Chapter 23 SEED PLANTS II: ANGIOSPERMS
advertisement

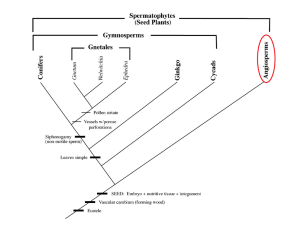
Chapter 23 SEED PLANTS: ANGIOSPERMS All flowering plants are classified in a single Division, Magnoliophyta. Angiosperms are commonly known as “flowering plants.” This division contains the greatest number of living species, about 257,000. They are characterized by producing flowers, a synapomorphy that unifies them. First appeared in the Cretaceous about 130 million years ago. The angiosperms have developed numerous methods of interaction with animals that are beneficial to both. They have developed many antiherbivore compounds that defend them against plant eating animals and parasites. CHARACTERISTICS OF ANGIOSPERMS Woody or herbaceous. Presence of vessels. Axial parenchyma; parenchyma cells mixed with vessels and tracheids. Produce flowers. Pollinated by wind or animals. Double fertilization: egg + sperm embryo and 2 polar nuclei + sperm endosperm. Seeds enclosed in a fruit. EVOLUTION OF THE FLOWER Angiosperm stamens and carpels developed from the gymnosperm sporophylls. Gymnosperm sporophylls are flat and spirally arranged, and microsporophylls never occur with megasporophylls. Flowering plants with flat spirally arranged flower parts are thought to be most ancient. Basal angiosperms have flat stamens without distinct filament and anther portions, arranged in spirals, and their sporogenous tissues (microspore mother cells) form relatively large, prominent internal masses. Amborella, Nymphaeales, Austrobaileyales, and Magnoliales. In most angiosperm carpels, the edges of sporophyll grow shut against one another, sometimes leaving a suture, sometimes not. These are called closed carpels. Closed carpels develop into fruits that enclose the embryos as they develop into seeds. Carpels in the basal angiosperms are leaf-like; resembling young leaves whose blades have not yet opened. Basal angiosperms lack stigma and style; instead, the ovary edges have rows of secretory hairs that secrete a thick liquid that both seals the seam and functions as a stigmatic surface. The stigmatic surface acts as a selector of pollen grains eliminating pollen from another species or preventing self-fertilization. THE FLOWER The flower is a reproductive shoot or branch. It has four parts arranged in whorls or circles on a stalk or peduncle. The parts of the flower are the sepals (calyx), petals (corolla), stamens and carpels. Stamens consist of a filament and an anther. Carpels are also referred to as pistils. They consist of an ovary, a style and a stigma. Flowers may be borne singly or in clusters called inflorescence. Flower parts are considered modified leaves. The ability to produce bisexual flowers must have occurred very early. All flowering plant clades including basal angiosperms have the megasporophylls located above the microsporophylls on a single axis. The fusion of carpels into a single structure (pistil) and the fusion of petals into one corolla (sympetaly), and floral parts that are bilaterally symmetrical rather than radially symmetrical (zygomorphy). DOUBLE FERTILIZATION It is characteristic of flowering plants and universally present. Double fertilization results in the formation of a diploid zygote and a triploid endosperm. The female gametophyte or embryo sac has an egg nucleus and two polar nuclei. One sperm fertilizes the egg nucleus and forms the zygote, 2n. Another sperm joins the two polar nuclei forming the triploid (3n) nutritive tissue called the endosperm. Double fertilization also occurs in gnetophytes but there is a disagreement if it is a case of a shared ancestral feature (symplesiomorphy) or convergent evolution (homoplasy). OTHER ANGIOSPERM APOMORPHIES Study Table 23.1 on page 526 of your textbook. 1. Vessel elements possibly evolved after the flowers because two basal species, Amborella and Drimys, lack them. Lack of vessels was thought to be a primitive characteristic of some basal angiosperms. Other features, however, were inconsistent with the idea that these were primitive plants. The current hypothesis is that Amborella is primitive (primitively vesselless) and never had vessels, while others arose after vessels had developed and then lost them. These are referred to as secondarily vesselless. 2. Sieve tubes probably originated next. Several species still have sieve cells in their phloem. 3. All gymnosperms, seed ferns and basal angiosperms are or were woody plants. The wood of basal angiosperms has many gymnosperms characteristics. Ancestral flowering plants were woody perennial in all probability. 