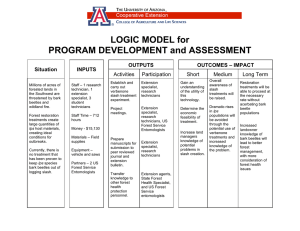
Drought Stress and Bark Beetle Outbreaks in the Climate Change Adaptation
advertisement

Drought Stress and Bark Beetle Outbreaks in the Future Forest: Extending an Existing Model to Inform Climate Change Adaptation1 Final Report November 2014 SNPLMA (Southern Nevada Public Lands Management Act) project (P086) PI: Robert M. Schellera Co-PIs: E. Louise Loudermilka,b, Matthew D. Hurteauc, Peter J. Weisbergd, Agency Collaborator: Carl Skinnere Project Researchers: Alec M. Kretchuna,f, Soumaya Belmecherig Final Report developed by E.L. Loudermilk and A.M. Kretchun aPortland State University Environmental Science and Management Department Portland State University PO Box 751, Portland, OR 97207 USA bUSDA Forest Service, Center for Forest Disturbance Science Southern Research Station 320 Green St., Athens, GA, 30606 USA cPennsylvania State University Department of Ecosystem Science and Management 306 Forest Resources Building, University Park, PA, 16802 dUniversity of Nevada, Reno Department of Natural Resources and Environmental Science 1664 N. Virginia St., Mail Stop 186, Reno, Nevada 89557 USA eUSDA Forest Service Pacific Southwest Research Station 3644 Avtech Parkway, Redding, CA 96002 USA fUSDA Forest Service Pacific Northwest Research Station 1220 SW 3rd Ave, Portland, OR 97204USA gPennsylvania State University Department of Meteorology 503 Walker Building, University Park, PA, 16802 1 Citation: Loudermilk, E. Louise; Kretchun, Alec M.; Scheller, Robert M.; Hurteau, Matthew D.; Weisberg, Peter J., Skinner, Carl; Belmecheri, Soumaya. 2014. Drought stress and bark beetle outbreaks in the future forest: Extending an existing model to inform climate change adaptation. Final Report for Southern Nevada Public Lands Management Act project P086, Pacific Southwest Research Station, Tahoe Center for Environmental Studies, Incline Village, NV. Table of Contents Key Findings................................................................................................................................................... 4 Introduction .................................................................................................................................................. 5 Study objectives ............................................................................................................................................ 7 Methods ........................................................................................................................................................ 8 Study Area ................................................................................................................................................. 8 Model parameterization and calibration ................................................................................................... 8 Landscape forest composition............................................................................................................... 9 Climate change data .............................................................................................................................. 9 Modeling C ............................................................................................................................................ 9 Modeling wildfires ............................................................................................................................... 10 Modeling forest fuel treatments ......................................................................................................... 10 Simulating bark beetles with BDA extension ....................................................................................... 11 ANPP data & scaling ................................................................................................................................ 12 Tree-core data: field data collection &scaling ..................................................................................... 12 Landscape simulations& analysis ........................................................................................................ 12 Experimental design for ANPP analysis ............................................................................................... 13 Tree Ring and Model Estimate Comparison ........................................................................................ 13 Long-term simulations of multiple disturbances ..................................................................................... 14 Results ......................................................................................................................................................... 15 ANPP Scaling analysis .............................................................................................................................. 15 Tree ring ANPP estimates .................................................................................................................... 15 LANDIS-II Bark beetle modeling ............................................................................................................... 15 C trajectories ....................................................................................................................................... 16 Species composition shifts .................................................................................................................. 16 Likelihood of continued C sequestration ............................................................................................. 17 Fire behavior ....................................................................................................................................... 17 Bark beetle outbreak dynamics ........................................................................................................... 18 Discussion .................................................................................................................................................... 19 ANPP scaling analysis............................................................................................................................... 19 LANDIS-II bark beetle & fuel treatment modeling ................................................................................... 20 2 Bark Beetles and C ............................................................................................................................... 20 Ecological Mechanisms........................................................................................................................ 20 Climate Change Implications ............................................................................................................... 21 Uncertainties ....................................................................................................................................... 21 Management Implications ................................................................................................................... 21 Policy Implications ............................................................................................................................... 21 Acknowledgements ..................................................................................................................................... 22 Figures ......................................................................................................................................................... 23 References................................................................................................................................................... 38 APPENDIX .................................................................................................................................................... 43 3 KEY FINDINGS In the final report of the first phase of the project (P049, Loudermilk et al. 2012), we presented key findings associated with simulated interactions among future climate, forest growth, C sequestration, wildfires (including changing ignition patterns), and fuel treatments in the Lake Tahoe Basin (LTB). Here, we present more in-depth findings concerning effects from bark beetles and drought, future effectiveness of fuel treatments, as well as results from the ANPP scaling analysis. Simulated interactions with bark beetles, climate, and fuel treatments Under baseline climate, bark beetles do not significantly reduce total ecosystem carbon over the simulated century beyond the reductions cause by fuel treatments Projected increases in bark beetle activity in the A2 climate (high emissions scenario) generally increased C emissions due to heightened tree mortality in the latter half of the century compared to simulations assuming a contemporary climate. In addition, these outbreaks caused greater uncertainty in forest composition and carbon dynamics. Bark beetle outbreaks and wildfire activity did not overlap significantly in our simulations, regardless of climate scenario applied. This is in large part due to wildfires generally occurring near the WUI, and BB outbreaks occurring basin-wide, including more remote and high elevation areas. In the A2 climate, fuel treatments remained effective at reducing wildfire activity, but were not particularly effective at reducing bark beetle outbreaks, due to the great magnitude of bark beetle outbreaks and their widespread distribution throughout the basin. A Log Odds Ratio analysis on Net Ecosystem C Balance indicated that bark beetles significantly increased the probability of the landscape becoming a C source after mid-century with climate change. The effects of bark beetles and fuel treatments had compensatory effects on species interactions, where, e.g., fire tolerant or less targeted species (by beetles or management) regenerated in areas affected by these disturbances. Under high emissions, this compensatory capacity was reduced in the latter half of the century, when effects from bark beetles were severe and forest recovery lagged behind outbreak frequency and intensity. The beetle outbreak area varied by species, with the fir engraver causing the most damage. The damage potential of the fir engraver was reduced when fir populations declined from significant beetle kill under high emissions. Under high emissions climate, fir engraver outbreaks were 35% greater in area when fuel treatments were not simulated, indicating that simulated fuel treatments have the capacity to reduce outbreak size of fir engraver. ANPP scaling analysis Two significant drought periods occurred during the historical study period of 1987-2006; these droughts caused reductions in median ANPP when estimated by both increment coring and LANDIS-II, particularly in the sites with the highest potential productivity. Moisture sensitivity and bark beetles both caused reductions in aboveground productivity, moisture sensitivity via slowed growth and bark beetles via mortality, particularly in older larger tree cohorts. LANDIS-II captured the underlying dynamics of tree growth well. Scenarios that included bark beetle outbreaks and moderate moisture sensitivity matched the increment core productivity estimates best. 4 INTRODUCTION Lake Tahoe Basin (LTB) managers are faced with managing a forest disturbed by multiple and interacting forces (climate, insects, drought, wildfire) that are highly dynamic across space and time. Climate change is of particular concern because of the overarching impacts on Basin wide processes, such as forest productivity(Battles et al. 2008), resilience(Millar et al. 2007), carbon (C) sequestration potential (Hurteau and Brooks 2011), fire and insect risk(Coops and Waring 2011), amongst others (e.g., water quality). Throughout the western U.S., increasing temperature and decreasing precipitation have been identified as the root cause of increasing tree mortality(van Mantgem et al. 2009). Climate projections for California however, suggest little change in annual total precipitation until late-century, although temperatures are expected to continue increasing(Cayan et al. 2008). Increasing temperature amplifies drought stress in plants because of the direct effects on evapotranspiration rates(Seager et al. 2007). In the dominant Sierran mixed-conifer forest, future climate is projected to decrease productivity and increase mortality (Battles et al. 2008). Large-scale droughts are already occurring in some systems. In southwestern piñonjuniper woodlands, prolonged water stress has increased susceptibility to other disturbance agents and large mortality events(Breshears et al. 2009). A five-fold increase in drought related mortality is expected to occur in southwestern U.S. forests by the end of the century(Adams et al. 2009). Across the U.S., more than 70 million acres of forested land are at risk of mortality from 26 different insects and diseases(US Department of Agriculture 2004). Bark beetles (e.g., Dendroctonus spp., Ips spp., Scolytus spp.) in particular, have been an especially important factor contributing to tree mortality in the U.S.