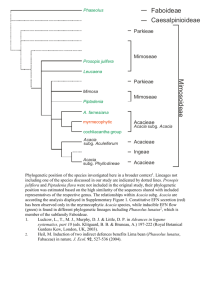
SPECIES DENSITY OF THE SOUTHERN LESSER BUSHBABY (GALAGO MOHOLI)
advertisement
")
SPECIES DENSITY OF THE SOUTHERN LESSER BUSHBABY (GALAGO MOHOLI) AT LOSKOP DAM NATURE RESERVE, MPUMALANGA, SOUTH AFRICA, WITH NOTES ON HABITAT PREFERENCE A THESIS SUBMITTED TO THE GRADUATE SCHOOL IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE MASTER OF ARTS BY IAN S. RAY DR. EVELYN BOWERS, CHAIRPERSON BALL STATE UNIVERSITY MUNCIE, INDIANA MAY 2014 SPECIES DENSITY OF THE SOUTHERN LESSER BUSHBABY (GALAGO MOHOLI) AT LOSKOP DAM NATURE RESERVE, MPUMALANGA, SOUTH AFRICA, WITH NOTES ON HABITAT PREFERENCE A THESIS SUBMITTED TO THE GRADUATE SCHOOL IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE MASTER OF ARTS BY IAN S. RAY Committee Approval: ____________________________________ Committee Chairperson ________________________ Date ____________________________________ Committee Member ________________________ Date ____________________________________ Committee Member ________________________ Date Departmental Approval: ____________________________________ Department Chairperson ________________________ Date ____________________________________ Dean of Graduate School ________________________ Date BALL STATE UNIVERSITY MUNCIE, INDIANA MAY 2014 TABLE OF CONTENTS 1. ABSTRACT. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . iii 2. ACKNOWLEDGEMENTS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . iv 3. LIST OF TABLES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .v 4. LIST OF FIGURES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vi 5. LIST OF APPENDICES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .vii 6. INTRODUCTION. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .1 a. BACKGROUND AND THEORY. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 b. LITERATURE REVIEW. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2 i. HABITAT. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 ii. MORPHOLOGY. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .5 iii. MOLECULAR BIOLOGY. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7 iv. REPRODUCTION. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .8 v. SOCIALITY. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10 vi. DIET. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 vii. LOCOMOTION. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .12 c. OBJECTIVES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13 7. MATERIALS AND METHODS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .15 a. STUDY SITE. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .15 b. DATA COLLECTION. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16 c. DATA ANLYSES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .16 8. RESULTS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20 a. SPECIES DENSITY. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20 i b. ASSOCIATED PLANT SPECIES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21 9. DISCUSSION. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24 a. SPECIES DENSITY. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24 b. HABITAT PREFERENCE. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25 10. CONCLUSION. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28 11. REFERENCES CITED. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29 12. APPENDICES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33 ii ABSTRACT THESIS: Species Density of the Southern Lesser Bush Baby (Galago moholi) at Loskop Dam Nature Reserve, Mpumalanga, South Africa with notes on habitat preference. STUDENT: Ian S. Ray DEGREE: Master of Arts COLLEGE: Sciences and Humanities DATE: May 2014 PAGES: 68 A population survey was conducted on Galago moholi along the road system at Loskop Dam Nature Reserve, Mpumalanga province, South Africa. The data were analyzed using the maximum perpendicular distance, mean perpendicular distance, and maximum reliable perpendicular distance methods. Vegetation sample plots were constructed at the location of each individual sighted in order to analyze the species’ habitat use. The results indicate that the species density of G. moholi is significantly lower at Loskop Dam Nature Reserve than previously reported at other sites within South Africa. G. moholi was found to prefer areas with high concentrations of Dichrostachys sp., Combretum sp., or Acacia sp. One individual was observed consuming vegetative matter, which may indicate that the population within the reserve is utilizing available resources in a different way than populations in other parts of southern Africa. iii ACKNOWLEDGEMENTS I will be eternally grateful to all who helped to make this project possible... My adviser and committee chairperson, Dr. Evelyn Bowers, deserves much gratitude for supplying me with a seemingly endless amount of advice and resources, both in the classroom and in the field. I thank my other committee members, Dr. S. Homes Hogue and Dr. Ronald Hicks, for their continued support throughout my academic career. Furthermore, I would like to extend my thanks to the full staff of Ball State University’s Department of Anthropology for their genuine compassion and dedication to the discipline. I would like to thank Dr. Brandi Wren for her help both in field training and throughout my field research, as well as Ruby Malzoni and Michele Mingini for their assistance in conducting the population survey. I am grateful to the aid provided by Jannie Coetzee and the staff of the Applied Behavioural Ecology and Ecosystem Research Unit (ABEERU) of the University of South Africa (UNISA), and for being allowed to work at Loskop Dam Nature Reserve. iv LIST OF TABLES TABLE 1: ESTIMATED SPECIES DENSITY OF G. MOHOLI ALONG VARIOUS TRANSECT SEGMENTS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .20 TABLE 2: ESTIMATED SPECIES DENSITY OF G. MOHOLI AT LOSKOP DAM NATURE RESERVE. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .21 TALBE 3: PLANT SPECIES IDENTIFIED AND NUMBER OF EACH SPECIES. . . . . . 22 TABLE 4: MAJOR PLANT GENUS AND PERCENT OF TOTAL WOODY SPECIES. . 23 TABLE 5: PERCENTAGE OF TREES BY SIZE CLASS. . . . . . . . . . . . . . . . . . . . . . . . . . .23 TABLE 6: PERCENTAGE OF CENTER TREE DBH BY SIZE CLASS. . . . . . . . . . . . . . .23 TABLE 7: PERCENT CANOPY COVER BY CLASS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 v LIST OF FIGURES FIGURE 1: MAP DISPLAYING THE PROVINCES OF SOUTH AFRICA. . . . . . . . . . . . . . 2 FIGURE 2: MAP DISPLAYING THE RANGE OF O. CRASSICAUDATUS AND G. MOHOLI. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 FIGURE 3: LOCATION OF THE LOSKOP DAM NATURE RESERVE IN THE MPUMALANGA PROVINCE OF SOUTH AFRICA. . . . . . . . . . . . . . . . . . . . .15 FIGURE 4: TRANSECTS LABELED BY COLOR. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18 FIGURE 5: LOCATION OF EACH INDIVIDUAL AT INITIAL SIGHTING. . . . . . . . . . . 19 FIGURE 6: G. MOHOLI CONSUMING THE FRUIT OF D. CINEREA. . . . . . . . . . . . . . . 27 vi LIST OF APPENDICES APPENDIX 1: TRANSECT DATA. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .33 APPENDIX 2: VEGETATION SAMPLE PLOT DATA. . . . . . . . . . . . . . . . . . . . . . . . . 44 vii INTRODUCTION Background and Theory Speciation by means of natural selection was first proposed by Darwin (2009). The concept of natural selection has been modified, resulting in the Synthetic Theory of evolution (Huxley 1939). This synthetic theory combines natural selection, genetics, and random processes (Huxley 1939). It is necessary to understand a population’s environment in order to understand the selective pressures to which the population is exposed. Thus, ecological studies are an important step towards understanding the evolution of a species, both past and present. The study of ecology would be impossible without the concept of a niche (Colwell and Rangel 2009; Wake et al. 2009). A niche is commonly described as a multidimensional hypervolume of space having the required resources and environmental conditions allowing a species to live (Colwell and Rangel 2009). In the face of climate change, niches change and populations may shift to new territory, adapt, or go into decline (Colwell and Rangel 2009; Wake et al. 2009). These niche changes produce differing pressures upon which natural selection acts, providing the mechanism for evolution (Darwin 2009; Wake et al. 2009) Two generally recognized species of prosimians are found within South Africa, Galago moholi A. Smith 1836 and Otolemur crassicaudatus E. Geoffroy 1812 (Ravosa et al. 2010). These species are partially sympatric and may display similar ecological adaptations, thus their exact evolutionary relationship is not entirely clear. The goal of this research is to determine what ecological niche G. moholi occupies and whether this niche could overlap with that of a population of O. crassicaudatus should one be introduced to Loskop Dam Nature Reserve, through either natural processes or artificial releases. 