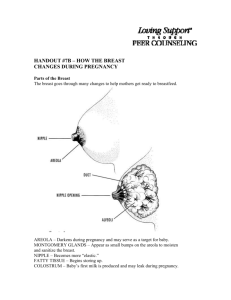
Helder thesis semi-final111111111111111
advertisement

University Code: 分 类 号: Registration No.: 2012393028 密级:秘密 Master Thesis IDENTIFICATION AND CHARACTERIZATION OF NATURAL ANTI-BREAST CANCER COMPOUND: COSTUNOLIDE By HELDER OLIVEIRA Supervised by Dr. XIAOMENG LI Professor, Institute of Genetics and Cytology School of Life Sciences Northeast Normal University School of Life Sciences, Northeast Normal University Changchun, Jilin, P.R. China APRIL, 2014 学校代码: 分 类 号: 研究生学号:2012393028 密级:秘密 Master Thesis IDENTIFICATION AND CHARACTERIZATION OF NATURAL ANTI-BREAST CANCER COMPOUND: COSTUNOLIDE 作者:HELDER 指导教师:Dr. OLIVEIRA Xiaomeng Li,Professor 学科专业:Biochemistry and Molecular Biology 研究方向:Cancer Biology 学位类型:Master’s Degree 东北师范大学学位评定委员会 2014年4月 2 CERTIFICATE This is to certify that Mr. HELDER OLIVEIRA have carried out research work embodied in this thesis entitled “Identification and Characterization of Natural anti-Breast Cancer Compound: Costunolide” in The Key Laboratory of Molecular Epigenetic of MOE, Institute of Genetics and Cytology, School of Life Sciences, Northeast Normal University, Changchun, Jilin, China. The work presented in this thesis is creative and distinctive. His exposition is admirable of presentation to School of Life Sciences, Northeast Normal University, Changchun, Jilin, China for the award of degree of Master of Science in Biochemistry and Molecular biology. Supervisor Dr. Xiaomeng Li, Professor, The Key Laboratory of Molecular Epigenetic of MOE, Institute of Genetics and Cytology, School of Life Sciences, Northeast Normal University, Changchun, Jilin, China 3 ABSTRACT Breast cancer is the most common cancer among women, 23% (1.3 million) of the total of new cases and the second leading cause of cancer death in women exceeded only by lung cancer. Natural medicines have been proven to be a central source of narrative agents with a pharmaceutical potential. Costunolide is sesquiterpene lactones consisting of diverse plant chemicals that exhibit anti cancer action through cytotoxic effects on various cancer cells. The objectives of present study were to explore the effects of natural compounds on the proliferation of MCF-7 cells and to determine the role of ROS in natural compounds-induced apoptosis in breast cancer cells with a therapeutic potential. Results showed that costunolide screened, possess potent anticancer properties against breast cancer MCF-7 cells, Costunolide was observed as strong anti-proliferative agent with IC50 = 50µM. The anti-proliferative effect of costunolide on MCF cells was confirmed by live/dead assay using fluorescent probes calcein AV/PI. The results demonstrated that treatment of cells with costunolide decreased the viability of MCF-7 cells in a dose-dependent manner. To determine the costunolide-induced apoptosis, flow cytometric analysis was carried out. The results showed that costunolide induced apoptosis in a dose-dependent manner in breast cancer MCF-7cells. ROS are well known mediators of intracellular signaling of cascades. The excessive generation of ROS can induce oxidative stress, loss of cell functioning, and apoptosis. In the present study, we assumed that costunolide might arouse ROS level, which could be involved in induction of apoptosis. Therefore, the intracellular ROS level was measured using the ROS-detecting fluorescence dye 2, 7-dichlorofluorescein diacetate (DCF-DA). Interestingly 4 these effects were significantly abrogated when the cells were pretreated with N-acetyl- cysteine (NAC), a specific ROS inhibitor. Costunolide induces apoptosis through extrinsic pathway in MCF-7 breast cancer cells, In order to examine whether costunolide suppresses cell growth inducing apoptotic cell death, we analyzed DNA contents and apoptosis-related proteins expression level by flow cytometry and western blot, respectively in MCF-7 breast cancer cells we investigated whether costunolide activates extrinsic apoptotic pathway. We examined the expression levels of death receptor signaling-related proteins, caspase-3, and PARP. The results showed that procaspase-3 was cleaved to yield 17 and 20kDa fragments and activation of PARP in treated cells with 25 and 50μM of costunolide. Costunolide induce apoptosis through intrinsic mitochondria pathway in MCF-7 breast cancer Cells. We examined the expression levels of mitochondrial apoptotic pathway related proteins such as anti-apoptotic protein, B-cell lymphoma protein-2 (Bcl2), and pro-apoptotic protein Bax. Costunolide involved in the down regulation of Bcl-2 and up regulation of Bax. These results suggest that costunolide may have beneficial effects for the reduction of breast cancer growth, and new therapeutic strategy for the treatment of human cancers. Keywords: Breast cancer, costunolide, apoptosis, reactive oxygen species. 5 乳腺癌是女性最常见的恶性肿瘤之一,在新增癌症病例中占23%。乳腺癌在女性癌症 患者中的致死率位居第二位,仅次于肺癌。天然化合物与合成化合物相比具有低毒的特点 ,是许多疾病的治疗药物生产的最初来源。木香烯内酯是一种倍半萜内酯,由多种植物化 学成分组成,通过细胞毒素作用对多种癌细胞发挥抗癌效应;但木香烯内酯在乳腺癌治疗 中的可能作用尚不明确。本研究旨在探索天然化合物对MCF7细胞增殖的影响,研究ROS 在由天然化合物诱导的乳腺癌细胞凋亡中潜在的治疗作用。 我们首先检测了木香烯内酯对乳腺癌MCF7细胞增殖的影响,并证明其具有高效的抗 癌能力。我们发现,木香烯内酯的IC50值为50µM,是强有力的细胞增殖抑制剂。利用荧 光探针钙绿化素AV/PI活体实验,我们发现木香烯内酯对MCF细胞的增殖抑制和凋亡诱导 效应,降低了MCF-7细胞在剂量依赖下的生存能力。 为了研究木香烯内酯诱导的细胞凋亡,我们采用了流式细胞检测技术。结果显示,在 剂量依赖下,木香烯内酯诱导乳腺癌MCF7细胞凋亡。由于ROS的过度表达会诱导氧化压力,导致细胞功能丧失和细胞凋亡。因此 ,我们利用荧光染色的ROS追踪素-双氯荧光乙酸乙酯来检测细胞内ROS水平,并发现木 香烯内酯显著提高ROS水平。有趣的是,细胞被一种特殊的ROS抑制剂—— 乙酰半胱氨酸预处理之后,ROS介导的细胞凋亡效应被消除。所以在本研究中,我们证明 木香烯内酯通过提高ROS水平,进而诱导细胞凋亡。 6 然后我们研究了木香烯内酯作为外源诱导剂对乳腺癌MCF7细胞凋亡的影响。通过流式细 胞检测技术和蛋白质免疫印记分析了DNA成分和细胞凋亡相关的蛋白质表达水平。在乳 腺癌MCF7细胞中,我们检测了木香烯内酯对凋亡外源途径(死亡受体途径)激活及对凋 亡效应蛋白质Caspase-3和PARP的活化作用。结果显示, 在25uM和50uM木香烯内酯处理的细胞中,蛋白质Caspase-3分裂为一个17kDa 和一个20kDa的片段,蛋白质PARP也被活化剪切。在此基础上,我们进一步检测了木香 烯内酯乳腺癌MCF7细胞凋亡过程中对凋亡内源途径(线粒体途径)的激活效应。我们检 测了线粒体凋亡途径中抗凋亡蛋白质BcL2和促凋亡蛋白Bax的表达水平,并证明木香烯内 酯可下调BcL2蛋白表达和上调Bax蛋白表达,从而证明木香烯内酯通过活化两种凋亡途径 从而诱导乳腺癌MCF7细胞产生凋亡。我们的研究结果表明,木香烯内酯对乳腺癌细胞具 有增殖抑制和凋亡促进效应,为人类癌症治疗提供了一种可能的新策略。 关键词: 乳腺癌,木香烯内酯,细胞凋亡,活性氧 7 Contents ABSTRACT .................................................................................................................................... 4 摘 要...............................................................................................Error! Bookmark not defined. CHAPTER I: LITERATURE REVIEW .................................................................................. 10 1.1 Introduction ......................................................................................................................... 10 1.2 Breast cancer ....................................................................................................................... 11 1.3 Types of breast cancer ......................................................................................................... 13 1.4 Epidiliology and risk factors ............................................................................................... 15 1.5 Apoptosis............................................................................................................................. 16 1.6 Reactive oxygen species...................................................................................................... 18 1.7 Cell cycle progression ............................................................................................................. 19 1.8 Mitochondria menbrane potential (ΔΨm) ........................................................................... 21 1.9 Western Bloting................................................................................................................... 23 1.10 Hypothesis of research work ............................................................................................. 25 1.11 Objectives of present study ............................................................................................... 25 CHAPTER II: NATURAL COMPOUNDS CHEMOPREVENTIVE AGENTS ................. 26 2.1 Medicinal Plants .................................................................................................................. 26 2.2 Alkaloids exert anti-proliferative activity on cancer cells .................................................. 27 2.3 Costunolide.......................................................................................................................... 29 2.4 Molecular targets of costunolide ......................................................................................... 30 CHAPTER III: MATERIAL AND METHODS ..................................................................... 32 3.1 Chemicals and Reagents...................................................................................................... 32 3.2 Cell Culture ......................................................................................................................... 32 8 3.3 Cell Proliferation Assay (MTT) .......................................................................................... 32 3.4 Flow Cytometric Analysis of Cell Cycle ............................................................................ 33 3.5 Flow Cytometric Determination of Apoptosis .................................................................... 33 3.6 Flow Cytometric Determination of Reactive Oxygen Species (ROS) ................................ 34 3.7 Flow Cytometric Determination of Mitochondrial Membrane Potential (ΔΨm) ............... 34 3.8 Western Blotting ................................................................................................................. 34 3.9 Statistical Analysis of Data ................................................................................................. 35 CHAPTER IV: RESULTS AND DISCUSSION ...................................................................... 36 4.1 Anti-proliferative effects of Costunolide on breast cancer cells ......................................... 36 4.2 Costunolide Induced G2/M Cell Cycle Arrest on breast cancer MCF7 Cells .................... 37 4.3 Costunolide Induced Apoptotic Cell Death in Breast Cancer MCF7 Cells ........................ 38 4.4 Costunolide Increased Generation of ROS in MCF7 Cells................................................. 40 4.5 Costunolide Decreased Mitochondrial Membrane Potential in MCF7 cells....................... 41 4.6 Costunolide Regulated Apoptosis-Related Proteins in MCF-7 Cells ................................. 42 4.7 CONCLUSION ................................................................................................................... 44 4.8 Novel findings and significance .......................................................................................... 44 5.3 List of abreviations .............................................................................................................. 46 References ................................................................................................................................. 47 ACKNOWLEDGEMENTS .......................................................................................................... 66 9 Chapter I: Literature review 1.1 Introduction Trillions of living cells formed in the human body grow, divide and die in an orderly fashion. Normal people during the early years, cells divide faster to allow the person to grow. When the person becomes an adult, most cells divide only to replace worn-out or dying cells or to repair injuries. Cancer begins when cells in a part of the body start to grow out of control. There are many kinds of cancer, but they all start because of out-of-control growth of abnormal cells [1]. Cancer comprises at least 100 different diseases, and each is classified by the type of cell that is initially affected. Breast cancer is a type of cancer originating from breast tissue, most commonly from the inner lining of milk ducts or the lobules that supply the ducts with milk. Cancers originating from ducts are known as ductal carcinomas, while those originating from lobules are known as lobular carcinomas. Breast cancer occurs in humans and other mammals [3].remainder of the breast is made up of fatty, connective, and lymphatic tissue. Most masses are benign; that is, they are not cancerous, do not grow uncontrollably or spread, and are not lifethreatening [2]. Plants have been long since use in the treatment of cancer and there are higher than 3000 plant species that have served in the treatment of cancer [4]. Traditional Chinese medicine and a practice that already takes 500 years of tradition, and still maintains an important attention in primary health in rural areas of China and is also very apparent in urban areas, nowadays the Chinese government has made enormous effort to modernize this practice investing capital in scientific research and economic exploitation of the same. In the Western world, interest in TCM 10 is born of the hope that it might complement Western medicine. The history, philosophy, theory and practice of TCM have been recently reviewed [5]. Regulation of apoptosis and cell cycle is an important process to preserve cell homeostasis between cell death and cell proliferation [6], which means that the induction of apoptosis and suppression of cell cycle is only an advantageous strategy for cancer therapy. In addition, the maximum effects and minimize side effects at low concentrations of drugs represent efficiency for cancer therapy. The purpose of this study was to evaluate the effect of costunolide on cell growth, cell cycle progression and induction of ROS-mediated apoptosis in MCF-7 breast cancer cells. 