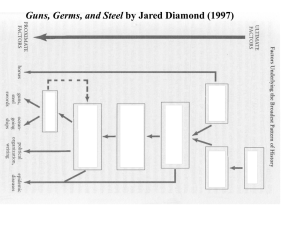
Cattle Before Crops - Department of Anthropology
advertisement

P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 C 2002) Journal of World Prehistory, Vol. 16, No. 2, June 2002 (° Cattle Before Crops: The Beginnings of Food Production in Africa Fiona Marshall1,2 and Elisabeth Hildebrand1 In many areas of the world, current theories for agricultural origins emphasize yield as a major concern during intensification. In Africa, however, the need for scheduled consumption shaped the development of food production. African cattle were domesticated during the tenth millennium BP by delayedreturn Saharan hunter-gatherers in unstable, marginal environments where predictable access to resources was a more significant problem than absolute abundance. Pastoralism spread patchily across the continent according to regional variations in the relative predictability of herding versus hunting and gathering. Domestication of African plants was late (after 4000 BP) because of the high mobility of herders, and risk associated with cultivation in arid environments. Renewed attention to predictability may contribute to understanding the circumstances that led to domestication in other regions of the world. KEY WORDS: archaeology; Africa; predictability; cattle domestication. INTRODUCTION It is generally agreed that pathways to food production have varied greatly from place to place around the world (reviews in Cowan and Watson, 1992; Diamond, 1997; Gebauer and Price, 1992; Harris, 1996a; Harris and Hillman, 1989; Piperno and Pearsall, 1998; Price and Gebauer, 1995; Smith, 1998). Agriculture appears to have arisen recently (<10,000 years ago) and 1 Department of Anthropology, Washington University in St. Louis, St. Louis, MO. whom correspondence should be addressed at Department of Anthropology, Campus Box 1114, Washington University in St. Louis, One Brookings Drive, St. Louis, MO 63130; e-mail: fmarshal@artsci.wustl.edu. 2 To 99 C 2002 Plenum Publishing Corporation 0892-7537/02/0600-0099/0 ° P1: GLQ Journal of World Prehistory [jowo] 100 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand rarely, in only a few independent transitions (Harris, 1996b; Smith, 1998). The profound, long-lasting effects of agriculture on human societies include higher population densities and more urban-based, stratified social systems. Today’s global distribution of wealth and political power may reflect variation in the characteristics and timing of early food production (Diamond, 1997), but the precise nature of, and reasons for, this variation are not yet well understood. Some of the most intensively studied regions where indigenous food producing economies developed include Southwest Asia, Mesoamerica, and Eastern North America. Recent syntheses drawing on data from some of these areas emphasize post-Pleistocene climatic change, the domestication of plants before animals, and the role of settled hunter-gatherers in the development of early food production (Bar-Yosef, 1998; Harris, 1996b,c; Price and Gebauer, 1995; but see Piperno and Pearsall, 1998). They also note that domestication occurred in well-watered places with relatively abundant resources, rather than in marginal settings (Price and Gebauer, 1995; Smith, 1998). For some regions, scholars point to centers from which entire crop complexes spread through colonization (Harris, 1996b,c). Elsewhere, more attention is given to diffusion and the involvement of local hunter-gatherers in the spread of food production (Price and Gebauer, 1995). Africa is less well known than the regions from which most syntheses have been drawn, but research published in the last decade has clarified African data on early food production (Blench and MacDonald, 2000; van der Veen, 1999). Rather than fitting into broad patterns known from other parts of the world, pathways to food production in Africa are distinctive. New genetic data support archaeological hypotheses of early Holocene domestication of cattle in northeast Africa. The earliest African food producers were mobile herders, not sedentary farmers. Herding developed in marginal areas, and then spread patchily across the Sahara and to the south as climatic conditions deteriorated. Although complex strategies for plant use developed early in Africa (c. 17,000 BP), plant domestication was late (after 4000 BP), and occurred in many different environments. To date, no models have addressed the multiple pathways to food production in the African continent. In this paper, we examine reasons for three of the most distinctive African themes: early domestication of cattle in northeast Africa, patchy spread of food production, and late domestication of African plants. We argue that a desire for increased predictability had a significant influence on all three of these African patterns. In the following section, we develop an ethnoarchaeologically based model that links the need for predictability in daily food supply to specific contexts in which domestication is likely. A better understanding of distinct African patterns and a new examination of the importance of short-term predictability as a P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 101 motivator for domestication may contribute to understanding factors influencing subsistence intensification and variation in trajectories toward food production in other regions of the world. THE IMPORTANCE OF DAY-TO-DAY PREDICTABILITY AND SCHEDULED CONSUMPTION Since Lee’s research of the 1960s, which suggested that hunter-gatherers often work less than farmers (Lee, 1968; but see Hawkes and O’Connell, 1981), archaeologists have emphasized the productivity of farming (it can support many people per unit area) as the major advantage of agriculture over hunting and gathering. Current theories for agricultural origins focus on population growth, environmental deterioration, and social or economic gain (reviews in Cowan and Watson, 1992; Price and Gebauer, 1995); despite diverging specific emphases, they share the premise that domestication occurred because humans wanted larger quantities of certain resources. A number of scholars have emphasized risk and the need for a predictable food supply (Flannery, 1986; Redding, 1988; Smith, 1998; Wills, 1995; Winterhalder and Goland, 1997). But the more common view, well expressed in current textbooks, is that understanding the reason for increased food yields is key to understanding the origins of agriculture (Crabtree and Campana, 2001, p. 242). Surprisingly little research has focused on individual human motivations during early stages of the domestication process, however. The end results of agriculture—visible today as larger yields, higher carrying capacity, denser stands of crops, larger seeds and seed heads, or greater animal productivity—were not necessarily realized during the earliest phases of the domestication process. Human motivations for initial manipulation of plants and animals thus remain an open question. Historic, anthropological, and ethnobotanical accounts of tending and transplanting wild plants by hunter-gatherers are potentially valuable sources of information on this issue (e.g. Steward, 1938; Turner and Kuhnlein, 1982; reviews in Harris and Hillman, 1989; Lourandros, 1997). In many cases, however, domestication was not the focus of research, and specific reasons for intensification are unclear (Hayden, 1990, versus Keeley, 1995). Ethnoarchaeological research seems the most promising source of new data on processes of domestication. Studies have identified modes of selection that lead to domestication, such as the use of sickles to harvest cereals (Hillman and Davies, 1990) or breeding only the most tamable individual animals (Trut, 1999). Researchers recognize that planting selected plants and captive breeding of desired animals are essential to maintaining selection and to the domestication process. Yet, ethnoarchaeological research has not focused P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 102 10:22 Style file version June 30th, 2002 Marshall and Hildebrand on the precise contexts in which people living in small-scale societies find it advantageous to capture and breed plants or animals. In this section, we use ethnoarchaeological data to address the question of why, during the domestication process, people perform the critical steps of sowing, relocating, or capturing plants and animals from the wild. We follow Harris’s conceptual framework for degrees of intensification in interactions between humans and plants and animals (Harris, 1989, 1996b; see also Hillman and Davies, 1990). Our ethnoarchaeological studies focus on motives for domestication among hunter-gatherers and extensive farmers currently adopting wild resources into domestic contexts. Marshall (2001) lived among Okiek, hunter-gatherers of Kenya during 1989–90, many of whom were farming at the time. Hildebrand (2001) studied shifting cultivators of forests and grasslands of southwest Ethiopia; she lived with the northeast Sheko during 1998–2000, and collected data on local crops and land use in nearby Gura Ferda and northern Dizi areas. In both studies, we found that people want to be able to schedule use of certain resources, and so manipulate plants and animals to ensure predictable access. We think that the motivations we observed for the capture and breeding of plants and animals are relevant (sensu Wylie, 1985) to those of any group of people contemplating the initial manipulation of wild resources. Knowledge of horticulture does not negate the relevance of these decisions, and presentday motivations for plant manipulation can inform considerations of early herding. This is true because, although decisions to capture or manipulate plants and animals may lead to domestication or adoption of domesticates, domestication is not necessarily the goal envisaged at the time of initial manipulation. Small, discrete steps in the domestication process reflect more general decisions about resource intensification. These arise in particular contexts (time, place, and social setting) of resource use, and are made by hunter-gatherers, horticulturists, and pastoralists alike. Understanding such contexts is key to understanding the reasons for capture and breeding of plants and animals by ancient people. The Case Studies Sheko and northern Dizi farmers who harvest yams (Dioscorea cayenensis Lam. complex) in wild areas often leave them to regrow, but transplant them to home gardens in contexts where the need for yams is unusually great, or yams are more difficult to find. In northeast Sheko, spontaneous yams are mostly transplanted by bachelors who value them as an easily prepared food. People with predictable access to food cooked by female kin transplant spontaneous yams much less frequently. In nearby northern P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 103 Dizi areas, families historically transplanted yams from sparsely populated lowlands, where spontaneous yams are abundant, to denser upland settlements, where they are rare. Yams growing spontaneously near upland settlements are left in place. Northern Dizi yam transplantation thus focuses on making resources distant from settlements more accessible. Adoptive transplantation is almost never practiced among the people of Gura Ferda, a third subgroup of the Sheko–Dizi ethnic entity. Demands for increased predictability at Gura Ferda are low because spontaneous yams are abundant and easy to find; furthermore, residential mobility in Gura Ferda is much higher in terms of both frequency and distance of moves, so that transplantation would not make access to yams much easier or more predictable in the long term. These examples show that the need for predictable access to certain foods can motivate people to capture and propagate plants; this is especially true for individuals who have an exceptional need for the plant, live in locations where the desired plant is relatively scarce, or invest in a long-term occupation site. Moving either a plant or an animal near a house guarantees predictable access, so that people can monitor the resource and schedule its consumption. Piik ap Oom Okiek also manipulate the distribution of wild plants, and plant some of them in domestic settings. Okiek families eat wild and weedy greens every day, but it is not the most desired greens that are sown or transplanted. Instead, effort is put into manipulating only those plants that are uncommon or not conveniently located (Marshall, 2001). Basella, for example, is an edible wild climbing shrub most common in steep ravines. Families often transplant one or two plants to the doorways of their homes. Similarly, Gynandropsis is a weed of disturbed ground, which families may sow if it does not spontaneously reseed near houses. Some less mobile Piik ap Oom families now also transplant a tree (Dracaena sp.), a creeper (Periploca sp.), and a high-altitude grass (Eleusine sp.) used in initiation and marriage ceremonies. All cases of planting and manipulation of these resources affect distribution more than abundance. Planting edible wild plants where they are accessible is a form of living storage. Through day-to-day monitoring of the depredations of pests, predators, and neighbors, individuals know the status of the plant and its availability for consumption or use. Concern is for predictable availability of plants eaten regularly or used at important ceremonies, not for increased yield. Manipulation is likely in contexts where families are staying in one place for a long period, where the distribution of desired plants is not convenient, or where ceremonies are about to take place. The need for precise scheduling also shapes Okiek choices of animal species for different consumption events. Okiek modes of meat acquisition (trapping, hunting with spears, or use of domestic animals) differ in the P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 104 10:22 Style file version June 30th, 2002 Marshall and Hildebrand predictability of their outcome. When Piik ap Oom families eat meat, it is usually wild (giant forest hog or bushbuck), but at ceremonies or gatherings, domestic sheep, goat, or cattle are used exclusively. Domestic animals provide no more meat than wild animals, and have no ritual significance. In fact, the “wild” is especially important in Okiek ceremonies (Kratz, 1994), but domestic stock is favored for ceremonies and gatherings because of its predictable availability. Okiek use of cattle provides examples of contexts in which close control over animals may be extremely desirable. Thus cattle, although domesticated, help us to understand potential circumstances in which wild animals are likely to have been domesticated prehistorically. Scheduled ceremonies create a clear need for easily and predictably available meat, which is best supplied by penned animals. The relation between scheduled consumption events, an increased need for predictablity, and capture of wild animals is supported by recently reported practices of the Conibo-Shipibo of the Peruvian Amazon, who capture, pen, and feed manatees, peccaries, and monkeys in preparation for feasts (DeBoer, 2001). Despite great differences in socioeconomic organization and environment among these ethnoarchaeological cases, they reveal similar contexts and motivations for manipulating resources. People often manipulate plants and animals in situations where the need for predictable access or scheduled consumption is especially great. We argue that these needs shaped prehistoric decisions to manipulate resources in ways that led to domestication. Although higher yield is a popularly cited advantage of food production and does affect later phases of agricultural innovation, the need for predictable access may have been a more important catalyst in many domestication events. The Model Intensification and domestication are structurally linked to the need for predictable day-to-day access and scheduled consumption. Understanding this connection makes it possible to move beyond the particulars of our studies to model a broader range of contexts in which predictable access to specific resources can become an especially prominent concern. We begin by identifying contexts that make predictable access to resources desirable, and go on to examine the relationship between these contexts and the large-scale social, economic, and ecological processes that create them. This approach fits the large anthropological literature on the risk of going without food in spatially and temporally variable environments (Cashdan, 1990; Halstead and O’Shea, 1989; Kelly, 