Project Description - Optical Oceanography Laboratory
advertisement

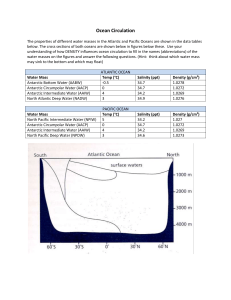
Ocean Circulation and Ecosystem Dynamics in the Vicinity of the Antarctic Peninsula: An Assessment Coupling Multiple Remote Sensing, In Situ Observation, and Modeling PROJECT DESCRIPTION INTRODUCTION The Antarctic Peninsula region is influenced by large-scale atmospheric forcing, such as the El Nino-Southern Oscillation, the Antarctic Circumpolar Wave, and the circumpolar trough of low pressure, as well as by the Antarctic Circumpolar Current (ACC) and interannual variability in sea ice extent. Despite relatively modest primary productivity throughout much of the Southern Ocean (Jacques 1989), waters to the west of the Antarctic Peninsula historically have supported an unusually high production of Antarctic krill and, thus, have been a favorable habitat for krill predators (e.g., fish, penguins, seals, and whales) (Marr 1962). Krill have a large interannual variability in recruitment, which has been correlated with the timing and extent of sea ice formation (Kawaguchi and Satake 1994, Siegel and Loeb 1995). A strong recruitment year, however, may be intervened by 4 - 6 years of moderate or poor year classes. Dramatic environmental changes are currently taking place along the Antarctic Peninsula (Vaughan et al. 2003, Smith et al. 1999). The most significant is warming air temperatures of about 0.1ºC per year during winter over recent decades (King et al. 2003). Given that krill recruitment is highly variable and possibly linked to sea ice extent, changing ice conditions may reduce krill populations and, through direct and indirect effects, upper trophic level populations as well (Fraser and Hofmann 2003). Owing to the remote location of the Southern Ocean and the difficulty of sampling in sea ice, however, our knowledge of ocean and ecosystem dynamics remains limited. As part of the Southern Ocean GLOBEC program, we collected field data during autumn and winter that indicated a large recruitment of krill occurred during 2001 in waters over the shelf west of the Antarctic Peninsula (Fig. 1), whereas there was poor recruitment in 2000 and low recruitment in 2002 (Fig. 2). The combination of environmental conditions that supported the high krill recruitment in 2001 cannot be deduced from sparse shipboard sampling. Satellite technology vastly increases the spatio-temporal sampling for these remote regions. Imagery from the Coastal Zone Color Scanner (CZCS) and SeaWiFS have been successfully used to investigate the distribution of phytoplankton blooms in different regions of the Southern Ocean (e.g., Comiso et al. 1990, Sullivan et al. 1993, Arrigo and McClain 1994, Moore et al. 1999). These data suggest that estimates of primary productivity need to be revised upwards. In addition, various geophysical ice parameters, sea surface temperature, cloud cover, and wind velocity are available for detailed investigations of ocean processes in polar regions at relatively high temporal and spatial resolutions (Comiso 1991). Here, we propose to use data from a suite of satellite sensors, in situ ocean observations, and an ocean circulation model to investigate different scales of forcing on ocean circulation and the relationship between different forcing mechanisms and krill recruitment for a period of 1997 to present in order to improve our ability to predict marine ecosystem response to climate variability in this dynamic region. BACKGROUND The annual advance and retreat of sea ice (>16 million km2) in the Southern Ocean is one of the most profound changes affecting ecosystems on this planet. Sea ice plays a central role in the temporal structuring of the ecosystem (Daly and Macaulay 1991) and by sustaining different communities within sea ice compared with that in the underlying water (Eicken 1992). In addition, sea ice extent and ocean circulation affects ecosystem structure and biological activity by forming boundaries defining different marine regimes (Nicol et al. 2000). Interannual variability in sea ice is influenced by a number of large-scale processes, such as the Southern Oscillation (Kwok and Comiso 2002) and the Antarctic Circumpolar Wave (ACW; White & Peterson 1996), and interactions between them. The ACW may take the form of a wavenumber-2 perturbation of sea surface temperature (SST), surface air pressure, sea-surface height, windstress, and sea ice extent, circling eastward around Antarctica with a period of around 4 years. In general, sea ice forms during May and June