Lecture 28 Human Evolution II
advertisement

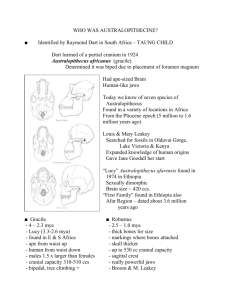
Mammals Grew 1,000 Times Larger After the Demise of the Dinosaurs The largest land mammals that ever lived, Indricotherium and Deinotherium, would have towered over the living African Elephant. Indricotherium lived during the Eocene to the Oligocene Epoch (37 to 23 million years ago) and reached a mass of 15,000 kg, while Deinotherium was around from the late-Miocene until the early Pleistocene (8.5 to 2.7 million years ago) and weighed as much as 17,000 kg. Fig. 3 Global fluctuations in maximum body size and various abiotic factors over the Cenozoic. F A Smith et al. Science 2010;330:1216-1219 Published by AAAS millions of years ago gibbon orangutan gorilla chimpanzee human 0 Hominidae 6 Hylobatidae 12 Pongidae Are gorillas and chimps each other's closest relatives? Filling the African “Ape Gap” from14- to 7-Myr Over the last year a number of new genera of great apes related to the modern Gorilla have been described that fill the gap. Ouranopithecus Samburupithecus Nakalipithecus Chororapithecus 9.6 –8.7 Myr ago. 9.6 Myr ago 9.9-9.8 Myr ago 10–11 Myr ago. First fossil chimpanzee Sally McBrearty and Nina G. Jablonski First unequivocal chimp fossils dated to ~545,000 bp. Contemporary with Homo erectus from the same site. Nature 437, 105-108 (1 September 2005) Placing confidence limits on the molecular age of the human– chimpanzee divergence Data from 167 nuclear proteincoding genes, correcting for rate heterogeneity and using 23.8-35 MYA as the date for the Old world Monkey – Ape split as a calibration time, suggest that the 95% confidence interval for the human - chimpanzee divergence is: 4.98 – 7.02 MYA Kumar et al. 2005 PNAS 102:18842-18847 Ardipithecus ramidus 4.5 mya Soon after the human-chimp split More ape-like than Australopithecus Ardipithecus fossils from 5.3 - 5.8 mya On the lineage to modern chimpanzee??? 2002 discovery of hominid from Chad with a mosaic of primitive (very small brain) and derived (small canines) features Sahelanthropus tchadensis 6-7 mya Orrorin tugenensis Femoral Morphology and the Evolution of Hominin Bipedalism Richmond & Jungers Morphological comparisons among femora of or attributed to (A) P. troglodytes, (B) O. tugenensis (BAR 1002'00), (C and D) Paranthropus robustus (SK 97 and SK 82, reversed), (E) A. afarensis (A.L. 288-1ap), (F) Paranthropus boisei (KNMER 1503, reversed), (G) early Homo (KNM-ER 1481), and (H) modern H. sapiens. Like other early hominin femora (C to F), BAR 1002'00 (B) is distinct from those of modern humans (H) and great apes (A) in having a long, anteroposteriorly narrow neck and wide proximal shaft. Early Homo femora (G) have larger heads and broader necks compared to early hominins. In addition to these features, modern human femora (H) have short necks and mediolaterally narrow shafts. Scale bar, 2 cm Bipedalism is a key human adaptation and a defining feature of the hominin clade. Fossil femora discovered in Kenya and attributed to Orrorin tugenensis, at 6 million years ago, purportedly provide the earliest postcranial evidence of hominin bipedalism, but their functional and phylogenetic affinities are controversial. We show that the O. tugenensis femur differs from those of apes and Homo and most strongly resembles those of Australopithecus and Paranthropus, indicating that O. tugenensis was bipedal but is not more closely related to Homo than to Australopithecus. Femoral morphology indicates that O. tugenensis shared distinctive hip biomechanics with australopiths, suggesting that this complex evolved early in human evolution and persisted for almost 4 million years until modifications of the hip appeared in the late Pliocene in early Homo. Science 21 March 2008: Vol. 319. no. 5870, pp. 1662 - 1665 Homo Australopithecus Earliest Homonids Australopithecus anamensis 3.9 – 4.2 mya A. afarensis ‘Lucy” 3.0 – 3.9 mya A. africanus & A. garhi 2.4 – 2.8 mya A. robustus / boisei / aethiopicus 1.0 – 2.7 mya “Lucy” Australopithecus afarensis 3 mya Laetoli Tracks - 3 mya Lack of a splayed big toe suggests full bipedalism by 3 mya Lack of a splayed big toe suggests full bipedalism by 3 mya = bipedality in A. afarensis Lucy Modern Human Chimp VERTEBRAL RECONSTRUCTION A. arafensis Recent analysis of fossil wrist bones suggest the A. afarensis may have occasionally used knuckle walking like modern chimps and gorillas. RECONSTRUCTIONS BASED ON MUSCLE ATTACHMENTS An early Australopithecus afarensis postcranium from Woranso-Mille, Ethiopia Haile-Selassie et al. Dated to 3.6 million years ago,the robust male stood between 1.5 and 1.7 meters tall, about 30% larger than Lucy. Isolated bones of other individuals suggest that some males were even larger, so the new skeleton doesn’t settle a long-standing debate over just how much sexual dimorphism there was in A. afarensis PNAS | July 