1995). It also echoes the archaeological literature on P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 105 risk and domestication cited earlier, but differs in scale and in emphasis. We focus on day-to-day, rather than seasonal, uncertainty about the availability of key resources. We are concerned more with the initial risk and social cost of not having what you want when you want it, than the subsequent danger of starvation. Day-to-day predictability of wild resources may become a major concern in settings where sedentism is increasing, plant abundance is decreasing, or prey mobility is increasing. Increasing sedentism can make distant resources difficult to collect, so that access becomes unpredictable. Transplanting, sowing, or otherwise moving the resource to the locus of human occupation restores the compromised predictability of that resource. Decreased abundance of certain plants, or increased mobility of prey can also bring about a crisis in predictability. In both situations, a formerly dependable resource becomes difficult to rely on, and humans may respond by focusing more on other resources (if available), or by finding ways to better manage either the resource itself or their access to it. In the case of plants, this may mean transplanting or sowing the plant in an especially favorable location where it can be weeded or tended. In the case of animals, it may mean allocating a certain portion of the human group to follow or guide stock along optimal routes for grazing, to find water, and to protect the herd from predators or competing human groups. These proximate factors (changes in sedentism, plant abundance, or prey mobility) may be caused by a number of broader social or ecological conditions. Sedentism is often thought to arise where key resources are abundant and concentrated. Decreased or patchy distribution of plants may be due to natural ecological differences, environmental degradation, overexploitation, or novel patterns of predation or disease. Prey mobility can increase as rainfall becomes lower and more variable, and the distribution of plants becomes more spatially and temporally stochastic. Any of these highly varied circumstances may raise search costs for key resources, so that knowledge about the location or state of a resource becomes less certain. One potential response to this crisis in predictable access is to manipulate the distribution of the resource to satisfy human needs for day-to-day or ceremonial events. If people knowingly or inadvertently cause morphological selection in tandem with manipulation of plant or animal distributions, then domestication may result. Domestication is linked to broad social or ecological processes, proximate contexts in which predictability becomes crucial, and manipulation practices meant to restore or enhance predictable access and scheduled consumption. We think the utility of this scheduled consumption–predictability model for domestication is broad: it can be applied to plants or animals in pristine settings of domestication, and to circumstances in which P1: GLQ Journal of World Prehistory [jowo] 106 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand domesticates were adopted from elsewhere. In Africa, this model can explain much about the domestication and spread of cattle, the patchy spread of food production, and the late domestication of plants. SCHEDULED CONSUMPTION AND THE DOMESTICATION OF CATTLE What circumstances might have prompted North Africans to capture and domesticate cattle? In western Asia, where studies of the domestication of cattle have a long history, scholars have emphasized religious motives (reviewed in Isaac, 1962). These theories are compatible with our emphasis on scheduling, and identification of ceremonies as settings in which domestication is likely. But the scheduled consumption model allows us to identify a broader range of settings in which capturing and domestication of cattle would have been advantageous—contexts in which people need to maintain or increase predictable access to key resources. These are most likely to arise amid ecological perturbations, such as drought or disease, or among sedentary populations clustered around concentrations of resources. We examine the influence of these factors on North African hunter-gatherers immediately prior to domestication, and develop a theory for the domestication of African cattle in the eastern Sahara. Climate and Society Before Food Production It is widely recognized that the more arid a region, the greater the variability in the amount, location, and timing of rainfall, both within seasons and between years (Coppock, 1993; Nicholson, 1980). Nonequilibrium systems exist in arid regions of Africa today (less than 300 mm p.a.), where the coefficient of variation in rainfall often exceeds 30%. Unpredictable rainfall causes great variation in the productivity of African savanna ecosystems (Behnke et al., 1993; Ellis and Swift, 1988). Fluctuations have been especially marked in North Africa, where regional feedback mechanisms prolong and amplify climatic perturbations such as droughts (Nicholson, 1989, 1994). The instability caused by repeated cycles of aridity is likely to have augmented concerns about predictability among North African hunter-gatherers during several key periods in prehistory. Hyperarid conditions prevailed across North Africa during the last glacial maximum. Very dry conditions c. 20,000 BP began to ameliorate c. 12,500 BP, giving way to oscillating wet and dry conditions that resulted from major systemic changes in atmospheric circulation at the end of the P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 107 last glaciation (Grove, 1993; Hassan, 1997, 2000; Nicholson and Flohn, 1980; Petit-Maire, 1991; Street and Gasse, 1981). The moist climatic regime, punctuated by a cold, dry phase c. 11,350–10,250 BP, peaked c. 9500–8500 BP. Rainfall decreased gradually across North Africa after c. 8000 BP, with regional variation in timing and severity. Wendorf et al. (2001) recognize six humid phases in the eastern Sahara between c. 9500 and 3800 BP. During wetter periods, much of the Sahara was covered with grasslands, the mountain ranges of the Sahara supported mediterranean vegetation, and water levels were high in lakes and rivers (Fig. 1). The eastern Sahara, however, was arid at all times, with severe droughts c. 9500 BP and c. 8700–8600 BP (Hassan, 2000; Wendorf et al., 1984, 2001). The Saharan region saw many short arid phases, and two marked ones c. 8000/7500–7000/6500 BP and 4500– 3000 BP (Grove, 1993; Hassan, 1997). More localized droughts occurred c. 9500–9000 BP at Adrar Bous in northwest Niger, and sporadically between c. 8500 and 7000 BP in the Chad Basin (Barich, 1998). The effects of these fluctuations on subsistence would have been especially pronounced in areas that consistently received scant rainfall, such as the eastern Sahara. Changes in hunter-gatherer technology and social organization also created contexts that favored domestication. During the last glacial maximum, the Sahara was deserted and population was dense in the Nile Valley. In some cases, as at Wadi Kubbaniya c. 17,000 BP, settlement remained in one place for several seasons of the year and resource use was intensive. Plants were harvested in ways that may have increased their abundance and diversity, and processed on grindstones. Occupants of such sites also fished, and hunted wild cattle, hartebeest, and gazelle (Wendorf et al., 1989). But these activities did not result in domestication. Later, c. 13,000–12,500 BP, inhabitants of the Nile Valley used plants in similar ways at Tushka, where grindstones are common. At this site, burials with skulls of wild cattle suggest that these animals had symbolic significance prior to domestication (Wendorf, 1968; Wendorf and Schild, 1976). After being deserted during the last glacial maximum, the Sahara was repopulated c. 9500 BP by hunter-gatherers who used ceramics with distinctive wavy-line decorative motifs. This cultural complex, scattered across North Africa, is variously referred to as Khartoum Mesolithic (Arkell, 1949), Epipaleolithic (Close, 1995), or Aqualithic (Sutton, 1977). Sites are concentrated in relatively well-watered massifs and lake basins. Some hunter-gatherers were fairly sedentary and harvested wild plants intensively, especially cereals. At Ti-n-Torha East rockshelter in the Acacus, people built stone structures (Barich, 1987). At this and nearby sites of Uan Afuda and Uan Tabu, they gathered and ground wild grasses, and hunted and possibly managed Barbary sheep (Ammotragus lervia) (Barich, 1987; Cremaschi et al., 1996; Di Lernia, 1999, 2001; Garcea, 2001). Farther south in the Khartoum Nile, Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 10:22 Fig. 1. Rainfall and physical features of North Africa. Present-day isohyets are shaded; the 200 mm isohyet is widely recognized as the border between the Sahara (desert) and Sahel (grassland). Estimated Sahara–Sahel boundaries for 9000 and 18,000 bp are also shown. Information from Banks (1984), Gautier (1987a), Goudie (1996), and Petit-Maire (1989). These and all other radiocarbon dates are uncalibrated. P1: GLQ Style file version June 30th, 2002 P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Cattle Before Crops 109 sites of the tenth to ninth millennia BP preserve abundant ceramics, as well as wild ungulates, fish, molluscs, grindstones, and wild grasses (Caneva, 1988; Haaland, 1987; Magid and Caneva, 1998; Peters, 1986). With concepts of ownership based on storage facilities and ceramics, some of these groups in the central and southern Sahara probably followed a delayed-return strategy of hunting and gathering (Barich, 1998; Dale et al., in press; Di Lernia, 2001). Domestication of Cattle in North Africa: Timing and Location Cattle were the earliest domesticates in Africa. Recent studies suggest that they were probably domesticated from North African populations of wild Bos primigenius by hunter-gatherers of the eastern Sahara 10,000–8000 BP. Their origins are still controversial, but Gautier (1980, 1987a, 2001) and Wendorf (Close and Wendorf, 1992; Wendorf et al., 1984, 2001; Wendorf and Schild, 1980) argue for domestic cattle in the eastern Sahara at Bir Kiseiba c. 9500 BP, and Nabta Playa c. 8840 BP (Figs. 1 and 2). Because these sites preserved few cattle bones, evidence for morphological change is difficult to evaluate (Grigson, 2000; Smith, 1986). Wendorf and colleagues buttress the admittedly scarce morphological data with an ecological argument: without human intervention, survival of wild cattle in the arid eastern Sahara would have been unlikely (Close and Wendorf, 1992; Wendorf et al., 1984; Wendorf and Schild, 1998). Cattle are present to the west at Enneri Bardagué in the Tibesti by c. 7400 BP and in the Acacus by c. 7400–6700 BP (Garcea, 1995; Gautier, 1987a). There are no early domestic cattle in the Nile Valley. Recent morphological and genetic research provides some support for Wendorf’s hypothesis. Grigson’s morphological study (Grigson, 1991, 2000) shows that Egyptian cattle of the fifth millennium BP had long, slender limbs morphologically distinct from those of Eurasian humpless cattle (Bos taurus) and Zebu (Bos indicus). On this basis, she suggests that African cattle may have been domesticated from wild Bos primigenius in Africa. Recent research on genetic variation in breeds of present-day African cattle points to a similar conclusion (Bradley et al., 1996; Bradley and Loftus, 2000; Hanotte et al., 2002). Genetic distance between African cattle and Asian Bos taurus is sufficient to define the two as discrete genetic populations (Bradley et al., 1996; Bradley and Loftus, 2000). Bradley and colleagues argue that wild cattle in Eurasia, Bos primigenius primigenius, and in Africa, Bos primigenius opisthonomus, diverged by 22,000 years ago, and propose that African populations of wild cattle were domesticated in Africa. Together, archaeological and recent genetic evidence indicate a single geographic origin for domestic cattle in the eastern Sahara (Gautier, 1987a; Hanotte et al., 2002). P1: GLQ Journal of World Prehistory [jowo] 110 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand Fig. 2. Distribution of sites with early domestic cattle. The earliest domestic sheep and goat in Africa appear c. 7000–6700 BP in the eastern Sahara and the Red Sea Hills (Close, 1992, 2002; Gautier, 1987a; Vermeersch et al., 1996). They almost certainly come from western Asia (Gautier, 1984a), because there are no wild ancestors for sheep and goat in Africa. Close (2002) argues that sheep and goat came to Africa via the southern Sinai before Near Eastern crop complex, which is thought (Wetterstrom, 1993) to have entered the continent through the Nile Valley. The fact that sheep and goats postdate domestic cattle is further evidence for indigenous domestication of cattle in North Africa. More archaeological data P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Cattle Before Crops 111 are needed, however, including large, well-dated faunal samples, enclosures with cattle dung, and sites that immediately predate domestic cattle in the eastern Sahara. The origins of African cattle are thus still controversial, but genetic and morphological data together strongly suggest independent domestication in Africa. Given the sparseness of faunal data from Nabta and Kiseiba and the absence of slightly older sites, specific application of the scheduled consumption model to small-scaled analyses of sites or household economies at the time of domestication is not yet possible. Application of the scheduled consumption model in more general terms is worthwhile, however, because it allows us to explore specific relations between short-term predictability and resource intensification in North Africa. Domestication of Cattle and the Scheduled Consumption Model Archaeological evidence suggests that cattle were domesticated in the eastern Sahara during the tenth millennium BP. Nabta, located in the driest part of the Sahara, received too little rainfall at this time (less than 300 mm p. a.) to sustain wild cattle. Domestication probably took place slightly farther west, in areas capable of supporting cattle. In marginal areas of the eastern Sahara, human populations concentrated in playa basins (Close, 2001; Close and Wendorf, 1992; Wendorf et al., 1984, 2001; Wendorf and Schild, 1998) or massifs. We argue that hunter-gatherers in these settings domesticated cattle to ensure their predictable availability as a food source. Both ecological perturbations, especially recurring cycles of aridity, and concentration of resources and people in playa basins, could have precipitated an increased need for predictability. Ritual use of cattle may also have provided a specific context in which scheduled consumption would have been especially desirable. Given that aridity is a recurring theme in North Africa, the question remains: why did domestication occur c. 9000 BP in marginal circumstances, rather than amidst arid conditions c. 17,000 BP, when human populations concentrated in the Nile Valley? We think that subtle variation in rainfall is more important to day-to-day predictability than acute aridity. During the last glacial maximum, environments were so extreme that conditions were quite predictable in most places. The Sahara was predictably dry, and uninhabited. People lived in the Nile Valley, close to water, where other resources were also dependable. In contrast, conditions in parts of the eastern Sahara c. 9000 BP were only marginal. There was sufficient rainfall for arid-adapted resources, but not enough for their distribution to be reliable. Although humans could survive under these circumstances, planning was P1: GLQ Journal of World Prehistory [jowo] 112 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand difficult because the time and place of rainfall and herd movements could not be foreseen. Another factor that differentiates the tenth millennium from earlier arid periods is the presence of ceramic-using, delayed-return hunter-gatherers in North Africa. Rights to resources and concepts of ownership associated with such groups are important preconditions for herding (Brooks et al., 1984; Meillassoux, 1972; North, 1981). Herding presents more scheduling conflicts for hunter-gatherers than cultivation, because animals, unlike plants, cannot be left for more than a few hours at a time (Marshall, 2000). Without concepts of ownership, individuals would have been unlikely to contribute the extra labor necessary for