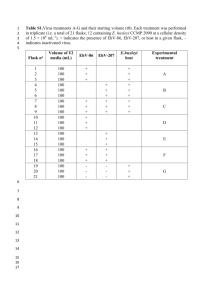
Table S1: List of primers used for qPCR and cloning of reporter constructs
Application
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
qPCR
Cloning
Cloning
Cloning
Mutagenesis
Mutagenesis
Name
Duox2-F3
Duox2-R3
L32-F
L32-R
Alpi_F
Alpi_R
Apoa4_F
Apoa4_R
Lyz1_F
Lyz1_R
Defa5_F
Defa5_R
Krt20_F
Krt20_R
Ki67_F
Ki67_R
Ephb2_F
Ephb2_R
Duox2_P_f1_SacI
Duox2_P_f3_SacI
Duox2_P_r_MluI
Duox2_1kbmut_F
Duox2_1kbmut_R
Sequence 5’3’
ACGCAGCTCTGTGTCAAAGGT
TGATGAACGAGACTCGACAGC
CCTCTGGTGAAGCCCAAGATC
TCTGGGTTTCCGCCAGTTT
CTGCCAAGAAGCTGCAGCCCA
GGCTAGGGGTGTCTCCGGTCC
GCCCAGTGAGGAGCCCAGGA
CCACATTGGCCACCTGGTCCG
ACTGAGGAGCAGCCAGGGGA
ACGCGTTCTCTTCTTTTGCAGCC
AGCTGGCTGACTGGGTGTGT
CGGTGCTTCGGTCTCCACGG
GGATTCGAGGTTCAAGTCACGG
TCTAGGTTGCGCTCCAGAGACT
CAATGTGCCTCGCAGTAAGA
GCATCTTTGGGGTTTTCTCA
CAACGGTGTGATCCTGGACTAC
CACCTGGAAGACATAGATGGCG
AAAAGAGCTCCAGGCAAGATGAACAGGAAGCTAG
AAAAGAGCTCGGTCATGGCACTGTGCAAGG
TTTTACGCGTGGGACTTGTTGGAAGCATGCTG
AGAACAAACCCCATAACCTGCAGGGCTGCGCTCCTGAC
GTCAGGAGCGCAGCCCTGCAGGTTATGGGGTTTGTTCT
Reference
Lipinski et al., 2009
Lipinski et al., 2009
Sayin et al., 2013
Sayin et al., 2013
References:
Lipinski, S., A. Till, C. Sina, A. Arlt, H. Grasberger, S. Schreiber and P. Rosenstiel (2009). "DUOX2-derived reactive oxygen species are
effectors of NOD2-mediated antibacterial responses." J Cell Sci 122(Pt 19): 3522-3530.
Sayin, S. I., A. Wahlstrom, J. Felin, S. Jantti, H. U. Marschall, K. Bamberg, B. Angelin, T. Hyotylainen, M. Oresic and F. Backhed (2013).
"Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist."
Cell Metab 17(2): 225-235.
Figure and Table legends
Figure S1: Characterization of LCM-harvested epithelial fractions.
Expression of the tip specific genes Alpi, Apoa4 and Krt20 and the crypt specific genes Lyz1,
Defa5, Ki67 and Ephb2 in LCM-harvested tip and crypt epithelial fractions of ileum and
colon and whole tissue. Data show a representative experiment. All values were normalized to
expression in whole tissue.
Figure S2: Microbial induction of Duox2 expression in duodenum and jejunum.
Duox2 expression in (A) duodenum and (B) jejunum of GF and CONV-R C57BL6 mice.
n=5-10 per group. Data show mean ± SEM; * p<0.05 and *** p<0.001 vs GF. Notably, the
absolute Duox2 expression in duodenum and jejunum were 10-100-fold lower as compared to
ileum and colon.
Figure S3: Ileal and colonic Tnfa expression during colonization of GF mice with a
normal microbiota.
n=4-5 per group. Data show mean ± SEM; *** p<0.001 and **** p<0.0001 vs GF.
Figure S4: Microbial induction of Duox2 expression is independent of MAMPs.
Duox2 expression in ileum and colon of (A) GF and CONV-R C57BL6 mice and mice
colonized with Escherichia coli, Bifidobacterium longum or Bacteroides thetaiotaomicron for
two weeks or (B) GF and CONV-R Swiss Webster mice and mice stably colonized with a
community of ten commensal bacterial strains (COM) for 4 months. n=5-10 per group. Data
show mean ± SEM; ** p<0.01 and *** p<0.001 vs GF.
Figure S5: NFκB binding sites are dispensable for IL-1β-mediated induction of Duox2
expression in Caco-2 cells.
A) Schematic overview of Duox2 promoter, predicted NFκB binding sites and luciferase
reporter vector. B) Luminescence of lysates from Caco-2 cells transfected with luciferase
reporter constructs treated with medium or 20 ng/ml IL-1β for 6 h. Constructs contained
either 1 kb or 3 kb of Duox2 wild type promoter or 1 kb promoter with mutagenized NFκB
binding site (1kb mut). n=3 per group. Data show mean ± SEM; *** p<0.001 vs medium.
Table S1: List of primers used for qPCR and cloning of reporter constructs.
 0
0