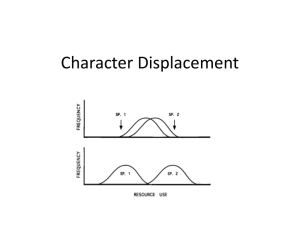
ele12043-sup-0001-AppendixS1-TableS1-S8
advertisement

Supporting Information Species interactions constrain geographic range expansion through evolutionary time Alex L. Pigot* and Joseph A. Tobias *To whom correspondence should be addressed. E-mail alex.pigot@zoo.ox.ac.uk Contents Expanded Materials and Methods (Appendix S1) Supplementary References Supplementary Figures (S1–S4) Data Tables (S1) Statistical Tables (S2–S8) 1 Appendix S1 Expanded Materials and Methods Sister species ages We obtained sister species pairs and estimated divergence times from a recently published molecular phylogeny of Furnariidae (Derryberry et al. 2011). This is unusually well sampled for a diverse radiation, containing 97% of currently described species (N = 285), and highly resolved, with more than 80% of nodes having >0.95 posterior probability (Table S1). Derryberry et al. (2011) generated a maximum clade credibility (MCC) tree based on multiple mitochondrial and nuclear genes and dated nodes using a relaxed clock Bayesian approach in BEAST v. 1.5.2. Substitution model, rate heterogeneity, and base frequencies were unlinked across partitions. No restrictions were placed on the topology so that topological uncertainty was factored into the divergence date estimates. In the absence of a detailed fossil record, Derryberry et al. (2011) used biogeographic events to place priors on divergence times, including the closure of the Panamanian Isthmus (3 million years ago [Ma]) and uplift of the Andes Cordillera (3.6 Ma). Priors were applied to nodes separating sister species with current distributions either side of these barriers. While absolute rates of secondary sympatry will be sensitive to inaccuracies in divergence date estimates, this will apply equally across all lineages and thus our assessment of how rates of secondary sympatry vary across lineages in response to divergence in the α-niche are unlikely to be biased. Systematic biases in the estimation of divergence dates are also unlikely to explain our key findings regarding the temporal dynamics of secondary sympatry. If the timing of recent nodes were systematically underestimated then this could potentially lead to an apparent speed-up in the rate of secondary sympatry with time since divergence. However, under this scenario rates of secondary sympatry would be expected to have accelerated with time since speciation across all species pairs regardless of habitat affiliations. Instead, our 2 analysis shows that rates of secondary sympatry have only accelerated with time since divergence in sister species with conserved β-niches, a pattern only expected if biotic interactions limit sympatry. In fact, it is generally expected that divergence times for recent nodes will tend to be overestimated compared to older nodes because of the comparatively large effect of ancestral polymorphism for young species (Edwards & Beerli 2000; Weir 2006). Inaccuracies in divergence estimates are therefore likely to make our analysis conservative with respect to detecting speed-ups in the rate of secondary sympatry with time since speciation. Geographic data Sister species were assigned as either sympatric (with overlapping breeding distributions) or allopatric (with non-overlapping breeding distributions) (Mayr 1942) according to vector polygon range maps kindly provided by C.D.L Orme and I.P.F Owens (Orme et al. 2005). Avian distribution maps were based on the avian taxonomy of Sibley and Monroe (1990) and so we updated species ranges to reflect recent taxonomic revisions, matching the distribution data to the lineages represented in the Furnaridae tree. Range polygons were edited in ArcMap v10. We used a binary classification of sympatry, rather than the magnitude of overlap, because we were interested simply in whether species are able to co-exist or not (Weir & Price 2011). We did not distinguish between cases of parapatry (abutting geographic ranges) or allopatry (geographically isolated) because for the purpose of our analysis both have the same pattern of non-overlapping ranges. While the range maps we use provide the most detailed information on species distributions available at these scales, they tend to overestimate the extents of species occurrence, particularly in regions with complex topography (Hurlbert & Jetz 2007). We therefore followed previous studies (e.g. Weir & 3 Price 2011) in using additional literature searches to further refine estimates of range overlap. This removed spurious cases of sympatry (N = 32) caused by mapping inaccuracies, or low mapping resolution. In most of these cases, the degree of sympatry predicted by range polygons was limited to only very localised co-occurrence (< 10% of the smaller species range) (Fig S1). In a few cases, however, range overlap estimated from polygon data was high while secondary sources revealed that the species were not in sympatry (e.g. Cinclodes palliatus and C. atacamensis). This was usually associated with occurrence in the Andes, where turnover in species distributions occurs over finer spatial scales than the resolution of polygon maps. Sister species range sizes and their areas of overlap (km2) were calculated using the mapTools and PBSmapping libraries in R version 2.10.1 (Team, 2010). Species Β-niches We quantified β-niches using information on species macro-habitat affinities and elevational range limits (Table S1) taken from Stotz et al. (1996) and supplemented with additional sources for species representing recent taxonomic revisions. To maximise sample size, we used three relatively coarse habitat categories (forest [F1–F15], scrub/grassland [N1–N14] and aquatic habitats, including lakes/rivers/coasts [A1–A12]) when assigning β-niches (codes refer to habitat types given in Stotz et al. [1996]). We used the first two categories listed by Stotz et al. (1996) for each species, and treated these as primary and secondary habitat types. We used additional sources from primary literature to update data for a small number of poorly known species, or for recent taxonomic revisions. We note that our results remained qualitatively unchanged when using the finer habitat classifications within the broad categories outlined above (Table S2, S3). 