herding, because any member of the group could have slaughtered an animal at any time. Like the “perfect storm,” the precise conditions that precipitated domestication occurred rarely in North Africa. They converged during the tenth millennium in areas of the eastern Sahara that were wet enough for wild cattle but dry enough to be risky, and were populated by hunter-gatherers with social organization conducive to resource intensification. Archaeological, genetic, and climatic evidence together suggest that domestic cattle spread from a point origin—perhaps a small playa near the Jebel Marra massif in northwest Sudan, or east of the Tibesti in northeastern Chad— during the tenth–ninth millennium BP. Hunter-gatherers of this region faced a nonequilibrium rainfall system with generally unpredictable rainfall, as well as short-term climatic fluctuations. We think that these circumstances were challenging, not in terms of absolute abundance of food, but in terms of the predictability with which food could be acquired. In this setting, where plant abundance and prey mobility varied stochastically, the scheduled consumption model predicts manipulation of favored resources during arid episodes. We suggest that local hunter-gatherers intensified their use of wild animal herds rather than their harvesting of wild plants for several reasons. Plant productivity is especially vulnerable to variation in rainfall, because the timing of rainfall relative to plant growth phases is crucial (Le Houérou et al., 1988; Mortimore, 1998). During droughts, ungulates are a more reliable resource than plants because their populations are maintained through movements that exploit local differences in topography, vegetation, and rainfall (Behnke et al., 1993). Following wild ungulates would have been a particularly attractive strategy for hunter-gatherers of the southeast Sahara during the tenth–ninth millennia BP, who faced variability in the amount, location, and timing of rainfall. The alternative, increasing mobility combined with more generalized use of plants, might not have been possible. Generalization would have carried the risk of lowered foraging efficiency (Winterhalder, 1986, p. 378), and the plant component of the diet was already generalized, requiring use of relatively P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 113 low-rank resources such as grains, which necessitated cumbersome grindstones for processing. Following resilient herds of large wild ungulates, such as cattle or hartebeest, would have reduced risk and constituted generalization by proxy, because the animals processed more diverse plant resources than humans could. In such arid conditions, however, locating herds of ungulates would have been difficult, and access to animal products would have remained unpredictable because of erratic rainfall and the high mobilities and low densities of wild herds. Sporadic access to herds would have impeded attempts to monitor herd size, composition, nutritional status, and the effects of disease and carnivore predation. This would have limited knowledge of their condition, and made scheduled consumption events (from large ceremonies to daily family meals) difficult to plan. Such scheduling would have been especially important to delayed-return hunter-gatherers of the early Holocene in the Sahara, because broad social networks, consolidated by periodic gatherings, would have helped to spread risks in an uncertain environment. A ceremonial role for cattle at such gatherings would have provided social, as well as dietary, motivation for humans to achieve or maintain predictable access to cattle through control of herds. Cattle would have been the logical focus of intensification for many reasons. Wild cattle would have been the main meat source for Saharan hunter-gatherers (Hassan, 2000). They were grassland-adapted herd animals (Gautier and van Neer, 1989), and would have been the easiest large North African ungulate to tame. Barbary sheep are territorial and found in small groups, and gazelle and other antelope are notoriously difficult to domesticate (Clutton-Brock, 1981; Diamond, 1997; Haltenorth and Diller, 1988, p. 105; Lewis, 1977), whereas cattle have proven amenable to domestication multiple times in different parts of the world (Bradley et al., 1996; Grigson, 1989; Meadow, 1996). This is probably due to their size (energetic efficiency), rapid growth, and behavioral characteristics (ease of breeding in captivity, lack of territoriality, and well-developed dominance hierarchy) (Clutton-Brock, 1981; Diamond, 1997). Tame animals could have been controlled by corralling them overnight in brush enclosures. Provisioning (wells or salt), taming, bleeding, milking, and selective breeding would have followed. Taming cattle and protecting them from predators at night would have required substantial commitment of labor, however (Marshall, 2000), and the sustained effort required is likely to have occurred only among delayedreturn hunter-gatherers with storage technology (ceramics) and concepts of ownership. We have stressed the importance of predictability in the domestication of cattle. During droughts, yield and predictability both would have declined, however. Might people have corralled cattle to increase yield rather P1: GLQ Journal of World Prehistory [jowo] 114 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand than predictability? Precise figures for the number of large animals that hunter-gatherers would have been able to kill under these circumstances are difficult to obtain, but a !Kung hunter in a similarly semiarid environment kills up to five large animals per year (Lee, 1979, p. 231). Hadza hunters kill substantially more animals: six to nine large animals per hunter per year (O’Connell et al., 1992). In contrast, African pastoralists today harvest only 4–8% of the cattle herd a year; in areas where drought and disease are common, this represents the surplus to the growth needs of the herd (Dahl and Hjort, 1976). To kill as many animals a year as does a single !Kung hunter would require keeping a herd of at least 125 cattle, an improbable scenario in the early stages of domestication. Given the slow rate of growth of cattle herds, and the fact that early domesticates are smaller, not larger, than their wild ancestors, it is unlikely that cattle were domesticated to increase yield. We hypothesize that delayed-return Saharan hunter-gatherers of the tenth–ninth millennium followed herds, and subsequently domesticated cattle in order to increase day-to-day predictability and reduce risk by manipulating a resource that could move to exploit localized favorable conditions. Controlling movements diminishes the risk of not finding animals, allows evaluation of condition and predictable access, and creates a dense, movable concentration of resources. Day-to-day control of herds and subsequent domestication of cattle also fit with well-known strategies for increasing longterm predictability: storage, mobility, and sharing (Halstead and O’Shea, 1989). Keeping cattle is a form of storage on the hoof (Close and Wendorf, 1992; Legge, 1989), and mobility and sharing of resources such as water and grazing are probable features of early pastoralism in the Sahara. [Our use of the term “pastoralism” follows Dyson-Hudson and Dyson-Hudson’s (1980) definition of pastoralists as people who rely heavily on production from domestic herds, and move herds to pasture.] Nascent cooperative social and political links among far-flung early herding groups would have passed on information and established safety networks (Legge, 1989; Robinson, 1989; Ryan et al., 2000). These could have developed through resource sharing, stock loans, ceremonies, and other social bonds. Cattle domestication in North Africa established mobile herding, and pastoralism rather than settled agriculture, as the earliest form of food production. In the following sections, we discuss the wide-ranging influence of herding on the subsequent spread and development of food producing economies in Africa. THE SPREAD OF FOOD PRODUCTION IN AFRICA: WHY SO PATCHY? The spread of food production on the African continent was strikingly uneven: hunter-gatherers and food producers coexisted in all regions long P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 Cattle Before Crops 10:22 Style file version June 30th, 2002 115 after initial contact or colonization by food producers. This was due to different factors in different regions within Africa, most of which related ultimately to the relative predictability of herding versus hunting and gathering. There was variation in the timing as well as the spread of early food production in Africa. Herding spread rapidly but incompletely across the Sahara during the mid-Holocene. As climatic conditions deteriorated during the sixth millenium BP, pastoralists moved south to better-watered regions, such as the Sudanese Nile. Small groups of pastoralists moved into eastern Africa slightly later. In southern Africa, early food production is largely associated with movements of Iron Age mixed farmers into the region c. 2000 years ago. In this section, we focus only on regions that are central to understanding continent-wide variability in pathways to food production. Parts of the western Sahara and the African rainforests that are important to the domestication of plants—and where livestock adoption follows patterns similar to those in other regions—are discussed in a subsequent section on African plant domestication. Northeast and Northwest Africa Pastoral occupation at Nabta c. 8000 BP provides an early example of a characteristic African pattern of early food production. The settlement is highly structured but mobile and seasonal, rather than village-based. Site E75–6 has at least two rows of hut floors with associated cooking holes, storage pits, many grindstones, and two large wells that could have been used to water herds of cattle (Banks, 1984; Close and Wendorf, 1992; Wendorf and Schild, 1998). A wide range of wild plants were collected. Use of wild sorghum may have been intensive (Wasylikowa et al., 1993, 1997), but E75–6 was only occupied seasonally (Close and Wendorf, 1992; Królik and Schild, 2001). As rainfall became lower and more variable in North Africa, pastoralism spread unevenly from the eastern Sahara to the Tibesti and Acacus massifs and the west African Sahara between c. 7000 and 5000 BP (Fig. 2). Even after the introduction of stock, the new subsistence economy was generalized, and Saharan pastoralists hunted and sometimes fished (Clark et al., 1973; Gautier, 1987a; Smith, 1980). Early Holocene patterns of plant use also persisted at Uan Muhuggiag and Adrar Bous (Smith, 1980; Wasylikowa, 1993), although smaller quantities of grindstones at some sites suggest less processing of wild plants than previously (Barich, 1987). Localized, unpredictable rainfall necessitated long-distance movements to exploit variable topography and vegetation for water and pasture (Muzzolini, 1993; Smith, 1992a). Herders at Adrar Bous and Acacus sites P1: GLQ Journal of World Prehistory [jowo] 116 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand were more mobile than earlier hunter-gatherers (Barich, 1987; Gautier, 1987b; Smith, 1992a). Upper levels at Torha North and Uan Muhuggiag preserve discontinuous laminae of dung, suggesting seasonal use of the shelters as livestock pens (Cremaschi et al., 1996; Gautier and van Neer, 1977– 82). Although herders seasonally occupied large sites near playas at Nabta and Dakhleh c. 7900–5500 BP (Close, 1992; McDonald, 1998a), ephemeral sites are more common throughout the Sahara (Barich, 1998; Close, 1990; Gabriel, 1987). Pastoral rock art attests to the symbolic importance of domestic cattle (Holl, 1999; Muzzolini, 1993; Smith, 1992), as do cattle burials c. 6500 BP near Adrar Bous and at Nabta (Applegate et al., 2001; Malville et al., 1998; Paris, 2000; Wendorf and Królik, 2001; Wendorf and Schild, 1998). Rituals associated with cattle may have occurred at seasonal meetings of pastoral groups or lineages, and helped to consolidate emerging social and political networks. Highly mobile pastoral land use minimized competition with huntergatherers. Even after herding was widespread, hunter-gatherers lived near pastoral groups until c. 7000 BP in the eastern Sahara at Dakhleh Bashendi-A (McDonald, 1998a), and until c. 5500 BP in the west-central Sahara at Amekni (Camps, 1968). Spatial variation in conditions made sedentary hunting and gathering unsustainable in different places at different times, and contributed to the patchy spread of herding. After c. 6500 BP, rainfall decreased. Broader pastoral contacts and exchange systems (McDonald, 1992; Smith, 1980, 1992a) may have buffered deteriorating conditions. Green vitric tuff and Amazon stone were traded thousands of kilometers (Clark, 1970; McDonald, 1992). Nevertheless, successful herding was short-term in some places: in the late-middle sixth millennium BP pastoral occupation ceased, at least temporarily, at Dakhleh BashendiB, Abu Ballas, and in the Nabta-Kiseiba area (McDonald, 1998a,b). Some herders responded to increasing aridity by emphasizing small stock, increasing mobility (Gautier, 1987a), and moving south to better-watered areas. Much of the eastern Sahara was depopulated by c. 5500 BP, and central Saharan montane sites such as Uan Muhuggiag were abandoned by c. 5000 BP (Barich, 1998; Close, 1992). Herders entered the Khartoum Nile by c. 5500 BP (Gautier, 1987a; Peters, 1986), the west African Sahel by c. 4500–3500 BP (Breunig et al., 1996; Holl, 1998; MacDonald and MacDonald, 2000; Smith, 1992a), and the forest margin by c. 3000 BP (Stahl, 1985; van Neer, 2000). The Sudanese Central Nile and Greater Eastern Africa Chronology and relations between Sudanese and Saharan areas (Paris, 2000; Smith, 1992a) suggest that domestic stock were introduced from the P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 117 Sahara as it became drier (Haaland, 1992; Hassan, 1997). Cattle, sheep, and goats appear by the sixth millennium BP (Gautier, 1984b,c; Peters, 1986) (Fig. 2). Local assemblages of lithics and ceramics show continuity (Caneva, 1987, 1989; Haaland, 1995; Marks and Mohammed-Ali, 1991), indicating that any movement of Saharans into the region was small-scale, and culture contact was more important than migration to socioeconomic change. Entry of Saharans may have been eased by prior social links with the Sudan, indicated by trade and common ceramic styles. Compared to the original Saharan herding environments, the Sudanese Nile offered more dependable, productive resources. This area also posed no particular problems for cattle, as it lies within their wild range. Like earlier local hunter-gatherers, pastoralists used large, semipermanent camps near the Nile, as at Esh Shaheinab and Geili (Caneva, 1988; Haaland, 1995; Krzyzaniak, 1991). Domestic animals are the dominant large mammals at many sites, such as Kadero c. 5000–4000 BP, but were added to a wide range of wild animals used by earlier hunter-gatherers (Gautier, 1984c; Haaland, 1992). Unlike Saharan pastoralists, herders in this better-watered landscape are thought to have used plants more intensively than their hunter-gatherer predecessors. Site structure and increased use of grindstones at Kadero 1, Um Direiwa, and Zakiab indicate to Haaland (1981, 1992) that, as early as 5000 BP, pastoral groups were cultivating sorghum that was morphologically wild (Stemler, 1990). Social differentiation appeared among Sudanese herders by the sixth millennium BP. Clusters of especially rich graves of men, women, and children at Kadero 1 argue for differences in wealth (Krzyzaniak, 1991), but there is no evidence for social stratification. Pastoral intensification and a decrease in wild animal use is also evident at some sites in the Middle Nile after 5300 BP. Despite these developments, the spread of herding was patchy: at Shaqadud, east of the Nile, subsistence focused on wild resources as late as 4000 BP (Marks and Mohammed-Ali, 1991; Peters, 1991). Farther to the east near the Eritrean border, cattle and small stock appear at Atbai sites during the fifth and fourth millennium BP (Fattovich, 1993; Sadr, 1991, pp. 53, 138). As herders continued to spread east and south of the central Nile, they moved beyond the natural distribution of wild cattle in North Africa and encountered new environmental and epizootic challenges. The earliest domestic cattle in the Horn of Africa date to c. 3500–2500 BP at Lake Besaka and Gobedra (Brandt, 1984; Phillipson, 1977). They spread slowly because