(Fettig et al. 2007). The link between bark beetle outbreaks and drought is significant (Guarín and Taylor 2005, Dobbertin et al. 2007). This is especially true in areas that have been fire suppressed and have high tree densities; the stress of resource competition from surrounding trees coupled with warm temperatures creates a suitable environment for beetle infestation(Parker et al. 2006), notwithstanding improved conditions for longer or multiple insect life cycles and dispersal capacity(Hicke et al. 2006). Beetle outbreaks are expected to increase in the near future due to prolonged dry seasons and increased drought stress caused by climate change. It is projected that bark beetle infestation risk will increase by 2.5-5 times in western forests by the end of the 21stcentury (Gan 2004, Hicke et al. 2006). The coniferous forests of the western U.S. have already experienced unprecedented levels (> 20 million ha annually, including pathogens (Dale et al. 2001)) of bark beetle infestations driven by the compounding factors of fire suppression, high tree density, increasing temperatures, and extended drought(Hicke et al. 2006, Fettig et al. 2007). This is also evident in the LTB where an extended drought period (1986-1992) caused landscape-level drought and bark beetle driven mortality patterns, damaging >260,000 m2of tree basal area (Macomber and Woodcock 1994). During this drought period, thirty percent of the conifers in the LTB were killed by bark beetles, with concentrated mortality rates as high as 80% in some areas. The intersection with the Wildland Urban Interface increased fire risk (i.e., higher dead fuel loads) and reduced the forest’s aesthetic quality (Macomber and Woodcock 1994, Donaldson and Seybold 1998a, b). 5 The excessive fuel accumulation resulting from nearly a century of fire suppression(Stephens and Ruth 2005)and several decades of climate warming(Westerling et al. 2006) has also caused increasingly greater area burned in U.S. National Forests. The amount of U.S. forest lands affected by wildfires and phytophagous insects are similar and often overlap in area(Hicke et al. 2006). The overlap is due to the feedbacks associated with tree stress and fuel loading. Even if trees survive a fire, they are stressed and become more susceptible to beetle infestation. An area not recently burned may still experience bark beetle attack (e.g., from drought stress) and will subsequently increase dead fuel loading in outbreak areas(Parker et al. 2006). Where prescribed burning is applied and fire intensities are lowered, trees may be fire-stressed and may therefore become prone to subsequent bark beetle invasion. Specific to the LTB, (Bradley and Tueller 2001)found that bole charring, duff consumption, and crown scorch were significant predictors of beetle attack after a prescribed burn and over 24% of the trees were infested in prescribed burn plots. These feedbacks between bark beetle outbreaks and wildland fire are, however, dependent on disturbance intensity, extent, timing, and location, as well as tree physiological response to each disturbance. With increasing temperatures in the west, these multifaceted tree stressors weaken individual tree resistance and reduce overall forest resilience to drought, bark beetles, and wildfires. As a consequence, LTB forests may be more vulnerable to rapid change than previously assumed, with the potential for a climate regime shift to non-analogue vegetation(Hurteau and Brooks 2011). For example, if a Jeffrey pine stand experiences drought, the resulting mortality and stress may trigger subsequent insect outbreaks and/or provide more fuel for wildfire. If a large, contiguous area is similarly affected, Jeffrey pine seed rain will diminish and, if the drought continues, germination and establishment will be reduced. The result would be a large area dominated by chaparral and shrubs with substantially lower embodied C and reduced ability to continue to uptake C. Furthermore, if conditions conducive to subsequent fire occur more frequently, establishment of tree regeneration may be further limited because of the increase in fire-induced mortality(McGinnis et al. 2010). Forest resilience can however, be maintained and even enhanced through active management that reduces resource competition and that limits insect mortality and the risk of high-intensity wildfire(Fettig et al. 2007). Although climate, drought, and insects are external factors that LTB managers cannot directly control, it is nonetheless important to understand the inherent processes and projected effects on the forest. This knowledge provides the foundation for developing strategic adaptive planning, especially related to invasive field applications, such as forest thinning. In addition to reducing fire risk, forest thinning has the potential to reduce drought stress and bark beetle susceptibility(Fettig et al. 2007). Research and extension(Donaldson and Seybold 1998b) has suggested ‘integrated pest management’ techniques to mitigate bark beetle infestation, including thinning and sanitation methods. These techniques are geared towards maintaining the health of un-infested trees and the removal of infested ones. Mitigation strategies, such as fuel treatments, are a necessity in western U.S. forests, where a course of ‘no action’ is not without consequence. The clear relationship of tree density and drought stress to bark beetle 6 susceptibility (as well as fire risk) drives the management motivation to implement extensive fuel treatments in the LTB(Fettig et al. 2007). Effective forest management requires a better understanding of a) the potential for amplified drought and beetle induced mortality, b) the interactions among climate-driven processes (wildfire, drought, beetle outbreaks, forest succession), c) the effects of current and future management activities (i.e., forest treatments) on mitigating the severity of drought and insect mortality, d) additional management alternatives for maintaining forest resilience, while remaining logistically flexible given the large uncertainty about the future of the LTB. A landscape-scale approach is critical to addressing these issues – the LTB is a tightly linked system and projections beyond the next decade require an integrated, landscape-scale approach. Furthermore, understanding the long-term effectiveness of various thinning strategies is most feasibly done using a modeling approach, given the difficulty of designing and maintaining long-term field experiments that incorporate multiple disturbance types and inherent feedbacks among disturbances, succession, forest insects and pathogens, and climate change. This project used an existing modeling framework (developed for P049, Loudermilk et al. 2012) of forest landscape processes (wildfires, climate change, succession, and fuel treatments) to include effects from drought and bark beetle outbreaks. More specifically, for P049, we evaluated climate change effects on net ecosystem C balance (Loudermilk et al. 2013), as well as a suite of fuel treatment scenarios to assess treatment effectiveness for reducing wildfire C emissions and maximizing total landscape C storage(Loudermilk et al. 2014). For this project (P086), we integrated the effects of drought and insects into our estimates of C dynamics and tree community response and feedbacks over the next 100 years, b) examined model accuracy of forest productivity (i.e., ANPP) with empirical estimates, and c) examined fuel treatment effectiveness for mitigating the effects from the interacting disturbances, i.e., climate (including drought), wildfires, and bark beetle outbreaks. STUDY OBJECTIVES Our primary objectives were to evaluate climate change effects associated with drought stress (reduced forest productivity), bark beetle outbreaks, and management mitigation options across the forested landscape of the Lake Tahoe Basin, CA and NV, USA (Fig. 1, 2). In addition, we compared in situ estimates of scaled ANPP from tree-ring data with model outputs across a coincident 20-year period (1987-2006) to climate to investigate model accuracy of forest productivity, as influenced by moisture sensitivity and bark beetle outbreaks. This research was an extension of our previous SNPLMA project [P049,(Loudermilk et al. 2012)] which assessed the impacts of climate change on wildfire and above and belowground forest C dynamics (Loudermilk et al. 2013), and the effectiveness of fuel treatment options for reducing fire emissions and maintaining forest C (Loudermilk et al. 2014). This research leveraged the spatial data, model parameterization, and analysis for P049, while incorporating drought-impact growth estimates from site-level data collected from another SNPLMA project (P029, Hurteau et al. 2014). A conceptual diagram of interacting disturbances and simulation approach are in Fig. 1. 