Inclusions - Ставропольский государственный медицинский
advertisement

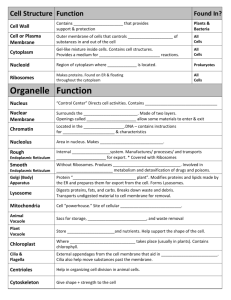
MINISTRY OF HEALTH OF THE RUSSIAN FEDERATION THE STATE BUDGET EDUCATIONAL ESTABLISHMENT OF HIGHER PROFESSIONAL EDUCATION “STAVROPOL STATE MEDICAL UNIVERSITY” Biology Department Makarenko E.N., Erina N.V., Kopteva T.S., Nikolenko T.S. Methodical recommendations for students of the first course of the English-speaking Medium of Medical University Stavropol, 2015 -2- Государственное бюджетное образовательное учреждение высшего профессионального образования «Ставропольский государственный медицинский университет» Министерства здравоохранения Российской Федерации Кафедра биологии Макаренко Э.Н., Ерина Н.В., Коптева Т.С., Николенко Т.С. КЛЕТКА КАК СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ЕДИНИЦА ЖИЗНИ Методические рекомендации для студентов англоязычного отделения медицинского вуза (на английском языке) Ставрополь, 2015 -3- УДК: 576. 3/. 4 : 577 (07) ББК 28. 05 я73 М 15 Макаренко Э.Н. КЛЕТКА КАК СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ЕДИНИЦА ЖИЗНИ: методические рекомендации для студентов англоязычного отделения медицинского вуза (на английском языке) / Э.Н. Макаренко, Н.В. Ерина, Т.С. Коптева, Т.С. Николенко – Ставрополь: Изд-во СтГМУ. – 2015 – 42 с. Рекомендации составлены в соответствии с федеральным государственным образовательным стандартом и включают базовые вопросы по цитологии. В методических рекомендациях на современном учебнометодическом уровне изложены материалы об организации прокариотической и эукариотической клетки, подробно рассмотрено строение эукариотической животной клетки. Рекомендации содержат много иллюстраций, диаграмм и таблиц, что поможет иностранным студентам при подготовке к практическим занятиям и экзамену по биологии. Рецензенты: Ильюх М.П., профессор кафедры ботаники, зоологии, общей биологии Северо-Кавказского федерального университета, доктор биологических наук. Заикина И.А., доцент кафедры ботаники, зоологии, общей биологии Северо-Кавказского федерального университета, кандидат биологических наук. Пискарёва Е.И., доцент кафедры гистологии Ставропольского государственного медицинского университета, кандидат медицинских наук. УДК: 576. 3/. 4 : 577 (07) ББК 28. 05 я73 М 15 Рекомендовано к печати редакционно-издательским советом СтГМУ. © Ставропольский государственный медицинский университет, 2015 -4- INTRODUCTION BIOLOGIA is a science that studies all living organisms. There are many life forms (diagram 1) existing on our planet, but all of them are divided on cellular forms and non-cellular forms. Diagram1 CELLULAR AND NON-CELLULAR FORMS OF LIFE According to a cell theory all living organisms consist of cells. Cell is the structural and functional unit of life. Study of a cell begins with discovery of a microscope. Two Dutch brothers Janssen and Francis constructed the first compound microscope in the year of 1590. BRIEF HISTORY OF CELL DISCOVERY All animals and plants are constituted of certain structural units, which are repeated in each animal or plant. Such a concept was originally put forward by Aristotle (384-322 B.C.). Many centuries later, in 1665 Robert Hooke reported honeycomb like structures in a very thin slice of cork (Fig. 1) and coined the term “cell” to describe them. He published his findings in the form of a book titled “Micrographia”. The next year he presented his findings to the Royal Society of London. In 1672, cells in the form of cavities with cellulose walls were observed in different plants by Grew and Marcello Malpighi. Anton Von Leeuwenhoek, for the first time, reported the discovery of protozoans, bacteria, sperm cell and red blood corpuscles in the year of 1674. -5- Fig. 1. Robert Hooke and honeycomb like structures. At the beginning of 19-th century (1802-1826), several workers including a Frenchman, H.J. Dutrochet gave the idea of the cell theory. In 1830 Purkinje coined the term protoplast to describe the cellular substance. Robert Brown discovered a thick, rounded structure in a cell and gave it the name of nucleus in 1831. The same year Dujarin not only recognized the importance of cell organelles but also called nucleus as sarcode. The credit for proposing “cell theory” goes to two German scientists, Mathias Schleiden and Theodore Schwann. In 1838 Schleiden reported that all the plants are made of cells only. The following year, i.e., 1839, Schwann reported that all animals were also made of cells. Schwann also proposed that tissues were composed of cells and the cells were the functional units of all living organisms. Unfortunately both of them wrongly believed that cells originated from non-living substances. Robert Remak and Rudolf Virchow reported that cells always originated from pre-existing cells only. Our knowledge of a cell has reached molecular level with the discovery of the electron microscope by Knoll and Ruska in 1932. PROKARYOTIC CELL Eukaryotes are unicellular and multicellular organisms, but the structured nucleus is always present in their cells (table 1.). Prokaryotes are unicellular organisms that do not have an organized nucleus. Prokaryotic cell is an integral primitive organism (fig. 2), where -6- the surface apparatus of cell is formed. It includes the cell wall or murein envelope, plasma membrane and mucous capsule. The cell membrane forms invaginations or mesosomes. They perform the function of missing cell organelles, such as mitochondria, Golgi complex and other. The cytoplasm is the internal environment of the cell. It includes hyaloplasm, ribosomes and inclusions. The locomotion organelles are present in motile prokaryotes for example flagella. Fig. 2. Structural organization of prokaryotic cells. Genetic apparatus of the cell or nucleus is absent. Nucleoid or genophore is genetic structure of prokaryotic cells, which are responsible for the storage and transmission of hereditary information. Nucleoid is a circular DNA molecule is not associated with proteins. It lies directly in hyaloplasm. Moreover extra-ring DNA fragments or plasmids are meeting in the cytoplasm responsible for cytoplasmic heredity. -7- Table 1 SIMILARITIES AND DIFFERENCES BETWEEN PROKARYOTIC AND EUKARYOTIC CELLS Parts of a cell Structural components Over membrane s e n a g r O T Y O P e L l A l S M Hyaloplasm C Type of the cells EukaryotIc Animal Plasmalemma