A genomic and phylogenetic perspective on endosymbiosis and algal origin
advertisement

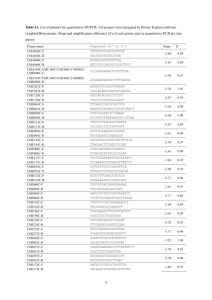
Journal of Applied Phycology (2006) 18: 475–481 DOI: 10.1007/s10811-006-9054-6 C Springer 2006 A genomic and phylogenetic perspective on endosymbiosis and algal origin Hwan Su Yoon, Jeremiah D. Hackett & Debashish Bhattacharya∗ Department of Biological Sciences and Roy J. Carver Center for Comparative Genomics, University of Iowa, 446 Biology Building, Iowa City, Iowa 52242-1324 ∗ Author for correspondence: e-mail: debashi-bhattacharya@uiowa.edu; fax: (319) 335-1069 Received 4 August 2004; accepted 17 November 2004 Key words: algal evolution, chromalveolates, endosymbiosis, gene transfer, plastid Abstract Accounting for the diversity of photosynthetic eukaryotes is an important challenge in microbial biology. It has now become clear that endosymbiosis explains the origin of the photosynthetic organelle (plastid) in different algal groups. The first plastid originated from a primary endosymbiosis, whereby a previously non-photosynthetic protist engulfed and enslaved a cyanobacterium. This alga then gave rise to the red, green, and glaucophyte lineages. Algae such as the chlorophyll c-containing chromists gained their plastid through secondary endosymbiosis, in which an existing eukaryotic alga (in this case, a rhodophyte) was engulfed. Another chlorophyll c-containing algal group, the dinoflagellates, is a member of the alveolates that is postulated to be sister to chromists. The plastid in these algae has followed a radically different path of evolution. The peridinin-containing dinoflagellates underwent an unprecedented level of plastid genome reduction with the ca. 16 remaining genes encoded on 1–3 gene minicircles. In this short review, we examine algal plastid diversity using phylogenetic and genomic methods and show endosymbiosis to be a major force in algal evolution. In particular, we focus on the evolution of targeting signals that facilitate the import of nuclear-encoded photosynthetic proteins into the plastid. Introduction The eukaryotic photosynthetic organelle (plastid) is critical to life on our planet because of its contribution to global primary production. Ten different types of plastids are known and are found in evolutionarily divergent eukaryotic clades (Baldaif et al., 2000; Bhattacharya et al., 2004). The endosymbiosis hypothesis was put forth to explain the origin of plastids and mitochondria (Margulis, 1970; Mereschkowsky, 1905) and has been extensively supported with modern molecular evolutionary analyses. In plastid primary endosymbiosis, a non-photosynthetic protist engulfed a cyanobacterium and converted it into a permanent photosynthetic organelle. This photosynthetic eukaryote gave rise to the red, green, and glaucophyte algae that have a plastid bound by two membranes. Thereafter, plastids were horizontally spread into the remaining photosynthetic protist groups through secondary endosymbiosis, in which non-photosynthetic cells engulfed an existing (red or green) alga. This proces resulted in the plastid of cryptophytes, haptophytes, stramenopiles, apicomplexans, dinoflagellates (red algal endosymbiont), euglenophytes, and chlorarachniophytes (green algal endosymbiont, Bhattacharya & Medlin, 1995; Cavalier-Smith, 1986; Douglas, 1998; Douglas et al., 1991; Gibbs, 1978; McFadden et al., 1994; Yoon et al., 2002b; Zhang et al., 1999). However, endosymbiosis did not stop there because in dinoflagellates the existing plastid of red algal origin was replaced on multiple independent occasions with this organelle from an alga containing a secondary plastid (a cryptophyte, haptophyte or stramenopile: tertiary endosymbiosis) or a primary plastid (a green alga) (Chesnick et al., 1997; Hackett et al., 2003; Ishida & Green, 2002; Tengs et al., 2000; Watanabe et al., 1990; Yoon et al., 2002a). The development of large scale sequencing and genomic approaches has greatly augmented our understanding of algal