked by a black and red triangle. Note that the zero time reference of the period histogram was the positive peak voltage of 2.5 kHz earphone drive signal, and "PZC time" in the x-axis label refers to the time after the positive peak voltage of 2.5 kHz earphone drive signal; (Right) Bias-tone level function of the synchrony index and phase of excitation to the tail-frequency tone. In the left panel of Fig. 12 for the example fiber, the significant levels of the bias-tone identified from the previous analysis procedures are marked as blue, red and green text for the suppression threshold for the first harmonic, the suppression threshold for the second harmonic and the excitation threshold respectively. The excitation phase at the minimum level of the bias-tone at 70 dB SPL is marked by a black triangle, and the excitation phase at 100 dB SPL, i.e., the suppression threshold for the second harmonic, is marked by a red triangle. For the example fiber, there was a phase lag of 17.50 across these two levels of the bias-tone although the difference was not beyond the standard errors of the phase estimates. Note also that increasing levels of the bias-tone lowered the synchrony index to the tail-frequency tone with drastic reductions at and above the excitation threshold to the bias-tone alone. III. Results Bias-tone effects on AN responses to low-level CF-tones and tail-frequency tone at 2.5 kHz were recorded from the total of 27 fibers over 6 cats. As expected from previous studies on bias-tone effects on low-level CF-tone responses [Patuzzi et al, 1984a; Cai and Geisler, 1996a], significant suppression of low-level CF-tone responses were found from all of the recorded fibers. As for responses to the tailfrequency tone, significant suppression was found from 10 of the total 27 fibers recorded including the fiber, CT026_U050, which was examined in detail in the methods section. The distribution of the fiber count over the CF of the fiber for all of the fibers and the 10 fibers with significant suppression of tailfrequency responses are plotted in Fig. 13 over the three octave bands from 5 kHz to 30 kHz. CF Distribution of the Fiber Count: Total Recorded VS Significant Suppressions on 2.5 kHz Tail 14 - - - - - Total Fiber Count Significant Suppression on 2.5 kHz Tail-tone Response 10 8 U- 4 2 01 5 10 CF of the AN fiber, kHz 20 30 Fig. 13 AN fiber count over the three octave bands of CF from 5 kHz to 20 kHz: Total recorded fibers are plotted in green, and the fibers with significant suppression of tail-frequency responses are plotted in red. Detailed results from five additional example fibers with significant suppression of tail-frequency responses are presented as follows: A. B. C. D. E. Example Example Example Example Example Result Result Result Result Result from from from from from CT023._U066: CT031_U078: CT026_U009: CT026_U041: CT028_U044: CF=7.16 kHz, 80 sps CF=7.37 kHz, SR=13.2 sps CF=12.8 kHz, SR=1.7 sps CF=13.43 kHz, SR=78.7 sps CF=19.37 kHz, SR=0.6 sps In the section F, the phase of excitation by the 50 Hz bias-tone alone is compared to the phase of major suppression of low-level CF-tone response from the cumulative data pool. In section G, comparisons of the bias-tone effects on low-level CF-tone responses VS near-threshold-level tailfrequency tone at 2.5 kHz are presented. Then, in the section H, the bias-tone effects on the phase of AN responses to the tail-frequency tone at 2.5 kHz are explored. A. Example Result from CT023_U066: CF=7.16 kHz, 80 sps Note that for this fiber, bias-tone effects on the tail frequency of 1 kHz in addition to 2.5 kHz were recorded. Also note that runs for the bias-tone effects on tail-frequency responses were limited to bias-tone effects on probe-tone level series without the subsequent runs on bias-tone level series. Those details are noted by the plot of the level series of 2.5 kHz and 1 kHz tones together with the tuning curve of the fiber in Fig. 14. 120 Tuning Curve cf CT23, Unit 66: CF=7.16 kHz, Th=2.5 dB SPL IL 100 llx + 0 TC CF-tone 2.5 kHz tone X 1.0 kHz tone 0 X 80 : 0 60 40 I 20 0 10 1 1 I 1 I 100 Tone Frequency (kHz) Fig. 14 CT023_U066: CF=7.16 kHz, SR=80 sps. Threshold TC of the fiber is plotted together with the CF-tone and tail-frequency tones applied on this fiber. Note that on this fiber, bias-tone effects on AN responses to tail frequency tones at 1.0 kHz in addition to 2.5 kHz were recorded. Also note that trials on the tail frequency responses were limited to probe-tone level series without the subsequent bias-tone level series. From the bias-tone effects on low-level CF-tone responses plotted in Fig. 15 and Fig. 16, the typical pattern of twice-per-bias-tone-period suppression was found with the suppression threshold at 90 dB SPL. The major suppression phase was approximately in the middle of the period. Note that the zero phase reference of the bias-tone period histograms was the positive peak voltage of 50 Hz drive of the earphone. Suppression effects of 50 Hz bias-tone at the fixed level of 90 dB SPL on a level series of tailfrequency tone at 2.5 kHz are shown in Fig. 17. Note that 90 dB SPL of the bias-tone was the suppression threshold of the low-level CF-tone responses for this fiber. Note that for this fiber, biastone runs on tail-frequency responses were limited to probe-tone level series runs. For such fibers, the suppression data were selected from the period histogram with the maximum level of the synchrony index from the runs with significant suppression. For this fiber, significant suppression for both first and second harmonic was met at the probe-tone level of 60, 70 and 80 dB SPL. From the three runs, the maxim level of synchrony index was reached at the probe-tone level of 70 dB SPL. Therefore, the suppression data were calculated from the runs at 70 dB SPL of the probe-tone, which was 55 dB higher than the level of the CF-tone. Note that the overall shape of the period histogram at that level was quite similar to the suppression pattern on low-level CF-tone responses from Fig. 15 including the typical twoper-cycle pattern and the similar location of the major suppression phase. Fig. 18 shows the effects of the bias-tone at 90 dB SPL on AN responses to a level series of tail frequency tone at 1 kHz. Significant suppression was found over the range of 60 - 80 dB SPL of the probe-tone level. Note