Surface apparatus Prokaryotic Endoplasmatic reticulum (ER) Mitochondria Golgi apparatus (GA) Peroxysomes Lysosomes Ribosomes Cell center Microtubules Microfilaments Plastids Vacuoles Cilia Flagellae Inclusions Present. It forms invaginations, like mesosomes et al. Cell wall (murein). Sometimes capsule (mucopolysacharides) Plant Present Glycocalyx (glycolipids, glycoproteins) Present Present Mainly granular ─ Cell wall (cellulose) Mainly smooth Present ─ Absent Flattened piles of cisternae (dictyosomes) Present only in higher plants Phagosomes Autophagosomes 70 S – mitochondria 80 S – hyaloplasm and ER Present Present only in lower plants Present Rare Present Absent ─ ─ 70 S Absent Absent Absent Absent Present in some species Proteins, lipids, carbohydrates -8- Forms piles of cisternae and vesicles Present Absent Absent Present Present Proteins, lipids, carbohy- Present Present Absent Absent. Present in some species (algae) Lipids, carbohydrates (glycogen), polyphosphates (volutin’s granules) Cytoskeleton Hereditary apparatus Absent Nucleoid and Plasmids drates (glycogen), secretory granules, pigment et al. Microtubules, microfilaments and microtrabecular fibers Nucleus (starch), protein (gluten), calcium oxalate crystals Occasional microtubules Nucleus EUKARYOTIC CELL The Main Components of any Eukaryotic Cell are (Fig. 3): 1) Plasma membrane - Cytoplasmic membrane - Plasmalemma - Cell membrane 2) Nucleus 3) Cytoplasm Fig. 3. Eukaryotic animal cell. -9- DISTINCTIONS ANIMAL FROM PLANT CELL: The plant cells include (Fig. 4) 1) Cell wall (or cellulose envelope) 2) Plastids: chloroplasts, chromoplasts, and leykoplasts 3) Vacuoles. In animal cell these structures are absent. Fig. 4. Eukaryotic plant cell. PLASMA MEMBRANE – PLASMALEMMA Plasma membrane is the thin film surrounding a cell. Thickness: 7-10 nm. Structure: 1. Composed of lipid and protein molecules. 2. Arrangement of molecules – current postulate: a) “Sandwiched” model proposed by Davson and Danielli (1935) The bimolecular leaflet model proposed by Davson and Danielli is an important step in understanding of the cell membrane structure. It also formed the basis for all the subsequent models. As for this model, membrane lipids are arranged in two layers, in such a way that their polar ends face outwards and non-polar ends face inwards (Fig. 5). The proteins occur as globular proteins and form a continuous layer on either side of the lipid - 10 - bilayer. In this arrangement the lipid bilayer appears sandwiched between the globular protein layers. Fig. 5. “Sandwiched” model. b) “Protein-crystal” model proposed by G.Vanderkooi and D.Е. Green (1970) The structure of biological membranes is similar to the structure of protein crystals with the only difference that lipids play the role of solvent in the membranes. The basis of the membrane is bimolecular layer placed freely of protein globules. The cavity between the globules filled with phospholipids. The polar heads of the lipids are in contact with water on the membrane surface (Fig. 6). The main role in the binding of proteins and lipids in this model is given hydrophobic interactions. The basis for the assertion that the membrane matrix is protein, served as the experimental data of many authors obtained primarily on mitochondrial membranes. Mitochondrial membrane after solvent extraction of almost all lipids kept the form that looks similar to the shape of the membrane to the action of the solvent, and had the same thickness. - 11 - Fig. 6. “Protein-crystal” model. “Fluid mosaic” model proposed by Singer and Nicolson (1972) Of all the models, Fluid-Mosaic Model, proposed by Singer and Nicolson, has the greatest acceptance among the scientists. According to this model the plasma membrane is made of a lipid bilayer, but the proteins do not form a continuous layer (Fig. 7). Instead of it they penetrate into the lipid bilayer partially or completely. As a result, the plasma membrane acquires dynamic quality. The proteins are inserted into the double layer and are not merely attached to it. Some of them span the whole membrane's thickness. The hydrophobic parts are in contact with the hydrophobic tails of the lipid molecules. The polar groups jut out from the membrane. c) Fig. 7. “Fluid mosaic” model. Both the lipid and the protein molecules have full freedom of mobility and are capable of translateral movements. Proteins appear like floating icebergs in a sea of lipid. This model also explains the mechanism involved in the transport of proteins and enzymes across the cell membrane. It also - 12 - describes how the wounds are closed and self healed due to translateral movements of lipids. In this model, there are 3 types of proteins: peripherical, semiintegral and integral proteins. Functions: 1) Barrier function: it helps in isolating the cell interior from the external environment. Besides, it is the compartmentalization. 2) Transport – some, but not all, substances move or are moved through membranes by various mechanism. 3) Receptor function: notably hormones from endocrine gland cells and neurotransmitters from nerve cells bind to binding sites (receptors) of specific surface proteins of plasmalemma thereby initiating changes in cell’s activities. 4) Metabolic function: many of the proteins in cytoplasmic membrane are any enzymes that accelerate (catalyze) an enormous variety of chemical reactions. Enzymes make possible the chemical reactions that keep our cells and our bodies alive. 