4. The herbaceous habit resulted from the loss of vascular cambium, and this has occurred in several clades. 5. The annual habit is a unique angiosperm characteristic. Some groups of angiosperms like grasses, bromeliads and orchids are still changing and evolving. CHANGING CONCEPTS ABOUT EARLY ANGIOSPERMS C.E. Bessey developed the hypothesis of the ranalean flower about 100 years ago. This generalized type of flower has: 1. All parts, sepals, petals, stamens and carpels. 2. These are arranged spirally. 3. The carpels are superior, that is above all other flower parts. This is the Magnolia-type of flower. Most botanists consider the angiosperms to be monophyletic. The transition from gymnosperms to angiosperm it is believe to have happened in the Jurassic (200-145 m.y.a.) and Lower Cretaceous (~135 m.y.a.) periods of the Mesozoic Era. The earliest fossils to be considered those of angiosperms are from the Lower Cretaceous (~130 m.y.a.). They represent both dicot and monocot leaves. These fossils are rare making about 2% of the plant fossils. In the Upper Cretaceous (ending about 65 m.y.a.), angiosperm fossils are more common, outnumbering the fossils of gymnosperms and ferns. Upper Cretaceous and Tertiary flora contains fossils that resemble modern genera but are not identical. The oldest angiosperm wood comes the Aptian Epoch of Japan (~125 m.y.a.). Flowers and fruits occurred in the Lower Cretaceous. CLASSIFICATION OF FLOWERING PLANTS Flowering plants used to be divided into two groups: monocots and dicots. This division has been largely abandoned because of new information that has become available mainly through molecular and cytological studies. In general, monocots can be recognized by the following traits: MONOCOTS have floral parts in multiples of three and the seed contains one cotyledon. The endosperm provides the food for the embryo. Venation is usually parallel (there are exceptions). Their vascular bundles are scattered throughout the ground tissue. The root system is fibrous. DICOTS have floral parts in multiples of four or five, and their seeds contain two cotyledons. The cotyledons usually absorb the food from the endosperm first, and then provide the food for the embryo. Venation is netted. The vascular bundles in the stem cross-section are arranged in circles (rings). They usually have a taproot system for at leas part of their life. Current research supports the hypothesis that monocots are monophyletic and form a single clade. The dicots, however, appear to be a polyphyletic grouping. Most of the dicots belong to the eudicots. Three lineages of the remaining dicots form what is called the basal angiosperms because they include what are considered to be the oldest lineages. Another group is called the magnoliids. MONOCOTS have floral parts in multiples of three and the seed contains one cotyledon. The endosperm provides the food for the embryo. Venation is usually parallel (there are exceptions). Their vascular bundles are scattered throughout the ground tissue. The root system is fibrous. DICOTS have floral parts in multiples of four or five, and their seeds contain two cotyledons. The cotyledons usually absorb the food from the endosperm first, and then provide the food for the embryo. Venation is netted. The vascular bundles in the stem cross-section are arranged in circles (rings). They usually have a taproot system for at leas part of their life. Current research supports the hypothesis that monocots are monophyletic and form a single clade. The dicots, however, appear to be a polyphyletic grouping. Most of the dicots belong to the eudicots. Three lineages of the remaining dicots form what is called the basal angiosperms because they include what are considered to be the oldest lineages. Another group is called the magnoliids. MONOCOTS have floral parts in multiples of three and the seed contains one cotyledon. The endosperm provides the food for the embryo. Venation is usually parallel (there are exceptions). Their vascular bundles are scattered throughout the ground tissue. The root system is fibrous. DICOTS have floral parts in multiples of four or five, and their seeds contain two cotyledons. The cotyledons usually absorb the food from the endosperm first, and then provide the food for the embryo. Venation is netted. The vascular bundles in the stem cross-section are arranged in circles (rings). They usually have a taproot system for at leas part of their life. Current research supports the