along the Antarctic Peninsula and starts to retreat near the northern end in August. Most of the Bellingshausen Sea to the west of the Peninsula, however, experienced a shortening of the sea ice season between 1979-1999 (Parkinson 2002), which was related to the Southern Oscillation index (Kwok and Comiso 2002). Little sea ice was present during 1999 and 2000 along the Peninsula, whereas during 2001 and 2002, sea ice conditions were extensive in our study area (Fig. 3). Anomalously high atmospheric pressure over the south Atlantic and anomalous lows over the Bellingshausen, also resulted in exceptionally heavy ice conditions during summer 2001/2002 in the Weddell Sea, east of the Antarctic Peninsula (Turner et al. 2002). The Antarctic Peninsula is the only part Fig. 3. Color-coded sea ice concentration maps of the Antarctic where the circumpolar during winter maximum of sea ice extent. Arrow trough crosses the continent. This lowshows location of GLOBEC study area. pressure trough rings the southern [removed because Comiso is co-I here. Fig. hemisphere between 60 and 70º S and is a Needs to be zoomed to show legend] region of higher cyclogenesis, particularly in the Bellingshausen Sea (Turner et al. 1998). The relative position of the circumpolar trough across the Antarctic Peninsula influences the variability in the seasonal cycle of atmospheric temperature, pressure, wind, precipitation, cyclonic activity and the sea ice distribution in this region (Smith et al. 1999). Warm winters along the Antarctic Peninsula have been associated with negative sea surface pressure anomalies in the Bellingshausen Sea and with positive anomalies for cold winters (Marshall and King 1998). The Southern Ocean also is characterized by low seawater temperatures, high nutrient concentrations, and extreme seasonal changes in incoming solar radiation, which results in highly seasonal primary production. Although much of the Southern Ocean is a high nutrient-low biomass environment, elevated concentrations of phytoplankton and zooplankton occur, particularly in coastal regions (Smith et al. 1996). The Antarctic krill, Euphausia superba, occupies a key role in the Southern Ocean ecosystem as the major pelagic herbivore and prey for most upper trophic level predators. It is not known why the waters west of the Antarctic Peninsula support one of the largest concentrations of krill in the Southern Ocean. A large krill recruitment, however, depends on several factors: (1) an early and sustained reproduction (i.e., November – March), (2) survival and high growth rates of larvae during summer [austral summer?], and (3) survival of larvae overwinter. The correlation between sea ice and recruitment may be due to the fact that ice edge blooms of phytoplankton support early adult reproduction and provide a food source for early feeding larvae (Daly and Macaulay 1991), while during winter sea ice may provide a food source (i.e., sea ice biota) and a haven from predation for overwintering larval krill (Daly 1990). Along the Antarctic Peninsula, adult krill migrate offshelf into the eastward flowing ACC during spring to spawn (Siegel 1989), their eggs sink into the underlying, warmer Upper Circumpolar Deepwater (UCDW) where they hatch, and then the larvae swim back up to the surface (Hofmann et al. 1992). Most larvae in offshore waters are probably advected eastward in the ACC towards South Georgia and are likely the source population for that area (Hofmann et al. 1998). For krill to have a large recruitment on-shelf and maintain a population along the Peninsula, offshore larvae must advected onto the shelf and then retained on the shelf until the following spring. Capella et al. (1992) used a model to demonstrate that surface flow was a primary factor governing the final location of larvae and, that due to seasonal changes in wind stress fields, favorable conditions for onshore transport of larvae were more likely early in the spawning period. Offshore larvae occur between 0 – 500 m depth, but the depth of maximum abundance offshelf and on-shelf is in the upper 30 m (Fig. 4, Daly in press). Recent results indicate that gyres, which could retain larvae and phytoplankton, occur in several locations on the shelf west of the Peninsula (Klinck ref?). The small internal Rossby radius of deformation at these high latitudes (ca. 5 –15 km?) results in narrow currents and relatively small mesoscale eddies and gyres. Thus, our central hypothesis is: [I move the "SeaWiFS" paragraph in the work plan/method part, as it reads abrupt