1 Literature Review The family Galagidae, or the bush babies, is composed of small nocturnal primates that inhabit the forests, woodlands, and savannahs of the African continent (Stephenson et al. 2010). Galagidae, along with the sister family Lorisidae, form the order Lorisiformes (Seiffert et al. 2003). In turn, Lorisiformes is a sister group to the Lemuriformes of Madagascar (Seiffert et al. 2003). The clade of the Galagidae, the African Lorisiformes, is of interest not only because ancestral populations are a likely source of the Madagascar Lemuriformes, but also because of the role of the animals within their own ecosystem (Stankiewicz et al. 2006). Many species of galago have been described and considerable debate exists over which populations belong to each species (Stephenson et al. 2010). Many species depend heavily on tree exudates, with saps and gums forming up to 75 percent of the diet of some species (Stephenson et al. 2010). The establishment of species currently is based predominantly on behavioral and genetic distinctions rather than on physical similarities between populations (Lawes 2005). This is Figure 1: Map displaying the provinces of South Africa (Skinner and Chimimba 2005:XVII) contrary to the morphological observations that have previously been used to distinguish galago species. Within South Africa, two generally accepted species are found. Galago moholi (the South African galago) is found in Limpopo, Mpumalanga, Gauteng, and northeast KwaZulu-Natal 2 provinces (Lawes 2005). The South African galago was grouped with Galago senegalensis É. Geoffroy 1796 as Galago senegalensis moholi (Anderson 1998; Nash et al. 1989). Otolemur crassicaudatus (the thick-tailed galago) is found in Limpopo, eastern Mpumalanga, and only at lower elevations in KwaZuluNatal provinces (Lawes 2005). Bearder (1974) suggested that the discontinuous distribution of both species indicates that the forests of South Africa have not been continuous since bushbabies first evolved. The two species occur sympatrically in some areas but at lower overall densities than in areas where they are separate (Bearder and Doyle 1974). These are the only members of the Lorisidae that are known to be sympatric outside of the tropics (Crompton 1980). This may be due to niche overlap or to sympatric environments being suboptimal habitat for both species (Bearder and Doyle 1974). Figure 2: Maps displaying the distribution of O. crassicaudatus (top) and G. moholi (bottom) (Lawes 2005:212, 214) 3 Habitat O. crassicaudatus has been said to prefer riverine savannah and woodlands (Nash et al. 1989). O. crassicaudatus is found from sea level to over 1,800 m (Lawes 2005). In Limpopo and Mpumalanga provinces, they often are found in dry woodlands that are adjacent to riverine areas (Lawes 2005).They have been found to rest in plantations of nonnative Eucalyptus spp. and Pinus spp. (Bearder and Doyle 1972; Lawes 2005). G. moholi is also found in riverine bush but can be found in more dry areas as well (Nash et al. 1989). It is most often found in association with Acacia sp. woodlands and the highest densities are found in Acacia karoo thickets (Bearder and Doyle 1972). G. moholi is independent of water sources, allowing it to inhabit areas away from rivers and streams (Lawes 2005). They occur only on the fringes of true forests, seeming to prefer slightly more open habitat (Lawes 2005). Outside of South Africa, they are replaced by Galagoides grantii at elevations under 200m (Lawes 2005). The population density of both South African galago species varies depending on the habitat. For O. crassicaudatus, Bearder and Doyle (1972) report a density of 88 individuals per square kilometer in riparian bush and scrub, 110 per km2 in mixed woodland and plantation forests, and 125per km2 in dune forest. This is of interest because the plantation forests are typically considered sterile areas where wildlife does not thrive (Bearder and Doyle 1972). The density of G. moholi is greatest in A. karoo woodlands, with up to 500 animals per square kilometer being reported (Bearder and Doyle 1972). In riparian bush and scrub G. moholi are found at a density of 95 individuals per km2, while the density decreases to 87 per km2 in areas of mixed woodlands and plantations (Bearder and Doyle 1972). 4 Morphology O. crassicaudatus is the most massive of the galagos, being 1131g on average (Nash et al. 1989). O. crassicaudatus occurs sympatrically with many other galago species over its extensive range, including Otolemur garnettii, G. senegalensis, and G. moholi (Nash et al. 1989). The average length of the head and body is 313mm, plus an average tail length of 410mm (Nash et al. 1989). The pelage of O. crassicaudatus varies in color from brown to grey (Nash et al. 1989). O. crassicaudatus has larger ears, a more robust snout, and a larger overall body size than the similar O. garnettii (Masters and Bragg 2000). The hind foot of O. crassicaudatus, however, is smaller in proportion to overall body size when compared with O. garnettii (Masters and Bragg 2000). G. moholi is significantly smaller than O. crassicaudatus, averaging only 158g (Nash et al. 1989). It is largely sympatric with O. crassicaudatus, O. garnettii, and has been shown to occasionally be sympatric with G. senegalensis (Nash et al. 1989). G. moholi averages only 438mm in total length, with a 288mm tail and a 150mm head and body (Nash et al. 1989). The pelage of G. moholi varies dorsally from brownish-gray to light brown. The flanks and the limbs have a distinctive yellow tinge and dark facial markings are present (Nash et al. 1989). The differences in penile morphology are of taxonomic interest due to the promiscuous mating system of galagos (Dixon 1989). Varying genital morphology may lead to differing reproductive rates, thus providing a basis on which natural selection can act. The penile length of O. crassicaudatus averages 20 mm with a maximum width of 6.5 mm (Anderson 1998). The baculum can be seen protruding from the end of the penis (Anderson 1998). This differs from O. garnettii which has a unique set of curves not seen in other galago species (Anderson 1998). Like G. senegalensis, G. moholi on average has a 10 mm penis (Anderson 1998). However, G. 5 moholi has a uniformly thick shaft of 3mm with a visible baculum and thick spines pointing towards the body (Anderson 1998). G. senegalensis has thin penile spines and no visible baculum (Anderson 1998). G. moholi was split as a species from the similar G. senegalensis on the basis of sympatry in Tanzania (Masters and Bragg 2000). G. moholi has a smaller, narrower snout than G. senegalensis with longer ears and different skull morphology (Masters and Bragg 2000). Although these are cryptic species, Masters and Bragg (2000) found they could distinguish preserved specimens with an accuracy of 89 percent using snout width and the length of the tail, ear, hind foot, and palate. The hair of G. moholi is 0.08mm in diameter with scales every 0.02mm on average (Anderson 1998). The hand and foot pads of G. moholi differ from those of G. senegalensis (Stephenson et al. 2010). This further helps to confirm the separation of G. moholi from G. senegalensis. The dentition of both species follows the standard primate dental formula of 2/2 1/1 3/3 3/3 (Lawes 2005). Like most galagos, the lower incisors and canines are modified to form a tooth-scraper (Stephenson et al. 2010). This is thought to aid in the collection of exudates from trees (Stephenson et al. 2010). In line with this, the ascending ramus of O. crassicaudatus develops at a faster rate than the rest of the mandible, allowing for the mouth to be opened extremely widely to utilize this tooth scraper (Ravosa et al. 2010). Ravosa et al. (2010) found that the ascending ramus of G. moholi develops at the same rate as seen in non-exudivore galagids, implying that the ability to open the mouth extremely wide to feed on gum is not necessary for gumivorous prosimians. Interestingly, the jaw has not been found to be more 6 robust in species like O. crassicaudatus and G. moholi than in other non-exudivorious galagids (Ravosa et al. 2010). Molecular Biology Molecular studies have been used to establish phylogenetic relationships and to estimate the timing of speciation events (Masters et al. 2007; Seiffert et al. 2003). As technology has progressed there has been a transition from using karyotyping methods to using more precise DNA sequence data. The taxonomy of the galagos is well supported at the family and genus levels by mitochondrial DNA sequences (Masters et al. 2007). Both Lorisidae and Galagidae are supported as monophyletic families by the molecular analysis and previous morphological analyses (Masters et al. 2007). Within the Galagidae, the sequencing supports the monophyly of the genera Galago and Otolemur (Masters et al. 2007). This supports the previous removal of O. crassicaudatus from the genus Galago (Anderson 1998). Ying and Butler (1971) reported that five chromosome polymorphisms were found in what they considered a single species of G. senegalensis. They constructed karyotypes for each of 11 individuals studied from groups they termed G. s. zanzibaricus, G.s. moholi, and G.s. braccatus (Ying and Butler 1971). It was found that all individuals of G.s. moholi had a diploid chromosome number of 38, while G.s. zanzibaricus had a diploid number of 36 (Ying and Butler 1971). The third group, G.s. braccatus, was found to have diploid numbers of 36, 37, and 38 in different individuals (Ying and Butler 1971). Individuals were assigned to these subspecies groupings using pelage color, size, weight, and geographic origin (Ying and Butler 1971). Individuals assigned to G.s. braccatus come from an area recognized today as a zone of sympatry between G. moholi and G. senegalensis, thus possibly accounting for the discrepancies in diploid chromosome number within the G.s. braccatus group (Nash et al. 1989; Ying and 7 Butler 1971). The hybrid with 37 chromosomes may be a result of nondisjunction of chromosomes rather than of direct hybridization between n=36 and n=38 chromosome parents (Ying and Butler 1971). While such hybrids are possible in the wild, the two species are unlikely to interbreed due to the differing reproductive timing of G. moholi and G. senegalensis (Nash et al. 1989). This is supported by the observation that all offspring from n=37 individuals had only 36 chromosomes (Ying and Butler 1971) Reproduction Mating in O. crassicaudatus occurs during two weeks at the beginning of the cold, dry winter in June (Clark 1985). Gestation lasts until late October or early November with weaning occurring three months after birth (Clark 