5) Adhesive function: all cells are linked through specific contacts – tight and loose; in plant cell – plasmodesmae, in animal cell – desmosomes. CELL WALL In plants and mushrooms (including bacteria) a cell is always surrounded by a cell wall lined throughout with plasmalemma (Fig. 8). While the bacterial cell wall consists of murein (protein-lipid-polysaccharide complexes), the cell wall of plants mainly consists of cellulose, hemicelluloses and compounds of pectin acid (table 2). Fig. 8. Cell wall in cell of plants and mushrooms. The presence or absence of cell wall is one of the most important distinctions between plants and animals because the presence or absence of chlorophyll is not as important. Many plants may become heterotrophic due - 13 - to the loss of chlorophyll, but they still have a definite cell wall, which is absent in case of animal cells. The function of cell wall in plants is believed mainly to provide mechanical strength. Particularly in higher plants having vascular system, cell wall provide main supporting framework. Since in animals, some sort of exoskeleton or endoskeleton is present, perhaps there is hardly any necessity of definite cell wall found in plants. Therefore besides others, main function of the cell wall is the same as that of a skeleton in animals. Table 2 OUTLINE SUMMARY ABOUT THE SURFACE APPARATUS OF THE CELL Over membrane complex Types of cells Cell wall Glycocalyx: carbohydrate molecules, linked with proteins (glycoproteins), with lipids (glycolipids) of plasmalemma Mucus capsule (mucopolysacharides) Plasmalemma: Bilipid layer, integral, semiintegral and peripherical proteins Eukaryotic cells Prokaryotic cell Plant cell + Cellulose envelope + Murein envelope Mushro om cell + Chitin envelope Functions: Animal cell Absent - osmotic regulation; - frame; - protection. + A B + S E N ± Present. Forms invaginations, like mesosomes et al. T A B S E N T P R E S E N T - receptor - protection - barrier (defensive, boundary) - transport - receptor - metabolic - adhesive (in multicellular organisms) NUCLEUS Nucleus is the most important component of a cell (table 3). In the nucleus of eukaryotes four parts are identified. They are (Fig. 9): 1) nucleus surface apparatus = nuclear envelope + pore complex + solid plate (lamina) 2) nuclear sap or karyoplasm = karyolymph + nuclear matrix - 14 - 3) nucleolus 4) chromatin or chromosomes. Fig. 9. Nucleus as hereditary apparatus of eukaryotic cell. The nuclear envelope is composed of two membranes, separated from each other by a perinuclear space of 100-500 A˚ widths. Outer membrane bears ribosomes (Fig. 10). Fig. 10. Interaction of a nucleus with cytoplasm. - 15 - Nuclear pores, having a diameter of 400-1000 A˚, perforate the nuclear envelope. Pore complex is protein globules, connected by fibrillar proteins (Fig. 11). Globular proteins in the pore site are settled in 3 rows 8 globules, each frequently 1 central globule is seen (8x3)+1. The nuclear envelope helps in isolating the nuclear material from the cytoplasm, but the nuclear pores represent the places, where substances are exchanged between the nucleus and cytoplasm. Fig. 11. Details of the organization pore complexes: A – top view, B – on the cut (1 – central globule, 2 – peripheral globule, 3 – binding fibrillar fibres, 4 – the fibres in the cytosol, 5 – solid plate,6 – perinuclear space). Solid plate or lamina is amorphic protein formation as compact layer, connected with inner membrane. Nuclear lamina supports the nuclear membrane and comes into contact with the chromatin and nuclear RNA. Nucleolus is non-constant component of nucleus. At the beginning of nuclear division, the nucleolus disappears only to reappear again, at the end of the nuclear division. Nucleoli are formed in the area of secondary constrictions of chromosomes. There are fibrillar and granular components. DNA is absent in the nucleolus, but RNA is present in the form of ribonucleic proteins. Along with phospholipids, a variety of enzymes also occurs in the nucleolus, but histone proteins are absent. Participation in the biosynthesis of ribosome sub-units is the main function of nucleolus. The fluid substance presented in the nucleus is called nuclear sap or karyoplasm. It is colloid solution of protein, nucleic acids and other organic substances. Chromatin threads or granules, nucleolus are suspended in this sap. Nuclear matrix is fibrillar proteins, forming a dense net in the entire volume of nucleus. A number of enzymes in nuclear matrix – DNApolymerase, RNA-polymerase, ribonuclease, alkaline phosphatase etc. also occur in the nuclear sap. They participate in functions like DNA-replication, transcription and polymerization of messenger RNA. - 16 - Chromatin threads and granules in nondividing cells or chromosomes in early stage of a cell division present very important, constant components of nucleus. There are euchromatin and heterochromatin in interphase chromosomes (chromatin). The chemical nature of the chromatin is the DNP. DNP (deoxiribonucleoproteid) consists of DNA=40%, proteins= 60% (85% – histones, 15% – nonhistones), 1% – RNA. Proteins covering the DNA molecules are of two types: the basic histones and the acid nonhistone proteins. Functions of chromosomes are functions of DNA molecules; they determine both the structure and the function of cells and heredity. Besides they passed this heredity information to daughter cells in the process of cell division. Table 3 Nucleus Surface apparatus OUTLINE SUMMARY ABOUT NUCLEUS FUNCTIONS Nuclear envelope: outer (connection with ribosomes) and inner membrane, perinuclear space Pore complex - barrier (protection of the genetic apparatus); - transport (inside the nucleus histones, enzymes for replication, transcription, ribosomal proteins, nucleotides); Solid plate (lamina) Karyoplasm Nuclear matrix Chromatin Nucleolus - internal medium for different processes - support (“skeleton” of nucleus) - participation in transcription and replication - storage or hereditary information; - transport of hereditary information; - itself reproduction or replication - synthesis of r-RNA - assembly of ribosomal subunits - 17 - CYTOPLASM Cytoplasm is the inner environment of cell (Fig. 12). In the composition of the cytoplasm consists of the following structures: 1. Cytoplasmic matrix or cytosol (hyaloplasm), 2. Inclusions, 3. Cytoskeleton, 4. Various types of cell organelles. Fig. 12. The contents of the internal environment of cells: 1 – plasmalemma, 2 – hyaloplasm, 3 – filaments of cytoskeleton, 4 – lipid inclusions, 5 – cell center, 6 – ribosomes, 7 – nucleus, 8 – nuclear pores, 9 – nuclear envelope, 10 – nucleolus, 11 – Golgi apparatus, 12 – mitochondria. CYTOSOL is a translucent, heterogeneous colloidal solution (table 4). Biopolymers with water form a colloidal system, which may be denser (in the form of a gel) or a liquid (in the form of sol) throughout the cytoplasm and in its separate parts according to conditions of external environment. Cytoplasmic matrix filled the space between the organelles and inclusions. - 18 - Table 4 THE CHEMICAL NATURE OF HYALOPLASM H2O Biopolymers Mineral salts Low-molecular organic substances 90 % 7% 0.9 % ≈2% Proteins predominate, Nucleic