here] The central western Antarctic Peninsula experiences a unique combination of environmental conditions that contributes to enhanced krill reproduction, growth, winter survivorship and recruitment. These conditions include early-forming and extensive sea ice coverage during fall and winter, predictable ice edge blooms both offshelf and on-shelf during spring, intrusions of Antarctic Circumpolar Current-derived Upper Circumpolar Deep Water onto the shelf which supplies nutrients that sustain phytoplankton blooms on the shelf, and a sluggish cyclonic shelf circulation that retains krill larvae. Atmospheric forcing, sea ice distribution, and oceanographic processes dictate the timing and location of phytoplankton blooms, the timing and magnitude of krill reproduction, the advective transport of larvae, the overwintering survival of larvae, and subsequent recruitment to juveniles during spring. Specifically, our objectives are to determine the suite of environmental conditions present during 2001 that would favorably support krill recruitment in comparison with conditions during other years. The environmental conditions include timing and extent of sea ice, atmospheric pressure, wind speed and direction, sea surface temperature, ocean currents, and the timing, location, and magnitude of chlorophyll concentrations; determine interannual changes in regional and large-scale oceanographic conditions and investigate the associated large-scale forcings to develop a better understanding of how climate variability influences ecosystem dynamics in the Antarctic Peninsula. To meet these objectives we will: Collate and statistically analyze in situ and remotely sensed data, develop climatologies, and assess anomalies for 1999 – 2003; Validate ocean color data with in situ chlorophyll concentrations; refine algorithms as necessary using in situ chlorophyll, CDOM, and transmissometer measurements; Study the correlation between phytoplankton (concentration, distribution, and timing) and krill recruitment Fine-tune and validate an ocean circulation model with satellite derived winds and sea ice cover; Use the ocean circulation model to test hypotheses related to sea ice distribution and krill recruitment and the transport and retention of larval krill on the Antarctic Peninsula shelf. [We also will investigate the potential impact of a reduction in ice habitat on krill populations - John I am not sure what more can be done with the model beyond advection questions - have I overstepped here?] Estimate the percent of krill larvae advected downstream in the ACC vs. the proportion advection onto the shelf west of the Antarctic Peninsula under different environmental conditions. Investigate how large-scale atmospheric and oceanographic forcings influence interannual variability in krill recruitment using statistical analyses (what statistical tests?) of remotely sensed data. Data sets Remote sensing The remote sensors primarily used in this study will include: SeaWiFS (1997 - present); ocean color (spectral water-leaving radiance, chlorophyll concentration, diffuse attenuation) MODIS (Terra: 1999 – present; Aqua: 2002 - present); ocean color and sea surface temperature (for quality control MODIS/Aqua data may be used only) AVHRR pathfinder (1985 - present); sea surface temperature QuikScat (1999 - present); surface wind TOPEX/POSEIDON (T/P, 1996 - present): surface height anomaly from 70oS to 70oN EPTOMS (1996 - present); total ozone thickness NCEP; blended meteorological data (surface pressure, water vapor, humidity, wind) with remote and in situ sensors and models. Sea ice sensors?????? Historical data from CZCS (1978-1986) and AVHRR (1985 - present) will be used for decadal comparisons. We recognize that CZCS ocean color data (pigment concentration) may not be comparable to those obtained from SeaWiFS and MODIS due to differences in sensor calibration and data processing algorithms, therefore only distribution patterns will be studied. In situ measurement Historical and recent in situ data will be obtained from a variety of sources. We have available to us all of the underway data collected by NSF/Polar Programs vessels, the RV L.M. Gould and the RVIB N.B. Palmer from their shipboard surface flow-through systems as well as biogeochemical data and hydrographic data collected at cruise stations as part of the several field program. These data from the field programs are listed in Table 1. These will be quality controlled [I think we need to focus on a subset and explain why we need data type X] and used