1985). According to a paper by Welker and SchaferWitt (1988), twins or triplets are born and newborns cling to and climb on the mother shortly after birth. A study at the Duke University Primate Center found that, with captive O. crassicaudatus, twins or triplets were found in approximately 50 percent of all pregnancies (Izard and Simons 1986). There was not a statistically significant difference in infant survival between wild-born females and captive-born females, or between females who previously had successful pregnancies and first-time mothers (Izard and Simons 1986). A statistically significant male-biased sex ratio was found within all three species of bushbaby studied including O. crassicaudatus and G. moholi, with respective p-values of p=0.0336 and p=0.0091 (Izard and Simons 1986). G. moholi has two breeding season per year and a gestation period of 122 to 125 days. This results in their giving birth in late September or October and in early February (Doyle et al. 1967; Pullen et al. 2000). This corresponds to the beginning and end of the summer wet season (Pullen et al. 2000). In a study at Duke University, twinning occurred in 60 percent of all 8 pregnancies with no triplets occurring (Izard and Simons 1986). Izard and Simons (1986) found a male-bias sex ratio in G. moholi. This may be due to the dispersal pattern of the genus Galago, with males traveling farther and females remaining matrilocal. The pattern allows male offspring to spread their genes farther and more rapidly than female offspring, creating a selective pressure favoring male offspring (Izard and Simons 1986). The nests of O. crassicaudatus are only constructed by females with young, although sleeping in open sites does occur both with and without young (Bearder et al. 2003; Clark 1985). By May, offspring gain up to 75 percent of their adult weight but do not grow further until the next spring (Clark 1985). Dispersal occurs the year after adult size is reached (Clark 1985). Female O. crassicaudatus often retain home ranges near to or overlapping with their mother’s (Clark 1985). Cannibalism has been observed in captivity, where it was likely the stress of experimental conditions that led mothers to cannibalize their infants (Tartabini 1991). G. moholi males have been found to be significantly heavier during breeding seasons than during other times of the year (Pullen et al. 2000). Generally, larger males are more successful in copulation and out-compete smaller males, giving evolutionary significance to this finding (Pullen et al. 2000). Within captive breeding groups of G. moholi, frequent short-duration copulations of 10-15 seconds were observed with a few longer duration mounts up to 450 seconds (Doyle et al. 1967). In the wild, only long duration mounts have been observed lasting 540 seconds on average with the maximum copulation time being 53 minutes in length (Pullen et al. 2000). It is possible that this extended copulation time gives selective influence to the penile morphology, supporting the observations made by Anderson (1998) of differences in penile morphology between G. senegalensis and G. moholi. 9 The nests of G. moholi are built from leaves and form a sleeping platform (Bearder 1969). Old birds’ nests are sometimes used as sleeping sites, with weaver birds’ nests in particular being utilized (Bearder 1969). Nests may be used for months, with favored nests sites being changed at different times of the year (Bearder 1969). Data from captive individuals indicates that nests are built by females and shared with their group, which may include a male and the female’s offspring (Bearder 1969). Sociality Scent marking and vocalizations are often observed in both species of South African Galago (Bearder and Doyle 1972; Bearder et al. 2003; Clark 1985; Welker and Schafer-Witt 1988). O. crassicaudatus males have been shown to group together after a loud call, moving from as far away as 0.3 km (Clark 1985). Juvenile O. crassicaudatus are known to emit a buzzing noise when contact with the foraging group is lost (Bearder et al. 2003). Many individuals of both species have been known to scent mark at specific sites, likely because information such as fertility can be exchanged through scent (Bearder and Doyle 1972; Clark 1985). Although often described as solitary, social interactions do consume a significant portion of O. crassicaudatus nightly activity (Bearder et al. 2003). Social interactions range from grooming and play to agonistic actions taken to cause a conspecific to leave the immediate area (Clark 1985). Groups have been observed foraging together at night, with each group containing a female, her offspring, and occasionally a single male (Bearder et al. 2003). On a nightly basis, the social interactions of O. crassicaudatus occur most frequently at areas of home range overlap where gum trees are present (Clark 1985). Females, on average, have smaller home ranges than males (Clark 1985). Males disperse farther and continue to move 10 later in life than females, resulting in females being the stable unit of socialization (Clark 1985; Welker and Schafer-Witt 1988). The territory of one male G. moholi averages 11 hectares and overlaps with the smaller territory of several females, each averaging 6.7 hectares (Harcourt and Bearder 1989). Dispersal is similar to O. crassicaudatus, with males traveling farther away than females, who remain close to their birth territory (Harcourt and Bearder 1989). Many different calls are used by G. moholi, possibly conveying different meanings ranging from food to territorial calls (Bearder and Doyle 1972). It has been observed that, in captive specimens, a call will be uttered by the dominant animal during play (Bearder and Doyle 1972). In captive G. moholi, all age-class and sex combinations were tolerant of juveniles, with grooming being the most common social interaction followed by displacement and play (Bearder and Doyle 1972; Nash and Flinn 1978). While group formation occurs in captivity, it is noted that G. moholi is on average more solitary than O. crassicaudatus in the wild (Bearder and Doyle 1972). Also, it is noted that aggression seen in captivity may be a result of confinement rather than of a dominance conflict, as in the wild two individuals having conflict could easily separate (Nash and Flinn 1978). Groups of wild G. moholi have been observed in numbers from two to six, with very few observations of animals in groups of four or more (Bearder 1969). Diet The diet of both species of South African bushbaby is similar, consisting of many invertebrates and tree sap (Harcourt 1986). The average height for feeding on Acacia sap was 20 feet for G. moholi and varied for O. crassicaudatus from a low of around five feet during the summer to a high of around 25 feet during the winter (Harcourt 1986). G. moholi has never been reported consuming fruit in its natural habitat, while O. crassicaudatus has been observed 11 feeding on the berries of Zizyphus mucronata and fecal samples have contained Euclea sp. seeds and Combretum sp. flowers (Harcourt 1980; Harcourt 1986; Harcourt and Bearder 1989). O. crassicaudatus has been said to consume vertebrates, including domestic chickens, but this activity is not known from many well studied populations (Bearder 1974; Harcourt 1980). The consumption of invertebrates by both species is similar during the summer months, and O. crassicaudatus has been known to prey on large millipedes of up to 150mm long (Harcourt 1986). O. crassicaudatus consumes larger insects than G. moholi during summer months, although both species feed on similarly sized prey during the winter months. (Harcourt 1980). O. crassicaudatus was observed to reduce its’ consumption of insects during the winter months and instead spent time feeding on gum, likely because fewer insects were available at the study site during the winter (Harcourt 1986). O. crassicaudatus relies more heavily on gum than G. moholi, although both species consume more gum during winter months than during other times of the year (Harcourt 1986). G. moholi was found to feed on gum for around 50 seconds per night during the summer to 3360 seconds during the winter while O. crassicaudatus was found to increase from 40 sec during the summer to 2280 seconds during the winter (Harcourt 1986). However the time spent feeding on gum does not mean the overall intake was greater, as gum becomes more crystalline during the winter. G. moholi may have a more difficult time than O. crassicaudatus taking in this harder winter gum (Harcourt 1986). Locomotion G. moholi is known to utilize leaping as the primary means of locomotion, accounting for over 50 percent of locomotion in one study (Crompton 1980). Climbing accounted for a further 20 percent of locomotor activity, with walking, running, and descending accounting for less than 12 15 percent each (Crompton 1980). Leaping was found to account for approximately 700m for every km traveled, with all other types of locomotion accounting for no more than 150m each (Crompton 1980). The majority of movement occurred from 1933h to 2033h nightly with a sharp decline thereafter (Crompton 1980). Locomotion during the day is rare but has been reported in the winter months (Bearder 1969). This is in line with behavior observed at Loskop Dam Nature Reserve, where several individuals were observed up to one hour before sunset and no animals were observed after 2100h throughout the study period. Conversely, O. crassicaudatus uses walking and climbing as the primary means of locomotion (Crompton 1980). It was found that climbing and walking each account for a third of O. crassicaudatus’ movement, while running makes up 10 percent of all movements (Crompton 1980). Walking accounts for 300m for every km traveled while climbing and running each account for approximately 250m (Crompton 1980). Leaping also accounts for 200m per km traveled (Crompton 1980). The movements of O. crassicaudatus have been said to resemble those of Perodicticus potto, likely because both species utilize similar resources (Crompton 1980). Objectives The purpose of this research was to find the answers to two major questions. First, what behavioral niche does G. moholi occupy? Second, could Loskop Dam Nature Reserve be expected to support a population of O. crassicaudatus? Limitations to this study included difficulty finding or observing the nocturnal primates, potentially small amounts of data being collected over the duration of the study, and limited utility of statistical analyses on the collected data. Also, published accounts of O. crassicaudatus did not provide enough information to determine the suitability of Loskop Dam Nature Reserve 13 as a potential habitat. Contrary to what was expected, G. moholi was not observed in habitat centering exclusively on gum producing trees located in areas more distant from park boundaries and campsites. 