Human Evolution
advertisement

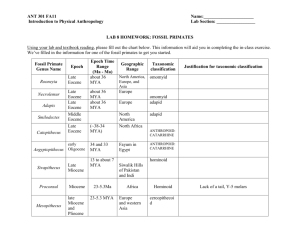
March 12, 2010 Bioe 109 Winter 2010 Lecture 26 Human evolution Hominoid phylogeny - the past 40 years have seen a dramatic re-appraisal of the age of the human species. - in the 1950’s and 1960’s, physical anthropologists had estimated the separation of the human lineage from the other ape groups at anywhere from 20 to 30 million years ago (mya). - in the late 1960’s Allan Wilson’s lab at Berkeley estimated, on the basis of immunological distance, that the human lineage split from the chimp lineage at about 5 mya. - this conclusion was highly controversial at its time - many anthropologists simply refused to accept the molecular evidence. - it is remarkable how well this initial estimate has stood the test of time - many additional markers and techniques corroborated this date of about 5-6 my. - despite the general consensus of when this split occurred, the precise structure of the tree linking humans with chimpanzees and apes has been extremely controversial. - gibbons and orangutans are clearly sister groups to humans, chimps and gorillas. - the two competing trees have humans and chimps more closely related in one, and chimps and gorillas more closely related in the other. - the first mtDNA analyses carried out in the early 1980’s by Alan Templeton was interpreted as supporting the grouping of chimps and gorillas. - however, more recent data based on nucleotide sequences and DNA-DNA hybridization has supported the grouping of humans and chimps. - a cladogram of the group has gibbons diverging earliest (15-20 mya) followed by orangutan (13 mya), gorillas (6-8 mya), and humans (~ 5 mya). - the two chimpanzee species (the bonobo, or pygmy chimp, and the common chimp) split at about 2 mya. Comparison of the human and chimpanzee genomes - the chimpanzee genome (published September 1, 2005) was obtained from a single individual called “Clint”. - Clint was born in captivity and belongs to the West African subspecies Pan troglodytes verus. - the genome data confirms what we already knew – humans and chimps are extremely similar genetically. - the genome-wide nucleotide divergence between humans and chimps is 1.23% - correcting for intra-species polymorphism, the fixed divergence rate is about 1.06%. - humans and chimps differ by 1 chromosomal fusion and at least 9 pericentric inversions. - human chromosome 2 represents a fusion of two ancestral chromosomes that have remained separate in chimpanzees. - the precise fusion point has been mapped. - 13,454 human and chimp genes were assembled that had unambiguous homology. - overall, 29% of all proteins compared were identical. - the median numbers of synonymous and nonsynonymous substitutions are 3 and 2, respectively. - a total of 585 genes (4.4%) show ka/ks ratios > 1 suggesting positive selection. - however, given the low divergence between most genes there is a large stochastic variance of this ratio. - are there genes that have experienced rapid evolution in the human lineage? - the single strongest outliers are transcription factors (including homeotic, forkhead, etc.) that play key roles in early development. - this needs further work since the numbers of genes is small. - in contrast, there are no excess of accelerated genes in the chimpanzee lineage. The hominid fossil record - the primate fossil record is very poor. - primates first appear in the fossil record in the late Cretaceous about 70 mya. - the first fossil fragment of an anthropoid ape (the lineage which will produce apes and humans) dates to about 50 my in Algeria. - in May, 2009 scientists reported the most spectacular primate fossil that has ever been found. - it came from the Messel deposit in Germany, a site well known for producing a number of spectacular fossils. - it was named in honor of Darwin’s 200th birthday as Darwinius masillae and given the nickname “Ida”. - she was a small, nocturnal anthropoid about 6-9 months in age. - she was almost it was almost mature and her adult teeth were being formed. - she is clearly not ancestor of lemurs because she lacks a tooth comb and a grooming claw on her second toe. - the preservation is so good that we can see her fur and the remnants of her last meal (some fruit). - fossils of primitive anthropoid apes have recently been identified in Egypt about 30 mya (called Aegyptopithecus). - additional fossils appear in Kenya about 25 mya. - there is another gap of about 10 my - at 15 mya there are several species of apes roaming Africa and Asia. - these are small species, equivalent in size to a 7 year-old human boy. - fossils corresponding to the lineage that ultimately produced humans are largely absent from the period of 5 to 14 mya in Africa. - the following sequence of fossils may trace our ancestry over the past 5 million years in Africa: 1. Ardipethicus ramidus at about 4.4 to 4.2 mya 2. Australopithecus anamensis at 4.2 to 3.9 mya 3. Australopithecus afarensis at 3.9 to 3.0 mya (“Lucy”) 4. Australopithecus africanus at 2.8 to 2.4 mya 5. Homo habilis at 2.5 to 1.6 mya. 6. Homo ergaster at 1.8 to 1.5 mya 7. Homo erectus at 1.2 to 0.4 mya 8. Homo heidelbergensis at 0.6 to 0.2 mya 9. Homo sapiens at 0.15 mya to present - there are three points to make about this succession of fossils 1. the exact path of descent is unclear - many new fossils are being discovered and more are likely to follow. 