acids, Polysaccharides, Lipids Amino acids, Monosaccharides and other INCLUSIONS are non-permanent structural components of the cytoplasm. There are 5 kinds of inclusions (diagram 2). Diagram 2 TYPES OF INCLUSIONS Trophic inclusions are reserve of nutrients: lecithin in egg cells, glycogen, and lipids. They are present in all cells almost. Pigment inclusions are melanin, haemoglobin, lipofuscin, bilirubin and etc. These inclusions have a certain color and are characteristic only for some types of cells (melanin – in melanocytes, the haemoglobin – in red blood cells, however, lipofuscin can accumulate in many cell types usually when they are aging). Secretory inclusions are present in secretion cells (zymogene granules in exocrine pancreatic cells, secretory granules in endocrine glands and other. Excretory inclusions: substances that must be removed from the body (for example, granules of uric acid in the epithelium of the renal tubules). Sporadic inclusions are medicaments, particles of carbon, silicon, etc. They are accumulated in some cells in the process of life. CYTOSKELETON of a cell consists of microfilaments, microtubules and microtrabecular fibers (table 5) that lay free in the cytoplasm. - 19 - Table 5 COMPONENTS OF THE CYTOSKELETON Description Microtubules Microtrabecular fibers Microfilaments Diameter Chemical composition Protein nature 20-30 nm globular protein 8-10nm keratin, desmin, vimentin and other fibrillar proteins 4-6 nm actin, rarely second myosin globular protein (actin) Physicochemical properties labile proteins stable proteins Functions 1) Support; 2) Form-building; 3) Regulation of current substances in the cell Elastic frame → (solidity of cell, give it rigidity and elasticity) labile protein (actin) Locomotor → the contraction provides movement of substances in the cell tubulin They are protein structures and play a role in cytoplasmic streaming or cyclosis and in other movements, which occur almost universally in all eukaryotic cells, besides the formation of the cell’s shape. Microtrabecular fibers are small seen fibers, which have a width of about 8 to 10 nm. They form a three-dimensional, irregularly shaped lattice that extends throughout cytoplasm, supports various organelles (ER, mitochondria and so-called free ribosomes). Fig. 13. Cytoskeleton of cell. - 20 - Slightly large fibers are suspended in the lattice. They are called microtubules (200-300 A˚). Microtubules, microfilaments (40-50 A˚) and the microtrabecular lattice together form a supporting framework for the cell or cytoskeleton (fig. 13). Microfilaments also serve as cellular “muscles” (actin and myosin, major contractile proteins of muscle cells, have been identified in cytoskeleton). By contracting and expanding, microfilaments control a cell shape and produce internal cell movements (cyclosis). Microtubules are the material from which centrioles, basal bodies, cilia, flagellum and etc. are constructed (Fig. 14). Fig. 14. Microtubules in structure of cilia and centriole. Thus, their functions are summarized as follows: they play a role in cyclosis, transport of substances and cell division, besides contributing to the shape of the cell. ORGANELLES are constant components of cytoplasm. There are 2 types of organelles (diagram 3). Main cell organelles are present in any eukaryotic cell. Special cell organelles may be in certain types of cells: Microvilli – projections of cytoplasm and plasma membrane; increase surface area of cells whose function is absorption. Cilia – hair like projections of cytoplasm and plasma membrane; each cilium is a tiny cylinder made up of nine double microtubules arranged around two single microtubules; one cell may have a hundred or - 21 - more cilia; they propel fluid in one direction over surface of cell, for example, upward in respiratory tract. Flagellum – single hair like projection from cells surface for example, flagellum of spermatozoon propels it forward in its fluid environment. Diagram 3 CONSTANT COMPONENTS OF CYTOPLASM CENTROSOMES OR THE CELL CENTER Structure: centrosomes are a small, spherical zone close to the nucleus. In the center of the centrosome a pair of cylindrical structures, arranged at right angle to each other is present. They are known as centrioles. Under the light microscope, centrioles appear as two dots located near the nucleus. They are 0,4 μm long with a diameter of 0,2 μm. The electron microscope, however, reveals them not as mere dots but as tiny cylinders of microtubules, with three tubules in each bundle – (9х3)+0. The rest of the centrosome is known as centrosphere (Fig. 15). - 22 - Function: at the beginning of the nuclear division the centrioles replicate and move to the opposite poles. They are responsible for the formation of mitotic spindle or the mitotic apparatus (achromatic apparatus). Fig. 15. Centrosome. RIBOSOMES Ribosomes are exclusively confined to the cytoplasm. They are not found in the nucleus. Their names are ribonucleoprotein particles (RNP). They are mainly composed of proteins and ribonucleic acid (RNA). The ribosomes account for about 85% of the RNA present in a cell. They occur either freely in the cytoplasm, mitochondrial matrix and stroma of plastids or bound to the outer surface of the endoplasmic reticulum. Fig. 16. Sub-units of ribosome. - 23 - Ribosomes are of spheroid structures. They are made of two subunits – one small and one big (Fig. 16). Sub-units of ribosomes synthesize in nucleolus, but their combination in the entire ribosome takes place in cytoplasm. When the concentration of magnesium ions (Mg 2+) is 0.001 M, the sub-units of a ribosome are joined together. They dissociate from each other when the Mg2+ concentration is less than 0.001 M. When the Mg2+ ion concentration is increased ten times, i.e. raised to 0.01 M, two ribosome join together, producing a “dimmer” or “twin particle”. During protein synthesis many ribosomes are attached to the m-RNA strand like beads on a string and the chain of beads move along the m-RNA strand. This chain of ribosomes is called polyribosome or polysome (Fig. 17). They form characteristic whorled pattern on the endoplasmic reticulum. Fig. 17. Polyribosome. - 24 - Function: participation in the protein synthesis is the main function of ribosomes – “protein factories”. During protein synthesis, in addition to serving as workbenches, they