14 MATERIALS AND METHODS Study Site Loskop Dam Nature Reserve is a Provincial Nature Reserve located 52 km north of Middleburg in Mpumalanga, South Africa (Barrett et al. 2010; Ferrar and Lotter 2007). Ferrar and Lotter (2007) state that Loskop Dam Nature Reserve is currently 22,900 hectares in size. Barrett et al. (2010) give the location between longitude 29°15’00” and 29°40’00” east and latitude 25°34’00” and 25°56’00” south. Minimum temperature for July averages 8.5°C, with yearly temperatures ranging from -8°C to 40°C (Barrett et al. 2010). Figure 3: Location of the Loskop Dam Nature Reserve in the Mpumalanga Province of South Africa (Barrett et al. 2010:3) 15 Data Collection Transects were driven throughout the tourist road system of Loskop Dam Nature Reserve between the hours of 1800 and 2200 on 12 days between June 29 and July 12, 2011. The system was broken down into segments based on major landmarks, including junctions in the road. The transects included the following road segments: Landberg road, Blesbok loop, T-junction 1 to TJunction 2, T-Junction 2 to picnic site, Camp to T-Junction 1, and the Water Pump road (see Figure 4). Additional transects had been planned but were not performed because access to a four-wheel-drive vehicle could not be secured. Speed varied between four and 13 km per hour with a target speed of 10 km per hour. The following were recorded for each sighting: the degrees off of transect, distance to the individual, a brief description of the tree the individual was in, estimated height in vegetation, and GPS coordinates of the vehicle. Vegetation sample plots were constructed during the following days at the location of each individual (see Figure 5). The center of the plot was the tree in which the individual was sighted. A tape measurer was used to determine 5m from the center tree in all directions. Any vegetation with over one half of its mass within the 5m radius was considered to be inside the plot. Species and diameter at breast height (DBH) were recorded for all woody species over 2.5cm in diameter. Woody species were identified using Schmidt et al. (2002) and Van Wyk and Van Wyk (2011). Canopy cover was determined by using a densitometer at 1m intervals from the center point in each cardinal direction. Data Analyses Transect data was analyzed using six methods presented by the National Research Council in Techniques for the Study of Primate Population Ecology (1981). Mean perpendicular 16 distance, maximum perpendicular distance, and maximum reliable perpendicular distance were used to extrapolate a population estimate for the entire reserve. Mean observer-to-animal distance, maximum observer-to-animal distance, and maximum reliable observer-to-animal distances were also used to estimate the population of the reserve (NRC 1981). Each transect segment was analyzed using the same techniques. However the sample was too small to accurately determine maximum reliable perpendicular or maximum reliable observer-to-animal distances (NRC 1981) Vegetation Analyses Data from the vegetation sample plots were analyzed in order to determine the following: total plants observed, percentage of the total each species accounted for, and the percentage of the total that potential fruiting species accounted for. Percent canopy cover was calculated for each sample plot and placed into four classes, 0 to 25 percent, 30 to 50 percent, 55 to 75 percent, and 80 to 100 percent. DBH data was counted to determine the average DBH of each sample plot and of all plots. The DBH of each identified woody species was also placed into one of five size classes; 2.5 cm to five cm, five cm to 10 cm, 10 cm to 20 cm, 20 cm to 35 cm and greater than 35 cm. This was then analyzed to determine the most prevalent size class for each center tree and across all plots. 17 Figure 4: Transects Labeled By Color Blue: Water Pump Road Green: T-junction 1 to Camp Yellow: Landberg Road Red: T-junction 1 to T-junction 2 Purple: Blesbok Loop Black: T-junction 2 to Picnic Site 18 Figure 5: Location of Each Individual at Sighting 19 RESULTS Species Density The transect segments varied in estimated density from a minimum of 0.58 animals per kilometer to a maximum of 8.5 animals per kilometer (Table 1). The most frequent estimate across all analytical methods was approximately three individuals per km (Table 1). Table 1: Estimated Species Density of G. moholi Along Various Transect Segments (animals/km2) Transect Segment Camp to Tjunction 1 T-junction 1 to T-junction 2 Picnic Road Landberg Road Blesbok Loop Maximum Mean Maximum Mean ObserverPerpendicular Perpendicular Observer-toto-Animal Distance Method Distance Method Animal Distance Distance Method Method 3.735 8.5 3.384 6.895 3.3258 8.4 2.016 5.593 1.27 2.48 1.27 8.147 0.5846 1.521 0.5846 3.722 2.13 2.169 1.11 1.965 The estimated population of G. moholi at Loskop Dam Nature Reserve ranges from a minimum of 196 individuals to a maximum of 1029 individuals depending on the analytical method used (Table 2). The mean perpendicular distance method and mean observer to animal distance method yielded estimates of 1028.9 and 609.0 animals throughout the reserve. The maximum perpendicular distance and maximum observer to animal distance methods provided an estimate of 323.9 and 196.5 animals, respectively. The maximum reliable perpendicular distance method provided an estimate of 772.8 animals with a cut off of 15m, but only 515 animals with a cut off of 30m. The maximum reliable observer to animal distance provided an estimate of 365.1 animals with a cut off of 30m and 296.2 animals with a cut off of 50m. 20 Table 2: Estimated Species Density of G. moholi at Loskop Dam Nature Reserve Analytical Method Maximum Perpendicular Distance Maximum Reliable Perpendicular Distance (15m) Maximum Reliable Perpendicular Distance (30m) Maximum Observer to Animal Distance Maximum Reliable Observer to Animal Distance (30m) Maximum Reliable Observer to Animal Distance (50m) Mean Perpendicular Distance Mean Observer to Animal Distance Estimated Density Estimated Population of (animals/km2) Reserve (animals) 1.58 323.9 3.77 772.85 2.51 515 0.959 196.595 1.781 365.105 1.445 296.225 5.019 2.971 1028.99 609.055 Associated Plant Species Fifty plant species were identified within a 5m radius of each sighting (Table 3). Of these, the most frequently identified belonged to the genus Acacia, Combretum, and Dichrostachys (Table 4). DBH of each woody plant over 2.5cm was measured with multiple stems of the same species being added together to yield one overall DBH. Overall, approximately 29 percent of woody species were between 2.5 and 5.0 cm in DBH, 32 percent were between 5.1 and 10.0 cm DBH, 20 percent were between 20.1 and 35.0 cm DBH, and only six percent were over 35cm DBH (Table 5). However, the DBH trees in which individuals were initially spotted occurred at different frequencies. Approximately 3.1 percent of sightings occurred in the 2.5 to 5.0 cm range, with 9.3 percent occurring in the 5.1 to 10.0 range (Table 6). The 20.1 to 35.0 cm range had the most occurrences, with 46.8 percent, followed by 10.1 to 20.0 with 25 percent of sightings and 15.6 percent in trees with an overall DBH of greater than 35.0 cm. 21 Percent canopy cover ranged from 10 percent to 90 percent. The greatest number (46.8 percent) of sightings occurred in areas with between 30 percent and 50 percent canopy cover (Table 7). Sightings in areas of 55 to 75 percent were also common (21.8 percent), as were 80 percent to 100 percent (28.1 percent). The lowest number of sightings occurred in areas of 0 to 25 percent canopy cover (6.2 percent) Table 3: Plant Species Identified and Number of Each Species Plant Species Acacia burkei Acacia caffra Acacia karoo Acacia nigrescens Acacia nilotica Acacia sp. Acacia tortillis Berchemia zaire Bridellia mollis Burkea Africana Combretum apiculatum Combretum kraussii Combretum molle Combretum zeyheri Corelia ovalis Croton megalobotrys Dichrostachys cinerea Dombeya rotundifolia Dyplorhyncus condylocarpon Ealodendron transvaalense Ehretia amoena Ehretia crispa Ehretia rigida Ehretia obtusfolia Euclea crispa Euclea undulate Individuals Identified 23 1 25 12 10 6 7 1 3 8 52 1 25 18 1 2 193 48 3 28 1 4 4 1 54 1 Flueggea virosa Grewia monticola Grewia flavescens Gymnosporia senegalensis Gymnosporia tenuispina Lippa javanica Lycium cinercum Maytenus undata Ozoroa paniculosa Peltophorum africanum Protea caffra Rhus leptodictia Rhus pyroides Rhus zaire Salvodora persica Sandpaper bushwillow Sclerocarpa birrea Strychnos madagascariensis Strychnos spinosa Terminalia prunoides Teinnea rhodesiana Vangueira parvifolia Warburgia salutaris Ziziphus mucronata unknown Total Plants 3 22 12 1 2 42 1 2 5 1 11 10 13 1 2 1 13 3 2 11 3 1 4 5 32 735 22 Table 4: Major Plant