Supplementary Tables (doc 112K)
advertisement
")
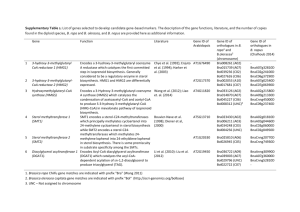
Supplementary Tables Table S1 Primers applied in this study. Primers HvUBQ5-F HvUBQ5-R HvPR1b-F HvPR1b-R HvPR10-F HvPR10-R HvJIP23-F HvJIP23-R HvKS1-F HvKS1-R HvKSL4-F HvKSL4-R HvDXS-F HvDXS-R TaAlpha-tubulin-F TaAlpha-tubulin-R ITS _Rh-F ITS_ Rh-R AtPDF1.2-F AtPDF1.2-R AtPR1-F AtPR1-R AtERF1-F AtERF1-R AtUBQ5-4-F AtUBQ5-4-R AtVSP2-F AtVSP2-R Sequence 5`- 3` ACCCTCGCCGACTACAACAT CAGTAGTGGCGGTCGAAGTG GGACTACGACTACGGCTCCA GGCTCGTAGTTGCAGGTGAT GGAGGGCGACAAGGTAAGTG CGTCCAGCCTCTCGTACTCT GGAGTGTTTGGTACCCCCAT GGCACCAGTGGCATTGTAGA GACAGCCAAGGCTTTGAGAG TGCACATCTTCCAGAACAGC CGTCACCTTCTCCGAGACAT GACGAACCTTCCTTGGGAGT ACTTGCAGTGTTGGCACAAG CACCAAACCCTCCTGTGACT ATCTCCAACTCCACCAGTGTCG TCATCGCCCTCATCACCGTC TCAGCACATAACCACACCAATCGCG TGCTTTGTACGCTCGGTAAGAAGGG GTTTGCTTCCATCATCACCC GGGACGTAACAGATACACTTG AAGAGGCAACTGCAGACTCA TCTCGCTAACCCACATGTTC CGAGAAGCTCGGGTGGTAGT GCCGTGCATCCTTTTCC CCAAGCCGAAGAAGATCAAG ATGACTCGCCATGAAAGTCC CAAACTAAACAATAAACCATACCATAA GCCAAGAGCAAGAGAAGTGA 5 1 Reference Imani et al. (2011) Imani et al. (2011) Schäfer et al. (2009) Schäfer et al. (2009) Schäfer et al. (2009) Schäfer et al. (2009) Schäfer et al. (2009) Schäfer et al. (2009) Li (2012) Li (2012) Li (2012) Li (2012) this study this study Wen et al. (2013) Wen et al. (2013) Sharma et al. (2008) Sharma et al. (2008) Schikora et al. (2012) Schikora et al. (2012) Trusov et al. (2009) Trusov et al. (2009) Trusov et al. (2009) Trusov et al. (2009) Jacobs et al. (2011) Jacobs et al. (2011) Jacobs et al. (2011) Jacobs et al. (2011) Table S2 Comparison of the gene content of the genomes of RrF4 and C58 performed in the EDGAR software (Blom et al., 2009). Presence of genomovar G8 specific gene clusters (SpG8 gene clusters) as described by Lassalle et al. (2011). Gene C58 CDSs1 cluster G8-specific regions SpG8-1a Atu1398 - Atu1408 SpG8-1b SpG8-2a SpG8-2b Atu1409 - Atu1423 Atu3054 - Atu3059 Atu3069 - Atu3073 SpG8-3 Atu3663 - Atu3691 SpG8-4 SpG8-5 Atu3808 - Atu3830 Atu3947 - Atu3952 SpG8-6a SpG8-6b Atu4196 - Atu4206 Atu4213 - Atu4221 SpG8-7a Atu4285 - Atu4294 SpG8-7b Atu4295 - Atu4307 Specific to G58 sporadic in G8 Cluster 1 Cluster 2 Atu1183 - Atu1194 Atu4606 – Atu4615 Cluster 3 Atu4864 – Atu4896 Prophages, sporadic in G8 ATPP-1 Atu0436-Atu0471 ATPP-2 Atu1183 - Atu1194 ATPP-3 Atu3831 – Atu3858 Predicted function1 RrF4 CDSs SY94_1294 - SY94_1304 Sugar and amino acid transport; sugar metabolism SY94_1305- SY94_1319 Ferulic acid uptake and catabolism SY94_3050 - SY94_3054 Curdlan EPS biosynthesis SY94_3064 - SY94_3068 Secondary metabolite biosynthesis Siderophore biosynthesis; ironSY94_3658 - SY94_3682 siderophore uptake Ribose transport; monosaccharide SY94_3809 - SY94_3828 catabolism and SY94_3921 - SY94_3925 Opine-like compounds catabolism Drug/toxic (tetracycline) SY94_4165 - SY94_4174 resistance SY94_4183 - SY94_4190 Drug/toxic resistance SY94_4254 - Environmental signal SY94_4263 sensing/transduction Environmental signal SY94_4264 - SY94_4276 sensing/transduction SY94_1089 - Prophage SY94_10932 SY94_4568 - SY94_4574 Lipopolysaccharide biosynthesis SY94_4815 SY94_48353 Absent Prophage SY94_1089- SY94_10932 Prophage SY94_3831 - Prophage SY94_38372 10 1 Adapted from Lassalle et al. (2011) 2 C58 homologous genes are only partially present 3 C58 homologous genes partially present and rearranged 2 15 Table S3 Overview of chromosomally coded genes of RrF4 that encode proteins/enzymes which are