the singing brain - McGill University
advertisement

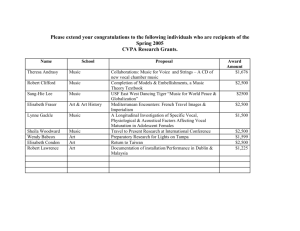
NEURAL CORRELATES OF VOCAL PITCH REGULATION IN SINGING JEAN MARY ZARATE Dept. of Neurology & Neurosurgery McGill University INTRODUCTION Precise vocal pitch regulation necessary for speech and song Vocal pitch regulation requires integration between: Stable vocal motor system Auditory feedback Interface between these two components not well-understood Used singing to find neural substrates for audio-vocal integration Elicit learned vocalizations Initiate vocalizations EXP 1: Experience-dependent neural substrates involved in vocal pitch regulation (Zarate & Zatorre, 2008) 12 non-musicians (6 ♀), 12 singers (6 ♀) HYPOTHESES: SIMPLE: basic network for singing (Perry et al., 1999) IGN: ↑ attention areas, ↓ auditory cortical activity COMP: audio-vocal integration = ACC, STG, insula? Singers: Singing tasks: singers > non-musicians Experience-dependent modulation in basic network for singing, audio-vocal integration error (cents) response magnitude (cents) 150 125 100 75 50 25 0 -25 -50 -75 -100 -125 -150 -175 300 275 250 225 200 175 150 125 100 75 50 25 0 -25 SIMPLE – PERC (SINGER ∩ NON-MUS) SIMPLE M1 NON-MUS SINGER * * b Cbl * b SMA Th STG/ INS PAC/ STG ACC 6.1 D# F G# B IGNORE x=0 C# ! D# F ! G# ! ! B C# z=0 2.5 IGN – SIMP (SINGER > NON-MUS) NON-MUS SINGER PT/ STG INCORRECT ! y = -14 STS 4.0 y = -22 2.0 COMPENSATE NON-MUS SINGER CORRECT D# F G# B C# dPMC 3.7 z = 68 2.0 SINGER > NON-MUS 300 275 250 225 200 175 150 125 100 75 50 25 0 -25 NON-MUS > SINGER response magnitude (cents) COMP – SIMP (GROUP DIFF) pSTS RCZa 4.4 x = -10 x = 50 2.4 EXP 1: KEY FINDINGS IGN: non-mus had pitch-shift responses Pitch-shift response = vocal stabilization system Training needed to suppress stabilization Audio-vocal integration: Non-mus: dPMC (sensorimotor association) Singers: RCZa, pSTS EXPERIMENT 2: Neural networks involved in voluntary and involuntary vocal pitch regulation in experienced singers (Zarate et al., submitted) 9 singers (6 ♀) SIMPLE; IGN/COMP 200c and 25c pitch shifts COMP200c = voluntary vocal pitch regulation Pitch-shift response in IGN25c = PAG? Unable to verify role of PAG due to temporal resolution limitations of fMRI FUNCTIONAL CONNECTIVITY: COMP200 EFFECTIVE CONNECTIVITY: COMP200 (vs. SIMPLE) pSTS seed EFFECTIVE CONNECTIVITY: IGN200 (vs. SIMPLE) pSTS seed EXP 1 & 2: RCZa, pSTS, anterior insula Recruited after vocal training Functionally connected to each other pSTS interacts with IPS to monitor feedback EXP 3: Training effects in non-musicians (Modulation of functional network for singing after auditory training) Better auditory skills = better vocal accuracy? Better vocal accuracy modulations in singing networks Melodies: Singing tasks: 50c & 100c melodies, simple singing Perception: micromelody discrimination (<100c interval) 110 PRE-TRAINING 110 100 percent correct (%) percent correct (%) 100 90 80 70 60 50 TRAINED CONTROL 40 * 90 * * * * 80 70 60 50 TRAINED CONTROL 40 30 30 5c 10c 15c 20c 30c 40c interval 40 * 30 20 10 0 PRE 60c 5c 10c 15c 20c 30c 40c interval SIMPLE PERFORMANCE (BEHAVIORAL SESSIONS) pitch variability (cents) POST-TRAINING POST FUNC. CONNECTIVITY (POST – PRE) right PT seed 60c EXP 3: CONCLUSIONS Short-term auditory training training effects with micromelody discrimination no training effects on vocal production no neural modulations specifically induced by training-enhanced vocal production