Genus and Percent of Total Major Plant Genus Acacia sp. Combretum sp. Dichrostachys sp. Percentage of Total 11.4754 13.1148 26.3661 Table 5: Percentage of Trees By Size Class Size Class (cm) Percent of All Trees (n=249) 28.9 31.7 20.5 12.9 6.0 2.5 to 5.0 5.1 to 10.0 10.1 to 20.0 20.1 to 35.0 Greater than 35.0 Table 6: Percentage of Center Tree DBH by Size Class Size Class (cm) Percent of Center Trees (n=32) 3.1 9.4 25 46.9 15.6 2.5 to 5.0 5.1 to 10.0 10.1 to 20.0 20.1 to 35.0 Greater than 35.0 Table 7: Percent Canopy Cover by Class Percent Canopy Cover 0 to 25 30 to 50 55 to 75 80 to 100 Percentage of Sample Plots (n=32) 6.3 46.9 21.9 28.1 23 DISCUSSION Species Density The National Research Council of the United States of America (1981) determined the most reliable methods for the maximum reliable observer-to-animal distance method and the maximum reliable perpendicular distance method. These methods were employed to estimate the population density of G. moholi throughout the reserve (Table 1). The maximum reliable perpendicular distance method with a cutoff of 15m yielded an estimate of 3.77 animals per square kilometer, or 772 animals throughout the reserve. If a 30 m cutoff is used, that estimate drops to 2.51 animals per square kilometer or 515 animals throughout the reserve. The maximum reliable observer-to-animal distance method may be the most reliable estimate (NRC 1981). If a cutoff of 30 m is used, the estimated population density is 1.78 animals per square kilometer or 365 animals throughout the reserve. If a cutoff of 50 m is used, the estimate drops to 1.44 animals per square kilometer or 296 animals throughout the reserve. The various methods used yielded a range of estimates of 0.58 individuals per square kilometer to 8.5 individuals per square kilometer (Table 2). According to Techniques for the Study of Primate Population Ecology, both the mean perpendicular distance method and the mean observer to animal distance method yield estimates of density off by as much as 85 percent (NRC 1981). Due to the low number of sightings, it was not possible to determine a maximum reliable sighting distance for each individual transect. The next most reliable methods are the maximum perpendicular distance and the maximum observer-to-animal distance methods (NRC 1981). These yielded estimates of between 3.7 and 0.58 individuals per square kilometer for various transects 24 Regardless of the method used, the estimated density of G. moholi at Loskop Dam Nature Reserve is lower than the estimate of 95 animals per square kilometer reported by Bearder and Doyle (1972). Because of the cold temperatures associated with higher latitudes and the lower activity of G. moholi during winter months, the population of the reserve may be underestimated. It is not likely that the season alone accounts for the substantial difference in population estimates. Rather, the population at Loskop Dam Nature Reserve is likely under significantly increased ecological stress compared populations at lower latitudes. Habitat Preference The habitat preference of G. moholi could not be statistically demonstrated due to a lack of published information on the areas of the reserve where G. moholi was observed. However, the majority of sightings occurred in areas where Dichrostachys cinerea, Acacia sp., or Combretum sp. were present (Table 4). Filmalter (2010) surveyed the vegetation in the Honderkraal section of Loskop Dam Nature Reserve and Barrett et al. (2010) surveyed the vegetation in the home ranges of Chlorocebus pygerythrus F. Cuvier 1821. Neither of these areas overlapped with the observed locations of G. moholi. The majority of sightings occurred in areas of 30 to 50 percent canopy cover, but this may represent observer bias rather than the preference of G. moholi for more open areas. G. moholi may prefer to locomote between trees of at least 10.0 cm in DBH, as only nine percent of sightings occurred on trees of below 10.0 cm DBH. This is not likely due to observer bias, as sighting an individual on a smaller tree was no more difficult than spotting in a larger tree (Figure 2). On July fifth at approximately 16:40h, an individual G. moholi was observed consuming the fruit of a Dichrostachys cinerea (Figure 3). The individual consumed the entire seed pod 25 before moving out of observable range. Because of the marginal habitat available at Loskop Dam Nature Reserve, this may represent a unique dietary adaptation used by the population around the University of South Africa Research Camp. 26 Figure 4: G. moholi consuming the fruit of D. cinerea 27 CONCLUSION The density of G. moholi at Loskop Dam Nature Reserve is extremely low compared to the density observed in other areas (Bearder 1972; Bearder and Doyle 1974; Harcourt and Bearder 1989). This may indicate the need for a species management plan to ensure G. moholi does not become locally extinct. Additional population surveys are needed to determine the population density in areas of the reserve not surveyed by the present study. The consumption of vegetative matter by G. moholi is a previously unobserved phenomenon and may have significant implications as a fallback food source. The population of G. moholi at Loskop Dam Nature Reserve may be exploiting a different niche than other populations of the same species due to the elevation and the location of the reserve at the edge of the species’ range. O. crassicaudatus may be capable of surviving at Loskop Dam Nature Reserve, but may be out competed by G. moholi due to increased niche overlap. Additionally, the high percentage of Combretum sp. found in the vegetation sample plots may indicate that Dichrostachys cinerea is not the only vegetative matter being consumed by G. moholi. Many Combretum sp. were observed to be fruiting during the study period. Further study is needed to demonstrate the extent to which G. moholi is utilizing vegetative matter as a food source. Although outside the current range of O. crassicaudatus, Loskop Dam Nature Reserve was hypothesized to be capable of supporting a population of O. crassicaudatus as well as G. moholi. An analysis of the potential impact of the introduction of O. crassicaudatus would have been needed if Loskop Dam Nature Reserve was found to be capable of supporting a population. However, the low population of G. moholi combined with the observation of the ingestion of fruit likely indicates that Loskop Dam Nature Reserve is suboptimal habitat for G. moholi, and would not be expected to support a population of the larger O. crassicaudatus. 28 REFERENCES CITED Anderson, Matthew J. 1998 Comparative Morphology and Speciation in Galagos. Folia Primatologica 69(1):325-331. Barrett, Alan S., Leslie R. Brown, Lousie Barrett, and Peter Henzi 2010 A Floristic Description and utilization of Two Home Ranges by Vervet Monkeys in Loskop Dam Nature Reserve, South Africa. Koedoe 52(1):1-12. Bearder, S. K. 1969 Territorial and Intergroup Behaviour of the Lesser Bushbaby, Galago senegalensis moholi in Semi-Natural Conditions and in the Field. MsC thesis, University of the Witwaterstrand. Bearder, Simon Kenneth 1974 Aspects of the Ecology and Behaviour of the Thick-Tailed Bushbaby Galago crassicaudatus. PhD thesis, University of the Witwaterstrand. Bearder, Simon K., Lesley Ambrose, Caroline Harcourt, Paul Honess, Andrew Perkin, Elizabeth Pimley, Samantha Pullen, and Nadien Svoboda 2003 Species-Typical Patterns of Infant Contact, Sleeping Site Use and Social Cohesion among Nocturnal Primates in Africa. Folia Primatologica 74:337-354. Bearder, Simon K. and G. A. Doyle 1972 Field and Laboratory Studies of Social Organization in Bushbabies (Galago senegalensis). Journal of Human Evolution 3:37-50. Bearder, S.K., and G.A. Doyle 1974 Ecology of Bushbabies Galago senegalensis and Galago crassicaudatus. In Prosimian Biology. Martin, Robert D., Gerald A. Doyle, and Alan C. Walker, eds. Pp. 109-130. Pittsburg: University of Pittsburg Press. Clark, Anne B. 1985 Sociality in a Nocturnal “Solitary” Prosimian: Galago crassicaudatus. International Journal of Primatology 6(6):581-600. Colwell, Robert K., and Thiago F. Rangel 2009 Huchinson’s duality: The once and future niche. Proceedings of the national Academy of Sciences 106(2):19651-19658. Crompton, Robin H. 1980 A Leap in the Dark: Locomotor Behaviour and Ecology in Galago senegalensis and Galago crassicaudatus. PhD thesis. Harvard University. 29 Darwin, Charles. 2009 The Origin of Species. 150th Anniversary ed. Alachua, FL: Bridge Logos. Originally published as On the origin of species by means of natural selection or the preservation of favored races in the struggle for life (London: John Murray, 1859). Dixon, A.F. 1989 Sexual Selection, Genital Morphology and Copulatory Behavior in Male Galagos. International Journal of Primatology 10(1):47-55. Doyle, G.A., A. Pelletier, and T. Bekker 1967 Courtship, Mating and Partuition in the Lesser Bushbaby (Galago senegalensis moholi) Under Semi-Natural Conditions. Folia Primatologica 7:169-197. Ferrar, Tony A., and Mervyn C. Lotter 2007 Mpumalanga Biodiversity Conservation Plan Handbook. Mpumalanga Parks and Tourism Agency, Nelspruit. Filmalter, Nicolene 2010 A Vegetation Classification and Management Plan for the Honderkraal Section of the Loskop Dam Nature Reserve. MTech dissertation. University of South Africa. Harcourt, Caroline Susan 1980 Behavioural adaptations in South African Galagos. MsC dissertation. University of the Witwaterstrand. Harcourt, Caroline 1986. Seasonal Variation in the Diet of South African Galagos. International Journal of Primatology 7(5):491-506. Harcourt, Caroline S., and Simon K. Bearder 1989 Comparison of Galago moholi in South Africa with Galago zanzibaricus in Kenya. International Journal of Primatology 10(1):35-45. Huxley, J.S. 1939 Ecology and Taxonomic Differentiation. Journal of Ecology 27(5):408-420. Izard, M. Kay, and Elwyn L. Simons 1986 Infant Survival and Liter Size in Primigravid and Multigravid Galagos. Journal of Medical Primatology 15:27-35. Lawes, M. J. 2005 XVI. Family Galagidae. In The Mammals of the Southern African Subregion. John D. Skinner and Christian T. Chimimba eds. Pp 210-217. Cape Town: Cambridge University Press. 