that the major suppression phases of all three types of responses, i.e., CF-tone, 2.5 kHz and 1 kHz tail-frequency tones, were similarly located in the middle of the period. Also note that the minor suppression phase of the suppression pattern of 1 kHz tail frequency (e.g., see the secondharmonic synchrony index) response was significantly less prominent than the other response types. Note that bias-tone effects on the phase of AN responses to tail-frequency tones were not included in the data pool for this fiber and for other fibers that did not have results from of a level series of bias-tone on tail-frequency responses. CT23, U66: CF=7.16 kHz, SR=80 sps (b)Bias on CF-tone: 7.16kHz / 15 dB SPL (a)Bias-tone Only BIAS TONE LEVEL dB SPL 110 100 jL E K 71 1,- cc) II 0.2 0.2 0 0 5 10 PZC time (ms) 15 70 0 I 0 5 :I , 10 LV I 15 - 20 70 PZC time (ms) Fig. 15 CT023_U066: Bias-tone level series of the bias-tone period histograms for (a)Bias-tone alone runs: for this fiber, the excitation threshold was not reached by not meeting the rate-threshold criterion; (b) Bias-tone effects on low-level CF-tone responses: suppression threshold for the second harmonic was reached at 90 dB SPL. Although there appears to be excitation at 110 dB SPL in the bias-tone-alone period histogram, this excitation is not important because the suppression thresholds of the tone responses were all well below 110 dB SPL. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero-time reference. CT23, U66: CF=7.16 kHz Analysis of Bias on CF-tone Spike Rate: BT-on-ProbeTone & BT-alone 200 Bias on CF Bias Alone 100 0 70 100 90 80 SPL Tone leveldB Bias Synchrony Index 110 1st Harmonic 2nd Harmonic 0.5 0 70 100 90 80 Bias Tone level dB SPL Suppression Phase Estimate 110 1 Significant 1st Har Significant 2nd Har 0.5 0 100 90 80 Bias Tone level dB SPL 110 Fig. 16 CT023_U066: The detailed data analysis on the bias-tone level functions of the suppression effects on CFtone responses from CT023 U066. Top: the rate-level function for the run with the bias-tone alone and for the bias on the probe tone run. Middle: bias-tone level functions of the synchrony index for 1s,& 2 nd harmonics. Note the sharp increase of the synchrony indices at the suppression thresholds. Bottom: level function of the suppression phase estimates for 1 st & 2nd harmonic with their standard error. The data points meeting the error bound are noted with markers labeled as "Significant 1s Harmonic" and "Significant 2 Harmonic". CT23, U66, CF=7.16 kHz: Bias on 2.5 kHz Tail-tone Probe-tone level series, Bias-tone at 90 dB SPL Spike Rate 200 I'll Probe-tone Level U) 100 dB SPL n 90 60 I lI 70 80 90 Probe Tone level, dB SPL Synchrony Index 1st Harmonic 2nd Harmonic U) 0.5 Cn 0 80 0 60 70 80 9( Probe Tone level dB SPL Suppression Phase Estimate 1 f, Significant 1st Har Significant 2nd Har CO 60 0.5 00 5 10 15 PZC time (ms) 20 60 70 80 Probe Tone level dB SPL Fig. 17 CT023_U066: Suppression effects of 50 Hz bias-tone at the fixed level of 90 dB SPIL on a level series of tail-frequency tone at 2.5 kHz. See main text for details. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. CT23, U66, CF=7.16kHz: Bias on 1 kHz Tail-tone Probe-tone Level Series: Bias at 90 dB SPL Probe-tone Level dB SPL Spike Rate: BT -on-ProbeTone - CD200 o 100 0 90 I I 60 b ~ 80 70 80 Probe Tone leveldB SPL Synchrony Index 1st Harmonic 2nd Harmonic 0.5 C ciD 0 70 1 70 60 70 80 90 Probe Tone level dB SPL Suppression Phase Estimate Significant 1st Har Significant 2nd Har C 60 0 5 10 PZC time (ms) CU 60 70 80 Probe Tone level dB SPL Fig. 18 CT023_U066: Suppressive effects of 50 Hz bias-tone on probe-tone level series of AN responses to 1 kHz tone. The 50 Hz bias-tone was presented at the fixed level of 90 dB SPL which was the suppression threshold on low-level CF-tone responses. Significant suppression was found over the range of 60 - 80 dB SPIL of the probetone level. Note the similarity of the major suppression phase among the suppressive patterns on the low-level CF-tone and the responses to tail-frequency tones at 2.5 kHz and 1 kHz. Also note that the minor suppression phase was not clearly visible from period histograms in contrast to the pattern on 2.5 kHz from Fig. 17. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. B. Example Result from CT031_U078: CF=7.37 kHz, SR=13.2 sps The tuning curve and the frequency-level points of the probe-tone stimuli used on this fiber are plotted in Fig. 19 CT031_U078: CF=7.37 kHz, SR=13.2 sps. Threshold TC of the fiber is plotted together with the CF-tone and tail-frequency tones applied on this fiber. A typical pattern of suppression was found from the bias-tone effects on low-level CF-tone responses from this fiber as shown in Fig. 20(b) and Fig. 21 with the suppression threshold for the second harmonic at 90 dB SPL and with the major suppression phase at a similar location as with the other example fibers. From the bias-tone effects on the probe-tone level series of 2.5 kHz tail-tone, the tail-frequency tone level of 70 dB SPL was found to be the most sensitive to bias-tone induced suppression effects. As shown in Fig. 19, that level of the tail-frequency tone was very near the neural threshold of the fiber. The effects of a level series of bias-tone on AN responses to the tail-frequency tone are shown in Fig. 22. Suppression threshold for the first harmonic was met at the bias-tone level of 90 dB SPL. Note that rate suppression is also noticeable with a dip in the rate-level curve at the suppression threshold of 90 dB SPL as shown in the right panel of Fig. 22. Fig. 23 shows the effects of the bias-tone on the excitation phase of AN responses to the tailfrequency tone. Increasing level of the bias-tone introduced additional delay to the excitation phase. Note, however, the phase shifts were not beyond standard error of the data points. Tuning Curve of CT31, Unit 78: CF=7.37 kHz, Th=1 1.7 dB SPL 120 I 100 I I I I I I I I I I I I I I CF-tone 2.5kHz Tail-tone 60 0 01 0-1 100 Tone Frequency (kHz) Fig. 19 CT031_U078: CF=7.37 kHz, SR=13.2 sps. Threshold TC of the fiber is plotted together with the CF-tone and tail-frequency tones applied on this fiber. CT31, U78, CF=7.37 kH, SR=13.2 sps (a) Bias-tone Only (b) Bias on CF-tone BIAS TONE LEVEL dB SPL 1hi I[ It 110 1 [II 100 - FI I - II - E 110 100 CL, U Iii CD III 11 90 C C'n I ... I I I [ I 5 10 15 PZC time (ms) II 20 I If I, I] , 70 0 5 10 PZC time (ms) Fig. 20 CT031_U078: Bias-tone level series of period histograms. (a) Bias-tone alone: excitation threshold was reached at 110 dB SPL. The excitation phase is noted by a green triangle; (b) Bias-tone effects on low-level CFtone: suppression threshold for the second harmonic was reached at 90 dB SPL with a similar phase of major suppression as the other example fibers from Fig. 15. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. CT31, U78, CF=7.37 kHz: Analysis of Bias on CF-tone Spike Rate: BT-on-ProbeTone & BT-alone i200 Bias on CF Bias alone o 100 0- 90 80 70 (1) 100 110 Bias Tone leveldB SPL Synchrony Index x 1 - - -1 c 2 0.5 </) 0 - 1st Harmonic 2nd Harmonic 70 08 100 90 80 Bias Tone leval dB SPL Suppression Phase Estimate 110 .C C 0 - 1C 0.5 70 0- - Significant 1st Har Significant 2nd Har 90 80 100 110 Bias Tone level dB SPL Fig. 21 CT031_U078: The detailed data analysis on the bias-tone level functions of the suppression effects for CF-tone responses from CT031_U078. Top: the rate-level function for the run with the bias-tone alone and for the bias on the probe tone run. Middle: bias-tone level function of the synchrony index for 1 st & 2nd harmonics. Note the sharp increase of the synchrony indices at the suppression thresholds. Bottom: level function of the suppression phase estimates for 1 & 2 harmonic with their standard error. The data points meeting the error bound are noted with markers labeled as "Significant 1st Harmonic" and "Significant 2 Harmonic". CT31, U78, CF=7.37 kHz: Bias on 2.5 kHz Tail-tone Spik e Rate: BT-on-ProbeTone & BT-alone Bias-tone Level (D 20 dB SPL () C') 110 10 e n I Bias on 2.5 kHz Ton Bias alone 70 80 90 100 110 Bias Tone level dB SPL Synchrony Index k 100 1st Harmonic 2nd Harmonic 0.5 II 14 0 90 L Ul) _0 70 80 90 100 110 Bias Tone level dB SPL Suppression Phase Estimate 1 0 0.05 C 0 () CO r ,I 0 t - 10 15 PZC time (ms) 70 CO -C 0.5 Significant 1st Har Significant 2nd Har 0 80 90 100 110 Bias Tone lewl dB SPL Fig. 22 CT031_U078: Bias-tone effects on responses to 2.5 kHz tail-frequency tone at 70 dB SPL, which was very near the threshold level from the tuning curve as shown in Fig. 19. Suppression threshold for the first harmonic was reached at 90 dB SPL of the bias-tone with the major suppression phase in the middle of the period. Note that the suppressive effect can also be seen from the rate-level function as a dip at the suppression threshold of 90 dB SPL. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. CT31, U78, CF=7.37 kHz: Bias-tone Effects on 2.5 kHz Tail-tone Synchrony Bias-tone Level dB SPL Synchrony Index Significant Excite Phase II I II 110 II K 100 aC 0 I ii 90 0 Ch .C CL 0.05 0 0 0.1 A 0.2 PZC time (ms) 0 70 90 100 80 Bias Tone level dB SPL Fig. 23 CT031_U078: Bias-tone effects on the phase of AN responses to near-threshold tail-frequency tone. 2.5 kHz tone at 70 dB SPL was presented together with a bias-tone level series with the range of 70 - 120 dB SPL. (Left) Bias-tone level series of probe-tone period histograms. The suppression threshold of 90 dB SPL and excitation threshold by 50 Hz alone are noted in blue and green respectively. The phase of excitation by the tailfrequency tone at the minimum level of the bias-tone is noted by a black triangle at 70 dB SPL. The phase of excitation under suppressive effects of the bias-tone is noted by a red triangle at 100 dB SPL of the bias-tone. Note that the zero time reference of the period histogram was the positive peak voltage of the 2.5 kHz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference; (Right) Bias-tone level series of the synchrony index and phase of excitation in response to the 2.5 kHz tail-frequency tone. Note the sharp dip in the synchrony at 110 dB SPL of the bias-tone, the excitation threshold by the bias-tone alone. Also note the trend of phase delay with increasing level of the bias-tone although the phase deltas were not statistically significant. The effect of the bias-tone on the phase of tail-frequency response was taken as the difference between the phase at 100 dB SPL VS 70 dB SPL. C. Example Result from CT026_U009: CF=12.8 kHz, SR=1.7 sps The tuning curve and the frequency-level points of the probe-tone stimuli used on this fiber are plotted in Fig. 24. As shown in Fig. 25, the suppression threshold on low-level CF-tone responses was reached at 90 dB SPL and 100 dB SPL for the first and second harmonic criterion respectively. The shape of the suppression pattern including the major suppression was similar to the results from the earlier example fibers. At the bias-tone level of 110 dB SPL, i.e., 20 dB above the suppression threshold of CF-tone responses, significant suppression was found from the probe-tone level series of the 2.5 kHz tailfrequency tone responses at 80 dB SPL of the tail-frequency tone as shown in Fig. 26. Note that the major suppression phase of the tail-frequency tone responses was similar to that of CF-tone responses as in earlier example fibers. Note that for this fiber, the results from the bias-tone only runs are unavailable. Such data points are marked separately in cumulative data plots in Fig. 37, Fig. 38, Fig. 39, and Fig. 40. 120 Tuning Curve of CT26, Unit 9: CF=12.88 kHz, Th=5.9 dB SPL - + 100, 0 0 TC CF-tone 2.5 kHz Tail-tone 80 60 I I I 40 + 0 I0- 100 Tone Frequency (kHz) Fig. 24 CT026_U009: CF=12.8 kHz, SR=1.7 sps. Threshold TC of the fiber isplotted together with the CF-tone and tail-frequency tones applied on this fiber. CT026, U009, CF=12.8 kHz, SR=1.7 sps: Bias on CF-tone Responses Bias-tone LeveI dB SPL Spike Rate: BT-on-ProbeTone 12 100 a,5 120 0 0 C,) 70 110 x 1 -[1 C: 0 90) -V 120 C 0.5 100 hI - 80 90 100 110 Bias Tone level dB SPL Synchrony Index 1st Harmonic 2nd Harmonic CD - - 70 90 C) 100 110 120 90 80 Bias Tone level dB SPL Suppression Phase Estimate 1- 0 40.5 70 0 0 5 15 10 PZC time (ms) 20 -C TU Significant 1st Har Significant 2nd Har u 9I TUU 1 0 120 Bias Tone level dB SPL Fig. 25 CT026_U009, Bias on CF-tone Responses: Suppression threshold for the first harmonic and second harmonic was reached at the bias-tone level of 90 and 100 dB SPL respectively. The shape of the suppression pattern including the major suppression phase was similar to those of earlier examples. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. CT026, U009: Bias on 2.5 kHz Tail-frequency Responses: Probe-tone level series Bias-tone at 110 dB SPL Spike Rate: BT-on-ProbeTone U) -i 1 Probe-tone LeveI 300 i 3 200 dB SPL 100 CD 100 I 7-5 0 - ' 7Q 1st Harmonic 2nd Harmonic I 70 1,1 - 1- 1 V 80 90 100 Probe Tone leveldB SPL Synchrony Index 1 1 80 90 100 Probe Tone level dB SPL Suppression Phase Estimate 0.5 Significant 1st Harmonic 5 10 15 PZC time (ms) 20 0 70 80 90 100 Probe Tone IeveI dB SPL Fig. 26 CT026_Uo09, Bias on 2.5 kHz Tail-frequency Responses: (Left) Effects of bias-tone at the fixed level of 110 dB SPL on a probe-tone level series of 2.5 kHz tail-frequency responses. Significant suppression for the first harmonic was found at 80 dB SPL of the tail-frequency tone which was very close to the neural threshold from the tuning curve shown in Fig. 24. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. D. Example Result from CT026_U041: CF=13.43 kHz, SR=78.7 sps The tuning curve and the frequency-level points of the probe-tone stimuli used on this fiber are plotted in Fig. 27. The suppression threshold of the low-level CF-tone responses was reached at 90 dB SPL of the bias-tone as shown in Fig. 28. The suppression pattern followed the typical pattern seen from the other fibers including the similar location of the major suppression phase. The effects of a bias-tone level series on AN responses to the tail-frequency tone at 2.5 kHz and 80 dB SPL are shown in Fig. 29. As with other example fibers, the level of the tail-frequency tone identified from the probe-tone level series was very near the neural threshold level as shown in Fig. 27. The suppression threshold for the second harmonic was reached at 90 dB SPL of the bias-tone, and the typical pattern of bias-tone induced suppression is clearly visible at the bias-tone level of 110 dB SPL. The effects of a bias-tone level series on the phase of AN responses to the tail-frequency tone are shown in Fig. 30. The bias-tone introduced a phase lag of ~10 deg in the AN responses to the tailfrequency tone between the bias-tone level of 70 dB SPL and 110 dB SPL. Note that the results from bias-tone alone runs are unavailable from this fiber. 120 Tuning Curve of CT26, Unit 41: CF=13.43 kHz, Th=11.1 dB SPL TC CF-tone 2.5 kHz Tail-tone 100 0' 80 60 40 20, 0 10 100 Tone Frequency (kHz) Fig. 27 CT026_U041: CF=13.43 kHz, SR=78.7 sps. Threshold TC of the fiber is plotted together with the CF-tone and tail-frequency tones applied on this fiber CT026 U041, CF=13.43 kHz, SR=78.7 sps: Bias on CF-tone Responses BIAS TONE LEVEL dB SPL - 120 Spike Rate: BT-on-ProbeTone 100 50 80 90 100 110 120 Bias Tone level dB SPL Synchrony Index 1st Harmonic 2nd Harmonic 100 II II 0.5 0 ~1 1 70 80 90 100 110 Bias Tone level dB SPL 120 Suppression Phase Estimate 0.5 5 10 PZC time (ms) 15 20 0 70 Significant 1st Har Significant 2nd Har 80 90 100 110 Bias Tone level dB SPL 120 Fig. 28 CT026_U041, CF=13.43 kHz: Bias-tone effects on low-level CF-tone responses. Suppression threshold was reached at 90 dB SPL of the bias-tone for the second harmonic criterion. The suppression pattern including the major suppression phase was similar to the other fibers. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. CT026 U041, CF=13.43 kHz: Bias on 2.5 kHz Tail-frequency Responses Probe-tone at 80dB SPL Spike Rate: BT-on-ProbeTone s 60Bias-tone Level () - 40 dB SPL ' CD I 20 120 0 - 11 110 I C,, E 0. IIIhr' IlIIl C,, 0.5 0 ii I 1st Harmonic 2nd Harmonic 100 U) I 120 100 80 Bias Tone level dB SPL Synchrony Index [I t 1111 90 C,, 120 100 80 Bias Tone level dB SPL Suppression Phase Estimate _0 0 -C : 1 I 80 0 C,, .C, 0 0 5 5 10 15 PZC time (ms) 20 70 - Significant 1st Har Significant 2nd Har 120 100 80 Bias Tone level dB SPL Fig. 29 CT026_UO41, CF=13.43 kHz: Bias-tone effects on AN responses to 2.5 kHz tone at 80 dB SPL. Suppression threshold for the second harmonic was reached at 90 dB SPL of the bias-tone. The suppression pattern at 110 dB SPL clearly shows the typical bias-tone induced pattern of twice-per-period suppression. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. CT026 U041, CF=13.43 kHz: Bias-tone Effects on 2.5 kHz Tail-tone Response Phase Bias-tone Level dB SPL 120 I I I Synchrony Index Significant Excite Phase _1 1 110 ~0 a- 100 111 -1 0.1 IIIII 0.2 0.3 PZC time (ms) 0 ; C/) 90 0.4 cc aD-- 0 60 I-- - 80 100 120 Bias Tone leveI dB SPL Fig. 30 CT026_UO41, CF=13.43 kHz: Bias-tone effects on the phase of AN responses to 2.5 kHz tone at 80 dB SPL. The suppression threshold of 90 dB SPL identified in Fig. 29 is marked in red. The phase delta introduced by the bias-tone was calculated as the difference between the phase of excitation at the minimum level of the biastone (marked with a black triangle) and at 110 dB SPL (marked with a red triangle). The bias-tone introduced ~10 deg of phase lag. Note that the difference was not beyond the standard error. Note that the zero time reference of the period histogram was the positive peak voltage of the 2.5 kHz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. E. Example Result from CT028_U044: CF=19.37 kHz, SR=0.6 sps The tuning curve and the frequency-level points of the probe-tone stimuli used on this fiber are plotted in Fig. 27. The suppression threshold of the low-level CF-tone responses was reached at 100 dB SPL of the bias-tone as shown in Fig. 32. The suppression pattern followed the typical pattern seen from the other fibers including the similar location of the major suppression phase. The effects of a bias-tone level series on AN responses to the tail-frequency tone at 2.5 kHz and 90 dB SPL are shown in Fig. 34. The suppression threshold was reached at 120 dB SPL of the bias-tone; however, the minor suppression phase was not apparent unlike other example fibers. Note that for this fiber the excitation threshold for bias-tone alone runs was not reached. The effects of a bias-tone level series on the phase of AN responses to the tail-frequency tone are shown in Fig. 35. The bias-tone introduced a phase lag of ~40 deg in the AN responses to the tailfrequency tone between the bias-tone level of 70 dB SPL and 120 dB SPL. 120 Tuning Curve of CT28, Unit 44: CF= 19.37 kHz, Th= 19.8 dB SPL + 0 100, TC CF-tone 2.5 kHz Tail-tone 0 80 60 40 20 0 1041 100 Tone Frequency (kHz) Fig. 31 CT028_U044, CF=19.37 kHz, SR=0.6 sps: Threshold TC of the fiber isplotted together with the CF-tone and tail-frequency tone applied on this fiber CT028, U044: CF=19.37 kHz, SR=0.6 sps (a) Bias-tone Alone (b) Bias on CF-tone BIAS TONE LEVEL dB SPL 120 120 110 E (D _W 100 II I J II 100 Ca 90 Cl) 90 0.05 0 0 5 10 - PZC time (ms) 15 20 5 10 15 20 PZC time (ms) Fig. 32 CT028, U044: CF=19.37 kHz. Bias-tone level series of bias-tone period histograms. (a) Bias-tone only. Excitation threshold was not reached; (b)Bias on low-level CF-tone responses. Suppression threshold for the second harmonic was reached at the bias-tone level of 100 dB SPL. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. CT028, U044: CF=19.37 kHz, Analysis of Bias on CF-tone Spike Rate: BT-on-ProbeTone & BT-alone (n 100 (D 50 Bias on CF -,Bias alone n (I) 80 110 100 90 SPL Bias Tone le'.eldB Synchrony Index 120 x ~0 1st Harmonic 2nd Harmonic C c 01... 0.5 C., c C,) 0 70 Co V 0 a) 0~ a) 100 110 90 80 Bias Tone level dB SPL Suppression Phase Estimate 120 1 C 0 U, 0.5 Significant 1st Har Significant 2nd Har a) m ai U) a) 0~ 0 70 80 110 100 90 SPL dB level Tone Bias 120 Fig. 33 CT028_U44: The detailed data analysis on the bias-tone level functions of the suppression effects for CFtone responses. Top: the rate-level function for the run with the bias-tone alone and for the bias on the probe tone run. Middle: bias-tone level functions of the synchrony index for 1 & 2 harmonics. Bottom: level functions of the suppression phase estimates for 1st & 2nd harmonics with their standard errors. The data points meeting the error bound are noted with markers labeled as "Significant 14 Harmonic" and "Significant 2"n Harmonic. CT028, U044: Bias on 2.5 kHz Tail-frequency Responses Probe-tone at 90 dB SPL Bias-tone Spike Rate: BT-on-ProbeTone & BT-alone - Level dB SPL ai 40 Bias on 2.5 kHz Tone Bias alone 20 120 (D) 70 80 90 1oo 110 Bias Tone leveldB SPL 120 110 Synchrony Index 1 E 1st Harmonic 2nd Harmonic 2 0.5 100 C U) 0 0.1 0 70 80 90 100 110 Bias Tone level dB SPL 120 Suppression Phase Estimate (n _0 0 80 a 0 0.5 CU U) 5 10 15 PZC time (ms) 20 70 C 70 Significant 1st Har Significant 2nd Har L __.L _ ...__L 80 90 100 110 Bias Tone level dB SPL 120 Fig. 34 CT028_U44: Bias-tone effects on AN responses to 2.5 kHz tail-frequency tone at 90 dB SPL. The suppression thresholds for both the first and second harmonics were reached at the bias-tone level of 120 dB SPL. Since the synchrony index for the second harmonic was half of the first harmonic synchrony index at the suppression threshold, the major suppression phase was estimated with the phase of the first harmonic. Note also qualitatively that the minor suppression phase was not apparent for this fiber. Note that the zero time reference of the period histogram was the positive peak voltage of the 50 Hz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. CT028, U044: Bias-tone Effects on Phase of AN Responses to 2.5 kHz Tail-frequency Tone 1 II Bias-tone Level dB SPL 0.9! 120 110 . 0 (D a- 0.6 C 0.5 0.4 0.9 Synchrony Index Significant Excite Phase 0 (D .0 0 100 lr' - LI an 0.3 0.2 0.1 0 0 W, A 0.1 0.3 0.2 PZC time (ms) 0.4 100 80 Bias Tone Ievel dB SPL Fig. 35 CT028_U044: Bias-tone effects on the phase of AN responses to 2.5 kHz tail-frequency tone at 90 dB SPL. The suppression threshold of 120 dB SPL identified in Fig. 34 is marked in red. The phase delta introduced by the bias-tone was calculated as the difference between the phase of excitation at the minimum level of the biastone (marked with a black triangle) and at 120 dB SPL (marked with a red triangle). The bias-tone introduced ~40 deg of phase lag. Note that the zero time reference of the period histogram was the positive peak voltage of the 2.5 kHz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. 53 F. Phase of Excitation by 50 Hz Bias-tone Alone and Phase of Suppression on Low-level CF-tone Responses The phase of excitation by the 50 Hz bias-tone alone is plotted versus the CF for fibers with CFs from 1 kHz to 30 kHz in Fig. 36. Also plotted in Fig. 36 is the phase of major suppression on low-level CFtone responses at the suppression threshold level. The standard error on all phase estimates was within ±30* which is noted on the plot with the blue error bar. Note that a part of these data appeared previously in [Nam, 2011]. The phase of excitation by the bias-tone alone provides a phase which is thought to be based on cochlear mechanics. Specifically, previous studies in guinea pigs have found that the phase of excitaiton by a low-frequency tone such as 50 Hz is aligned with the phase of peak velocity of BM displacement toward scalar vestibuli (SV) [Sellick et al, 1982]. The results from this study in Fig. 36 show that the phase of suppression on low-level CF-tone responses remained relatively flat throughout the CFs tested, and the span of phase values was approximately within one standard error of the phase estimate except for some fibers with reversed phase of supression for CF > 20 kHz. Such reversals of major suppression phase have also been reported in [Cai and Geisler, 1996a]. Also note that the phase of major suppression of low-level CF-tone responses was ~1/4 cycle away from the phase of excitation by the bias-tone alone. Similar results have been reported previously and interpreted as the phase of suppression by the bias-tone is alinged with the peak displacment of BM whereas the phase of excitation is aligned with the peak velocity of BM movement [Sellick et al, 1982; Cai and Geisler, 1996a]. 