potentially involved in the interaction of RrF4 with host plants and belong to functional groups of genes which are well known to be involved in the beneficial effect of PGPRs (e.g. as reviews by Lugtenberg and Kamilova, 2009). The gene homologs of C58 are 20 also given. Predicted functions of genes detected in the circular and linear chromosomes were based on literature data, based on genome analysis of C58 (Goodner et al., 2001; Wood et al., 2001), or KEGG passed analysis performed in GenDB (Meyer et al., 2003). The functionality of gene was not proofed experimentally. Location of RrF4´s genes: CDS <SY94_3000 are located at the circular chromosome; CDSs between SY94_ 3000 and SY94_ 5000 are located 25 on the linear chromosome. Gene RrF4 CDS C58 CDS Predicted function Reference of experimental evidence* Matrix components for attachment and biofilm ctpABCDEFGHI SY94_0156SY94_0148 Atu0224Atu0216 Flp-type pili: form thin Wang et al. filaments, thinner than flagella, (2014) arranged around the cell surface and sheeted into the external milieu celABCG and celDE SY94_3304SY94_3302, SY94_3301; Atu3309Atu3307, Atu8186; Cellulose synthesis gene (two opera); Involved in surface attachment; and binding of plant cells Matthysse (1995); Matthysse et al. (2005) SY94_3298SY94_3299 Atu3302Atu3303 uppABCDEF SY94_1137SY94_1142 Atu1235Atu1240 Polysaccharide biosynthesis cluster - UPP: Unipolar polysaccharide (required for surface binding and biofilm formation) Xu et al. (2013) exoC (pgm) SY94_4051 Atu4074 Required for the synthesis of extracellular ß-1, 2-glucan and succinoglucan polysaccharides (phosphoglucomutase) Changelosi et al. (1987); Bahat-Samet et al. (2004) 3 chvAB SY94_2559, SY94_2561 Atu2728; Atu2730 Required for synthesis of extracellular beta-1,2-glucan – involved in binding of plant cells; Cangelosi et al. (1987) chvA: beta-1, 2-glucan export ATP-binding protein; chvB: beta-1, 2-glucan biosynthesis protein SY94_3050- Atu3054– Atu3059 (SpG8-2a1) Curdlan (beta-1,3-glycan) EPS biosynthesis Lassalle et al. (2011) Atu3056 in SpG8-2a Putative beta-1,3-glucan synthase (curdlan synthase, CrdS) Lassalle et al. (2011) Siderophore biosynthesis; Rondon et al. (2004) SY94_3682 Atu3663– Atu3691 (SpG8-31) SY94_3675 SY94_3682 Atu3684– Atu3691 Homologous to fecABCDE genes involved in TonB dependent reuptake of the siderophores if chelated to iron Braun et al. (2006) SY94_3675 Atu3684, Atu3692, FecAIR seemed to form a cell surface signaling system, whose homolog in Escherichia coli was proposed to regulate the whole system of biosynthesis, release, and reuptake of siderophores Braun et al. (2006) SY94_3054 SY94_3052 Siderophore biosynthesis SY94_3680- SY94_3683 Atu3693 SY94_3684 iron-siderophore uptake Metabolism (nutrient uptake and metabolism/catabolism) SY94_1294 SY94_1304 Atu1398– Atu1408 (SpG8-1a1) Sugar and amino acid transport; sugar metabolism Lassalle et al. (2011) SY94_3809 SY94_3828 Atu3808– Atu3830 (SpG8-41) Ribose transport; monosaccharide catabolism and carbohydrate metabolism Lassalle et al. (2011), but without experimental proof SY94_1305 SY94_1319 Atu1409– Atu1423 Ferulic acid uptake and catabolism (uptake and catabolism of plant derived Lassalle et al. (2011) 4 β-ketoadipate pathway Linear chromosome (SpG8-1b1) phenolic compounds) Present at the linear chromosome Pathway for aromatic compound degradation (degradation of plant derived compounds) Goodner et al. (2001) and references therein Cell wall degradation, degradation of