30 Masters, Judith C., Michele Boniotto, Sergeio Crovella, Christian Roos, Lucia Pozzi, and Massimiliano Delpero 2007 Phylogenetic Relationships Among the Lorisoidea As Indicated by Craniodental Morphology and Mitochondrial Sequence Data. American Journal of Primatology 69:6-15. Masters, Judith C., and Nigel P. Bragg 2000 Morphological Correlates of Speciation in Bush Babies. International Journal of Primatology 21(5):793-813. Nash, Leanne T., Simon K. Bearder, and Todd R. Olson 1989 Synopsis of Galago Species Characteristics. International Journal of Primatology 10(1):57-80. Nash, Leanne T., and Lynn Flinn 1978 Group Formation in Captive Lesser Galagos (Galago senegalensis). Primates 19(3):493-503. National Research Council Subcommittee on Conservation of Natural Populations 1981 Techniques for the Study of Primate Population Ecology. Washington, D.C: National Academy Press. Pullen, S.L., S.K. Bearder, and A.F. Dixson 2000 Preliminary Observations on Sexual Behavior and the mating System in Freeranging Lesser Galagos (Galago moholi). American Journal of Primatology 51:78-88 Ravosa, Matthew J., Ashley N. Daniel, and Destiny B. Costly 2010 Allometry and Evolution in the Galago Skull. Folia Primatologica 81:177-196. Schmidt, Ernst, Mervyn Lotter, and Warren McCleland 2002 Trees and Shrubs of Mpumalanga and Kruger National Park. Johannesburg: Jacana Media. Seiffert, Erik R., Elwyn L. Simons, and Yousry Attia 2003 Fossil evidence for an ancient divergence of lorises and galagos. Nature 44:421424. Skinner, John D, and Christain T. Chimimba 2005 Explainatory Notes. In The Mammals of the Southern African Subregion. John D. Skinner and Christian T. Chimimba eds. Pp XII. Cape Town: Cambridge University Press. Stankiewicz, J., C. Thiart, J.C. Masters, and M.J. de Wit 2006 Did lemurs have sweepstake tickets? An exploration of Simpson’s model for the colonization of Madagascar by mammals. Journal of Biogeography 33:221-235. 31 Stephenson, Isobel R., Simon K. Bearder, Guiseppe Donati, and Johann Karlsson 2010 A Guide to Galago diversity: Getting a Grip on How Best to Chew Gum. In: The Evolution of Exudativory in Primates. Anne M. Burrows and Leanne T. Nash, eds. Pp 235-255. New York: Springer Science. Tartabini, Angelo 1991 Mother-infant Cannibalism in Thick-tailed Bushbabies (Galago crassicaudatus umbrosus). Primates 32(3): 379-383. Van Wyk, Braam, and Piet Van Wyk 2011 Field Guide to Trees of Southern Africa. Cape Town: Struik Publishers Wake, David B., Elizabeth A. Hadly, and David D. Ackerly 2009 Biogeography, changing climates, and niche evolution. Proceedings of the National Academy of Sciences 106(2):19631-19636. Welker, C., and C. Schaver-Witt 1988 Preliminary Observations on Behavioral Differences Among Thick-Tailed Bushbabies. International Journal of Primatology 9(6):507-518. Ying, K.L., and H. Butler 1971 Chromosomal Polymorphism in the Lesser Bush Babies (Galago senegalensis). Canadian Journal of Genetics and Cytology 13:793-800. 32 Transect Start Time Start Location Route Sighting Transect Start Time Start Location 1 19:46 S E Water Pump Road None 2 20:21 Route S E Blesbok Loop Sighting None Transect Start Time Start Location 25°24.5431' 29°20.9685' Appendix 1: Transect Data Date 6/30/2011 End Time 19:56 End Location S 25°24.0880' E 29°21.0390' Distance 873m 25°25.1297' 29°17.6832' Route Sighting None 6/30/2011 20:58 S E 25°25.2865' 29°17.9593' 5.27km Distance 3 21:02 S E T2 to camp Date End Time End Location 25°25.1263' 29°17.6813' Date End Time End Location Distance 6/30/2011 22:02 S E 25°24.4571' 29°21.0448' 3.5km 33 Transect Start Time Start Location Route Sighting Transect Start Time Start Location Route 4 Date 18:55 End Time S 25°25.1293' End Location E 29°17.6791' Blesbok Loop Distance Time GPS Coordinates 1 19:10 S 25°25.2902' 2 19:16 S 25°25.8526' 5 19:55 S E T2 to T1 25°25.1257' 29°17.6782' Time Sighting Transect Start Time Start Location Route 3 25°24.4377' 29°19.0889' Time Sighting Distance GPS Coordinates 20:18 S 25°24.6606' 6 20:45 S E Langberg 4 5 Date End Time End Location Date End Time End Location Distance GPS Coordinates 21:11 S 25°23.7967' 21:36 S 25°24.2260' 7/1/2011 19:45 S E 2.59km 25°25.2902' 29°17.9873' Distance E 29°18.0666' E 29°18.1345' 43m 25m Degrees off transect 272° 245° Height (est) 10m 2m 7/1/2011 S E 3.51km 25°24.4377' 29°19.0892' Distance E 29°19.0892' 18m Degrees off transect 338° Height (est) 1m 7/1/2011 22:11 S E 7.28km 25°24.6541' 29°20.7159' Distance E 29°19.6454' E 29°20.7792' 15m 43m Degrees off transect 35° 284° Height (est) 2m 2.5m 34 Transect Start Time Start Location Route 7 17:58 S E Langberg 25°24.6568' 29°20.7196' Time Sighting Transect Start Time Start Location Route 6 7 25°24.4367' 29°19.0917' Time Sighting Transect Start Time Start Location Distance GPS Coordinates 18:02 S 25°24.4770' 18:15 S 25°24.4770' 8 19:05 S E T1 to T2 8 Route Sighting None Date End Time End Location Distance GPS Coordinates 19:40 S 25°24.7977' 9 19:40 S E Blesbok Loop Date End Time End Location 25°25.1249' 29°17.6793' Date End Time End Location Distance 7/3/2011 18:56 S E 7.28km 25°24.4367' 29°19.0917' Distance E 29°20.8126' E 29°20.8126' 60m 36m Degrees off transect 0° 7° Height (est) 8m 3m 7/3/2011 19:30 S E 3.44km 25°25.1289' 29°17.6805' Distance E 29°18.8458' 15m 7/3/2011 20:10 S E 5.19km 25°25.2978' 29°17.9941 Degrees off transect 123° Height (est) 15m 35 Transect Start Time Start Location 10 20:10 Route S E T1 to Camp Sighting None Transect Start Time Start Location Route 25°24.4518' 29°19.0691' Distance 11 17:51 S E Blesbok Loop 25°25.1291' 29°17.6826' Time Sighting Transect Start Time Start Location 9 Route Sighting None Date End Time End Location Distance GPS Coordinates 18:20 S 25°25.1446' 12 18:36 S E Picnic In Date End Time End Location 25°25.1306' 29°17.6755' Date End Time End Location Distance 7/3/2011 20:54 S E 4.49km 25°24.4270' 29°21.0113' 7/4/2011 18:28 S E 5.29km 25°25.2924' 29°17.9850' Distance E 29°18.4181' 5m 7/4/2011 19:22 S E 7km 25°26.9066' 29°15.7612' Degrees off transect 323° Height (est) 0.5m 36 Transect Start Time Start Location Route 13 19:59 S E T2 to T1 25°25.1262' 29°17.6779' Time Sighting Transect Start Time Start Location Route 10 25°24.4368' 29°19.1003' Time Sighting Transect Start Time Start Location Distance GPS Coordinates 20:16 S 25°24.552' 14 20:29 S E T1 to Camp 11 Route Sighting None Date End Time End Location Distance GPS Coordinates 20:48 S 25°24.6681' 15 21:06 S E Waterpump Date End Time End Location 25°24.5093' 29°20.9728' Date End Time End Location Distance 7/4/2011 20:25 S E 3.49km 25°24.4386' 29°19.1005' Distance E 29°19.1752' 95m Degrees off transect 355° Height (est) 8m 7/4/2011 21:00 S E 4.54km 25°24.4573' 29°21.0447' Distance E 29°20.6992' 25m 7/4/2011 21:12 S E 866m 25°24.0870' 29°21.0402' Degrees off transect 75° Height (est) 2m 37 Transect Start Time Start Location Route 16 18:00 S E Camp to T1 25°24.4598' 29°21.0468' Time Sighting Transect Start Time Start Location Route 12 13 25°24.4385' 29°19.0900' Time Sighting Transect Start Time Start Location Distance GPS Coordinates 18:07 S 25°24.8473' 18:20 S 25°25.0282' 17 18:47 S E T1 to T2 14 15 16 Route Sighting None Date End Time End Location Distance GPS Coordinates 18:48 S 25°24.4531' 19:00 S 25°24.6125' 19:30 S 25°24.9950' 18 19:43 S E Blesbok Loop Date End Time End Location 25°25.1270' 29°17.6824' Date End Time End Location Distance 7/6/2011 18:43 S E 4.56km 25°24.4369' 29°19.0898' Distance E 29°20.4168' E 29°20.3030' 33m 13m Degrees off transect 343° 90° Height (est) 1.5m 2m 7/6/2011 19:47 S E 3.47km 25°25.1304' 29°17.6825' Distance E 29°19.0862' E 29°19.0329' E 29°17.9791' 15m 19m 68m 7/6/2011 20:15 S E 5.16km 25°25.2938' 29°17.9918' Degrees off transect 321° 338° 122° Height (est) 1m 3m 1.5m 38 Transect Start Time Start Location 19 20:23 Route S E Picnic Out Sighting None Transect Start Time Start Location Route Sighting None Route Distance 20 21:55 S E Langberg in Transect Start Time Start Location 25°25.1273' 29°17.6814' 25°24.4356 29°19.0917 Date End Time End Location Distance 21 18:10 S E Langberg out 25°24.6514' 29°20.7152' Time Sighting Date End Time End Location 17 Date End Time End Location Distance GPS Coordinates 18:16 S 25°24.3271' 7/6/2011 21:17 S E 7.09km 25°26.9012' 29°15.7578' 7/6/2011 22:35 S E 7.15km 25°24.6542' 29°20.7198' 7/7/2011 19:11 S E 7.19km 25°24.4355' 29°19.0915' Distance E 29°20.7715' 19m Degrees off transect 285° Height (est) 1.5m 39 Transect Start Time Start Location Route 22 19:14 S E T1 to T2 25°24.4368' 29°19.0924' Time Sighting Transect Start Time Start Location 18 19 Route Sighting None Transect Start Time Start Location Route Distance GPS Coordinates 19:31 S 25°24.8282' 19:42 S 25°24.9700' 23 20:01 S E Picnic Out 25°25.1263' 29°17.6771' Date End Time End Location Distance 24 18:10 S E Camp to T1 25°24.5218' 29°21.0158' Time Sighting Date End Time End Location 20 21 22 23 18:15 18:28 18:34 18:42 Date End Time End Location Distance GPS Coordinates S 25°24.6516' S 25°24.7895' S 25°24.8258' S 25°25.0305' 7/7/2011 19:59 S E 3.47km 25°25.1278' 29°17.6758' Distance E 29°18.8129' E 29°18.3440' 24m 39m Degrees off transect 60° 310° Height (est) 2.5m 0.5m 7/7/2011 21:02 S E 7.17km 25°26.9057' 29°15.7613' 7/8/2011 19:08 S E 25°24.4367' 29°19.0918' 5.21 Distance E 29°20.7517' E 29°20.4585' E 29°20.4275' E 29°20.2947' 11m 44m 14m 55m Degrees off transect 80° 55° 45° 65° Height (est) 0.5m 7m 0.5m 3m 40 Transect Start Time Start Location Route 25 19:17 S E Langberg In 25°24.4383' 