1 Bias-tone Excitation Phase VS Suppression Phase of CF-tone IAI + A A++ 09 + .0.8-A A 0.8& A A A *0.7 0.6 + A + + 0 &++, + 0.5 I... 0.4 + A + + + + + + ++. + A A 0 *0.3 + + A CU 0.2 0.1 Max Std Err 1 A + Bias-tone Excite phase CF-tone Suppress Phase 5 CF, kHz 10 30 Fig. 36 Phase of excitation by the "bias-tone alone" and the phase of major suppression on low-level CF-tone responses versus the CF of the fiber. The zero phase reference of the plot is the phase of the positive peak of the 50 Hz drive voltage to the earphone. G. Comparison of Bias-tone Effects on AN Responses to Low-level CFtone Responses VS Near-threshold-level Tail-frequency Tone at 2.5 kHz As the results of the example fibers demonstrate, the typical pattern of twice-per-bias-tone period suppression was consistently found from the bias-tone effects on low-level CF-tone responses. As for the bias-tone effects on AN response to tail-frequency tones, suppressive effects were found from 10 fibers out of the total of 27 fibers recorded. Further, from the six example fibers examined in Fig. 14 Fig. 35, the suppression patterns of AN responses to tail-frequency tone at 2.5 kHz were overall qualitatively similar to those of CF-tone responses. The pattern of suppression by the 50 Hz bias-tone on these two responses types was compared quantitatively by first examining the major suppression phases as shown in Fig. 37. Overall, the major suppression phases of the two response types were quite similar, particularly for fibers with CF < 15 kHz. The suppression amplitude differences between the major and minor phases for the two response types was examined by plotting the log ratio of the second harmonic synchrony index over the first harmonic synchrony index at the suppression threshold as shown in Fig. 38. Higher values of this ratio indicate a stronger presence of the minor suppression phase thereby indicating a more symmetric location of the operating point at rest of the OHC transduction function driving the AN responses. The trend of higher level of symmetry at higher CF of the fiber was apparent for the bias-tone effects on lowlevel CF-tone responses. A similar pattern has been reported in [Cai and Geisler, 1996a]. As for the biastone effects on tail frequency responses, such a trend was less evident; however, the range of values for the two response types were not drastically different. Phase of Suppression: CF-tone VS 2.5 kHz tail tone Max Std Err 0.91 0.8 0 lo 0.7 a. +0 0.6 0 + 0 ++ Cu cn 0.5 + 0.4 CU 0.3 a. 0.2 0.1 + CF-tone o 2.5 kHz Tail-tone Tail-tone NO bias-tone + 10 CF, kHz Fig. 37 The major suppression phase of the bias-tone effects on AN responses to low-level CF-tones compared to AN responses to near-threshold-level tail-frequency tones at 2.5 kHz. Note that 3 of the 10 fibers were without the bias-tone-alone runs; these data are marked with filled circles for the tail-frequency data points. The major suppression phase of the two response types was quite similar except for a fiber with a CF of 16.7 kHz. Note that the major suppression phase for the tail-frequency response of the fiber with the CF of 22.6 kHz was ~1/2 period from most other suppression phases, but it was similar to the suppression phase for low-level CF-tone responses of this fiber and some other fibers with similar CFs. 50Hz Bias Tone Effects on CF VS Tail Tone Burst: Log Ratio of Syncldx2 / Syncldxl 10 5 X - Sync Index 2 Larger 0 0 ++ + g % 0 x + + +-Cton + ++ + 00 S0 + + SyncIndex 2 o ot 0 + Larger F + + +o 0 + o CF-tone Tail 2.5kHz Tail 2.5kHz NO bias-alone -15 I 5 10 30 OF, kHz Fig. 38 Plot of the ratio of the 2f. over the Its harmonic synchrony index at the suppression threshold VS the CF of the fiber. Higher value of the ratio indicates more symmetry in the suppression pattern between the two suppression dips; therefore, higher value of this ratio indicates more symmetrical location of the operating point on the OHC mechano-electric transdauction function. Fig. 39 compares the 50 Hz bias-tone level at the threshold of suppression on the low-level CFtone responses and tail-frequency tone responses. Fig. 39(a) shows the distributions of the fiber count versus the suppression threshold. The median of the distribution for the CF-tone responses was 10 dB lower than that of the tail-frequency responses. Fig. 39(b) displays the suppression thresholds versus the CF of the fiber. For both response types, the suppression threshold increased with the CF of the fiber at the rate of "-10 dB per octave. Also as in the fiber count distribution, the suppression threshold for tail-frequency responses was found to be -'10-15 dB higher than that of low-level CF-tone responses. Since the suppression threshold for low-level CF-tone responses approached the maximum level of the bias tone for fibers with CF > 15 kHz, the higher level of suppression threshold for tail-frequency responses provides a potential explanation for the relatively small number of fibers with significant suppression of tail-frequency responses for CF > 10 kHz as shown in Fig. 13. (a) Histogram of Fiber Count over BT-suppression Threshold Level 1 50 - .4.... CF-tone 28 Fibers .g,...Tail 2.5 kHz 40 . 10 Fibers - 0 -- 320 - 0 ~10 (b) Threshold of Significant Suppression over CF a) 0 + CF-tone 110 0 Tail 2.5kHz Tail 2.5kHz No Bias-alone 0 9- 100 90 6 70 2 60 ~0 + -E + 41 ~80 +0 5 + - +1- + + + + + + + + 0 120 10 Cl) jH 120 110 100 90 80 BT Level of Suppression Threshold, dB SPL 70 60 + + + 30 30 10 CF, kHz Fig. 39 The threshold of suppression for AN responses to low-level CF-tones and tail-frequency tones at 2.5 kHz: (a) Histogram of the fiber count versus the suppression threshold; (b)the plot of the suppression thresholds versus CF. The total fiber counts were 28 and 10 for the CF-tone and tail-frequency tone respectively. These data show that the suppression threshold of the CF-tone responses was typically slightly lower than that of the tail-frequency responses. Bias-tone Effects on the Phase of AN Responses to Tail-frequency H. tone at 2.5 kHz Bias-tone induced shift in the phase of AN responses to a fixed level of 2.5 kHz tail-frequency tone was measured from 5 of the 10 fibers with significant suppression of tail frequency-tone responses. As described in the Example Result from CT028__U044: CF=19.37 kHz, SR=0.6 sps the phase shift was calculated as the difference between the minimum level of the bias-tone and a bias-tone level at or above the suppression threshold. The standard error for a phase difference was calculated as the square root of the sum of the squares of the standard error of the phase estimates. The results are plotted versus the CF of the fiber in Fig. 40. A phase delay of 10 - 40 degrees was found from 4 of the 5 fibers. These results are in general agreement with the effects of MOC efferent stimulation on tailfrequency responses where the average of 15 degree of phase delay was reported [Stankovic and Guinan, 2000]. Note that the phase shift measurements from the other five fibers were not included in the data pool since their suppression the data for those fibers did not include results from a bias-tone level series on tail-frequency responses. For those fibers, significant suppression on tail-frequency responses