Early Hominins Hominins ultimately distinguished by brain size, bipedal
advertisement

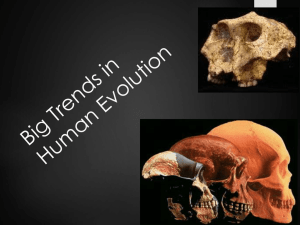
Early Hominins Hominins ultimately distinguished by brain size, bipedal locomotion and toolmaking behavior But these did not develop simultaneously: mosaic evolution The only reliable indicator of earliest hominin status: bipedal locomotion Miocene forms (earliest Australopiths) • Sahelanthropus tchadensis (7-6 mya) • Small brain, huge supraorbital torus • Face not prognathic and canines not large • Foramen magnum more anteriorly positioned than in apes • Orrorin tugenensis (6 mya) • Postcranial evidence suggests bipedal, but questions remain • Ardipithecus kadabba (5.8-5.2) • Mix of apelike and hominin dental traits • Postcranial remains imply bipedal, similar to later form A. afarensis All these forms quite primitive, as expected. But appear to have been bipedal. Key: changes in environment. Cooler, drier. Forests less continuous. Hypothesis: some late Miocene hominoids: exploited areas that included some grasslands. These become hominins. Other hominoids: exploited wetter, more densely forested areas: ancestors of African apes. Why did hominins become bipedal? Ecological theories: patchier forests But, ground-dwelling doesn’t require bipedalism! Also, hominins did not suddenly become completely terrestrial. A. kadabba associated with a wet, wooded area 2 with some wooded grassland. S. tchadensis associated with lakeside gallery forest and some grasslands. Various • • • • • • • scenarios: Carrying (objects, infants) Scavenging Seed and nut gathering Feeding from bushes Surveillance Long-distance walking Male provisioning Darwininian model has been used: natural selection for increasingly better adaptation to a specific habitat: stresses consistency of selection over time Alternative View of Adaptive Change: Variability Selection What if a population faces many remodelings of its habitat over geologic time? May evolve in response to wide range of adaptive conditions – selection may be inconsistent over long time spans Environmental records: worldwide fluctuations increasingly extreme from Miocene to Recent: glacial-interglacial, aridmoist cycles over longer and longer intervals, up to 100,000+ years during last half of Pleistocene. Hypothesis: significant changes during hominin evolution may not have resulted from adaptation to any one habitat type or general environmental trend: First: environmental variability Æ strong disparities in adaptive settings over time Second: resulting inconsistencies in natural selection had powerful influence on evolutionary change Consider: generally two ways of responding to habitat change: 1. Mobility or dispersal: track habitat or key resource 2. Broaden range of conditions in which can live How can adaptive flexibility be achieved? 3 Several ways: genetic polymorphism, phenotypic plasticity and… Third way: complex structures or behaviors designed to respond to novel and unpredictable adaptive settings: Variability Selection V.S. refers to evolution of such adaptive mechanisms. V.S. emphasizes effect of inconsistent selection over long spans to time. Examples: • Locomotor system designed to allow wide repertoire of movement • Particular dental structure or foraging strategy that enhances shift to newly available food types • Large brain or specific neurological structure – effective data processing and complex cognitive responses • Species-specific social behaviors Existential poker, once again: Remember key: maximizing homeostatic ability In case of V.S. alleles favored by V.S. must avoid losing the game (i.e., extinction) in each specific environmental setting they encounter. BUT—these alleles may not be associated with highest reproductive success in any given setting! What really matters: alleles permit survivorship in a given setting AND episodic change in surroundings ultimately lower fitness of habitat-specific alleles that once had greater advantage. V.S. a process by which adaptive mechanisms emerge that buffer episodic change in an environmental setting Effects on Hominin Evolution Early australopith body plan: terrestrial bipedalism and tree climbing: a mosaic of human-like and ape-like characters: Pelvic and knee modifications for two-legged striding Powerful forelimbs and joint maneuverability indicating arboreal activity 4 Fossil evidence also suggests: foot more flexible than ours with respect to climbing This theme endures for 2+ million years Gives locomotor versatility Early australopith environments: variable: forest, savanna, woodland Testing hypothesis of V.S. for locomotor system: what were the habitats like where australopithecine locomotor system first evolved? The Bipedal Adaptation Striding gait: weight placed alternately on single, fully extended hind limb. (Both feet on ground only 25% of time, decreases with increasing speed) Drastic structural/anatomical alterations in basic primate quadrupedal pattern: • • • • • • • Hominin pelvis comparatively much shorter and broader and extends to side: to stabilize line of weight transmission from lower back to hip joint Broadening two sides an extend around to produce basinshaped structure to support abdominal organs • These also reposition attachments of key muscles that act on hip and leg: especially gluteus maximus Gluteus maximus in quadruped: pulls thigh to side; in biped it acts, along with hamstring, to extend thigh, pulling to rear when walking and running Foramen magnum repositioned farther under skull. Skull balanced on spine Spine has two distinct curves – thoracic and lumbar. Keep trunk (and weight) centered above pelvis Lower limbs elongated: in humans 20% of body height, only 11% in gorillas) Femur angles inward; modified knee permits full extension of this joint Big toe enlarged and brought in line with others (adducted); distinctive longitudinal arch: absorbs shock and adds propulsive spring Hominin bipedalism becomes habitual and obligate. 5 All major structural changes required for bipedalism seen in early hominins from East and South Africa. In particular, pelvis dramatically remodeled. Pliocene forms Set I: Early, primitive Australopiths • Ardipithecus ramidus • Dated to 4.4 mya • In partial cranium, foramen magnum positioned further forward than in quadrupeds • Humerus differs from quadrupeds: didn’t function to support weight in locomotion • Above evidence suggests obligate bipedalism • But, clearly primitive: flattening of cranial base and relatively thin enamel caps on molars; canines large relative to postcanines • Estimated weight: 93 lbs. • Australopithecus anamensis • Dated to 4.2 – 3.9 mya • Thick tooth enamel like all later Australopithecines • Canines slightly larger than in A. afarensis but with humanlike vertical roots • Postcanine tooth rows parallel • Very small acoustic meatus as in Ar. ramidus • Postcranial skeleton similar to A. afarensis • Probably had relatively long forearms compared to modern humans • Possibly descended from Ar. ramidus and ancestral to A. afarensis • Teeth and supporting bones suggest change in diet: from one like chimp in Ar. ramidus to harder foods. • Australopithecus afarensis • Dated 4.2 – 2.5 mya • Heavily enameled back teeth • Postcanine tooth rows range from nearly parallel to slightly divergent • Foramen magnum under skull • Footprints from Laetoli (3.7 mya) believed from this species – bipedal, perhaps “strolling” with short stride • Canines large, lower first premolar semisectorial 6 • • Cranium: hint of sagittal crest; 380-500 ml cranial capacity, with average 420 ml (chimp avg. 395, gorilla 506) • Kenyanthropus platyops • Dated 3.5 – 3.2 mya • Site environment comparable to Laetoli and Hadar (mixed grassland/wooded) • Skull: mosaic of features – thick enamel on molars (like A. afarensis) as well as derived characteristics such as flatter mid-facial region and tall, vertically oriented cheek region • Australopithecus bahrelghazalia • Dated 3.5 – 3.0 mya • Found in Chad, in north-central Africa • Extends known range of hominins of period 1500 miles west Set II: Later, more derived Australopiths • Australopithecus africanus • Dated 3.5 – 1.7 mya • Presumably descended from A. afarensis • Retains long arms • Large, thickly enameled back teeth • Retains flexibility of foot • Australopithecus garhi • Dated 2.5 mya • Femur longer than other australopiths – step towards long legs of modern humans • Possible use of tools to process animals for food • Paranthropus • Dated 2.4 – 1.3 mya • P. aethiopicus ancestor to later P. robustus and P. boisei • Exceptionally large back teeth with thick enamel • Specialized diet of hard foods: seeds, nuts but also meat • Extremely powerful chewing apparatus: massive jaws, broad faces, sagittal crest • Use of bone digging tools, possible stone tools 7 • Set III: Early Homo (first hominans) • Homo habilis • Dated 2.3 – 1.6 mya • • Homo rudolfensis Dated 2.4 – 1.6 mya • • • • • These forms the first to show encephalization Altered cranial shape – higher vault, more rounded Smaller teeth (especially molars); narrower premolars Diet more omnivorous Toolmaker: Oldowan industry