hypothesis that monocots are monophyletic and form a single clade. The dicots, however, appear to be a polyphyletic grouping. Most of the dicots belong to the eudicots. Three lineages of the remaining dicots form what is called the basal angiosperms because they include what are considered to be the oldest lineages. Another group is called the magnoliids. Amborella and Austrobaileyales greatly resemble eudicots; however, they cannot be classified as eudicots because the common ancestor that included both of them the eudicots also includes the monocots. See fig.23.9. The same would be the case with the Nymphaeales. BASAL ANGIOSPERMS Basal angiosperms originated when the angiosperms were still a young clade. Their ancestors became reproductively isolated from the other early flowering plants before distinctive angiosperm traits had originated. These groups continued to evolve as the other clade evolved. Amborellaceae, Nymphaeaceae and Austrobaileyales are the extant descendants of that original divergence. The Nyphaeaceae are the water lilies. They lack wood, are pollinated by animals and their vascular bundles are scattered like those of the monocots. Amborella trichopoda is the only species in this clade. They are trees in New Caledonia; their wood lacks vessels and has little parenchyma. The species is dioecious, with staminate plants and carpel late plants with staminate, atrophied stamens. This indicates that they evolved from bisexual plants. Both flowers are small with 5 to 11 tepals (not differentiated into sepals and petals). Staminate flowers have numerous stamens; carpellate flowers have 5 or 6 carpels whose edges do not seal tightly but instead have interlocking, secretory hairs that act as a stigma. Austrobaileyales are trees with bisexual flowers, with stamens and carpels being similar to those of Amborella. All these three have evolved considerably since their clade originated. The magnoliids clade evolved later than Amborella and Austrobaileyales according to DNA evidence. The morphology of the magnoliids is not very difference from these two earlier clades. The family Magnoliaceae lacks vessels, fibers and axial parenchyma in their wood. Magnolia has many stamens and carpels arranged in spiral. The carpels are not fused together into a pistil. The pollen has a single germination pore (uniaperturate). MONOCOTS It is commonly accepted that monocots evolved from early angiosperms between 80 and 100 m.y.a., perhaps even 120 m.y.a. Their ancestors were probably herbs with no or little cambial activity. The gynoecia consist usually of three carpels that are either free or slightly fused. The perianth usually consists of three outer and three inner members and often looks very similar than they are called tepals. Tepals rarely fuse together to form a tube; they remain free. Nymphaeales have similar features. Nymphaeales and monocots may have a common ancestor. Parallel venation may have arisen from aquatic ancestors that have broad leaves. These leaves became reduced and lost the lamina, a condition better adapted to submerged life. Some of these plants moved into drier habitats where their leaves were exposed to the air and a broad or long lamina would be advantageous. Basal meristem of leaves and in some, marginal meristem evolved. It was located at the end of the residual leaf, resulting in the formation of what appears to be a petiole and lamina. ALISMATALES Many members of the Alismatales are aquatic herbs found in swamps and marshes, partly or entirely submerged. Air chambers make the plants buoyant; mutations that prevent the formation of lignified fibers are advantageous. They have very little xylem and sclerenchyma. Tissues have been lost. Emergent members of the Alismataceae are less modified because their bodies are partially or entirely exposed during part of the year. These plants have considerable amount of fibers and a thick cuticle. The flowers of the Alismatales are large and showy, with three sepals and petals. Flowers that do not emerge from the water are highly modified and lack perianth. Family Araceae This family has a very distinct inflorescence. Tiny flowers, either unisexual or perfect, develop embedded in a thick axis called spadix. Staminate flowers grow on upper part of the spadix, and carpellate flowers below. A bract, the spathe, protects the entire inflorescence. Several common houseplants belong to this family: Philodendron, Anthurium, and Dieffenbachia (dumb cane). LILIALES Recent DNA evidence indicates that Liliales is a