The Appendicular Skeleton
advertisement

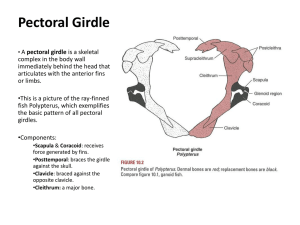
The Appendicular Skeleton Chapter 9 Introduction • Transitions from water to land and from land to air have allowed for a great diversity of forms within the appendicular skeleton. • As with many other designs, form closely follows function. • In aquatic animals fins serve different purposes – Some maneuver, some provide lift, and some are lost or modified • Not all birds use the air the same way, and some don’t fly at all. • Tetrapods use land differently – Some crawl, some run, some dig and some climb. Basic Components • The appendicular skeleton is composed of paired fins, or limbs, and their girdles. • The anterior girdle is the pectoral girdle, or shoulder. • The posterior girdle is the pelvic girdle or hip. Fins • Fins are membranous or webbed processes internally strengthened by radiating, thin fin rays. – They are formed from epidermis, but sink inward to the dermis. – In elasmobranchs rays are slender keratinized rods – Fins rays in bony fishes are usually ossified tiny elements that strengthen the fin web. • The proximal part of the fin is supported by pterygiophores of two types: – The enlarged basals, within the proximal fin – The slender radials that extend into the middle portion of the fin. • Fins occur singly, except for the paired pectoral and pelvic fins. • The basal pterygiophores of these projecting fins articulate with and are braced with girdles Limbs • Limbs, or chiridium, are muscular appendages with well defined joints bearing digits. • In tetrapods the fore and hind limbs are built on the same pattern, with three regions. – The distal autopodium consisting of the wrist and ankle • The manus, hand, and pes, foot, are supported at this joint. – The middle zeugopodium with two supporting elements • The radius and ulna, or tibia and fibula respectively. – The proximal stylopodium with a single element • The humerus and femur. Origin of Paired Fins • The body of fishes are susceptible to deflections from their line of travel. – They may swing (yaw), rock on the long axis (roll), or buck forward and back (pitch) • Dorsal and lateral fins control the body by resisting deflections around the center of mass. • Pectoral fins do not produce significant lift, instead they are used for maneuvering within tight spaces • As early fishes became more active they would have experienced instability in motion. – This would favor any bony projection that resisted pitch, roll, or yaw. And led to the evolution of the paired fins. – The associated girdles stabilized the fins and provided sites for muscle attachment. • In gnathostome fishes two type of fin developed from two different arrangements of the axis. – The archipterygial and metapterygial fins Origin Theories • Gill Arch theory: – Paired fins arose from gill arches – Specifically the endoskeletal girdle arose from the gill arch and the archipterygial fin from the gill rays – Does not explain evolution of pelvic fins or girdle. • Fin-Fold Theory: – Paired fins arose from a paired, continuous set of ventrolateral folds in the body wall that were stiffened by endoskeletal pterygiophores, – Inward extensions of the basals and their fusion around the midline produced the supporting girdles. – Dermal bone, from the bony armor, later added strength to these fins. Phylogeny Agnathans • Two early vertebrates from the Cambrian possessed lateral fin folds but lacked paired fins. • Paired fins are also absent in modern hagfishes and lamprey. • Ostracoderms had unpaired medial fins and rudimentary pectoral fins – Like sharks they lack a swim bladder and they would have been denser that the surrounding water – Pectoral fins or spines, along with a flattened head shield, would have provided some some lift as they swam. Placoderms • Both pectoral and pelvic girdles were present. • The pelvic girdle appears to have a single endoskeletal element. • The more complex pectoral girdle consisted of various fused dermal elements and braced the scapolucoracoid. Chondrichthyans • Early sharks possessed pectoral and pelvic fins that were primarily stabilizers. – They consisted of basal elements and tightly packed radials. – The girdle was a single basal element. • In later sharks the paired basal elements of the pectoral and pelvic girdles became extended across the midline to fuse into a U-shaped scapulacoracoid and puboishiac bar respectively. Actinopterygians • The pectoral girdle is partly endodermal, but mostly dermal. • An air bladder, or lung is common. • Fins function mainly in close maneuvering, adjustments of body position, or breaking. • The dermal shoulder girdle forms a U-shaped collar of bone around the posterior gill chamber and braces the small scapulacoracoid. Sarcopterygians • Called lobed fin fishes in reference to the muscles and internal supportive elements that project from the body to form a fleshy dermal fin. • Among living genera fins are considerable reduced. • Ripidistans possessed pectoral and pelvic appendages that internally possess bones above the wrist/ankle that are homologous to those in modern tetrapods. Tetrapods • The first tetrapods retained or quickly changed the appendicular skeleton in correlation with locomotion. • Appearing in the sarcopterygian fish Tiktaalik, and retained by tetrapods was the loss of the attachment of the pectoral girdle to the skull – A feature that allowed increased cranial movement. • Girdles and limbs became stronger, more robust, and completely ossified. Pectoral Girdle • Tetrapods have a shoulder girdle that is structurally and functionally detached from the skull. • Leaving a dermal shoulder girdle composed of the remaining ventral elements, – The cleithrum and clavicle, and an unpaired interclavicle that joins both halves across the midline. • In modern amphibians the dermal bones are lost (salamanders) or reduced (frogs) – The endoskeletal scapulocoracoid is the prominent girdle element. • In primitive amniotes, the clavicle and interclavicle persist, but the cleithrum is absent. • In birds, the paired clavicle usually fuses with the interclavicle to form the furcula. • A single, unossified scapulocoracoid is soon replaced by two articulated, but distinct elements. – Giving rise to a scapula (dorsally) and coracoid (ventrally) Pelvic Girdle • From its first appearance the pelvic girdle is exclusively endoskeletal. • In most fishes and early tetrapods it is