evolution. These methods have been applied to generate complete genome or expressed sequence tag (EST) databases of model algae or protists 476 as well as to generate broadly sampled multi-gene phylogenies. In this paper, we discuss algal diversity from the perspective of plastid endosymbiosis, and present a brief summary of recent findings from genomic and phylogenetic approaches. In addition, we examine the leader sequences of nuclear-encoded plastid genes that have resulted from intracellular gene transfer and that make possible plastid targeting. Endosymbiosis is an important driving force in algal evolution Primary endosymbiosis Rhodophyta, Viridiplantae (green algae and land plants), and Glauco(cysto)phyta contain plastids surrounded by a double membrane that very likely originated through a single primary endosymbiosis in the common ancestor of these taxa (Bhattacharya & Medlin, 1995; Delwiche et al., 1995; Gray, 1992; McFadden, 2001; Moreira et al., 2000; Matsuzaki et al., 2004; McFadden & van Dooren, 2004). Molecular clock analysis using a concatenated data set of six plastid genes and multi-fossil calibrations suggest that the primary endosymbiosis occurred around 1.6 billion years ago (Yoon et al., 2004). This estimate has been independently confirmed by multi-protein analyses of nuclear loci that suggest a date of 1.6–1.5 BY for primary plastid origin (Hedges et al., 2004; Hackett et al., 2006). Despite their ancient origin, the monophyly of Plantae is moderately supported by recent molecular phylogenetic studies using nuclear and mitochondrial genes (Baldauf et al., 2000; Moreira et al., 2000; Palmer, 2003; Rodriguez-Ezpeleta et al., 2005). A broadly sampled tree of microbial eukaryotes is urgently needed to test the monophyly of Plantae (and other groups – see below). Secondary endosymbiosis The putative lineage Chromista, which comprises the cryptophytes, haptophytes, and stramenopiles, contain chlorophyll c in their 4-membrane bound plastid (Cavalier-Smith, 1986). The chromist plastid is not located in the cytosol but rather within the rough endoplasmic reticulum (RER), which is connected to the outermost membrane of the plastid and is referred to as the chloroplast endoplasmic reticulum (CER). Secondary endosymbiosis, in which the nonphotosynthetic ancestor of chromists engulfed an existing red alga, explains plastid origin in this group (Bhattacharya & Medlin, 1995; Douglas et al., 1991; Fast et al., 2001; Gibbs, 1981; Harper & Keeling, 2003). Evidence for this secondary endosymbiosis comes from the cryptophytes that retain the remnant nucleus of the red algal endosymbiont, the nucleomorph, between the two inner and two outer plastid membranes. The haptophytes and stramenopiles have presumably lost the nucleomorph after their divergence from the cryptophytes. Our molecular clock analysis suggests a minimum age of 1.3 BY for this secondary endosymbiotic event and around 1.2 BY for the divergence of cryptophytes from the other chromists and 1 BA for the split of haptophytes and stramenopiles (Yoon et al., 2004). Alveolates, which comprise the dinoflagellates, apicomplexans, and ciliates, are postulated to be sister to the chromists (together, the chromalveolates; CavalierSmith, 1999 [see Fast et al., 2001; Harper & Keeling, 2003; Bhattacharya et al., 2004]). The chromalveolate common ancestor most likely contained a red algal secondary endosymbiont (Cavalier-Smith, 1999) that was apparently lost in the ciliates. In the apicomplexans, such as the well-known human parasite Plasmodium falciparum Welch, the remnant plastid (called the apicoplast) genome was reduced to a 35 Kb circle (Williamson et al., 1994). However, the phylogenetic history of apicoplasts remains unclear because of the high divergence of the encoded sequences that usually results in long branch artifacts in trees (Funes et al., 2002; Waller et al., 2003; Zhang et al., 1999, 2000). An important data set that supports chromalveolate monophyly is the presence of a unique