6, 2010 | vol. 107 | no. 27 | 12121–12126 Australopithecus afarensis ~4 ft. tall (sexually dimorphic?) Mixed Bipedal / climbing, curved phalanges Brain size MUCH closer to chimp than Modern Human Fig. 1 Craniodental elements of Au. sediba L. R. Berger et al., Science 328, 195-204 (2010) Published by AAAS Fig. 2 Associated skeletal elements of MH1 (left) and MH2 (right), in approximate anatomical position, superimposed over an illustration of an idealized Au L. R. Berger et al., Science 328, 195-204 (2010) Published by AAAS Australopithecus sediba at 1.977 Ma and Implications for the Origins of the Genus Homo The right forearm and hand (hand skeleton ∼12.3 centimeters long) of (Australopithecus sediba), specimen Malapa Hominin 2. Papers in this issue present a detailed look at the hands, feet, pelvis, brain endocast, and age of this hominid, which lived 2 million years ago, near the emergence of our genus, Homo. Pelvic tilt. The pelvic blades of Au. africanus (left) flare more widely than those of the younger Au. sediba from South Africa (right; reconstructed parts are in gray or white). Au. sediba's hand has some humanlike traits, but its arm is long and primitive. Fig. 3 Comparisons of virtual endocasts in (A) superior, (B) inferior, (C) anterior, and (D) left lateral views. K J Carlson et al. Science 2011;333:1402-1407 Published by AAAS Fig. 2 Comparison of the MH1 (left), Sts 14 (center), and MH2 (right, mirror-imaged) pelves in anteroinferior (top row) and anterosuperior (bottom row) views. Comparison of the MH1 (left), Sts 14 (center), and MH2 (right, mirror-imaged) pelves in anteroinferior (top row) and anterosuperior (bottom row) views. Areas represented in white or light gray in the MH1 and MH2 pelves represent reconstructed portions of the pelvis (SOM text S1). Sts 14 is attributed to Au. africanus and is represented by the virtual reconstruction of (41). Scale bar in centimeters [note that the anterosuperior view of Sts 14, as provided by (41), is in a slightly different orientation than those of MH1 and MH2]. An additional comparison is provided in fig. S7 J M Kibii et al. Science 2011;333:1407-1411 Published by AAAS Fig. 6 Relative length of the thumb in the Au. sediba MH2 hand. Relative length of the thumb in the Au. sediba MH2 hand. Shown is a box-and-whisker plot of the relative length of the thumb calculated as a ratio of the total length of the Mc1 and first proximal phalanx to the total length of the Mc3 and third proximal and intermediate phalanges within the same individual (bones highlighted in dark gray in outline of MH2 hand) in all taxa apart from Au. afarensis, for which that ratio is derived from multiple individuals from different sites (30, 38). MH2 has a relatively longer thumb than that of other hominins and falls outside the range of variation in modern humans (highlighted by shaded box). T L Kivell et al. Science 2011;333:1411-1417 Published by AAAS A. africanus 2.5 mya “Taung” child Approx. 3-4 years old A. africanus Asa Issie, Aramis and the origin of Australopithecus Tim D. White et al. The origin of Australopithecus, the genus widely interpreted as ancestral to Homo, is a central problem in human evolutionary studies. Australopithecus species differ markedly from extant African apes and candidate ancestral hominids such as Ardipithecus, Orrorin and Sahelanthropus. The earliest described Australopithecus species is Au. anamensis, the probable chronospecies ancestor of Au. afarensis. Here we describe newly discovered fossils from the Middle Awash study area that extend the known Au. anamensis range into northeastern Ethiopia. The new fossils are from chronometrically controlled stratigraphic sequences and date to about 4.1–4.2 million years ago. They include diagnostic craniodental remains, the largest hominid canine yet recovered, and the earliest Australopithecus femur. These new fossils are sampled from a woodland context. Temporal and anatomical intermediacy between Ar. ramidus and Au. afarensis suggest a relatively rapid shift from Ardipithecus to Australopithecus in this region of Africa, involving either replacement or accelerated phyletic evolution. Australopithecus “Lucy” Chimp Ardipithecus Nature 440, 883-889 (13 April 2006) Robust Australopithecines Genus: Paranthropus ROBUST SPECIES P. aethiopicus 2.6 mya P. boisei 2.6 - 1.0 mya P. robustus 2.0 – 1.2 mya Large sagittal crests, massive jaw muscles and teeth Small brains Number of biological species??? P. boisei 2.6 - 1.0 mya P. aethiopicus The “Black Skull” Kenya 2.5 mya Australophitecus garhi (n=1) Possible ancestor to Homo? 2.5 mya from eastern Africa NEW HOMONID FROM 3.5 MYA Mosaic of primitive and derive characters Flat face and smallish teeth Kenyanthropus platyops MULTIPLE HOMONID SPECIES BETWEEN 3.5 – 2.0 MYA K. platyops K. rudolfensis Multiple cooccurring hominid species Sahelanthropus tchadensis 6-7 mya