2. our evolution was largely continuous - there is no sudden “jumps” in morphology. -cranial capacity increased gradually from the 600-800 cc (in H. habilis) to the 1200 – 1400 cc range observed in anatomically modern humans. 3. many hominid species coexisted over this entire period. - not included in the list above are the “robust” Australopithecines that were present in Africa about 3 mya. Ardipethicus ramidus - on Oct. 2, 2009 a series of papers appeared in the journal Science describing detailed study of Ardipethicus ramidus. - the initial discovery of A. ramidus was made by Tim White in 1994, but it took 15 years to before these findings finally appeared in print. - White’s group collected fossils from an additional 35 A. ramidus individuals to complete their description of the species. - their results have forced a complete reappraisal of early hominid evolution. - up to now, the last common ancestor of the human and chimpanzee lineages was assumed to be intermediate between Homo and Pan. - this is obviously incorrect – the two extant chimpanzee have undergone extensive evolution that are unique to those lineages. - in fact, one could not have predicted the unusual mix of primitive and derived characters observed in A. ramidus. - it had a much smaller face than chimps but a similar cranial capacity (300 -350 cm3). - its hands were not adapted to knuckle walking or suspending from tree limbs. - the wrists and fingers were extremely flexible, allowing for careful grasping of tree branches. - its feet had divergent big toes like chimps but much more rigid, suggesting adaptations to walking upright. - its pelvis shows that A. ramidus was at home both on the ground and in trees, but it was a “careful climber” not a jumper. - its canines are much less developed than in chimps and are similar in both sexes. - this suggests early changes in social behavior typified by less aggression among males. - females apparently had concealed ovulation, a trait believed to have evolved much more recently. - it is unclear still if A. ramidus was the ancestor of Australopithecus but it does not possess any traits that would exclude it from this history. - it is still unclear how many Ardipethicus species lived in the window between 4.5 and 6 mya (two are currently described). - it is possible that A. ramidus represents an extinct side branch of the lineage eventually giving rise to Homo, but more fossils need to be discovered. The evolution of Neanderthals - H. sapiens neanderthalensis lived in Western Europe between 400,000 and 30,000 ya. - Neanderthals inhabited southern Britain, northern Germany, Kiev and as far east as the Caspian Sea. - their average height was about 5’4’’ but they would have weighed about 20 lbs more than H. sapiens, due to extra muscle. - although their brain size was about 10% larger than H. sapiens, there are many things that Neanderthals lacked. - they left no art objects, they lacked needles and any evidence of sewing, they lacked boats, and they did not practice overland trade as did H. sapiens. - they did not develop any increased sophistication in their tools over the period of 125,000 to 40,000 quite unlike modern H. sapiens. - in 1997, Neanderthal mtDNA (the control region, or “d-loop”) has been successfully amplified and sequenced. - the source of DNA was actually from the “type” Neanderthal skeleton first discovered in 1856 in Germany. - over the past 9 years, an additional 11 mtDNA sequences have been obtained from bones collected throughout western Europe. - all sequences place the Neanderthal lineage outside of present-day humans. - this suggests that no Neanderthal mtDNA has introgressed into humans. - what about nuclear genes? - in 2006, two articles appeared in which significantly more Neanderthal DNA had been sequenced (1 million base pairs in the Nature article) - from this data, it is still unclear whether there is any evidence for Neanderthal sequences in contemporary human populations. - however, there is a hint of the opposite (i.e., human introgression into Neanderthals) based on a higher divergence of the Neanderthal X chromosome compared to other autosomes. - there is a project ongoing at the moment to sequence the entire Neanderthal genome. - from the data collected to date, the Neanderthal lineage diverged from the modern human lineage about 600,000-800,00 years ago (which is very similar to that provided by the earlier mtDNA studies). - similar to modern humans, the effective population size estimated for the Neanderthal lineage is very small (about 3,000 compared to about 13,000 for us). - when used as an outgroup to root the human mtDNA genealogy, the Neanderthal sequence shows African sequences to be ancestral, thus confirming yet again that Africa was the likely origin of modern humans. The origin of modern Homo sapiens - three models have been proposed to account for the evolution of modern humans. 1. the African replacement (AR) model. - this model proposes that two radiations of humans occurred out of Africa. - the first took place about 1.8 mya when H. erectus appears throughout the far east and China. - the AR model proposes very recently (within the last 200,000 years) a second wave of modern humans emerged from Africa and displaced previously existing humans throughout the globe. 2. the hybridization/assimilation (HA) model. - the HA model also proposes two waves of human migration out of Africa. - unlike the African replacement model, it proposes that there was some hybridization between the modern and archaic forms. - therefore, existing human populations are predicted to be some mixture of recent and ancient alleles. 