glyceraldehyde3-phosphate dehydrogenase (GAPDH) gene replacement shared by these taxa (Fast et al., 2001; Harper & Keeling, 2003). In addition, because the plastid genes of dinoflagellates were re-organized into mini-circles and most of the plastid genes were transferred to the nucleus (followed by high sequence divergence rates in many of these coding regions), it is difficult to accurately infer the phylogeny of dinoflagellate plastids (Bachvaroff et al., 2004; Hackett et al., 2004; Zhang et al., 1999). To resolve this issue, we sequenced five minicircle-encoded plastid proteins from a handful of peridinin dinoflagellates and from fucoxanthincontaining taxa. These latter taxa presumably gained their plastid through a haptophyte tertiary endosymbisois (Ishida & Green, 2002; Tengs et al., 2000; Yoon et al., 2005). In the resulting trees, the evolutionary origin of the peridinin plastids remains unclear but this clade is clearly positioned as a monophyletic lineage within the red algae with a weak sister group 477 relationship to the stramenopiles. This result is consistent with the chromalveolate hypothesis and potentially explains the monophyletic origin of the plastid (and by extension, the host cells; Cavalier-Smith, 1999; Fast et al., 2001; Harper & Keeling, 2003). However, it is critical to verify this hypothesis with a broad taxon sampling and a multi-gene analysis of the host cells. Euglenophytes and chlorarachniophytes acquired their green algal plastid through independent secondary endosymbioses (Archibald & Keeling, 2002; Baldauf, 2003; Bhattacharya et al., 2004; Palmer, 2003). However, if secondary plastid loss has occurred more frequently then we postulate, genomic analysis may be the best, and perhaps only, approach to identify the number of plastid endosymbioses that have occurred during eukaryotic evolution. The finding of nuclearencoded genes of photosynthetic function in presently aplastidial cells will, for example, allow us to more accurately map the ancestral plastid distribution on the host tree. Moestrup which gained its plastid through a green algal secondary endosymbiosis (Archibald et al., 2003). EST analysis of this taxon showed numerous lateral transfers of genes from streptophyte, stramenopile, red algal, and bacterial sources. Furthermore, the whole genome sequence from the apicomplexan parasite P. falciparum reveals that 11% (581/5268 proteins) of genes of plastid function are still maintained in the nucleus (Gardner et al., 2002). It is interesting that these genes are absent from another apicomplexan, Cryptosporidium parvum Tyzzer, that apparently lacks an apicoplast (Abrahamsen et al., 2004). Taken together, genomic approaches are becoming ever more popular and result in an unprecedented quantity and quality of data that provide critical evolutionary information. However, it is important to keep in mind that single sequenced taxa (model or otherwise) do not adequately represent the evolutionary diversity of eukaryotes and taxonomically broadly sampled genomics projects hold the greatest promise for clarifying algal evolution. Genomic approaches for clarifying algal evolution Protein import system: Leader sequence Genomic methods such as whole genome random shotgun and EST approaches have recently been used to significantly improve our understanding of algal evolution. In one study Martin et al. (2002) found that ca. 18% of the nuclear genes in Arabidopsis thaliana (L.) Heynh. (of both photosynthetic and non-photosynthetic function) originated through intracellular gene transfer from the original cyanobacterial primary endosymbiont. This suggests that primary endosymbiosis resulted in massive lateral gene transfer from the endosymbiont to the nucleus and subsequently resulted in the enrichment and potential re-organization of the nuclear genome. Three EST studies have thus far been done with dinoflagellates (i.e., Alexandrium tamarense (Lebour.) E. Balech, Amphidinium carterae Hulburth, and Lingulodinium polyedrum (F. Stein) J. D. Dodge)) and many others are underway. These studies have identified a massive transfer of plastid genes to the nucleus (Bachvaroff et al., 2004; Hackett et al., 2004) and intriguingly some of these genes are of green algal origin. This suggests that that there has either been multiple lateral gene transfers from green algal sources or, less parsimoniously, an as yet unsubstantiated green algal endosymbiosis (Hackett et al., 2004). Other examples of gene transfer have been reported in the chlorarachniophyte alga, Bigelowiella natans Following transfer to the nucleus, the proper function of the proteins involved in plastid function relies on their successful import into this organelle. This process occurs because of the presence of a N-terminal extension on each protein that specifies organellar import (Martin & Herrmann, 1998). The nature of these “leader” sequences depends on the ultrastructure of the plastid, such as the number of bounding membranes, and the location of the organelle in the cytoplasm (for detailed review, see Kilian & Kroth, 2003). Two-membrane plastids located in the cytosol The cyanobacterial origin of plastids bound by two membranes is strongly supported by the finding of homology of the protein import channel of the translocon of the inner/outer envelope of the plastid (i.e., Tic20, Tic55, and Toc75) among plants and cyanobacteria (Eckart et al., 2002; Heins et al., 1998; Kilian & Kroth, 2003). The green, red, and glaucophyte algae contain leader sequences (Figure 1A) to target proteins to the plastidial Tic/Toc system (see Matsuzaki et al., 2004; McFadden & van Dooren, 2004). These residues are encoded on the 5 -terminus of the open reading frame and are of length 25–125 amino acids (Cavalier-Smith, 2000; McFadden, 1999; Nassoury et al., 2003; Waller et al., 1998). Within the plastid, an endopeptidase 478 Figure 1. Plastid origin and the protein import system in photosynthetic eukaryotes. The protein import system reflects the plastid ultrastructure rather than the source of the organelle. (A) After primary endosymbiosis, massive plastid gene transfer (GT) occurred to the nucleus (Nu). A transit peptide (TP) that modified the N-terminus targets the functional proteins to the 2-membrane bound plastid (P) through the translocon of the inner/outer plastid envelope (TIC/TOC). This import system is found in algae containing primary endosymbionts, however, it may also be present in the 2-membrane bound plastid of the dinoflagellates Dinophysis and Lepidodinium. (B) Chromista that contain 4-membrane plastids with a chloroplast endoplasmic reticulum (CER) have a modified bipartite leader sequence, which targets the proteins to the CER with the signal peptide (SP) in addition to typical transit peptide. (C) The secretory pathway (ER and Golgi apparatus) is involved in the protein import system in the apicomplexa and chlorarachniophytes. The small circle represents microsomes (m) that contain the transit peptide and the functional protein via the secretory pathway. (D) The 3-membrane bound plastid in the dinoflagellates and euglenophytes contain a tripartite leader sequence. The second hydrophobic region (ST) acts as a stop transfer signal that generates a functional protein in the cytoplasm (m). CB, cyanobacterium; CR, cryptophyte; HA, haptophyte; RH, rhodophyte; VI, Viridiplantae. cleaves the leader sequence (transit peptide). The twomembrane bound plastid in the dinoflagellates, Dinophysis spp. and Lepidodinium viride M. Watanabe, S. Suda, I. Inoye, T. Sawaguchi and M. Chihara which did not originate through primary endosymbiosis (rather via cryptophyte and green algal plastid replacements, respectively), most likely use the Tic import pathway of the inner plastid membrane that has been found in all algae and plants (McFadden & van Dooren, 2004). Four-membrane bound plastids located in the CER of the lumen In addition to the two inner membranes, chromists contain an additional two membranes that necessitate a more