participate in the catalytic actions of chain initiation, elongation and termination of a polypeptide chain. ENDOPLASMIC RETICULUM Studies carried out with the electron microscope have revealed the presence of a complex membranous labyrinth in the cytoplasm. It is called endoplasmic reticulum. It even spreads throughout the cell, as a complex interlacing membranous structure. It may establish contact with the nuclear membrane on one side and the plasma membrane on the other side. Sometimes, it is associated with the Goldi complex also. The endoplasmic reticulum is composed of three types of membranous structures (Fig. 18): - Cisternae, - Vesicles, - Tubules. Fig. 18. Membranous structures of the endoplasmic reticulum. Cisternae are the most important structures of endoplasmic reticulum. They appear a stack of parallely arranged structures in the cross section. Each cisterna is 50-60 μm thick. The cisternae are highly developed in - 25 - the cells involved in protein synthesis and secretory activity (e.g. cells of pancreas, liver, neurons etc.). Vesicles are found in abundant numbers in cells of organs like pancreas and do not possess a definite shape and size. They have a diameter of 40-500 millimicrons. Tubules are highly branching and anastomosing structures, with a diameter of 50-190 µm. They are associated with both the cisternae and vesicles. Depending upon the metabolic state of a cell, these membranous structures exhibit transformation from one form to another (i.e. cisternae to vesicles, vesicles to tubules, vesicles to cisternae etc.) The membrane structure of the endoplasmic reticulum is variable in different species. Even within the same species variability exists in different types of cells. But a similar structural pattern is seen in all the cells carrying out the same physiological function. Fig. 19. Types of the endoplasmic reticulum. Types (Fig. 19): the endoplasmic reticulum occurs in two broad types – rough endoplasmic reticulum (RER) and smooth endoplasmic reticulum (SER). The morphological distinction between RER and SER depends on the distribution of ribosomes. When ribosomes are attached to the outer surface of the endoplasmic reticulum it appears rough or granular and so, described as rough endoplasmic reticulum. In the absence of ribosomes, it appears smooth and known as smooth endoplasmic reticulum. - 26 - Cisternae, vesicles and tubules are well developed in RER. In SER, tubules are well developed forming an interlacing system. In a large number of cells RER and SER are observed interconnected and transport of materials from RER to SER has been noticed. As per the need of a cell, RER and SER are interchangeable. Functions: 1. SER participates in the synthesis of lipids (steroids). 2. SER participates in the synthesis of carbohydrates (glucose), secretes the enzyme glucose-6-phosphatase and converts glucose-6-phosphate into glucose in the liver. (Plasma membrane prevents escape of glucose-6phosphate but allows glucose to leave the cell.). 3. RER participates in the synthesis of proteins and enzymes. 4. ER plays an important role in the structural and transportation systems of a cell: canals of reticulum serve for the cell as its inner circulatory system, for example, proteins move through canals on way to Golgi complex. GOLGI APPARATUS Golgi complex was first discovered and described by Camillo Golgi in 1898.This specialized organelle, in the cells, exhibits some degree of variability in structure, in different types of cells. It is well developed in the cells involved in high secretory activity. Golgi apparatus generally occupies a position near the pole or between the pole and the nucleus. However, there are cells in which, it is distributed anywhere in the cytoplasm. Fig. 20. Membranous structures of dictyosome. The structural and functional unit of GA is dictyosome. In plant cell the number of dictyosomes more than in an animal cell. Every dictyosome appears as an assemblage of three types of membranous structures (Fig. 20): - 27 - - Cisternae or flattened sacs, - Vesicles or secretory granules, - Microvesicles. Cisternae are flattened sac-like structures with a diameter of 0.5-100 A˚. Of these, four to eight are arranged one above the other, like a stack of pancakes. The number of such stacks varies from one to a few thousands, depending upon the nature of the cell. The stacks are concave structures, with a shallow cup-like appearance. The cisternae of a stack are separated from each other by a gap of 250-300 A˚. Interconnections between the cisternae have not been observed. Golgi complex is closely associated with the SER. Cisternae of the Golgi complex closest to the SER is called cis-face or outer-edge of the stack. The membrane thickness of this cisterna is 50-60 A˚ and it closely resembles the ER in structure. The farthest cisterna of the stack (from the SER) is known as the trans-face or inner edge of the stack. Structurally, the membrane of this cistern resembles plasma membrane and it has a thickness of 75-100 A˚. In most of the cells, the edges of the cisternae, particularly the ones towards the trans-face, appear dilated. It is believed that the vesicles are budded off from these dilations. As ribosomes are absent, the membrane of Golgi complex appears smooth. The precise relationship between the ER and GA is often unclear. In case of pancreatic cells, RER is found associated with SER and continuous formation of micro vesicles has been observed from the edges of SER. These vesicles probably act as transport vesicles carrying the substances synthesized in the RER to the GA. At the cis-face of GA, the vesicles fuse forming new cisternae. For this reason the cis-face is also known as “forming face”. Thus, as the new cisternae are being added at the cis-face, the cisternae at the trans-face break up into vesicles. So, the trans-face is also known as “maturing face” (Fig. 21). For carrying out the studies on polypeptide synthesis, through pulsechase experiments labelled aminoacids are employed. Such experiments have shown that, newly synthesized pancreatic enzymes take 10-20 minutes time to reach the Golgi complex, from the site of their synthesis in RER, about 40 minutes to enter the condensing vesicles and about 2 hours to leave the cell. In some cells, existences of direct connections between the ER and GA have been noticed. The condensing vesicles develop from the maturing face of the GA. They are described as secretory granules, after the process of condensation is over. The secretory granules travel towards the cell membrane, fuse with it and release their contents into the cell exterior. - 28 - Therefore, Golgi complex is considered as a transitional organelle between the endoplasmic reticulum (ER) and plasma membrane. Fig. 21. Cis-face and trans-face of Golgi complex. Functions: 1. Participates in the processing, packing and distribution of the substances synthesized in the RER. 2. Synthesizes large carbohydrate molecules (cellulose). 