The Chemical Senses
advertisement

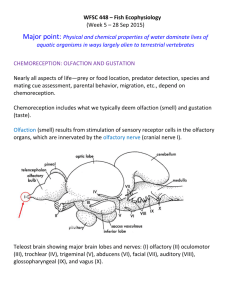
NBIO 401 Fall 2013 The Chemical Senses Class 26 – November 25th 2013 DiNino The olfactory and gustatory systems convert chemical stimuli into our perceptions of smell and taste. In humans, these systems are fully developed at birth and have played a significant role in the survival of mankind. The Olfactory System Objectives Be able to describe the pathway that odorant information takes through the olfactory system, from the nasal cavity to olfactory cortex Be able to describe how individual odorants are coded in the olfactory epithelium, the olfactory bulb, and primary olfactory cortex Be able to name cortical regions that receive projections from primary olfactory cortex and describe how smell plays a role in the functions of those areas Be able to list three reasons why the olfactory system is unique relative to other sensory systems The olfactory system detects and transduces odorants - chemicals that easily turn into a gas - enabling us to perceive these volatile chemicals as smells in the environment. Each odorant has a different chemical structure, as shown in the picture below. Many everyday smells are composed of multiple odorants: for example, the smell of coffee is a combination of about 650 odorants. Olfaction is one of the most primitive senses. Humans have evolved to rely less on olfaction than other animals do, and as a result the number of olfactory receptors and the percentage of cortex dedicated to olfaction in humans is smaller than those of other animals. Still, about 3% of the human genome is dedicated to coding for olfactory receptor genes. Humans are able to detect more than 10,000 different odorants, even at very low concentrations. Olfactory sensory neurons (OSNs) are embedded in the olfactory epithelium, which is a small area of tissue located on the roof of the nasal cavity and the upper portion of the nasal septum. These neurons are unique relative to other types of sensory neurons because they only live for one to two months and are continuously replaced from basal stem cells. The dendrite of an OSN has many hair-like structures called cilia, which protrude into the mucus that covers the olfactory epithelium. The olfactory receptors are located on the cilia of OSN dendrites. Molecules of an odorant enter the nasal cavity and bind to these olfactory receptors. These are G-protein coupled receptors, meaning that the binding of a specific odorant initiates a cascade of events that leads to depolarization of the OSNs. There are many different types of olfactory receptors. All of the cilia on the dendrite of an individual OSN contain only one type of olfactory receptor. However, since there are thousands of odorants in the environment, it would be extremely difficult for the olfactory system to have a receptor for every odorant. Instead, each type of receptor can bind multiple types of odorants, and each odorant can bind to multiple types of receptors, illustrated by the picture below. Therefore, each odorant is detected by a unique combination of receptors. Since each OSN contains only one type of olfactory receptor, individual odorants are encoded by a particular combination of OSN activity. Depolarized OSNs generate action potentials which transmit odorant information to the brain. The axons of OSNs comprise the olfactory nerve, cranial nerve I. The olfactory nerve passes through small holes in a bone that separates the nasal cavity from the brain, called the cribriform plate, and terminates in the ipsilateral olfactory bulb in the cortex. The olfactory bulb is visible when viewing the underside of the brain. Olfactory nerve fibers terminate on mitral cells in the olfactory bulb. These neurons refine olfactory information. The axons of OSNs and the dendrites of mitral cells form spherical structures inside the olfactory bulb called glomeruli. All mitral cells in an individual glomerulus receive information from only one type of olfactory receptor. Since a single odorant activates multiple types of olfactory receptors, an odorant is coded in the olfactory bulb by a unique combination of activated mitral cells. This allows the neural coding of an odorant to be preserved along the olfactory pathway and maintained for very long periods of time - we are still able to recognize odorants that we have not experienced in years. Unlike other sensory systems, olfactory information does not travel through the thalamus before reaching primary sensory cortex. Mitral cells in the olfactory bulb project through the olfactory tract directly to ipsilateral primary olfactory cortex. Primary olfactory cortex is defined as regions of cortex that receive direct input from the olfactory bulb. Several regions of cortex meet this criterion, including parts of the amygdala and entorhinal cortex, which is why odors can elicit emotional responses as well as trigger memories that are associated with that particular smell. These responses must be conditioned, however – such connections are formed in primary olfactory cortex when an odorant is detected simultaneously with a specific experienced event. Piriform cortex is the region of primary olfactory cortex that is considered to be the main olfactory processing area. This region is paleocortex, meaning it is evolutionarily older than other cortical regions and only has three layers. Similar to the odorant coding in the olfactory epithelium and the olfactory bulb, neurons in piriform cortex are activated by more than one odorant, and one odorant activates a distinct pattern of neurons. However, unlike other primary sensory cortical areas, there is no known topographical organization in primary olfactory cortex. Neurons responding to specific odorants seem to be scattered