complex targeting signal (Figure 1B). Because the plastid of chromists is located in the CER, targeting into this membrane requires a classic signal peptide that has a hydrophobic region (Apt et al., 2002). This bipartite leader sequence subsequently targets the protein across the inner two membranes with a downstream transit peptide. Four-membrane plastids located in the cytosol Although the plastids of apicomplexans and chlorarachniophytes are of independent origins (from a red and a green alga, respectively), they both contain a four-membrane bound plastid located in the cytosol (unlike the chromists, Figure 1C). The bi-partite leader sequence, that specifies a signal peptide and a transit peptide, target the proteins into the cytosolic RER where the signal peptide is cleaved (McFadden, 1999; 479 Waller et al., 1998). During passage through the Golgi system, the secretory vesicles fuse with the outermost membrane of the plastid and thereafter, the transit peptide directs the protein through the Tic/Toc system. A secretory system-dependent transport system has been found in the four-membrane bound apicoplast in P. falciparum and surprisingly, as well as in the threemembrane bound plastid in Euglena gracilis G. A. Klebs and Gonyaulax polyedra F. Stein that do not contain a CER (Nassoury et al., 2003; Sulli et al., 1999; Waller et al., 2000). Three membrane-bound plastids located in the cytosol Dinoflagellates and euglenophytes contain plastids bound by three membranes that do not have a connection between the outer plastid membrane and the endomembranes (Figure 1D). Tripartite leader sequences, which consist of a hydrophobic signal peptide, a transit peptide, and a second hydrophobic region, were found in these taxa (Hackett et al., 2004; Nassoury et al., 2003; Sulli et al., 1999). The second hydrophobic region acts as a “stop transfer signal” and the functional protein is located in the cytoplasmic side of the ER after cleavage of the signal peptide (Nassoury et al., 2003). Microsomes that contain the functional transit peptide pass through the membranes via a secretory pathway (Golgi apparatus) followed by subsequent vesicular transport across the cytoplasm before entering the plastids (Nassoury et al., 2003; Sulli et al., 1999). A tripartite leader sequence in the psbO gene has been identified from the fucoxanthin-containing dinoflagellate, Karenia brevis (C. C. Davis) G. Hansen & Ø. Moestrup (Ishida & Green, 2002). springs (Lopez-Garcia et al., 2001; Moon-van der Staay et al., 2001; Ciniglia et al., 2004). Furthermore, the picoplankton in both coastal and open ocean environments promises to be a potentially endless source of novel taxa. Because many of these lineages are positioned basal in trees, the environmental PCR method provides a powerful tool for understanding early algal evolution. In this regard, the finding that many basal lineages such as the marine stramenopiles and alveolates are heterotrophic, forces us to postulate multiple secondary plastid losses in chromalveolates to be consistent with the ideas presented here. Recently, an environmental meta-genomic approach using shotgun whole genome sequencing was used to study bacterial diversity in the oligotrophic Sargasso Sea (Venter et al., 2004). This breakthrough work identified at least 1800 genomic species including 148 previously unknown bacterial phylotypes and over 1.2 million previously unknown genes. Although an imprecise and incomplete approach to generating complete genome sequences, this remarkable body of data challenges all biologists to account more rigorously for microbial diversity when studying eukaryotic and in particular, algal evolution. Clearly, we are just beginning to understand the complex history of microbial eukaryotes and the future holds great promise in clarifying the phylogeny of algae and their place in the tree of life. Acknowledgements This work was primarily supported by grants from the United States National Science Foundation to D.B. (grants DEB 