Chapter 7: Nerve Cells and Electrical Signaling
advertisement

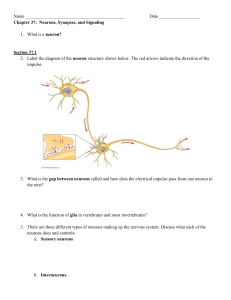
Kristen Taylor: 2420 Lecture Notes Chapter 7 Chapter 7: Nerve Cells and Electrical Signaling Before you tackle this stuff, make sure you really understand electrochemical gradients, ion channels and types of transporters (especially the Na+/K+ pump!). In chapter 6 we began a discussion about our organism-wide slow/sustained signaling system: the endocrine system. Now, in chapters 7-11, we are going to begin our body’s fast communication system: the nervous system. There are going to be a ton of details that you need to know about this system but do NOT lose sight of the big picture: rapid physiological responses are mediated by this one! There are really only 4 major concepts for the nervous system: 1) Graded Potentials (GPs) – Chapter 7 2) Action Potentials (APs) – Chapter 7 3) Synapses (how neurons communicate with other cells) – Chapter 8 4) Neurotransmitters (Remember the signaling molecule classes from chapter 5?) – Chapter 8 These 4 processes are how neurons work! Let’s start with the anatomy (which makes sense because of the structure function correlation!) Nervous System Overview: Figure 7.1 page 167 1) Central Nervous System (CNS): consists of the brain and spinal cord. The function of the CNS is to receive and process information – this is your integrating center (chapter 1). The CNS receives information from the external environment (sensory information through the afferent pathway) as well as the internal environment (visceral information through the efferent pathway). The really ambiguous part of this system is how it integrates the incoming information and sends out appropriate signals to effectors in order to maintain homeostasis (alluded to in chapters 9 and 11). 2) Peripheral Nervous System (PNS): consists of the nervous system everywhere else but the CNS. Subdivided into two subdivisions: A) Afferent (from outside to the CNS): this subdivision innervates organs, which transmit sensory and visceral information. This sensory information includes innervation from the somatic senses (from skin, muscles, and joints etc), the special senses (hearing, sight etc) and internal visceral receptors (pH, blood pressure, stomach fullness etc). [hint: remember that A comes before E so Afferent information comes first then the CNS “thinks” about it then sends its response via the Efferent pathways] More to come on this in chapter 10 1 B) Efferent (from CNS to outside): this subdivision transmits info from the CNS to the organs (effector organs – such as muscles and glands). This subdivision can be further divided into two branches: 1) Somatic Nervous System (SNS): this one consists of motor neurons innervating skeletal muscle (most of which are under voluntary control) 2) Autonomic Nervous System (ANS): this subdivision regulates the working of internal organs and other involuntary structures [are they really totally involuntary or can we have a conscious effect on them?] such as sweat glands, blood vessel, and smooth muscle. The ANS can be further subdivided into 2 other divisions (more to come in chapter 11): a) Parasympathetic: Rest and Digest responses b) Sympathetic: Fight or Flight responses C) Enteric Nervous System: this division can be thought of as its own system. However, I’ve grouped it with the PNS so our heads would not explode with yet another subdivision. This system innervates our digestive tract and is REALLY poorly understood! (we’ll talk a little about this towards the end of the semester when we discussion chapter 20) Cells of the Nervous System (NS): Pages 169-171 There are 2 main classes of cells: 1) Neurons: these are the fundamental functional unit of the NS. Neurons are excitable cells, which means that they are capable of producing action potentials (APs) which are, in essence, large, rapidly changing, electrical signals (if you want a preview see page 183). Neurons are a highly derived cell type (meaning very specialized) so we need to discuss some cellular structures: A neuronal cell can be divided into three distinctly functional parts: A) Cell Body (soma): this is the part of the cell that “looks” like a normal cell – it contains the nucleus and organelles so is able to carry out normal metabolic functions (see page 171). This part of the cell can receive inputs of information in the form of chemical, electrical, or nonchemical stimuli (light, sound etc). Interesting Note: when most CNS neurons become fully mature (G0 stage) they lose the ability to divide because they get rid of their centrioles (see the cool reading in the Discovery panel on Page 170)! B) Dendrites: these are small branches (foldings of the plasma membrane) that usually (depending on neuron type) branch off of the soma. These structures are specialized to receive input in the form of neurotransmitters (so contain a lot of ligand-gated receptors!). C) Axon (nerve fiber): these long strands of mostly plasma membrane are specialized to transmit information. Most neurons have one main axon but these axons can branch (called collaterals) and transmit information to more than one site. The portion of the axon that comes 2 off of the cell body is called the axon hillock and is specialized in the initiation of action potentials (APs). The end of the axon is aptly called the axon terminal and is