29°19.0921' Time Sighting Transect Start Time Start Location Route 24 25 26 25°26.9014' 29°15.7463' Time Sighting Transect Start Time Start Location Route 27 S E Waterpump 25°24.5056' 29°20.9710' 28 Date End Time End Location Distance GPS Coordinates 18:45 S 25°26.0207' 27 19:45 Time Sighting Distance GPS Coordinates 19:55 S 25°24.0658' 19:55 S 25°24.0658' 20:10 S 25°24.3191' 26 18:25 S E Picnic in Date End Time End Location Date End Time End Location Distance GPS Coordinates 19:52 S 25°24.3478' 7/8/2011 20:20 S E 7.22km 25°24.6542' 29°20.7187 Distance E 29°20.7500' E 29°20.7500' E 29°20.7726' 5m 18m 20m Degrees off transect 270° 50° 313° Height (est) 4m 0m 4.5m 7/9/2011 19:21 S E 7.41km 25°25.1269' 29°17.6760' Distance E 29°15.4519' 53m Degrees off transect 10° Height (est) 1.5m 7/9/2011 20:03 S E 873m 25°24.0874' 29°21.0374' Distance E 29°21.0374' 39m Degrees off transect 310° Height (est) 3m 41 Transect Start Time Start Location Route 28 18:19 S E Langberg Out 25°24.6534' 29°20.7198' Time Sighting Transect Start Time Start Location 29 Route Sighting None Transect Start Time Start Location Distance GPS Coordinates 18:32 S 25°23.9261 29 19:26 S E Picnic In 25°25.1248' 29°17.6760' Route Sighting None Date End Time End Location Distance 30 18:27 S E Picnic In Date End Time End Location 25°26.9031' 29°15.7570' Date End Time End Location Distance 7/10/2011 19:12 S E 7.37km 25°24.4360' 29°19.0912' Distance E 29°19.0912' 68m 7/10/2011 20:13 S E 7.09km 25°26.9033' 29°15.7613' 7/11/2011 19:11 S E 7.00km 25°25.1280' 29°17.6759' Degrees off transect 350° Height (est) 0m 42 Transect Start Time Start Location 31 19:20 Route S E Blesbok Loop Sighting None Transect Start Time Start Location 25°25.1288' 29°17.6778' Distance 32 18:03 Route S E Picnic In Sighting None Date End Time End Location 25°26.8969' 29°15.7492' Date End Time End Location Distance 7/11/2011 19:52 S E 5.2km 25°25.2955' 29°17.9920' 7/12/2011 18:43 S E 6.97km 25°25.1266' 29°17.6786' 43 Appendix 2: Vegetation Sample Plot Data Sighting GPS Center Point Center Tree Strychnos spinosa 1 S 25°25.8233' E 29°18.0700' 45% Hits Misses DBH (cm) 125.1 Other Species Rhus leptodyctia Rhus leptodyctia Dichrostachys cinerea Grewia monticola Maytenus undata Maytenus undata Burkea africana DBH (cm) 7.1 10 5.8 4.2 8.8 6.8 6 Sighting GPS Center Point Center Tree Burkea africana 2 S 25°25.8388' E 29°18.1277' 35% Hits Misses DBH (cm) 18.2 Other Species Burkea africana Burkea africana Burkea africana Burkea africana Burkea africana Burkea africana DBH (cm) 28.1 9.2 12.3 30.7 22.8 10.2 Sighting GPS Center Point 3 S 25°24.6514' E 29°19.0021 65% Hits Misses DBH (cm) 35 Percent Canopy Cover Percent Canopy Cover Percent Canopy Cover Center Tree Grewia monticola 9 11 7 13 13 7 44 Other Species Combretum molle Elaeodendron transvaalense Elaeodendron transvaalense Grewia monticola Euclea crispa Rhus leptodictea Rhus leptodictea Acacia karoo Acacia karoo Acacia karoo Unknown Unknown DBH (cm) 9.7 6.9 2.8 4 4 12.6 1.1 15.5 22.1 18.7 2.8 7.6 Sighting GPS Center Point Center Tree Diplorhyncus condylocarpon 4 S 25°23.7917' E 29°19.6422' 25% Hits Misses DBH (cm) 18.3 Other Species Combretum apiculatum Combretum apiculatum Sclerocarya birrea Grewia monticola Grewia monticola Acacia burkei Acacia burkei Acacia burkei Combretum apiculatum DBH (cm) 1.7 too small 4.6 too small too small too small too small 25.4 3 Sighting GPS Center Point 5 S 25°24.2316' E 29°20.7997 50% Hits Misses DBH (cm) 15.7 Percent Canopy Cover Percent Canopy Cover Center Tree Sclerocarya birrea 5 15 10 10 45 Other Species Grewia flavescense Ozora sphaerocarpa Sclerocarya birrea Bridelia mollis Combretum apiculatum Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea DBH (cm) too small 6.4 8.4 29.5 35.2 too small 2.5 too small 11.6 2.5 3.1 Sighting GPS Center Point Center Tree Acacia caffra 6 S 25°24.4482' E 29°20.8226' 70% Hits Misses DBH (cm) 32.2 Other Species Combretum molle Dichrostachys cinerea Dead tree DBH (cm) 45.1 6.5 16.5 Sighting GPS Center Point Center Tree Ehretia obtusifolia 7 S 25°24.0576' E 29°20.7453' 80% Hits Misses DBH (cm) 36 Other Species Combretum molle Combretum molle Combretum molle Combretum molle Combretum molle DBH (cm) 2.7 46.5 6 too small too small Percent Canopy Cover Percent Canopy Cover 14 6 16 4 46 Euclea crispa Euclea crispa Euclea crispa Euclea crispa Grewia monticola Grewia monticola Grewia monticola Grewia monticola Rhus leptodictea Rhus leptodictea Acacia karoo Elaeodendron transvalense Dichrostachys cinerea Sighting GPS Center Point too small too small too small too small too small too small too small too small 30.2 2.5 4.4 too small too small Center Tree Combretum molle 8 S 25°24.7864' E 29°18.8492' 85% Hits Misses DBH (cm) 41.5 Other Species Combretum molle Combretum molle Combretum molle Rhus leptodictea Rhus leptodictea Dichrostachys cinerea Protea caffra DBH (cm) 7 2.7 too small too small too small too small 14.3 Sighting GPS Center Point Center Tree Acacia karoo 9 S 25°25.1429 E 29°18.4347 40% Hits Misses DBH (cm) 8.7 Other Species Acacia nilotica DBH (cm) 38.4 Percent Canopy Cover Percent Canopy Cover 17 3 8 12 47 Acacia nilotica Acacia nilotica Acacia nilotica Acacia nilotica Acacia nilotica Acacia nilotica Euclea crispa Euclea crispa Acacia burkei Acacia tortillis Acacia tortillis Acacia tortillis Dichrostachys cinerea Sighting GPS Center Point 4 9.9 10.6 9.5 13.2 2.6 too small too small 7.2 11 13.4 too small 8.3 Center Tree Acacia burkei 10 S 25°24.5182' E 29°19.1461' 70% Hits Misses DBH (cm) 29.8 Other Species Grewia monticola Acacia burkei Euclea crispa Euclea crispa Ziziphus mucronata Ziziphus mucronata Ziziphus mucronata Ziziphus mucronata Terminalia prunoides Terminalia prunoides Terminalia prunoides Terminalia prunoides Terminalia prunoides Terminalia prunoides Terminalia prunoides Terminalia prunoides Terminalia prunoides Terminalia prunoides DBH (cm) 20.8 too small too small too small 2.7 too small 7.9 5.2 2.8 too small too small 2.5 too small too small too small too small 5 7.3 Percent Canopy Cover 14 6 48 Grewia flavecens Grewia flavecens Sighting GPS Center Point too small too small Center Tree Acacia nigrescens 11 S 25°24.6740' E 29°20.7040' 50% Hits Misses DBH (cm) 28.4 Other Species Acacia nigrescens Acacia nigrescens Combretum molle Combretum apiculatum Lippa javanica Lippa javanica Dombeya rotundifolia Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea DBH (cm) 21.8 6.4 too small too small too small too small 4.4 too small too small too small too small too small too small too small too small too small too small too small 7.4 3.5 Sighting GPS Center Point 12 S 25°24.5647' E 29°20.4156' 40% Hits Misses DBH (cm) 13.2 Percent Canopy Cover Percent Canopy Cover Center Tree Combretum zeyheri 10 10 8 12 49 Other Species Combretum zeyheri Combretum zeyheri Combretum zeyheri Combretum zeyheri Combretum zeyheri Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Gymnosporia senegalensis Ozoroa paniculosa Acacia burkei DBH (cm) 21.9 5.6 8.9 5.9 14 2.8 3.4 too small too small too small too small too small too small too small too small 4.5 3 24.3 Sighting GPS Center Point Center Tree Combretum zeyheri 13 S 25°25.0235' E 29°20.3015' 45% Hits Misses DBH (cm) 23 Other Species Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Grewia monticola Lippa javanica Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Euclea crispa DBH (cm) 19.8 16.2 18.6 too small too small too small too small too small too small too small too small Percent Canopy Cover 9 11 50 Sighting GPS Center Point Center Tree Acacia karoo 14 S 25°24.4608' E 29°19.0876' 65% Hits Misses DBH (cm) 31.5 Other Species Acacia sp. Acacia sp. Acacia sp. Acacia sp. Acacia sp. Acacia sp. Unknown Unknown DBH (cm) 3 too small too small 4.8 6.7 6.4 3.5 10.8 Sighting GPS Center Point Center Tree Acacia karoo 15 S 25°24.6185' E 29°19.0235 45% Hits Misses DBH (cm) 32.5 Other Species Grewia monticola Grewia monticola Grewia monticola Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Acacia karoo Acacia karoo Acacia karoo Euclea crispa DBH (cm) 11.5 too small 9.4 4.5 6 8.7 too small too small 2.2 5.9 Percent Canopy Cover Percent Canopy Cover 13 7 9 11 51 Sighting GPS Center Point Center Tree Protea caffra 16 S 25°24.9654' E 29°18.0033' 10% Hits Misses DBH (cm) 8.4 Other Species Vangueira parvifolia Sclerocarya birrea Sclerocarya birrea Sclerocarya birrea Protea caffra DBH (cm) too small too small too small 26.9 too small Sighting GPS Center Point Center Tree Combretum zeyheri 17 S 25°24.3253' E 29°20.7630' 85% Hits Misses DBH (cm) 20 Other Species Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Acacia nigrescens Acacia nigrescens Acacia nigrescens Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Bridelia mollis Bridelia mollis Salvadora persica DBH (cm) 4.9 too small 11.4 2.8 too small 4 too small 7.3 too small too small too small too small too small too small 11.4 3.8 too small Percent Canopy Cover Percent Canopy Cover 2 18 17 3 52 Salvadora persica Flueggea virosa Cordia ovalis Tinnea rhodesiana Tinnea rhodesiana Tinnea rhodesiana Gymnosporia tenuispina Gymnosporia tenuispina Ehretia amoena Combretum zeyheri Acacia burkei Elaeodendron transvaalense Elaeodendron transvaalense Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Unknown Unknown Sighting GPS Center Point too small too small 19 too small too small too small 2.8 too small too small 13.5 26.1 too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small 11.5 too small Center Tree Acacia burkei 18 S 25°24.8283' E 29°18.8042' 60% Hits Misses DBH (cm) 19.1 Other Species Combretum apiculatum DBH (cm) 28.3 Percent Canopy Cover 12 8 53 Combretum apiculatum Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Rhus pyroides Rhus pyroides Strychnos madagascariensis Strychnos madagascariensis Dichrostachys cinerea Dichrostachys cinerea Combretum molle Combretum