1.2 Breast cancer Breast cancer is a malignant tumor that is initiated in the cells of the breast. A malignant tumor is a collection of cancer cells that can grow into (invade) surrounding tissues or spread (metastasize) to distant areas of the body. The disease occurs almost entirely in women, but men can get it too [8]. Among females the most frequently diagnosed cancer and the leading of cancer death is breast cancer, 23% (1.3million) of the total new cases and14% (458,400) of total cancer deaths in 2008 worldwide is second only to lung cancer. About half the breast cancer cases and 60% of the deaths are estimated to occur in economically developing countries. In general, incidence rates are high in Western and Northern Europe, Australia/New Zealand, and North America; intermediate in South America, the Caribbean, and Northern Africa; and low in sub-Saharan Africa and Asia figure 1 [2]. 11 Figure 1: Incidence rates for total cancers by gender and country. Source: GLOBOCAN 2008 The female breast is made up mainly of lobules (milk-producing glands), ducts (tiny tubes that carry the milk from the lobules to the nipple), and stroma (fatty tissue and connective tissue surrounding the ducts and lobules (figure 2), blood vessels, and lymphatic vessels). Most breast cancers start in the cells that line the ducts (ductal cancers). Some begin in cells that line the lobules (lobular cancers), while a small number start in other tissues [8,1]. 12 1.3 Types of breast cancer The histological classification of breast cancer includes adenocarcinoma, cancers that originate in the glandular tissue, which include the ducts and lobules and sarcomas, cancers that originate in the connective tissue of muscle, fat or blood vessels. Carcinoma in situ (CIS) is an early stage form of cancer where the tumor is confined to the layer of the cells where the cancer began and it has not invaded deeper breast tissue or spread to other areas of the body [9]. There are several types of breast cancer, but some of them are quite rare. In some cases a single breast tumor can be a combination of these types or be a mixture of invasive and in situ cancer [10]. Ductal carcinoma in situ (DCIS) is the most common type of non-invasive cancer in women, where cancer cells have not spread beyond the duct walls into surrounding breast tissue. The prevalence of DCIS is strongly correlated with mammographic screening and in countries such as the US, can be as high as 18% of all newly diagnosed [11] but in countries such as India, represents a very low proportion of total disease since most cases present in late stage [12]. Invasive or infiltrating ductal carcinoma originates in the breast duct, has broken through the wall of the duct into surrounding fatty tissue of the breast and is capable of metastasizing to other organs of the body through the lymphatic system and bloodstream. This represents about 80% of breast cancers [13]. Lobular carcinoma in situ (LCIS) is not cancer but is sometimes classified as a noninvasive breast cancer and women who have this condition are more likely to develop invasive breast cancer in the future [14]. 13 Invasive or infiltrating lobular carcinoma originates in the milk-producing glands or lobules of the breast and can spread to other parts of the body. This is less common and represents about 1 in 10 breast cancer diagnoses [15]. Other (less common) types of breast cancer: inflammatory breast cancer (1-3% of all breast cancers), triple negative breast cancers, mixed tumors, medullary carcinoma (3-5% of all breast cancers), metaplastic carcinoma, mucinous carcinoma, paget disease of the nipple, tubular carcinoma, papillary carcinoma, adenoid cystic carcinoma (adenocystic carcinoma), phyllodes tumor and angiosarcoma [14,15]. Estrogen receptor (ER) and Progesterone receptor (PR) status Confirmed carcinomas of the breast are subjected to a test to determine the estrogen receptor (ER) and progesterone receptor (PR) status. Breast cancers that contain estrogen receptors on the outside surface of their cells are ER-positive cancers while those with progesterone receptors are called PR-positive cancers. Women who are positive for either or both receptors generally have a better prognosis because they are more responsive to hormone therapy than without any of these receptors [14]. 14 Figure 2: Normal breast with non–invasive ductal carcinoma in situ (DCIS) in an enlarged cross–section of the duct. A = ducts, B = lobules, C = dilated section of duct to hold milk, D = nipple, E = fat, F = pectoralis major muscle, G = chest wall/rib cage. Enlargement: A = normal duct cells, B = basement membrane, C = lumen (center of duct) 1.4 Epidiliology and risk factors The factors that contribute to the international variation in incidence rates largely stem from differences in reproductive and hormonal factors and the availability of early detection services [7]. Reproductive factors that increase risk include a long menstrual history, nulliparity, recent use of postmenopausal hormone therapy or oral contraceptives, and late age at first birth [16]. Alcohol consumption also increases the risk of breast cancer [17,18]. The breast cancer incidence increases observed in many Western countries in the late 1980s and 1990sb likely result from changes in reproductive factors (including the increased use of postmenopausal hormone therapy) as well as an increased screening intensity [19]. Incidence rates in some of these countries, including the United States, United Kingdom, France, and Australia, sharply decreased from the beginning of the millennium, partly due to lower use of combined postmenopausal hormone therapy [20,21]. In contrast, breast cancer death rates have been decreasing in North America and several European countries over the past 25 years, largely as a result of early detection through mammography and improved treatment [7,22,23]. In many African and Asian countries however, including Uganda, South Korea, and India, incidence and mortality rates have been rising [24;25], with changes in reproductive patterns, physical inactivity, and obesity being the main contributory factors [7,26,27]; increases in breast cancer awareness and screening activity may be partially responsible for the rising incidence in these populations. Maintaining a healthy body weight, increasing physical activity, and minimizing alcohol intake are the best available strategies to reduce the risk of developing breast cancer [28]. Early detection through mammography has been shown to increase treatment options and save lives, although this approach is cost prohibitive and not feasible in most economically developing countries [29]. Recommended early detection strategies in these countries include the promotion of awareness of early signs and symptoms and screening by clinical breast examination [30]. 15 1.5 Apoptosis Extreme and synchronized way of cell death is defined by apoptosis. It is characterized by distinct morphological features, including chromatin condensation and nuclear fragmentation [31, 32]. The importance of signaling has been recognized in cell regulation during normal and disease [33, 34]. Cancer, a complex genetic disease resulting from mutation of oncogenes or tumor suppressor genes, can be developed due to alteration of signaling pathways; it has been well known to have numerous links to PCD [35]. Programmed cell death (PCD) is regulated by proteolytic enzymes called caspases; Bcl-2 and IAP protein families. Caspases activates apoptosis by cleaving precise protein in cytoplasm and nucleus. In cells, caspases exist as inactive precursors or pro-caspases, which are generally stimulated by cleavage by other caspases, generating proteolytic cascade of caspases [36]. Bcl-2 and IAP protein families are involved in activation of caspases. In the regulation of apoptosis and growth arrest p53, the tumor suppressor protein, plays a critical role [37]. The process of apoptosis is activated by two major pathways: the so-called, receptormediated extrinsic and mitochondria-mediated intrinsic pathway [38]. The extrinsic apoptotic pathway is induced by the binding of extracellular ligands (for example, FasL) to transmembrane death receptors (for example, Fas) on the cell surface leading to activation of caspase-8 [39]. On the other hand, the intrinsic apoptotic pathway is induced by changing the permeability of the outer mitochondrial membrane, reducing mitochondrial membrane potential (Δψm), and releasing mitochondrial pro-apoptotic factors including cytochrome c and Apaf-1 into the cytoplasm leading to activation of caspase-9 [40,41]. Caspase-8 of extrinsic pathway and caspase-9 of intrinsic pathway activate caspase-3 and cleave poly(ADP-ribose) polymerase (PARP), thus resulting in apoptosis [42]. 16 Inhibition of cell cycle progression of cancer cells is an important step in cancer therapy because abnormal cell proliferation is observed in cancer cells [43]. Cell proliferation is regulated by multiple checkpoints in cell cycle progression. The transition between G1-S and G2-M phases checkpoints is activated after DNA damage in the mammalian cells leading to cell cycle arrest and is regulated by cyclins, cylin-dependent kinase (CDKs), and CDK inhibitors (CKIs) [44,45]. Figure 1: Schematic representation of the main molecular pathways leading to apoptosis. In the extrinsic pathway upon ligand binding to specific receptors the DISC complex is formed and caspase-8 activated. In the intrinsic pathway release of cyt-c from the mitochondria result in the formation of the apoptosome and activation of caspases-9. Caspase-8 and -9 then activate downstream caspases such as caspases-3 resulting in cell death. The two pathways are connected via the cleavage of the BH3 only protein BID. 17 1.6 Reactive oxygen species Oxidative stress and cell-cycle regulation are two essential elements in the apoptosis process. Reactive oxygen species are emerging as critical signaling molecules [46-47]. The term reactive oxygen species (ROS) encompasses a wide range of molecules. Free radicals are chemical species containing one or more unpaired electrons. Examples include the hydrogen atom, with one unpaired electron, most transition metal ions, nitric oxide, and oxygen, which have two unpaired electrons [46]. The unpaired electrons of oxygen react to form partially reduced highly reactive species that are classified as ROS, including superoxide (O2 -), hydrogen peroxide (H2O2), hydroxyl radical, and peroxynitrite. Various enzyme systems produce ROS, including the mitochondrial electron transport chain, cytochrome P450, lipoxygenase, cyclooxygenase, the NADPH oxidase complex, xanthine oxidase, and peroxisomes [48]. Mitochondrial oxygen metabolism is the dominant source of O2- that results from incomplete coupling of electrons and H+ with oxygen in the electron transport chain. Under normoxic conditions, ROS are maintained within narrow boundaries by scavenging systems, as would be expected where fluxes of such species are involved in cell signaling [49, 50]. Redox balance, the ratio between oxidizing and reducing species within the cell, plays a significant role in the regulation of signaling pathways, including kinase and phosphatase activity and gene expression through modulation of transcription factor function [51,52]. Redox balance is achieved by various enzyme systems that neutralize toxic oxidants, such as ROS. Superoxide dismutases (SOD) catalyze the conversion of O2- to H2O2, which can then be converted to water by catalase or glutathione (GSH) peroxidase coupled with glutathione reductase. Other relevant scavengers include thioredoxin coupled with thioredoxin reductase, and glutaredoxin, which uses GSH as a substrate. GSH plays a central role in maintaining redox homeostasis, and the GSH to 18 oxidized glutathione ratio provides an estimate of cellular redox buffering capacity [53]. Apoptosis may be triggered by oxidative insults [54]. Reactive oxygen species (ROS) are important chemical messengers in normal cells. They keep the balance with antioxidants in healthy cells [55]. When the accumulation of ROS is more than what defense systems can accommodate, ROS can induce damage to DNA, protein and lipids [56]. Overproduction of ROS results in oxidative stress, which mostly results in cell apoptosis [57]. In addition, cell-cycle arrest is tightly associated with apoptosis. Because cancer involves deregulated cell proliferation and survival, inducing cell-cycle arrest is an available treatment to forestall continued tumor proliferation [58]. Cell-cycle regulation and apoptosis share some regulation proteins [59]. Many chemical compounds trigger apoptosis in tumor cells, with accompanying cell-cycle arrest [60, 61]. This schematic depicts the varied caspase pathways involved in apoptosis. The intrinsic or mitochondrial-mediating pathway involves loss of mitochondrial membrane potential and cytochrome c release leading to activation of caspase 9 followed by downstream effecter caspase-3 activation and resultant cell death. The extrinsic pathway involves stimulation of pathways such as Fas (CD95) leading to activation of upstream caspase-8 with resultant stimulation of effecter caspases. Reactive oxygen species (ROS) may act as an extracellular intermediate directly stimulating the mitochondria and/or Fas cell death pathways [62]. 