cellulose, hemicellulose, pectin, and/or lignin Pectinase (kdgF) SY94_3140 Atu3145 Pectine degrading protein Wood et al. (2001) Ligninase (ligE) SY94_1026 Atu1121 Lignin-degrading protein Wood et al. (2001) Beta-endoglucanase (celC) SY94_3302 Atu3307 Encoded within the cellulose synthesis gene cluster Goodner et al. (2001) (linear chr.) Xylanase (xynA) SY94_2222 Atu2371 Endo-1,4-beta-xylanase Regulators of pectinase and cellulase (pecS/M) SY94_0202/ SY94_0203 Atu0272/ Atu0273 transcriptional regulator, MarR family 1 G8-specific regions defined by Lassalle et al. (2011). * not tested/proofed for RrF4 5 30 Table S4 Quantification of root colonization by RrF4 using qPCR targeting the ITS of the ribosomal operon. RrF4 cells were calculated per g root FW (three ITS targets per cell). Data represent mean values and standard deviations (in brackets) of three independent biological replicates. The upper 4 cm of roots were investigated. FW: fresh weight; nd: under detection 35 limit. Time Barley Wheat -1 [dpi] [cells g FW ] 0 2.1 (±0.4) × 10 5 1.4 (±0.3) × 10 7 2.1 (±0.7) × 10 14 8.1 (±1.4) × 10 Arabidopsis -1 [cells g FW-1] [cells g FW ] 8 9 9 9 3. 3 (±2.0) × 10 1.4 (±0.2) × 10 3.1 (±0.6) × 10 6.0 (±2.5) × 10 8 9 9 9 nd 5.0 (±0.3) × 10 1.2 (±0.6) × 10 2.9 (±0.5) × 10 8 9 9 References: Bahat Samet E, Castro-Sowinski S, Okon Y (2004). Arabinose content of extracellular polysaccharide 40 plays a role in cell aggregation of Azospirillum brasilense. FEMS Microbiol Lett 237:195-203. Braun V, Mahren S, Sauter A. (2006). Gene regulation by transmembrane signaling. Biometals. 19:103–113. Cangelosi GA, Hung L, Puvanesarajah V, Stacey G, Ozga DA, Leigh JA, Nester EW. (1987). Common loci for Agrobacterium tumefaciens and Rhizobium meliloti exopolysaccharide synthesis and 45 their roles in plant interactions. J Bacteriol 169:2086-2091. 6 Lassalle F, Campillo T, Vial L, Baude J, Costechareyre D, Chapulliot D et al. (2011). Genomic species are ecological species as revealed by comparative genomics in Agrobacterium tumefaciens. Genome Biol Evol 3:762–781. Lugtenberg B, Kamilova F. (2009). Plant-growth-promoting rhizobacteria. Annu Rev Microbiol. 63: 50 541-556. Matthysse AG, Marry M, Krall L, Kaye M, Ramey BE, Fuqua C. et. al. (2005). The effect of cellulose overproduction on binding and biofilm formation on roots by Agrobacterium tumefaciens. Mol.Plant Microbe Interact. 18:1002–1010. Matthysse AG, White S, Lightfoot R (1995). Genes required for cellulose synthesis in Agrobacterium 55 tumefaciens. J. Bacteriol. 177:1069–1075. Meyer F, Goesmann A, McHardy AC, Bartels D, Bekel T, Clausen J et al. (2003). GenDB--an open source genome annotation system for prokaryote genomes. Nucleic Acids Res 31:2187-2195. Rondon MR, Ballering KS, Thomas MG. (2004). Identification and analysis of a siderophore biosynthetic gene cluster from Agrobacterium tumefaciens C58. Microbiology. 150:3857–3866. 60 Wang Y, Haitjema CH, Fuqua C (2014). The Ctp type IVb pilus locus of Agrobacterium tumefaciens directs formation of the common pili and contributes to reversible surface attachment. J Bacteriol. 196:2979-2988. Wood DW et al. (2001). The genome of the natural genetic engineer Agrobacterium tumefaciens C58. Science. 294:2317–2323. 65 Xu J, Kim J, Koestler BJ, Choi JH, Waters CM, Fuqua C. (2013). Genetic analysis of Agrobacterium tumefaciens unipolar polysaccharide production reveals complex integrated control of the motile-tosessile switch. Mol Microbiol. 89:929-948. 7