Problem #3: spindle positioning; chromosomal directional instability; mono- oriented chromosomal oscillations
advertisement

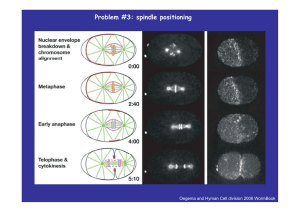
Problem #3: spindle positioning; chromosomal directional instability; monooriented chromosomal oscillations Oegema and Hyman Cell division 2006 WormBook How many motors pull spindle to the cortex? Grill et al 2003 Science 301 518 Grill and Hyman Dev Cell 8 461 2005 Spindle oscillates as it moves in the posterior direction Pecreaux et al Curr Biol 2006 16 2111 Kozlowski et al Cell 2007 129 499 Interplay between spindle pole movements and microtubule dynamics Microtubules Contact the Cortex Briefly before Undergoing Catastrophe Dynamic Microtubules Associate with Each Other to Form Persistent Astral Fibers Microtubule Fibers Contact the Cortex at Fixed Positions more MT contacts are made on the side of the cortex t that th t is i approached h db by th the aster t th than on the receding side. Kozlowski et al Cell 129 499 2007 N+ y dN ± v ⎛ = k1 ⎜1 ± dt ⎝ V ⎞ ⎟ − k2 N ± ⎠ V d ( N + − N − ) 2k1 v − k2 ( N + − N − ) = dt V v N− y ΔN ± V ± v ) Δt ( = Vδ t dy =v dt dn 2k1 = n = N+ − N− , v − k2 n dt V f K f K v = ( N+ − N− ) − y = n − y ζ ζ ζ ζ ζ K n = v+ y f f ζ dv ⎛ζ K dy 2k1 K + = v − k2 ⎜ v + f dt f dt V f ⎝ f ⎞ y⎟ ⎠ 2k1 k2ζ ⎞ dy k2 K +⎜ − + y=0 ⎟ + 2 f dt V f ⎠ dt f ⎝ f ⎛K 2 fk1 ⎞ dy k2 K d2y = − ⎜ + k2 − y ⎟ − 2 ζ V ⎠ dt ζ dt ⎝ζ ζ d2y ⎛ K ξ ( 1 y ~ e , λ = −ξ ± ξ 2 − 4k 2 λt k ) Things g to think about: dependence p of p period and bifurcation on model p parameters; what limits the amplitude of oscillations? ‘Tug of war’ between opposing force generators The key idea behind the model is that the rate of detachment of the force generators from MTs is load dependent Grill et al Phys Rev Lett 2005 94 108104 Pecreaux et al Curr Biol 2006 16 2111 ζ y = − Ky + ( f + − f − ) y + p+ = − koff p+ + kon (1 − p+ ) − p− = −koff p− + kon (1 − p− ) p− , f − ⎛ v⎞ f = f 0 ⎜1 − ⎟ ⎝ v0 ⎠ f ± = p± ( f 0 ∓ ϕ y ) ± kon = const ; koff = k off(0) exp [ f ± / f c ] ≈ k off(0) (1 ∓ α y ) ζ y = − Ky + θ ( p+ − p− ) − σ y ( p+ + p− ) p+ ≈ k − ( a − by ) p+ p = p+ + p− z = p+ − p− p− ≈ k − ( a + by ) p− ζ y = − Ky + θ z − σ yp p = 2k − ap + byz z ≈ − az + byp ζ y = − Ky + θ z − σ yp p = 2k − ap + byz z ≈ − az + byp λ ~ ( s1 − s2 ) ± s1 , s2 , s3 > 0 y = 0, z = 0, p = 2k / a ⎡ y ⎤ ⎡ y ⎤ ⎡ y0 ⎤ ⎢ p ⎥ = ⎢ p ⎥ + ⎢ p ⎥ eλt ⎢ ⎥ ⎢ ⎥ ⎢ 0⎥ ⎢⎣ z ⎥⎦ ⎢⎣ z ⎥⎦ ⎢⎣ z0 ⎥⎦ ( s1 − s2 ) 2 − s32 exp ( γ t ) × sin ( wt ) Things to think about: linearize and do carefully linear perturbation analysis; dependence of period and bifurcation on model parameters; what limits the amplitude of oscillations? Read both papers and think about testing the model Another example of oscillations: Directional instability phenomenon Joglekar and Hunt Biophys J 2002 83 42 Gardner and Odde Cur Opin Cell Biol 2006 18:639 plus, rules for MT dynamic instability… Civelekoglu-Scholey et al., Biophys J. 2006 90:3966-82. High kinetochore dynein activity is deployed to dampen metaphase oscillations; Modulation of the MT rescue frequency by the kinetochore-associated kinesin-13 depolymerase promotes metaphase chromosome oscillations As only a part of the force imbalance is accounted for by the viscous drag arising from chromosome velocity, a large portion of this force imbalance is stored as kinetochore resistance, which depends on both the chromosome position and chromosome velocity Liu et al. (2007) PNAS 104, 16104-16109 Relative amount of bound motors at which AP and P forces exactly balance effect of chromosome friction sensitivity of motor unbinding to an external load Campàs O, Sens P. Phys Rev Lett. 2006 97(12):128102.