1.7 Cell cycle progression Animal development from a single-cell zygote to fertile adult requires many rounds of cell division. During each division, cells complete an ordered series of events that collectively make up the "cell cycle". This cycle includes two consecutive processes, mainly characterized by 19 DNA replication and segregation of replicated chromosomes into two separate cells. Originally, cell division was divided into two stages: mitosis (M), i.e. the process of nuclear division; and interphase, the interlude between two M phases. Stages of mitosis include prophase, metaphase, anaphase and telophase. Interphase includes G1, S and G2 phases [63]. Replication of DNA occurs in a restricted part of the interphase called S phase. S phase is preceded by a gap called G1 during which the cell is preparing for DNA synthesis and is followed by a gap called G2 during which the cell is prepared for mitosis. Cells in G1 can, before commitment to DNA replication, enter a resting state called G0. Cells in G0 account for the major part of the non-growing, non-proliferating cells in the human body [64]. Figure 5: The stages of the cell cycle progression. Different proteins regulate transition from one cell cycle phase to another in an orderly way. Cyclin-dependent kinases (CDK), a family of serine/threonine protein kinases that are activated at specific points of the cell cycle, require association with a cyclin subunit for their activation. Cyclin D1, cyclin D2, cyclin D3 is required to CDK4 and to CDK6 and CDK-cyclin D complexes are essential for entry in G1 phase , Another G1 cyclin is cyclin E, Cyclin A binds with CDK2 and this complex is required during 20 S phase, In late G2 and early M, cyclin A complexes with CDK1 to promote entry into M. Mitosis is further regulated by cyclin B in complex with CDK1, CDK7-cyclin H complex, also called CAK, all cell cycle phases. In cancer, there are fundamental alterations in the genetic control of cell division, resulting in an unrestrained cell proliferation. Mutations mainly take place in two classes of genes: proto-oncogenes and tumor suppressor genes. In normal cells, the products of protooncogenes to act at different levels along the pathways that stimulate cell proliferation. Mutated versions of proto-oncogenes or oncogenes can promote tumor growth. Inactivation of tumor suppressor genes like pRb and p53 results in dysfunction of proteins that normally inhibit cell cycle progression. Cell cycle deregulation associated with cancer occurs through mutation of proteins important at different levels of the cell cycle. In cancer, mutations have been observed in genes encoding CDK, cyclins, CDK-activating enzymes, CKI, CDK substrates, and checkpoint proteins [86,87]. 1.8 Mitochondria menbrane potential (ΔΨm) Mitochondria have been described to play a key role and perhaps even a central role in the apoptotic process due in part to the mitochondria being the ‘junction’ of at least two distinct signaling pathways. Mitochondrial control of apoptosis has been described at several levels: (1) maintenance of ATP production and (2) mitochondrial membrane potential and mitochondrial membrane permeability for the release of certain apoptogenic factors from the intermembrane space into the cytosol [88]. Mitochondria are very specialized organelles containing an outer membrane (OM) separated from an inner membrane (IM) by an intermembrane space (IMS) containing many proteins implicated in cell death induction following their release from 21 mitochondria. These IMS proteins include caspase-independent death effectors such as nucleases and/or proteases, as well as caspase activators. In addition, certain pro-apoptotic proteins seem to be sequestered in the mitochondrial cristae [89], the pleiomorphic involutions of the IM that increase the surface area for the electron transport chain. Cytochrome c, the first molecule shown to be released from mitochondria in the induction of apoptosis, normally functions in energy production but, upon release from mitochondria, complexes with apoptosis protease-activating factor 1 (Apaf-1), dATP, and procaspase-9 to form the apoptosome [90,91]. Figure 6: Schematic representation of apoptogenic proteins from the mitochondrial inter- membrane space mediating the mitochondrial death decision. Many cell death stimuli converge at mitochondrial level to induce mitochondrial outer membrane permeabilization (MOMP) andsubsequent release of pro-apoptotic proteins including AIF, cytochrome c, Endo G, Omi/HtrA2, and Smac/DIABLO. However, the execution of apoptosis mainly involves activation of a distinct cascade of cysteine-aspartic acid proteases (caspases) that are present as inactive zymogens, causing 22 selective degradation of cell components, and the activation of further caspases. Cell surface death receptor (such as FasR, TRAIL, TNFα7,8) activation typically involves the cleavage and activation of caspase-8 and/or -10, and the subsequent activation of effectors caspases like caspase-3, which is the central caspase responsible for the proteolytic cascade leading to cell death. The caspase cascade may induce the activation of more than just other caspases. Caspase3 can also be activated via a feedback loop involving the mitochondria, amplifying the caspase signal, but how this occurs remains to be elucidated. Besides a direct proteolytic action on the mitochondria, one possible mechanism to account for the mitochondrial amplification/activation of caspases involves apoptotic signaling through Bcl-2 superfamily members: Bax, Bak and Bid, or via changes in calcium homeostasis [92,93,94]. Instead of directly activating caspases, Bax, Bak and Bid induce caspase signaling through an as yet unidentified mechanism involving the mitochondrial release of several proapoptotic proteins, namely cytochrome c, Smac/Diablo [95,96]. Cytochrome c released during apoptosis binds to the adaptor molecule Apaf-1, and leads to the activation of caspase-9 through the formation of the multimeric apoptosome complex [97,98]. Caspase-9 then activates the central caspase in the apoptotic signaling machinery, caspase-3. Once caspase-3 is in action, apoptotic cell death is inevitable [99]. 1.9 Western Bloting The term "blotting" refers to the transfer of biological samples from a gel to a membrane and their subsequent detection on the surface of the membrane. Western blotting (also called immunoblotting because an antibody is used to specifically detect its antigen) was introduced by Towbin, et al. in 1979 and is now a routine technique for protein analysis. The specificity of the antibody-antigen interaction enables a target protein to be identified in the midst of a complex 23 protein mixture. Western blotting can produce qualitative and semi-quantitative data about that protein.[100]. Gel electrophoresis is a technique in which charged molecules, such as protein or DNA, are separated according to physical properties as they are forced through a gel by an electrical current. Proteins are commonly separated using polyacrylamide gel electrophoresis (PAGE) to characterize individual proteins in a complex sample or to examine multiple proteins within a single sample. When combined with Western blotting, PAGE is a powerful analytical tool providing information on the mass, charge, purity or presence of a protein. Several forms of PAGE exist and can provide different types of information about the protein [101]. Following electrophoresis, the protein must be transferred from the electrophoresis gel to a membrane. There are a variety of methods that have been used for this process, including diffusion transfer, capillary transfer, heat-accelerated convectional transfer, vacuum blotting transfer and electro-elution. The transfer method that is most commonly used for proteins is electro-elution or electrophoresis transfer because of its speed and transfer efficiency [102]. Therefore, after the transfer of the proteins from the gel, it is important to block the remaining surface of the membrane to prevent nonspecific binding of the detection antibodies during subsequent steps. A variety of blocking buffers ranging from milk or normal serum to highly purified proteins have been used to block free sites on a membrane. The blocking buffer should improve the sensitivity of the assay by reducing background interference and improving the signal to noise ratio [103]. Western blotting consists of a series of incubations with different immunochemical reagents separated by wash steps to remove unbound reagents and reduce background, thereby increasing the signal: noise ratio. Occasionally, wash buffer formulations consist of only a 24 physiological buffer such as Tris buffered saline (TBS) or phosphate buffered saline (PBS) without any additives [104]. In general, the primary antibody which recognizes the target protein in a Western blot is not directly detectable. Therefore, tagged secondary antibodies or other detection reagents are used as the means of ultimately detecting the target antigen (indirect detection). A wide variety of labeled secondary detection reagents can be used for Western blot detection. While there are many different tags that can be conjugated to a secondary or primary antibody, the detection method used will limit the choice of what can be used in a Western blotting assay [105]. 1.10 Hypothesis of research work To determine the role of ROS in natural compounds- costunolide induced apoptosis in breast cancer cells with a therapeutic potential. 1.11 Objectives of present study To screen natural compound from Chinese Traditional Medicine Herbs (CTMH) library against breast cancer MCF7 cells. To explore the effects of natural compounds on the proliferation of breast cancer MCF-7 cells. To find the novel anti-cancer agents for treatment of breast cancer through screening of Chinese herbal compounds. 25 CHAPTER II: NATURAL COMPOUNDS CHEMOPREVENTIVE AGENTS 2.1 Medicinal Plants Drug resistance is a multi-factorial phenomenon; it involves the expression of defense factors and, or detoxification mechanisms, in addition, it also alters drug target interactions and cellular responses to specific cytotoxic lesions [106]. Recently the attention in medicinal plants and their biologically dynamic derivatives has augmented in relation to the possible development of narrative potential drugs for several pathologies of relevant social impact [107]. Indeed, it is well known that medicinal plants are reported and used in prenatal care, gynecology, obstetrics, skin disorders, respiratory disorders, cardiac diseases, muscular disorders, nervous and mental health [108, 109]. The promising applications of medicines with respect to potential anti-tumor activity for cancer prevention have been recently described [110]. All over the history of realm, as a primary source of medicine the humans have relied on natural products. Herbal medicines have been proven to be a central source of narrative agents with a pharmaceutical potential. The history of traditional Chinese medicine goes back to 2000 to 3000 years. Dating from the Sui Dynasty (610 AD), oral pseudo membranous candidacies was recorded in infants in General Treatise on the Causes and Symptoms of Diseases, written by Yuan-Fang Chao, who named the condition thrush [111]. Traditionally natural products provided a prosperous source of drugs for many diseases including cancer, and plants are vital source of novel natural products [112]. In exploring new anti-cancer drugs plants and plants-constitutes have always played a vital role [106]. Many anticancer drugs in use are either natural products or derivates of natural products. Until now, among all approved anticancer drugs, about 73% drugs are either products derived from natural 26 sources or developed based on knowledge prevailed from natural products [43]. Herbal medicines, such as paclitaxel, vinca alkaloids, amptothecin, and etoposide hold great potential as hopeful agents for the treatment of cancer [113]. In 2008, 225 anticancer drugs have been developed among these drugs 164 were from natural sources, and 108 were derived from plants, 24 from animal sources, 25 from bacterial sources and 7 were from fungal sources. Among 108 plant-based drugs, 46 were in preclinical development, in phase I, 41 in phase II, 5 in phase III and 2 had already reached preregistration stage Among 155 FDA-approved small molecule anti-cancer drugs, 47% were either natural products or directly derived from natural [114]. 