of the vertical relief and closed woodlands of highland areas. Dramatic rock paintings of cattle herds in the Horn probably date to this period, and may reflect ceremonies and seasonal gatherings of frontier pastoral groups (Brandt and Carder, 1987; Joussaume, 1981). P1: GLQ Journal of World Prehistory [jowo] 118 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand Before 4000 BP, small numbers of herders migrated into Kenya from increasingly arid areas of Sudan and Ethiopia (Fig. 2), but herding was not widespread until c. 3000 BP, and extended only to northern Tanzania. Arid conditions c. 6000–3300 BP (Ambrose, 1998) and wild animal diseases (Gifford-Gonzalez, 2000) may have slowed the spread of herding in southern Kenya, and made cattle a less predictable source of food than in more northern areas. Cattle moving into areas with wildebeest and buffalo were exposed for the first time to Bovine Malignant Catarrhal and East Coast Fevers. In this frontier context, the low density of herders would have made seasonal aggregations more important, because it would have constrained other mechanisms for risk reduction, such as intergroup exchange networks, stock loans, and gifts (Gifford-Gonzalez, 1998, 2000), as well as the availability of breeding stock. Hunter-gatherers of the fifth millenium BP near Lake Turkana and in central Kenya are thought to have added herding to local hunting or fishing strategies, because lithics show continuity with earlier East African traditions (Ambrose, 1984a; Bartheleme, 1985). Use of the Pastoral Neolithic funerary complex at the northern Kenyan site of Jarigole (Nelson, personal communication, 1998; Gifford-Gonzalez, 2000) may have reinforced extensive social networks among dispersed early pastoral groups. Farther south in the central Rift Valley, the earliest domestic stock are found at very low densities in a hunter-gatherer occupation, RBL2.1, c. 4000 BP at Enkapune Ya Muto rockshelter (Marean, 1992). A mosaic of pastoral and hunter-gatherer groups coexisted in southern Kenya and parts of northern Tanzania from >4000 BP onwards. After 3500 BP, two distinct specialized pastoral cultures emerged: the Elmenteitan at sites like Ngamuriak, and the Savanna Pastoral Neolithic at Narosera and Crescent Island Main (Bower, 1991; Robertshaw, 1990). Both cultures relied on intensive use of livestock, and made little use of abundant wild ungulates (Gifford-Gonzalez, 2000; Marshall, 2000). At the site of Enkapune Ya Muto, contemporary Eburran 5 hunter-gatherers had lithic technology and microlith styles similar to those of earlier hunter-gatherers, and consumed large quantities of wild fauna and limited stock (Ambrose, 1984b). The few domestic animals are attributed to gifts from pastoral neighbors, raiding, or limited herding (Marean, 1992). Nderit ceramics similar to those found on pastoral sites also attest to interaction between Eburran hunter-gatherers and nearby herders (Ambrose, 1998). Pastoral use of the landscape was mobile and extensive, did not destroy hunter-gatherer habitat, and allowed local hunter-gatherer subsistence and social organization to continue (Gifford-Gonzalez, 2000; Marshall, 1986, pp. 248–249). Gifford-Gonzalez (1998) argues that a likely pastoral strategy for reducing the risk of moving into new areas would have been to integrate P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 Cattle Before Crops 10:22 Style file version June 30th, 2002 119 with local hunter-gatherer groups, perhaps through marriage alliances. In this way, some hunter-gatherers would have assimilated into herding groups, but herders could fall back among hunter-gatherer groups in case of stock loss to drought or disease. Social and economic systems continued to be fluid in the central Rift Valley until recent times: hunter-gatherer and pastoral groups interacted regularly, some hunter-gatherers adopted food production, and pastoralists periodically suffered stock losses (Marshall, 1994; Mutundu, 1999; Spear and Waller, 1993). Southern Africa Adoption of food production in southern Africa followed a different trajectory from that in the northern half of the continent. Although small groups of pastoralists are thought to have entered parts of southern Africa early, disease was a barrier to domestic stock. The introduction of domestic animals and plants is closely associated with the rapid spread of Early Iron Age farmers c. 1600 BP. Mixed agriculture became common in the extreme east, and herding in the west of South Africa. South of the Orange River, hunting and gathering continued across the interior of the subcontinent. The earliest pastoralists may have been so mobile and patchily distributed as to be archaeologically invisible in some places. It has long been thought that early herders spread from Zimbabwe and Zambia south, and that stone-using Khoisan groups may have brought sheep and pottery from the Zambezi through Namibia to the Cape by 2000 BP (Klein, 1984; Smith, 2000). Some scholars suggest, however, that ceramics and small stock both spread south through exchange networks between hunter-gatherers and Iron Age farmers, rather than via Khoi migration (Mitchell, 1996; Sadr, 1998). There are few sites dating to this period, but sheep are directly dated at Spoegrivier in Namibia c. 2105 BP, and Blombos Cave in the southern Cape c. 2000 BP (Henshilwood, 1996; Sealy and Yates, 1994, 1996) (Fig. 2). Faunal evidence from other sites, previously thought to indicate early herding, may in fact be more recent. Pastoral sites occur at low densities, and show a generalized subsistence based on sheep herding and variable use of wild animals and plants. Early Iron Age groups in southern Africa after 2000 BP lived in fairly permanent settlements and relied on livestock, especially sheep and goats, and African grains and pulses (Maggs, 1984). Cattle became more numerous after c. 1500 BP, but fish, molluscs, tortoises, and wild mammals continued to be important (Plug and Voigt, 1985; Voigt, 1987). Mixed farming was not continuously distributed across far southern Africa as it was just south of the equator. Because of livestock diseases, winter rainfall, and desert areas, P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 120 10:22 Style file version June 30th, 2002 Marshall and Hildebrand farming was confined to the eastern half of South Africa. Away from the continental margins, immediate-return hunter-gatherers continued to exploit flexible and predictable resources such as tubers and mongongo nuts, which were more evenly distributed in space and time than were cereals in North Africa. In areas where hunter-gatherers and food producers coexisted, they had variable relations, ranging from trade (Bousman, 1998; Kinahan, 1996; Smith et al., 1991, 1996; Wadley, 1996) to clientship (Denbow and Wilmsen, 1986; Schrire, 1992). Over much of southernmost Africa, agriculture was not adopted until recent times (Deacon, 1984a,b; Smith, 1992a). Pathways to food production in northern and southern Africa offer an interesting contrast. Diamond (1997, p. 132) suggests that southern regions lacked a critical mass of potential domesticates, and that domestic plants spread south slowly because of Africa’s north–south axis. Although the ranges of wild sorghum and rice extend into southern Africa (Harlan, 1992a), Diamond notes that cattle and most African cereal crops are not found wild south of the equator. Factors affecting continental patterns of rainfall also differ between Sahel and southern Africa in ways that may have affected pathways to food production. Despite significant arid areas within southern Africa, the subcontinent as a whole is generally cooler and wetter, and lacks the local feedback mechanisms that prolong droughts in the Sahel (Nicholson, 1989, 1994). Increasingly unpredictable rainfall and the resultant stresses on Saharan hunter-gatherer groups during the Holocene may not have had parallels in the Southern Hemisphere. Another interesting contrast with northern Africa is that southern regions have no evidence for early ceramic use, and little for delayed-return subsistence strategies (but see Sadr, 1998). Digging sticks and digging stick weights for harvesting underground storage organs are found in the archaeological record from early periods (Deacon 1984a,b). Holocene huntergatherers in southern Africa were able to exploit high-ranked tubers and nuts, resources that are consumed immediately. Thus, factors that promote use and storage of relatively low-ranked resources such as wild grasses may not have operated in southern Africa. Finally, the southward spread of a fully developed Early Iron Age agricultural complex made the domestication of local plants less likely than in other parts of Africa, where early herders without domestic plants were the first food producers in many regions. Why So Patchy? As during the initial domestication of cattle, concerns about predictable access to animal products would have shaped decisions to adopt livestock, or to move them to new locales. The relative predictability of herding versus P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 121 hunting and gathering varied from place to place, however, depending on local food resources, terrain, diseases, and social settings. African cattle were well-adapted to the grasslands of north Africa that constituted the original wild range of Bos primigenius: wetter parts of the Sahara and northern Sudan. There, pastoralism spread quickly because cattle allowed hunter-gatherers to use a broad range of vegetation types, intensifying subsistence while exploiting localized environmental instability. Herding was more sustainable than sedentary hunting and gathering in an arid landscape with unpredictable resources, and social relations among huntergatherer groups were well established. Trading networks between the Sahara and the Sudanese Nile had long existed, and in the Sudan cattle had access to a dependable water source. In contrast, herders moving into eastern Africa had no previous relations with local hunter-gatherers, and limited access to other herders, breeding stock, and social safety networks. Eastern Africa also lies outside the wild range of cattle, and wildlife diseases made livestock a less predictable source of food. In part because of these diseases, pastoralism was not widespread farther south until it was integrated with crop cultivation. Pastoralists tend to cope with climatic variability by exploiting spatial heterogeneity, rather than by modifying the landscape. Early herders thus affected hunter-gatherer resources less than settled agriculturalists would have done, and extensive, shifting land use allowed pastoralists and huntergatherers to coexist, contributing to the continuation of hunting and gathering. Furthermore, in many areas, herding offered no particular advantages over existing strategies of hunting and gathering, but would have required much more labor. Despite environmental deterioration in the Sahara, hunting and gathering persisted for millennia after stock became widely distributed. The Sudanese Nile offered reliable access to plants and fish without investing in herding. The abundance of game in East Africa would have offered higher returns from hunting than herding. In eastern and southern Africa, the sustained commitment of time and labor required by herding would not have fit well with preexisting immediate-return hunting and gathering strategies supported by the region’s comparatively predictable nuts, fruits, tubers, and game. In Africa, hunting and gathering continued both as an independent strategy and as a component of generalized pastoralism. This pattern contrasts with the spread of farming in many other regions of the world. Despite the success of herding as the earliest form of food production in Africa, several characteristics of herding (as compared with agriculture) contributed to the uneven spread of food production, and ensured the continuation of hunting and gathering groups. Pastoralists are mobile, with relatively low population densities. They are quite specialized, depending mainly on P1: GLQ Journal of World Prehistory [jowo] 122 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand three domestic species. In addition, the continous labor of herding presents more scheduling conflicts than farming does for hunter-gatherers adopting food production: domestic plants can be more easily left than animals (Marshall, 2000). The form of the earliest food production in Africa, pastoralism, and the spatial variation in relative predictability of hunting and gathering versus food production, both contributed to the patchy spread of food production so distinctive of Africa. DOMESTICATION OF PLANTS: WHY SO LATE? In most regions of the world, plants were domesticated before animals. In Africa, plants were domesticated long after herding was first established. Delays in domestication of African plants are due to a number of factors, many of which relate to scheduled consumption, predictability, and the mobile nature of early pastoral societies. We discuss general reasons for the late domestication of plants; reasons for domestication of particular plant species are beyond the scope of this paper. The first domestic plants in Africa were southwest Asian crops c. 7000 BP, which were confined to the Nile Valley by their need for winter rain (Wetterstrom, 1993). Indigenous African crops can be grouped into three complexes (Table I): savanna, forest margin, and Ethiopian (Harlan 1982, 1992b). On the basis of the geographic distribution of wild progenitors of African crops, Harlan (1971) argues that in Africa, unlike many other regions of the world, domestication of plants was noncentric: crops did not spread from a single geographic point of origin (Fig. 3). Rather, domestication occurred under a variety of conditions in widely dispersed regions of Africa. The first indigenous domestic grains appear after early movement of Saharan groups into the west African grasslands c. 4000–3500 BP (Smith 1980, 1984) (Fig. 4). Domestic pearl millet is found as impressions in sherds at Dhars Tichitt and Oualata in Mauritania from c. 3500 BP (Amblard, 1996; Amblard and Pernès, 1989). Charred grains of domestic pearl millet are directly dated to c. 3460 BP at Birimi in northern Ghana (D’Andrea et al., 2001), c. 2840 BP at Ti-n-Akof in northern Burkina Faso, and c. 2930 BP at Gajiganna and c. 2430 BP at Kursakata in the Chad basin of northeast Nigeria (Neumann et al., 1996). Recent studies have resolved the longstanding question of whether pearl millet was domesticated by hunter-gatherers, early mobile herders, or later, more sedentary herders (Clark, 1976; Shaw, 1977). It now appears that domestication took place among semisedentary herders in west African savannas, well after the first appearance of pastoralists (D’Andrea et al., 2001; Holl, 1985; Neumann, 1999). Domestic pearl Latin name Cola acuminata Schott et Endl. Cola nitida (Vent.) Schott et Endl. Fruit, leaves, seeds Cereal Seeds: stimulant (Robusta coffee) Fruits: edible Fruits: caffeine Tropical west Africa Highlands, Guinea Lowland forests, west Africa to Uganda West Africa, forests West Africa, forests August 23, 2002 Cola Crops of the Forest Margin Complex Okra Abelmoschus esculentus (L.) Guinea millet Brachiaria deflexa (Schum.) Hubb. Robusta coffee Coffea canephora Pierre Highlands, Guinea Dry lowland savanna, south and east Africa Widespread in lowlands Senegal to Lake Chad Savanna, Togo to Nigeria West Africa Savanna Widely used Savanna, far west Africa Dry savanna, Sudan to Senegal Savanna, Sudan to Chad Savanna, west Africa to east Nigeria Habitat and/or location PP573-jowo-378641 Cereal Pulse Cereal Fruit, leaves, seed, oil: water source in deserts Leaves: pot herb Cereal Cereal Leaves: pot herb, seeds Leaves, calyces: pot herb Pericarp: containers Cereal Cereal Edible parts and uses Journal of World Prehistory [jowo] Crops of the Savanna Complex Guinea millet Brachiaria deflexa African watermelon Citrullus lanatus (Thumb.) Masf., Colocynthis citrullus (L.) O. Kuntze Tossa jute Corchorus olitorius Linn. Fonio Digitaria exilis (Kipp.) Stapf Black fonio Digitaria iburua Stapf Kenaf Hibiscus cannabinus Linn. Roselle Hibiscus sabdariffa Linn. Bottle gourd Lagenaria siceraria Standl. African rice Oryza glaberrima Steud. Pearl millet, Pennisetum glaucum (L.) R. Br., Bulrush millet formerly P. americanum Sorghum Sorghum bicolor (L.) Moench Bambara Voandzeia subterranea groundnut (L.) Thouars Vernacular name Table I. Selected African Crops, Their Uses and Original Distributionsa P1: GLQ 10:22 Style file version June 30th, 2002 Cattle Before Crops 123 10:22 et al. (1991), FAO (1988), Harlan (1992a), Harlan et al. (1976), Purseglove (1968, 1972), and Zeven and Zhukovsky (1975). Ethiopia, Kenya, Somalia Mid/highland forest, Ethiopia Highland Ethiopia, east Africa Mid/highland forest, Ethiopia Semiarid mid/highland Ethiopia Low/midland Ethiopia, east Africa West and north Ethiopia highlands Midlands, west Ethiopia West Africa Forest margin, west Africa August 23, 2002 a Engels Leaves: stimulant Seeds, leaves: stimulant Cereal Corm, pseudostem starch Cereal Tuber Oil seed Wild fruits, domestic tubers Fruit; seeds: oil Pulse, pot herb Forest, west Africa Midlands and highlands, Ethiopia, east and south Africa Southern Sahel, east/south Africa Forest margins, west Africa Savanna, west Africa to east Nigeria Savanna, east Africa West Africa West Africa Habitat and/or location 124 Crops of the Ethiopian/East African Highlands Complex Chat Catha edulis Forsk. Coffee Coffea arabica Linn. Finger millet Eleusine coracana (L.) Gaertner Enset Ensete ventricosum (Welw.) Cheesman Tef Eragrostis tef (Zucc.) Trotter Yam Dioscorea cayenensis Lam. complex Noog Guizotia abyssinica Cass. Anchote Coccinia abyssinica (W. & A.) Cogn. Fluted gourd Cowpea Legume: seeds, tubers Pulse Tuber Tuber Subterranean and aerial tubers Tuber Fruit: oil; male flowers: wine Pulse Edible parts and uses PP573-jowo-378641 Yam pea Dioscorea cayenensis Lam. complex Elaeis guineensis Jacq. Kerstingiella geocarpa Harms Yam Oil palm Kersting’s groundnut Hyacinth bean Hausa potato Piasa Lablab niger Medik. Plectranthus esculentus N. E. Br. Solenostemon rotundifolius (Poir.) J.K. Morton Sphenostylis stenocarpa (Hochst.) Harms Telfairia occidentalis Hook. f. Vigna unguiculata (L.) Walp. Dioscorea bulbifera L. Latin name Journal of World Prehistory [jowo] Air potato Vernacular name Table I. (Continued ) P1: GLQ Style file version June 30th, 2002 Marshall and Hildebrand P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 125 Fig. 3. Proposed areas of domestication of African plants (after Harlan, 1971). 