01-07754, MCB 02-36631). Conclusions Great progress has recently been made in generating the outline of the eukaryotic tree of life using molecular phylogenies (e.g., Baldauf, 2003). The deep branches of the tree remain however unsubstantiated and await a rigorous multi-gene approach with a broad taxonomic sampling. This type of analysis will ultimately resolve the main splits in the algal tree and establish the timing of algal origins. The methods of phylogenetics and genomics provide significant data but the challenge remains to extensively sample representatives of all the major algal groups and relevant non-algal groups. Our understanding of algal biodiversity has also been significantly changed by environmental PCR analyses of extreme environments such as the deep sea and hot References Abrahamsen MS, Templeton TJ, Enomoto S, Abrahante JE, Zhu G, Lancto CA, Deng M, Liu C, Widmer G, Tzipori S, Buck GA, Xu P, Bankier AT, Dear PH, Konfortov BA, Spriggs HF, Iyer L, Anantharaman V, Aravind L, Kapur V (2004) Complete genome sequence of the apicomplexan, Cryptosporidium parvum. Science 304: 441–445. Apt KE, Zaslavkaia L, Lippmeier JC, Lang M, Kilian O, Wetherbee R, Grossman AR, Kroth PG (2002) In vivo characterization of diatom multipartite plastid targeting signals. J. Cell Sci. 115: 4061–4069. Archibald JM, Keeling PJ (2002) Recycled plastids: A ‘green movement’ in eukaryotic evolution. Trends in Genet. 18: 577–584. Archibald JM, Rogers MB, Toop M, Ishida K, Keeling PJ (2003) Lateral gene transfer and the evolution of plastid-targeted proteins in 480 the secondary plastid-containing alga Bigelowiella natans. PNAS 100: 7678–7683. Bachvaroff TR, Concepcion GT, Rogers CR, Herman EM, Delwiche CF (2004) Dinoflagellate expressed sequence tag data indicate massive transfer of chloroplast genes to the nuclear genome. Protist 155: 65–78. Baldauf SL (2003) The deep roots of eukaryotes. Science 300: 1703– 1706. Baldauf SL, Roger AJ, Wenk-Siefert I, Doolittle WF (2000) A kingdom-level phylogeny of eukaryotes based on combined protein data. Science 290: 972–977. Bhattacharya D, Medlin L (1995) The phylogeny of plastids: A review based on comparisons of small-subunit ribosomal RNA coding regions. J. Phycol. 31: 489–498. Bhattacharya D, Yoon HS, Hackett JD (2004) Photosynthetic eukaryotes unite: Endosymbiosis connects the dots. BioEssays 26: 50–60. Cavalier-Smith T (1986) The kingdon chromista: Origin and systematics. In: Round FE, Chapman DJ (eds.) Progress in Phycological Research No. 4, Biopress, Bristol, pp 309–347. Cavalier-Smith T (1998) A revised six-kingdom system of life. Biol. Rev. 73: 203–266. Cavalier-Smith T (1999) Principles of protein and lipid targeting in secondary symbiogenesis: Euglenoid, dinoflagellate, and sporozoan plastid origins and the eukaryote family tree. J. Eukaryot. Microbiol. 46: 347–366. Cavalier-Smith T (2000) Membrane heredity and early chloroplast evolution. Trends in Plant Sci. 5: 174–182. Chesnick JM, Kooistra WH, Wellbrock U, Medlin LK (1997) Ribosomal RNA analysis indicates a benthic pennate diatom ancestry for the endosymbionts of the dinoflagellates Peridinium foliaceum and Peridinium balticum (Pyrrhophyta). J. Eukaryot. Microbiol. 44: 314–320. Ciniglia C, Yoon HS, Pollio A, Pinto G, Bhattacharya D (2004) Hidden biodiodiversity of the extremophilic Cyanidiales red algae. Mol. Ecol. 13: 1827–1838. Delwiche CF, Kuhsel M, Palmer J. D (1995) Phylogenetic analysis of tufA sequences indicates a cyanobacterial origin of all plastids. Mol. Phylogenet. Evol. 4: 110–128. Douglas SE (1998) Plastid evolution: Origins, diversity, trends. Curr. Opin. Genet. Dev. 8: 655–661. Douglas SE, Murphy CA, Spencer DF, Gray MW (1991) Cryptomonad algae are evolutionary chimaeras of two phylogenetically distinct unicellular eukaryotes. Nature 350: 148– 151. Eckart K, Eichacker L, Sohrt K, Schleiff E, Heins L, Soll J (2002) A toc75-like protein import channel is abundant in chloroplasts. EMBO Reports 3: 557–562. Fast NM, Kissinger JC, Roos DS, Keeling