2.2 Alkaloids exert anti-proliferative activity on cancer cells Alkaloids are an extremely diverse group of compounds containing a ring structure and nitrogen atom located inside the heterocyclic ring structure. Different categories of alkaloids have been classified on the basis of their biosynthetic pathways [115]. Alkaloids have been reported to exist in a wide distribution of higher plants, such as Ranunculaceae, Leguminosae, Papaveraceae, Menispermaceae, and Loganiaceae kingdoms [115]. Furthermore, several alkaloids demonstrate considerable biological activities, for instance the relieving action of ephedrine for asthma, the analgesic action of morphine, and the anticancer effects of vinblastine [115-116]. Indeed, alkaloids are the most dynamic components in natural herbs, and few of these compounds have already been effectively developed into chemotherapeutic drugs [116,117]. The alkaloids derived from natural sources for instance evodiamine; berberine, piperine, matrine, sanguinarine, and tetrandrine have comparatively more anticancer studies [118]. Berberine an 27 isoquinoline alkaloid is an extensively prescribed Chinese herb, and is broadly distributed in natural herbs, including Rhizoma Coptidis [119]. Berberine possesses an extensive array of bioactivities, like anti-inflammatory, anti-diabetes, sedation, expansion of blood vessels, hepatoprotective, anti-bacterial, anti-ulcer, protection of myocardial ischemia-reperfusion injury, inhibition of platelet aggregation and neuroprotective effects [120-121]. Matrine, a major alkaloid found in several Sophora plants including Sophora flavescens Ait. [122], exhibits a broad range of pharmacological properties such as antiviral, anti-bacterial, anti-asthmatic, anti-inflammatory, anti-obesity, anti-arrhythmic, diuretic, choleretic, anticancer, hepatoprotective, cardioprotective, and nephroprotective effects [123-124]. Piperine a piperidine alkaloid found in famous spices that have been used for centuries, it is extracted from Piper nigrum and Piper longum [125]. Piperine reveals anti-oxidant, anti-diarrheal, anti-convulsant, anti-inflammatory, hypolipidemic, anti-mutagenic, endorsing bile secretion, and tumor inhibitory activities [126, 127]. Sanguinarine a benzophenanthridine alkaloid extracted from the Papaveracea family including Sanguinaria canadensis L. and Chelidonium majus L. [128, 129], possesses antibacterial, anti-schistosomal, antifungal, anti-platelet, and anti-inflammatory properties [130-131], and is used for schistosomiasis control. Tetrandrine bisbenzylisoquinoline alkaloid isolated from the root of Stephania tetrandra reveals a wide range of pharmacological activities, such as immunomodulating, antiinflammatory, anti-arrhythmic, anti-hepatofibrogenetic, anti-portal, anti-cancer, hypertension, and neuroprotective activities [132]. The appearance of drug resistance with poor patient compliance, undesirable side effects of chemotherapy and the higher cost of combination therapy, reveals an extreme demand for a therapeutic regimen having the same or higher beneficial properties of anticancer agents but with low side effects. The use of natural herbs is 28 one possible approach. Alkaloids demonstrate considerable biological activities and can be used as potent anti-cancer agents. To identify a novel and specific inducer of ROS mediated apoptosis in bladder cancer cells, evodiamine, an alkaloid, was screened in the presence or absence of NAC, a specific ROS scavenger, using the MTT assay [133]. 2.3 Costunolide Costunolide, an illustrious sesquiterpene, is an active compound found in medicinal plants and it is isolated from crude extract of Chinese Traditional medicinal herb Saussurea lappa roots [134]. Accrued data shows that Costunolide was extracted from several plant species such as [135], Magnolia kobus [136], Eupatorium lindleyanum [137], Magnolia ovata [138], Laurus nobilis [139], Laurus novocanariensis [140], Cosmos pringlei [141], Magnolia grandiflora [142], Magnolia sieboldii [143], Tsoongiodendron odorum Chun [144], Podachaenium eminens [145], and Michelia floribunda [146]. In medication, costunolide is used as a popular herbal medicine, having antiinflammatory [147-148], anti-ulcer [149], anti-fungal [150, 151], anti-viral [134], and anti-cancer activities [143, 152, 153]. Costunolide was also reported to have antipyretic, anti-mycobacterial activities [154], pro-differentiation, hypolipidemic [155], and inhibitory effects against cellular production of melanin [156]. For the first time, the anticancer property of costunolide was demonstrated in azoxymethane induced rat intestinal carcinogenesis model and then supported by consequent study using hamster buccal pouch carcinogenesis model induced by DMBA [157]. After these two in vivo experiments extensive research was conducted to understand the mechanism accountable for anti-cancer activity of costunolide. Additionally, costunolide has 29 been demonstrated to possess anti-cancer properties on diverse cancer types including leukemia [158], intestinal [159], melanoma [160], and breast cancer [161]. 2.4 Molecular targets of costunolide Reported that two main pathways are involved to initiate apoptosis, the intrinsic or mitochondrial, and the extrinsic or death receptor pathway; both pathways eventually activate the same effectors caspases and apoptosis effectors molecules. In cancer cells costunolide is a strong apoptosis inducer via various pathways, it has been demonstrated that costunolide readily diminish intracellular GSH and suspend the cellular redox balance [162]. The interaction between members of the Bcl-2 protein family regulates the apoptosis through mitochondrial pathway. ROS can also be involved in the process of lipid peroxidatio and/or the cross-linking of thiol groups in proteins; both of these processes can induce the opening of the mitochondrial permeability transition pore (PTP) [163, 164]. Formerly, in human leukemia cells, it has been reported that costunolide induces apoptosis by regulation of mitochondria permeability transition and consequent cytochrome c release [165], and via reactive oxygen species and Bcl-2-dependent mitochondrial transition [165, 162]. When Bcl-2 protein was over expressed it attenuated apoptosis induced by costunolide [147]. The caspases, a family of cysteine proteases, are one of the focal executors of the apoptotic process via triggering of the death receptors and mitochondrial pathways to accomplish the programmed cell death [61]. Caspases are present in the form of inactive zymogens that are activated during apoptosis. Among them, caspase-3 is a frequently activated death protease, catalyzing the specific cleavage of many key cellular proteins [166,167]. Several studies reveal effects of coustunolide on expression of caspases [168,169], and coustunolide induces apoptosis 30 in bladder and gastric adenocarcinoma cells by activation of caspase- 3 and modulating the level of Bcl-2 protein family [127,169]. Moreover, Lei et al. demonstrate that caspase-8 activation is associated with alteration of Bid in liver cancer cells [51a]. Furthermore, the cleavage of specific substrates for caspase- 3, poly (ADP-ribose) polymerase (PARP), is involved costunolide-induced apoptosis in various cancer cells [127,168,176]. Protein Bax induces the release of cytochrome c from the mitochondria. It is considered an important event that affects apoptosis mediated by mitochondrial pathway [168]. Table 1. Molecular targets of costunolide in different cancer types EC50/ Type of cancer Cell lines Concent. Targets References NFκB↓; ↑MMP09↓;PARP ↓;BCL2 ↑;BAX↓; P53↓;TNFα↓;p27Kip1↓; CDK2↓;Fas, caspase8, caspase-3↑, and degradation of PARP; Cdc2↓; cyclin B1↓; tubulin↓;McTN↓;ROS↑ [147,161,162,1 63,176,167] MDA-MB436;231;157 and Bt549 46 10 μM, 25 μM;50 µg/mL Bladder r T24 46.5 µM BAX ↑;BCL2↓;survivin↓;caspase-3↑; PARP ↑; ↑; NAC(i); ROS↑ Ovarian SKOV3, A2780, MPSC1 cells 5 0 µM caspase-3, -8, and -9↑; BCL2↓; BAX ↑; human hepatocellular carcinoma HA22T/VGH 100 µM Chk2 (Thr 68) ↑;Cdc25c (Ser 216) ↑;Cdk1 (Tyr 15) ↑;cyclin B1↑ Prostate LNCaP; (PC-3 and DU-145 36 µM p21↑;cyclin E↑; Ca(2+), cyclin dependent kinase↑ Leukemia U937 cells; HL-60 10, 40, 50 µM Bcl-2↑;JNK(i); GSH↓ Breast r [169] [153,169 . [101] [100] [162, 158 160,101] ↓: Downregulation; ↑: Upregulation 31 CHAPTER III: MATERIAL AND METHODS 3.1 Chemicals and Reagents Cell culture medium, Dulbecco's modified Eagle's medium (DMEM) and MTT (3′-(4, 5dimethyl-thiazol-2-yl)-2, 5-diphenyl tetrazolium bromide), propidium iodide (PI), and dimethyl sulfoxide (DMSO) were purchased from Sigma. Fetal bovine serum (FBS) was purchased from the Hangzhou Sijiqing Biological Engineering Materials Co., Ltd. An annexin V-FITC apoptosis detection kit was purchased from Beyotime Institute of Biotechnology (Shanghai, China). Rabbit polyclonal anti-human Bcl-2, Bax, cleaved caspase-3 and PARP antibodies were purchased from Wuhan Boster Biological Technology Co., Ltd. (Wuhan, China). Mouse anti-β-actin and antirabbit antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Ponceou and cell lysis buffer for western blots and IP were purchased from Bio SS Beijing (Beijing, China). Rhodamine 123 was purchased from Invitrogen (Eugene, OR, USA). 3.2 Cell Culture MCF-7 cells were cultured in Dulbecco's modified Eagle's medium (Gibco, USA) supplemented with 10% fetal bovine serum (Gibco, USA), and maintained in a humidified atmosphere of 5% CO2 in air at 37 °C. Cells were seeded in 10 cm culture, the medium was changed every 2 days, and cells were allowed to grow to approximately 70% confluence before experimentation. 3.3 Cell Proliferation Assay (MTT) MMT assay was used to determine the cytotoxic effects of the Costunolide on the MCF-7 breast cancer cells. Briefly, cells were seeded at a density of 1×104 cells per well in 96-well microtiter plates and were allowed to grow overnight. Cells were incubated with 100µL of 32 complete culture medium containing Costunolide, after 24 hours incubation, cell growth was measured by adding 10µL MTT (5mg/mL in phosphate buffered saline) to each well and incubated for 4 hour at 37o C. After 4 hours incubation medium was removed and 150µL DMSO was added to each well following shaking gently and carefully. The absorbance was read at a wavelength of 490 nm in a plate reader (ELX 800, BIO-TEK Instruments Inc., Winooski, VT, USA). 3.4 Flow Cytometric Analysis of Cell Cycle For cell cycle analysis, Breast cancer cells were seeded in 6-well plates for overnight and then treated with Costunolide, for 24 hours. After 24 h incubation MCF-7 cells were trypsinized and fixed in Ice-cold ethanol (70%) at 4oC for 10 minutes. After incubation cell pellet was washed and re-suspend in propidium iodide (PI) staining buffer and incubated at 37oC for 15 minutes and then the percentages of cells in the different phases of cell cycle were evaluated by determining the PI stained DNA contents by flow cytometry (Beckman Coulter, Epics XL, Miami, FL, USA). 3.5 Flow Cytometric Determination of Apoptosis Apoptotic cell death was measured by double staining annexin V-FITC and PI using the annexin V-FITC apoptosis detection kit (Beyotime Biotechnology, Shanghai, China) according to the manufacturer’s instructions. For these cells were cultured in 12-well plates and allowed to attach overnight following treatment with indicated concentration of Costunolide, for 24 hours. After 24 hour apoptotic cells were collected, washed in cold PBS and 1×105 cells were resuspended in 100μL annexin-V binding buffer and 5μL annexin V-FITC for 10 minutes in dark at room temperature, then 5μL PI was added. Flow cytometric analysis was performed 33 immediately after staining. Data acquisition and analysis were performed by flow cytometry using the Cell Quest software. 