were found from runs with a probe-tone level series as in the Example Result from CT023_U066: CF=7.16 kHz, 80 sps and the Example Result from CT026_U009: CF=12.8 kHz, SR=1.7 sps Since the phase of AN responses to a level series of tail-frequency tones was found to exhibit phase delays over increasing tone intensity [Stankovic and Guinan, 2000], phase shift measurements from runs with a probe-tone level series would mainly reflect phase shift induced by the changes in the intensity of the tail-frequency tone rather than by the suppression effects of a low-frequency bias-tone. Bias-tone Induced Shift in the Phase of AN Responses to Tail-frequency tone at 2.5 kHz 20 Lead 10 0 -10 -20 IT A Lag -30 -40: -50 -60' 30 5 CF, kHz Fig. 40 Bias-tone induced shift in the phase of AN responses to 2.5 kHz tail-frequency tone. Phase delay of 10 40 degree was found from 4 of the 5 fibers. IV. Discussion A. Concerns for Potential Artifacts Caused by Direct Stimulation of IHC Stereocilia by the Bias-tone Although the 50 Hz bias-tone used in this study is expected to stimulate the OHC stereocilia with higher amplitude than the IHC stereocilia [Russell and Sellick, 1983], it has been found in this study and elsewhere [Sellick et al, 1982; Patuzzi et al, 1984a; Cai and Geisler, 1996a] that AN responses with biastone alone do show both firing rate elevation and phasic modulation of period histograms at high enough levels of the bias-tone. In order to guard against such artifacts from bias-tone-induced suppression patterns, the threshold of excitation by the bias-tone alone was measured, and the suppression records which were invoked at a bias-tone level at or above the excitation threshold were not included in the data pool. Note that the cumulative data plots included the three fibers which were without the results from the bias-tone alone runs. Artifact rejection test could not be run on these fibers, and they are separately marked in Fig. 37, Fig. 38, Fig. 39, and Fig. 40. This excitation-threshold-based artifact rejection strategy is discussed in detail in [Nam, 2011], and found to be effective for the suppression data on low-level CF-tone responses. As for the three fibers in the cumulative data plots that did not have bias-tone-alone runs, they have been included in Figs. 37-40 because the suppression thresholds for these fibers were below the typical bias-tone-alone excitation threshold. Since their data did not deviate significantly from other data points with similar CFs, the potential for artifacts in their data is unlikely to be high. B. Bias-tone Induced Suppression Patterns on Tail-frequency Tone Responses Significant suppression of AN responses to tail-frequency tones at 2.5 kHz was found from 10 fibers out of a total of 27 fibers recorded. Those 10 fibers came from 6 different cats and covered the range of CF from 7.16 kHz to 22.6 kHz. Among those 10 fibers, the typical suppression pattern associated with the compressive non-linearity of mechano-electric transduction function of OHCs was found for fibers with CF < 15 kHz. These results directly show that the mechano-electric transduction function of OHCs is involved in generation of AN responses to 2.5 kHz tone for fibers with CF < 15 kHz in a similar way to how it is involved in producing cochlear amplification for low-level CF-tones. There are some reservations on the above conclusion, however. Specifically, as shown in Fig. 13, significant suppression was found from only 5 out of 22 fibers recorded from the fibers with CF > 10 kHz. These results bring up a question as to whether the involvement of the mechano-electric transduction function of OHCs is either limited to CF < 10 kHz or limited to certain fibers or a certain class of fibers. As mentioned in the methods section, one explanation for the relatively low proportion of fibers with significant suppression at tail frequencies is that, for tail frequency tones, there is only a narrow range of levels near the threshold that is sensitive to the suppressive effects of a bias-tone, and tests of suppression were done on a limited range of tail-frequency sound levels. Similar results have been reported in the inhibitive effects of MOC efferent stimulation [Stankovic and Guinan, 2000]. The major suppression phase and the symmetry between the major and minor suppression phases of the AN responses to low-level CF-tones and tail-frequency tone were compared in order to determine potential differences in the shape or the operating point at rest of the mechano-electric transduction function producing the two response types. The results overall did not reveal significant differences as shown in Fig. 37 and Fig. 38 thereby indicating that the detailed mechanism of the mechano-electric transduction function producing the two responses types is similar. Note, however, that this conclusion does not either support or refute the hypothesis that the mechanism of OHC involvement in AN response generation for tail-frequency tones must be somehow different from that of the low-level CF-tone responses [Stankovic and Guinan, 1999]. It does, however, indicate that any potential differences in the OHC mechanism between these response types do not involve the mechanoelectric transduction function of OHCs. C. Bias-tone Induced Phase Shift in Tail-frequency Tone Responses The shift in the phase of AN responses to tail-frequency tones at 2.5 kHz induced by a bias-tone ranged from -45' (phase lag) to +5* (phase lead) as shown in Fig. 40. A phase lag was found from 4 of the 5 fibers. These results are in general agreement with the MOC efferent effects on the phase of AN responses which ranged from -80' to +600 with the average of -15', a phase lag [Stankovic and Guinan, 2000]. The general agreement between the results of this study and the MOC efferent effects seems to suggest that the MOC efferents and a low-frequency bias-tone affect the phase of AN responses to a tail-frequency tone through a similar mechanism by lowering the gain of the transduction function of OHCs, i.e., the forward transduction function for the case of bias-tone and the reverse transduction function for MOC efferents. Note, however, that the mechanism behind the bias-tone effects on the phase responses demands further consideration. Specifically, high-levels of a bias-tone may affect the phase response to a tail-frequency