1. Guinea millet; 2. Fiono and black fonio; 3. African rice; 4. Yam (Dioscorea cayenensis complex); 5. Enset; 6. Tef; 7. Groundnuts (Kerstingiella and Vooandzeia); 8. Sorghum; 9. Bullrush/pearl millet; 10. Finger millet. millet appears abruptly 600 years after early pastoralists enter Burkina Faso and Nigeria (Neumann, 1999). In the middle Niger region of Mali, domestic pearl millet, sorghum, and African rice are known from the beginning of the occupation sequence at Jenné Jenno c. 2060 BP. Even after farming becomes widespread, wild plants remain important: use of wild panicoid grasses persists in the Chad basin (Neumann, 1999), and wild rice, Brachiaria, Panicum, Echinocloa, and greens are found at Jenné Jenno (McIntosh, 1995, 1997). P1: GLQ Journal of World Prehistory [jowo] 126 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand Fig. 4. Distribution of sites with early cultivated or domestic plants. Sites in the eastern Sahara and Sudanese Nile reveal evidence for early intensive use of sorghum, but for late morphological change. Sorghum dating to c. 7950–8020 BP at Nabta has lipids that differ from the wild form, but it is morphologically wild (Wasylikowa and Dahlberg, 1999; Wasylikowa, 2001; Wendorf et al., 1998). Its abundance indicates intensive use (Close, 2001) or perhaps occasional cultivation of wild sorghum (Wasylikowa et al., 1997). Impressions of wild sorghum appear in sherds at Um Direiwa, Kadero 1, and Zakiab (Stemler, 1990). On the basis of these, and on the high frequency of grindstones (e.g., 30,000 at Um Direiwa), Abdel-Magid (1989) P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 Cattle Before Crops 10:22 Style file version June 30th, 2002 127 and Haaland (1981, 1992, 1999) argue that wild sorghum was cultivated by c. 5000 BP. Secure dates for domestic sorghum are late: c. 2060 BP at Jenné Jenno (McIntosh, 1995) and 20–250 AD at the historic sites of Meroë, Jebel Tomat, and Qasr Ibrim in the Sudan (Clark and Stemler, 1975; Rowley-Conwy et al., 1999; Stemler and Falk, 1981) (Fig. 4). Morphologically domestic sorghum (race bicolor) from Qasr Ibrim is genetically identical to both wild and domestic modern sorghum in the area of the genome studied, supporting the idea of recent domestication (Rowley-Conwy et al., 1999). Despite the importance of the Horn of Africa as a center for agricultural origins, little is known about domestication processes there. Recent excavations in the Aksum area (Fig. 4) have found domestic tef in historic periods: c. 500 BC at sites D and K (Boardman, 1999) and during the fifth century AD at Bieta Giyorgis (Bard et al., 1997). Sorghum and noog oilseeds appear during the sixth century AD (Boardman, 1999). Ethnoarchaeological studies of crop processing stages for tef (D’Andrea et al., 1999) and of differences between wild and domestic enset (Hildebrand, 2001) will facilitate future archaeological investigation. The relative antiquity of indigenous versus exotic Near Eastern crops in Ethiopia has yet to be fully explored. East Africa has long been regarded as the locus for domestication of finger millet (Harlan, 1992a). Little research has been conducted on Later Stone Age sites in Uganda, but domestic finger millet occurs by the early seventh century AD at Aksum (Boardman, 1999), and c. 1185 BP at Deloraine Farm in Kenya (Ambrose, 1984c) (Fig. 4). The forests and forest margins of central and western Africa have yielded little archaeological data on the domestication of plants. After domestic sheep and goat appeared c. 3500 BP at Kintampo, use of local legumes and oil palm may have increased (but see Maley, 2001), yet local wild resources such as Canarium and small wild animals remained important (Anquandah, 1993; Flight, 1976; Stahl, 1985, 1993). Domestic cowpeas and Bambara groundnuts are first found in Iron Age contexts in western Africa (Vogelsang et al., 1999). No indigenous domesticates are known from the southern half of the continent. Rather, pearl and finger millet, sorghum, and domestic pulses appear in conjunction with Early Iron Age mixed farming in Zambia, Zimbabwe, and South Africa (Maggs, 1984). The earliest grain, domestic Pennisetum, dates to c. 270 AD at Silver Leaves (Klapwijk, 1974; Klapwijk and Huffman, 1996) (Fig. 4). Why So Late? Many African cereals are amenable to domestication because of their appealing taste, rapid growth, high yields, drought tolerance, good storage P1: GLQ Journal of World Prehistory [jowo] 128 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand potential, and single-locus genetic inheritance of key traits (Diamond, 1997; Harlan, 1992a). The absence of evidence for early domestic plants has been attributed to several causes. Young and Thompson (1999) argue that poor preservation of seeds and cereals in tropical east Africa results from rapid carbon cycling, wetting and drying cycles, and a high level of soil microbial activity, and contributes to a lack of evidence for early crops. Insufficient archaeobotanical research is also an issue (Bar-Yosef, 1998; Wetterstrom, 1998). Poor preservation and lack of research do not completely account for the absence of morphologically domestic plant remains before 4000 BP, however. A growing body of paleoethnobotanical data from northern Africa attests to the use of wild grasses and other plants by hunter-gatherers and early herders (Abdel-Magid, 1989; Magid and Caneva, 1998; Stemler, 1990; Wasylikowa, 1993; Wasylikowa et al., 1993, 1997). Possible reasons for the late onset of morphological change among African crops include harvesting practices, plant biology (Abdel-Magid, 1989; Haaland, 1992), the unpredictable plant productivity of the Sahara and its margins, and the mobility of early herders. Uprooting plants or cutting seed heads with sickles can select for indehiscence, synchronous ripening, and loss of dormancy. Not all harvesting methods result in domestication, however (Harlan, 1992a). Tuareg and Zaghawa pastoralists today collect Panicum, Cenchrus, and other wild African grains by hand-stripping, by using a swinging basket, or by beating (Harlan, 1989, 1992c; Nicolaisen, 1963). These methods select for brittle rachises, and hence will not result in domestication. Evidence for prehistoric selection processes is mixed. Blades or flakes with gloss, possible indicators of sickle harvesting, are absent at Zakiab, Kadero 1, and Um Direiwa (Haaland, 1992), but appear during the mid-Holocene at Dakhleh (McDonald, 1998a), Adrar Bous III (Roset, 1987), and Farafra (Barich, 1998), and at Laga Oda in Ethiopia during the last 3000 years (Clark and Prince, 1978). More detailed microwear studies are needed to identify the precise cause of the gloss (e.g., grain cutting or hide preparation); if, however, these pieces are indeed found to be sickles, they might be indicative of harvesting practices that could have led to domestication. Even if harvesting methods favor tough rachises, grains must be replanted and genetically isolated for selection to result in morphological change (Harlan, 1989, 1992c; Hillman and Davies, 1990). Late domestication of sorghum and pearl millet has been attributed to their ability to outcross, which can impede genetic isolation. According to this view, domestication of sorghum became possible only when early Sudanese pastoralists took it outside its natural range (Abdel-Magid, 1989; Haaland, 1992, 1995). Outcrossing rates vary among African cereals, however (National Research Council, 1996): finger millet is almost entirely self-pollinating, sorghum has highly P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 129 variable rates of outcrossing, and pearl millet outcrosses the most but appears to have been domesticated the earliest. Although genetic isolation is a well-known general requirement of domestication, the actual mechanisms for isolation during prehistoric times are far from clear. Climate and ecology in the southern Sahara may have fostered interactions between people and plants that would not have led to domestication. As already noted, low and variable rainfall affects productivity of plants more than that of animals, because the timing of rainfall relative to plant growth phases is crucial, and plants have fixed positions, whereas animals can move or disperse. All of these factors make cultivation of grains in marginal environments more risky than plant collecting. Even today, much of the northern Sahel can support pastoralism but not farming. Areas with less than 558 mm p. a. average one crop failure in 10 years, and farming is rarely attempted with less than 348 mm p. a. (Mortimore, 1998, p. 77). If hunter-gatherers or pastoralists with marginal subsistence due to low and variable rainfall sought more predictable access to food, then emphasizing livestock would have been more logical than undertaking cultivation. Planting would have entailed gambling on when and where rain would fall and which cereals the rainfall distribution would suit. Different grains have different growth cycles: pearl millet matures quickly, whereas sorghum grows slowly but can use residual moisture (Mortimore, 1998, p. 89). By focusing on stock, herders could move animals to pasture and continue exploiting wild grass stands wherever they occurred in any given year. From initial stages of cattle domestication until the fourth millenium BP, pastoral strategies provided more predictable access to food than did intensification of plants. Mobile pastoral strategies precluded steady selection on populations of useful plants. When pastoralists moved into wetter grasslands and became more sedentary, then selection pressures on plants became sufficiently constant to cause morphological change. Domestication requires a constellation of cultural plant management practices, such as reaping with a sickle and replanting harvested grain that is either self-fertilizing or genetically isolated year after year. If not all of the requisite practices or conditions are in place, then selection is not maintained, and morphological change does not take place. In Africa, suitable sets of factors for domestication came together late, rarely, and in highly varied circumstances. This is largely because the unpredictable environments of the early middle Holocene Sahara, and the mobile pastoral lifestyles they fostered, together created circumstances in which humans would not have exercised continuous, directional selection on cereals. Continuing intensive use of wild plants indigenous to different parts of Africa led to the continent’s noncentric pattern of domestication. P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 130 10:22 Style file version June 30th, 2002 Marshall and Hildebrand DISCUSSION African patterns of early food production contrast with those in other parts of the globe in interesting ways. In Africa, domestic animals were present several thousand years before domestic plants, and the earliest food producers were mobile cattle herders. In other regions of the world, plants were domesticated first, animals were domesticated later, among settled, village-based communities (Grigson, 2000; Smith, 1998; see also Hole, 1996), and nomadic pastoralism emerged still later, as a specialized strategy complementing settled agriculture (Bar-Yosef and Khazanov, 1992). African cattle and some African cereals were domesticated in high-risk arid and semiarid environments, settings not consistent with current emphases on domestication in relatively resource-rich environments (Harris, 1996a; Price and Gebauer, 1995; Smith, 1998). The desire to schedule consumption of resources can lead people to manipulate plants and animals, and may have prompted domestication of African cattle amid low and variable rainfall in the Sahara during the early Holocene. Herding did not develop in predictably abundant areas such as the Nile Valley, or in predictably harsh environments such as the late Pleistocene Sahara. Rather, it developed in marginal environments where predictable access to resources was important, and where mobile animals were less vulnerable than plants to localized, short-term droughts. Prerequisites for labor investment in herding included delayed-return strategies of hunting and gathering, and concepts of ownership. As pastoralism spread across the Sahara and subsequent desiccation prompted herders to move south, adoption of food production was patchy despite the overall success of herding. This is largely due to spatial variation in the relative predictability of herding versus hunting and gathering. Difficult terrain impeded the spread of stock into some areas; elsewhere, cattle diseases made pastoralism a risky endeavor. Where herding became established, mobile use of the landscape by small pastoral populations left many local resources intact. Thus, in areas where wild resources were predictable, local groups could continue to hunt and gather well after the arrival of domesticates. Herding is more difficult to adopt than cultivation, especially among immediate-return hunter-gatherers, because of ownership and scheduling issues. All of these factors led to a distinctively African pattern of slow, patchy spread of food production. Other Old World agricultural complexes often competed more directly with local hunter-gatherer subsistence strategies, so that food production was adopted broadly along a rapidly moving frontier. The development of herding before mixed agriculture started Africans on a distinctive path of subsistence intensification. Mobility lightened P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 131 selective pressure on local plant populations; harvesting practices and plant biology may also have delayed morphological change. Prolonged, intensive use of wild plants across the African continent led to a noncentric pattern of late domestication, and continued use of wild plants by foodproducing societies. In many early farming societies outside Africa, a complex of crops such as cereals, pulses, and livestock often spread as a package from a point-origin (Harlan, 1971; Harris, 1996b), and the importance of local wild resources diminished sharply once the suite of domesticates was established. Early mobile, animal-intensive food production may have had important consequences for subsequent trajectories of social change in Africa. Pastoral groups require communal access to distant water sources and pastures and are often associated with social structures such as age sets that promote wide ranging relationships (Spear, 1993). Many of the traits of early African herders are also those thought to be important