3.6 Flow Cytometric Determination of Reactive Oxygen Species (ROS) In order to determine the intracellular changes in ROS generation, MCF-7 cells were stained with 2', 7'-dichlorofluorescein-diacetate (DCFH-DA). The fluorescent dye DCFH-DA is cell membrane permeable and is converted into the cell membrane impermeable non-fluorescent compound DCFH by intracellular esterases. Oxidation of DCFH by reactive oxygen species produces highly fluorescent DCF. The fluorescence intensity of DCF inside the cells is proportional to the amount of peroxide produced. Briefly, cells were treated with indicated concentration of natural compounds for 24 hours. After treatment, cells were further incubated with 10μM final concentration of DCFH-DA at 37 °C for 30 min. Consequently, cells were collected, washed, re-suspended in PBS, filtered with 300 apertures and analyzed for 2',7'dichlorofluorescein (DCF) fluorescence by flow cytometry (Beckman Coulter, Epics XL). 3.7 Flow Cytometric Determination of Mitochondrial Membrane Potential (ΔΨm) To probe the changes in ΔΨm, MCF-7 cells were stained with rhodamine 123 (1μM) after treatment with indicated concentration of costunolide, and control group for 24 hours. The fluorescence of Rhodamine 123 was measured by flow cytometry with excitation and emission wavelengths of 488 and 530 nm. 3.8 Western Blotting Western Blot analysis was used to determine the mechanism of the apoptotic effect of Costunolide on apoptotic associated proteins. Briefly, MCF-7 cells were incubated with 34 indicated concentration of Costunolide for 24 h. Cells were trypsinized, collected in 1.5mL centrifuge tube and washed with PBS. The cell pellets were re-suspended in cell lysis buffer and were lysed on ice for 30 min. After centrifugation for 15 min, the supernatant fluids were collected and the protein content of the supernatant was measured by the Nano-Drop 1000 spectrophotometer (Thermo Scientific, Waltham, MA, USA). The protein lysates were separated by electrophoresis on 10% SDS-polyacrylamide gel and transferred to a PVDF membrane (Amersham Biosciences, Piscataway, NJ, USA). The membranes were soaked in blocking buffer (5% skimmed milk) for 2 h. To probe for BAX, Bcl-2, cleaved caspase-3, PARP, and β-actin; membranes were incubated overnight at 4°C with relevant antibodies, followed by appropriate HRP conjugated secondary antibodies and ECL detection 3.9 Statistical Analysis of Data For the statistical analysis of data, comparisons between results from different groups were analyzed with SPSS for Window Version 15.0. Student’s t-test was employed to determine the statistical significance of the difference between different experimental groups and control group. p < 0.05 value was defined as statistically significant. All experiments were repeated at least two times. Data was presented as mean ± standard deviation (S.D). 35 CHAPTER IV: RESULTS AND DISCUSSION 4.1 Anti-proliferative effects of Costunolide on breast cancer cells We evaluated the effects of costunolide (fig 7) on the growth of MCF-7, we performed cell viability assay. MCF-7 cell line was used to assess the anti-proliferative effects of costunolide. The cells were treated with Costunolide for 24 h. MTT assay showed that costunolide inhibited cell growth in a dose- and time-dependent manner. Previous results showed that costunolide have antiproliferative effects on breast cancer cells [151,152,153] and in others cancer cells such as colon, melanoma, ovary, lung, leukemia [164, 165], pancreatic carcinoma cells, MK-1, HeLa and B16F10 cells [144]. Pretreatment with 5mM NAC restored the viability of cells indicating that costunolide exerts cytotoxic effect on cell viability through ROS generation. cytotoxic effects on various cancer cells such as human leukemia cell line [30, 31], lung, ovary, colon, melanoma cancer cells[153], human bladder cancer [169], and human breast cancer [,166,167,168]. 36 Figure 7: Costunolide inhibited the cell growth and induced cell death. MCF-7 Cells were treated with indicated doses of costunolide in the presence or absence of NAC for 24 h and cell viability was measured by MTT assay. Data are expressed as Mean ± SD (n = 3). Columns not sharing the same superscript letter differ significantly (p < 0.05). 4.2 Costunolide Induced G2/M Cell Cycle Arrest on breast cancer MCF7 Cells Cell cycle arrest is one of the major causes of cell death, many studies show that cells with malfunctions checkpoint are more vulnerable to anticancer agents [119]. Cells can arrest at cell cycle checkpoints temporarily to allow for: (i) cellular damage to be repaired; (ii) the dissipation of an exogenous cellular stress signal; or (iii) availability of essential growth factors, hormones, or nutrients. Checkpoint signaling may also result in activation of pathways leading to programmed cell death if cellular damage cannot be properly repaired. Defects in cell cycle checkpoints can result in gene mutations, chromosome damage, and aneuploidy, all of which can contribute to tumorigenesis. Many laboratories are now searching for compounds that interfere with cell cycle checkpoints, in the hope that such agents will be more effective in anticancer therapy [180]. The result showed that costunolide arrested cell cycle at G2/M phase and the percentage of accumulation of cells in the G2/M phase was increased from 4.9% in control group to 20.4% and 25.9% in the cells treated with 25 and 50μM of costunolide for 24 hours respectively. This increase was coupled with the decreased percentage of cells in G0/G1 phase as showed in Figure 8. Acumulated data reported that costunolide induced G1 phase cell cycle arrest in human prostate cancer cells [151] and G2/M phase arrest in human hepatocellular carcinoma cells , Soft tissue sarcomas (STS) [162], human breast cancer [156] Human bladder cancer T24 cells [161]. 37 control 25µM 50µM Figure 8: Flow cytometry analysis of cell cycle phase distribution. Flow cytometry analysis of cell cycle phase distribution in MCF-7 cells treated with 25 and 50μM costunolide for 24 hours. Data are expressed as Mean ± SD (n = 3). Columns not sharing the same superscript letter differ significantly (p < 0.05). 4.3 Costunolide Induced Apoptotic Cell Death in Breast Cancer MCF7 Cells Apoptosis, autophagy, and necrosis are the major types of cell death [163]. Among the three major pathways of cell death, apoptosis is most well planned and orderly mode of cell death [13, 164]. More than 50% of neoplasms undergo aberrations in the apoptotic machinery which leads to abnormal cell proliferation [155, 156]. The regulation of apoptosis is, therefore, 38 the most important in the treatment of cancer [126-168]. Accumulated evidences indicated that the most of chemotherapeutic agents halt tumor cells proliferation via induction of apoptosis [119, 190, 121]. We wanted to know if costunolide inhibits MCF7 cells through induction of apoptosis Costunolide-induced apoptosis was determined by flow cytometric analysis. Cells were sown in 12 well plates. After incubation of cells with costunolide for 24 hours, cells were collected in centrifuge tubes and stained with annexin V-FITC and PI double staining as described in the Experimental section. The results of flow cytometric analysis showed that the rates of apoptosis were 20.77 ± 1.36% and 52 ± 1.53% and in the cells treated with 25 and 50μM of costunolide respectively for 24 hours as compared to the 4.68 ± 0.42% in control cells. Pretreatment with NAC completely blocked the apoptotic effect of costunolide indicating that induction of apoptosis is a ROS-dependent manner. Costunolide-induced apoptosis in breast cancer cells was compatible with previously reported studies [139, 159,166, 147]. Figure 9: Apoptosis induced by costunolide in MCF-7 cells. (A) MCF-7 cells were treated with 25 and 50 μM of costunolide for 24 hours in the presence or absence of NAC. Then cells were stained with 39 FITC-conjugated Annexin V and PI for flow cytometric analysis. The flow cytometry profile represents Annexin V-FITC staining in x axis and PI in y axis. (B) Data are expressed as Mean ± SD (n = 3). Columns not sharing the same superscript letter differ significantly (p < 0.05). 4.4 Costunolide Increased Generation of ROS in MCF7 Cells Oxidative stress is the cellular status associated with enhanced production of intracellular ROS and/or impaired function of the cellular anti-oxidant defense system [162]. The intracellular redox status is a precise balance between oxidative stress and endogenous thiol buffers present in the cell [133]. It has been well-established that the intracellular redox status plays an important role in cell survival and death [144]. Unbalanced intra-cellular redox states trigger downstream cellular events, such as alterations of mitochondrial function and cell signaling pathways [195], which lead to apoptotic cell death. Many cancer therapeutic drugs can induce apoptosis by disrupting the redox balance. Formerly, in human leukemia cells, it has been reported that costunolide induces apoptosis by regulation of mitochondria permeability transition and consequent cytochrome c release [75], and via reactive oxygen species and Bcl-2-dependent mitochondrial transition [166,158]. In our study, we addressed the question of whether or not costunolide could induce apoptosis and increase ROS generation in breast cancer cells. The intracellular ROS level of costunolide treated with MCF-7 cells was measured using the ROS-detecting fluorescence dye 2, 7-dichlorofluorescein diacetate (DCF-DA). The level of ROS was significantly increased after treating the cells with indicated dose of costunolide for 24 hours. As shown in Figure 10 A, B, the ratio of DCF-positive cells, treated with 25 and 50 μM costunolide was significantly higher (19.52 ± 1.73 and 35.94 ± 1.83 vs. 1.07 ± 0.53 in control group, p < 0.05). The findings evidenced that costunolide had enhanced the generation of ROS in Breast cancer cells, which corroborated with a previous report about Breast cancer [126]. 40 Figure 10: Flow cytometry analysis of ROS generation. (A) MCF-7 cells were treated with 25 and 50μM costunolide in the presence or absence of 5mM NAC for 24 h. (B) Data are expressed as mean ± SD (n=3). Columns not sharing the same superscript letter differ significantly (p < 0.05). 4.5 Costunolide Decreased Mitochondrial Membrane Potential in MCF7 cells Mitochondria have been described to play a key role and perhaps even a central role in the apoptosis process due in part to the mitochondria being the ‘junction’ of at least two distinct signaling pathways, which contains pro-apoptotic proteins (e.g., cytochrome c) [104]. It has been elucidated that upon the depolarization of the mitochondrial membrane potential results in mitochondrial swelling and subsequent release of cytochrome c from the intermitochondrial membrane space into the cytosol [105]. It is becoming increasingly apparent that the mitochondria play a key role in the processes leading to cell death [169]. The effects of costunolide on the mitochondrial membrane potential of MCF-7 cells were determined by flow cytometry using Rhodamine 123 staining. The rates of depletion of mitochondrial membrane potential were 80.1% and 71.3% in the cells treated with 25 and 50μM 41 of costunolide, respectively, for 24 h as compared to 94.6 % in the control group. To further confirm the involvement of ROS in disruption of mitochondrial membrane potential, cells were treated with 5mM NAC. Pretreatment with NAC completely prevented dissipation of mitochondrial membrane potential, indicating that this was ROS-dependent as shown in figure 11 A B Figure 11: The effects of costunolide on mitochondrial membrane potential of MCF7 cells. The effects of costunolide on mitochondrial membrane potential of MCF7 cells were determined by flow cytometry. (A) The values indicates the percentages of Rhodamine 123 fluorescence in the MCF7 cells treated without and with 25 and 50μM of costunolide for 24 hours in the presence or absence of NAC. (B) Data are expressed as Mean ± SD (n = 3). Columns not sharing the same superscript letter differ significantly (p < 0.05). 