specialized in releasing neurotransmitters in response to action potentials traveling down the axon. Channels in Neurons: (hopefully this will be a review) Leak Channels (non-gated channels): are found on all parts of a neuronal cell and are always open (allowing the maintenance of the resting potential). Ligand-gated channels: these guys open/close in response to the binding of a specific ligand, which in neurons, is most often a neurotransmitter. This type of channel is very dense on the cell body and dendrites since these channels act to receive the information. Voltage-gated channels: either open or close in response to changes in the membrane potential. Voltage-gated Na+ and K+ channels are found all over the cell but most are tightly packed on the axon especially within the axon hillock area. Voltage-gated Ca2+ channels are also present especially on the axon terminal and open in response to an action potential. Triggering this type of Ca2+ channel triggers the release of neurotransmitter. Structural Classes of Neurons: Page 172 Bipolar: have two projections dendrites and an axon. This type is most often associated with sensory structures such as olfaction and vision. It has one subclass: Pseudounipolar which is a type where the dendrites and axon appear as if they are one continual structure, which, in fact, they are! The dendrites are called a peripheral axon and the true axon is called the axon process (which always ends in the CNS). Almost ALL sensory neurons are pseudounipolar (so the Afferent nervous system uses these)! Multipolar: these are the ones that you see most often in drawings and have multiple dendrites projecting from the cell body and one axon coming off the cell body. Second Type of Cell Typically found in the NS: Page 173 2) Glial Cells: these cells make up 90% of the nervous system and are critical for maintenance of normal neuronal function – look at their name: glia means “glue”. There are 5 types of glial cells: astrocytes, ependymal cells, microglia, oligodendrocytes and Schwann cells. We’ll only cover the last two in this chapter: a) Oligodendrocytes: these are found only in the CNS and wrap their plasma membrane around neurons in order to form an insulating wrap called myelin. This cell type can reach out many plasma membrane projections in order to myelinate multiple neurons. b) Schwann cells: these are found only in the PNS and work just like oligodendrocytes. However, one Schwann cell can only form one myelin sheath for one neuron. 3 Why form these Myelin sheaths? Easy: the plasma membrane (pm) is hydrophobic so ions cannot move through it so these wrappings of plasma membrane act to insulate the neuron and allow much faster conduction of APs (like you would with an insulated wire). Please note: action potentials can only propagate between myelin sheaths at locations called the Nodes of Ranvier, which are the sites between myelin wrappings that contain the voltage-gated Na+ and K+ channels. For a discussion on the functions of the other types of glial cells please see pages 216-223 in chapter 9) Resting Membrane Potential (a little review of chapter 4) pgs 174-179 We talked about this in chapter 4 but now we are really going to get into the ins and outs! Reading through the toolbox on page 176 it reviews Ohm’s Law! Some terms: Current (I): the movement of electrical charges (in our case, ions) Resistance (R): How hard it is to move the charges (dependent on the properties of the pm, how many channels for each ion and how fast ions can move through them Conductance (g): the inverse of resistance or how easy it is to move charges Potential Difference (E in our book but V anywhere else): Also known as voltage (everywhere but in obscure physiology textbooks ) Ohm’s Law states the relationship between these: V=IR Rearrange it a bit and you get: I=V/R or I=Vg Remember that resting membrane potential is negative and is expressed ALWAYS as the potential of the inside of the cell versus the outside of the cell! The resting Vm of neurons is -70 mV and is dependent upon the relative [ions] (especially Na+ and K+) and the number of ion channels in the plasma membrane. Please remember the following: there is a lot more Na+ outside the cell and it is balanced by the amount of Cl- outside; there is a lot more K+ inside the cell and the charged is balanced by organic anions within the cell (such as phosphate groups)! Determining Equilibrium Potential for Na+ and K+ If you remember the following two rules then this section should be fairly understandable: The resting membrane potential of any cell is dependent upon two factors: 1) The concentration gradient between ions (in this case Na+ and K+) across the plasma membrane 2) The presence, and number, of ion channels in the plasma membrane In the End: “the net electrochemical force on an ion tends to move that ion across the membrane in the direction that will move the membrane potential toward the ion’s equilibrium potential – that is to bring the ion closer to equilibrium.” Reading through and understanding pages 173-178 (Figures 7.6, 7.7 and 7.8) are critical for a good understanding of how the resting membrane potential in neurons work! Overall point: Because neurons are 25X more permeable to K+ (meaning they have more K+ leak channels) than to Na+ the resting membrane potential will be 4 much closer to the K+ equilibrium potential (-94 mV) than to the Na+ equilibrium potential (+60 mV). Remember that neither ion will ever come to equilibrium because the movement of one opposes the other and the voltage of the living cell, despite the presence of leak channels for both Na+ and K+, will never reach zero because of the action of the Na+/K+ pump. We cannot say that the cell at rest is at equilibrium because there are a lot of forces driving the many ions in play but we do say that the cell is at steady state. See the GHK Equation on page 178. Electrical Signaling through Changes in Membrane Potential (Vm) We need some basic terminology to be able to proceed: Gated Ion channels: these open or close in response to some stimuli and when they do they allow movement of a specific ion which changes the membrane potential towards that of the equilibrium potential for that ion Three types of gated ion channels: 1) Voltage-gated channels: these open or close in response to changes in voltage (membrane potential) 2) Ligand-gated channels: these open or close in response to the binding of a chemical signal (ligand – which is almost always a neurotransmitter in neurons!) 3) Mechanically-gated channels: these open or close in response to mechanical stress and are associated with sensory responses (keep these in mind for chapter 10) Page 179: The change in membrane potential is named after the direction of change relative to that membrane’s resting potential! Note: because there is a voltage difference across the membrane we say that the membrane is polarized. Depolarization: the change in membrane potential towards less negative (or fully positive) potential Hyperpolarization: the change in membrane potential towards a more negative potential Repolarization: when the membrane returns to the resting potential There are two main types of electrical signals in neurons that result from the opening or closing of gated ion channels: 1) Graded Potentials [GPs]: (figure 7.10 – 7.12) Page 180-182 This type of potential is small, short range and reduces in size over distance. Most often these potentials are a result of neurotransmitter molecules binding to ligand-gated receptors on the dendrites – many of which are a result of a sensory stimulus (such as touch, sound, light etc). Why are they called graded? Because the magnitude of the response is proportional to the magnitude of the stimulus – it is Graded in strength. These potentials can be either hyperpolarizations (inhibitory) or depolarizations (stimulatory) and are greatly dependent upon the stimulus type, the signaling molecule, the particular type of neuron, and the specific ion channels that are targeted. Graded potentials determine if the cell will generate an AP – which will only be generated if the graded potential reaches a 5 predetermined threshold level (the threshold levels are determined by how depolarized the cell must be before activation of the Na+ voltage-gated channels and their positive feedback loop – see APs below). These potentials also decrease in strength with increasing distance from the signaling point (meaning decremental in nature) because of the increasing loss of current through the leak channels as the graded potential spreads away from its origin. The current (voltage) moves through the intercellular medium as the spreading of ions and is called electrotonic conduction. Single graded responses are usually too small to achieve threshold and begin an AP but multiple graded potentials which overlap in time (stimulated again and again) – called temporal summation - can evoke a response, as well as different sources of current (multiple stimuli received) – called spatial summation – can cause a response (see figure 7.13). Please listen to the Graded Potential Audio Lecture found on our Exam 2 materials page on our Canvas site! 2) Action Potentials: (figure 7.14, 7.16) Page 182-188 This potential change is very large and can travel long distances without any reduction in size. (See figure 7.15) The changes in permeability associated with an AP is due to the opening and/or closing of voltage-gated Na+ and K+ channels on the axon hillock which then propagates down the axon. These are “all-or-nothing” means either they happen or they don’t and when they do occur (which they do billions of times a second in your nervous system) they always look the same. The ionic basis of an action potential consist of 3 distinct phases: a) Depolarization: in about 1 msec the membrane potential changes from -70 mV to +30 mV because the cell becomes momentarily very permeable to Na+ (so the cell rapidly approached the Na+ equilibrium potential). Na+ Votage-gated channels have 2 gates: an activation gate and an inactivation gate (see figure 7.15). At rest, the Na+ activation gate is closed and the Na+ inactivation gate is open (Figure 7.14). During a depolarization the stimulus allows the activation gate to open (which causes a rapid increase in permeability to Na+ decreased R and increased g) but then after about 1 msec the Na+ inactivation gate swings shut (decreasing Na+ permeability – probably due to the change in voltage). Note: This Na+ inactivation gate cannot reopen until the membrane potential returns to resting state (below threshold)! So during repolarization the activation gate slowly closes again and most of the inactivation gates are still shut so no Na+ current flows. The opening of Na+ activation gates are regenerative, meaning that once one Na+ gate is open it causes more depolarization, which opens more Na+ activation gates to open etc (can we say positive feedback loop? – how does this end?). Going back to graded potentials for a second: a subthreshold stimulus is one that is capable of opening a few Na+ voltage-gated channels but not enough to be greater than the outward flow of K+ through leak channels. 