4 Estimating rates of secondary sympatry We modelled the process of secondary sympatry using continuous time multi-state Markov models. This class of models is appropriate for processes that can be described as a set of transition events between states but where the exact times of these events are not actually observed. The approach has been widely used in the phylogenetic literature to model the evolution of discrete character traits (e.g. Pagel 1994; Maddison & Maddison 2011). Here we use it to model sympatry as a combined trait of two sister species rather than a property of an individual lineage. Each sister pair contributes two observations: the geographic state at the time of population divergence and that of the present day. We assume all species pairs were initially allopatric. This assumption is valid for birds because there is substantial evidence that allopatry (or parapatry) is the predominant mode of speciation, with sympatric speciation being very rare (Coyne & Price 2000; Phillimore et al. 2008; Price 2008). We considered a model with two states, allopatry and sympatry, and use maximum likelihood to estimate the rate of transition from allopatry to sympatry. All models were implemented in R using the packages msm (Jackson 2011) and nhm (Titman 2011). An advantage of using multi-state Markov models to estimate rates of sympatry amongst sister species is that it is robust to differences in the distribution of species ages arising from variation in speciation and/or extinction rates. Weir and Price (2011) used an alternative method based on the ages of the youngest and oldest sympatric and allopatric nodes in the phylogenetic tree to provide upper and lower bounds on the time required to attain sympatry. However, this method may bias estimated times to sympatry because (1) the observed age of sympatric nodes is constrained by the overall distribution of divergence times, and (2) sympatry need only be attained by any two member species of sister clades potentially confounding rate estimates with clade richness. Our method avoids these limitations because we estimate transition rates across the age distribution of both sympatric 5 and allopatric lineages, and because we focus only on sister species pairs. Importantly, this has the further advantage of allowing us to examine the temporal patterns of secondary sympatry. Estimating the return to allopatry The instantaneous probabilities of moving between states are governed by a set of transition intensities (qallo-sym and qsym-allo), where the order of subscripted states denotes the direction of movement (allopatry to sympatry, and vice versa). In our standard models presented in the main text (Constant-Rate, Rate-Switch and Time-Variable models) we treat the movement from allopatry to sympatry as an irreversible process by fixing qsym-allo = 0. Following the attainment of sympatry, however, some species potentially undergo a contraction or shift in their geographic ranges leading to a new phase of allopatry. We explored this possibility using a State-Reversible model in which we allowed qsym-allo to vary and thus estimated the rate at which species pairs return to allopatry following a period in sympatry. Estimated return rates to allopatry were extremely slow and did not differ from zero at the 95% confidence level (Table S5). We compared the fit of the State-Reversible model to our Constant-Rate model using a likelihood ratio test and failed to reject our initial assumption that the transition rate from sympatry to allopatry is zero. This was true regardless of whether we included all species pairs or only those with conserved β-niches, and was robust to variations in the threshold defining distances between species ranges (Table S5). Across Furnariid sister pairs, secondary sympatry therefore appears to be a one-way process. Quantifying the effects of species interactions and dispersal constraints We examined the role of biotic interactions on the rate of secondary sympatry using two complementary methods. First, we tested whether rates of secondary sympatry depend on the morphological distance separating species. We note that although present day species 6 contrasts will tend to exceed those prevailing during the course of divergence, here we are simply interested in the shape of the relationship between sympatry rates and morphological distances. Our models therefore assume that while rates of morphological divergence may vary across lineages, species pairs share a common model of trait divergence. Second, we tested whether rates of secondary sympatry vary with time since speciation. We focus on a simple ‘Rate-Switch model’ in which an initial constant rate (S1), switches to a different rate (S2) at time ts (Figure S2). We estimated the maximum likelihood break point by fitting our models using different values of ts in increments of 100 kyr from 0.1 Myr to a maximum value (tmax) determined by the distribution of sister species ages (Figure S3). Specifically, we only considered break point ages where at least 10 sister pairs occurred after the break point because smaller sample sizes prevented model convergence and reliable parameter estimates (Table S2). We highlight