7 METHODS STUDY AREA Our study area comprised approximately 85,000 ha of forested land in the Lake Tahoe Basin (Loudermilk et al. 2013, Fig. 2). The climate is Mediterranean with a summer drought period. The basin-like topography and elevation range (ca. 1897 to 3320 m) control local temperature and precipitation patterns. The majority of precipitation falls as snow between October and May and snowpack persists into the summer, dependent on elevation. The western portion of the Basin has a higher water balance and greater productivity than the northern and eastern portions of the Basin. Mean annual precipitation on the west shore at Tahoe City, CA is 80 cm and mean annual snowfall is 483 cm. Mean annual precipitation on the east shore at Glenbrook, NV is 46 cm and mean annual snowfall is 236 cm. At Tahoe City, average January high temperature is 6 °C and the low is -6 °C. Summers are mild with an average high temperature of 26 °C in August and a low of 6 °C (Western Regional Climate Center 2012). Soils are classified as shallow Entisols or Inceptisols and the more developed soils are Alfisols. The substrate is mainly granite with ancient volcanic bedrock lining the north shore(Rogers 1974). Tree species distribution in the LTB is controlled by elevation and precipitation (Barbour 2002). The lower montane zone in the west Basin is primarily a mixed conifer forest consisting of up to six co-dominant species including white and red fir (Abies concolor, A. magnifica), incense cedar (Calocedrus decurrens), and Jeffrey, sugar, and lodgepole pine (Pinus Jeffreyi, P. lamertiana, P. contorta). The east side montane zone is dominated by Jeffrey pine, red fir, and/or white fir. The subalpine zone consists of whitebark pine (P. ablicaulis), western white pine (P. monticola), and mountain hemlock (Tsuga mertensiana). Almost 70% of the lower montane zone in the LTB was clearcut during the Comstock logging era beginning around 1870 and continuing through the turn of the century. Timber harvested from the LTB was used for shoring up mineshafts, creating timber flumes, and extending rail lines to nearby Virginia City, NV. The timber harvest and subsequent fire suppression throughout the last century has led to a shift in the age and size distribution of the forest from a characteristic old-growth canopy, with an open mid-story, to a more dense forest of younger age-cohorts (<120 years old) and more closed mid-story (Barbour et al. 2002, Nagel and Taylor 2005). This shift has allowed more surface and ladder forest fuels to accumulate and has increased fire risk(Beaty and Taylor 2008). In addition, shade tolerant trees like white fir and incense cedar have increased disproportionately over fire-adapted species like Jeffrey and sugar pine (Nagel and Taylor 2005). MODEL PARAMETERIZATION AND CALIBRATION This project (P086) is a continuation of the previous modeling efforts of project P049 (Management Options for Reducing Wildfire Risk and Maximizing C Storage under Future Climate Changes, Ignition Patterns, and Forest Treatments). As such, extensive details on model parameterization, calibration, and validation are elsewhere (Loudermilk et al. 2013, 2014), including the original project’s final report (Loudermilk et al. 2012). As such, details on initial forest composition, climate change data (including downscaling), C dynamics, wildfires, and fuel treatments are written in brief. For the methods in this final report, we extend these modeling efforts by providing full details on modeling bark beetles, as well as 8 examining model drought sensitivity and beetle outbreaks with empirical data (tree-cores) on forest growth. We used the Landscape Disturbance and Succession model LANDIS- II (v.6.0). The LANDIS-II model has been used throughout the U.S. (Scheller et al. 2011a, Scheller et al. 2011b), b) and elsewhere (Cantarello et al. 2011, Steenberg et al. 2011)for landscape climate change research (Scheller and Mladenoff 2008, Xu et al. 2009), as well as and feedbacks associated with wildfire (Sturtevant et al. 2009), fuel treatments (Syphard et al. 2011, Loudermilk et al. 2014), and insect outbreaks (Kretchun et al. 2014). LANDIS- II offers the flexibility to integrate various ecosystem processes and disturbances that interact across large spatial extents and long time periods, ideal for projecting forest succession and responses to climatic variability. Input parameters for LANDIS-II modeling for the LTB are found in the Appendix. LANDSCAPE FOREST COMPOSITION To characterize the initial forest communities across the landscape, we utilized a database and a map of age-cohorts of trees and shrubs developed for the LTB based on the Fuel Characteristic Classification System and the existing vegetation map (CALVEG) from the GIS Clearinghouse of the Pacific Southwest Region (Ottmar and Safford 2011). The resulting forest community map was coupled with Forest Inventory Analysis data from the Basin and nearby Sierra Nevada forests to provide ground estimates of species composition and age distribution by forest type (e.g., mixed conifer), similar to (Syphard et al. 2011). This map was refined to account for the largest wildfires from years 2002 to 2010. CLIMATE CHANGE DATA Study site climate projections were processed using existing downscaled Geophysical Fluid Dynamics Laboratory (GFDL) General Circulation Models (GCM) of high (A2) and moderate(B1) global CO2 emissions scenarios of climate up to year 2100, specific to the LTB(Coats et al. 2010). These downscaled daily precipitation and temperature values (12 km ) were further processed using PRISM data (Parameterelevation Regressions on Independent Slopes Model, http://www.prism.oregonstate.edu/, PRISM 30-year Normals (1971–2000), 800 m ) to provide area-weighted averages across each fire region and ecoregion. For this project (P086), we only used the high (A2) emissions scenario of future climate. For the Century and Dynamic Fire extensions of LANDIS-II (discussed below), the climate variables were updated every 5 years (e.g., 2020–2024), and influenced monthly growth, decomposition, establishment ability as well as seasonal fire weather, respectively, within each 5-year-time step. 2 2 MODELING C Ecosystem C dynamics were modeled using the LANDIS-II Century Succession extension (Scheller et al. 2011a), which is based on the original CENTURY soil model (Parton et al. 1983). This extension (hereon called ‘Century’) integrates aboveground processes of stand dynamics with C and nitrogen cycling as well as decomposition and accumulation of soil C. Pool flows and interactions with climate (e.g., effects from temperature and changes in soil moisture), as well as further parameter descriptions and examples of calibration procedures for Century, are found elsewhere(Scheller et al. 2011a, Scheller et al. 2011b)including the Century User’s Guide (http://www.landis-ii.org/exts/Century-succession). Six target model outputs were chosen to calibrate and validate Century parameters based on available literature and expert opinion. These include aboveground net primary productivity (ANPP), NPP, net ecosystem 9 production (NEP), aboveground live biomass, soil organic C (SOC), and soil inorganic nitrogen (Mineral N). Full details on Century parameterization, calibration, and validation are found elsewhere (Loudermilk et al. 2012, 2013, 2014). MODELING WILDFIRES The Dynamic Fire and Fuels extension (hereon called ‘Dynamic Fire’) simulates fire behavior and fire effects based on a parameterized fire regime and forest fuel types as well as input fire weather and topography (Sturtevant et al., 2009). The LTB was divided into three fire regions, representing distinct fire regime characteristics, and associated with varying elevation, climate, and ignition density estimations. Each fire region had unique characteristics associated with ignitions and fire weather that influenced fire ignition and spread. Details on fire region classification and parameters, as well as calibration based on local wildfire data are found elsewhere (Loudermilk et al. 2012, 2013, 2014). MODELING FOREST FUEL TREATMENTS Fuel treatment (i.e., forest thinning) prescriptions and scenarios were developed using an expertknowledge approach similar to Syphard et al. (2011) and (Collins et al. 2010), where agency personnel at the federal, state, and local level provided information on fuel treatment implementation and tactics, including treatment efficacy. From these communications, we developed fuel treatment strategies that represented their current and anticipated management activities in the LTB at the stand to landscape level. Full details on fuel treatment prescriptions, fuel treatment areas and stand selection approach, as well as fuel treatment scenarios are in Loudermilk et al. (2012, 2014). Fuel treatments were simulated using the Leaf Biomass Harvest extension (v. 2.0.1) of LANDIS-II, that has been successfully used in other fuel treatment (Scheller et al., 2011b; Syphard et al., 2011) and forest harvesting studies (Scheller et al., 2011a). The LeafBiomass Harvest extension was designed specifically to link with Century to simulate removal of aboveground live leaf and woody biomass of designated species age-cohorts within selected areas and with Dynamic Fire to simulate post-treatment effects on firebehavior and subsequent fire effects. Similar to Syphard et al. (2011), we used this extension to simulate forest thinning from below (i.e., fuel treatments), where much of the older cohort biomass was left intact. Simulated fuel treatments represented the basic prescriptions deployed in the LTB including hand and mechanical thinning of understory and mid-story trees up to specified diameter limits. The light thinning prescription (Syphard et al., 2011) was designed to represent hand-thinning of understory and mid-story trees up to 14 in. (35.6 cm) in diameter. The moderate thinning prescription (Syphard et al., 2011) was designed to represent a more intense mechanical thinning of understory and mid-story trees up to 30 in. (76.2 cm) in diameter. Fuel treatments were simulated within three designated treatment areas (Marlow et al., 2007): the defensible space, defense zone, and extended wildland urban interface (WUI). These treatment areas were further divided into treatment stands, representing an area that was completely treated if chosen based on selection criteria and treatment interval. Stands were selected for treatment based on estimated ‘‘fire hazard’’. The ‘‘fire hazard’’ stand selection method (found in v. 2.1.1 of the Base Harvest extension) was created based on a management approach of stand selection that assesses forest and fuel characteristics that describe a stand’s latent wildfire risk. Although multiple fuel treatment 10 scenarios were developed under the original project (P049), we only used one fuel treatment scenario for this project; A ‘Continued Intensity’ scenario was designed to apply fuel treatments on a designated 15 rotation period continuously through time through all three management areas. SIMULATING BARK BEETLES WITH BDA EXTENSION Bark beetle outbreaks were simulated using the Biological Disturbance Agent (BDA) extension for LANDISII (Sturtevant et al. 2004). A conceptual diagram representing the interactions between climate, the Century extension, and the BDA extension for modeling bark beetles effects on C dynamics are in Fig. 3. This extension simulates tree mortality resulting from outbreaks of insects and disease that are significant enough to influence forest succession, fire disturbance, or harvest disturbance at the landscape scale. Outbreaks are probabilistic at the site level, where the probability of a cell being disturbed is based on the available hosts at that site. Individual host species are ranked as one of four possible levels: primary, secondary, minor, and non-host. These four categories are described as species and age categories. For example, in the LTB, the Jeffrey pine beetle (Dendroctonus jeffreyi) is an obligate pest of Jeffrey pine, though it prefers older cohorts (>60 years, primary host) much more than younger cohorts (<20 years old, minor host). These host categorizations help determine ‘site vulnerability’ of a given site(Sturtevant et al. 2004). The severity of outbreaks that occurs is a direct function of site vulnerability, classified as one of three categories: light, moderate, and severe. ‘Light’ outbreaks kill all vulnerable cohorts; ‘moderate’ outbreaks kill all tolerant and vulnerable cohorts; and ‘severe’ outbreaks kill resistant, tolerant, and vulnerable cohorts. Outbreaks are synchronous, and the user can bound the severity by defining a minimum and maximum possible outbreak level. For this study, we were matching a historical outbreak considered to be severe compared to others in recent history; therefore the outbreak level within the BDA was restricted to only the ‘severe’ level. The BDA extension reduces site and landscape ANPP through mortality of affected cohorts rather than direct reductions in cohort growth rates. Three bark beetle species were modeled: the Jeffrey Pine Beetle (Dendroctonus jefrreyi, JPB), the Mountain Pine beetle (Dendroctonus ponderosae, MPB), and the Fir engraver beetle (Scolytus ventralis, FE). Although there are other beetles active in the area (e.g. Red turpentine beetle (Dendrocotnus valens)), these three beetles are responsible for the vast majority of the