6 b) Repolarization: again, this stage happens very quickly and changes the membrane potential from +30 mV back to -70 mV (resting) because Na+ permeability greatly decreases and K+ permeability increases. About 1 msec after depolarization the Na+ inactivation gates close (decreasing Na+ permeability) and this triggers the K+ voltage gated channels to open (increasing the permeability to K+) which allows K+ to flow outward and restore the resting potential. Please note: our model of K+ voltage gated channels only predict that they have one gate in contrast to the two-gated Na+ voltage-gated channels. As the cell repolarizes the K+ voltage gated channels close in a classic negative feedback loop. c) After-Hyperpolarization: I always learned this as simple hyperpolarization but we’ll go with your book. At this period, because increased K+ permeability remains elevated for a while, due to the slow nature of the K+ channels as they close, the membrane becomes more negative than the resting potential. However the K+ voltage gated channels do all eventually close so the membrane will return to resting Vm because of the leaks and pumps. So now all the channels are reset: the Na+ voltage-gated channel has its inactivation gate open and its activation gate closed and the K+ voltage-gated channels are closed. You are ready to fire another AP! Please listen to the Action Potential Audio Lecture on our Exam 2 materials page on our Canvas site. Refractory Period (pg 187): the period just following an AP the membrane is much less excitable and can be divided into 2 phases: 1) Absolute refractory period: this last for around 1-2 msec and spans the depolarization period and most of the repolarization period. The first reason for this is the positive feedback loop that the voltage gated Na+ channels have which will last until their inactivation gates close. The second reason is that the Na+ inactivation gates cannot themselves inactivate until the cell returns to the resting potential. You can see how these two reasons would preclude the cell from firing another AP during this time! Because of this period, a neuron can only fire as fast as 5001000 APs per second (Hz). 2) Relative refractory period: this period lasts around 5-15 msec. It is possible during this time to generate another AP but in order to do so the stimulus must be suprathreshold. Why? By this time most of the Na+ inactivation gates are inactivated and capable of opening again, however, the voltage-gated K+ channels are still open or partially open (which means the cell is still highly permeable to K+) so the potential is hyperpolarized for a short while so there would have to be a LOT of Na+ come in, in order to initiate another AP at this membrane potential. (Read on page 180 about TTX and TEA and their effects on voltage-gated ion channels!) 7 Refractory periods are the reason that APs cannot sum, are all-ornothing and they only move in one direction (does this make sense to you?) So How do APs code for the magnitude of the stimulus? -Unlike graded potentials they code for the magnitude in the frequency of APs generated. This statement means that a large stimulus creates more frequent APs and a smaller stimulus generates less frequent APs (really important to remember for chapter 10). We call this frequency coding. How do APs Propagate? Page 189-191 Remember that the ECF and ICF both have very low resistance to current flow so an area of depolarization can travel, via electrotonic conduction, away from the site of depolarization and trigger the voltage-gated Na+ channels nearby to open. This means that the entire axon does not depolarize at once but small area by small area as the current changes the Na+ channels open and once they open, more in that area open etc. The larger the diameter of the axon the faster the conduction, kind of like the larger diameter of hose you have, the faster water can flow through it (the less resistance it encounters). Plus, if you have myelinated axons, ions flow through the myelin is greatly impeded (has high resistance) because of its hydrophobic nature but the areas in between the myelin sheaths – the Nodes of Ranvier - are full of Na+ and K+ voltage gated channels. So in this case, the change in membrane potential spreads by electrotonic conduction and dissipates with distance like a graded potential (which isn’t a bit deal since Nodes are very close together) until it reaches a Node of Ranvier and the Na+ channels there and triggers them to initiate depolarization and then that depolarization spreads from there etc. Can you see how myelination can greatly increase the speed of conduction by working through local depolarizations instead of one continuous one as in unmyelinated neurons? So now you can deduce that those neurons that have to be very fast in order to function properly are going to have wide, myelinated axons (such as motor neurons) 8