molle Combretum molle Elaeodendron transvaalensis Acacia karoo Dyplorhyncus condylocarpon Sighting GPS Center Point 4 too small too small too small too small too small too small too small 7.1 19.7 2.9 20.8 too small too small too small too small too small 6.6 4.8 6.3 too small too small too small too small 99.4 Center Tree Euclea crispa 19 S 25°24.9931' E 29°18.3405' 50% Hits Misses DBH (cm) 7 Other Species Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Acacia karoo DBH (cm) too small too small too small too small too small 30.2 Percent Canopy Cover 10 10 54 Acacia karoo Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Dichrostachys cinerea Unknown dead tree Sighting GPS Center Point too small too small too small too small too small too small too small too small too small 5.6 12.2 Center Tree Combretum zeyheri 20 S 25°24.6407' E 29°20.7522' 80% Hits Misses DBH (cm) 13.4 Other Species Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Acacia nilotica Acacia nilotica Acacia burkei Acacia burkei Acacia burkei Acacia burkei Sclerocarya birrea Sclerocarya birrea Combretum zeyheri Dichrostachys cinerea DBH (cm) 10.5 5.2 6.7 9 too small too small too small too small 8.9 9.3 13.9 too small 31.7 8.3 3.6 20.7 9 29.9 too small 29.6 Percent Canopy Cover 16 4 55 Dichrostachys cinerea Dichrostachys cinerea Ozora paniculosa Acacia karoo Combretum kraussii Rhus pyroides Unknown Unknown Unknown Unknown Sighting GPS Center Point too small too small 5.8 5 too small too small too small too small too small too small Center Tree Combretum apiculatum 21 S 25°24.7848' E 29°20.4343' 40% Hits Misses DBH (cm) 29.7 Other Species Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea DBH (cm) 21.5 11 too small 6.5 13.2 2.5 3.4 9.2 too small too small too small too small too small too small too small too small too small too small too small too small too small Percent Canopy Cover 8 12 56 Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Sclerocarya birrea Combretum zeyheri Combretum zeyheri Unknown Unknown Unknown Sighting GPS Center Point too small too small too small 12.7 7.4 14.8 2.5 18.2 too small 14.3 too small 2.9 too small too small Center Tree Dombeya rotundifolia 22 S 25°24.8258' E 29°20.4275' 30% Hits Misses DBH (cm) 4.5 Other Species Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Acacia burkei Acacia burkei Elodendron transvalensis Elodendron transvalensis Elodendron transvalensis Rhus pyroides Acacia karoo Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum DBH (cm) 7.9 4.8 too small too small 17.3 21 too small too small too small too small 4.1 too small too small too small too small too small too small Percent Canopy Cover 6 14 57 Combretum apiculatum Ehretia rigida Ehretia rigida Euclea crispa Euclea crispa Sclerocarya birrea Lippa javanica Unknown Unknown Unknown Unknown Sighting GPS Center Point 8.3 too small too small too small too small 21.9 too small 4.1 too small too small too small Center Tree Combretum zeyheri 23 S 25°25.0247' E 29°20.2721' 40% Hits Misses DBH (cm) 28.2 Other Species Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea DBH (cm) too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small Percent Canopy Cover 8 12 58 Dichrostachys cinerea Dichrostachys cinerea Grewia monticola Grewia monticola Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Lippa javanica Combretum zeyheri Combretum zeyheri Euclea crispa Dombeya rotundifolia Sighting GPS Center Point too small too small 10 too small too small too small too small too small too small too small too small 27.5 26.6 too small 22.2 Center Tree Terminalia prunoides 24 S 25°24.0646' E 29°20.7518' 60% Hits Misses DBH (cm) 22 Other Species Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Euclea crispa Euclea crispa Euclea crispa Euclea crispa DBH (cm) too small too small too small too small too small 48.9 9.7 21.7 too small too small too small too small 4.5 too small too small too small Percent Canopy Cover 12 8 59 Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Grewia monticola Grewia monticola Grewia monticola Grewia monticola Combretum molle Eleodendron transvaalensis Eleodendron transvaalensis Euclea undulata Berchemia zaire Ozoroa paniculosa Acacia karoo Acacia karoo Acacia karoo Acacia karoo Rhus pyroides Acacia burkei Acacia burkei Warburgia saltaris Sighting GPS Center Point too small too small too small too small too small 16.9 8.1 too small too small 44.8 too small too small too small too small too small 6 too small too small too small too small too small too small too small Center Tree Combretum zeyheri 25 S 25°24.0658' E 29°20.7500 40% Hits Misses DBH (cm) 32.1 Other Species Croton menyharthii Croton menyharthii Ziziphus mucronata Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Acacia karoo DBH (cm) too small too small too small 7.3 too small too small too small 21.5 Percent Canopy Cover 8 12 60 Acacia karoo Acacia karoo Euclea crispa Euclea crispa Euclea crispa Rhus pyroides Combretum apiculatum Sighting GPS Center Point 13.4 too small too small too small too small too small 29.4 Center Tree Combretum molle 26 S 25°24.3323' E 29°20.7814' 80% Hits Misses DBH (cm) 32.9 Other Species Grewia flavescens Grewia flavescens Grewia flavescens Grewia flavescens Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Dombeya rotundifolia Diplorhyncus condylocarpon Acacia karoo Acacia karoo Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea DBH (cm) too small too small too small too small too small too small too small too small too small too small 34.7 too small too small 2.5 20.6 4.6 3.6 5.3 too small too small too small too small too small too small Percent Canopy Cover 16 4 61 Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Dichorstachys cinerea Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Acacia burkei Combretum zeyheri Lippa javanica Rhus zaire Rhus zaire Grewia monticola Warburgia salutaris Warburgia salutaris Warburgia salutaris Flueggea virosa Flueggea virosa Combretum molle Unknown Sighting GPS Center Point too small too small too small too small too small too small too small too small too small too small too small too small 2.6 too small too small too small too small 34.9 too small too small too small 10.8 too small too small too small too small too small 41 too small Center Tree Acacia karoo 27 S 25°26.0198' E 29°15.4815' 90% Hits Misses DBH (cm) 92.6 Other Species Rhus pyroides Rhus pyroides DBH (cm) too small too small Percent Canopy Cover 18 2 62 Rhus pyroides Rhus pyroides Rhus pyroides Combretum apiculatum Euclea crispa Euclea crispa Euclea crispa Euclea crispa Dichrostachys cinerea Rhus leptodictea Rhus leptodictea Elaeodendron transvaalensis Unknown Unknown Unknown Sighting GPS Center Point too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small Center Tree Sclerocarya birrea 28 S 25°24.3325' E 29°20.9699' 90% Hits Misses DBH (cm) 39 Other Species Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea DBH (cm) 3.1 5.2 8 5 6.9 3.5 7.5 8.8 too small too small too small too small too small too small too small too small Percent Canopy Cover 18 2 63 Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small 64 Dichrostachys cinerea Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Elaeodendron transvaalensis Combretum molle Combretum molle Combretum molle Combretum molle Combretum molle Combretum molle Combretum molle Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Euclea crispa Ehretia rigida Ehretia rigida Grewia flavescens Grewia flavescens Grewia flavescens Grewia flavescens Dombeya rotundifolia Dombeya rotundifolia Acacia burkei Acacia burkei Strychnos madagascariensis Strychnos spinosa Sclerocarya birrea too small 4.4 too small too small too small too small too small too small too small too small too small too small too small 7.9 4.5 8.5 too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small too small 3.4 2.9 too small too small too small too small too small 65 Rhus pyroides Unknown Unknown Unknown Unknown Unknown Unknown Unknown Unknown Unknown Unknown Sighting GPS Center Point too small 3 9.1 18.7 5.2 too small too small too small too small too small too small Center Tree Acacia nigrescens 29 S 25°23.9305' E 29°20.5247' 30% Hits Misses DBH (cm) 24.7 Other Species Acacia nigrescens Acacia nigrescens Acacia nigrescens Acacia nigrescens Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Combretum apiculatum Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Lycium cinereum DBH (cm) 8.3 8.4 25.8 too small 6.2 4.5 5.6 too small too small too small 2.7 5 too small too small Percent Canopy Cover Sighting GPS Center Point Percent Canopy Cover 30 S 25°24.9987' E 29°18.0172' 40% Hits Misses 6 14 8 12 66 Center Tree Sclerocarya birrea DBH (cm) 29.7 Other Species Dombeya rotundifolia Dombeya rotundifolia Acacia burkei Protea caffra Protea caffra Protea caffra Protea caffra Protea caffra Protea caffra Protea caffra Protea caffra DBH (cm) 24.3 7.3 7.2 too small too small 20.9 4.3 6.5 5.7 6.1 14.4 Sighting GPS Center Point Center Tree Dicrhostachys cinerea 31 S 25°24.7356' E 29°20.5844' 70% Hits Misses DBH (cm) 15.3 Other Species Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Acacia nilotica Acacia tortillis Acacia tortillis Acacia tortillis Acacia tortillis Rhus pyroides DBH (cm) 6.2 3.6 4.9 36.6 2.7 too small too small too small too small too small too small 2.5 19.4 too small too small too small Percent Canopy Cover 14 6 67 Sighting GPS Center Point Center Tree Dichrostachys cinerea 32 S 25°24.4588' E 29°21.0310' 90% Hits Misses DBH (cm) 20.4 Other Species Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Dichrostachys cinerea Acacia burkei Grewia monticola Ozoroa paniculosa Combretum apiculatum Grewia flavescens Peltophorum africanum DBH (cm) 9.3 21.2 7.8 9.8 51.4 14.3 11.5 too small too small too small too small 4.4 46.1 too small 10.7 Percent Canopy Cover 18 2 68