3. the multiregional (MR) model. - this model assumes that modern humans emerging from Africa hybridized with pre-existing populations throughout Europe and Asia. - various modifications of this model have been proposed that differ on the extent of migration, selection and hybridization among regions. - the AR model makes three predictions that can be tested with molecular data: 1. An African ancestry should exist for all “modern” alleles (i.e., those sampled from existing populations). 2. Modern populations should be recently formed (less than 200,000 years). 3. The greatest allelic diversity should exist in Africa. - all three predictions have been met for a variety of genes (mitochondrial DNA, Y chromosomes, and other nuclear genes). - in the original work on “mitochondrial eve”, the ancestor of all modern mtDNAs was placed in Africa between 140,000 and 280,000 years ago. - more sophisticated work accomplished after this initial study has been able to date the origin of modern humans much more precisely. - the date has been confirmed with higher precision to be about 150,000 years. - more recent work has focused on the Y chromosome which is the male counterpart to mtDNA. - it is inherited down paternal lines and should reveal the appearance of Y chromosome Adam. - perhaps surprisingly, the Y chromosome data closely matches the mtDNA data. - the origin of all Y chromosomes can be traced again to Africa (having a common ancestor about 60,000 years ago) and to have largely replaced previously existing Y chromosomes. - recent studies have suggested that Y chromosomes were reintroduced back into Africa from Asia. - however, there was not a replacement of previously existing Y chromosomes - this in part explains the unusually high levels of Y chromosome polymorphism in Africa. - the “Eve” hypothesis confuses gene genealogies and individual genealogies. - gene genealogies gradually coalesce towards a unique DNA sequence - individual genealogies increase by a factor of two each generation. - the observation of a single mitochondrial lineage existing in Africa 150,000 years ago does not mean that only one female lived at that time! - if we assume 20 years for a human generation time throughout the Pleistocene period, we can estimate that the number of females present at the root of this tree at about 4,000-5,000. Recent evidence refuting the African Replacement hypothesis - in 2005, Alan Templeton reconstructed the history of human populations by analyzing 25 gene trees. - his analysis rejects the complete replacement hypothesis with a probability equal to10-17! - Templeton’s analysis detected three successive waves of migration out of Africa. - the first occurred about 1.7 MYA by the expansion of Homo erectus out of Africa into Europe and Asia. - a second wave was detected about 700,000 years ago. - interestingly, this corresponds to a period of high rainfall in eastern Africa that converted much of the Sahara Desert to savannah. - the third wave occurred 150,000 years ago. - unlike previous analyses, Templeton claims to have detected the introgression of African genes into European and Asian populations that significantly predate the most recent “replacement”. - if true, these results completely undermine the African replacement model. - many more genes will need to be examined from populations throughout the world before it becomes widely accepted. The discovery of Homo floresiensis - in the fall of 2004, one of the most exciting discoveries in paleoanthropology in the past 50 years was published – the discovery of Homo floresiensis. - the skeleton of a small (1 m) skeleton of an adult females (called LB1) was discovered on the island of Flores – an island that lies to the east of Java in Indonesia. - primitive stone tools had been previously found on Flores in 1998 that suggested that Homo erectus had colonized the island (which is unusual because a major ocean barrier had to have been overcome). - one of the many startling features about this skeleton was that it dates to only ~18,000 years ago! - unlike pygmies (that mature at a similar size but have normal cranial capacities) the cranial capacity of LB1 was only 380 cc – this is similar to the size of australopithecines from Africa 2 to 3 mya! - however, LB1 is clearly not an australopithecine as she has cranial and mandibular characteristics that clearly indicate she belongs to the genus Homo. - this observation suggests that the proximate causes leading to dwarfism are not the same as those in pygmies (i.e., reduced levels of and/or sensitivity to insulin-like growth factor 1). - she has an interesting mix of primitive, unique and derived features suggesting that she is a descendant of H. erectus that evolved into an insular dwarf species. - the same tendency towards the evolution of dwarfism on islands is common in large mammals – some of the best known involve dwarf elephants on many islands in the Mediterranean. - the selective pressures that may favor dwarfism include reduced interspecific competition, a reduced resource base, and the absence of predators. - the discovery of H. floresiensis suggests that ancestral humans have been much more adaptable to local conditions than previously believed. - there was a large predator on Flores, however, the Komodo dragon!