to fine-tune and validate remote sensing algorithms and circulation models (see Methods). Table 1. Catalogue of available in situ data Measurements AMLR Vertical Profiles Temperature Oxygen Depth PAR Fluorescence Transmissometer 1995 - 2003 1995 - 2002 1995 - 2003 1995 - 2003 1995 - 2003 1995 - 2003 Underway data Sea surface temperature Sea surface salinity Fluorescence Air temperature Pressure Air humidity Wind speed and direction 1999 – 2002 1999 – 2002 1999 – 2002 1999 – 2002 1999 – 2002 1999 – 2002 1999 – 2002 Phytoplankton Chlorophyll a concentration PAR HPLC POC/PON Phycoerythrins (PE) Beam attenuation (transmissometer) Fluorescence Up/Downwelling irradiance (Ed + Eu) Ap + As (coeffcicients) Seawifs imagery Species composition Cell size spectrum Primary production 1995 – 2003 1999 – 2002 1999 – 2002 1995 – 2002 1999 – 2002 1999 – 2002 1999 – 2002 1999 – 2002 1999 – 2002 1999 – 2002 1995 - 1997 1995 - 1997 1995 - 1997 Photoadaptational state of cells 1996 - 1997 Krill abundance/stage/size 1995 – 2003 METHODS Ocean color data analyses (timing and magnitude of food availability) Phytoplankton provides food for all higher trophic levels. Therefore one of our major efforts will be on quantifying its concentration, distribution, and temporal changes by combining in situ and remote measurements. CZCS and SeaWiFS-derived chlorophyll data have been used to document distributions of phytoplankton in the Polar regions (e.g., Sullivan et al., 1993; Moore et al., 1999), yet these pioneering studies are limited in the time span, therefore do not provide a comprehensive image. Our own preliminary study with low-resolution (9-km) SeaWiF chlorophyll data from 1997 to 2003 indicates that spring blooms west of the Antarctic Peninsula start offshelf in the ACC then progress into coastal waters, where relatively intense blooms occur in different areas throughout the summer, particularly at the southern end of the Peninsula in the vicinity of the GLOBEC study site near Marguerite Bay (Fig. 5). During austral summer, the ACC has low chlorophyll concentrations, while coastal regions and frontal zones are food-rich environments. Chlorophyll maxima are usually near surface in waters over the continental shelf and there is a tight coupling between surface chlorophyll and chlorophyll integrated to depth (Smith et al. 1996). Chlorophyll concentrations near surface are usually > 1.0 mg m-3 between October and April (austral spring – fall) and, on average, about 5 mg m-3 during austral summer, with maxima up to 40 mg m-3. Prézelin et al. (2000) demonstrated that diatom blooms primarily occur in areas on the shelf that have topographically-driven upwelling of UCDW from the meandering southern boundary of the ACC. Seasonal and interannual changes in the location and strength of the ACC will influence the flushing of subpycnocline waters on the shelf. Wind-driven upwelling also may occur at some sites; however, summer winds (predominantly from the north-northeast) generally favor downwelling conditions. The areas where upwelling of UCDW occurs, nutrients are replenished near surface permitting sustained phytoplankton production. Elsewhere on the shelf, blooms may be dominated by prymnesiophytes, pelagophytes, or cryptophytes in regions influenced by glacial melt. Intrusions of UCDW were observed in the GLOBEC study site where a deep canyon intersected the shelf break (Klinck et al. in press?), and large blooms were dominated by the diatoms, Chaetoceros criophilum and Synedra antarctica, and phaeodarian radiolarians throughout Marguerite Bay (Daly in press). A preliminary comparison between SeaWiFS- and ship-derived chlorophyll values (Fig. X bottom panel) shows that SeaWiFS (Version 4 processing) tends to underestimate chlorophyll at low concentrations, but overestimate it in higher concentrations. This is consistent with findings from the Arctic region (Cota et al., 2004), but different from results obtained by Moore et al. (1999), where SeaWiFS data from earlier version of processing. Certainly, the comparison is not strict as each SeaWiFS pixel used here is about 99 km2 and the ship sampling may not be concurrent (up to one month difference). Yet, the availability of larger in situ dataset, as well as higher-resolution SeaWiFS data (daily and weekly), makes it possible for an extensive comparison, based on which a regional algorithm may be developed and validated with independent in situ measurement. Ocean color estimates from satellite are sub to uncertainties, especially in the high-latitude regions