Problem #3: spindle positioning Oegema and Hyman Cell division 2006 WormBook
advertisement

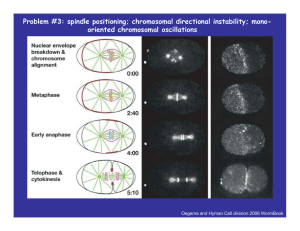
Problem #3: spindle positioning Oegema and Hyman Cell division 2006 WormBook How many motors pull spindle to the cortex? Grill et al 2003 Science 301 518 Grill and Hyman Dev Cell 8 461 2005 Spindle oscillates as it moves in the posterior direction Pecreaux et al Curr Biol 2006 16 2111 Kozlowski et al Cell 2007 129 499 ‘Tug of war’ between opposing force generators The key idea behind the model is that the rate of detachment of the force generators from MTs is load dependent Grill et al Phys Rev Lett 2005 94 108104 Pecreaux et al Curr Biol 2006 16 2111 ζ y& = − Ky + ( f + − f − ) y p& + = − koff p+ + kon (1 − p+ ) p& − = −koff p− + kon (1 − p− ) p− , f − ⎛ v⎞ f = f 0 ⎜1 − ⎟ ⎝ v0 ⎠ f ± = f 0 m ϕ y& kon = const ; koff = k off(0) exp [ f / f c ] ≈ k off(0) (1 − α y& ) ζ y& = − Ky + α ( p+ − p− ) − αβ y& ( p+ + p− ) p& + ≈ k − ( a − by& ) p+ p& − ≈ k − ( a − byy& ) p− λ ~ (α s1 − s2 ) ± s1 , s2 , s3 > 0 (α s1 − s2 ) 2 − s32 y = 0, p+ = p− = k / a ⎡ y ⎤ ⎡ y ⎤ ⎡ y0 ⎤ ⎢ p ⎥ = ⎢ p ⎥ + ⎢ p ⎥ eλt ⎢ +⎥ ⎢ +⎥ ⎢ 1⎥ ⎢⎣ p− ⎥⎦ ⎢⎣ p− ⎥⎦ ⎢⎣ p2 ⎥⎦ exp ( γ t ) × sin ( wt ) Interplay between spindle pole movements and microtubule dynamics Microtubules Contact the Cortex Briefly before Undergoing Catastrophe Dynamic Microtubules Associate with Each Other to Form Persistent Astral Fibers Microtubule Fibers Contact the Cortex at Fixed Positions more MT contacts are made on the side of the cortex that is approached by the aster than on the receding side. Kozlowski et al Cell 129 499 2007 f v y 0.6 2 y 1.5 f ( y& ) 1 0.4 0.2 0.5 y& 0 0 y& = − Ky + f ( y& ) -0.2 -0.5 -0.4 -1 -0.6 -1.5 -2 -20 -15 -10 -5 0 5 10 15 20 -0.8 -2 1 ⎡ f ( y& ) − y& ⎤⎦ K⎣ -1.5 -1 -0.5 0 0.5 1 1.5 2 Another example of oscillations: Directional instability phenomenon Joglekar and Hunt Biophys J 2002 83 42 Gardner and Odde Cur Opin Cell Biol 2006 18:639 Problem #4: spindle length f v f v Two Opposing Motors v Can two opposing motors compete? - FK k Klp v FN n Ncd + ⎛ v ⎞ nf n ⎜1 − ⎟ = kζ k v ⎝ Vn ⎠ Vn x n ,x = v= x + ε n (1 − x ) n+k εn = ζ kVn fn ≈ 4.5 ⎛ v ⎞ kf k ⎜1 − ⎟ = nζ n v ⎝ Vk ⎠ V (1 − x ) n ,x = v= k n+k (1 − x ) + ε k x εk = ζ nVk fk ≈ 0.25 ζ k ~ ζ n ~ kt ~ 0.05 pN×sec/nm t ~ 0.01/sec, k ~ 1 pN/nm S2 ς S3 dS3 ς = k5 − k 6 S 3 dt ς Nédélec J Cell Biol 2002 158 1005 dS 2 = k3 − k 4 S 2 dt dS4 = k7 ( L − S 4 ) − k8 S 4 dt L S1 dS1 ς = ⎡⎣ k1 ( L − S1 ) − k2 S1 ⎤⎦ dt Important here is that due to transport properties of the motors there is a selection of the motors of the same p polarity y at p parallel overlapping pp g MT p pairs. Nédélec J Cell Biol 2002 158 1005 Nédélec J Cell Biol 2002 158 1005 Ambrose et al Mol Biol Cell 2005 16 1584 Force balance models Motors 45 4,5 MT dynamics 1,6 , “spindle matrix” 3 cortex forces 2 Extrinsic mechanisms What determines stable length of bi-polar p spindle? p depletion of molecules 1 Concentration gradient of morphogens models Intrinsic mechanisms Balance of dynein (outward) and ncd (inward) forces explains pole separation and transient steady state in interphase - prophase S Fdyn Fncd Sharp p et al., Mol. Biol. Cell. 2000 11:241 Cytrynbaum et al., Biophys. J. 2003 84:757 Geometry questions: where are dynein, ncd, actin? Are MT asters asymmetric y and how are they made asymmetric? Mechanical q questions: how strong are the forces? dS 2 = ( Fdyn y − k ncd S ) dt ς S ≈ S0 (1 − e − t / T ) ?