Dissociation between perceptual and production skills? different time-courses of behavioral improvement auditory-motor training necessary Consolidated after adequate audiovocal training Short-term auditory training does not engage or consolidate network ACKNOWLEDGMENTS Robert J. Zatorre Advisory Committee: D. Louis Collins Alan Evans David Ostry Université de Montréal / BRAMS / CIRMMT: James Bergstra Douglas Eck Sean Wood McGill / MNI: Pierre Ahad Patrick Bermudez Marc Bouffard André Cormier Karine Delhommeau Michael Ferreira Nicholas Foster Talya Grumberg Funding: Canadian Institutes of Health Research (CIHR) Eileen Peters McGill Majors Fellowship Centre for Interdisciplinary Research in Music Media and Technology (CIRMMT) New York: Henry McDonagh III Members of the Z-Lab FUTURE DIRECTIONS A-V network specific to vocal pitch? MEG, EEG/ERP: pitch-shift response Auditory training vocal accuracy manipulate other features (e.g., formants) training effects: foreign language students more testing sessions of vocal production longer auditory training Similar network with other perturbations? somatosensory feedback EXP 1: Audio-vocal integration SINGERS & NON-MUS EXP 2: Voluntary/involuntary vocal pitch regulation SINGERS EXP 3: Vocal pitch regulation after auditory training NON-MUS SIMPLE: Sing back single note PITCH-SHIFTED TASKS: ignore/compensate for ± 200c-shift Deviation from target note (cents) Deviation from target n 125 100 100 75 75 TARGET OWN FEEDBACK 50 50 25 25 0 0 -25 -25 -50 -50 -75 -75 -100-100 -1.0 -125 -0.5 0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 IGNORE TIME(s) -150 -175 -200 -225 -250 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 2.0 2.5 3.0 3.5 4.0 TIME(s) 250 225 Deviation from target note (cents) 200 175 150 125 100 75 50 25 0 -25 -0.5 0.0 0.5 1.0 1.5 -50 -75 -100 -125 -150 -175 -200 -225 -250 TIME(s) TARGET OWN FEEDBACK COMPENSATE error (cents) 150 125 100 75 50 25 0 -25 -50 -75 -100 -125 -150 -175 SIMPLE NON-MUS SINGER D# F * * b * b G# B C# SIMPLE – PERC (SINGER ∩ NON-MUS) M1 STG/ INS SMA Cbl Th PAC/ STG ACC 6.1 2.5 x=0 y = -14 z=0 response magnitude (cents) 300 275 250 225 200 175 150 125 100 75 50 25 0 -25 IGNORE NON-MUS SINGER INCORRECT ! ! D# F ! G# ! ! B C# IPS SMG 4.2 2.0 y = -50 SINGER > NON-MUS CONJUNCTION IGNORE - SIMPLE STS PT/ STG 4.0 2.0 y = -22 response magnitude (cents) 300 275 250 225 200 175 150 125 100 75 50 25 0 -25 COMPENSATE NON-MUS SINGER CORRECT D# F G# B C# COMPENSATE – SIMPLE ACC IPS 3.9 SMG 2.0 y = -44 dPMC 3.7 z = 68 2.0 SINGER > NON-MUS x=4 NON-MUS > SINGER CONJUNCTION rACC x = -10 pSTS 4.4 2.4 x = 50 EXP 1: Experience-dependent neural substrates involved in vocal pitch regulation (Zarate & Zatorre, 2008) Behavioral tasks: SIMPLE, IGN: singers > non-mus COMP: both groups successful Programmed to stabilize systems against disturbances Training needed to suppress stabilization mechanisms fMRI results SIMPLE: singers ≈ non-musicians COMP/IGN: ↑ auditory activity in singers Audio-vocal integration: Non-mus: dPMC Singers: rACC, pSTS Deviation from target note (cents) Deviation from target note (cents) 200 175 150 IGNORE 125 100 100 75 75 TARGET OWN FEEDBACK 50 50 25 25 0 0 -25 -25 -50 -50 -75 -75 -100 -100 -1.0 -125 -0.5 0.0 0.5 1.0 pitch-shift 1.5 2.0 response TIME(s) 2.5 3.0 3.5 4.0 2.5 3.0 3.5 4.0 -150 -175 -200 -225 -250 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0 TIME(s) Responses to long pitch shifts (>500ms): – Early: ~100-150ms, more automatic – Late: ~300ms, may be subject to voluntary control EXPERIMENT 2: Neural networks involved in voluntary and involuntary vocal pitch regulation in experienced singers (Zarate et al., submitted) 9 singers (6 ♀) SIMPLE; IGN/COMP 200c and 25c pitch shifts HYPOTHESES: Resp. magnitude: COMP200c > IGN 200c Singers cannot suppress pitch-shift responses to small shifts: COMP25c = IGN25c IGN/COMP200c networks similar to exp1 PAG pitch-shift response in IGN/COMP25c? 200 * IGN 200c INCORRECT COMP 200c CORRECT 0 response magnitude (cents) 150 125 100 75 50 25 ! IGN 25c INCORRECT COMP 25c CORRECT -25 IGN 25c COMP 200c IGN 25c INCORRECT COMP 25c CORRECT IGN 200c IGN 25c COMP 200c -75 -100 -125 -150 COMP 25c -200 -25 150 140 130 120 110 FULL VOLUNTARY CORRECTION (COMP) 100 FULL INVOLUNTARY CORRECTION (IGN) 90 80 70 60 50 40 30 20 10 0 IGN 200c IGN 25c COMP 200c IGN 200c INCORRECT COMP 200c CORRECT ! # # * COMP 25c COMP 25c + -50 -175 0 percent response magnitude (%) response magnitude (cents) 175 IGN 200c * EXP 2: CONCLUSIONS Pitch-shift responses to IGN25c under less voluntary control than IGN200c Role of PAG in pitch-shift response: not verified part of stabilization system occurs in milliseconds, fMRI temporal resolution in seconds MEG, EEG/ERP: temporal interaction during A-V integration Voluntary vocal corrections: same network for different magnitudes: rACC, pSTS, anterior insula functionally connected to each other pSTS interacts with IPS to monitor shifted feedback EXPERIMENT 3: Modulation of functional network for singing after auditory training (Zarate et al., in prep) HYPOTHESES: Auditory training with pure tones ↑ micromelody discrimination (pure- and vocal-tone) ↑ vocal accuracy Melodic singing requires audio-vocal integration: similar regions seen in Exp 1, 2 auditory working memory (e.g., inf. frontal) Modulation of regions after training: singing network audio-vocal integration • Perception: – 2 micromelodies: same/different? – Trained/tested with micromelodies (pure & vocal tones) • Production: simple singing & 5-note melodies – Middle note ≈ 250 Hz – Intervals: 50 and 100 cents EXP 3: ORDER OF TASKS beh pre Trained 9 subj (6 ♀) Production: Simple Melodies Perception: Micromelody discrimination Control 10 subj (6 ♀) Production: Simple Melodies Perception: Micromelody discrimination fMRI pre Production: Simple Melodies TRAINING (2 weeks) YES NO fMRI post Production: Simple Melodies beh post Perception: Micromelody discrimination Production: Simple Melodies Perception: Micromelody discrimination Production: Simple Melodies SIMPLE – PERC (PRE) SMA PAC / STG / PT INS sensorimotor (mouth) ACC 7.8 2.5 x=2 y = -16 z = 10 MEL(50+100) – SIMPLE (PRE) PT/STG 6.6 2.6 y = -12 z=4 POST – PRE SCANNER PARAMETERS Exp 1 & 3: 1.5 Tesla TR = 10s, TE = 85ms voxel = 5 mm3 25 slices (whole head) Matrix: 64x64 Exp 2: 3 Tesla, cardiac gating TR = 10.3s, TE = 60ms voxel = 3.5 mm3 40 slices (whole head) Matrix: 64x64 DUAL-STREAM MODEL OF AUDITORY PROCESSING Rauschecker/Tian 2000: Ventral: “what” – auditory object info Dorsal: “where” – auditory spatial info Belin/Zatorre 2000: Dorsal = “how” Warren et al. 2005: Dorsal = “do” Updated model: Dorsal = how / do SINGING NETWORKS IN OTHER STUDIES Hickok et al. 2003: covert speech vs. covert humming Schultz et al. 2005: voiced vs. whispered speech Toyomura et al. 2007: COMP EXP 1: PUTAMINAL ACTIVITY IGNORE - SIMPLE COMP - SIMPLE Put pSTS Put 4.0 4.4 2.0 z = 10 2.4 z=2