for a number a reasons, one of which is lack of reliable optical measurements in these remote regions. Although the relative distribution and the temporal changes of the distribution rather than the absolute chlorophyll concentration are more important in our study, it is critical to fine-tune the algorithms to produce at least consistent, if not accurate (this is our ultimate goal, though), time-series of chlorophyll distributions. Otherwise it is unknown if a distribution pattern and its temporal change are real, due to several factors. For example, another important water constituent is colored dissolved organic matter (CDOM, also called Gelbstoff), which is often interpreted as chlorophyll because they both strongly absorb blue light and the band-ratio OC4 bio-optical algorithm (O'Reilly et al., 2000) does not distinguish the two. Also, the spectral water-leaving radiance data (Lw()) from SeaWiFS or MODIS in this region have never been validated in this region due to lack of reliable in situ Lw() measurements. However, starting with the satellite-derived Lw() and assuming that they are consistently derived, a regional bio-optical inversion algorithm may be derived with the in situ chlorophyll and other measurement. Specifically, we plan to collect in situ data (historical and ongoing) that are related to ocean color (see Data Sets), for example chlorophyll, CDOM, transmission/attenuation, and light profiles obtain Lw() and other data products (chlorophyll, attenuation) from SeaWiFS and MODIS at higher spatial (4- and 1-km) and temporal (daily to weekly) resolutions from NASA DAAC and GSFC validate the satellite parameters by comparing the concurrent matching pairs from satellite and in situ measurements (e.g., Hu et al., 2003), and if necessary (very likely, as indicated by Fig. X) develop and fine-tune a regional algorithm by using the satellite Lw() and in situ parameters, similar to the approach used by Kahru and Mitchell (2001) This effort is of particular importance to NASA, as both SeaWiFS and MODIS show unacceptable bias in high latitude waters (e.g., Cota et al., 2004) even after several rounds of data reprocessing. Meanwhile the southern ocean plays a significant role in the global carbon cycle, but it is at present difficult to quantify the uncertainties associated with the satellite estimates in a systematic fashion, primarily due to lack effort in consolidating all related in situ data collected in recent years (see Data Sets) with satellite data. Our studies will address these issues and ultimately improve global carbon cycle models, which depend critically on reliable chlorophyll estimates. Sea ice analyses Ocean circulation model SIGNIFICANCE OF RESEARCH PROJECT COORDINATION AND TIMELINE Kendra Daly - primary contact, organization, in situ data, supervise grad student, co-supervise post-doc Chuanmin Hu - responsible for all remote sensing products except sea ice, co-supervise postdoc Joey Comiso - analysis of sea ice products, coordinate with Chunamin on other satellite data and with John on model John Klinck - model development, validation, hypothesis testing, supervise grad student, work with USF post-doc? REFERENCES Aoki S, Yoritaka M, Masuyama A. 2003. Multidecadal warming of subsurface temperature in the Indian Ocean sector of the Southern Ocean. Journal of Geophysical Research-Oceans 108 (C4):8081. Arrigo KR, McClain CR. 1994. Spring phytoplankton in the western Ross Sea. Science 266: 261-263. Capella JE, Quetin LB, Hofmann EE, Ross RM. 1992. Models of the early life history of Euphausia superba – Part II. Lagrangian calculations. Deep-Sea Research 39: 1201-1220. Comiso JC. 1991. Satellite remote sensing of the polar oceans. Journal of Marine Science 2: 395-434. Comiso JC, Maynard NG, Smith WO, Jr, Sullivan CW. 1990. Satellite ocean color studies of Antarctic ice edges in summer and autumn. Journal of Geophysical Research 95 (C6): 94819496. Cota GF, Wang J, Comiso JC. 2004. Transformation of global satellite chlorophyll retrievals with a regionally tuned algorithm. Remote Sensing of Environment 90: 373-377. Daly KL. 1990. Overwintering development, growth, and feeding of larval Euphausia superba in the Antarctic marginal ice zone. Limnology and Oceanography 35: 1564-1576. Daly KL. Overwintering growth and development of larval Euphausia superba: An interannual comparison under varying environmental conditions west of the Antarctic Peninsula. Deep-Sea Research, in press. Daly KL, Macaulay MC. 1991. Influence of physical and biological mesoscale dynamics on the