that the timing of the shift in the rate of secondary sympatry is relative to the timing of speciation and not an absolute date in the past. In a selection of sister species of varying age these rate shifts will therefore have occurred at various times in the past (Figure S2). Assuming that the strength of ecological interactions between species is inversely related to the time since their divergence (Figure S4a), it may be possible to use the rates estimated under the Rate-Switch model to partition the delay in the timing of secondary sympatry arising due to biotic interactions and dispersal constraints (Figure S4). The rates (S1) and waiting times to secondary sympatry (W1) amongst recently diverged species prior to the break point will be influenced by the combined effects of both dispersal constraints and strong biotic interactions (Figure S4a–c). In contrast, the rates (S2) and waiting times to secondary sympatry (W2) amongst long diverged species after the break point will be influenced primarily by dispersal constraints because biotic interactions will presumably be weaker (Figure S4a–c). The difference between the observed waiting time to sympatry (W) 7 and that expected due to dispersal constraints (W2) thus provides an estimate for the delay in secondary sympatry arising due to biotic interactions (ΔW = W – W2) (Figure S4d). We note that estimates of ΔW were robust to data selection procedures, including the geographic distance separating sister species (Table S2). Clade-wide shifts in rates of sympatry It is plausible that a single clade-wide and contemporaneous shift in the rate of secondary sympatry could generate a pattern analogous to that expected from species interactions. To test this idea we compared the fit of the Constant-Rate model to a Clade-Rate-Switch model in which an initial rate S1 can switch to a higher or lower rate S2 at time ta in the past (Figure S2b). We highlight that in contrast to the Rate-Switch model (Figure S2a), the timing of the shift in the rate of secondary sympatry occurs at an absolute date in the past. In a selection of sister species of varying age these rate shifts will therefore have occurred at various times relative to the timing of speciation (Figure S2b). We denote the timing of the rate shift as ta (rather than ts in the Rate-Switch model [Figure S2a]) to highlight that this represents an absolute date in the past where all sister pairs in existence experienced a shift in rate. We explored different values of ta from 10 Ma to 1 Ma in increments of 1 Myr, selecting the switch point with the highest likelihood as the best model. In addition to considering a single shift in rates, we also fit a Clade-Time-Variable model in which rates are allowed to change continuously through time. Models in which the rate of secondary sympatry has undergone a clade-wide decline provide a better fit to the date that assuming a constant arte of secondary sympatry (Table S8). However, we view biogeographical explanations for this finding as highly unlikely for two main reasons. First, the furnariids are spread across the Neotropics, from Tierra del Fuego to Mesoamerica and from sea level to over 5000m in elevation (Derryberry et al. 8 2011). Species pairs are thus separated by a multitude of different environmental gradients and/or biogeographic barriers with contrasting histories (Table S1). Second, bird species are known to respond to these barriers individualistically, with speciation and recolonisation occurring at different rates according to variation in ecology, and in particular dispersal limitation (Burney & Brumfield 2009). For example, avian lineages dispersing both southwards and northwards after the closure of the Isthmus of Panama have tended to produce speciation and recolonisation events at seemingly random intervals (Dacosta & Klicka 2008; Miller et al. 2008). This lack of synchronicity is unlikely to generate bursts of secondary sympatry amongst sister species at a given point in time, and instead is expected to generate heterogeneity in rates of secondary sympatry over time (Figure 1b). A more plausible explanation for any possible clade-wide slowdown in the rate of sympatry could be related to niche filling in highly diverse and ecologically saturated communities (Rundell & Price 2009). Although to our knowledge this pattern has never been detected, our finding that rates of secondary sympatry are reduced amongst ecologically similar species is consistent with the idea that range expansions are likely to become increasingly constrained as niche space is saturated. 9 Supplemental References Burney C.W. & Brumfield R.T. (2009). Ecology predicts levels of genetic differentiation in Neotropical birds. Am. Nat., 174, 358-368. Coyne J.A. & Price T.D. (2000). Little evidence for sympatric speciation in island birds. Evolution, 54, 2166-2171. Dacosta J.M. & Klicka J. (2008). The Great American Interchange in birds: a phylogenetic perspective with the genus Trogon. Mol. Ecol., 17, 1328-1343. Derryberry E.P., Claramunt S., Derryberry G., Chesser R.T., Cracraft J., Aleixo A. et al. (2011). Lineage diversification and morphological evolution in a large-scale continental radiation: the Neotropical ovenbirds and woodcreepers (Aves: Furnariidae). Evolution, 65, 2973-2986. Edwards S.V. & Beerli P. (2000). Perspective: gene divergence, population divergence, and the variance in coalescence time in phylogeographic studies. Evolution, 54, 18391854. Hurlbert A.H. & Jetz W. (2007). Species richness, hotspots, and the scale dependence of range maps in ecology and conservation. P. Natl. Acad. Sci. USA, 104, 13384-13389. Jackson C.H. (2011). Multi-state models for panel data: The msm package for R. J. Stat. Softw., 38, 