4.6 Costunolide Regulated Apoptosis-Related Proteins in MCF-7 Cells Costunolide induces apoptosis through extrinsic pathway in MCF-7 breast cancer cells, In order to examine whether costunolide suppresses cell growth inducing apoptotic cell death, we analyzed DNA contents and apoptosis-related proteins expression level by flow cytometry and 42 western blot, respectively in MCF-7 breast cancer cells we investigated whether costunolide activates extrinsic apoptotic pathway we examined the expression levels of death receptor signaling-related proteins including caspase-3, and PARP. The results showed that procaspase-3 was cleaved .Da fragments and activation of PARP in treated cells with 25 and 50 μM of costunolide after 24 hours as compared to that of control cells as shown in Figure 12B. To determine whether costunolide induces intrinsic mitochondria apoptotic pathway in MCF-7 breast cancer cells, we examined the expression levels of mitochondrial apoptotic pathway related proteins such as anti-apoptotic protein, B-cell lymphoma protein-2 (Bcl2), and proapoptotic protein Bax It was observed that costunolide involved in the down regulation of Bcl-2 in a dose-dependent manner, and up regulation of BAX as shown in Figure 11. These results are similar with previously reported studies [162,116, 169] A B Figure 12: The effect of costunolide on the expression of major apoptosis regulatory proteins. MCF-7 cells were exposed to 25 and 50μM of costunolide for 24 hours. Equal amounts of lysate protein were subjected to gel electrophoresis. (A,B) Expression levels of Bax, Bcl-2, caspase-3 and PARP were monitored by western blot assay. β-actin was used as loading control 43 4.7 CONCLUSION In this study we reported the anti-proliferative potential and apoptotic effect of costunolide against human breast cancer MCF-7 cells. Ours results showed that costunolide is effectively inhibits the proliferation of breast cancer cells through induction of ROS-mediated apoptosis. We also observed that costunolide treatment accumulated the cell population at G2/M phase, costunolide induces apoptosis through extrinsic pathway in Breast cancer cell, This event was confirmed with the appearance of cleaved caspase-3 and PARP and costunolide also induce apoptosis through intrinsic mitochondria pathway due induced the up-regulation of BAX and the parallel down-regulation Bcl-2 expression. Costunolide Decreased Mitochondrial Membrane Potential. To further confirm the involvement of ROS in disruption of mitochondrial membrane potential, cells were treated with 5mM NAC. Pretreatment with NAC completely prevented dissipation of mitochondrial membrane potential, indicating that the mechanism was ROSdependent. These results suggest that costunolide may have beneficial effects for the reduction of breast cancer growth 4.8 Novel findings and significance In this study, we reported anti-proliferative activities of Costunolide in human breast cancer MCF-7 cells. We reported that Costunolide, induced apoptosis in dose dependent manner in human breast cancer MCF-7 cells. 44 In this study, for the first time we reported that costunolide, exerts its growth inhibitory effect through ROS generation, and G2/M phase cell cycle arrest human breast cancer MCF-7 cells. In this study, we reported the ROS-dependent induction of apoptosis by costunolide in human breast cancer MCF-7 cells. We reported that costunolide involved in the down regulation of anti-apoptotic proteins, Bcl-2, and the activation of pro-apoptotic protein, PARP in a dose-dependent manner. These findings provide the rationale for further molecular mechanistic and in vivo investigation of costunolide, against human breast cancer MCF-7 cells. 45 5.3 List of abreviations CDK- Cyclin-dependent kinases CKIs -Cyclin-dependent Kinase Inhibitors ATM - Ataxia-telangiectasia-mutated ATR- Rad3-related kinase DNA-PK -DNA protein kinase DNA- Deoxyribonucleic acid TCM-Traditional Chinese medicine ROS-Reactive oxygen species CIS - Carcinoma in situ DCIS- Ductal carcinoma in situ LCIS -Lobular carcinoma in situ ER- Estrogen receptor PR -Progesterone receptor PCD- programmed cell death PARP-poly(ADP-ribose) polymerase H2O2--), hydrogen peroxide SOD-Superoxide dismutases IM- inner membrane OM - outer membrane IMS- intermembrane space MTT-Cell Proliferation Assay NAC- N-acetyl-L-cysteine DMSO- Dimethyl sulfoxide FBS -Fetal bovine serum 46 References 1-American Cancer Society. Cancer Facts and Figures 2013. Atlanta, Ga: American Cancer Society; 2013. 2- Jemal A, Bray F, Center MM, et al. Global cancer statistics. CA Cancer J Clin 2011; 61:69. 3- Sariego J (2010). "Breast cancer in the young patient". The American surgeon 76 (12): 1397– 1401. PMID 21265355 4- F. E. Koehn and G. T. Carter,(2005) “The evolving role of natural products in drug discovery,” Nature Reviews Drug Discovery, vol. 4, no. 3, pp. 206–220, 2005. 5- Zhou, J. et al. (2004) Traditional Chinese Medicine: Molecular Structures, Natural Sources and Applications (2nd edn), Wiley & Sons 6- Hengartner MO (2000) The biochemistry of apoptosis. Nature 407:770–776 7- Jemal A, Center MM, Desantis C, .( 2010) WardEMGlobal patterns of cancer incidence and mortality rates and trends. Cancer Epidemiol Biomarkers Prev.;19:1893-1907. 8- Abeloff MD, Armitage JO, Lichter AS, et al, (2008) eds. Clinical Oncology. 4th ed. Philadelphia, Pa: Elsevier;: 1875–1943. 9- Armstrong D. Ovaries and fallopian tubes. In: Abeloff MD, Armitage JO, Lichter AS, Niederhuber JE, Kastan MB, McKenna WG, eds(2008). Clinical Oncology. 4th ed. Philadelphia, PA: Elsevier;. 10- Schottenfield DaF, Jr., Joseph F (2006). Cancer Epidemiology and Prevention, Third Edition. New York: Oxford University Press; 11- Simpson PT, Gale T, Fulford LG, Reis-Filho JS, Lakhani SR(2003). The diagnosis and management of pre-invasive breast disease: pathology of atypical lobular hyperplasia and lobular carcinoma in situ. Breast Cancer Research. 5(5):258-62. 12- Moore MA, Ariyaratne Y, Badar F, et al. (2010) Cancer epidemiology in South Asia - past, present and future. Asian Pac J Cancer Prev.;2:49-66. 47 13- Abeloff MD, Wolff AC, Weber BL, et al. Cancer of the Breast. In: Abeloff MD, Armitage JO, Lichter AS, et al, eds. Clinical Oncology. 4th ed. Philadelphia, Pa: Elsevier; 2008. 14 -Avis N, Crawford S, Manuel J, et al. (2005)Quality of life among younger women with breast cancer. J Clin Oncol.;23-:3322–30. 15- Altekruse SF KC, Krapcho M, et al. (2009)SEER Cancer Statistics Review, 1975-2007. Bethesda, MD: National Cancer Institute;. 16- Hulka BS, Moorman PG. Breast cancer (2001): hormones and other risk factors. Maturitas.;38:103-113; discussion 113-116. 17- Baan R, Straif K, Grosse Y, et al.( 2007)Carcinogenicity of alcoholic beverages. Lancet Oncol.;8:292-293. 18- Key J, Hodgson S, Omar RZ, et al. (2006) Metaanalysis of studies of alcohol and breast cancer with consideration of the methodologicalissues. Cancer Causes Control.;17:759-770. 19- Althuis MD, Dozier JD, Anderson WF, Devesa SS, Brinton LA.( 2005)Global trends in breast cancer incidence and mortality 1973- 1997. Int J Epidemiol.;34:405-412. 20- Ravdin PM, Cronin KA, Howlader N, et al. (2007) The decrease in breast-cancer incidence in 2003 in the United States. N Engl J Med.;356:1670-1674. 21. Canfell K, Banks E, Moa AM, Beral V. Decrease in breast cancer incidence following a rapid fall in use of hormone replacement therapy in Australia. Med J Aust. 2008;188:641-644. 22- Althuis MD, Dozier JD, Anderson WF, Devesa SS, Brinton LA. Global trends in breast cancer incidence and mortality 1973- 1997. Int J Epidemiol. 2005;34:405-412. 23- Autier P, Boniol M, La Vecchia C, et al. Disparities in breast cancer mortality trends between 30 European countries: retrospective trend analysis of WHO mortality database. BMJ. 2010;341:c3620. 24- Parkin DM, Nambooze S, Wabwire-Mangen F, Wabinga HR. Changing cancer incidence 48 in Kampala, Uganda, 1991-2006. Int J Cancer. 2010;126:1187-1195. 25- Parkin DM, Whelan S, Ferlay J, Storm H, eds. Cancer Incidence in Five Continents. Vol I to VIII. Cancer Base No. 7. Lyon: IARC Press; 2005. 26- Colditz GA, Sellers TA, Trapido E. Epidemiology- identifying the causes and preventability of cancer Nat Rev. 2006;6:75-83. 27- Ito Y, Ioka A, Tanaka M, Nakayama T, Tsukuma H. Trends in cancer incidence and mortality in Osaka, Japan: evaluation of cancer control activities. Cancer Sci. 2009;100:23902395. 28- Kushi LH, Byers T, Doyle C, et al. American Cancer Society Guidelines on Nutrition and Physical Activity for cancer prevention: reducing the risk of cancer with healthy food choices and physical activity. CA Cancer J Clin. 2006;56:254-281; quiz 313-314. 29- Anderson BO, Yip CH, Ramsey SD, et al. Breast cancer in limited-resource countries: health care systems and public policy. Breast J. 2006;12(suppl 1):S54-S69. 30- Anderson BO, Yip CH, Smith RA, et al. Guideline implementation for breast healthcare in low-income and middle-income countries: overview of the Breast Health Global Initiative Global Summit 2007. Cancer. 2008; 113(8 suppl):2221-2243. 30. 31- S. Elmore,(2007). “Apoptosis: a review of programmed cell death,” Toxicologic Pathology, vol. 35, no. 4, pp. 495–516, 32- M. O. Hengartner,( 2000) “The biochemistry of apoptosis,” Nature, vol. 407, no. 6805, pp. 770–776,. 33- G. I. Evan and K. H. Vousden, (2001) “Proliferation, cell cycle and apoptosis in cancer,” Nature, vol. 411, no. 6835, pp. 342–348, 34- D.Hanahan andR. A.Weinberg, (2000). “Thehallmarks of cancer,” Cell, vol. 100, no. 1, pp. 57–70, 49 35- Amelio I, Melino G, Knight RA (2011) Cell death pathology: cross-talk with autophagy and its clinical implications. Biochem. Biophys. Res. Commun. 414, 277–281. 36- Alberts B JA, Lewis J,. Molecular Biology of the Cell. 4th edition. New York: Garland Science. Programmed Cell Death (Apoptosis) 2002. 37- Vousden KH. Activation of the p53 tumor suppressor protein. Biochim Biophys Acta 2002; 1602: 47-59. 38- Konopleva M, Zhao S, Xie Z, Segall H, Younes A, Claxton DF, Estrov Z, Kornblau SM, Andreeff M (1999) Apoptosis. Molecules and mechanisms. Adv Exp Med Biol 457:217–236 39- Waring P, Mullbacher A (1999) Cell death induced by the Fas/ Fas ligand pathway and its role in pathology. Immunol Cell Biol 77:312–317 40- Martinou JC, Green DR (2001) Breaking the mitochondrial barrier. Natl Rev Mol Cell Biol 2:63–67 41- Reed JC (1997) Cytochrome c: can’t live with it–can’t live without it. Cell 91:559–562 42- Gupta S (2003) Molecular signaling in death receptor and mitochondrial pathways of apoptosis (Review). Int J Oncol 22:15–20 43- Lopez-Saez JF, de la Torre C, Pincheira J, Gimenez-Martin G (1998) Cell proliferation and cancer. Histol Histopathol 13:1197–1214 44- Cox LS, Lane DP (1995) Tumour suppressors, kinases and clamps: how p53 regulates the cell cycle in response to DNA damage. Bioessays 17:501–508 45- Nasmyth K (1996) Viewpoint: putting the cell cycle in order. Science 274:1643–1645 46- Halliwell B. Reactive oxygen species in living systems: source, biochemistry, and role in human disease. Am J Med 1991; 91: 14S-22S. 47- Kamata H, Hirata H. Redox regulation of cellular signalling. Cell Signal 1999; 11: 1-14. 50 48- Inoue M, Sato EF, Nishikawa M, Park AM, Kira Y, Imada I, Utsumi K. (2003) Mitochondrial generation of reactive oxygen species and its role in aerobic life. Curr Med Chem; 10: 2495-2505. 49- Linnane AW, Eastwood H. (2006) Cellular redox regulation and prooxidant signaling systems: a new perspective on the free radical theory of aging. Ann N Y Acad Sci; 1067: 47-55. 50- Kinnula VL, Crapo JD. (2004) Superoxide dismutases in malignant cells and human tumors. Free Radic Biol Med; 36: 718-744 51- Biswas S, Chida AS, Rahman I. Redox modifications of protein-thiols: emerging roles in cell signaling. Biochem Pharmacol 2006; 71: 551-564. 52-Thannickal VJ, Fanburg BL. Reactive oxygen species in cell signaling. Am J Physiol Lung Cell Mol Physiol 2000; 279: L1005-1028. 53- Schafer FQ, Buettner GR. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic Biol Med 2001; 30: 1191-1212. 54- Lee DH, Rhee JG, Lee YJ. Reactive oxygen species up-regulate p53 and Puma; a possible mechanism for apoptosis during combined treatment with TRAIL and wogonin. Br J Pharmacol 2009; 157: 1189-1202. 55- Sauer H, Wartenberg M, Hescheler J. Reactive oxygen species as intracellular messengers during cell growth and differentiation. Cell Physiol Biochem 2001; 11: 173-186. 56- Martindale JL, Holbrook NJ. Cellular response to oxidative stress: signaling for suicide and survival. J Cell Physiol 2002; 192: 1-15. 57- Chandra J, Samali A, Orrenius S. Triggering and modulation of apoptosis by oxidative stress. Free Radic Biol Med 2000; 29: 323-333. 58- Evan GI, Vousden KH. Proliferation, cell cycle and apoptosis in cancer. Nature 2001; 411: 342-348. 51 59- Vermeulen K, Berneman ZN, Van Bockstaele DR. Cell cycle and apoptosis. Cell Prolif 2003; 36: 165-175. 60- Choi JA, Kim JY, Lee JY, Kang CM, Kwon HJ, Yoo YD, Kim TW, Lee YS, Lee SJ. Induction of cell cycle arrest and apoptosis in human breast cancer cells by quercetin. Int J Oncol 2001; 19: 837-844. 