recorded damage in the Basin and there is very little overlap in host species between the three. Empirical data from the literature and expert opinion was used to determine host species and ages most preferred by each of the three modeled beetle species (Kretchun et al. in review Ecosystems). JPB and FE are limited in their primary host selection (Jeffrey pine and red/white fir respectively), whereas MPB is much more of a generalist, impacting a variety of pine species across the basin. Beetle dispersal is modeled within BDA, defined at an annual rate (m yr-1). This is unique for each species, and reflects the physiological differences in each species’ ability to spread across a landscape. A widespread outbreak of bark beetles occurred in the region, concurrent with a severe drought that began in 1988. USFS Aerial Detection Survey (ADS) maps of the basin indicated >15,000 ha of damaged area. These maps attribute damage to specific agents, meaning that area affected by beetles could be identified on an annual basis. Maximum damage captured by these maps for each beetle species was: mountain pine beetle (933.01 ha), Jeffrey pine beetle (3,126.2 ha), and Fir engraver beetle (11,726.30 11 ha). From the ADS flyover maps and expert opinion (Bulaon, personal communication), total acreage affected by Fir engraver beetle required some correction for two reasons. Fir engraver beetles are typically less aggressive (lower mortality percentage amongst affected stands) than species of genus Dendroctonus and they are also more restricted to areas dominated by their hosts. This means that areas identified by the flyover maps may be overstating the total area affected by these beetles. To correct for this possible overestimation, stand dominance by tree species was determined using biomass estimates within a 5 ha moving window across modeled sites. Stands that contained >75% red and white fir were within an identified outbreak zone were determined to have likely been impacted, which could then be totaled to calculate total area affected by fir engraver beetle. This correction factor was not applied to area affected by the two species of Dendroctonus beetles, as it is believed the flyover maps area reasonable estimates of damage. Therefore, area affected by each beetle species was calibrated to mountain pine beetle (933.01 ha), Jeffrey pine beetle (3,126.2 ha), and fir engraver (8,794.5 ha). Total area affected in the peak outbreak year of 1991 within the model was 10,417 ha, compared to remotely sensed estimates of 15,783 ha. Outbreaks were simulated from 1990-1998, matching flyover maps and other records. The outbreaks were deterministic in length, lasting for 8 years. ANPP DATA & SCALING TREE-CORE DATA: FIELD DATA COLLECTION &SCALING Field data were collected at two to four plots in each of 21 creek drainages ranging from 1900-2200m elevation (Fig. 4). Forest structural attributes were measured using a nested design in which all trees > 5cm dbh were measured in a 1/50th ha subplot, all trees > 50 cm dbh were measured in a 1/10th ha subplot, and all trees >80 cm dbh were measured in a 1/5th ha plot. Within each plot two to three individual trees were selected for coring from the five smallest and five largest individuals (Hurteau et al. 2014). Annual ring width was measured to the nearest 0.001 mm using Windendro and error prone cores were re-measured using a Unislide TA measuring system (Velmex, Bloomfield, NY). Raw tree-ring widths were then used to calculate the radius of each tree to account for cores that missed pith. The inferred radius was then used to estimate diameter at breast height (DBH) for each tree at each annual increment. Annual DBH values were then used to estimate annual biomass production using allometric equations from(Jenkins et al. 2004). To scale tree-level estimates of annual biomass production to the plot level, individuals measured at DBH, but not cored, were matched with cored individuals of the same species and similar diameter from the nearest plot. Annual growth measurements from the cored individuals and genus-specific allometric equations were then used to estimate annual biomass production for the trees that were not cored. Plot level estimates of annual ANPP were then scaled to the hectare level using the appropriate scaling factor (e.g., a tree > 50cm DBH represents 50 trees ha-1) from the nested plot design. LANDSCAPE SIMULATIONS& ANALYSIS We selected the period from 1987-2006 because it had the highest number of available tree core samples and it captured two of the most significant droughts of the latter 20 th century which were accompanied 12 by large bark beetle outbreaks. Over the study period, maximum summer temperature ranged from 14 to 18.5⁰C, and minimum January temperature ranged from 0.8 to -4.2⁰C. The two drought periods occurred from 1988 to1992 and 2001 to 2002, with the earlier of these two droughts generally being regarded as more severe. The drought of 2001 was characterized by below average wintertime (December-February) precipitation and slightly above average temperatures, whereas the drought of 1988 was characterized primarily by extreme below average wintertime precipitation (Fig. 5). We chose the Palmer Drought Severity Index (PDSI) (Palmer 1965) to identify distinct drought periods for the years of study. PDSI is a supply-demand model of soil moisture, which has been widely applied as a general measure of drought(Zhao et al. 1986, Nicault et al. 2008). Values above zero indicate above average moisture; values below zero indicate a drier period. More extreme values signify increased levels of wetness or drought, respectively. The two drought periods on record for LTB for our study period are reflected in the low PDSI (-2.08 and -2.2 respectively) values for 1988 and 2002 (Fig. 5). EXPERIMENTAL DESIGN FOR ANPP ANALYSIS Using LANDIS-II, we ran 20-year simulations with historical climate from 1987-2006. Monthly temperature and precipitation values for 1986-2006 were from the PRISM dataset(Daly et al. 1997) for the LTB. There were no records or physical evidence in the field of any fire (wildfire or prescribed burning) or forest thinning within the study sites where tree-cores were collected. Although there may have been some forest thinning operations and wildfires that occurred in the LTB during that timeframe, they were not simulated to be congruent with disturbances evident at the field study sites. Therefore, our fullfactorial design included two levels of conifer drought sensitivity (moderate and high sensitivity) and two levels of bark beetle outbreak (with and without beetles). Because the moisture sensitivity parameters are not empirically derived values, the two levels of moisture sensitivity were determined by incrementally increasing the DroughtB parameter by the minimum amount anticipated to have an effect (0.1). The scenarios were as follows: low moisture sensitivity with no beetles (LowM-NoBB), low moisture sensitivity with beetles (LowM-BB), high moisture sensitivity with no beetles (HiM-NoBB), and high moisture sensitivity with beetles (HiM-BB). ANPP values from these four model scenarios were then compared with empirical ANPP values. TREE RING AND MODEL ESTIMATE COMPARISON In order to make meaningful comparisons of model and tree-ring estimates of ANPP throughout the study period, we bootstrapped both the empirical and modeled ANPP estimates using 500 draws from each population to calculate the median and 95% confidence intervals. To visualize the relative densities of ANPP between empirical and modeled data, kernel density estimates were constructed for each year of the 20-year study period. Kernel density estimation is a non-parametric approach to estimate a probability density function of a random variable; in this case, ANPP (Gagolewski 2013). Within these plots, each year is represented by density distributions for empirical and modeled ANPP. Increasing yvalues (i.e. ‘peaks’ in the distributions) correspond to greater frequency of ANPP. For instance, if the highest point, aka ‘peak’, of a distribution lay at 200 g C m 2 on the a-axis, that is the most frequent ANPP value found across the study area. All statistical and graphical analyses were done using the R statistical software package (R Core Team 2013). 13 LONG-TERM SIMULATIONS OF MULTIPLE DISTURBANCES We used a scenario approach to assess the multiple climate-disturbance interactions and their influence on long-term forest change and ecosystem C dynamics. Each scenario was simulated for 100 years (20102110). The landscape was simulated at a 1 ha spatial resolution, and each scenario was replicated five times to account for the stochastic variation (due to climate, wildfire, insects, treatments, and regeneration) among replicates. These scenarios of climate and disturbance were run in an iterative process of incorporating bark beetles and fuel treatments: High emissions climate and base climate (no change in climate) were simulated without effects from bark beetles or fuel treatments Bark beetles were simulated using the base and high emissions climate Fuel treatments were simulated using the base and high emissions climate Bark beetles and fuel treatments were simulated together using the base and high emissions climate 14 RESULTS Here we present results on the ANPP scaling analysis of 20 year (1987-2006) tree-core data and coincident LANDIS-II model runs, as well as LANDIS-II projection modeling of climate and bark beetle impacts over the next 100 years (2010-2110). ANPP SCALING ANALYSIS TREE RING ANPP ESTIMATES Median ANPP derived from tree-cores was generally below 200 g C m-2 (Fig. 6), with increasing variance over the 20-year study period. Over the simulation period, median ANPP values generally increased; from 118.5 g C m2 in 1987 to 207.1 g C m2 in 2006, with temporal fluctuations throughout the 20 year period. Following two drought events median ANPP dropped from 118.5 g C m2 to 93.26 g C m2 (years 19871988) and from 229.4 g C m2 to 150.15 g C m2 (years 2000-2001). Changes in median ANPP values closely followed changes in PDSI and total winter precipitation (Fig. 5). Simulations that included bark beetle presence (LowM-BB and HiM-BB) experienced declines in both aboveground biomass and total ecosystem C. Aboveground biomass was reduced by 9.7 ± 0.09% on average for the two scenarios and total ecosystem C (all C pools combined including live, detrital, and soil) was reduced by 8.59 ± 0.085%. Simulated median ANPP values for the moderate drought sensitivity with beetles (LowM-BB) scenario and high drought sensitivity (Hi) scenario fell within the bootstrapped confidence intervals of the empirical data within 16 of the 20 years (Fig. 6). The LowM-BB and HiM-noBB median ANPP values showed a consistently closer match than the LowM-noBB and HiM-BB scenarios to the empirical values in a majority of years. Agreement between all model scenarios and the empirical data was lowest in the early simulation years (1988-1993). A sharp decline in tree ring and all modeled median ANPP values occurred in 2001 (Fig. 6; 229.4 g C/m2 to 150.2 g C/m2 in tree ring scenario), which corresponded with a similarly steep drop in PDSI (Fig. 5; -0.07 to -1.75). Confidence intervals for the empirical data were much larger than those of any of the model scenarios, likely because of the discrepancy in sample size between the empirical (n=41) and modeled data (n≈31,000 grid cells). Kernel density distributions illustrated similar temporal ANPP values among all model scenarios and empirical data (Fig. 7). In years of severe drought (1988, 2001), ANPP values in the empirical data rarely exceeded 400 g C m2; yet during years with more favorable growing conditions (more precipitation, less beetle kill), ANPP values above 400 g C m2 were more