small clade with approximately 11 families and about 1300 species. The Liliales are characterized by the presence of spots or lines on the petals and nectaries present at the base of tepals or sepals. Family Liliaceae This family contains many ornamental plants with bulbs. Some of the common members are the lilies and tulips. Family Colchicaceae The flame lily and Colchicum belong to this family. Colchicum produces the alkaloid colchicine that prevents microtubule polymerization. ASPARAGALES The carpels of many Asparagales fuse side by side starting at their bases, the fused sides being called. Septa. They do not fuse all the way up to the style, however, and the open areas secrete nectar; these are called septal nectaries. This group has a very diverse morphology: onions, orchids, hyacinths, irises, yuccas, and daffodils. They have also diversified biochemically. Onions contain sulfur compounds; the Amaryllidaceae produces alkaloids. Family Orchidaceae This is the largest and most diverse family of the Asparagales. Many are epiphytic or terrestrial; one species is a subterranean parasite. Their zygomorphic flowers are large, colorful and showy for the most part. Many types of insect pollination have evolved, many including diverse aromas. The plants produce thousands of dust-like seeds that must form a symbiosis with a fungus in order to survive long enough to form roots and leaves. DIOSCOREALES This order has only one family, Dioscoreaceae. Some species produce a yam, a starchy root that is the staple food in many tropical areas like Polynesia. This family has reticulate venation and is often mistaken for dicots. COMMELINOID MONOCOTS The following fours orders are called “commelinoid” because they share several unusual synapomorphies. 1. 2. 3. 4. 5. Unique type of epicuticular wax. Walls with unusual type of hemicelluloses and ultraviolet-fluorescent compounds. Pollen contains starch. Endosperm contains starch. Molecular studies of rbcL, atpB, and other DNA regions support this clade. Arecales The family Arecaceae contains the palms. The old name of the family is Palmae. There are about 3500 species of palms. Solitary trunk, which varies from 1 m wide to 1 cm. Scattered vascular bundles. Leaves near the shoot of the apex. Leaves pinnate or fan-shaped. Some species have a prostrate trunk or are vines. Poales This order contains the grasses, cattails, sedges and bromeliads. The family Poaceae (old name Graminae) contains about 8,000 species. They are one of the most important families from the economic point of view. It includes the most important staple foods of humans: wheat, rice, corn, barley, oats, sorghum, rye, and sugar cane. They also include the bamboos, an important building material in the tropics. Grasses abound in open, flat and dry regions of the world: North America’s prairies, steppes of Ukraine and Russia and the Argentinean pampas Grasses form tight clumps or produce rhizomes. Meristem is located near the ground protected from fire and grazing animals. Wind pollinated. Reduced sepals and petals. Three fused carpels with separate stigma each. One ovule present in the ovary. Seed are produced with a tight coat fused firmly to the fruit wall; this fruit is called a caryopsis. The sedges belong to the family Cyperaceae, with about 4,000 species. The family Juncaceae contains the rushes. Species of both families are common in marshy areas. They produce tiny flowers and small dry fruits. Sedges stems are triangular; rushed stems are round and solid; those of grasses are round and hollow. Cattails belong to the family Typhaceae, and bromeliads to the Bromeliaceae. Cattails grow in ponds and marshy areas. They produce an underground stem called a rhizome, with axillary buds that produce more rhizomes. Other axillary buds grow upwards and produce the familiar cattail plant, which in reality are only a single branch and not the whole plant. Their inflorescence consists of many staminate flowers and thousand of carpellate flowers. The Bromeliaceae contain some of the most beautiful tropical epiphytes with brightly colored inflorescence. Some species occur in the subtropics, Florida and Texas, forming the familiar Spanish and ball mosses. Some species are terrestrial and often xerophytic, like the pineapple and the Puya of Peru. The terrestrial habit is considered to be relictual. This is a typical New World family that probably evolved after South America separated from Africa about 80 million years ago. Had they evolved earlier, there would be species in Africa. Zingiberales Many houseplants belong to this order: bananas, cannas, gingers, Maranta, etc. The flowers of the Zingiberales are large and showy pollinated by insects, birds or bats. Flower sepals and petals are fused into tubes, and are bilaterally symmetrical. The gynoecium consists of three fused carpels and often inferior to sepals and petals. The family Zingiberaceae has bout 1,000 species. Most species in this family are tropical and herbaceous. The leaves are broad and have a petiole. These plants grow in heavy shade where broad leaves are advantageous. EUDICOTS The eudicots is a large clade with many different orders and families. 