1-28. Maddison W.P. & Maddison D.R. (2011). Mesquite: a modular system for evolutionary analysis. v2.75. http://mesquiteproject.org. Mayr E. (1942). Systematics and the origin of species. Columbia University Press, New York. Miller M.J., Bermingham E., Klicka J., Escalante P., Do Amaral F.S.R., Weir J.T. et al. (2008). Out of Amazonia again and again: episodic crossing of the Andes promotes diversification in a lowland forest flycatcher. Proc. R. Soc. B, 275, 1133-1142. Orme C.D.L., Davies R.G., Burgess M., Eigenbrod F., Pickup N., Olson V.A. et al. (2005). Global hotspots of species richness are not congruent with endemism or threat. Nature, 436, 1016-1019. Pagel M. (1994). Detecting correlated evolution on phylogenies: a general method for the comparative analysis of discrete characters. Proc. R. Soc. B, 255, 37-45. Phillimore A.B., Orme C.D.L., Thomas G.H., Blackburn T.M., Bennett P.M., Gaston K.J. et al. (2008). Sympatric speciation in birds is rare: insights from range data and simulations. Am. Nat., 171, 646-657. 10 Price T. (2008). Speciation in birds. Roberts and Company, Colorado. Revell L.J. (2009). Size-correction and prinicpal components for interspecific comparative studies. Evolution, 63, 3258-3268. Rundell R.J. & Price T.D. (2009). Adaptive radiation, nonadaptive radiation, ecological speciation and nonecological speciation. Trends Ecol. Evol., 24, 394-399. Sibley C.G. & Monroe B.L. (1990). Distribution and taxonomy of the birds of the world. Yale University Press, New Haven. Stotz D.F., Fitzpatrick J.W., Parker T.A. & Moskovits D.K. (1996). Neotropical birds: ecology and conservation. The University of Chicago Press Chicago. R Development Core Team (2010). R: a language and environment for statistical computing. Vienne, Austria: R Foundation for Statistical Computing. Retrieved from http://www.R-project.org. Titman A.C. (2011). Flexible nonhomogeneous markov models for panel observed data. Biometrics, 67, 780-787. Weir J.T. (2006). Divergent timing and patterns of species accumulation in lowland and highland Neotropical birds. Evolution, 60, 842-855. Weir J.T. & Price T.D. (2011). Limits to speciation inferred from times to secondary sympatry and ages of hybridizing species along a latitudinal gradient. Am. Nat., 177, 462-469. 11 Figure S1 Range overlap (%) calculated from polygon range maps across a) all sister species pairs, b) confirmed sympatric species and c) cases of spurious or localised sympatry. 12 Figure S2 Modelling of changes in the rate of secondary sympatry through time (S) under the a) RateSwitch model and b) Clade-Rate-Switch model. Species divergence times and present day geographic states (green rectangles) are shown for four sister species pairs. Red bars on accompanying graphs show the rate of secondary sympatry prior to (S1) and after (S2) the break point. In a), ts is time following speciation when there is a significant shift in S. The relative timing of the shift after speciation is identical across lineages, but the absolute date of the shift is variable because sister pairs vary in age. As the age of the top pair is < ts it has yet to undergo a shift in S. In b), ta represents the time before the present when there was a shift in S. This shift occurs at the same absolute time across all species pairs that were present at the time of the shift, but the timing of the shift relative to the timing of speciation varies, again because sister pairs vary in age. Pairs arising after the rate shift (i.e. closer to the present, so that age < ta) are characterised purely by S2. In a), the increase in rates after the putative break point corresponds to that estimated across furnariids. In b) a decrease in rates is shown because this leads to a similar pattern of sympatry across present day sister species (Table S8). 13 Figure S3 Likelihood of different break points (ts) in the Rate-Switch model for a) all sister pairs sisters with conserved β-niches, and sisters with conserved β-niches and that were separated by b) < 1000 km, c) < 500 km, d) < 250 km and e) < 125 km. Break points not significantly different from the maximum log-likelihood value (black triangle) at the 95% confidence level (i.e. within 1.92 loglikelihood units) are highlighted in red. N = sample size. 14 Figure S4 Conceptual model for estimating the delay in secondary sympatry arising due to dispersal limitation (W2) and species interactions (ΔW) between sister lineages (see ‘An evolutionary model of species co-occurrence’ in the main text for justifications). We assume (a) that the strength of species interactions (indicated by the width of the dark green triangle) declines with time since speciation, while the influence of dispersal limitation is constant or increasing; and (b) that weakening competition as species diverge ecologically may lead to an upward shift in the rate of secondary sympatry (S) from S1 to S2 at time ts indicated by the vertical dashed line. These different rate estimates generate (c) variation in the expected distribution (coloured curves, with colours corresponding to those in [b]) and average waiting times (vertical lines) to sympatry. This allows us to partition the overall waiting time to sympatry (W) into components due to dispersal limitation and species interactions (d). While the overall waiting time to sympatry (W) will be driven by both dispersal limitation and species interactions, the waiting time following the rate shift (W) will primarily be due to dispersal limitation. 