common. LANDIS-II BARK BEETLE MODELING We estimated PDSI for a suite of 6 high-emission GCMs and found similar patterns in all, though projections were highly variable, as expected. Values below the zero line indicate a dry period, with more severe droughts indicated by more negative numbers. Regular dry and wet periods were commonly projected by all GCMs. Generally speaking, average PDSI (indicated by the smoothed black spline in Fig. 8) is expected to remain steady until about mid-century. From ~2050 on, projected PDSI shows a slow, 15 steady decline. The GCM chosen for our simulations, the GFDL A2 scenario, was among the most extreme with regards to the increasing frequency and persistence of drought conditions, as defined by PDSI. Using this GCM, there are several notable dips in PDSI towards the end of the century. C TRAJECTORIES In the baseline climate, simulated total ecosystem C, including above and below-ground live C, detrital C, and soil C, was reduced with fuel treatments and bark beetles (Fig. 9), with bark beetles creating more uncertainty (more variability between simulation replicates). This had similar effects within management areas and simulated total ecosystem C was strongly affected by the combined effects of climate change and bark beetles (Fig. 10). Total ecosystem C increased over the 100 year simulations regardless of climate or disturbance scenario. Bark beetles (with wildfire and harvesting) and without climate change had only a minor effect on total C. Total C was generally lower under high emissions climate, whether bark beetles were included or not. However, when bark beetles were included under high emissions climate, the persistent drought conditions created the conditions necessary for large and repeated bark beetle outbreaks and total C was reduced by ~25% after 2070 . SPECIES COMPOSITION SHIFTS Baseline climate The effects of climate, fuel treatments, and bark beetles differed according to species, particularly with regards to the two species of primary management concern, Jeffrey pine and white fir (Fig.11, 12). The effect of bark beetles on the competitive balance between Jeffrey pine and white pine, indicates that without climate change, bark beetles would only have very small effects. As white fir is reduced due to thinning and prescribed fire, Jeffrey pine increases, a compensatory mechanism that maintains forest productivity (Loudermilk et al. 2014). The primary impact of bark beetle outbreaks on Jeffrey pine under baseline climate is an increase in variability of aboveground biomass. Though the competitive dynamics between Jeffrey pine and white fir are not noticeably changed by bark beetle outbreak under baseline climate, bark beetle outbreaks do have an effect on white fir biomass. This is true when focusing on individual management units (Fig. 11, 12), between which fuel treatment prescriptions are highly variable, as well as landscape averages (Fig. 13). The primary impact of bark beetles on white fir under baseline climate is a reduction of aboveground biomass via outbreak related mortality. This is true in all management units, but the reductions are proportional to overall white fir biomass. For instance, the largest white fir biomass reductions are seen in the Defense zone and extended WUI, the two management units with the most white fir. High Emissions climate Under high emissions climate, the impacts of bark beetles and fuel treatments on the two species of primary management focus, white fir and Jeffrey pine, are more pronounced and variable than under baseline climate. When looking at landscape averages, Jeffrey pine shows increases in biomass with fuel treatments (Fig. 14). These gains are severely reduced by bark beetle outbreaks in the second half of the simulated century, as outbreaks become more frequent and severe. Variability is also significantly increased. This same pattern is borne out when focusing on individual management units (Fig. 15, 16). As biomass of Jeffrey pine increases through thinning and prescribed fire, the reductions in biomass caused 16 by bark beetles, particularly the Jeffrey pine beetle, become larger as well (Fig. 15). When fuel treatments are implemented however, there is still an upward trajectory of Jeffrey pine biomass after beetle outbreaks suggesting an ability to recover after beetle-caused losses. White fir showed similar patterns to those of Jeffrey pine - under high emissions climate, bark beetle outbreak decreased aboveground biomass in all management units as well as across the entire landscape (Fig. 16). Unlike Jeffrey pine however, bark beetle outbreak accelerated the white fir biomass losses caused by the implementation of fuel treatments. Outbreaks, primarily from the fir engraver beetle, caused significant biomass reductions with an attendant increase in variability. There was less evidence of post-outbreak recovery in white fir suggesting a greater uncertainty in C stability and continued sequestration in white fir. LIKELIHOOD OF CONTINUED C SEQUESTRATION The probability of the LTB forests becoming net C sources to the atmosphere was sensitive to bark beetles and climate change. A Log Odds Ratio (LOR) above zero indicates there is a higher likelihood of the landscape switching from a net C sink to a net C source in any given year during that particular quarter century when comparing two scenarios (Fig. 17). Both climate scenarios indicate the odds of switching from a C sink to a C source are higher with bark beetles, although the uncertainty and temporal patterns differ between the two. Under baseline emissions, there is some temporal change in the likelihood of a sink/source switch. The LOR for NECB indicates that bark beetles significantly increase the probability of the landscape becoming a C source after mid-century with climate change. However, bark beetles increase the probability in the first quarter century under baseline climate, in part due to the higher PDSI under our chosen climate projection. FIRE BEHAVIOR In order to estimate the effect of bark beetles as compared to climate change on area burned, we conducted an ANOVA of total area burned (ha) across all years. Neither insects nor climate change were significantly correlated with total area burned (p = 0.19 and 0.12, respectively) (see also Loudermilk et al. 2013). Mean area burned in response to fuel treatment and bark beetle outbreaks was also analyzed by year, climate scenario, and management unit. Patterns of area burned were similar under both baseline and high emissions climate scenarios (see also Loudermilk et al 2014). Both the defensible space and defense zone, the two units with the most active fire management programs due to their proximity to the wildland urban interface, burned around 200 ha yr-1 on average with no fuel treatment (Fig. 18, 19). When fuel treatments were simulated, average hectares burned dropped significantly, sometimes into the single digits per year. This pattern did not significantly change with the scenarios that included bark beetle outbreaks. This is true of years with and without active outbreaks on the landscape. Fire and bark beetles generally did not spatially overlap often. At its maximum across all years and replicates, fire only occurred on 8% of sites that had experienced a bark beetle outbreak in the prior 15 years. 17 BARK BEETLE OUTBREAK DYNAMICS The area of defoliation varied by bark beetle species with fir engraver defoliating the largest area (Fig. 20). The differing area of outbreaks by beetle was largely a result of the prevalence of that beetle’s preferred hosts. Jeffrey pine beetle outbreaks affected the second most cumulative area, followed by mountain pine beetle with the least area affected. This followed our calibration and the general pattern held across all model scenarios. Fir engraver outbreaks were 35% higher when fuel treatments were not simulated (data not shown), indicating that simulated fuel treatments have the capacity to reduce outbreak size of fir engraver. 18 DISCUSSION ANPP SCALING ANALYSIS Productivity is influenced by a number of biotic and abiotic factors in the conifer forests of the Sierra Nevada. Available soil moisture drives tree growth and ANPP(Dolanc et al. 2013), notwithstanding the effects from natural disturbances (bark beetle outbreaks, wildfire), as well as land-use legacies (past clear-cutting) and management (forest thinning for fuels reduction). In our study, we estimated ANPP from tree-ring data and scaled these tree-level data to a per unit area basis. These validation data were similar to simulation runs of ANPP, illustrating comparable distributions of ANPP within and between years in response to drought and insect attack. While empirical estimates of ANPP are useful for quantifying inter-annual variability as a function of both biotic and abiotic factors, isolating the contribution of individual factors that can occur simultaneously presents a challenge. Using model simulation to isolate the effects of different influential factors on ANPP can improve our understanding of the mechanisms driving a particular empirical response. The two scenarios that most closely approximated the empirical ANPP data were the ‘LowM-BB’ and ‘HiM-noBB’ scenarios (Fig. 6). Although both scenarios fell within the empirically-derived ANPP confidence intervals, a fundamental difference in approach and inherent feedbacks exists between the two – climate alone drives ANPP patterns in the ‘HiM-noBB’ scenario, whereas climate and beetle-induced mortality explained these patterns in ‘LowM-BB’. Tree mortality was the primary effect of simulated bark beetle outbreaks, removing cohorts of susceptible tree species and reducing ANPP. This mortality is captured within the LowM-BB scenario, where median ANPP declines are seen in years of high bark beetle activity (e.g. 1993; Fig. 6). However, the HiM-noBB scenario ignores bark beetle mortality, explaining intra-annual ANPP patterns only through physiological responses to climate. The more conservative drought sensitivity and bark beetle scenario (LowM-BB scenario) was a better representation of the coupled processes affecting forest productivity during this timeframe because of the clear biological link of drought and beetle attack(Guarín and Taylor 2005, Hebertson and Jenkins 2008, Creeden et al. 2014)and the prevalence of bark beetles in the LTB(Bradley and Tueller 2001, Walker et al. 2007, Egan et al. 2010). Given the occurrence of beetle outbreaks, excluding bark beetles and driving the decline in ANPP with higher drought sensitivity (e.g. HiM-noBB) less accurately represents realistic feedbacks in the system; a dynamic that is especially important for long-term simulations of climate-forest dynamics, where drought-induced bark beetle outbreaks are common. Focusing more specifically on individual bark beetle dynamics within our model scenarios, the Fir engraver beetle affected the largest area across the LTB, resulting in widespread mortality of their preferred hosts, white and red fir. This is consistent with remotely sensed data from the study period (USDA Forest Service Pacific Southwest Region 2013). The large reductions in ANPP (Fig 5; -79.2 g C m-2 in year 2001) highlight the sensitivity of these fir-dominated communities to drought conditions. This also highlights the additive impacts of disturbance and climate on fir dominated areas – many of the plots with the highest potential ANPP are highly stocked white fir stands, which are highly productive and prolific seeders, but are most sensitive to drought conditions compared to most other species in the LTB(Hurteau et al. 2007, Earles et al. 2014). 