PJ (2001) Nuclearencoded, plastid-targeted genes suggest a single common origin for apicomplexan and dinoflagellate plastids. Mol. Biol. Evol. 18: 418–426. Funes S, Davidson E, Reyes-Prieto A, Magallon S, Herion P, King MP, Gonzalez-Halphen D (2002) A green algal apicoplast ancestor. Science 298: 2155. Gardner MJ, Hall N, Fung E, White O, Berriman M, Hyman RW, Carlton JM, Pain A, Nelson KE, Bowman S, Paulsen IT, James K, Eisen JA, Rutherford K, Salzberg SL, Craig A, Kyes S, Chan MS, Nene V, Shallom SJ, Suh B, Peterson J, Angiuoli S, Pertea M, Allen J, Selengut J, Haft D, Mather MW, Vaidya AB, Martin DM, Fairlamb AH, Fraunholz MJ, Roos DS, Ralph SA, McFadden GI, Cummings LM, Subramanian GM, Mungall C, Venter JC, Carucci DJ, Hoffman SL, Newbold C, Davis RW, Fraser CM, Barrell B (2002) Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 419: 498–511. Gibbs SP (1978) The chloroplasts of Euglena may have evolved from symbiotic green algae. Can. J. Bot. 56: 2883–2889. Gibbs SP (1981) The chloroplasts of some algal groups may have evolved from endosymbiotic eukaryotic algae. Annal. NY Acad. Sci. 361: 193–208. Gray MW (1992) The endosymbiont hypothesis revisited. Int. Rev. Cytol. 141: 233–357. Hackett JD, Maranda L, Yoon HS, Bhattacharya D (2003) Phylogenetic evidence for the cryptophyte origin of the plastid of Dinophysis (dinophysiales, dinophyceae). J. Phycol. 39: 440– 448. Hackett JD, Yoon HS, Butterfield NJ, Sanderson MJ, Bhattacharya D (2006) Plastid endosymbiosis: Origins and timing of events. In: Falkowski P, Knoll A (eds.) Evolution of Aquatic Photoautotrophs, Academic Press, in press. Hackett JD, Yoon HS, Soares MB, Bonaldo MF, Casavant TL, Scheetz TE, Nosenko T, Bhattacharya D (2004) Migration of the plastid genome to the nucleus in a peridinin dinoflagellate. Curr. Biol. 14: 213–218. Harper JT, Keeling PJ (2003) Nucleus-encoded, plastid-targeted glyceraldehyde-3-phosphate dehydrogenase (GAPDH) indicates a single origin for chromalveolate plastids. Mol. Biol. Evol. 20: 1730–1735. Hedges SB, Blair JE, Venturi ML, Shoe JL (2004) A molecular timescale of eukaryote evolution and the rise of complex multicellular life. BMC Evol. Biol. 4: 2. Heins L, Soll J, Collinson I (1998) The protein translocation apparatus of chloroplast envelopes. Trends in Plant Sci. 3: 56–61. Ishida K, Green BR (2002) Second- and third-hand chloroplasts in dinoflagellates: Phylogeny of oxygen-evolving enhancer 1 (psbO) protein reveals replacement of a nuclear-encoded plastid gene by that of a haptophyte tertiary endosymbiont. PNAS 99: 9294– 9299. Kilian O, Kroth PG (2003) Evolution of protein targeting into “complex” plastids: The “secretory transport hypothesis”. Plant Biol. 5: 350–358. Lopez-Garcia P, Rodriguez-Valera F, Pedros-Alio C, Moreira D (2001) Unexpected diversity of small eukaryotes in deep-sea Antarctic plankton. Nature 409: 603–607. Margulis L (1970) Origin of eukaryotic cells. Yale University Press, New Haven, CT. Martin W, Herrmann RG (1998) Gene transfer from organelles to the nucleus: How much, what happens, and why? Plant Physiol. 118: 9–17. Martin W, Rujan T, Richly E, Hansen A, Cornelsen S, Lins T, Leister D, Stoebe B, Hasegawa M, Penny D (2002) Evolutionary analysis of Aarabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. PNAS 99: 12246–12251. Matsuzaki M, Misumi O, Shin-i T, Maruyama S, Takahara M, Miyagishima S, Mori T, Nishida K, Yagisawa F, Nishida K, Yoshida Y, Nishimura Y, Nakao S, Kobayashi T, Momoyama Y, Higashiyama T, Minoda A, Sano M, Nomoto H, Oishi K, Hayashi H, Ohta F, Nishizaka S, Haga S, Miura S, Morishita T, Kabeya Y, Terasawa K, Suzuki Y, Ishii Y, Asakawa S, Takano H, Ohta N, Kuroiwa H, Tanaka K, Shimizu N, Sugano S, Sato N, Nozaki H, Ogasawara N, Kohara Y, Kuroiwa T (2004) Genome 481 sequence of the ultrasmall unicellular red alga Cyanidioschyzon merolae 10D. Nature 428: 653–657. McFadden GI (1999). Plastids and protein targeting. J. Eukaryot. Microbiol. 46: 339–346. McFadden GI (2001) Primary and secondary endosymbiosis and the origin of plastids. J. Phycol. 