15 Table S1 Furnariid sister species pairs included in the analysis, with details on their estimated times of divergence, geographic states (allopatry/sympatry), habitats and elevation ranges. Sister Species Age Sympatric (Myr) 1 Anabacerthia amaurotis 1 Philydor lichtensteini 2 Anabazenops fuscus 2 A. dorsalis 3 Asthenes anthoides 3 A. hudsoni 4 Asthenes dorbignyi 4 A. baeri 5 Asthenes humilis 5 A. modesta 6 Asthenes maculicauda 6 A. virgata 7 Asthenes ottonis 7 Schizoeaca palpebralis 8 Asthenes pudibunda 8 Schizoeaca vilcabambae 9 Asthenes pyrrholeuca 9 Schizoeaca harterti 10 Asthenes sclateri 10 A. wyatti 11 Automolus leucophthalmus 11 A. lammi 12 Automolus rubiginosus Distance between ranges (km) conserved Primary F4 β-niche 5.43 1 0 1 6.12 0 1575 1 4.45 0 718 1 1.49 0 0 0 Habitat Elevation (m) Secondary 3 3 F1 F3 0 1100 4 N7 N1 0 1500 5 0 0 3 F6 1800 4300 11 N1 0 0 4 3500 4800 4 N2 1.49 0 10 1 N10 N9 61 1 N7 0 1634 1 1.23 0 0 0 0 217 1 1.54 0 0 1 3 4300 3 3300 4300 5 N2 N3 2800 3700 3 3000 3500 3 N2 F6 2500 4000 5 N10 F5 2800 3400 4 N1 N2 0 3000 3 N10 F5 2900 3500 3 2000 2900 2 3000 4300 8 0 1000 3 0 1000 2 0 2400 9 N7 N9 F1 F1 16 4600 N9 N10 0.19 0 2900 N10 N10 3.63 3 950 N9 0 1500 1250 1 1.83 600 0 0 1 data (n) 350 0 87 Max F4 2.19 0 Min F1 N7 1.33 Morphological F4 F1 12 Hylocryptus erythrocephalus 13 Automolus rufipileatus 13 A. melanopezus 14 Campylorhamphus procurvoides 14 C. trochilirostris 15 Certhiaxis mustelinus 15 C. cinnamomeus 16 Cinclodes albiventris 16 C. olrogi 17 Cinclodes antarcticus 17 C. fuscus 18 Cinclodes aricomae 18 C. excelsior 19 Cinclodes nigrofumosus 19 C. taczanowskii 20 Cinclodes palliatus 20 C. atacamensis 21 Coryphistera alaudina 21 Anumbius annumbi 22 Cranioleuca albiceps 22 C. marcapatae 23 Cranioleuca antisiensis 23 C. curtata 24 Cranioleuca muelleri 24 C. vulpina 25 Cranioleuca pallida 25 C. pyrrhophia 26 Cranioleuca subcristata 26 C. hellmayri 27 Cranioleuca sulphurifera F7 3.36 3.57 1 0 0 0 1 1 0.31 1.03 2.19 0.12 2.70 10.49 1.74 1 0 1 0 0 0 1 0 0 989 0 1255 104 0 0 240 1 1 0 1 1 1 1 1 1.67 0 1 0 0 1 1 F2 0 850 6 F1 0 0 4 0 500 7 F4 0 1200 21 A1 A8 0 0 3 A1 F14 0 1000 4 N10 0 5000 3 N2 1600 2800 3 F1 0 0 5 N10 A4 N9 0 4900 3 F6 N3 3500 4600 2 N10 N3 3200 5200 3 A4 0 0 3 A4 0 0 4 A10 N9 4400 5000 5 N9 A9 2800 4900 4 N1 0 700 4 N6 0 1000 7 F4 2200 3300 4 2400 3300 3 F4 1200 3100 6 F4 800 2500 5 F2 0 0 3 0 800 5 F8 0.20 0.54 2.24 0 0 1 847 62 0 1 1 1 17 5 F2 F4 0.30 1800 F3 F2 6.11 600 F5 N12 F4 800 2200 3 F1 F7 0 2500 7 F4 F1 300 2300 3 F4 F15 1500 3000 2 0 0 3 A1 27 Limnoctites rectirostris 28 Dendrocincla fuliginosa 28 D. anabatina 29 Dendrocincla merula 29 D. tyrannina 30 Dendrocolaptes certhia 30 D. sanctithomae 31 Dendrocolaptes picumnus 31 D. hoffmannsi 32 Dendroplex kienerii 32 D. picus 33 Drymornis bridgesii 33 Drymotoxeres pucherani 34 Furnarius cristatus 34 F. rufus 35 Furnarius figulus 35 F. leucopus tricolor 36 Geositta crassirostris 36 G. rufipennis 37 Geositta isabellina 37 G. saxicolina 38 Geositta punensis 38 G. cunicularia 39 Hylexetastes perrotii 39 H. stresemanni 40 Hylocryptus rectirostris 40 Clibanornis dendrocolaptoides 41 Lepidocolaptes angustirostris 41 L. albolineatus 42 Lepidocolaptes leucogaster 4.57 10.50 4.29 1.17 0 0 0 0 0 0 162 0 1 0 1 1 A1 0 1000 3 F1 0 1200 12 F1 0 1250 6 F1 0 600 4 F4 1300 2500 4 F1 0 1300 13 F1 0 1000 3 0 2800 5 0 0 3 F1 F4 F1 7.33 8.54 1 0 0 1117 1 0 F2 0 0 5 F3 F7 0 900 4 F7 N1 0 1000 3 2100 2950 4 F4 2.91 3.54 10.18 1 1 0 0 0 351 1 1 0 0 1000 3 N14 N1 N13 0 3500 4 F3 F15 0 900 3 F2 F3 0 800 10 600 2500 4 N2 3100 4400 8 N9 1800 3100 3 N9 3700 4900 8 3200 4600 9 0 4800 18 F1 0 0 11 F1 0 0 3 F8 600 1200 3 F1 0 1000 3 0 1200 4 0 1100 13 950 4000 4 N2 N9 5.67 4.00 0 1 1500 0 0 1 N9 N7 2.92 3.76 3.69 0 0 0 0 0 0 1 1 1 F7 N9 N4 F1 2.32 0 0 1 18 F11 F10 42 L. affinis 43 Leptasthenura andicola 43 L. aegithaloides 44 Leptasthenura striata 44 L. pileata 45 Leptasthenura striolata 45 L. platensis 46 Margarornis bellulus 46 M. stellatus 47 Nasica longirostris 47 Dendrexetastes rufigula 48 Ochetorhynchus phoenicurus 48 O. ruficaudus 49 Phacellodomus dorsalis 49 P. maculipectus 50 Phacellodomus sibilatrix 50 P. striaticeps 51 Phacellodomus striaticollis 51 P. ruber 52 Philydor erythrocercum 52 P. fuscipenne 53 Philydor pyrrhodes 53 Heliobletus contaminatus 54 Philydor ruficaudatum 54 Anabacerthia variegaticeps 55 Philydor rufum 55 P. erythropterum 56 Phleocryptes melanops 56 Limnornis curvirostris 57 Premnoplex tatei 2.76 3.47 0.27 0.36 9.39 3.81 0 1 0 0 1 0 38 0 374 198 0 0 1 1 1 1 1 0 F4 F11 1000 3100 3 N2 N3 3200 4700 3 N1 N2 0 4300 7 N2 F6 1550 3800 5 N2 F6 2800 4400 4 F9 N11 750 1200 2 F8 N11 0 800 3 F4 F5 1350 1600 2 F4 F5 900 2200 3 F2 F3 0 500 5 F1 F2 0 950 5 N1 N7 0 1200 5 1800 4200 4 N2 1.48 1.67 1.58 2.63 7.70 4.08 4.32 0 0 0 0 0 0 1 1701 0 0 90 1130 101 0 1 0 1 1 1 1 1 N2 2050 2800 4 N2 F7 1000 2500 4 F7 N1 0 0 3 N2 F6 2600 4000 3 N11 F8 0 800 4 N11 N14 0 700 4 F1 F4 0 700 5 F1 F4 0 1400 3 F1 F2 0 750 4 F4 F1 750 1800 3 0 1050 3 F1 F4 F1 700 2100 7 F4 F1 0 2200 4 0 900 3 F1 10.86 7.62 1 0 0 261 1 1 19 A1 A1 0 4300 5 A1 A3 0 0 3 800 2400 3 F4 57 P. brunnescens 58 Pseudasthenes steinbachi 58 P. cactorum 59 Pseudocolaptes lawrencii 59 P. boissonneautii 60 Pseudoseisura gutturalis 60 P. lophotes 61 Schizoeaca griseomurina 61 S. fuliginosa 62 Schizoeaca perijana 62 S. coryi 63 Sclerurus caudacutus 63 S. guatemalensis 64 Sclerurus rufigularis 64 S. mexicanus 65 Sclerurus scansor 65 S. albigularis 66 Simoxenops ucayalae 66 S. striatus 67 Sittasomus griseicapillus 67 Deconychura longicauda 68 Sylviorthorhynchus desmursii 68 Leptasthenura yanacensis 69 Synallaxis albilora 69 S. maranonica 70 Synallaxis brachyura 70 S. subpudica 71 Synallaxis cabanisi 71 S. moesta 72 Synallaxis candei 3.38 4.06 0 0 1103 0 1 1 F4 950 2500 11 N2 800 3000 3 N2 0 2400 5 F4 1800 3100 4 1400 3400 7 0 1000 3 0 800 4 2750 3200 4 3000 4300 8 N10 3000 3400 1 N10 2800 4100 3 F1 0 1100 8 0 1250 5 F4 1.75 0 0 0 N1 F7 0.16 0 104 1 5.18 0 0 192 152 