19 Although we explored two processes that influence ANPP at multiple scales, many critical processes were excluded by design or necessity. Wildfires and forest thinning were not included in our simulations because there was no evidence of recent fire or thinning practices within the stands selected for tree coring. As in much of the western US, the hydrology of the LTB is snowpack driven; melting snow is responsible for gradually recharging soil moisture in the spring and summer. As such, there is an inherent lag between snowfall and soil water availability. Although Century simulates spring snowmelt, the underlying hydrology model within Century does not fully capture long-term snowmelt dynamics into the summer months. Previous comparisons of snow hydrology models have revealed that representing snowpack dynamics within forested systems is particularly challenging; for example, there can be major differences in model performance between open and closed sites(Rutter et al. 2009). Though these issues persist, forests play an essential role in snow retention and therefore forest landscape models such as LANDIS-II would benefit from the inclusion of a prolonged seasonal snow melt. LANDIS-II BARK BEETLE & FUEL TREATMENT MODELING BARK BEETLES AND C Bark beetles have the capacity to substantially reduce ecosystem C storage capacity and can cause NECB to become negative (a C source) for any given year (Bright et al. 2012, Hicke et al. 2012a). Our simulations suggest that the probability of negative NECB will substantially increase due to climate change, particularly after 2050. (Kurz et al. 2008)similarly linked mountain pine beetle outbreaks in British Columbia to negative NECB, although their projections were near term (out to 2020) and did not include an explicit link between climate and insect outbreaks. Although our simulations were limited to the LTB, the implications for the interior western US are that current trends of increasing insect-caused forest mortality(Weed et al. 2013)will accelerate, particularly after mid-century when PDSI is forecast to become increasingly negative(Coats et al. 2013). ECOLOGICAL MECHANISMS The proximal mechanism by which bark beetles reduce NECB is a reduction of the compensatory growth (sensu (Connell et al. 1984) of the system. As compared to fuels management or low-severity fire in which the decline of one dominant species, (i.e., white fir) is compensated by release and growth of another (i.e., Jeffrey pine) our climate change and bark beetle simulations suggest substantially reduced projected growth of both. This is in part due to the confluence of multiple bark beetle species, both generalists and specialists, as simulated. Although other extant trees species and/or shrubs would be expected to occupy the newly available growing space, none has the fecundity or growth necessary to maintain extant C stocks. Earles et al. (2014) also suggest that fir dominated stands subjected to drought and wildfire would exhibit greater C instability. Bark beetle susceptibility increases after stands burn by prescription(Bradley and Tueller 2001), however, our simulations suggest that such effects are and will continue to be minor at landscape-scales due to current and foreseeable levels of fire suppression and minimal prescribed burning efforts. Bark beetle damage effects on subsequent fuel conditions may also be a consideration, but again the landscape effects are likely minor because of active fire suppression and disparate location and timing between wildfires and bark beetle outbreaks (Hicke et al. 2012b). 20 CLIMATE CHANGE IMPLICATIONS Over many decades, the spatial distribution of major insects are expected to shift with climate change as their preferred host distributions shift (H. Aukema et al. 2008, Bentz et al. 2010). In the near term, there may be a substantial time lag during which the hosts may be particularly susceptible due to drought stress (Bentz et al. 2010)and an expansion of niche-availability for insects(Ward and Masters 2007). For instance, MPB, will likely experience range expansion due to climatic warming and flexibility in life history strategies, but could become limited to higher elevations in some locations (Hicke et al. 2006, Bentz et al. 2010). Over time mortality caused by insects with high host specificity would be expected to decline because of reduced host density (Negrón and Popp 2004, H. Aukema et al. 2008). Although this effect was not apparent overall in our 100 year simulations, this was evident in our simulations, where increased white fir mortality from wildfires in the high emissions climate and targeting of white fir for treatment in both climate scenarios removed large portions of the insect host, causing a decline in area affected by that host (Fig. 20). UNCERTAINTIES Our simulation approach integrated many factors critical to projecting insect outbreaks and landscape C dynamics over the coming decades, including climate and host availability. There are, however, subtle mechanisms that were not explicitly incorporated and contribute to uncertainty in our results. For example, large variation in MPB outbreaks suggest additional factors were not accounted for by climate (Creeden et al. 2014). These may include local variation in host damage caused by thinning or prescribed fires (Bradley and Tueller 2001) or wind damage or aspect (Guarín and Taylor 2005). Nor did we evaluate the full life-cycle of each beetle population (e.g., overwintering mortality) and assumed that PDSI was a sufficient proxy for the spatiotemporal scales of interest. MANAGEMENT IMPLICATIONS Managing for fuels may substantially reduce outbreak area by reducing host density and altering forest structure. Conversely, climate change and insect outbreaks may reduce the need for fuels management by eliminating a recurring source of ladder fuels, namely white fir. Similarly, areas of ponderosa pine plantations in the Modoc National forest which were thinned prior to a MPB outbreak showed significantly lower mortality rates than untreated areas (Fettig et al. 2007, Egan et al. 2010). Work in other parts of the western US has also shown reductions in beetle-caused mortality in conifer forests as a result of thinning treatments, though outbreaks that reach ‘epidemic’ levels may nullify the benefits of such preventative measures (Fettig et al. 2007). POLICY IMPLICATIONS Developing strategic approaches to managing in these future conditions is essential. Field applications used in the basin, primarily forest thinning, are intended to reduce fire risk but may also have the potential to reduce drought stress and potential beetle-related mortality. This may be particularly important in a changing climate, where higher temperatures may exacerbate these conditions (Coats et al. 2010, Loudermilk et al. 2013). Management strategies to reduce outbreaks need to be preemptive, whereby forests are thinned to create a more resilient forest structure (Egan et al. 2010). Beetles would 21 be forced to travel farther between trees and are less likely to find suitable hosts because the remaining trees are often less drought stressed. ACKNOWLEDGEMENTS We would like to thank all of the Lake Tahoe Basin agency personnel as well as others from across the region who participated in our final project workshop in May 2014. Here, we discussed results and implications for beetle management in the Basin and across the Sierra Nevada forests. We especially thank Dave Fournier and the Lake Tahoe Basin Management Unit (USDA Forest Service) for hosting the workshop and providing invaluable feedback both at the workshop and throughout the project. We thank Beverly Bulaon (USDA Forest Service) for datasets and insight to bark beetle outbreaks and management as well as for presenting at the workshop. We graciously thank all of the 15 participants in the workshop. Everyone was very engaged, where we all discussed project results, management implications, future research directions, and potential collaborations. We would especially like to thank Tiffany Van Huysen of the USDA Forest Service Pacific Southwest Research Station for her support, administration of the grant, and participation in the project. This research was supported using funds provided by the Bureau of Land Management through the sale of public lands as authorized by the Southern Nevada Public Land Management Act, and was funded in part through grant 10 DG-11272170-038 from the USDA Forest Service Pacific Southwest Research Station. Writing on this report was funded in part by the Strategic Environmental Research and Development Program, project RC-2243. We thank Portland State University, Pennsylvania State University, and University of Nevada, Reno for their administrative support. The research team of the Dynamic Ecosystems and Landscapes Lab, Portland State University was especially important for the success of this project. We thank the Forestry Sciences Laboratory, Southern Research Station, USDA Forest Service, Athens, GA, for their support. The views in this report are those of the authors and do not necessary reflect those of the USDA Forest Service or Bureau of Land Management. 22 FIGURES Figure 1: Conceptual diagram of interaction between climate, forest disturbances, and forest carbon. Figure 2: Map of the study area for the LANDIS-II project in the Lake Tahoe Basin. Forest communities are divided into ecoregions that were used in the LANDIS-II simulations: Eastside forest and woodland (34,346 ha), upper montane (21,854 ha), subalpine (6,292 ha), montane shrubland (3,541 ha), and riparian areas (3,017 ha). 23 Figure 3: Conceptual diagram of LANDIS-II components, particularly related to the interactions between climate, the century extension, and the BDA extension for modeling bark beetles effects on C dynamics in the LTB. 24 Figure 4: Study area for ANPP scaling analysis. Dark red dots indicate sites at which tree core samples were taken; the orange area represents the modeling extent used in the LANDIS-II simulations; the grey area is the entirety of the Lake Tahoe Basin. 25 Figure 5: Palmer Drought Severity Index (PDSI), total winter (December-February) precipitation, and tree core-derived aboveground net primary productivity values for the Lake Tahoe Basin. The solid line within the ANPP figure shows the median, while the shaded ribbon is constructed from the 90 th and 10th percentile of the data respectively. Figure 6: Comparison of empirical ANPP data with 2 LANDIS-II model scenarios. The ‘LowM-BB’ scenario includes low moisture sensitivity and bark beetle outbreaks, ‘HiM-NoBB’ represents high moisture sensitivity and no bark beetle outbreaks. Each line represents the median ANPP, shaded areas around each median line represent bootstrapped 95% confidence intervals, 500 draws each. 26 Figure 7: Kernel density distributions of ANPP values for empirical and 4 modeled scenarios. Peaks in the distributions correspond to more frequent ANPP values in any given year. 27 Bark Beetle Climate Change Analysis Figure 8: Projected PDSI for 6 different GCM high emissions scenarios. The red line indicates the GFDL A2 scenario that was used to simulate climate change within our LANDIS-II simulations Figure 9: Mean carbon across the Lake Tahoe Basin landscape (g C m2) for baseline (low emissions) climate. The blue envelope represents total variation among five replicates for scenarios with no thinning and no bark beetles; cyan represents thinning and no bark beetles; yellow represents no thinning with bark beetles; red represents thinning with bark beetles. 28 Figure 10: Mean carbon across the Lake Tahoe Basin landscape (g C m 2) for baseline (low emissions) climate. The blue envelope represents total variation among five replicates for scenarios with no thinning and no bark beetles; cyan represents thinning and no bark beetles; yellow represents no thinning with bark beetles; red represents thinning with bark beetles. 