Powerpoint
advertisement

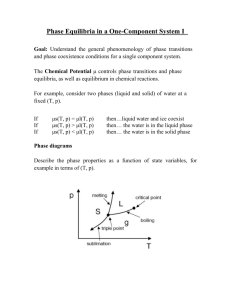
The nature of the plant community: a reductionist view JB Wilson Chapter 4 Mechanisms of coexistence and stability Liz Matthews and Matt Simon Introduction • How is coexistence between plant species achieved? What are the mechanisms? • Bastow’s fundamental statement: Plant communities comprise persistent, coexisting populations of several species • Fundamental problem = how is this achieved? • Resource (light, nutrients, etc) and competition (interference) limited • ‘Paradox of the plankton’ and monospecific stands of vegetation (Hutchinson 1941;1961) • HIGH diversity examples, coexisting seems to be working pretty well here… – Or are the niches just more abundant, ie resources? •“…we are concerned with mechanisms that allow species to coexist locally, i.e. mechanisms that are not due to imposed habitat heterogeneity within the area considered.” -JB JB Bastow’s 12 mechanisms of coexistence Stabilizing mechanisms Niche-differentiation 1. Alpha-niche Differentiation (type of resource and time of availability) 2. Environmental Fluctuation – season, decadal and gradual change Balances 3. Heterotroph challenges: Pest Pressure 4. Circular competitive networks Escape through movement 5. Allogenic Disturbance – disrupting growth mainly mechanically 6. Competition/dispersal Tradeoff 7. Initial Patch Composition 8. Cyclic Succession: movement of community phases: Equalizing mechanisms 9. Equal Chance (neutrality) 10. Inertia - Temporal and Spatial Inertia (aggregation) 11. Coevolution of Similar Interference Ability 12. Spatial Mass Effect (vicinism) What is a stabilizing coexistence mechanism vs. an Equalizing one? • Stabilizing mech. = increase-when-rare – Include negative abundance/density-dependence to counter species fluctuation – Alpha-niche differentiation, environmental fluctuation, balances, or escape through movement - these mechansims lead to true coexistence – If plant biomass is low then in order for it stay in the mix, it must increase its relative growth rate (fitness) – (Chesson in press) , this JB states is the “one necessary and sufficient phenomenon for maintaining a species in a mixture” Equalizing Mechanism • Mechanisms are: Equal Chance (neutrality), Inertia (temporal and spatial), Coevolution of Similar Interference Ability, Spatial Mass Effect (vicinism) • Ways in which species may persist for a time in unstable local coexistence, slowing exclusion by interference • A temporary competitive solution Alpha-niche Differentiation Mechanism No. 1 “impossible to disprove” • Increase-when rare occurs if a species is rare, its required resources will be more abundant = niche is not full, there’s room to grow – The potential to increase is there, but will this rare-species necessarily increase? • In contrast, populations are limited by full niche occupancy, coexistence occurs when resource requirements differ (niche seperation). • Stronger the niche differentiation, greater chance of coexistience between species…. Resource Gradients & Niche Differentiation Types and Temporal • Tilman (1976) “number of species able to coexist is equal to the number of resources, and then only if each species is limited by a different resource” is there no contradiction? • Vance (1984) coexistence on one resource occurs only “if each species interferes less with resource acquisition by the other than with resource acquisition by itself” – two species can coexist on one resource if: intraspecific competition for the resource > interspecific “ “ Resource Gradient cont. • How can a resource gradient be used as a niche differentiation mechanism? 1. 2. 3. Soil resources at different depths Seasonally pollinators Light-capture, canopy shapes (“subtle and devious ways...[of]…resource differentiation” JB) 4. • Temporal Resource gradient, two examples 1. 2. • Rooting depth Speed of reaction to resource availability, opportunistic species establish quickly when resources become available Seasonal: species’ growth patterns can separated by their response to length of day and temperature How much separation is needed? Heterotroph-imposed niches • Pollinators typically highly specialized – robbers, mimics, rewards, guides and warnings • Allele effect can alter the pollination niche and neutralize the increasewhen-rare mechanism • Mycorrhizal fungi assocations, niche differentiation is unlikely says JB. Do you agree? Chiloglottis trapeziformis, mimicry Niche extended by reaction • Species react to their environment and construct their own niche… • Species density patterns reflected in habitat create resource patchiness in the landscape, or is it the other way around? – Pelletier et al. (1999) found beech litter reduced soil [Ca] – Ehrenfeld et al. (2001) higher pH was found below two exotic species than native vaccinium – Other examples closer to home? Acidic coves…black walnut • Root plasticity and nutrient uptake • The tree canopy alters shade/temperature/humidity etc. Environmental Fluctuation Seasonal, annual and decadal change • In order for Env. Fluctuation to cause coexistence there has to be interaction between growth and resource supply • ‘Species can also separate along niche axes of vegetation phenology’ examples? • JB says “Separation in flowering times will reduce competition for pollinators, giving coexistence based on niche differentiation” – Difference between this and heterotroph-imposed niches? • ‘Flowering and fruiting niche gradient mediated by pollinators and dispersers’ Relative non-linearity vs. sub-additivity: two temporal variations leading to coexistence • Relative non-linearity: two species, two levels of response to resources – Fluctuating levels of a resource lead to increase-whenrare mechanism • Storage Effect - considerably stronger effect than relative non-linearity 1. 