1 1 F1 11.68 2.90 2.10 1 0 0 0 760 0 1 1 1 F8 N10 N10 3.11 F5 F5 F4 F1 0 900 4 F1 F4 0 1800 7 F1 F4 0 1250 4 F4 F1 700 2100 5 F1 F3 F1 11.64 1 0 1 F1 F2 F1 10.50 3.76 3.94 3.08 2.27 0 0 0 0 0 1158 2454 0 342 1414 0 1 0 1 1 20 0 1000 4 650 800 4 0 1550 19 0 1400 6 F9 F15 0 1000 3 F6 N3 3700 4500 3 F8 F15 0 600 5 F8 F15 0 1500 3 N14 N11 0 2000 6 F4 F15 1200 3200 1 F1 F15 0 1400 4 F3 F1 0 1200 4 0 1000 4 F7 72 S. erythrothorax 73 Synallaxis castanea 73 S. unirufa 74 Synallaxis cherriei 74 S. rutilans 75 Synallaxis courseni 75 S. azarae 76 Synallaxis hypospodia 76 S. spixi 77 Synallaxis scutata 77 S. cinerascens 78 Synallaxis stictothorax 78 S. zimmeri 79 Syndactyla dimidiatum 79 S. rufosuperciliata 80 Syndactyla ruficollis 80 S. subalaris 81 Tarphonomus certhioides 81 T. harterti 82 Thripadectes melanorhynchus 82 T. rufobrunneus 83 Thripadectes scrutator 83 T. flammulatus 84 Thripophaga fusciceps 84 T. cherriei 85 Upucerthia albigula 85 U. dumetaria 86 Upucerthia validirostris 86 U. jelskii 87 Xenerpestes singularis 4.06 3.10 0 1 318 0 1 1 F15 N14 0 1000 4 F4 F15 1300 2900 3 F4 F5 1700 3300 8 F1 F15 0 1050 3 0 900 10 F1 1.66 2.85 2.17 2.27 1.74 2.30 1.18 3.53 1.81 3.56 0 0 0 0 0 0 0 0 0 0 7 0 0 62 0 0 61 882 91 987 1 1 1 0 1 1 0 1 1 1 F4 F15 2700 3500 5 F4 F15 1250 3100 13 N6 N14 0 700 6 N14 N11 0 2200 5 F7 F8 0 1500 4 F1 F4 0 2000 3 N1 0 400 8 N2 1900 3000 3 F8 F1 0 1200 3 F1 F4 0 2600 7 F4 400 2900 5 F4 1000 2300 7 N1 0 1300 3 N2 1400 2900 3 F4 900 1750 3 F4 1200 2500 3 F4 F5 2100 3500 5 F4 F5 1400 3250 4 F2 F3 0 0 8 0 0 3 3050 3700 4 0 4000 4 2700 5000 4 3250 4600 3 1050 1700 3 F3 2.89 0 30 1 N2 N2 0.45 0 0 1 N9 N9 2.92 0 117 0 21 N1 F4 N2 87 X. minlosi 88 Xenops rutilans 88 X. tenuirostris 89 Xiphocolaptes major 89 X. promeropirhynchus 90 Xiphorhynchus eytoni 90 X. guttatus 91 Xiphorhynchus lachrymosus 91 X. flavigaster 92 Xiphorhynchus pardalotus 92 X. ocellatus 93 Xiphorhynchus spixii 93 X. elegans 94 Xiphorhynchus triangularis 94 X. erythropygius 5.87 1.22 3.02 2.66 2.54 4.61 4.15 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 F4 F1 0 1000 2 F4 F1 0 2400 8 F3 F2 0 0 4 F7 F8 0 1800 4 F4 F11 700 3100 8 F1 0 0 2 F1 F2 0 1100 5 F1 F14 0 1200 4 F1 F4 0 1500 5 F1 F4 0 1800 6 F1 F4 0 1600 5 F1 0 600 3 F1 0 500 4 F4 1100 2400 7 900 2200 4 F4 F1 Species habitat assignments and elevation range limits are from Stotz et al. (1996), supplemented with additional sources for species representing recent taxonomic revisions. Habitat codes are: forest [F1–F15], scrub/grassland [N1–N14] and aquatic (including lakes/rivers/coasts [A1–A12]). 22 Table S2 Rates of secondary sympatry across Furnariidae under alternative β-niche classifications and distance cut-offs. Constant-Rate Data N All pairs β-niche Conserved β-niche Conserved2 94 <1000km 71 <500km 65 <250km 61 <125km 55 79 61 S 0.07 (0.04,0.11) 0.09 (0.05,0.14) 0.07 (0.04,0.13) 0.1 (0.06,0.16) 0.11 (0.07,0.18) 0.12 (0.08,0.2) 0.14 (0.09,0.22) W 14.52 (9.23,22.85) 11.59 (7.25,18.51) 13.83 (7.81,24.51) 9.84 (6.14,15.75) 8.97 (5.59,14.39) 8.09 (5.03,13.01) 7.21 (4.47,11.64) Rate-Switch AIC 83.5 68.2 49.6 62.5 59.3 55.8 51.9 S1 0.06 (0.03,0.11) 0.06 (0.03,0.12) 0 (0,62182.17) 0.07 (0.04,0.13) 0.08 (0.04,0.15) 0.08 (0.04,0.15) 0.09 (0.05,0.18) W1 16.16 (8.92,29.29) 16.35 (8.58,31.17) 10.16x10-2 (0,6.42x10-10) 14.43 (7.54,27.64) 12.8 (6.69,24.49) 12.54 (6.52,24.14) 10.99 (5.71,21.17) S2 0.1 (0.03,0.36) 0.33 (0.12,0.9) 0.14 (0.08,0.26) 0.54 (0.18,1.61) 0.52 (0.17,1.6) 4.57 (0.55,37.97) 4.56 (0.14,151.75) W2 9.59 (2.75,33.39) 3.03 (1.11,8.26) 7.05 (3.89,12.76) 1.87 (0.62,5.6) 1.93 (0.63,5.96) 0.22 (0.03,1.82) 0.22 (0.01,7.29) Time-Variable ΔW TS Tmax AIC P 7.89 4.6 7.7 85.1 0.55 8.56 4.6 6.1 66.1 0.04 6.79 1.7 4.6 48.3 0.07 7.97 4.6 5.4 58.9 0.02 7.04 4.6 5.4 56.7 0.03 7.87 5.1 5.2 48.9 <0.01 6.99 4.6 4.6 45.2 <0.01 S0 0.05 (0.02,0.13) 0.04 (0.01,0.09) 0.03 (0.01,0.09) 0.04 (0.01,0.11) 0.05 (0.02,0.1) 0.03 (0.01,0.14) 0.03 (0,0.18) b 0.08 (-0.16,0.32) 0.31 (0.06,0.55) 0.31 (0.03,0.58) 0.35 (0.04,0.65) 0.33 (0.1,0.56) 0.58 (0.05,1.11) 0.67 (0,1.34) AIC P 85.1 0.54 65.4 0.03 47.8 0.05 59.1 0.02 56.6 0.03 50.8 0.01 46.5 0.01 Rates and waiting times (95% CI) were estimated across all furnariid sister pairs (‘All pairs’), across those sister pairs occupying the same β-niche defined using either coarse (β-niche conserved: forest, scrub/grassland, aquatic) or fine (β-niche conserved2: F1–F15, N1–N14, A1–A12) habitat classifications given in Stotz et al. (1996), and for sisters species separated by different distance cut-offs. N = number of sister pairs; S = rate of secondary sympatry per sister pair per million years (Myr); W = expected waiting time to secondary sympatry (Myr); S1, S2, W1 and W2 = the rates and waiting times to secondary sympatry estimated under the RateSwitch model before and after the break point (TS), respectively and ΔW is the estimated waiting time to sympatry arising from biotic interactions; Tmax = max break point age considered in the RateSwitch model, b = rate of change in transition rate with time since divergence estimated under the Time-Variable model; P = significance calculated from likelihood ratio tests comparing the Rate-Switch and Time-Variable models to a Constant-Rate model, respectively. 