randomly throughout piriform cortex. The primary olfactory cortical areas also send olfactory information the hypothalamus, which controls hunger, and to the orbitofrontal cortex, which is thought to play a role in odor discrimination as well as the integration of sensory information from multiple modalities. Neurons in orbitofrontal cortex have been found to respond to several different attributes of an object, such as the smell, sight, and taste of a fruit (Rolls & Baylis, 1994). The Gustatory System Objectives Be able to define the evolutionary purpose of the gustatory system Be able to describe the main pathways that tastant information takes through the gustatory system, from the mouth to gustatory cortex Be able to name the five categories of tastants that we can detect Be able to label the components of a taste bud Be able to describe the function of taste cells in tastant transduction The gustatory system allows us to perceive taste by transducing the chemicals that are present in the foods and beverages that we consume. Historically, the sense of taste has been especially important for survival - it encourages ingestion of the foods that help sustain life and discourages consumption of rotten or poisonous substances. Our perception of a food’s flavor, however, does not come purely from gustation. Flavor is a combination of gustatory, olfactory, and somatosensory inputs. In fact, a diminished ability to smell decreases the flavor that is perceived, causing food to seem bland. A tastant is any substance that excites the gustatory receptor cells. We can detect five types of tastants: sweet, salty, sour, bitter, and umami. The table below gives examples of the items we that consume that would fall into each category of tastant. Foods that taste sweet have lots of calories and foods that are umami contain protein, making these types of tastants important for survival. Conversely, poisonous plants taste bitter and spoiled food tastes sour, and thus consuming these tastants could be fatal. As a result, the taste of sweet and umami foods is more enjoyable than that of bitter or sour foods. This feature of the gustatory system has directed animals and early humans to eat the foods that will keep them alive and avoid eating foods that are potentially dangerous. The sensory receptors of the gustatory system are neuro-epithelial taste cells which are located in structures called taste buds. Most taste buds are on the tongue but some are also located on the palate, pharynx, epiglottis, and the upper part of the esophagus, labeled in the picture below. http://www.mayoclinic.com Each taste bud contains about 50 -100 taste cells. Similar to olfactory sensory neurons, taste cells only live for a short time and are continuously replaced by basal stem cells. The tastant receptors are located on tiny hair-like structures called microvilli that extend from the top of each taste cell through a hole in the taste bud called the taste pore. The microvilli, and thus the gustatory receptors, come into contact with food molecules dissolved in saliva on the surface of the tongue. ’ It was previously thought that each taste cell responds to one class of tastant, but new evidence suggests that individual taste cells are excited by multiple tastant categories. The coding of specific tastants in the gustatory system is therefore not yet fully known. Interaction with particular tastants depolarizes the taste cell, which then releases neurotransmitter. Each taste cell is innervated by fibers of peripheral branches of one of three cranial nerves. The facial nerve (CN VII) innervates taste cells on the anterior 2/3 of the tongue and the soft palate, the glossopharyngeal nerve (CN IX) innervates taste cells on the posterior 1/3 of the tongue and the pharynx, and the vagus nerve (CN X) innervates taste cells located on the epiglottis and the upper esophagus. The cell bodies of these peripheral cranial nerve fibers located are in the geniculate ganglion (for the facial nerve), the petrosal ganglion (for the glossopharyngeal nerve), and the nodose ganglion (for the vagus nerve), as shown in the picture below. Since taste cells synapse onto the dendrites of these peripheral ganglia neurons, the release of neurotransmitter by taste cells initiates action potentials in these neurons. The neurons in either the geniculate, petrosal, or nodose ganglia are activated depending on the location of the taste cell in the mouth. Despite the different locations of the peripheral ganglia, the axons of peripheral ganglion neurons all project to the ipsilateral nucleus of the solitary tract in the medulla. Projections from neurons in the nucleus of the solitary tract travel through the central tegmental tract to the ipsilateral ventral posteromedial nucleus (VPM) of the thalamus. VPM neurons then send gustatory information to ipsilateral primary gustatory cortex, which is located on the anterior insula and frontal operculum. Some neurons in gustatory cortex have been found to respond to only one particular class of tastant while others respond to multiple tastant types. Due to these distinctive responses, neurons in gustatory cortex are thought to encode both innate responses to tastants as well as blends of flavors. Gustatory cortex neurons send information to the orbitofrontal cortex, which integrates taste with other sensory information. The hypothalamus, amygdala, and hippocampus also receive gustatory information through pathways that are separate from the main gustatory pathways discussed here. The picture below shows the location of primary olfactory and gustatory cortices in relation to other areas of the cortex. Notice that you need to pull away parts of the lateral cortex in order to view the entire olfactory and gustatory areas.