2. 3. 4. • Species must be competing affected by an environmental factor and respond different covariance b/w env. factor and intensity of competition Subadditivity, interaction b/w env. factor and effect of competition Timescale of resource depletion determines scale at which env. fluctuation affect coexistence Resource level [R] Pest (pathogens and herbivores) Pressure Heterotroph challenges • This increase-when-rare mechanism can occur with pests if three conditions are met – Impact is significant – Pest is specific to a particular species – Abundance (density)-dependence, pressure must be less on sparse • Abundant species will be kept in check When sparse the species will benefit from lack of infestation & fitness will increase = increase-when-rare effect Pest Pressure, cont. Abundance-dependance • JB cites the following examples of abundance-dependance mechanisms (Boudreau and Mundt 1997): 1. Decreased abundance of palatable/susceptible plants, which inhibits the dispersal of herbivores, disease spores, or disease-vectors 2. Flypaper effect for disease spores and possibly for insect pests, virus vectors and hence for the viruses they carry, in which the pest is caught by a passive surface 3. Alteration in the air flow and microclimate ? 4. Chemicals from associated species that repel herbivorous insects 5. Promotion by an associated species of natural enemies of the herbivorous insects, i.e their predators - used in gardenning for multiple species, but what’s another example? Jb then give various seedling mortality examples in which the ‘JanzenConnell’ hypothesis may explain Pathogens the considerable impact of fungal pathogens.. • No doubt that the impact can be significant. • Can pathogens that act in the soil be species specific? How about aerial transmission? JB balks… How about sudden oak death, chestnut blight, dogwood blight… How are these considered coexistence mechanisms? Are they too catastrophic? Coast live oak in Marin County, CA, May 2000 • These are all probably more density-dependent, but some do seem to target a particular species. Some empirical evidence and arguments for host specificity • C.E. Mitchell et al. (2002 and 2003) – At the Tilman established Cedar Creek site, found that infection dropped as species richness increased • C.E. Mitchell and Power (2006) “the transmission of specialist pathogens can be highly sensitive to the identity of other host species in the community” • For below-ground pathogens this has not been proven, yet…has it? Herbivory, general • Potential mechanism for coexistence via herbivory is similar to that for pathogens • Grover (1994) used the keystone concept, subordinate species are able to survive because of a controlling herbivore munching down the dominant Yucca moth in Yucca flower, not herbivory… Herbivory, of disseminules and seedlings • Herbivores can impact adult population abundance by limiting the seed input to the seedbank – But is there specificity? • Herbivory has been shown to be densitydependent, but its effects on density are not always consistent • Overall a weak case for for 2 out 3 of pest pressure coexistence mechanisms – Can any of his arguments be strengthened? Vegetative Herbivory • • Impact can be considerable Selectivity varies even though preferences do exist – – • Butterflies are often specialists Cattle are selective, but still generalists Some clear examples: 1. 2. – Beetle plagues keeping goldenrod pops. in check Vertebrate grazing increasing species diversity BUT, says JB, “when the sward is higher, light competition is more important…this has nothing to do with any mechanism of coexistence between plant species” Pest Pressure Conclusions • JB concludes that this mechanism most likely acts via diseases • Also concludes with a pretty good example of coexistence and cyclic succession – a pioneer grass is impacted by nematodes early in the season (& succession) – the pathogens and nematodes, with a synergistic effect, reduce the growth of the next dominant grass – The result is a patchy landscape of both species, with different phases of vigor associated with pest phases (4) Circular Interference Networks species A species C species B Interference relations between a set of species is intransitive, i.e. it cannot be arranged in a pecking order such that a species higher in the order always competitively excludes a species lower in the order Precautions in the interpretation of Circular Interference Network Mechanism: • Competitive abilities change per environment • RGR of species may change as community composition changes • Past studies exploring competitive abilities have been flawed… – Comparisons of performances in mixtures with those in monoculture Precautions in interpretation of Circular Interference Network Mechanism: Table 4.2. Which species has the higher interference ability? The starting biomass for both species was 1.00 Species Biomass in monoculture Biomass in mixture A 3.00 Decreases to… 2.77 Winner in mixture B 2.64 Increases to… 2.71 Connolly 1997 Various experiment do not find intransitivity in plant communities: • • • • Mouquet et al (2004) Roxburgh and Wilson (2000) Keddy et al (1998) Silvertown et al (1992) and (1994) So, do circular interference networks exist??? Theoretical processes that could lead to intransitivity: (1) (2) A A is taller than B and shades B out C produces an allelopathic chemical, toxic to A C (3) A o Tree A is taller than shrub B and shades B out grass C lowers the temperature, and suppresses seedlings of A B B is taller than C A B C shrub B shades out grass C, and is not affected by lower temperature (4) A C is shadetolerant, and scavenges nitrogen A is taller than B and