61- Fimognari C, Nusse M, Cesari R, Iori R, Cantelli-Forti G, Hrelia P. Growth inhibition, cellcycle arrest and apoptosis in human T-cell leukemia by the isothiocyanate sulforaphane. Carcinogenesis 2002; 23: 581-586. 62- Engel RH, Evens AM. Oxidative stress and apoptosis: a new treatment paradigm in cancer. Front Biosci 2006; 11: 300-312. 63- Polo SE, Jackson SP. Dynamics of DNA damage response proteins at DNA breaks: a focus on protein modifications. Genes Dev. 2011;25(5):409–433. 64- Goto H, Izawa I, Li P, Inagaki M. Novel regulation of checkpoint kinase 1: is checkpoint kinase 1 a good candidate for anti-cancer therapy? Cancer Sci. 2012;103(7):1195–1200. 65- Cox LS, Lane DP (1995) Tumour suppressors, kinases and clamps: how p53 regulates the cell cycle in response to DNA damage. Bioessays 17:501–508 66- Nasmyth K (1996) Viewpoint: putting the cell cycle in order. Science 274:1643–1645 67- Sherr CJ (1996) Cancer cell cycles. Science 274, 1672. 68- Morgan DO (1995) Principles of CDK regulation. Nature 374, 131. 69- Lew DJ, Kornbluth S (1996) Regulatory roles of cyclin-dependent kinase phosphorylation in cell cycle control. Curr.Opin. Cell Biol. 8, 795. 70- Sherr CJ, Roberts JM (1995) Inhibitors of mammalian G1 cyclin-dependent kinases. Genes Dev. 9, 1149. 71-Carnero A, Hannon GJ (1998) The INK4 family of CDK inhibitors. Curr. Top. Microbiol. Immunol. 227, 43. 52 72-Polyak K, Lee MH, Erdjument BH, Koff A, Roberts JM, Tempst P, Massague J (1994) Cloning of p27Kip1, a cyclin-dependent kinase inhibitor and a potential mediator of extracellular antimitogenic signals. Cell 78, 59. 73- Harper JW, Elledge SJ, Keyomarsi K, Dynlacht B, Tsai LH, Zhang P, Dobrowolski S, Bai C, Connell CL, Swindell E, et al. (1995) Inhibition of cyclin-dependent kinases by p21. Mol. Biol. Cell 6, 387. 74-Lee MH, Reynisdottir I, Massague J (1995) Cloning of p57KIP2, a cyclin-dependent kinase inhibitor with unique domain structure and tissue distribution. Genes Dev. 9, 639. 75- el Deiry WS, Tokino T, Velculescu VE, Levy DB, Parsons R, Trent JM, Lin D, Mercer WE, Kinzler KW, Vogelstein B (1993) WAF1, a potential mediator of p53 tumor suppression. Cell 75, 817. 76- Hannon GJ, Beach D (1994) p15INK4B is a potential effector of TGF- -induced cell cycle arrest. Nature 371, 257. 77-Reynisdottir I, Polyak K, Iavarone Massague J (1995) Kip/cip and INK4 CDK inhibitors cooperate to induce cell cycle arrest in respons to TGF- Genes Dev. 9, 1831. 78- Agarwal ML, Taylor WR, Chernov MV, Chernova OB, Stark GR (1998) The p53 network. J. Biol. Chem. 273, 1. 79- Owen-Schaub LB, Zhang W, Cusack JC, Angelo LS, Santee SM, Fujiwara T, Roth JA, Deisseroth AB, Zhang WW, Kruzel EA (1995) Wild-type human p53 and a temperature-sensitive mutant induce Fas /APO-1 expression. Mol. Cell Biol. 15, 3032. 53 80- Polyak K, Xia Y, Zweier JL, Kinzler KW, Vogelstein B (1997) A model for p53-induced apoptosis. Nature 389,300. 81- Gottlieb TM, Oren M (1998) p53 and apoptosis. Semin. Cancer Biol. 8, 359. 82- Siliciano JD, Canman CE, Taya Y, Sakaguchi K, Appella E, Kastan MB (1997) DNA damage induces phosphorylation of the amino terminus of p53. Genes Dev. 11, 3471. 83- Burma S, Kurimasa A, Xie G, Taya Y, Araki R, Abe M, Crissman HA, Ouyang H, Li GC, Chen DJ (1999) DNA-dependent protein kinase-independent activation of p53 in response to DNA damage. J. Biol. Chem. 274, 17139. 84- Durocher D, Jackson SP (2001) DNA-PK, ATM and ATR as sensors of DNA damage: variations on a theme Curr. Opin. Cell Biol. 13, 225. 85- Hermeking H, Lengauer C, Polyak K, He TC, Zhang L, Thiagalingam S, Kinzler KW, Vogelstein B (1997) 14– -regulated inhibitor of G2/M progression. Mol. Cell 1, 3. 86- Taylor WR, Stark GR (2001) Regulation of the G2/M transition by p53. Oncogene 20, 1803. 87- McDonald ER III, el Deiry WS (2000) Cell cycle control as a basis for cancer drug development. Int. J. Oncol. 16, 871. 88- Azhar Rasul,Muhammad Khan,3 Muhammad Ali,4 Jiang Li, and Xiaomeng Li (2013) Targeting Apoptosis Pathways in Cancer with Alantolactone and IsoalantolactoneScientificWorldJournal. 2013 Oct 27;2013:248532 89- Scorrano, L., Ashiya, M., Buttle, K., Weiler, S., Oakes, S. A., Mannella, C. A., and Korsmeyer, S. J. (2002) Dev. Cell., 2, 55-67. 54 90- Liu, X., Kim, C. N., Yang, J., Jemmerson, R., and Wang, X. (1996) Cell, 86, 147-157. 91- Cain, K., Bratton, S. B., Langlais, C., Walker, G., Brown, D. G., Sun, X. M., and Cohen, G. M. (2000) J. Biol. Chem., 275, 6067-6070. 92- Silke J, Vaux DL. Cell death: Shadow baxing. Curr Biol 1998; 8: 528–531. 93- Korsmeyer SJ,Wie MC, Saito M,Weiler S,OhKJ, Schlesinger PH. Pro-apoptotic cascade activates BID, which oligomerizes BAK or BAX into pores that result in the release of cytochrome c . Cell Death Differ 2000; 7: 1166–1173. 94- Yin X-M. Signal transduction mediated by Bid, a pro-death Bcl-2 family proteins, connects the death receptor and mitochondria apoptosis pathways. Cell Res 2000; 10: 161–167. 95- Duchen MR. Mitochondria and calcium: From cell signaling to cell death. J Physiol 2000; 529: 57–68. 96- McConkey DJ, Orrenius S. The role of calcium in the regulation of apoptosis. J Leukocyte Biol 1996; 59: 775–783. 97- Creagh EM, Martin SJ. Caspases: Cellular demolition experts. Biochem Soc Trans 2001; 29: 696–702. 98- Zou H, Li Y, Liu X, Wang X. An APAF-1・cytochrome c multimeric complex is a functional apoptosome that activates procaspase-9. J Biol Chem 1999; 274: 11549–11556. 99- Acehan D, Jiang X, Morgan DG, Heuser JE,Wang X, Akey CW. Three-dimensional structure of the apoptosome: Implications for assembly, procaspase-9 binding, and activation. Mol Cell 2002; 9: 423–432. 106- Newman DJ, Cragg GM. Natural products as sources of new drugs over the last 25 years. J Nat Prod 2007; 70: 461-477. 107- Heinrich M, Gibbons S. Ethnopharmacology in drug discovery: an analysis of its role and potential contribution. J Pharm Pharmacol 2001; 53: 425-432. 55 108- Abo KA, Adeyemi AA, Adeite DA. Ethnobotanical survey of plants used in the treatment of infertility and sexually transmitted diseases in southwest Nigeria. Afr J Med Med Sci 2000; 29: 325-327. 109- Pinn G. Herbs used in obstetrics and gynaecology. Aust Fam Physician 2001; 30: 351-354, 356. 110- Katsube N, Iwashita K, Tsushida T, Yamaki K, Kobori M. Induction of apoptosis in cancer cells by Bilberry (Vaccinium myrtillus) and the anthocyanins. J Agric Food Chem 2003; 51: 6875. 111- Liu X, Han Y, Peng K, Liu Y, Li J, Liu H. Effect of traditional Chinese medicinal herbs on Candida spp. from patients with HIV/AIDS. Adv Dent Res 2011; 23: 56-60. 112- Amin AR, Kucuk O, Khuri FR, Shin DM. Perspectives for cancer prevention with natural compounds. J Clin Oncol 2009; 27: 2712-2725. 113- Cragg GM, Newman DJ. Plants as a source of anti-cancer agents. J Ethnopharmacol 2005; 100: 72-79. 114- Harvey AL. Natural products in drug discovery. Drug Discov Today 2008; 13: 894-901. 115- Lee MR. The history of Ephedra (ma-huang). J R Coll Physicians Edinb 2011; 41: 78-84. 116- Li W, Shao Y, Hu L, Zhang X, Chen Y, Tong L, Li C, Shen X, Ding J. BM6, a new semisynthetic vinca alkaloid, exhibits its potent in vivo anti-tumor activities via its high binding affinity for tubulin and improved pharmacokinetic profiles. Cancer Biol Ther 2007; 6: 787-794. 117- Huang M, Gao H, Chen Y, Zhu H, Cai Y, Zhang X, Miao Z, Jiang H, Zhang J, Shen H, Lin L, Lu W, Ding J. Chimmitecan, a novel 9-substituted camptothecin, with improved anticancer pharmacologic profiles in vitro and in vivo. Clin Cancer Res 2007; 13: 1298-1307. 118- Lu JJ, Bao JL, Chen XP, Huang M, Wang YT. Alkaloids isolated from natural herbs as the anticancer agents. Evid Based Complement Alternat Med 2012; 2012: 485042. 56 119- Chen J, Zhao H, Wang X, Lee FS, Yang H, Zheng L. Analysis of major alkaloids in Rhizoma coptidis by capillary electrophoresis-electrospray-time of flight mass spectrometry with different background electrolytes. Electrophoresis 2008; 29: 2135-2147. 120- Lau CW, Yao XQ, Chen ZY, Ko WH, Huang Y. Cardiovascular actions of berberine. Cardiovasc Drug Rev 2001; 19: 234-244. 121- Kulkarni SK, Dhir A. Berberine: a plant alkaloid with therapeutic potential for central nervous system disorders. Phytother Res 2010; 24: 317-324. 122- Lai JP, He XW, Jiang Y, Chen F. Preparative separation and determination of matrine from the Chinese medicinal plant Sophora flavescens Ait by molecularly imprinted solid-phase extraction. Anal Bioanal Chem 2003; 375: 264-269. 123- Han Y, Zhang S, Wu J, Yu K, Zhang Y, Yin L, Bi L. Matrine induces apoptosis of human multiple myeloma cells via activation of the mitochondrial pathway. Leuk Lymphoma 2010; 51: 1337-1346. 124- Zheng J, Zheng P, Zhou X, Yan L, Zhou R, Fu XY, Dai GD. Relaxant effects of matrine on aortic smooth muscles of guinea pigs. Biomed Environ Sci 2009; 22: 327-332. 125- Szallasi A. Piperine: researchers discover new flavor in an ancient spice. Trends Pharmacol Sci 2005; 26: 437-439. 126- Bae GS, Kim MS, Jung WS, Seo SW, Yun SW, Kim SG, Park RK, Kim EC, Song HJ, Park SJ. Inhibition of lipopolysaccharide-induced inflammatory responses by piperine. Eur J Pharmacol 2010; 642: 154-162. 127- Srinivasan K. Black pepper and its pungent principle-piperine: a review of diverse physiological effects. Crit Rev Food Sci Nutr 2007; 47: 735-748. 128- Mahady GB, Beecher CW. Quercetin-induced benzophenanthridine alkaloid production in suspension cell cultures of Sanguinaria canadensis. Planta Med 1994; 60: 553-557. 57 129- Vavreckova C, Gawlik I, Muller K. Benzophenanthridine alkaloids of Chelidonium majus; I. Inhibition of 5- and 12-lipoxygenase by a non-redox mechanism. Planta Med 1996; 62: 397401. 130- Beuria TK, Santra MK, Panda D. Sanguinarine blocks cytokinesis in bacteria by inhibiting FtsZ assembly and bundling. Biochemistry 2005; 44: 16584-16593. 131- Lenfeld J, Kroutil M, Marsalek E, Slavik J, Preininger V, Simanek V. Antiinflammatory activity of quaternary benzophenanthridine alkaloids from Chelidonium majus. Planta Med 1981; 43: 161-165. 132- Li DG, Wang ZR, Lu HM. Pharmacology of tetrandrine and its therapeutic use in digestive diseases. World J Gastroenterol 2001; 7: 627-629. 133- Lee SH, Son JK, Jeong BS, Jeong TC, Chang HW, Lee ES, Jahng Y. Progress in the studies on rutaecarpine. Molecules 2008; 13: 272-300. 134- Denicourt C, Dowdy SF. Medicine. Targeting apoptotic pathways in cancer cells. Science 2004; 305: 1411-1413. 135- Zhang Q, Cai D, Liu J. Matrix solid-phase dispersion extraction coupled with HPLC-diode array detection method for the analysis of sesquiterpene lactones in root of Saussurea lappa C.B.Clarke. J Chromatogr B Analyt Technol Biomed Life Sci 2011; 879: 2809-2814. 136- Park HW, Lee JH, Choi SU, Baek NI, Kim SH, Yang JH, Kim DK. Cytotoxic germacranolide sesquiterpenes from the bark of Magnolia kobus. Arch Pharm Res 2010; 33: 7174. 137- Yang NY, Duan JA, Shang EX, Tian LJ. (2010)Analysis of sesquiterpene lactones in Eupatorium lindleyanum by HPLC-PDA-ESI-MS/MS. Phytochem Anal; 21: 144-149. 138- Kassuya CA, Cremoneze A, Barros LF, Simas AS, Lapa Fda R, Mello-Silva R, Stefanello ME, Zampronio AR. Antipyretic and anti-inflammatory properties of the ethanolic extract, dichloromethane fraction and costunolide from Magnolia ovata (Magnoliaceae). J Ethnopharmacol 2009; 124: 369-376. 58 139- De Marino S, Borbone N, Zollo F, Ianaro A, Di Meglio P, Iorizzi M. New sesquiterpene lactones from Laurus nobilis leaves as inhibitors of nitric oxide production. Planta Med 2005; 71: 706-710. 140- Ferrari B, Castilho P, Tomi F, Rodrigues AI, do Ceu Costa M, Casanova J. Direct identification and quantitative determination of costunolide and dehydrocostuslactone in the fixed oil of Laurus novocanariensis by 13C-NMR spectroscopy. Phytochem Anal 2005; 16: 104107. 141- Mata R, Rivero-Cruz I, Rivero-Cruz B, Bye R, Timmermann BN. Sesquiterpene lactones and phenylpropanoids from Cosmos pringlei. J Nat Prod 2002; 65: 1030-1032. 142- Wu SH, Luo XD, Ma YB, Hao XJ, Zhou J, Wu DG. Two new germacranolides from Magnolia grandiflora. J Asian Nat Prod Res 2001; 3: 95-102. 143- Park SH, Choi SU, Lee CO, Yoo SE, Yoon SK, Kim YK, Ryu SY. Costunolide, a sesquiterpene from the stem bark of Magnolia sieboldii, inhibits the RAS-farnesylproteintransferase. Planta Med 2001; 67: 358-359. 144- Song XK, Tu PF, Wu LJ, Cai Y, Zhu H, Lu Y, Liu XY, Zheng QT. A new sesquiterpene lactone from Tsoongiodendron odorum Chun. J Asian Nat Prod Res 2001; 3: 285-291. 145- Castro V, Murillo R, Klaas CA, Meunier C, Mora G, Pahl HL, Merfort I. Inhibition of the transcription factor NF-kappa B by sesquiterpene lactones from Podachaenium eminens. Planta Med 2000; 66: 591-595. 146- Mondranondra IO, Che CT, Rimando AM, Vajrodaya S, Fong HH, Farnsworth NR. Sesquiterpene lactones and other constituents from a cytotoxic extract of Michelia floribunda. Pharm Res 1990; 7: 1269-1272. 