seasonal distribution and behavior of Euphausia superba in the Antarctic marginal ice zone. Marine Ecology Progress Series 79: 37-66. Eicken H. 1992. The role of sea ice in structuring Antarctic ecosystems. Polar Biology 12: 313. Fraser WR, Hofmann EE. 2003. A predator’s perspective on causal links between climate change, physical forcing and ecosystem response. Marine Ecology Progress Series 265: 1-15. Hofmann EE, Klinck JM, Locarnini RA, Fach B, Murphy E. 1998. Krill transport in the Scotia Sea and environs. Antarctic Science 10: 406-415. Hofmann EE, Capella JE, Ross RM, Quetin LB. 1992. Models of the early life history of Euphausia superba – Part I. Time and temperature dependence during the descent- ascent cycle. Deep-Sea Research 39: 1177-1200. Hu C, Muller-Karger FE, Biggs DC, Carder KL, Nababan B, Nadeau D, and Vanderbloemen J. 2003, Comparison of ship and satellite bio-optical measurements on the continental margin of the NE Gulf of Mexico, Int. J. Remote Sens. 24:2597-2612. Jacobs SS, Giulivi CF, Mele PA. 2002. Freshening of the Ross Sea during the late 20th century. Science 297: 386-389. Jaques G. 1989. Primary production in the open Antarctic Ocean during the austral summer: a review. Vie Milieu 39: 1-17. Kahru M, Mitchell BG. 2001. Seasonal and non-seasonal variability of satellite-derived chlorophyll and CDOM concentration in the California Current. J. Geophys. Res., 106(C2): 2517-2529. Kawaguchi S, Satake M. 1994. Relationship between recruitment of the Antarctic krill and the degree of ice cover near the South Shetland Islands. Fisheries Science 60: 123-124. King JC, Turner J, Marshall GJ, Connolley WM, Lachlan-Cope TA. 2003. Antarctic Peninsula climate variability and its causes as revealed by analysis of instrumental records. Antarctic Research Series 79: 17-30. Kwok R, Comiso JC. 2002. Southern Ocean climate and sea ice anomalies associated with the Southern Oscillation. Journal of Climate 15: 487-501. Marr JWS. 1962. The natural history and geography of the Antarctic krill, Euphausia superba Dana). Discovery Reports 322: 33-464. Marshall GJ, King JC. 1998. Southern hemisphere circulation anomalies associated with extreme Antarctic Peninsula winter temperatures. Geophysical Research Letters 25: 24372440. Moore JK, Abott MR, Richman JG, Smith WO, Cowles TJ Coale KH, Gardner WD, Barber RT. 1999. SeaWiFS satellite ocean color data from the Southern Ocean. Geophysical Research Letters 26: 1465-1468. Nicol S, Pauly T, Bindoff NL, Wright S, Thiele D, Hosie GW, Strutton PG, Woehler E. 2000. Ocean circulation off east Antarctica affects ecosystem structure and sea-ice extent. Nature 406: 504-507. O'Reilly, J. E., and 21 others, Ocean color chlorophyll a algorithms for SeaWiFS, OC2, and OC4: version 4. In: SeaWiFS Postlaunch Technical Report Series (S. B. Hooker & E. R. Firestone Eds.), NASA Technical Memorandum 2000-206892, Vol. 11. Greenbelt, Maryland: NASA Goddard Space Flight Center (51pp), 2000. Parkinson CL. 2002. Trends in the length of the Southern Ocean sea-ice season, 1979-99. Annals of Glaciology 34: 435-440. Siegel V. 1989. Winter and spring distributions and status of the krill stock in the Antarctic Peninsula area. Arch. FishWiss. 39: 45-72. Smith RC, Dierssen HM, Vernet M. 1996. Phytoplankton biomass and productivity in the western Antarctic Peninsula region. Antarctic Research Series 70: 333-356. Smith RC, Ainley D, Baker K, Domack E, Emslie S, Fraser B, Kennett J, Leventer A, Mosley-Thompson E, Stammerjohn S, Vernet M. 1999. Marine ecosystem sensitivity to climate change. BioScience 49: 393-404. Siegel V, Loeb V. 1995. Recruitment of Antarctic krill Euphausia superba and possible causes for its variability. Marine Ecology Progress Series 123: 45-56. Sullivan CW, Arrigo KR, McClain CR, Comiso JC, Firestone J. 1993. Distributions of phytoplankton blooms in the Southern Ocean. Science 262: 1832-1837. Turner J, Marshall GJ, Lachlan-Cope TA. 1998. Analysis of synoptic-scale low pressure systems within the Antarctic Peninsula sector of the circumpolar trough. International Journal of Climatology 18: 253-280. Turner J, Harangozo SA, Marshall GJ, King JC, Colwell SR. 2002. Anomalous atmospheric circulation over the Weddell Sea, Antarctica during the austral summer of 2001/2002 resulting in extreme sea ice conditions. Geophysical Research Letters 29: 2160-2163. Vaughan DG, Marshall GJ, Connolley WM, Parkinson C, Mulvaney R, Hodgson DA, King JC, Pudsey CJ, Turner J. 2003. Recent rapid regional climate warming on the Antarctic Peninsula. Climatic Change 60: 243-274.