37: 951–959. McFadden GI, van Dooren GG (2004) Evolution: Red algal genome affirms a common origin of all plastids. Curr. Biol. 14: R514– R516. McFadden GI, Gilson PR, Hofmann CJ, Adcock GJ, Maier UG (1994) Evidence that an amoeba acquired a chloroplast by retaining part of an engulfed eukaryotic alga. PNAS 91: 3690–3694. Mereschkowsky C (1905) Über Natur und Ursprung der Chromatophoren im Pflanzenreiche. Biol. Centralbl. 25: 593–604. Moon-van der Staay SY, Wachter RDe, Vaulot D (2001) Oceanic 18s rDNA sequences from picoplankton reveal unsuspected eukaryotic diversity. Nature 409: 607–610. Moreira D, Le Guyader H, Phillippe H (2000) The origin of red algae and the evolution of chloroplasts. Nature 405: 69–72. Nassoury N, Cappadocia M, Morse D (2003) Plastid ultrastructure defines the protein import pathway in dinoflagellates. J. Cell Sci. 116: 2867–2874. Palmer JD (2003) The symbiotic birth and spread of plastids: How many times and whodunit. J. Phycol. 39: 4–12. Rodriguez-Ezpeleta N, Brinkmann H, Burey SC, Roure B, Burger G, Loffelhardt W, Bohert HJ, Philippe H, Lang BF (2005) Monophyly of primary photosynthetic eukaryotes: green plants, red algae, and glaucophytes. Curr. Biol. 15: 1325–1330. Sulli C, Fang Z, Muchhal U, Schwartzbach SD (1999) Topology of Euglena chloroplast protein precursors within endoplasmic reticulum to Golgi to chloroplast transport vesicles. J. Biol. Chem. 274: 457–463. Tengs T, Dahlberg OJ, Shalchian-Tabrizi K, Klaveness D, Rudi K, Delwiche CF, Jakobsen KS (2000) Phylogenetic analyses indicate that the 19 hexanoyloxy-fucoxanthin- containing dinoflagellates have tertiary plastids of haptophyte origin. Mol. Biol. Evol. 17: 718–729. Venter JC, Remington K, Heidelberg JF, Halpern AL, Rusch D, Eisen JA, Wu D, Paulsen I, Nelson KE, Nelson W, Fouts DE, Levy S, Knap AH, Lomas MW, Nealson K, White O, Peterson J, Hoffman J, Parsons R, Baden-Tillson H, Pfannkoch C, Rogers Y-H, Smith HO (2004) Environmental genome shotgun sequencing of the Sargasso Sea. Science 304: 66– 74. Waller RF, Keeling PJ, Donald RG, Striepen B, Handman E, Lang-Unnasch N, Cowman AF, Besra GS, Roos DS, McFadden GI (1998) Nuclear-encoded proteins target to the plastid in Toxoplasma gondii and Plasmodium falciparum. PNAS 95: 12352–12357. Waller RF, Keeling PJ, van Dooren GG, McFadden GI (2003) Comment on “a green algal apicoplast ancestor”. Science 301: 49. Waller RF, Reed MB, Cowman AF, McFadden GI (2000) Protein trafficking to the plastid of Plasmodium falciparum is via the secretory pathway. EMBO J. 19: 1794–1802. Watanabe MM, Suda S, Inouye I, Sawaguchi T, Chihara M (1990) Lepidodinium viride gen. et sp. nov. (Gymnodiniales, Dinophyta), a green dinoflagellate with chlorophyll a- and bcontaining endosymbiont. J. Phycol. 26: 741–751. Williamson DH, Gardner MJ, Preiser P, Moore DJ, Rangachari K, Wilson RJ (1994) The evolutionary origin of the 35 kb circular DNA of Plasmodium falciparum: New evidence supports a possible rhodophyte ancestry. Mol. Gen. Genet. 243: 249– 252. Yoon HS, Hackett JD, Bhattacharya D (2002a) A single origin of the peridinin- and fucoxanthin-containing plastids in dinoflagellates through tertiary endosymbiosis. PNAS 99: 11724–11729. Yoon HS, Hackett JD, Ciniglia C, Pinto G, Bhattacharya D (2004) A molecular timeline for the origin of photosynthetic eukaryotes. Mol. Biol. Evol. 21: 809–818. Yoon HS, Hackett JD, Pinto G, Bhattacharya D (2002b) The single, ancient origin of chromist plastids. PNAS 99: 15507–15512. Yoon HS, Hackett JD, Van Dolah FM, Nosenko T, Lidie KL, Bhattacharya D (2005) Tertiary endosymbiosis driven genome evolution in dinoflagellate algae. Mol. Biol. Evol. 22: 1299–1308. Zhang Z, Green BR, Cavalier-Smith T (1999) Single gene circles in dinoflagellate chloroplast genomes. Nature 400: 155–159. Zhang Z, Green BR, Cavalier-Smith T (2000) Phylogeny of ultrarapidly evolving dinoflagellate chloroplast genes: A possible common origin for sporozoan and dinoflagellate plastids. J. Mol. Evol. 51: 26–40.