29 Baseline Climate Figure 11: Aboveground biomass of Jeffrey pine under baseline climate for the 4 distinct management units across the Lake Tahoe Basin. WUI stands for wildland-urban interface. 30 Baseline Climate Figure 12: Aboveground biomass of white fir under baseline climate for the 4 distinct management units across the Lake Tahoe Basin. WUI stands for wildland-urban interface. Figure 13: Mean aboveground biomass for the entire Lake Tahoe Basin under baseline climate for the two most prominent species of management concern (Jeffrey pine and white fir). 31 High Emissions Climate Figure 14: Mean aboveground biomass for the entire Lake Tahoe Basin under baseline climate for the two most prominent species of management concern (Jeffrey pine and white fir). Figure 15: Aboveground biomass of Jeffrey pine under high emissions climate for the 4 distinct management units across the Lake Tahoe Basin. WUI stands for wildland-urban interface. 32 High Emissions Climate Figure 16: Aboveground biomass of white fir under high emissions climate for the 4 distinct management units across the Lake Tahoe Basin. WUI stands for wildland-urban interface. 33 Figure 17: Log Odds ratio for NECB. Values above zero indicate the likelihood of the landscape switching from a net carbon sink to a net carbon source in any given year during that particular quarter century when comparing two scenarios. The LOR in Figure 17 compares scenarios with and without bark beetles; the solid line represents baseline climate, while the dotted line represents high emissions climate. Error bars within the figure represent the 95% confidence interval. Baseline climate is in the solid line while high emissions is in the dashed line. Both climate scenarios indicate the odds of switching from a C sink to a C source are higher with bark beetles and fuel treatments (FT), although the uncertainty and temporal patterns differ between the two. There also some evidence in the baseline of fuel treatments decreasing the likelihood of the landscape becoming a C source. Dotted line represents high emissions scenario, solid line no emissions 34 Figure 18: Mean area burned by management area under the baseline climate scenario. 35 Figure 19: Mean area burned by management area under the high emissions climate scenario. 36 Figure 20: Average hectares affected by individual bark beetle species during outbreak years. The patterns differ between fir engraver (whose preferred and only hosts are red and white fir) and jeffrey pine beetle (preferred host: Jeffrey pine)/mountain pine beetle (preferred host: generalist pines). 37 REFERENCES Adams, H. D., M. Guardiola-Claramonte, G. A. Barron-Gafford, J. C. Villegas, D. D. Breshears, C. B. Zou, P. A. Troch, and T. E. Huxman. 2009. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proceedings of the National Academy of Sciences 106:7063-7066. Barbour, M., E. Kelley, P. Maloney, D. Rizzo, E. Royce, and J. Fites-Kaufmann. 2002. Present and past OldGrowth Forests of the Lake Tahoe Basin, Sierra Nevada, US. Journal of Vegetation Science 13:461-472. Battles, J., T. Robards, A. Das, K. Waring, J. Gilless, G. Biging, and F. Schurr. 2008. Climate change impacts on forest growth and tree mortality: a data-driven modeling study in the mixed-conifer forest of the Sierra Nevada, California. Climatic Change 87:193-213. Beaty, R. M. and A. H. Taylor. 2008. Fire history and the structure and dynamics of a mixed conifer forest landscape in the northern Sierra Nevada, Lake Tahoe Basin, California, USA. Forest Ecology and Management 255:707-719. Bentz, B. J., J. Régnière, C. J. Fettig, E. M. Hansen, J. L. Hayes, J. a. Hicke, R. G. Kelsey, J. F. Negrón, and S. J. Seybold. 2010. Climate Change and Bark Beetles of the Western United States and Canada: Direct and Indirect Effects. BioScience 60:602-613. Bradley, T. and P. Tueller. 2001. Effects of fire on bark beetle presence on Jeffrey pine in the Lake Tahoe Basin. Forest Ecology and Management 142:205-214. Breshears, D. D., O. B. Myers, C. W. Meyer, F. J. Barnes, C. B. Zou, C. D. Allen, N. G. McDowell, and W. T. Pockman. 2009. Tree die-off in response to global change-type drought: mortality insights from a decade of plant water potential measurements. Front Ecol Environ 7:185-189. Bright, B., J. Hicke, and A. Hudak. 2012. Landscape-scale analysis of aboveground tree carbon stocks affected by mountain pine beetles in Idaho. Environmental Research Letters 7:045702. Cantarello, E., A. C. Newton, R. A. Hill, N. Tejedor-Garavito, G. Williams-Linera, F. López-Barrera, R. H. Manson, and D. J. Golicher. 2011. Simulating the potential for ecological restoration of dryland forests in Mexico under different disturbance regimes. Ecological Modelling 222:1112-1128. Cayan, D., E. Maurer, M. Dettinger, M. Tyree, and K. Hayhoe. 2008. Climate change scenarios for the California region. Climatic Change 87:21-42. Coats, R., M. Costa-Cabral, J. Riverson, J. Reuter, G. Sahoo, G. Schladow, and B. Wolfe. 2013. Projected 21st century trends in hydroclimatology of the Tahoe basin. Climatic Change 116:51-69. Coats, R., J. Reuter, M. Dettinger, J. Riverson, G. Sahoo, G. Schladow, B. Wolfe, and M. Costa-Cabral. 2010. Final Report: The effects of climate change on Lake Tahoe in the 21st century: meteorology, hydrology, loading and lake response. Pacific Southwest Research Station, Tahoe Center for Environmental Studies, Incline Village, NV. Collins, B. M., S. L. Stephens, J. J. Moghaddas, and J. Battles. 2010. Challenges and approaches in planning fuel treatments across fire-excluded forested landscapes. Journal of Forestry 108:2431. Connell, J. H., J. Tracey, and L. J. Webb. 1984. Compensatory recruitment, growth, and mortality as factors maintaining rain forest tree diversity. Ecological Monographs:142-164. Coops, N. C. and R. H. Waring. 2011. Estimating the vulnerability of fifteen tree species under changing climate in Northwest North America. Ecological Modelling 222:2119-2129. Creeden, E. P., J. a. Hicke, and P. C. Buotte. 2014. Climate, weather, and recent mountain pine beetle outbreaks in the western United States. Forest Ecology and Management 312:239-251. 38 Dale, V. H., L. A. Joyce, S. McNulty, R. P. Neilson, M. P. Ayres, M. D. Flannigan, P. J. Hanson, L. C. Irland, A. E. Lugo, C. J. Peterson, D. Simberloff, F. J. Swanson, B. J. Stocks, and B. Michael Wotton. 2001. Climate Change and Forest Disturbances. BioScience 51:723-734. Daly, C., G. Taylor, and W. Gibson. 1997. The Prism Approach to Mapping Precipitation and Temperature. Pages 1-4. Dobbertin, M., B. Wermelinger, C. Bigler, M. Bürgi, M. Carron, B. Forster, U. Gimmi, and A. Rigling. 2007. Linking Increasing Drought Stress to Scots Pine Mortality and Bark Beetle Infestations. The Scientific World Journal 7:231-239. Dolanc, C. R., J. H. Thorne, and H. D. Safford. 2013. Widespread shifts in the demographic structure of subalpine forests in the Sierra Nevada, California, 1934 to 2007. Global Ecology and Biogeography 22:264-276. Donaldson, S. and S. Seybold. 1998a. Bark Beetles in the Lake Tahoe Basin. Reno: University of Nevada Cooperative Extension, Reno, NV. Donaldson, S. and S. Seybold. 1998b. Thinning and Sanitation: Tools for the Management of Bark Beetles in the Lake Tahoe Basin. Reno: University of Nevada Cooperative Extension, Reno, NV. Earles, J. M., M. P. North, and M. D. Hurteau. 2014. Wildfire and drought dynamics destabilize carbon stores of fire-suppressed forests. Ecological Applications 24:732-740. Egan, J. M., W. R. Jacobi, J. F. Negron, S. L. Smith, and D. R. Cluck. 2010. Forest thinning and subsequent bark beetle-caused mortality in Northeastern California. Forest Ecology and Management 260:1832-1842. Fettig, C. J., K. D. Klepzig, R. F. Billings, A. S. Munson, T. E. Nebeker, J. F. Negrón, and J. T. Nowak. 2007. The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern United States. Forest Ecology and Management 238:24-53. Gagolewski, M. 2013. Computer Statistics with R. Gan, J. 2004. Risk and damage of southern pine beetle outbreaks under global climate change. Forest Ecology and Management 191:61-71. Guarín, A. and A. H. Taylor. 2005. Drought triggered tree mortality in mixed conifer forests in Yosemite National Park, California, USA. Forest Ecology and Management 218:229-244. H. Aukema, B., A. L. Carroll, Y. Zheng, J. Zhu, K. F. Raffa, R. Dan Moore, K. Stahl, and S. W. Taylor. 2008. Movement of outbreak populations of mountain pine beetle: influences of spatiotemporal patterns and climate. Ecography:080227084236895. Hebertson, E. G. and M. J. Jenkins. 2008. Climate Factors Associated with Historic Spruce Beetle ( Coleoptera : Curculionidae ) Outbreaks in Utah and Colorado Climate Factors Associated with Historic Spruce Beetle ( Coleoptera : Curculionidae ) Outbreaks in Utah and Colorado. 37:281292. Hicke, J. a., C. D. Allen, A. R. Desai, M. C. Dietze, R. J. Hall, E. H. Ted Hogg, D. M. Kashian, D. Moore, K. F. Raffa, R. N. Sturrock, and J. Vogelmann. 2012a. Effects of biotic disturbances on forest carbon cycling in the United States and Canada. Global Change Biology 18:7-34. Hicke, J. A., M. C. Johnson, J. L. Hayes, and H. K. Preisler. 2012b. Effects of bark beetle-caused tree mortality on wildfire. Forest Ecology and Management 271:81-90. Hicke, J. A., J. A. Logan, J. Powell, and D. S. Ojima. 2006. Changing temperatures influence suitability for modeled mountain pine beetle (Dendroctonus ponderosae) outbreaks in the western United States. Journal of Geophysical Research 111:1-12. Hurteau, M., H. Zald, and M. North. 2007. Species-specific response to climate reconstruction in upperelevation mixed-conifer forests of the western Sierra Nevada, California. Canadian Journal of Forest Research 37:1681-1691. 39 Hurteau, M. D. and M. L. Brooks. 2011. Short- and Long-term Effects of Fire on Carbon in US Dry Temperate Forest Systems. BioScience 61:139-146. Hurteau, M. D., T. a. Robards, D. Stevens, D. Saah, M. North, and G. W. Koch. 2014. Modeling climate and fuel reduction impacts on mixed-conifer forest carbon stocks in the Sierra Nevada, California. Forest Ecology and Management 315:30-42. Jenkins, J. C., D. C. Chojnacky, L. S. Heath, and R. A. Birdsey. 2004. Comprehensive Database of Diameter-based Biomass Regressions for North American Tree Species. USDA Forest Service General Technical Report NE-319. Kretchun, A. M., R. M. Scheller, M. S. Lucash, K. L. Clark, J. Hom, and S. Van Tuyl. 2014. Predicted Effects of Gypsy Moth Defoliation and Climate Change on Forest Carbon Dynamics in the New Jersey Pine Barrens. Plos One 9:e102531. Kurz, W. a., C. C. Dymond, G. Stinson, G. J. Rampley, E. T. Neilson, a. L. Carroll, T. Ebata, and L. Safranyik. 2008. Mountain pine beetle and forest carbon feedback to climate change. Nature 452:987-990. Loudermilk, E. L., R. M. Scheller, P. J. Weisberg, J. Yang, T. E. Dilts, S. L. Karam, and C. Skinner. 2013. Carbon dynamics in the future forest: the importance of long‐term successional legacy and climate–fire interactions. Global Change Biology 19:3502-3515. Loudermilk, E. L., A. Stanton, R. M. Scheller, T. E. Dilts, P. J. Weisberg, C. Skinner, and J. Yang. 2014. Effectiveness of fuel treatments for mitigating wildfire risk and sequestering forest carbon: A case study in the Lake Tahoe Basin. Forest Ecology and Management 323:114-125. Loudermilk, E. L., A. E. Stanton, R. M. Scheller, P. J. Weisberg, J. Yang, T. E. Dilts, and C. Skinner. 2012. Final Report: Management Options for Reducing Wildfire Risk and Maximizing Carbon Storage under Future Climate Changes, Ignition Patterns, and Forest Treatments P049, Pacific Southwest Research Station, Tahoe Center for Environmental Studies, Incline Village, NV. Macomber, S. A. and C. E. Woodcock. 1994. Mapping and monitoring conifer mortality using remote sensing in the Lake Tahoe Basin. Remote Sensing of Environment 50:255-266. McGinnis, T. W., J. E. Keeley, S. L. Stephens, and G. B. Roller. 