147- Bocca C, Gabriel L, Bozzo F, Miglietta A. A sesquiterpene lactone, costunolide, interacts with microtubule protein and inhibits the growth of MCF-7 cells. Chem Biol Interact 2004; 147: 79-86. 59 148- Park HJ, Jung WT, Basnet P, Kadota S, Namba T. Syringin 4-O-beta-glucoside, a new phenylpropanoid glycoside, and costunolide, a nitric oxide synthase inhibitor, from the stem bark of Magnolia sieboldii. J Nat Prod 1996; 59: 1128-1130. 149- Pandey MM, Rastogi S, Rawat AK. Saussurea costus: botanical, chemical and pharmacological review of an ayurvedic medicinal plant. J Ethnopharmacol 2007; 110: 379-390. 150- Kang JS, Yoon YD, Lee KH, Park SK, Kim HM. Costunolide inhibits interleukin-1beta expression by down-regulation of AP-1 and MAPK activity in LPS-stimulated RAW 264.7 cells. Biochem Biophys Res Commun 2004; 313: 171-177. 151- Wedge DE, Galindo JC, Macias FA. Fungicidal activity of natural and synthetic sesquiterpene lactone analogs. Phytochemistry 2000; 53: 747-757. 152- Park HJ, Kwon SH, Han YN, Choi JW, Miyamoto K, Lee SH, Lee KT. Apoptosis-Inducing costunolide and a novel acyclic monoterpene from the stem bark of Magnolia sieboldii. Arch Pharm Res 2001; 24: 342-348. 153- Sun CM, Syu WJ, Don MJ, Lu JJ, Lee GH. Cytotoxic sesquiterpene lactones from the root of Saussurea lappa. J Nat Prod 2003; 66: 1175-1180. 154- Luna-Herrera J, Costa MC, Gonzalez HG, Rodrigues AI, Castilho PC. Synergistic antimycobacterial activities of sesquiterpene lactones from Laurus spp. J Antimicrob Chemother 2007; 59: 548-552. 155- Eliza J, Daisy P, Ignacimuthu S, Duraipandiyan V. Normo-glycemic and hypolipidemic effect of costunolide isolated from Costus speciosus (Koen ex. Retz.)Sm. in streptozotocininduced diabetic rats. Chem Biol Interact 2009; 179: 329-334. 156- Choi JY, Choi EH, Jung HW, Oh JS, Lee WH, Lee JG, Son JK, Kim Y, Lee SH. Melanogenesis inhibitory compounds from Saussureae Radix. Arch Pharm Res 2008; 31: 294299. 157- Ohnishi M, Yoshimi N, Kawamori T, Ino N, Hirose Y, Tanaka T, Yamahara J, Miyata H, Mori H. Inhibitory effects of dietary protocatechuic acid and costunolide on 7,1260 dimethylbenz[a]anthracene-induced hamster cheek pouch carcinogenesis. Jpn J Cancer Res 1997; 88: 111-119. 158- Choi JH, Ha J, Park JH, Lee JY, Lee YS, Park HJ, Choi JW, Masuda Y, Nakaya K, Lee KT. Costunolide triggers apoptosis in human leukemia U937 cells by depleting intracellular thiols. Jpn J Cancer Res 2002; 93: 1327-1333. 159- Mori H, Kawamori T, Tanaka T, Ohnishi M, Yamahara J. Chemopreventive effect of costunolide, a constituent of oriental medicine, on azoxymethane-induced intestinal carcinogenesis in rats. Cancer Lett 1994; 83: 171-175. 160- Chen CN, Huang HH, Wu CL, Lin CP, Hsu JT, Hsieh HP, Chuang SE, Lai GM. Isocostunolide, a sesquiterpene lactone, induces mitochondrial membrane depolarization and caspase-dependent apoptosis in human melanoma cells. Cancer Lett 2007; 246: 237-252. 161- Choi SH, Im E, Kang HK, Lee JH, Kwak HS, Bae YT, Park HJ, Kim ND. Inhibitory effects of costunolide on the telomerase activity in human breast carcinoma cells. Cancer Lett 2005; 227: 153-162. 162- Choi JH, Ha J, Park JH, Lee JY, Lee YS, Park HJ, Choi JW, Masuda Y, Nakaya K, Lee KT. Costunolide triggers apoptosis in human leukemia U937 cells by depleting intracellular thiols. Jpn J Cancer Res 2002; 93: 1327-1333. 163- H. L. Jae, N.-I. Baek, S.-H. Kim et al., “A new cytotoxic prenylated chalcone from Sophora flavescens,” Archives of Pharmacal Research, vol. 30, no. 4, pp. 408–411, 2007. 164- A. E. Vercesi, A. J. Kowaltowski, M. T. Grijalba, A. R.Meinicke, and R. F. Castilho, “The role of reactive oxygen species in mitochondrial permeability transition,” Bioscience Reports, vol. 17, no. 1, pp. 43–52, 1997. 165- Lee MG, Lee KT, Chi SG, Park JH. Costunolide induces apoptosis by ROS-mediated mitochondrial permeability transition and cytochrome C release. Biol Pharm Bull 2001; 24: 303306. 61 166- J. M. Adams, “Ways of dying: multiple pathways to apoptosis,” Genes and Development, vol. 17, no. 20, pp. 2481–2495, 2003. 167- A. G. Porter and R. U. Janicke, “Emerging roles of caspase-3 in apoptosis,” Cell Death and Differentiation, vol. 6, no. 2, pp. 99– 104, 1999 168-Nadine Kretschmer1, Beate Rinner2, Nicole Stuendl3, Heike Kaltenegger3, Elisabeth Wolf 3, Olaf Kunert4, Herbert Boechzelt5, Andreas Leithner3, Rudolf Bauer1, Birgit Lohberger3(2012) Effect of Costunolide and Dehydrocostus Lactone on Cell Cycle, Apoptosis, and ABC Transporter Expression in Human Soft Tissue Sarcoma Cells Planta Med 2012; 78(16): 17491756 169-Rasul, A.; Bao, R.; Malhi, M.; Zhao, B.; Tsuji, I.; Li, J.; Li, X. Induction of Apoptosis by Costunolide in Bladder Cancer Cells is Mediated through ROS Generation and Mitochondrial Dysfunction. Molecules 2013, 18, 1418-1433 170- Dirsch VM, Stuppner H, Vollmar AM. Cytotoxic sesquiterpene lactones mediate their death-inducing effect in leukemia T cells by triggering apoptosis. Planta Med 2001; 67: 557-559. 171- Whipple RA, Vitolo MI, Boggs AE, Charpentier MS, Thompson K, Martin SS.(2013) Parthenolide and costunolide reduce microtentacles and tumor cell attachment by selectively targeting detyrosinated tubulin independent from NF-κB inhibition. Breast Cancer Res 2013;15(5):R83. 172- Choi YK1, Cho SG, Woo SM, Yun YJ, Jo J, Kim W, Shin YC, Ko SG(2013). Saussurea lappa Clarke-Derived Costunolide Prevents TNF α -Induced Breast Cancer Cell Migration and Invasion by Inhibiting NF- κ B Activity . Evid Based Complement Alternat Med.. 2013;2013:936257. 173- Peng ZX1, Wang Y, Gu X, Wen YY, Yan C...( 2013) A platform for fast screening potential anti-breast cancer compounds in traditional Chinese medicines. Biomed Chromatog. 2013 Dec;27(12):1759-66 174- Konishi T, Shimada Y, Nagao T, Okabe H, Konoshima T. Antiproliferative sesquiterpene lactones from the roots of Inula helenium. Biol Pharm Bull 2002; 25: 1370-1372. 62 175- Pal HC, Sehar I, Bhushan S, Gupta BD, Saxena AK. Activation of caspases and poly (ADPribose) polymerase cleavage to induce apoptosis in leukemia HL-60 cells by Inula racemosa. Toxicol In Vitro 2010; 24: 1599-1609. 176- Choi YK, Seo HS, Choi HS, Choi HS, Kim SR, Shin YC, Ko SG. .(2012) Induction of Fasmediated extrinsic apoptosis, p21WAF1-related G2/M cell cycle arrest and ROS generation by costunolide in estrogen receptor-negative breast cancer cells, MDA-MB-2 Mol Cell Biochem. 2012 Apr;363(1-2):119-28. 177- Choi SH , Im E, Kang HK, Lee JH, Kwak HS, Bae YT, Park HJ, Kim ND. (2005) Inhibitory effects of costunolide on the telomerase activity in human breast carcinoma cells. Cancer Lett. 2005 Sep 28;227(2):153-62. 178- Bocca C, Gabriel L, Bozzo F, Miglietta A. (2004) A sesquiterpene lactone, costunolide, interacts with microtubule protein and inhibits the growth of MCF-7 cells.Chem Biol Interact. 2004 Jan 15;147(1):79-86. 179- Hartwell and Weinert, nL.H. Hartwell, T.A. (1989) Weiner Checkpoints: controls that ensure the order of cell cycle events Science, 246 (1989), pp. 629–63 14. Altekruse SF KC, Krapcho 180- Pietenpol JA1, Stewart ZA.(2002) Cell cycle checkpoint signaling: cell cycle arrest versus apoptosis. Toxicology. 2002 Dec 27;181-182:475-81. 181- Hsu JL, Pan SL, Ho YF, Hwang TL, Kung FL, Guh JH. Costunolide induces apoptosis through nuclear calcium2+ overload and DNA damage response in human prostate cancer. J Urol; 185: 1967-1974. 182- Rasul AY, B. Yang, L. Arshad, M. Khan, M. Ma, T. Yang, H. Costunolide, a sesquiterpene lactone induces G2/M phase arrest and mitochondria-mediated apoptosis in human gastric adenocarcinoma SGC-7901 cells. J. Med. Plant. Res. 2012; 6: 1191-1200. 183- Lohberger B1, Rinner B, Stuendl N, Kaltenegger H, Steinecker-Frohnwieser B, Bernhart E, Bonyadi Rad E, Weinberg AM, Leithner A, Bauer R, Kretschmer N.(2013) Sesquiterpene 63 lactones downregulate G2/M cell cycle regulator proteins and affect the invasive potential of human soft tissue sarcoma cells. PLoS One. 2013 Jun 14;8(6):e66300. 184- Whipple RA, Vitolo MI, Boggs AE, Charpentier MS, Thompson K, Martin SS.(2013) Parthenolide and costunolide reduce microtentacles and tumor cell attachment by selectively targeting detyrosinated tubulin independent from NF-κB inhibition. Breast Cancer Res 2013;15(5):R83. 185- Leist M, Jaattela M. Four deaths and a funeral: from caspases to alternative mechanisms. Nat Rev Mol Cell Biol 2001; 2: 589-598. 186- Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol 2007; 35: 495 516. 187- Mashima T, Tsuruo T. Defects of the apoptotic pathway as therapeutic target against cancer. Drug Resist Updat 2005; 8: 339-343. 188- Pommier Y, Sordet O, Antony S, Hayward RL, Kohn KW. Apoptosis defects and chemotherapy resistance: molecular interaction maps and networks. Oncogene 2004; 23: 29342949. 189- Fulda S. Evasion of apoptosis as a cellular stress response in cancer. Int J Cell Biol 2010; 2010: 370835. 190-. Lawen A. Apoptosis-an introduction. Bioessays 2003; 25: 888-896. 191- Reed JC. Apoptosis-based therapies. Nat Rev Drug Discov 2002; 1: 111-121. 192- Buttke TM, Sandstrom PA. Oxidative stress as a mediator of apoptosis. Immunol Today 1994; 15: 7-10. 193- Davis W, Jr., Ronai Z, Tew KD. Cellular thiols and reactive oxygen species in druginduced apoptosis. J Pharmacol Exp Ther 2001; 296: 1-6. 194- Hampton MB, Orrenius S. Redox regulation of apoptotic cell death. Biofactors 1998; 8: 15. 64 195- Sandau KB, Brune B. Up-regulation of Bcl-2 by redox signals in glomerular mesangial cells. Cell Death Differ 2000; 7: 118-125. 196- Lee MG, Lee KT, Chi SG, Park JH. Costunolide induces apoptosis by ROS-mediated mitochondrial permeability transition and cytochrome C release. Biol Pharm Bull 2001; 24: 303306. 197- Robles M, Aregullin M, West J, Rodriguez E. Recent studies on the zoopharmacognosy, pharmacology and neurotoxicology of sesquiterpene lactones. Planta Med 1995; 61: 199-203. 198- Okugawa H, Ueda R, Matsumoto K, Kawanishi K, Kato A. Effect of jinkoh-eremol and agarospirol from agarwood on the central nervous system in mice. Planta Med 1996; 62: 2-6. 199- Yang YI, Kim JH, Lee KT, Choi JH. Costunolide induces apoptosis in platinum-resistant human ovarian cancer cells by generating reactive oxygen species. Gynecol Oncol 2011; 123: 588-596. 200- Jui-Ling Hsu, Shiow-Lin Pan, Yunn-Fang Ho, Tsong-Long Hwang, Fan-Lu Kung, JihHwa Guh(2011) Costunolide Induces Apoptosis Through Nuclear Calcium2+ Overload and DNA Damage Response in Human Prostate CancerThe Journal of UrologybVolume 185, Issue 5, May 2011, Pages 1967–1974 201- S.H. Kim, S.N. Kang, H.J. Kim, T.S. Kim (2002) Potentiation of 1, 25-dihydroxyvitamin D(3)-induced differentiation of human promyelocytic leukemia cells into monocytes by costunolide, a germacranolide sesquiterpene lactone Biochem. Pharmacol., 64 (2002), pp. 1233– 1242 202- Chia-Yuan Liu, Hsun-Shuo Chang, Ih-Sheng Chen,Chih-Jen Chen, Ming-Ling Hsu, ShuLing Fu, and Yu-Jen Chen(2011) Costunolide causes mitotic arrest and enhances radiosensitivity in human hepatocellular carcinoma cells Radiat Oncol. 2011; 6: 56. 203- Park HJ, Kwon SH, Han YN, Choi JW, Miyamoto K, Lee SH, Lee KT. Apoptosis-Inducing costunolide and a novel acyclic monoterpene from the stem bark of Magnolia sieboldii. Arch Pharm Res 2001; 24: 342-348. 65 204-Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol 2007; 35: 495-516 205-Buytaert E, Dewaele M, Agostinis P. Molecular effectors of multiple cell death pathways initiated by photodynamic therapy. Biochim Biophys Acta 2007; 1776: 86-107. 206- Wang X. The expanding role of mitochondria in apoptosis. Genes Dev 2001; 15: 29222933. ACKNOWLEDGEMENTS It is an honor to express my sincerest thanks to my very kind scholarly and compassionate supervisor Dr. Xiaomeng Li, Professor, The Key Laboratory of Molecular Epigenetic of MOE, Institute of Genetics and Cytology, School of Life Sciences, Northeast Normal University, for her precious encouragement, priceless suggestions, instructions and competent guidance without this my project would not be materialized. I am also broadening my thanks to my Co-supervisor Dr. Azhar Rasul for his guidance and support throughout the project, without of that this work would not have been brought to end. I am also grateful to express our sincere thanks to my all the lab-fellows and teachers from Northeast Normal University for their honorable assistance. I'd like to thank China Scholarship Council (CSC) and Ministry of Education of CApe verde. for financial support. 66 In the last but not least, I am also very thankful to my parents, mainly my Mother Josefa Lopes, my brothers Osvaldina, Edelson, Oteldino, Alexandra Milco and Tamires and all other family members due to their full support, encouragement and blessing during the study period. HELDER OLIVEIRA 67