to the development of relatively egalitarian societies over the long term: communal access to pastures, low population densities, and high mobility (Salzman, 1999 and CA comment). Although livestock-based inequalities of wealth are known to exist, they may be more difficult to create and maintain than agriculturally based inequalities (Little, 1999; Salzman, 1999; Schneider, 1979; but see Fratkin, 1999). Flexible camp groups and the importance of decision making by individual herd owners also tend to diffuse political authority (McCabe, 1999; Tavakolian, 1999). Some African patterns, such as the use of small-seeded crops, patchy adoption of food production, and continuation of hunter-gatherer societies, resemble those of eastern North America (Fritz, 1990; Smith, 1992b; Watson, 1989) more than those of western Asia (Bar-Yosef, 1998; Harris, 1996a). Others, such as domestication of animals before plants, may be similar to Andean patterns (Browman, 1989; see also Wheeler, 1984; Wing, 1986). African data also reinforce some commonalities noted for many loci of domestication. Arid conditions following the end of the Pleistocene are often thought to have catalyzed subsistence change (Bar-Yosef and Belfer-Cohen, 1989; Harris, 1996b; McCorriston and Hole, 1991; Moore and Hillman, 1992; Piperno and Pearsall, 1998). Finally, the African data support Harris’ contention that pristine domestication processes are rare, and require unusual combinations of biological and cultural circumstances (Harris, 1996c). We argue that concerns about predictable availability of resources, rather than increased yield, catalyzed domestication in Africa, and suggest that renewed attention to predictability may contribute to understanding the circumstances that led to domestication in other regions of the world. P1: GLQ Journal of World Prehistory [jowo] PP573-jowo-378641 August 23, 2002 132 10:22 Style file version June 30th, 2002 Marshall and Hildebrand ACKNOWLEDGMENTS This paper could not have been written without years of fieldwork by many Africanist scholars. We dedicate it to the memory of J. Desmond Clark. We are grateful to David Browman, Cathy D’Andrea, Gayle Fritz, Randi Haaland, Mary McDonald, Tom Pilgram, Patty Jo Watson, and Fred Wendorf for comments and information. We thank Angela Close and five anonymous reviewers, but are solely responsible for any errors. Diane Gifford-Gonzalez, Ruth Shahack-Gross, and Darla Dale contributed stimulating discussions of African archaeology. We are indebted to the institutions in Ethiopia and Kenya that have supported our research: the Kenya National Museums and the ARCCH and National Herbarium in Ethiopia. REFERENCES Abdel-Magid, A. (1989). Plant Domestication in the Middle Nile Basin: An Archaeobotanical Case Study, Cambridge Monographs in African Archaeology 35 (B.A.R. International Series 532), Oxford. Amblard, S. (1996). Agricultural evidence and its interpretation on the Dhars Tichitt and Oualata, south-eastern Mauritania. In Pwiti, G., and Soper, R. (eds.), Aspects of African Archaeology: Papers from the 10th Congress of the Pan African Association for Prehistory and Related Studies, University of Zimbabwe Publications, Harare, pp. 421–427. Amblard, S., and Pernès, J. (1989). The identification of cultivated pearl millet (Pennisetum) amongst plant impressions on pottery from Oued Chebbi (Dhar Oualata, Mauritania). African Archaeological Review 7: 117–126. Ambrose, S. H. (1984a). The introduction of pastoral adaptations to the highlands of East Africa. In Clark, J. D., and Brandt, S. A. (eds.), From Hunters to Farmers: The Causes and Consequences of Food Production in Africa, University of California Press, Berkeley, pp. 212–239. Ambrose, S. H. (1984b). Holocene Environments and Human Adaptation in the Central Rift Valley, Kenya, PhD Thesis, University of California, Berkeley. Ambrose, S. H. (1984c). Excavations at Deloraine, Rongai, 1978. Azania 20: 29–67. Ambrose, S. H. (1998). Chronology of the Later Stone Age and food production in East Africa. Journal of Archaeological Science 25: 377–392. Anquandah, J. (1993). The Kintampo complex: A case study of early sedentism and food production in sub-Sahelian West Africa. In Shaw, T., Sinclair, P., Bassey, A., and Okpoko, (eds.),The Archaeology of Africa: Food, Metals, and Towns, Routledge, London, pp. 255–260. Applegate, A., Gautier, A., and Duncan, S. (2001). The north tumuli of the Nabta Late Neolithic ceremonial complex. In Wendorf, F., Schild, R., and Associates (eds.), Holocene Settlement of the Egyptian Sahara, Vol. 1: The Archaeology of Nabta Playa. Plenum Publishers, New York, pp. 468–520. Arkell, A. J. (1949). Early Khartoum, Oxford University Press, London. Banks, K. M. (1984). Climates, Cultures and Cattle, Department of Anthropology, Institute for the Study of Earth and Man, Southern Methodist University, Dallas. Bar-Yosef, O. (1998). Introduction: Some comments on the history of research. The Review of Archaeology 19(2): 1–4. P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 133 Bar-Yosef, O., and Belfer-Cohen, A. (1989). The origins of sedentism and farming communities in the Levant. Journal of World Prehistory 3: 447–498. Bar-Yosef, O., and Khazanov, A., (1992). Pastoralism in the Levant: Archaeological Materials in Anthropological Perspectives, Prehistory Press, Madison. Bard, K., Fattovich, R., Manzo, A., and Perlinieri, C. (1997). Archaeological investigations at Bieta Giyorgis (Aksum), Ethiopia: 1993–1995 field seasons. Journal of Field Archaeology 24: 387–403. Barich, B. (ed.) (1987). Archaeology and Environment in the Libyan Sahara, 1978–1983, Cambridge Monographs in African Archaeology 23 (B.A.R. International Series 368), Oxford. Barich, B. (1998). People, Water, and Grain: The Beginnings of Domestication in the Sahara and the Nile Valley, L’Erma, di Bretschneider, Rome. Barthelme, J. (1985). Fisher-Hunters and Neolithic Pastoralists in East Turkana, Kenya, Cambridge Monographs in African Archaeology 13 (B.A.R. International Series 254), Oxford. Behnke Jr., R. H., Scoones, I., and Kerven, C. (1993). Range Ecology at Disequilibrium: New Models of Natural Variability and Pastoral Adaptation in African Savannas, Overseas Development Institute, London. Blench, R., and MacDonald, K. (eds.) (2000). The Origins and Develpment of African Livestock: Archaeology, Genetics, Linguistics, and Ethnography, UCL Press, London. Boardman, S. (1999). The agricultural foundation of the Aksumite empire, Ethiopia: An interim report. In van der Veen, M. (ed.), The Exploitation of Plant Resources in Ancient Africa, Plenum, New York, pp. 137–148. Bousman, C. B. (1998). The chronological evidence for the introduction of domestic stock into southern Africa. African Archaeological Review 15: 133–150. Bower, J. (1991). The pastoral Neolithic of East Africa. Journal of World Prehistory 5: 49–82. Bradley, D., and Loftus, R. T. (2000). Two Eves for Taurus? Bovine mitochondrial DNA and African cattle domestication. In Blench, R. M., and MacDonald, K. C. (eds.), The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics, and Ethnography, UCL Press, London, pp. 244–250. Bradley, D., MacHugh, D. E., Cunningham, P., and Loftus, R. T. (1996). Mitochondrial diversity and the origins of African and European cattle. Proceedings of the National Academy of Sciences 93: 5131–5135. Brandt, S. (1984). New perspectives on the origins of food production in Ethiopia. In Clark, J. D., and Brandt, S. A. (eds.), From Hunters to Farmers: Causes and Consequences of Food Production in Africa, University of California Press, Berkeley, pp. 173–190. Brandt, S. A., and Carder, N. (1987). Pastoral rock art in the Horn of Africa: Making sense of udder chaos. World Archaeology 19: 195–213. Breunig, P., Neumann, K., and van Neer, W. (1996). New research on the Holocene settlement and environment of the Chad Basin in Nigeria. African Archaeological Review 13: 111–145. Brooks, A., Gelburd, D. E., and Yellen, J. (1984). Food production and culture change among the !Kung San: Implications for prehistoric research. In Clark, J. D., and Brandt, S. A. (eds.), From Hunters to Farmers: Causes and Consequences of Food Production in Africa, University of California Press, Berkeley, pp. 293–310. Browman, D. (1989). Origins and development of Andean pastoralism: An overview of the past 6000 years. In Clutton-Brock, J. (ed.), The Walking Larder: Patterns of Domestication, Pastoralism and Predation, Unwin Hyman, London, pp. 256–267. Camps, G. (1968). Amekni: Néolithique ancien du Hoggar, Memoires X du Centre de Recherches Anthropologiques Préhistoriques et Ethnographiques, de Cooperation Scientifique en Algerie, Paris. Caneva, I. (1987). Pottery decoration in prehistoric Sahara and upper Nile: A new perspective. In Barich, B. E. (ed.), Archaeology and Environment in the Libyan Sahara: The Excavations in the Tadrart Acacus, 1978–1983, British Archaeological Reports, International Series 368, Oxford, pp. 231–254. Caneva, I. (ed.) (1988). El Geili: The History of a Middle Nile Environment 7000 B.C.–A.D. 1,500, Cambridge Monographs in African Archaeology 29 (B.A.R. International Series 424), Oxford. P1: GLQ Journal of World Prehistory [jowo] 134 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand Caneva, I. (ed.) (1989). Typological notes: The Sudanese case. In Krzyzaniak, L., and Kobusiewicz, M. (eds.), Late Prehistory of the Nile Basin and the Sahara, Polish Academy of Sciences, Poznan, pp. 375–379. Cashdan, E. (1990). Risk and Uncertainty in Tribal and Peasant Economics, Westview, Boulder. Clark, J. D. (1970). Prehistory of Africa, Thames and Hudson, Southampton. Clark, J. D. (1976). Prehistoric populations and pressures favoring plant domestication in Africa. In Harlan, J., de Wet, J. M. J., and Stemler, A. B. L. (eds.),Origins of African Plant Domestication, Mouton Publishers, The Hague, pp. 465–478. Clark, J. D., and Prince, G. R. (1978). Use-wear on Later Stone Age microliths from Laga Oda, Hararghi, Ethiopia, and possible functional interpretations. Azania 13: 101–110. Clark, J. D., and Stemler, A. B. (1975). Early domesticated sorghum from Central Sudan. Nature 25: 588–591. Clark, J. D., Williams, M. A. J., and Smith, A. B. (1973). The geomorphology and archaeology of Adrar Bous, central Sahara: A preliminary report. Quaternaria 18: 245–297. Close, A. (1990). Living on the edge: Neolithic herders in the eastern Sahara. Antiquity 64: 79–96. Close, A. (1992). Holocene occupation of the eastern Sahara. In Klees, F., and Kuper, R. (eds.), New Light on the Northeast African Past, Heinrich-Barth-Institut, Köln, pp. 157–183. Close, A. (1995). Few and far between: Early ceramics in North Africa. In Barnett, W.K., and Hoopes, J. W. (eds.), The Emergence of Pottery: Technological and Innovation in Ancient Societies, Smithsonian, Washington, DC, pp. 23–37. Close, A. (2001). Sites E-91–3 and E-91–4: The Early Neolithic of El-Adam type at Nabta Playa. In Wendorf, F., Schild, R., and Associates (eds.), Holocene Settlement of the Egyptian Sahara, Vol. 1: The Archaeology of Nabta Playa, Plenum, New York, pp. 71–96. Close, A. (2002). Sinai, Sahara, Sahel: The introduction of domestic caprines to Africa. In Jennerstrasse 8 (eds.), Tides of the Desert-Gezeiten der Wüste, Henrich-Barth-Institut, Cologne, pp. 459–469. Close, A., and Wendorf, F. (1992). The beginnings of food production in the eastern Sahara. In Gebauer, A. B., and Price, T. D. (eds.), Transitions to Agriculture in Prehistory, Prehistory Press, Madison, pp. 63–72. Clutton-Brock, J. (1981). Domesticated Animals from Early Times, British Museum, London. Coppock, D. L. (1993). Vegetation and pastoral dynamics in southern Ethiopian rangelands: Implications for theory and management. In Behnke Jr., R. H., Scoones, I., and Kerven, C. (eds.), Range Ecology at Disequilibrium: New Models of Natural Variability and Pastoral Adaptation in African Savannas, Overseas Development Institute, London, pp. 42–61. Cowan, C. W., and Watson, P. J. (eds.) (1992). The Origins of Agriculture: An International Perspective, Smithsonian, Washington, DC. Crabtree, P. J., and Campana, D. V. (2001). Archaeology and Prehistory, McGraw Hill, New York. Cremaschi, M., Di Lernia, S., and Trombino, L. (1996). From taming to pastoralism in a drying environment: Site formation processes in the shelters of the Tadrart Acacus massif (Libya, Central Sahara). In Castelletti, L., and Cremaschi, M. (eds.), Micromorphology of Deposits of Anthropogenic Origin, XIII Congress, UISPP, Forli, Edizioni, pp. 87–106. Dahl, G., and Hjort, A. (1976). Having Herds: Pastoral Herd Growth and Household Economy, Stockholm Studies in Social Anthropology 2, University of Stockholm, Stockholm. Dale, D., Marshall, F., and Pilgram, T. (in press). Delayed-return hunter-gatherers in Africa? Historic perspectives from the Okiek and archaeological perspectives from the Kansyore. In Crothers, G. M. (ed.), Hunter-Gatherers in Theory and Archaeology, Center for Archaeological Investigations Occasional Paper 21, Southern Illinois University, Carbondale. D’Andrea, A. C., Klee, M., and Casey, J. (2001). Archaeobotanical evidence for pearl millet (Pennisetum glaucum) in sub-Saharan Africa. Antiquity 75: 341–348. D’Andrea, C., Lyons, D., Haile, M., and Butler, A. (1999). Ethnoarchaeological approaches to the study of prehistoric agriculture in the highlands of Ethiopia. In van der Veen, M. (ed.),The Exploitation of Plant Resources in Ancient Africa, Plenum, New York, pp. 101– 122. P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 135 Deacon, J. (1984a). Later Stone Age of Southernmost Africa, Cambridge Monographs in African Archaeology 12 (B.A.R. International Series 213), Oxford. Deacon, J. (1984b). Later Stone Age people and their descendants in southern Africa. In Klein, R. G. (ed.), Southern African Prehistory and Environments, Balkema, Rotterdam, pp. 221– 328. DeBoer, W. R. (2001). The big drink: Feast and forum in the Upper Amazon. In Dietler, M., and Hayden, B. (eds.), Feasts, Smithsonian, Washington, DC, pp. 215–239. Denbow, J., and Wilmsen, E. (1986). Advent and course of pastoralism in the Kalahari. Science 234:1509–15. Diamond, J. (1997). Guns, Germs, and Steel, Norton, New York. Di Lernia, S. (ed.) (1999). The Uan Afuda Cave: Hunter-Gatherers Societies of Central Sahara, Arid Zone Archaeology Monographs 1, Università degli Studi di Roma “La Sapienza,” All’Insegna del Giglio, Firenze, Rome. Di Lernia, S. (2001). Dismantling dung: Delayed use of food resources among Early Holocene foragers of the Libyan Sahara. Journal of Anthropological Archaeology 20: 408–441. Dyson-Hudson, R., and Dyson-Hudson, N. (1980). Nomadic pastoralism. Annual Review of Anthropology 9: 15–61. Ellis, J. E., and Swift, D. M. (1988). Stability of African pastoral ecosystems: Alternate paradigms and implications for development. Journal of Range Management 41: 450–459. Engels, J. M. M., Hawkes, J. G., and Worede, M. (1991). Plant Genetic Resources of Ethiopia, Cambridge University Press, Cambridge. FAO (1988). Traditional Food Plants: A Resource Book for Promoting the Exploitation and Consumption of Food Plants in Arid, Semi-arid, and Sub-humid Lands of Eastern Africa, Food and Agriculture Organization of the United Nations, Rome. Fattovich, R. (1993). The Gash group of the eastern Sudan: An outline. In Krzyzaniak, L., Kobusiewicz, M., and Alexander J. (eds.), Environmental Change and Human Culture in the Nile Basin and Northern Africa Until the Second Millennium B.C., Archaeological Museum, Poznan, pp. 439–448. Flannery, K. V. (1986). Guilá Naquitz, Academic Press, Orlando. Flight, C. (1976). The Kintampo culture and its place in the economic prehistory of West Africa. In Harlan, J. R., DeWet, J. M. J., and Stemler, A. B. L. (eds.), Origins of African Plant Domestication, Mouton, The Hague, pp. 211–221. Fratkin, E. M. (1999). Comment on ‘Is inequality universal?’ by P. C. Salzman. Current Anthropology 40(1): 46–47. Fritz, G. J. (1990). Multiple pathways to farming in precontact Eastern North America. Journal of World Prehistory 4: 387–435. Gabriel, B. (1987). Palaeoecological evidence from Neolithic fireplaces in the Sahara. African Archaeological Review 5: 93–104. Garcea, E. (1995). New investigations in the Tadrart Acacus. Nyame Akuma 44: 35–37. Garcea, E. (ed.) (2001). Uan Tabu: In the settlement history of the Libyan Sahara, Arid Zone Archaeology Monographs 2, Università degli Studi di Roma “La Sapienza,” All’Insegnia del Giglio, Firenze, Rome. Gautier, A. (1980). Contributions to the archaeozoology of Egypt. In Wendorf, F., and Schild, R. (eds.), The Prehistory of the Eastern Sahara, Academic Press, New York, pp. 317– 344. Gautier, A. (1984a). Archaeozoology of the Bir Kiseiba region, eastern Sahara. In Wendorf, F., Schild, R., and Close, A. E. (eds.), Cattle Keepers of the Eastern Sahara: The Neolithic of Bir Kiseiba, Department of Anthropology, Institute for the Study of Earth and Man, Southern Methodist University, Dallas, pp. 49–72. Gautier, A. (1984b). Quaternary mammals and archaeozoology of Egypt and the Sudan: A survey. In Krzyzaniak, L., and Kobusiewicz, M. (eds.),Origin and Early Development of Food-Producing Cultures in North-Eastern Africa, Polish Academy of Sciences and Poznan Archaeological Museum, Poznan, pp. 43–56. Gautier, A. (1984c). The fauna of the Neolithic site of Kadero (Central Sudan). In Kryzaniak, L., and Kobusiewicz, M. (eds.), Origin and Early Development of Food-producing Cultures P1: GLQ Journal of World Prehistory [jowo] 136 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand in North-Eastern Africa, Polish Academy of Sciences and Poznan Archaeological Museum, Poznan, pp. 317–319. Gautier, A. (1987a). Prehistoric men and cattle in North Africa: A dearth of data and a surfeit of models. In Close, A. E. (ed.), Prehistory of Arid North Africa, Southern Methodist University Press, Dallas, pp. 163–187. Gautier, A. (1987b). The archaeological sequence of the Acacus. In Barich, B. E. (ed.), Archaeology and Environment in the Libyan Sahara, 1978–1983, Cambridge Monographs in African Archaeology 23 (B.A.R. International Series 368), Oxford, pp. 283–308. Gautier, A. (2001). The early to late Neolithic Archaeofaunas from Nabta and Bir Kiseiba. In Wendorf, F., Schild, R., and Associates (eds.), Holocene Settlement of the Egyptian Sahara, Vol. 1: The Archaeology of Nabta Playa, Plenum, New York, pp. 609–635. Gautier, A., and van Neer, W. (1977–1982). Prehistoric fauna from Ti-n-Torha (Tadrart Acacus, Libya). Origini XI: 87–127. Gautier, A., and van Neer, W. (1989). Animal remains from the late Paleolithic sequence at Wadi Kubbaniya. In Close, A., (ed.), and assembled by Wendorf, F., and Schild, R., The Prehistory of Wadi Kubbaniya, Vol. 2, Southern Methodist University Press, Dallas, pp. 119–161. Gebauer, A. B., and Price, T. D. (eds.) (1992). Transitions to Agriculture in Prehistory, Prehistory Press, Madison. Gifford-Gonzalez, D. P. (1998). Gender and early pastoralists in East Africa. In Kent, S. (ed.), Gender in African Prehistory, Altamira Press, Walnut Creek, CA, pp. 115–137. Gifford-Gonzalez, D. P. (2000). Animal disease challenges to the emergence of pastoralism in sub-Saharan Africa. African Archaeological Review 17(3): 95–139. Goudie, A. S. (1996). Climate: Past and present. In Adams, W. M., Goudie, A. S., and Orme, A. R. (eds.), The Physical Geography of Africa, Oxford University Press, Oxford, pp. 34–59. Grigson, C. (1989). Size and sex: Evidence for the domestication of cattle in the Near East. In Miles, A., Williams, D., and Gardner, N. (eds.), The Beginnings of Agriculture, B.A.R. International Series 496, Oxford, pp. 77–109. Grigson, C. (1991). An African origin for African cattle: Some archaeological evidence. African Archaeological Review 9: 119–144. Grigson, C. (2000). Bos africanus (Brehm)? Notes on the archaeozoology of the native cattle of Africa. In Blench, R. M., and MacDonald, K. C. (eds.), The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics, and Ethnography, UCL Press, London, pp. 38–60. Grove, A. T. (1993). Africa’s climate in the Holocene. In Shaw, T., Sinclair, P., Andah, B., and Okpoko, A. (eds.), The Archaeology of Africa: Food, Metals and Towns, Routledge, London, pp. 32–42. Haaland, R. (1981). Seasonality and division of labour: A case study from Neolithic sites in the Khartoum Nile environment. Norweigan Archaeological Review 14: 44–59. Haaland, R. (1987). Socio-Economic Differentiation in the Neolithic Sudan, Cambridge Monographs in African Archaeology 20 (B.A.R. International Series 350), Oxford. Haaland, R. (1992). Fish, pots and grain: Early and mid-Holocene adaptations in the central Sudan. African Archaeological Review 10: 43–64. Haaland, R. (1995). Sedentism, cultivation and plant domestication in the Holocene Middle Nile region. Journal of Field Archaeology 22: 158–174. Haaland, R. (1999). The puzzle of the late emergence of domesticated sorghum in the Nile Valley. In Gosden, C., and Hather, J. (eds.), The Prehistory of Food: Appetites for Change, Routledge, London, pp. 397– 418. Halstead, P., and O’Shea, J. (eds.) (1989). Bad Year Economics: Cultural Responses to Risk and Uncertainty, Cambridge University, Cambridge. Haltenorth, T., and Diller, H. (1988). A Field Guide to the Mammals of Africa, Collins, London. Hanotte, O., Bradley, D. G., Ochieng, J. W., Verjee, Y., Hill, E. W., and Rege, J. E. O. (2002). African pastoralism: Genetic imprints of origins and migrations. Science 296: 336–339. Harlan, J. R. (1971). Agricultural origins: Centers and noncenters. Science 174: 468–474. P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 137 Harlan, J. R. (1982). The origins of indigenous African agriculture. In Clark, J. D. (ed.), The Cambridge History of Africa, Vol. I, Cambridge University, Cambridge, pp. 624– 657. Harlan, J. R. (1989). Wild grass seed harvesting in the Sahara and sub-Sahara of Africa. In Harris, D. R., and Hillman, G. C. (eds.), Foraging and Farming: The Evolution of Plant Exploitation, Unwin Hyman, London, pp. 79–98. Harlan, J. R. (1992a). Crops and Man, 2nd edn., American Society of Agronomy, Inc., Madison. Harlan, J. R. (1992b). Indigenous African agriculture. In Cowen, C., and Watson, P. J. (eds.), The Origins of Agriculture: An International Perspective, Smithsonian, Washington, DC, pp. 59–70. Harlan, J. R. (1992c). Wild grass seed harvesting and implications for domestication. In Anderson, P. (ed.), Préhistoire de L’Agriculture: Nouvelles Approches Expérimentales et Ethnographiques, Monographe du CRA No. 6, CNRS, Paris, pp. 21–27. Harlan, J. R., de Wet, J. M. J., and Stemler, A. B. L. (eds.) (1976). Origins of African Plant Domestication, Mouton, The Hague. Harris, D. R. (1989). An evolutionary continuum of people–plant interaction. In Harris, D.R., and Hillman, G. C. (eds.), Foraging and Farming: The Evolution of Plant Exploitation, Unwin Hyman, London, pp. 11–26. Harris, D. R. (ed.) (1996a). The Origins and Spread of Agriculture and Pastoralism in Eurasia, Smithsonian Institution Press, Washington, DC. Harris, D. R. (1996b). The origins and spread of agriculture and pastoralism in Eurasia: An overview. In Harris, D. R. (ed.), The Origins and Spread of Agriculture and Pastoralism in Eurasia, Smithsonian, Washington, DC, pp. 552–573. Harris, D. R. (1996c). Introduction: Themes and concepts in the study of early agriculture. In Harris, D. R. (ed.), The Origins and Spread of Agriculture and Pastoralism in Eurasia, Smithsonian, Washington, DC, pp. 1–9. Harris, D. R., and Hillman, G. C. (eds.) (1989). Foraging and Farming: The Evolution of Plant Exploitation, Unwin Hyman, London. Hassan, F. A. (1997). Holocene paleoclimates of Africa. African Archaeological Review 14: 213–230. Hassan, F. A. (2000). Climate and cattle in North Africa: A first approximation. In Blench, R. M., and MacDonald, K. C. (eds.), The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics, and Ethnography, UCL Press, London, pp. 61–85. Hawkes, K., and O’Connell, J. (1981). Affluent hunters? Some comments in light of the Alyawara case. American Anthropologist 83: 622–626. Hayden, B. (1990). Nimrods, piscators, pluckers and planters: The emergence of food production. Journal of Anthroplogical Archaeology 9: 31–69. Henshilwood, C. (1996). A revised chronology for pastoralism in southermost Africa: New evidence for sheep at c. 2000 b.p. from Blombos Cave, South Africa. Antiquity 70: 945–949. Hildebrand, E. (2001). Morphological characterization of wild vs. forest-growing Ensete ventricosum (Welw.) Cheesman, Musaceae, in Sheko district, Bench-Maji Zone, southwest Ethiopia. In Friis, I., and Ryding, O. (eds.), Biodiversity Research in the Horn of Africa Region, Royal Danish National Academy of Sciences and Letters, Copenhagen, pp. 287–309. Hillman, G., and Davies, M. S. (1990). Measured domestication rates in wild wheats and barley under primitive cultivation, and their archaeological implications. Journal of World Prehistory 4: 157–222. Hole, F. (1996). The context of caprine domestication in the Zagros region. In Harris, D. R. (ed.), The Origins and Spread of Agriculture and Pastoralism in Eurasia, Smithsonian, Washington, DC, pp. 263–281. Holl, A. (1985). Subsistence patterns of the Dar Tichitt Neolithic, Mauretania. African Archaeological Review 3: 151–162. Holl, A. (1998). Livestock husbandry, pastoralisms, and territoriality: The West African record. Journal of Anthropological Archaeology 17: 143–165. Holl, A. (1999). Research on Tassilian pastoral iconography. Sahara 11: 21–34. Isaac, E. (1962). On the domestication of cattle. Science 137(3525): 195–204. P1: GLQ Journal of World Prehistory [jowo] 138 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand Joussaume, R. (1981). L’art rupestre de l’Éthiopie. In Roubet, C., Hugot, H. J., and Souville, G. (eds.), Préhistoire Africaine: Mélanges Offerts au Doyen Lionel Balout, A.D.P.F., Paris, pp. 159–175. Keeley, L. (1995). Protoagricultural practices among hunter-gatherers: A cross-cultural survey. In Price, T. D., and Gebauer, A. B. (eds.), Last Hunters-First Farmers, SAR Press, Santa Fe, pp. 243–272. Kelly, R. L. (1995). The Foraging Spectrum: Diversity in Hunter-Gatherer Lifeways, Smithsonian, Washington, DC. Kinahan, J. (1996). Alternative views on the acquisition of livestock by hunter-gatherers in southern Africa: A rejoinder to Smith, Yates and Jacobson. South African Archaeological Bulletin 51: 106–108. Klapwijk, M. (1974). A preliminary report on pottery from the north-eastern Transvaal, South Africa. South African Archaeological Bulletin 29: 19–23. Klapwijk, M., and Huffman, T. N. (1996). Excavations at Silver Leaves: A preliminary report. South African Archaeological Bulletin 51: 84–93. Klein, R. (1984). The prehistory of Stone Age herders in South Africa. In Clark, J. D., and Brandt, S. A. (eds.), From Hunters to Farmers: The Causes and Consequences of Food Production in Africa, University of California Press, Berkeley, pp. 281–289. Kratz, C. A. (1994). Affecting Performance: Meaning, Movement, and Experience in Okiek Women’s Initiation, Smithsonian, Washington, DC. Królik, H., and Schild, R. (2001). Site E75–6: An El Nabta and Al Jerar Village. In Wendorf, F., Schild, R., and Associates (eds.), Holocene Settlement of the Egyptian Sahara, Vol. 1: The Archaeology of Nabta Playa, Plenum, New York, pp. 111–146. Krzyzaniak, L. (1991). Early farming in the Middle Nile basin: Recent discoveries at Kadero (Central Sudan). Antiquity 65: 515–532. Lee, R. (1968). What hunters do for a living, or, how to make out on scarce resources. In Lee, R. B., and Devore, I. (eds.), Man the Hunter, Aldine, New York, pp. 30–48. Lee, R. (1979). The !Kung San: Men, Women, and Work in a Foraging Society, Cambridge University, Cambridge. Le Houérou, H. N., Bingham, R. L., and Skerbek, K. (1988). Relationships between the variability of primary production and the variability of annual precipitation in world arid lands. Journal of Arid Environments 15: 1–8. Legge, K. (1989). Changing responses to drought among the Wodaabe of Niger. In Halstead, P., and O’Shea, J. (eds.), Bad Year Economics: Cultural Responses to Risk and Uncertainty, Cambridge University, London, pp. 81–86. Lewis, J. G. (1977). Game domestication for animal production in Kenya: Activity patterns of eland, oryx, buffalo and zebu cattle. Journal of Agricultural Science 89: 551–563. Little, P. D. (1999). Comment on ‘Is inequality universal?’ by P. C. Salzman. Current Anthropology 40(1): 50–51. Lourandros, H. (1997). Continent of Hunter-Gatherers, Cambridge University Press, Cambridge. MacDonald, K., and MacDonald, R. H. (2000). The origins and development of domesticated animals in arid West Africa. In Blench, R. M., and MacDonald, K. C. (eds.), The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics, and Ethnography, UCL Press, London, pp. 127–162. Maggs, T. (1984). The Iron Age south of the Zambezi. In Klein, R. G. (ed.), Southern African Prehistory and Environments, Balkema, Rotterdam, pp. 329–361. Magid, A., and Caneva, I. (1998). Economic strategy based on food-plants in the early Holocene central Sudan: A reconsideration. In di Lernia, S., and Manzi, G. (eds.), Before Food Production in North Africa, Union Internationale des Sciences Prehistoriques et Protohistoriques, XIII World Congress, Forli, Edizioni, pp. 79–90. Maley, J. (2001). Elaeis guineensis Jacq. (oil palm) fluctuations in central Africa during the late Holocene: Climate or human driving forces for this pioneering species? Vegetation History and Archaeobotany 10: 117–120. Malville, J. M., Wendorf, F., Mazar, A. A., and Schild, R. (1998). Megaliths and Neolithic astronomy in southern Egypt. Nature 392: 488–491. P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 139 Marean, C. (1992). Hunter to herder: Large mammal remains from the hunter-gatherer occupation at Enkapune ya Muto rockshelter. African Archaeological Review 10: 65–127. Marks, A. E., and Mohammed-Ali, A. (eds.) (1991). The Late Prehistory of the Eastern Sahel: The Mesolithic and Neolithic of Shaqadud, Sudan, Southern Methodist University Press, Dallas. Marshall, F. (1986). Aspects of the Advent of Pastoral Economies in East Africa, PhD Dissertation, Department of Anthropology, University of California, Berkeley. Marshall, F. (1994). Archaeological perspectives on East African pastoralism. In Fratkin, E., Galvin, J., and Roth, E. (eds.), African Pastoralist Systems, Lynn Rienner, Boulder, CO, pp. 17–44. Marshall, F. (2000). The origins of domesticated animals in Eastern Africa. In Blench, R. M., and MacDonald, K. C. (eds.), The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics, and Ethnography, UCL Press, London, pp. 191–221. Marshall, F. (2001). Agriculture and use of wild and weedy vegetables by the Piik ap Oom Okiek of Kenya. Economic Botany 55: 32–46. McCabe, J. T. (1999). Comment on ‘Is inequality universal?’ by P. C. Salzman. Current Anthropology 40(1): 51–52. McCorriston, J., and Hole, F. (1991). The ecology of seasonal stress and the origins of agriculture in the Near East. American Anthropologist 93: 46–69. McDonald, M. M. A. (1992). Neolithic of Sudanese tradition or Saharo-Sudanese Neolithic? The view from Dakhla Oasis, south central Egypt. In Stenner, J., and David, N. (eds.), An African Commitment: Papers in Honour of Peter Louis Shinnie, University of Calgary Press, Calgary, pp. 51–70. McDonald, M. M. A. (1998a). Early African pastoralism: View from Dakhleh Oasis (south central Egypt). Journal of Anthropological Archaeology 17: 124–142. McDonald, M. M. A. (1998b). Adaptive variability in the eastern Sahara during the early Holocene. In di Lernia, S., and Manzi, G. (eds.), Before Food Production in North Africa, Union Internationale des Sciences Prehistoriques et Protohistoriques, XIII World Congress, Forli, Edizioni, pp. 127–136. McIntosh, R. (1997). Agricultural beginnings in sub-Saharan Africa. In Vogel, J. O. (ed.), Encyclopedia of Precolonial Africa, Altamira Press, Walnut Creek, CA, pp. 409–418. McIntosh, S. K. (1995). Paleobotanical and human osteological remains. In McIntosh, S. K. (ed.), Excavations at Jenné Jeno, Hambarketolo, and Kaniana (Inland Niger Delta, Mali, the 1981 Season), University of California Press, Berkeley, pp. 348–357. Meadow, R. H. (1996). The origins and spread of agriculture and pastoralism in northwestern South Asia. In Harris, D. R. (ed.), The Origins and Spread of Agriculture and Pastoralism in Eurasia, Smithsonian, Washington, DC, pp. 390–412. Meillassoux, C. (1972). On the mode of production of the hunting band. In Alexandre, P. (ed.), French Perspectives in African Studies, Oxford University Press, Oxford, pp. 39–58. Mitchell, P. J. (1996). Sehonghong: The late Holocene assemblages with pottery. South African Archaeological Bulletin 51: 17–25. Moore, A., and Hillman, G. (1992). The Pleistocene to Holocene transition and human economy in southwest Asia: The impact of the Younger Dryas. American Antiquity 57: 482–494. Mortimore, M. (1998). Roots in the African Dust, Cambridge University, Cambridge. Mutundu, K. (1999). Ethnohistoric Archaeology of the Mukogodo in North-Central Kenya, Cambridge Monographs in African Archaeology 47 (B.A.R. International Series 775), Archaeopress, Oxford. Muzzolini, A. (1993). The emergence of a food producing economy in the Sahara. In Shaw, T., Sinclair, P., Andah, B., and Okpoko, A. (eds.), The Archaeology of Africa: Food, Metals and Towns, Routledge, London, pp. 227–239. National Research Council (1996). Lost Crops of Africa, Volume 1: Grains, National Academy Press, Washington, DC. Neumann, K. (1999). Early plant food production in the West African Sahel: New evidence. In van der Veen, M. (ed.), The Exploitation of Plant Resources in Ancient Africa, Plenum, New York, pp. 73–80. P1: GLQ Journal of World Prehistory [jowo] 140 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand Neumann, K., Ballouche, A., and Klee, M. (1996). The emergence of plant food production in the West African Sahel: New evidence from northeast Nigeria and northern Burkina Faso. In Pwiti, G., and Soper, R. (eds.), Aspects of African Archaeology: Papers From the 10th Congress of the Pan African Association for Prehistory and Related Studies, University of Zimbabwe Publications, Harare, pp. 441–448. Nicholson, S. E. (1980). The nature of rainfall variability in Africa south of the equator. Journal of Climatology 6: 515–530. Nicholson, S. E. (1989). African drought: Characteristics, causal theories and global teleconnections. In Berger, A., Dickinson, R. E., and Kidson, J. W. (eds.), Understanding Climatic Change, Geophysical Monograph 52, International Union of Geodesy and Geophysics, Washington, DC, pp. 79–100. Nicholson, S. E. (1994). Recent rainfall fluctuations in Africa and their relationship to past conditions over the continent. The Holocene 4: 121–131. Nicholson, S., and Flohn, H. (1980). African environmental and climate changes and the general atmospheric circulation in the late Pleistocene and Holocene. Climatic Change 2: 313–348. Nicolaisen, J. (1963). Ecology and Culture of the Pastoral Tuareg, Copenhagen National Museum, Copenhagen. North, D. (1981). Structure and Change in Economic History, Norton, New York. O’Connell, J. F., Hawkes, K., and Blurton-Jones, N. G. (1992). Patterns in the distribution, site structure and assemblage composition of Hadza kill-butchering sites. Journal of Archaeological Science 19: 319–345. Paris, F. (2000). African livestock remains from Saharan mortuary contexts. In Blench, R. M., and MacDonald, K. C. (eds.), The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics, and Ethnography, UCL Press, London, pp. 111–126. Peters, J. (1986). A revision of the faunal remains from two Central Sudanese sites: Khartoum Hospital and Esh Shaheinab. In Archaeozoologia, 5th International Congress of Archaeozoologia, Bordeaux, pp. 11–35. Peters, J. (1991). The faunal remains from Shaqadud. In Marks, A. E., and Mohammed-Ali, A. (eds.), The Late Prehistory of the Eastern Sahel, Southern Methodist University Press, Dallas, pp. 197–235. Petit-Maire, N. (1989). Interglacial environments in presently hyperarid Sahara: Paleoclimatic implications. In Leinen, M., and Sarnthein, M. (eds.), Paleoclimatology and Paleometeorology: Modern and Past Patterns of Global Atmospheric Transport, Kluwer Academic, Dordrecht, The Netherlands, pp. 637–661. Petit-Maire, N. (1991). Paleoenvironments du Sahara: Lacs Holocenes à Taoudenni (Mali), CNRS, Paris. Phillipson, D. W. (1977). The excavation of Gobedra rock shelter, Axum: An early occurrence of cultivated finger-millet in northern Ethiopia. Azania 12: 53–82. Piperno, D. R., and Pearsall, D. M. (1998). The Origins of Agriculture in the Lowland Neotropics, Academic Press, San Diego. Plug, I., and Voigt, E. (1985). Archaeozoological studies of Iron Age communities in southern Africa. Advances in World Archaeology 4: 189–237. Price, T. D., and Gebauer, A. B. (eds.) (1995). Last Hunters-First Farmers: New Perspectives on the Prehistoric Transition to Agriculture, SAR Press, Santa Fe. Purseglove, J. W. (1968). Tropical Crops: Dicotyledons, Longman, London. Purseglove, J. W. (1972). Tropical Crops: Monocotyledons, Longman, London. Redding, R. W. (1988). A general explanation of subsistence change: From hunting and gathering to food production. Journal of Anthropological Archaeology 7: 56–97. Robertshaw, P. (1990). Early Pastoralists of South-Western Kenya, Memoir 11, British Institute in Eastern Africa, Nairobi. Robinson, P. W. (1989). Reconstructing Gabbra history and chronology: Time reckoning, the Gabbra calendar, and the cyclical view of life. In Downing, T. E., Gitu, K. W., and Kamau, C. M. (eds.), Coping with Drought in Kenya: National and Local Strategies, Lynne Rienner, Boulder, CO, pp. 151–168. Roset, J.-P. (1987). Néolithisation, néolithique et post-néolithique au Niger nord-oriental. International Journal of the French Quaternary Association 32: 203–214. P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 141 Rowley-Conwy, P., Deakin, W., and Shaw, C. H. (1999). Ancient DNA from sorghum: The evidence from Qasr Ibrim, Egyptian Nubia. In van der Veen, M. (ed.), The Exploitation of Plant Resources in Ancient Africa, Plenum, New York, pp. 55–62. Ryan, K., Munene, K., Kahinju, S. M., and Kunoni, P. N. (2000). Ethnographic perspectives on cattle management in semi-arid environments: A case study from Maasailand. In Blench, R. M., and MacDonald, K. C. (eds.), The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics, and Ethnography, UCL Press, London, pp. 462–477. Sadr, K. (1991). The Development of Nomadism in Ancient Northeast Africa, University of Pennslyvania Press, Philadelphia. Sadr, K. (1998). The first herders at the Cape of Good Hope. African Archaeological Review 15: 101–132. Salzman, P. C. (1999). Is inequality universal? Current Anthropology 40(1): 31–61. Schneider, H. K. (1979). Livestock and Equality in East Africa: The Economic Basis for Social Structure, Indiana University Press, Bloomington. Schrire, C. (1992). The archaeological identity of hunters and herders at the Cape over the last 2000 years: A critique. South African Archaeological Bulletin 47: 62–64. Sealy, J., and Yates, R. (1994). The chronology of the introduction of pastoralism to the Cape, South Africa. Antiquity 69: 58–67. Sealy, J., and Yates, R. (1996). Direct radiocarbon dating of early sheep bones: Two further results. South African Archaeological Bulletin 51: 109–110. Shaw, T. (1977). Hunters, gatherers and first farmers in West Africa. In Megaw, J. V. S. (ed.), Hunters, Gatherers and First Farmers Beyond Europe, Leicester University Press, Leicester, pp. 69–126. Smith, A. B. (1980). Domesticated cattle in the Sahara and their introduction into West Africa. In Williams, M. A. J., and Faure, H. (eds.), The Sahara and the Nile, A.A. Balkema, Rotterdam, pp. 489–503. Smith, A. B. (1984). Origins of the Neolithic in the Sahara. In Clark, J. D., and Brandt, S. A. (eds.), From Hunters to Farmers: The Causes and Consequences of Food Production in Africa, University of California Press, Berkeley, pp. 84–92. Smith, A. B. (1986). Cattle domestication in North Africa. African Archaeological Review 4: 197–203. Smith, A. B. (1992a). Pastoralism in Africa: Origins and Development Ecology, Hurst and Company, London. Smith, A. B. (2000). The origins of the domesticated animals of southern Africa. In Blench, R. M., and MacDonald, K. C. (eds.), The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics, and Ethnography, UCL Press, London, pp. 222–238. Smith, A. B., Sadr, K., Gribble, J., and Yates, R. (1991). Excavations in the south-western Cape, South Africa, and the archaeological identity of prehistoric hunter-gatherers within the last 2000 years. South African Archaeological Bulletin 46: 71–91. Smith, A. B., Yates, R., and Jacobson, L. (1996). Geduld contra Kinahan. South African Archaeological Bulletin 51: 36–39. Smith, B. D. (1992b). Rivers of Change: Essays on Early Agriculture in Eastern North America, Smithsonian, Washington, DC. Smith, B. D. (1998). The Emergence of Agriculture, Scientific American Library, New York. Spear, T. (1993). Being ‘Maasai’, but not ‘People of Cattle’: Arusha agricultural Maasai in the Nineteenth Century. In Spear, T., and Waller, R. (eds.). Being Maasai, James Currey Ltd., London, pp. 120–136. Spear, T., and Waller, R. (eds.) (1993). Being Maasai, James Currey Ltd., London. Stahl, A. B. (1985). Reinvestigations of Kintampo 6 rock shelter, Ghana: Implications for the nature of culture change. African Archaeological Review 3: 117–150. Stahl, A. B. (1993). Intensification in the West African Late Stone Age: A view from central Ghana. In Shaw, T., Sinclair, P., Andah, B., and Okpoko, A. (eds.), The Archaeology of Africa: Food, Metals and Towns, Routledge, London, pp. 261–273. Stemler, A. B. L. (1990). A scanning electron microscopic analysis of plant impressions in pottery from the sites of Kadero, El Zakiab, Um Direiwa and El Kadada. Archaeologie du Nil Moyen 4: 87–105. P1: GLQ Journal of World Prehistory [jowo] 142 PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 Marshall and Hildebrand Stemler, A. B. L., and Falk, R. H. (1981). SEM of archaeological plant remains. Scanning Electron Microscopy III: 191–196. Steward, J. H. (1938). Basin-Plateau Aboriginal Sociopolitical Groups, Bulletin 120, Bureau of American Ethnology, Washington, DC. Street, F. A., and Gasse, F. (1981). Recent developments in research into the quaternary climatic history of the Sahara. In Allen, J. A. (ed.), The Sahara: Ecological Change and Early Economic History, Menas, London, pp. 8–28. Sutton, J. E. G. (1977). The African aqualithic. Antiquity 51: 25–34. Tavakolian, B. (1999). Comment on ‘Is inequality universal?’ by P. C. Salzman. Current Anthropology 40(1): 54. Trut, L. N. (1999). Early canid domestication: The farm-fox experiment. American Scientist 87: 160–169. Turner, N., and Kuhnlein, H. (1982). Two important ‘root’ foods of the Northwest Coast Indians: Springbank clover (Trifolium wormskioldii) and Pacific silverweed (Potentilla anserina ssp. pacifica). Economic Botany 36: 411–432. van der Veen, M. (ed.) (1999). The Exploitation of Plant Resources in Ancient Africa, Plenum, New York. van Neer, W. (2000). Domestic animals from archaeological sites in central and west-central Africa. In Blench, R. M., and MacDonald, K. C. (eds.), The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics, and Ethnography, UCL Press, London, pp. 163–190. Vermeersch, P. M., van Peer, P., Moeyersons, J., and van Neer, W. (1996). Neolithic occupation of the Sodmein area, Red Sea mountains, Egypt. In Pwiti, G., and Soper, R. (eds.), Aspects of African Archeology: Papers From the 10th Congress of the Pan-African Association for Prehistory and Related Studies, University of Zimbabwe Publications , Harare, pp. 411– 419. Vogelsang, R., Albert, K.-D., and Kahlheber, S. (1999). Le sable savant: les cordons dunaires sahéliens au Burkina Faso comme archive archéologique et paléoecologique du Holocene. Sahara 11: 51–68. Voigt, E. A. (1987). The dispersion of domestic stock into southern Africa. Archaeozoologia 1: 149–159. Wadley, L. (1996). Changes in the social relations of precolonial hunter-gatherers after agropastoralist contact: An example from the Magaliesberg, South Africa. Journal of Anthropological Archaeology 15: 205–217. Wasylikowa, K. (1993). Plant macrofossils from the archaeological sites of Uan Muhuggiag and Ti-n-Torha, southwestern Libya. In Krzyzaniak, L., Kobusiewicz, M., and Alexander, J. (eds.), Environmental Change and Human Culture in the Nile Basin and Northern Africa Until the Second Millennium B.C., Poznan Archaeological Museum, Poznan, pp. 25–41. Wasylikowa, K., and Dahlberg, J. (1999). Sorghum in the economy of early Neolithic nomadic tribes at Nabta Playa, southern Egypt. In van der Veen, M. (ed.), The Exploitation of Plant Resources in Ancient Africa, Plenum, New York, pp. 11–32. Wasylikowa, K., Harlan, J., Evans, J., Wendorf, F., Schild, R., Close, A., Krolick, H., and Housley, R. (1993). Examination of botanical remains from Early Neolithic houses at Nabta Playa, Western Desert, Egypt, with special reference to sorghum grains. In Shaw, T., Sinclair, P., Andah, B., and Okpoko, A. (eds.), The Archaeology of Africa: Food, Metals and Towns, Routledge, London, pp. 154–164. Wasylikowa, K., Mitka, J., Wendorf, F., and Schild, R. (1997). Exploitation of wild plants by the early Neolithic hunter-gatherers of the Western Desert, Egypt: Nabta Playa as a case-study. Antiquity 71: 932–941. Wasylikowa, K. (assembler) (2001). Other botanical remains. In Wendorf, F., Schild, R., and Associates (eds.), Holocene Settlement of the Egyptian Sahara, Vol. 1: The Archaeology of Nabta Playa, Plenum, New York, pp. 592–608. Watson, P. J. (1989). Early plant cultivation in the eastern woodlands of North America. In Harris, D. R., and Hillman, G. C. (eds.), Foraging and Farming: The Evolution of Plant Exploitation, Unwin Hyman, London, pp. 555–565. P1: GLQ Journal of World Prehistory [jowo] Cattle Before Crops PP573-jowo-378641 August 23, 2002 10:22 Style file version June 30th, 2002 143 Wendorf, F. (1968). Late Paleolithic sites in Egyptian Nubia. In Wendorf, F. (ed.), The Prehistory of Nubia, Fort Burgwin Research Center and Southern Methodist Press, Dallas, pp. 954–995. Wendorf, F., and Królik, H. (2001). Site E-96–1: The complex structures or shrines. In Wendorf, F., Schild, R., and Associates (eds.), Holocene Settlement of the Egyptian Sahara, Vol. 1: The Archaeology of Nabta Playa, Plenum, New York, pp. 503–520. Wendorf, F., and Schild, R. (1976). The use of ground grain during the Late Paleolithic of the Lower Nile Valley, Egypt. In Harlan, J. R., de Wet, J. M. J., and Stemler, A. B. L. (eds.), Origins of African Plant Domestication, Mouton, The Hague, pp. 269–288. Wendorf, F., and Schild, R. (1980). Prehistory of the Eastern Sahara, Academic Press, New York. Wendorf, F., and Schild, R. (1998). Nabta Playa and its role in northeastern African prehistory. Journal of Anthropological Archaeology 17: 97–123. Wendorf, F., and Schild, R., (assemblers); Close, A., (ed.) (1984). Cattle Keepers of the Eastern Sahara: The Neolithic of Bir Kiseiba, Department of Anthropology, Southern Methodist University, Dallas. Wendorf, F., and Schild, R., (assemblers); Close, A., (ed.) (1989). The Prehistory of Wadi Kubbaniya, Vol. 2, SMU, Dallas. Wendorf, F., Schild, R., and Associates (2001). Holocene Settlement of the Egyptian Sahara, Vol. 1: The Archaeology of Nabta Playa, Plenum, New York. Wendorf, F., Schild, R., Wasylikowa, K., Dahlberg, J., Evans, J., and Biehl, E. (1998). The use of plants during the early Holocene in the Egyptian Sahara: Early Neolithic good economies. In di Lernia, S., and Manzi, G. (eds.), Before Food Production in North Africa, Union Internationale des Sciences Prehistoriques et Protohistoriques XIII World Congress, A.B.A.C.O., Forli, Edizioni, pp. 71–78. Wetterstrom, W. (1993). Foraging and farming in Egypt: The transition from hunting and gathering to horticulture in the Nile Valley. In Shaw, T., Sinclair, P., Bassey, A., and Okpoko, A. (eds.), The Archaeology of Africa: Food, Metals and Towns, Routledge, London, pp. 165–226. Wetterstrom, W. (1998). The origins of agriculture in Africa, with particular reference to sorghum and pearl millet. Review of Archaeology 19(2): 30–46. Wheeler, J. (1984). On the origin and development of camelid pastoralism in the Andes. In Clutton-Brock, J., and Grigson, C. (eds.), Animals and Archaeology: Early Herders and Their Flocks, B.A.R. International Series 202, Oxford, pp. 395–410. Wills, W. H. (1995). Archaic foraging and the beginning of food production in the American Southwest. In Price, T. D., and Gebauer, A. B. (eds.), Last Hunters-First Farmers, SAR Press, Santa Fe, pp. 215–242. Wing, E. (1986). Domestication of Andean mammals. In Vuilleumier, F., and Monasterio, M. (eds.), High Altitude Tropical Biogeography, Oxford University Press, Oxford, pp. 246– 264. Winterhalder, B. (1986). Diet choice, risk, and food sharing in a stochastic environment. Journal of Anthropological Archaeology 5: 369–392. Winterhalder, B., and Goland, C. (1997). An evolutionary ecology perspective on diet choice, risk, and plant domestication. In Gremillon, K. J. (ed.), People, Plants, and Landscapes: Studies in Paleoethnobotany, University of Alabama Press, Tuscaloosa, pp. 123–160. Wylie, A. (1985). The reaction against analogy. Advances in Archaeological Method and Theory 8: 63–110. Young, R., and Thompson, G. (1999). Missing plant foods? Where is the archaeobotanical evidence for sorghum and finger millet in East Africa? In van der Veen, M. (ed.), The Exploitation of Plant Resources in Ancient Africa, Plenum, New York, pp. 63–72. Zeven, A. C., and Zhukovsky, P. M. (1975). Dictionary of Cultivated Plants and Their Centres of Diversity, Centre for Agricultural Publishing and Documentation, Wageningen.