shades B out C with A is taller than it, and shades it out C B C B is taller than C, and fixes N A is taller than B and shades B out B B with C is taller than it, and shades C out Any real world examples of this mechanism? (5) Allogenic Disturbance (disrupting growth, mechanically) • A between-patch mechanism • Newly disturbed patches and recovered patches must exist…disturbance must be frequent enough to produce patches at various stages of recovery …should this be considered coexistence? If we accept “between-patch” patterns as coexistence… • Then allogenic disturbance mechanism assumes distinct pioneer and climax species – May not always be true: Peterson and Pickett (1995) – Autosuccession • According to W&A’s discussion, pioneer vs. climax species traits appear to be most evident in temperate forests – Most tropical species are intermediate in respect to these traits – Poulson and Pratt (1996) found size of gaps in Michigan did impact species composition of regeneration (6) Interference vs. Dispersal Tradeoff • competitive abilities vs. dispersal abilities • Increase-when-rare: – If our competitor is sparse, there are empty sites for it to occupy and outcompete (due to increased growth rate) our disperser – If our disperser is sparse, there are many sites left over by the dispersal-limited competitor for it to occupy • The assumption inherent in this trade-off appears to be common (7) Initial Patch Composition • By chance, small transient patches are occupied by species in different proportions • Species in the majority will suppress species in minority if inter-specific interference > intra-specific interference • Does this constitute a stabilizing coexistence mechanism? Is there an “increasewhen-rare” component? • Why include it in the list? (8) Cyclic Succession: movement of community phases • Similar to mechanism 4 (circular interference networks), but involves the whole community • Again, a between-patch scale is necessary (9) Equal chance: neutrality “…any one of a number of species is equally likely to occupy and pre-empt by reaction a particular microsite.” • Dispersal, for example, is so unpredictable in practice that we can invoke chance – Unpredictable climate – Unpredictable disturbance events W&A’s discussion of equal chance: • “One cause would be that the probability of a disseminule reaching a site is proportional to its abundance.” – Interference ability are equal, “equivalence of competitors” So this mechanism implies there are no trade-offs? The more abundant species has a “better” (non-equal) chance of establishing…is this equal chance? What brought about abundance in the first place…competitive abilities, dispersal abilities, etc? W&A’s discussion of equal chance: • Equal chance sometimes used when species composition of a community does not appear to be correlated with an environmental factor, past or present – Perhaps explanatory variables were not measured – “…used as an excuse for failing to find vegetation/environment correlations is the last resort of the scoundrel.” • Does equal chance have a place in describing coexistence mechanisms? Chance and IBG: • MacArthur and Wilson model (1963) based on probabilistic immigration and extinction • W&A claim this hasn’t been proved often in field experiments (10) Inertia • Equalizing mechanism • Exclusion by interference of a superior competitor is slow and may allow stabilizing mechanisms to occur (10) Inertia: temporal • Small-scale dispersal switch due to niche construction • May be a result of dispersal limitation – Dalling et al 1998 found that seedlings on BCI were denser beneath conspecific adults, but this correlation was weaker for small-seeded spp. • May be a result of a seed bank and the storage effect • May be due to a switch (10) Inertia: temporal • May not apply equally to all species in a community – Short lived vs. long lived species • Inertia as a mechanism does not explain original coexistence: “if there is no coexistence, inertia cannot prolong it.” (10) Inertia: spatial • Spatial aggregation may delay exclusion, since exclusion occurs only at patch boundaries (11) Coevolution of Similar Interference Abilities • Aarssen (1983): selection pressure on the less-competitive species would cause it to become the better competitor • “unbelievable because it involves continual increases in interference ability…” (12) Spatial Mass Effect (vicinism) • A population may be maintained by constant immigration into a patch where the species cannot otherwise maintain itself • Rarely quantified • Metapopulation dynamics Conclusions • Stabilizing vs. equalizing mechanisms – Short-term vs. long term coexistence • What is the relative importance of each mechanism? Conclusions “…if the difference between the interference abilities of species is large, even the presence of a stabilizing mechanism may not prevent exclusion by interference and in this situation an equalizing mechanism might reduce the difference in interference ability between two species so that the stabilizing mechanism is able to cause coexistence.” (Chesson 2000). Bastow and Agnew’s order of importance: • • • • • • • • • • • • Alpha-niche differentiation Environmental fluctuation Pest pressure Spatial mass effect Allogenic disturbance Interference/dispersal tradeoff Temporal and spatial inertia Cyclic succession Circular interference networks Equal chance Co-evolution of similar interference ability Intitial patch composition Do we agree with this order??? Any other comments or reactions to the chapter? Ideas for next week • Expand upon metapopulation ideas • Expand upon equal chance/neutral theory: – Hubbell, S.P. (2005) Neutral theory in community ecology and the hypothesis of functional equivalence. Functional Ecology, 19, 166172