tone through a non-linear mechanism of the cochlea other than the mechano-electric transduction function of OHCs. A possible test of this alternate explanation might be to examine the bias-tone-induced shifts in the phase of AN responses to a tail-frequency tone presented at a level well above the threshold so that the bias-tone did not introduce significant suppression (e.g. at bias-tone levels where there was no MOC inhibition, see Fig. 1). If the shifts in the phase responses were similar between the runs with and without significant suppressions induced by a bias-tone, serious doubts would be raised on the mechanism behind the phase shifts found in this study. One example of this test is presented through the fiber, CT013_U033, with the CF of 22.45 kHz. For this fiber, the tail-frequency tone at 2.5 kHz was presented at 85 dB SPL which was ~20 dB above the neural threshold, and a bias-tone level series spanning up to 110 dB SPL did not introduce significant suppression. The bias-tone effects on the phase and synchrony of AN responses to the tail-frequency tone are shown in Fig. 41. The bias-tone level function of the synchrony index and excitation phase in response to the tail-frequency tone did not change significantly over the bias-tone levels in contrast to the results shown in Fig. 12, Fig. 23 and Fig. 35 where the bias-tone introduced significant suppressions. Results of this test are limited to this single example. Investigations on this issue are left for future studies on this topic. Bias-tone Effects on the Phase of AN Responses to 2.5 kHz tone at 85 dB SPL CT013, U033: CF=22.45 kHz, SR=75.1 sps Bias-tone Level dB SPL Synchrony Index Significant Excite Phase 0.9 'IIW II h L Ij U) -O 100 0 0. C 0 a. ] to .c -C a. III I , , I 0.2 0.3 PZC time (ms) - 0.4 70 0 70 - 80 - 90 100 110 Bias Tone level dB SPL Fig. 41 Bias-tone effects on the phase and synchrony to a 2.5 kHz tail-frequency tone presented at a level well above the threshold. The data came from the fiber, CT013_U033, with the CF=22.45 kHz. The tail-frequency tone was presented at 85 dB SPL which was ~20 dB above the tuning curve threshold. A bias-tone level series did not introduce significant suppressions (Data not shown). Also, the bias-tone level series did not affect the synchrony or the phase of response to the tail-frequency tone. Note that the zero time reference of the period histogram was the positive peak voltage of the 2.5 kHz earphone drive signal, and "PZC time" in the x-axis label refers to the time after that zero time reference. V. References Cai Y, Geisler CD (1996c) Suppression in auditory-nerve fibers of cats using low-side suppressors. Ill. Model results. Hear Res. 1996 Jul;96(1-2):126-40. Cai Y, Geisler CD (1996a) Suppression in auditory-nerve fibers of cats using low-side suppressors. 1. Temporal aspects. Hear Res. 1996 Jul;96(1-2):94-112. Dallos, P., Cheatham, M. A., and Ferraro, J. (1974). "Cochlear mechanics, nonlinearities, and cochlear potentials," J. Acoust. Soc. Am. 55, 597-605. Goldberg, J. M., and Brown, P. B. (1969). "Response of binaural neurons of dog superior olivary complex to dichotictonal stimuli: some physiological mechanisms Of sound localization, "J. Neurophysiol. 32, 613-636. Guinan (1996) Chapter 8, Physiology of Olivocochlear Efferents, The Cochlea, 436-502, Dallos P, Popper AN, Fay FF., editors (Springer, New York) Guinan JJ Jr (2006) Olivocochlear efferents: anatomy, physiology, function, and the measurement of efferent effects in humans. Ear Hear. 2006 Dec;27(6):589-607. Review. Erratum in: Ear Hear. 2007 Feb;28(1):129 Johnson DH. (1980) The relationship between spike rate and synchrony in responses of auditory-nerve fibers to single tones. J Acoust Soc Am. 1980 Oct;68(4):1115-22. Kiang NYS, Watanabe T, Thomas EC, Clark LF (1965) Discharge Patterns of Single Fibers in the Cat's Auditory Nerve (MIT, Cambridge, MA) Kiang NY, Moxon EC (1974) Tails of tuning curves of auditory-nerve fibers. J Acoust Soc Am. 1974 Mar;55(3):620-30 Liberman,M . C., and Dodds,L . W. (1984). "Single-neuron labeling and chronic cochlear pathology.] 11. Stereocilia damage and alterations of threshold tuning curves," Hear. Res. 16, 55-74. Mardia KV (1972) Statistics of Directional Data (Academic Press, New York) Nam HS (2011) Low-frequency Bias-tone Effects on Auditory-Nerve Responses to Clicks and Tones: Investigating Multiple Outer-Hair-Cell Actions on Auditory-Nerve Firing . Ph.D. thesis, Massachusetts Institute of Technology Narayan SS, Temchin AN, Recio A, Ruggero MA. Frequency tuning of basilar membrane and auditory nerve fibers in the same cochleae. Science. 1998 Dec 4;282(5395):1882-4 Patuzzi R, Sellick PM, Johnstone BM (1984a) The modulation of the sensitivity of the mammalian cochlea by low frequency tones. 1. Primary afferent activity. Hear Res. 1984 Jan;13(1):1-8 Robles L, Ruggero MA. Mechanics of the mammalian cochlea. Physiol Rev. 2001 Jul;81(3):1305-52. Review Ruggero MA, Robles L, Rich NC (1992) Two-tone suppression in the basilar membrane of the cochlea: mechanical basis of auditory-nerve rate suppression. J Neurophysiol. 1992 Oct;68(4):1087-99 Ruggero MA, Rich NC, Recio A, Narayan SS, Robles L. (1997) Basilar-membrane responses to tones at the base of the chinchilla cochlea.J Acoust Soc Am. 1997 Apr;101(4):2151-63 Russell IJ,Sellick PM. (1983) Low-frequency characteristics of intracellularly recorded receptor potentials in guinea-pig cochlear hair cells. J Physiol. 1983 May;338:179-206. Schoonhoven R, Keijzer J, Versnel H, Prijs VF (1994) A dual filter model describing single-fiber responses to clicks in the normal and noise-damaged cochlea. J Acoust Soc Am. 1994 Apr;95(4):2104-21 Sellick PM, Patuzzi R, Johnstone (1982) BM Modulation of responses of spiral ganglion cells in the guinea pig cochlea by low-frequency sound. Hear Res. 1982 Jul;7(2):199-221 Stankovic KM, Guinan JJ Jr. (1999) Medial efferent effects on auditory-nerve responses to tail-frequency tones. 1. Rate reduction. J Acoust Soc Am. 1999 Aug;106(2):857-69 Stankovic KM, Guinan JJ Jr. (2000) Medial efferent effects on auditory-nerve responses to tail-frequency tones 11: alteration of phase. J Acoust Soc Am. 2000 Aug;108(2):664-78. Versnel H, Schoonhoven R, Prijs VF. (1992) Single-fibre and whole-nerve responses to clicks as a function of sound intensity in the guinea pig. Hear Res. 1992 May;59(2):138-56.