3. The chemical substances synthesized in the ER are subjected to cyclical changes in the GA: combines large carbohydrate molecules with proteins and secretes product (glycoproteins). 4. Participates in the condensation of secretory “products”. 5. Participates in the formation of lysosomes. 6. GA is involved in the formation of plasma membrane, in dividing of plant cell (cytokinesis), in producing a cap-like acrosome on the head of the sperm. The hydrolases present in the acrosome facilitate fertilization by dissolving the egg membranes. Also it partakes in the synthesis of yolk in the egg cells. LYSOSOMES In 1949, de Duve along with his colleagues discovered and named this cell organelle as lysosome. Lysosomes carry more than 40 acidhydrolases. - 29 - Lysosomes exhibit great variation in their shape and size. They are spherical, rod-shaped or irregular in shape and have a diameter of 250 A˚ to 1 µm. Lysosomes are temporary structures, surrounded by a lipoprotein membrane. The method of their formation closely resembles the formation of secretory granules in the Golgi apparatus. This process is summarized as follow: 1. The synthesis of lysosomal enzymes in the RER. 2. Their transportation to the GA. 3. Their packaging into lysosomes from the trans-face of the Golgi complex. Lysosomes exhibit polymorphism. Four types of lysosomes are identified (Fig. 22): Primary lysosomes, Digestive vacuoles or secondary lysosomes (heterophagosomes), Residual bodies, Autophagic vacuoles. Fig. 22. Types of lysosomes. Primary lysosomes are also known as storage granules. They are minute granular structures filled with hydrolases synthesized in the ribosome, attached to the RER. - 30 - Digestive vacuoles are also known as heterophagosomes or secondary lysosomes. Phagosomes or phagocytic vesicles are formed in the cells due to endocytosis of food or phagocytosis of harmful microorganism. When primary lysosomes fuse with these structures, digestive vacuoles (heterophagosomes) are formed. They contain substances at different stages of digestion. Residual bodies after digestion and absorption of digested food (amino acids, glucose, fatty acids and glycerol) into the neighboring cytoplasm, the digestive vacuoles are left with undigestible food. Now they are called residual bodies. They come to the cell surface, fuse with the cell membrane and discharge their contents to the outside (exocytosis) In case of vertebrates, a suitable mechanism is absent in the cells for the removal of these residual bodies. As a result, they get accumulated in the cytoplasm and are known as lipofuscin granules. Their number increases with the increasing age of the cell. This type of accumulation is very well represented in the nerve cells, which have a long life span. This may be the main cause for the ageing process in animals. Autophagic vesicles lysosomes also participate in the digestion of cell organelles like mitochondria and ribosomes, this phenomenon is called autophagy. Due to injury, poisoning old age or oxygen deficiency, lysosomes, rupture releasing their enzymes into the cell. Consequently, the cell itself is digested. This is called autolysis or self-destruction. Functions: participates in the digestion of the substances entering the cell. For this reason lysosomes are known as “suicidal bags”, “bags of destruction” or “autolytic vesicles”. Heterophagy: lysosomes participate in the digestion of material present in the phagosomes (vacuoles with solid food) and pinosomes (vacuoles with liquids) Autophagy: lysosomes are also involved in the digestion of other cell organelles like mitochondria, ribosomes, ER, etc. PEROXISOMES Peroxisomes are membrane-bounded sacs of enzymes (Fig. 23) that carry out oxidation reactions in which they combine oxygen with various substrates (oxidase). They are named for hydrogen peroxide (H2O2), which some of these enzymes make. Another peroxisome enzyme, catalase, uses hydrogen peroxide to detoxify harmful substances, especially in the liver and kidneys. For example, peroxisomes detoxify about half the ethanol we consume. - 31 - In modern eukaryotic cells most oxidation takes place in mitochondria during cellular respiration. Some biologists think peroxisomes are the remnants of ancient oxidizing organelles that have largely been supplanted by mitochondria. The advantage of mitochondria over peroxisomes is that the major product of the oxidation reactions is ATP, which the cell can use for energy. Fig. 23. Structure of peroxisome. MITOCHONDRIA The term mitochondrion means thread, granule (GK. “mito” – thread; “chondrion” – granule). It’s plural from is mitochondria. Kolliker observed them first in a muscle cell in 1850. Benda employed a new staining technique, gave a detailed description of its external structure and called it “mitochondrion” in 1898. Fig. 24. Structure of mitochondrion. - 32 - Mitochondria are very important cell organelles, as they carry a unique chemical and structural system for the synthesis of ATP. ATP is the chemical energy required for carrying out various metabolic activities in cells. Because of this, mitochondrion is rightly described as a “power plant” or “power house” of a cell. Mitochondria are also semiautonomous and selfreplicating cell organelles. New mitochondria always develop from preexisting mitochondria, through division. In 1890 Altmann expressed the view that mitochondria are probably bacteria leading a symbiotic life in the cells. Mitochondria display various shapes. Mitochondrial number depends on the functional status of a cell. Electron microscopic studies reveal that mitochondria are made of two membranes of different nature. They are outer and inner membranes. The outer membrane encircles the