2010. Fuel buildup and potential fire behavior after stand-replacing fires, logging fire-killed trees and herbicide shrub removal in Sierra Nevada forests. Forest Ecology and Management 260:22-35. Millar, C. I., N. L. Stephenson, and S. L. Stephens. 2007. Climate Change and Forests of the Future: Managing in the face of Uncertainty. Ecological Applications 17:2145-2151. Nagel, T. A. and A. H. Taylor. 2005. Fire and Persistence of Montane Chaparral in Mixed Conifer Forest Landscapes in the Northern Sierra Nevada, Lake Tahoe Basin, California, USA. Journal of the Torrey Botanical Society 132:442-457. Negrón, J. F. and J. B. Popp. 2004. Probability of ponderosa pine infestation by mountain pine beetle in the Colorado Front Range. Forest Ecology and Management 191:17-27. Nicault, a., S. Alleaume, S. Brewer, M. Carrer, P. Nola, and J. Guiot. 2008. Mediterranean drought fluctuation during the last 500 years based on tree-ring data. Climate Dynamics 31:227-245. Ottmar, R. and H. Safford. 2011. Final Report: FCCS Fuelbeds for the Lake Tahoe Basin Management Unit. Final Report P018, Pacific Southwest Research Station, Tahoe Center for Environmental Studies, Incline Village, NV. Parker, T. J., K. M. Clancy, and R. L. Mathiasen. 2006. Interactions among fire, insects and pathogens in coniferous forests of the interior western United States and Canada. Agricultural and Forest Entomology 8:167-189. Parton, W. J., D. W. Anderson, C. V. Cole, and J. W. B. Stewart. 1983. Simulation of soil organic matter formation and mineralization in semiarid agroecosystems. Pages 533-550 in R. L. T. R.R. Lowrance, L.E. Asmussen and R.A. Leonard, editor. Nutrient cycling in agricultural ecosystems. The University of Georgia, College of Agriculture Experimental Stations, Athens, Georgia. 40 Rogers, J. H. 1974. Soil survey Tahoe Basin Area: California and Nevada. U.S. Department of Agriculture, Soil Conservation Service, Washington, D.C. Rutter, N., R. Essery, J. Pomeroy, N. Altimir, K. Andreadis, I. Baker, A. Barr, P. Bartlett, A. Boone, H. Deng, H. Douville, E. Dutra, K. Elder, C. Ellis, X. Feng, A. Gelfan, A. Goodbody, Y. Gusev, D. Gustafsson, R. Hellström, Y. Hirabayashi, T. Hirota, T. Jonas, V. Koren, A. Kuragina, D. Lettenmaier, W.-P. Li, C. Luce, E. Martin, O. Nasonova, J. Pumpanen, R. D. Pyles, P. Samuelsson, M. Sandells, G. Schädler, A. Shmakin, T. G. Smirnova, M. Stähli, R. Stöckli, U. Strasser, H. Su, K. Suzuki, K. Takata, K. Tanaka, E. Thompson, T. Vesala, P. Viterbo, A. Wiltshire, K. Xia, Y. Xue, and T. Yamazaki. 2009. Evaluation of forest snow processes models (SnowMIP2). Journal of Geophysical Research 114:D06111. Scheller, R. and D. Mladenoff. 2008. Simulated effects of climate change, fragmentation, and interspecific competition on tree species migration in northern Wisconsin, USA. Climate Research 36:191-202. Scheller, R. M., D. Hua, P. V. Bolstad, R. A. Birdsey, and D. J. Mladenoff. 2011a. The effects of forest harvest intensity in combination with wind disturbance on carbon dynamics in Lake States Mesic Forests. Ecological Modelling 222:144-153. Scheller, R. M., S. Van Tuyl, K. Clark, J. Hom, and I. La Puma. 2011b. Carbon Sequestration in the New Jersey Pine Barrens Under Different Scenarios of Fire Management. Ecosystems 14:987-1004. Seager, R., M. Ting, I. Held, Y. Kushnir, J. Lu, G. Vecchi, H.-P. Huang, N. Harnik, A. Leetmaa, N.-C. Lau, C. Li, J. Velez, and N. Naik. 2007. Model Projections of an Imminent Transition to a More Arid Climate in Southwestern North America. Science 316:1181-1184. Steenberg, J. W. N., P. N. Duinker, and P. G. Bush. 2011. Exploring adaptation to climate change in the forests of central Nova Scotia, Canada. Forest Ecology and Management 262:2316-2327. Stephens, S. L. and L. W. Ruth. 2005. Federal forest-fire policy in the United States. Ecological Applications 15:532-542. Sturtevant, B. R., E. J. Gustafson, W. Li, and H. S. He. 2004. Modeling biological disturbances in LANDIS: a module description and demonstration using spruce budworm. Ecological Modelling 180:153174. Sturtevant, B. R., R. M. Scheller, B. R. Miranda, D. Shinneman, and A. Syphard. 2009. Simulating dynamic and mixed-severity fire regimes: A process-based fire extension for LANDIS-II. Ecological Modelling 220:3380-3393. Syphard, A. D., R. M. Scheller, B. C. Ward, W. D. Spencer, and J. R. Strittholt. 2011. Simulating landscapescale effects of fuels treatments in the Sierra Nevada, California, USA. International Journal of Wildland Fire 20:364-383. US Department of Agriculture, F. S. 2004. USDA Forest Service Strategic Plan for Fiscal Years 2004-2008. USDA Forest Service Pacific Southwest Region. 2013. Region 5 Aerial Detection Monitoring. van Mantgem, P. J., N. L. Stephenson, J. C. Byrne, L. D. Daniels, J. F. Franklin, P. Z. Fulé, M. E. Harmon, A. J. Larson, J. M. Smith, A. H. Taylor, and T. T. Veblen. 2009. Widespread Increase of Tree Mortality Rates in the Western United States. Science 323:521-524. Walker, R. F., R. M. Fecko, W. B. Frederick, D. W. Johnson, and W. W. Miller. 2007. Forest Health Impacts of Bark Beetles, Dwarf Mistletoe, and Blister Rust in a Lake Tahoe Basin Mixed Conifer Stand. Western North American Naturalist 67:562-571. Ward, N. L. and G. J. Masters. 2007. Linking climate change and species invasion: an illustration using insect herbivores. Global Change Biology 13:1605-1615. Weed, A. S., M. P. Ayres, and J. A. Hicke. 2013. Consequences of climate change for biotic disturbances in North American forests. Ecological Monographs 83:441-470. Westerling, A. L., H. G. Hidalgo, D. R. Cayan, and T. W. Swetnam. 2006. Warming and Earlier Spring Increase Western U.S. Forest Wildfire Activity. Science 313:940-943. 41 Xu, C., G. Z. Gertner, and R. M. Scheller. 2009. Uncertainties in the response of a forest landscape to global climatic change. Global Change Biology 15:116-131. Zhao, M., S. W. Running, and N. Oceanic. 1986. Drought-Induced Reduction in Global. 1667:2-5. 42 APPENDIX Tree species and functional group attributes used in LANDIS-II modeling of the Lake Tahoe Basin, CA, NV, USA. Species or functional group Longevity (yrs.) Age of Sexual Maturity (yrs.) Shade tolerance (1-5) Fire tolerance (1-5) Effective seeding distance (m) Maximum seeding distance (m) Vegetative Reproduction probability Minimum resprouting age Maximum resprouting age Post-fire regeneration Pinus jeffreyi 500 25 2 5 50 300 0 0 0 none Pinus lambertiana 550 20 3 5 30 400 0 0 0 none Calocedrus decurrens 500 30 4 5 30 2000 0 0 0 none Abies concolor 450 35 4 3 30 500 0 0 0 none Abies magnifica 500 40 3 4 30 500 0 0 0 none Pinus contorta 250 7 1 2 30 300 0 0 0 none Pinus monticola 550 18 3 4 30 800 0 0 0 none Tsuga mertensiana 800 20 5 1 30 800 0.0005 100 800 none Pinus albicaulis 900 30 3 2 30 5000 0.0001 100 900 none Populus tremuloides Non N-fixing resprouting shrubs Non N-fixing obligate seeding shrubs 175 15 1 2 30 1000 0.9 1 175 resprout 80 5 2 1 30 550 0.85 5 70 resprout 80 5 2 1 30 1000 0 0 0 none N-fixing resprouting shrubs N-fixing obligate seeding shrubs 80 5 1 1 30 500 0.75 5 70 resprout 80 5 1 1 30 800 0 0 0 none 43 Physiological parameters for 11 tree species and 4 shrub functional groups found in the Lake Tahoe Basin, CA, NV, USA. GDD: Growing Degree Days Species or functional group Functional type Nitrogen tolerance GDD min GDD max Min. Jan. Temp. (C) Max Drought Leaf longevity (yrs.) Leaf lignin (%) Fine root lignin (%) Wood lignin (%) Coarse root lignin (%) Leaf CN ratio Fine root C:N ratio Wood C:N ratio Coarse root C:N ratio Litter C:N ratio Pinus jeffreyi 1 N 555 2149 -5 0.94 6.0 0.28 0.2 0.25 0.25 48 48 250 167 100 Pinus lambertiana 1 N 815 2866 -5 0.90 2.5 0.17 0.2 0.25 0.25 53 53 278 185 100 Calocedrus decurrens 1 N 837 2938 18 0.99 4.0 0.1 0.2 0.25 0.25 48 48 500 333 100 Abies concolor 1 N 540 2670 -10 0.93 8.0 0.17 0.2 0.25 0.25 30 30 333 222 100 Abies magnifica 1 N 483 1144 -18 0.87 8.0 0.17 0.2 0.25 0.25 30 30 250 167 100 Pinus contorta 1 N 276 993 -18 0.87 3.5 0.25 0.2 0.25 0.25 48 48 500 333 100 Pinus monticola 1 N 155 1016 -18 0.82 7.0 0.31 0.2 0.25 0.25 37 37 500 333 100 Tsuga mertensiana 1 N 235 894 -18 0.80 4.5 0.24 0.2 0.25 0.25 80 80 333 222 100 Pinus albicaulis 1 N 230 950 -18 0.90 5.5 0.27 0.2 0.25 0.25 80 80 333 222 100 Populus tremuloides 2 N 600 3000 -10 0.82 1.0 0.18 0.2 0.25 0.25 62 62 333 222 100 Non N-fixing resprouting shrubs Non N-fixing obligate seeding shrubs 3 N 400 4000 -10 0.99 1.5 0.25 0.2 0.25 0.25 56 56 333 222 100 3 N 400 4000 -10 0.97 1.5 0.25 0.2 0.25 0.25 59 59 333 222 100 N-fixing resprouting shrubs 3 Y 400 4000 -10 0.97 1.5 0.25 0.2 0.25 0.25 20 28 333 222 50 N-fixing obligate seeding shrubs 3 Y 400 4000 -10 0.99 1.5 0.25 0.2 0.25 0.25 20 30 333 222 50 44 Functional type parameters for the species list in Table above. Name Functional type index PPDF1 mean PPDF2 max PPDF1 shape 1 PPDF1 shape 2 NPP leaf (%) BTOLAI KLAI MAXLAI PPRPTS2 PPRPTS3 Wood decay rate Wood mortality (%/mo.) Age mortality shape Leaf Drop Month Conifers 1 23.0 40.0 0.05 6.0 0.2 0.002 5000 10 0.2 0.1 1.0 0.002 10.0 9.0 Hardwoods 2 23.0 35.0 0.05 7.0 0.3 0.002 5000 20 0.2 0.1 1.0 0.002 10.0 9.0 Shrubs 3 23.0 32.0 0.05 10.0 0.3 0.002 500 5 0.2 0.1 1.0 0.002 10.0 9.0 Ecoregion fixed parameters for the Lake Tahoe Basin, CA, NV, USA. Ecoregion Soil depth (cm) % Clay fraction % Sand fraction Field capacity Wilting point Storm flow fraction Base flow fraction Drainage Atmospheric N (slope) Atmospheric N (intercept) Latitude Decay rate Surface soil Decay rate SOM1 Decay rate SOM2 Decay rate SOM3 Denitrification Eastside Forest & Woodland 100 0.070 0.714 0.109 0.055 0.4 0.4 0.691 0.08 0.005 39.02 0.4 1 0.02 0.0002 0.5 Upper Montane 100 0.050 0.786 0.090 0.046 0.4 0.4 0.815 0.08 0.005 39.02 0.4 1 0.02 0.0002 0.5 Subalpine 100 0.061 0.744 0.097 0.051 0.4 0.4 0.735 0.08 0.005 39.02 0.4 1 0.02 0.0002 0.5 Montane shrubland 100 0.078 0.718 0.090 0.060 0.4 0.4 0.826 0.08 0.005 39.02 0.4 1 0.02 0.0002 0.5 Riparian areas 100 0.068 0.704 0.123 0.053 0.4 0.4 0.523 0.08 0.005 39.02 0.4 1 0.02 0.0002 0.5 45 Initial values of Carbon and Nitrogen in various soil organic pools. Ecoregion SOM1 C (surface) SOM1 N (surface) SOM1 C (soil) SOM1 N (soil) SOM2 C SOM2 N SOM3 C SOM3 N Mineral N Eastside Forest & Woodland 75.0 3.0 100.0 10.0 3000.0 50.0 300.0 15.0 3.0 Upper Montane 75.0 3.0 100.0 10.0 3000.0 50.0 300.0 15.0 3.0 Subalpine 75.0 3.0 100.0 10.0 3000.0 50.0 300.0 15.0 3.0 Montane shrubland 75.0 3.0 100.0 10.0 3000.0 50.0 300.0 15.0 3.0 Riparian areas 75.0 3.0 100.0 10.0 3000.0 SOM1: Soil Organic Matter fast pool; SOM2: Soil Organic Matter fast pool; SOM3: Soil Organic Matter passive pool 50.0 300.0 15.0 3.0 Fire regime parameter inputs for the Dynamic Fire extension used for the Lake Tahoe Basin, CA, NV. FMC: Foliar moisture content. Fire Region Region size (ha) sigma Maximum Size or Duration Spring FMC Low Spring FMC High Spring High Proportion Summer FMC Low Summer FMC High Summer High Proportion mu Lower-elevation 9,603 5.2 0.32 4500 135 175 0.05 85 100 0.89 Mid-elevation Higher-elevation 28,777 5.3 0.32 4500 135 175 0.05 85 100 0.89 31,194 5.2 0.32 4500 135 175 0.05 85 100 0.89 46 Bark beetle species, their host tree species, and associated minor, moderate, and major host ages as input in the BDA extension used for the Lake Tahoe Basin, CA, NV. Bark beetle species Host tree species Minor host age Moderate host age Major host age Mountain pine beetle (Dendroctonus ponderosae) Pinus albicaulis 20 60 80 Pinus lambertiana 20 60 80 Pinus contorta 20 60 80 Pinus monticola 20 60 80 Calocedrus decurrens 20 60 80 Jeffrey pine beetle (Dendroctonus jeffreyi) Pinus jeffreyi 15 25 40 Fir engraver beetle (Scolytus ventralis) Abies concolor 15 30 60 Abies magnifica 15 30 60 47