1. They are characterized by their tricolpate pollen, that is, the pollen grain has three germination pores. 2. Their flower parts are arranged in whorls and not spirally. 3. The stamens usually have a well-defined filament and anther. BASAL EUDICOTS The Ranunculales, Proteales and Trochodendraceae are thought to have diverged early in the evolution of the eudicots. The family Ranunculaceae has flowers whose parts have very few fused parts. Buttercups (Anemone) and Clematis belong here. 1. Many stamens and carpels. 2. Stamens may fuse together into a tube or even fuse to the carpels. 3. Carpels remain free. The family Papaveraceae include the poppies. This family has numerous ornamental species. They either herbs or shrubs with very soft wood. Papaver somniferum is the opium poppy, the source of morphine and codeine. The hammamelid families are now believed to basal eudicots. They have dangling inflorescences with many staminate flowers and a few carpellate flowers. The family Platanaceae includes the sycamores. The sycamores produce tiny flowers that mature into spherical clusters. Caryophyllales Molecular evidence has been used to combine a large number of families into an order called Caryophyllales. Many of these families were considered in the past to be closely related and are referred now as “core Caryophyllales.” Some the “core Caryophyllales” are the Cactaceae (cactus), Aizoaceae (iceplant), portulaca (Portulacaceae), bougainvillea and four-o’clocks (Nygtagenaceae), spinach and beets (Amaranthaceae), and carnations and chickweed (Caryophyllaceae). The “core Caryophyllales” share many derived characters but one is particularly important. They produce a group of water-soluble pigments called betalains that are deposited in their flowers. Other flowering plants produce instead anthocyanin pigments. Many “core Caryophyllales” lack petals in their flowers. Some have petals that are thought to be modified sepals or stamens. Their endosperm develops only a little and then stops growing. Instead, the nucellus cells proliferate and form a nutritive tissue called perisperm, which surrounds the developing embryo. By the time the seed matures, the perisperm is absorbed almost completely. Another distinguishing characteristic that unifies this order is the nature of the sieve tube plastids. “Core Caryophyllales” phloem plastids contain deposits of fibrous protein located as a ring just interior to the plastid membrane. In other angiosperms, plastids contain either starch or crystalline protein with a different structure. The Caryophyllales possibly arose from ancestors similar to the Ranunculaceae. The Caryophyllales are herbaceous with no wood, very little wood or abnormal wood, so the ancestor is thought to have been herbaceous or shrubby that had lost the ability to produce extensive secondary growth. They may have evolved around 70 to 80 million years ago, a time of important continental drift. The southern continent of Gondwana existed at the time. The Caryophyllales may have developed in southern Gondwana and spread across the continent. Then continental drift broke Gondwana and Caryophyllales found themselves in South America, India, Antarctica and Australia. Families that evolved after the breakup of Gondwana are confined now to one continent, e.g. Cactaceae found only in the New World; the Didieriaceae arose in east Africa and Madagascar and is found only in that continent. Other families that are now placed in the Caryophyllales are the Polygonaceae (buckwheat), Plumbaginaceae (pokeweed), Droseraceae (Venus-fly-trap) and Nepenthaceae (pitcher plants). Santales This is a small order of highly modified species many of which are parasitic. The sandalwood tree Santalum (Santalaceae) produces fine root connections with other trees and parasitizes them. The mistletoes Viscum and Phoradendron produce chlorophyll and are