mitochondrion completely and serves as its outer boundary. The inner membrane produces a number of inward foldings known as “cristae”. The cristae increase the inner surface, providing space for the components of the respiratory chain. The inner cavity of mitochondrion is described as “inner chamber”. It is filled with a jelly-like substance, the matrix. Its jelly-like appearance is due to the presence of a high concentration of soluble proteins. The cavity presented between the outer and inner membranes is known as “inter- membrane space” or “outer chamber” (Fig. 24). It is filled with a watery fluid. The inner membrane space is continuous with the space present within the folds of cristae. Fig. 25. Oxysomes of mitochondrion. - 33 - With the help of electron microscope, Humberto-Fernandez-Moran observed a number of lollipop-like structures attached to the inner side of the inner membrane. They are known as oxysomes (Fig. 25), as they carry oxidative enzymes. They are also called elementary particles, F1 – particles or Fernandez-Moran particles. Each F1 – particles has three parts VIZ; the head piece, the stalk and the base piece. The mitochondrial matrix includes a variety of items not found in other cell organelles. They are ribosomes (smaller than those found in the cytoplasm), circular DNA molecules, filaments, dense granules and enzymes of Krebs’s cycle. The circular DNA molecules help in the synthesis of RNA and proteins required by the mitochondria. One type of dense granules is presented in the matrix store calcium ions, in the form of precipitates of calcium phosphate. The calcium ions play an important role in the regulation of numerous biochemical activities within the cell. Mitochondria thus act as calcium accumulators. Besides, mitochondria are also involved in heat generation. The heat generated by mitochondria helps in the maintenance of constant body temperature in homoiotherms or warm-blooded animals (aves and mammals). PLASTIDS These are the universal important organelles presented only in cells of plants. There are three kinds of plastids (Fig. 26): Chloroplasts Chromoplasts Leucoplasts They are different in the structure and functions, but all plastids take part in synthesis of carbohydrates. Like mitochondria, plastids carry some of their own hereditary material in the form of a circular molecule of DNA and ribosomes in stroma. They are duplicated by division. Chloroplasts are the green plastids. They act as photosynthetic apparatus. The entire process of photosynthesis is completed in each chloroplast. Hence, these are the sites of photosynthetic reactions. Leaves are the specialized photosynthetic organs and hence, they contain maximum number of chloroplasts in their mesophyll cells. Moreover, morphology and anatomy of leaves are most helpful during photosynthesis for getting maximum benefit of sunlight providing steady supply of water to green cells and allowing free exchange of CO2 and O2. Structure: the chloroplasts in higher plants are microscopic and mostly oval, spherical or discoid. Each chloroplast is bounded by two - 34 - smooth and selectively permeable cytoplasmic membranes with an intermembrane space. These membranes are composed of lipoprotein sub-units. Fig. 26. Kinds of plastids. The internal space of the chloroplast is filled with a colorless hydrophilic matrix called stroma. Numbers of grana are suspended in the stroma. Each granum is a stack (compact bundle) of thylakoids. These are membrane-bound flattened, disc-shaped vesicles. The thylakoid membranes are called grana lamellae. All grana are connected with one another by stroma lamellae, i.e. inter-grana lamellae or frets. Internal space of each thylakoid is called fret channel. The thylakoid lamellae are composed of alternating layers of lipids and aqueous proteins. There is a layer of chlorophyll and carotenoid molecules situated between the protein and lipid layers. The chlorophyll molecules are arranged in such a way that their hydrophilic heads extend into the aqueous protein layer while the lipophilic tails are embedded in the lipid layer. The pigments are organized into numerous photosynthetic units called quantasomes. Each quantasome contains about 230 to 300 chlorophyll molecules. Quantasomes are capable of trapping light energy and converting into chemical energy (ATP) during the photochemical reactions - 35 - (light reaction of photosynthesis). The grana also contain various coenzymes and electron acceptors necessary for the process. Hence, grana are the site of the light reaction (phase - I) in photosynthesis. The stroma contains various enzymes required for the dark reaction i.e. the biochemical reactions involving the reduction of CO 2 to form carbohydrates. Hence, stroma is the site of dark reaction (phase - II) of photosynthesis. The grana thylakoids and the stroma lamellae together form an intricate internal membrane system in the chloroplast. This system is derived from the inner limiting membrane during the development of chloroplast. Pigments in photosynthesis: the most common photosynthetic pigments presented in higher plants and green algae are: Chlorophyll-A (blue-green) ► C55H72O5N4Mg Chlorophyll-B (yellow-green) ► C55H70O6N4Mg Carotenoids: Carotenes (orange-red) ► C40H56 Xanthophylls (yellow) ► C40H56O2 Fig. 27. Structure of chloroplast. For photosynthesis, these pigments can absorb and use light belonging to the visible spectrum only. Both chlorophyll-A and B show light absorption maximum in red followed by blue and then in violet regions. Absorption of green light by the chlorophylls is negligible. In fact, chlorophylls reflect green light and hence appear green. - 36 - Carotenoids absorb light in the blue, green and violet regions. Carotenes reflect orange light therefore appear orange. Xanthophylls are yellow colored because they reflect yellow light. Carotenoids protect the chlorophyll from undergoing photo oxidation when exposed to very high light intensity. Chlorophyll-A is the essential pigment in photosynthesis, because only chl-A can utilize the absorbed light energy for the synthesis of chemical energy ATP. Other pigments act as accessory pigments. They collect the light energy and transfer it to chlorophyll-A for photosynthesis. Thylakoids in prokaryotes: In prokaryotes like cyanobacteria, purple bacteria, etc. thylakoids are present but they lie scattered and not organized into grana. Grana or the chloroplasts are absent. In prokaryotes, pigments are distributed uniformly on or in the lamellae. Functions of plastids: 1) The chloroplasts participate in the synthesis of primary carbohydrate (glucose). 