23 Table S3 Trait-dependent models of secondary sympatry fit to observed α-niche divergence (z) across species pairs (N = 48), showing the effects of more stringent β-niche classification. Variable Bill.PC1 Bill.PC2 Bill.PC3 Bill Euclidian Morph.PC1 Morph.PC2 Morph.PC3 Morph.PC4 Morph.PC5 Morphology Euclidian Proportional Change 2.87 (0.08,5.75) 6.51 (-1.58,15.27) 0.08 (-2.22,2.44) 4.25 (-0.13,8.83) 2.6 (-0.26,5.53) 5.16 (-1.39,12.15) 2.41 (0.18,4.68) -0.53 (-2.75,1.74) 3.36 (-2.38,9.44) 4.1 (-0.29,8.69) S (z=min(z)) W (z=min(z)) S (z=max(z)) W (z=max(z)) 0.05 (0.13,0.02) 0.05 (0.15,0.02) 0.1 (0.28,0.04) 0.04 (0.13,0.01) 0.05 (0.14,0.02) 0.06 (0.15,0.02) 0.05 (0.13,0.02) 0.12 (0.3,0.05) 0.06 (0.19,0.02) 0.04 (0.13,0.01) 21.25 (7.65,59.03) 18.73 (6.51,53.83) 9.98 (3.62,27.51) 23.47 (7.75,71.11) 19.31 (7.06,52.78) 17.2 (6.53,45.31) 20.01 (7.55,53.03) 8.21 (3.34,20.18) 17.86 (5.29,60.28) 23.18 (7.55,71.13) 0.8 (6.44,0.1) 29.36 (34589.41,0.02) 0.11 (0.54,0.02) 2.74 (88.86,0.08) 0.67 (5.79,0.08) 8.94 (2722.91,0.03) 0.54 (2.54,0.11) 0.07 (0.38,0.01) 1.53 (166.98,0.01) 2.41 (77.5,0.07) 1.25 (0.16,10.04) 0.03 (0,40.12) 9.22 (1.86,45.77) 0.36 (0.01,11.8) 1.49 (0.17,12.78) 0.11 (0,34.06) 1.86 (0.39,8.77) 13.91 (2.62,73.99) 0.65 (0.01,71.59) 0.42 (0.01,13.37) P 0.02 0.07 0.95 0.02 0.04 0.08 0.03 0.64 0.13 0.02 Species β-niches were based on habitat classifications given in Stotz et al. (1996) and elevational range overlap (see Appendix 1). To aid comparison, values of z across species were standardised from 0 (minimum divergence) to 100 (maximum divergence). Proportional Change indicates the % change in S per 1% increase in z. S and W = expected rates (per sister pair per million years [Myr]) and waiting times to secondary sympatry (Myr), respectively, corresponding to the minimum (z = 0) and maximum (z = 100) observed levels of α-niche divergence; P = significance calculated from a likelihood ratio test comparing the Trait-Dependent model to a Constant-Rate model. Models were only fitted to those sister pairs with geographical ranges less than 250 km apart. 24 Table S4 Trait-dependent models of secondary sympatry fit to observed α-niche divergence (z) across species pairs, showing the effects of varying distance cut-offs. Distance cut-off (km) 125 125 125 125 125 125 125 125 125 125 250 250 250 250 250 250 250 250 250 250 500 500 500 500 500 500 500 500 500 Variable Bill. PC1 Bill. PC2 Bill. PC3 Bill Euclidian Morph. PC1 Morph. PC2 Morph. PC3 Morph. PC4 Morph. PC5 Morph Euclidian Bill. PC1 Bill. PC2 Bill. PC3 Bill Euclidian Morph. PC1 Morph. PC2 Morph. PC3 Morph. PC4 Morph. PC5 Morph Euclidian Bill. PC1 Bill. PC2 Bill. PC3 Bill Euclidian Morph. PC1 Morph. PC2 Morph. PC3 Morph. PC4 Morph. Proportional Change S (z=min(z)) W (z=min(z)) S (z=max(z)) W (z=max(z)) 2.63 (0.17,5.15) 2.9 (-3.58,9.82) 1.07 (-0.96,3.15) 3.91 (0.02,7.95) 2.32 (-0.14,4.85) 2.55 (-3.29,8.74) 0.88 (-0.82,2.6) 0.18 (-1.49,1.88) 1.3 (-1.45,4.12) 3.64 (-0.13,7.55) 2.82 (0.38,5.32) 3.37 (-3.16,10.35) 0.49 (-1.4,2.42) 4.15 (0.28,8.18) 2.51 (0.06,5.01) 2.93 (-2.94,9.16) 1.06 (-0.63,2.79) -0.17 (-1.92,1.62) 1.56 (-1.29,4.48) 3.91 (0.15,7.81) 3.08 (0.61,5.61) 4.49 (-1.94,11.33) -0.33 (-2.24,1.62) 4.46 (0.52,8.54) 2.78 (0.3,5.32) 3.9 (-1.82,9.96) 1.16 (-0.56,2.92) -0.81 (-2.67,1.08) 0.07 (0.16,0.03) 0.1 (0.23,0.05) 0.1 (0.23,0.04) 0.07 (0.16,0.03) 0.08 (0.18,0.04) 0.11 (0.23,0.05) 0.11 (0.22,0.05) 0.13 (0.28,0.06) 0.11 (0.23,0.05) 0.07 (0.17,0.03) 0.06 (0.14,0.03) 0.09 (0.2,0.04) 0.11 (0.23,0.05) 0.06 (0.14,0.02) 0.07 (0.15,0.03) 0.09 (0.2,0.04) 0.09 (0.19,0.04) 0.13 (0.27,0.06) 0.09 (0.19,0.04) 0.06 (0.14,0.02) 0.05 (0.12,0.02) 0.07 (0.16,0.03) 0.12 (0.25,0.06) 0.05 (0.12,0.02) 0.06 (0.13,0.03) 0.08 (0.16,0.04) 0.08 (0.17,0.04) 0.14 (0.29,0.07) 13.53 (6.07,30.17) 9.7 (4.29,21.89) 10 (4.42,22.64) 14.91 (6.12,36.35) 12.43 (5.66,27.3) 9.46 (4.35,20.57) 9.29 (4.52,19.1) 7.66 (3.62,16.19) 9.42 (4.39,20.23) 14.75 (5.95,36.55) 15.72 (7.1,34.81) 11.35 (5.02,25.68) 9.49 (4.27,21.08) 17.43 (7.17,42.34) 14.48 (6.61,31.7) 10.99 (5.06,23.89) 10.93 (5.35,22.32) 7.66 (3.65,16.08) 11.11 (5.13,24.07) 17.34 (7.02,42.8) 18.2 (8.24,40.23) 13.72 (6.18,30.45) 8.13 (3.95,16.75) 20.3 (8.32,49.53) 16.84 (7.69,36.89) 13.18 (6.2,28.02) 12.44 (6.05,25.56) 7.04 (3.5,14.15) 0.99 (6.6,0.15) 1.8 (643.55,0.01) 0.29 (1.24,0.07) 3.11 (72.07,0.13) 0.8 (5.45,0.12) 1.31 (258.71,0.01) 0.26 (0.91,0.07) 0.16 (0.52,0.05) 0.39 (3.53,0.04) 2.41 (48.92,0.12) 1.03 (6.74,0.16) 2.42 (884.12,0.01) 0.17 (0.66,0.04) 3.36 (76.17,0.15) 0.82 (5.44,0.12) 1.64 (326.42,0.01) 0.26 (0.92,0.08) 0.11 (0.4,0.03) 0.42 (4.17,0.04) 2.66 (53.15,0.13) 1.14 (7.69,0.17) 5.87 (1822.32,0.02) 0.09 (0.38,0.02) 3.85 (90.67,0.16) 0.92 (6.31,0.13) 3.5 (583.37,0.02) 0.26 (0.91,0.07) 0.06 (0.27,0.01) 1.01 (0.15,6.72) 0.56 (0,198.57) 3.44 (0.8,14.7) 0.32 (0.01,7.46) 1.25 (0.18,8.5) 0.76 (0,150.23) 3.89 (1.1,13.68) 6.38 (1.91,21.3) 2.6 (0.28,23.82) 0.41 (0.02,8.41) 0.97 (0.15,6.35) 0.41 (0,150.48) 5.79 (1.51,22.27) 0.3 (0.01,6.74) 1.22 (0.18,8.09) 0.61 (0,121.74) 3.79 (1.08,13.28) 9.05 (2.47,33.13) 2.37 (0.24,23.41) 0.38 (0.02,7.5) 0.87 (0.13,5.87) 0.17 (0,52.84) 11.35 (2.61,49.27) 0.26 (0.01,6.11) 1.09 (0.16,7.45) 0.29 (0,47.75) 3.91 (1.1,13.94) 15.88 (3.7,68.17) 1.44 0.08 12.1 0.35 2.89 25 P 0.02 0.24 0.31 0.02 0.04 0.25 0.33 0.83 0.34 0.03 0.01 0.19 0.62 0.01 0.02 0.2 0.24 0.85 0.26 0.02 0.01 0.11 0.73 0.01 0.01 0.11 0.21 0.38 0.28 500 1000 1000 1000 1000 1000 1000 1000 1000 1000 1000 None None None None None None None None None None PC5 (-1.27,4.23) (0.18,0.04) (5.65,25.93) (3.03,0.04) (0.33,25.28) Morph Euclidian Bill. PC1 Bill. PC2 Bill. PC3 Bill Euclidian Morph. PC1 Morph. PC2 Morph. PC3 Morph. PC4 Morph. PC5 Morph Euclidian Bill. PC1 Bill. PC2 Bill .PC3 Bill Euclidian Morph .PC1 Morph. PC2 Morph. PC3 Morph. PC4 Morph. PC5 Morph Euclidian 4.26 (0.46,8.21) 2.83 (0.49,5.22) 3.58 (-2.7,10.26) -0.71 (-2.68,1.3) 3.99 (0.25,7.88) 2.46 (0.14,4.84) 3.21 (-2.34,9.08) 1.2 (-0.56,2.98) -1.11 (-3.01,0.83) 1.57 (-1.13,4.34) 3.75 (0.09,7.53) 1.88 (0.17,3.62) 2.54 (-3.13,8.53) -0.64 (-2.62,1.38) 2.57 (-0.15,5.35) 1.66 (-0.05,3.41) 2.35 (-2.4,7.33) 1.27 (-0.5,3.07) -1.2 (-3.12,0.76) 1.81 (-0.81,4.49) 2.43 (-0.12,5.04) 0.05 (0.12,0.02) 0.05 (0.11,0.02) 0.07 (0.16,0.03) 0.13 (0.26,0.06) 