formed of a single element. • In modern tetrapods it has three bones: – The Illium, Ishium, and Pubis. • The attachment of the illium to the vertebral column establishes, and defines the sacral region. Manus and Pes • The autopodium has undergone extensive modifications throughout evolution. – There are several digits, beginning with the metacarpals, or metatarsals, followed by a chain of phalanges. – The digits rest on a several bones called carpals or tarsals • Its unusual to find animals with more than 5 (pentadactylous) digits – In many animals the opposite has occurred • Although the expected pattern of the manus and pes give a starting point when looking at distal limb anatomy. • The actual morphology is considerably modified by modified by fusions, elongations, eliminations, and additions of new elements. • In the hindlimb, lateral digits are lost and medial metatarsals are fused into a composite ankle. • Within amniotes, three ankle joints exist – Mesotarsal Joint is a simple hinge between, found in birds and dinosaurs. – Intratarsal Joint, line of flexion passes between the calcaneum and astralagus, found in crocodilians and thecodonts. – A Crurotarsal Joint forms between the shank and proximal tarsals, found in mammals. Onto Land • The musculature of early rhipidistians was probably too weak to supply propulsion or bear the weight of the organism for long periods of time. – Well developed axial musculature allowed for lateral undulations that propelled the body around pin-like fins. • Not until the Permian did well developed, terrestrial tetrapods appear Why Leave Water? • Different hypothesis have been put forth: – One states that early rhipidistians crawled from pool to pool as evaporation forced them from their home. • This assumes that the limb was already strong enough for overland travel – Another states that movement onto land was an attempt to avoid predation by young animals. • Movement onto land does not require long journeys, just a few steps, so limbs not need be overly strong – No one is sure what selection pressure actually led to colonization of land. • But, the fossil record and variation of modern vertebrates clearly demonstrate its success. Form and Function • Changes in the skeletal system are the results of changing demands placed on the different parts of the body. • On land the main contributors to locomotion are the limbs, not the tail. – Therefore limbs undergo extensive and significant morphological change. • In addition, the shoulder and hip establish new associations with the axial skeleton. – In tetrapods the axial column is slung from the shoulder girdle by muscles and the hip attaches directly to column. • The shoulder moves on the thorax via these muscles which lessons the wobbling of the head and neck. • The hip is firmly attached and applies strong propulsive forces to the axial column. Swimming • Swimming motion in the aquatic environment is resisted by drag. • Streamlining prevents laminar flow , reduces drag, and improves performance. • Lateral undulations that pass along the body wall move the fish through the aquatic environment • This same mode of transportation still serves most modern amphibians and reptiles. Terrestrial Locomotion • The pattern of foot contact is called a gait. • One cycle is complete after all 4 limbs have been used. • One basic gait is a diagonal sequence, oppositely opposed feet strike the ground in unison. – A trot is based on the diagonal sequence – The line between the diagonal points of contact pass through the center of mass, improving support • Another basic gait is the lateral sequence gait where feet on the same side of the body strike at the same time. – During cycles of locomotion, the center of mass remains within the supportive configuration, never at the edge. • This gait occurs in salamanders and reptilians. • Unlike the gait produced by the fins of bottom walking fishes, terrestrial gaits include significant longitudinal of the stylopodium, • This makes the limb more active in locomotion. – Contributing a pulling and pushing force against the ground. – Rotation of the sylopodium can contribute to locomotion only after the development of a rightangle elbow. Limb Placement • Early tetrapods have limbs placed laterally in a sprawled stance, establishes pivot points. • Locomotion occurs through lateral undulations around this fixed point. • In terrestrial birds, dinosaurs, and many mammals the trend has been toward cursorial (running) motion • From the characteristic sprawled position of early tetrapods, modern tetrapods have limbs drawn under the body – A change in posture that increases efficiency of limb swing – This also restricts limb movement to a single, sagittal, plane. • Early tetrapods with sprawled postures must use an overarm swing after each propulsive stroke to reestablish contact with the ground. – With legs positioned below the body, limb recovery can be accomplished efficiently using pendulum motion. • In mammals a major shift in the functional participation of the vertebral column appears. – This is a change from lateral to ventral flexion. – With limbs under the body, lateral motion contributes little to locomotion. – Loss of ribs from the posterior trunk increases the flexibility of this region in conjunction with the flexible vertebral column. • Generally , as locomotion becomes used for more sustained, efficient, and rapid transport on land, many structures become modified. – The digits move forward and more in line with the limb – Limbs were moved under the girdles that support them – Vertical flexion of the vertebral column assists in limb placement Cursorial Locomotion • Along with increasing the efficiency of limb motion, many tetrapods become specialized in rapid terrestrial motion. – Rapid locomotion is present in both predator and prey • The speed attained by a vertebrate is produced by its stride length and stride rate • Stride Length: – Increased by increasing limb length; changes in foot posture – Increase the distance through which the limbs move while they are off the ground • Stride Rate – Velocity of travel also depends on the rate at which the limbs. • Larger, more efficient muscles increase rate of limb motion • Or, lightening of the fore limb reduces mass and allows the limb to reposition quicker. Gait • The gait an animal selects depends on the rate of travel, obstructions in terrain, maneuverability, and body size. END