2) Chromoplast and leucoplast take part in the synthesis of secondary carbohydrates (starch). Chromoplasts are contained in cells of plant colored organs (for example, flowers, fruits, etc.). The leucoplasts are placed in cells of plant colorless organs. There are seeds, spores or subterranean organs of plants (roots, bulbs, tubers, etc.). - 37 - OUTLINE SUMMARY ABOUT ORGANELLES Hyaloplasm (cytoplasmic matrix) M Endoplasmatic reticulum (ER) O r g T Y C Smooth ER – system of canals, formed with membranes Rough (granular) ER- system of flattened cisternae and canals bearing ribosomes on outer surface Outer- smooth membrane, inner – with crystae. Intermembrane space- matrix; DNA, ribosomes, proteins and inclusions. e l e n Golgi apparatus (Golgi body) a O P l L A s S Mitochondrion Colloid solution of proteins containing organic and mineral substances Lysosome Peroxisome Ribosome Microtubules System of flattened membranous sacks (cisternae), surrounded by a number of micro and macrovesicles (vacuoles). Cis-side of GA is located closer to nucleus, contains microvesicles. Trans-side of GA contains macrovesicles, forming vacuolar zone of GA. Vesicle, bounded by membrane, with homogenous contents (set of hydrolases) Vesicle, bounded by membrane, with crystal-like contents (catalases) Small and large subunits Hollow cylinder, formed with spirally packed dimmers of protein (tubulin) - 38 - - internal medium of the cell, providing process of metabolic reactions - synthesis of lipids - synthesis of oligosaccharides - transport -compartmentalization - synthesis of proteins - maturation of proteins - transport - compartmentalization - energy accumulation (synthesis of ATR) - synthetic (synthesis of proteins) - genetic (replication of DNA) - compartmentalization - maturation, reassortment of protein - formation of primary lysosomes - formation of secretory granules - synthesis of polysaccharides - synthesis of lipids - detoxification - compartmentalization - heterophagy - autophagy - compartmentalization - participation in peroxide oxidation - compartmentalization - synthesis of proteins (translation) - movement (basis of cilia and flagella); - participation in the division of the cells (basis of mitotic apparatus) Microfilaments Cell centre Cilia and flagella Inclusions Cytoskeleton Actin (main amount). Myosin Diplosome (2 centrioles) and centrosphere. Each centrioles is cylinder, consisting of 9 triplets of microtubules (9x3)+0 Cytoplasmatic protrusions (length 10-12μm – cilia, > 1000 μm – flagella), covered by plasmalemma Temporary intracellular structures, accumulated and used during metabolism Microtubules, microfilaments and microtrabecular fibers free lied in cytoplasm - 39 - - contractive -formation of desmosomes - participates in cell division (formation of mitotic apparatus) - movement of a cell (unicellular organisms); - transport of the particles and fluid - nutrition (carbohydrates, lipids) - secretory - pigment (melanin, haemoglobin) - “skeleton” of cell - cyclosis LITERATURE AND INTERNET RESOURCES: 1. Molecular Biology of the Cell by Bruce Alberts, Alexander Johnson, Julian Lewis , David Morgan, Garland Science; 6 edition, 2014. – 1464 pages. – ISBN-13: 978-0815344322. 2. Essential Cell Biology, by Bruce Alberts, Dennis Bray, Karen Hopkin, Alexander D Johnson. – 4th Edition, 2013. – 864 pages. – ISBN13: 978-0815344544. 3. Campbell Biology (10th Edition) by Jane B. Reece, Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Benjamin Cummings. – 10 edition, 2013. – 1488 pages. – ISBN-13: 978-0321775658. 4. Biology by Robert Brooker, Eric Widmaier, Linda Graham, Peter Stiling. – 2nd edition, 2010. – 1440 pages. – ISBN-13: 978-0077349967. 5. Cell Biology and Genetics (Biology: the Unity & Diversity of Life) Cecie Starr, Ralph Taggart, Christine Evers, Lisa Starr. – 14 edition, 2014. – 304 pages. – ISBN-13: 978-1305251243. 6. Biology by Sylvia Mader, Michael Windelspecht. – 11 edition, 2012. – 1024 pages. – ISBN-13: 978-0073525501. 7. Biology by Robert Brooker, Eric Widmaier, Linda Graham, Peter Stiling. – 3 edition,2013. – 1424 pages. – ISBN-13: 978-007353224. 8. Cell Structure & Function (Fundamentals of Biomedical Science) by Guy Orchard, Brian Nation. – 2014. – 448 pages. – ISBN-13: 9780199652471. 9. https://www.khanacademy.org/science/biology/cellular-molecularbiology 10. http://www.biology-online.org/dictionary/Cytology - 40 - CONTENTS Introduction…………………………………………….. Brief history of cell discovery…………………………. Prokaryotic cell………………………………………… Eukaryotic cell…………………………………………. Plasma membrane – Plasmalemma……………………. Cell wall………………………………………………... Nucleus………………………………………………….. Cytoplasm………………………………………………. Cytosol…………………………………………….. Inclusions…………………………………………… Cytoskeleton………………………………………… Organelles: …………………………………………. Centrosomes or the cell center………………... Ribosomes…………………………………….. Polysomes……………………………………… Endoplasmic reticulum………………………... Golgi apparatus……………………………….. Lysosomes……………………………………... Peroxisomes……………………………………. Mitochondria…………………………………… Plastids………………………………………… Outline summary about organelles…………….. Literature…………………………………………………. - 41 - 5 5 6 9 10 13 14 18 18 19 19 21 22 23 24 25 27 29 31 32 34 38 40 АВТОРЫ: Макаренко Элина Николаевна, кандидат медицинских наук, доцент кафедры биологии СтГМУ; Ерина Надежда Викторовна, ассистент кафедры биологии СтГМУ; Коптева Татьяна Сергеевна, ассистент кафедры биологии СтГМУ; Николенко Татьяна Сергеевна, ассистент кафедры биологии СтГМУ. Makarenko E.N., Erina N.V., Kopteva T.S., Nikolenko T.S. МАКАРЕНКО Э.Н., ЕРИНА Н.В., КОПТЕВА Т.С., НИКОЛЕНКО Т.С. Cell as structural and functional unit of life КЛЕТКА КАК СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ ЕДИНИЦА ЖИЗНИ Методические рекомендации для студентов англоязычного отделения медицинского вуза (на английском языке) Сдано в набор __________. Подписано в печать ___________. Бумага типог. № 1. Печать офсетная. Гарнитура офсетная. Усл. печ. л 5,8. Уч.-изд. л. 6,0. Заказ _______. Тираж 100. Ставропольский государственный медицинский университет, 355017, г. Ставрополь, ул. Мира, 310. - 42 -