0.05 (0.11,0.02) 0.06 (0.12,0.03) 0.07 (0.15,0.03) 0.07 (0.15,0.04) 0.14 (0.29,0.07) 0.07 (0.16,0.03) 0.05 (0.12,0.02) 0.05 (0.11,0.02) 0.07 (0.14,0.03) 0.1 (0.22,0.05) 0.05 (0.11,0.02) 0.06 (0.11,0.03) 0.07 (0.13,0.03) 0.06 (0.13,0.03) 0.13 (0.25,0.06) 0.06 (0.13,0.03) 0.05 (0.11,0.02) 20.35 (8.24,50.29) 19.56 (8.85,43.23) 14.07 (6.35,31.19) 7.92 (3.79,16.54) 21.36 (8.72,52.3) 17.81 (8.17,38.84) 13.72 (6.47,29.09) 13.79 (6.66,28.54) 6.98 (3.44,14.18) 13.6 (6.37,29) 21.12 (8.52,52.36) 19.46 (9.35,40.47) 15.3 (7.14,32.81) 9.56 (4.58,19.95) 20.33 (9.13,45.27) 18.1 (8.86,36.98) 15.12 (7.44,30.73) 16.56 (7.97,34.41) 7.98 (3.92,16.25) 16.64 (7.95,34.85) 20.2 (9.14,44.68) 3.19 (65.61,0.16) 0.83 (4.89,0.14) 2.39 (673.75,0.01) 0.06 (0.28,0.01) 2.35 (46.21,0.12) 0.64 (3.77,0.11) 1.72 (251.7,0.01) 0.24 (0.86,0.07) 0.05 (0.21,0.01) 0.35 (3.03,0.04) 1.87 (33.36,0.11) 0.33 (1.15,0.1) 0.8 (133.46,0) 0.06 (0.25,0.01) 0.62 (5.1,0.08) 0.29 (1.03,0.08) 0.67 (47.49,0.01) 0.21 (0.77,0.06) 0.04 (0.17,0.01) 0.36 (2.92,0.04) 0.54 (3.86,0.08) 0.31 (0.02,6.43) 1.2 (0.2,7.06) 0.42 (0,117.8) 16.14 (3.57,72.97) 0.43 (0.02,8.36) 1.56 (0.27,9.19) 0.58 (0,84.98) 4.19 (1.16,15.19) 21.32 (4.81,94.47) 2.86 (0.33,24.7) 0.53 (0.03,9.52) 3.02 (0.87,10.51) 1.25 (0.01,209.09) 18.18 (3.99,82.87) 1.61 (0.2,13.25) 3.47 (0.97,12.37) 1.49 (0.02,105.25) 4.7 (1.29,17.11) 26.7 (5.93,120.25) 2.77 (0.34,22.43) 1.84 (0.26,13.02) 0.01 0.01 0.14 0.47 0.01 0.02 0.14 0.21 0.24 0.24 0.02 0.03 0.18 0.52 0.04 0.06 0.18 0.18 0.21 0.16 0.04 To aid comparison, values of z across species were standardised from 0 (minimum divergence) to 100 (maximum divergence). Proportional Change indicates the % change in S per 1% increase in z. S and W = expected rates (per sister pair per million years (Myr)) and waiting times to secondary sympatry (Myr), respectively, corresponding to the minimum (z = 0) and maximum (z = 100) observed levels of α-niche divergence; P = significance calculated from a likelihood ratio test comparing the TraitDependent model to a Constant-Rate model. 26 Table S5 Results of the State-Reversible model used to estimate the rate at which species pairs return to allopatry following the attainment of sympatry. Constant-Rate Data N All pairs 94 β-nicheConserved1 79 β-nicheConserved2 61 s 0.07 (0.04,0.11) 0.09 (0.05,0.14) 0.07 (0.04,0.13) AIC 83.47 68.23 49.61 State-Reversible s 0.07 (0.04,0.11) 0.09 (0.05,0.14) 0.07 (0.04,0.13) a 0 (0,4.56x10-4) 0 (0,5.17x10-5) 0 (0,3.54x10-4) 0.1 (0.06,0.16) 0.11 (0.07,0.18) 0.12 (0.08,0.2) 0.14 (0.09,0.22) 0 (0,2.30x10-4) 0 (0,2.01x10-13) 0 (0,1.27x10-4) 0 (0,1.03x10-5) AIC P 85.49 1 70.25 1 51.63 1 64.55 1 61.34 1 57.84 1 53.94 1 Distance cut-off <1000km 71 <500km 65 <250km 61 <125km 55 0.1 (0.06,0.16) 0.11 (0.07,0.18) 0.12 (0.08,0.2) 0.14 (0.09,0.22) 62.52 59.34 55.81 51.92 Rates and waiting times (95% CI) were estimated across all furnariid sister pairs (‘All pairs’), across those sister pairs occupying the same β-niche defined using either coarse (β-niche conserved1: forest, scrub/grassland, aquatic) or fine (β-niche conserved2: F1–F15, N1–N14, A1–A12) habitat classifications given in Stotz et al. (1996), and for sisters species separated by different distance cutoffs. N = number of sister pairs; S = rate of secondary sympatry per sister pair per million years (Myr); a = rate of return to allopatry per sister pair per million years (Myr); P = significance calculated from likelihood ratio tests comparing the State-Reversible model to the Constant-Rate model. 27 Table S6 Morphological trait loadings for principle components of bill dimensions. Trait PC1 PC2 PC3 Bill length 0.908 0.419 -0.014 Bill width 0.743 -0.546 -0.387 Bill depth 0.808 -0.499 0.313 Results are from a phylogenetic Principal Components (PC) analysis (Revell et al. 2009) based on the covariance matrix following log-transformation of species trait values. Table S7 Morphological trait loadings for principle components of body dimensions. Trait PC1 PC2 PC3 PC4 PC5 Bill length -0.865 0.498 0.058 -0.014 -0.025 Bill width -0.773 -0.466 0.135 -0.407 -0.042 Bill depth -0.830 -0.410 0.232 0.260 0.147 Tarsus -0.769 -0.106 -0.544 -0.041 0.317 Wing -0.758 -0.335 -0.285 0.171 -0.451 Results are from a phylogenetic Principal Components (PC) analysis (Revell et al. 2009) based on the covariance matrix following log-transformation of species trait values. 28 Table S8 Clade-wide shifts in rates of secondary sympatry. Constant-Rate Data N All pairs 94 β-niche Conserved 79 S W 0.07 14.52 (0.04,0.11) (9.23,22.85) 0.09 11.59 (0.05,0.14) (7.25,18.51) Clade-Rate-Switch AIC 83.47 68.23 Clade-Time-Variable S1 W1 S2 W2 0.78 1.27 0.06 16.16 (0.11,5.5) (0.18,8.93) (0.04,0.1) (9.75,26.78) 3.96 0.25 0.07 14.3 (0,10992.07) (0,701.79) (0.04,0.12) (8.29,24.67) Ta 10 8 AIC 83.99 64.59 P 0.22 0.02 S0 b 0.14 -0.08 (0.02,1.3) (-0.32,0.16) 1.29 -0.29 (0.14,11.62) (-0.54,-0.05) AIC P 85.1 0.54 65.44 0.03 Rates and waiting times (95% CI) were estimated across all furnariid sister pairs (‘All pairs’) and across those sister pairs occupying the same β-niche (βniche conserved: forest, scrub/grassland, aquatic) using habitat classifications given in Stotz et al. (1996). N = number of sister pairs; S = rate of secondary sympatry per sister pair per million years (Myr); W = expected waiting time to secondary sympatry (Myr); S1, S2, W1 and W2 = the rates and waiting times to secondary sympatry estimated under the Clade-Rate-Switch model before and after the breakpoint (Ta), respectively; b = rate of change in transition rate through time estimated under the CladeTimeVariable model; P = significance calculated from likelihood ratio tests comparing the Clade-Rate-Switch and Clade-Time-Variable models to a Constant-Rate model, respectively. 29