photosynthetic; they are called hemiparasitic. Parasitic plants without chlorophyll are called holoparasites and depend completely on their host for survival The family Loranthaceae has about 900 species, all of which are parasitic. ROSID CLADE The Rosids are a large clade that includes families with many differences in morphology. It is based mostly on molecular evidence. The Rosids consists of several small orders and two large groups. The Vitales and Geraniales contain species of economic importance. Vitis (Vitaceae) is the grapevine, and the Geraniaceae contains Pelargonium, the geraniums. The two large clades are the fabids (eurosids I) and malvid (eurosids II). These two clades contain more than a hundred families that cannot be unified by morphology. The Rosids are more derived than basal tricolpates and have derived characters that differ from those of Caryophyllales. Rosids lack the highly relictual features found in basal angiosperms. Rosids have pinnately compound leaves. This is considered an ancestral feature. Some Rosids have simple leaves, which is a condition considered derived by suppression of leaflets. Simple leaves are an early, ancestral condition for the angiosperms as a whole, but they are a later derived condition for Rosids. There are 14 orders and over 50,000 species in the rosid clade. The family Rosaceae is very important economically because it contains numerous ornamentals like roses, photinias and mountain ash, but also important fruit trees and shrubs like apples, pears, almonds, plums, peach, apricots and many more. The family Fabaceae, the legume family, is a large family or herbaceous annuals to shrubs, vines and long-live trees. It is an economically important family that produces foods, drugs, dyes and woods. Beans, peas, lentils and peanuts belong to this family. Many species have associations with nitrogen-fixing bacteria and can grow in poor soils and produce protein rich seeds. ASTERID CLADE This is the most derived clade of the angiosperms. It is a sister clade to the rosids. It originated perhaps as recently as 60 million years ago. The asterids can be distinguished by the following three synapomorphies. 1. Sympetalous flowers (petals fused together into a tube). 2. A few stamens, no more than the petal lobes. 3. Stamens alternate with petals. The pollinators of asterids are very specialized. Asterids lack many chemicals found in other groups; they lack betalains (Caryophyllales), bezylisoquinoline alkaloids (magnoliids), ellagic acid or proanthocyanins (rosids). Instead many asterids have iridoid compounds rarely found outside this group. Certain families produce potent chemicals that deter or kill animals. The poison hemlock that killed Socrates came from Conicum, family Apiaceae. The asterids contain the greatest number of species, about 60,000 species, grouped into two small orders, Cornales and Ericales. The rest are grouped in two large groups of orders, lamiids (euasterids I) and campanulids (euasterids II). The family Asteraceae, formerly known as Compositae, contains about one third of the species and is the largest family of eudicots. Sunflowers and dandelions belong here. Other families are the Solanaceae, potato, tomato, peppers, tobacco, eggplant and others; the Lamiaceae includes the mints, thyme and lavender; Convolvulaceae, the morning glory family; and the Asclepiadaceae, the milkweeds. Many asterids are important medicinally. Vinca, family Apocynaceae, produces two potent anticancer drugs, vinblastine and vincristine. The family Rubiaceae includes Coffea produces coffee, and Cinchona, from which the antimalarial drug quinine is extracted; another rubiaceous drug is ipecac from Cephaelis is used to induce vomiting in case of oral poisoning. Digitalis, family Scrophulariaceae, produces a cardiac glycoside to treat heart disease. Lamiales This order is quite derived in floral characters; the flowers tend to be bilaterally symmetrical and of sizes and shapes that permit only certain insects to enter and effect pollination. The family Asteraceae contains 1,100 genera and over 20,000 species distributed worldwide and in all habitats except dense forests. The family includes important food plants like Lactuca, lettuce, to ornamentals. They produce the typical daisy-sunflower that makes them easily recognizable. The members of this family have strong chemicals used against herbivores. This is a young family probably no more than 30 million years old, which probably originated in the Oligocene Epoch of the Tertiary Period.