Life-moves-onto-dry
advertisement

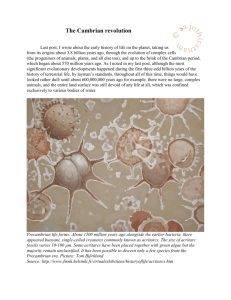
Life moves onto dry land Last post, I wrote about the so called Cambrian “revolution” (which began around 570 million years ago, and ended about 490 million years ago), a period of time during which organisms on the Earth underwent massive speciation, diversifying well beyond the bounds of the relatively very simple pre-Cambrian forms (which were mostly unicellular or fairly basic multi-cellular forms collectively called the Ediacaran flora and fauna). Reconstruction of the sea floor during the Vendian times when the Ediacaran organisms thrived. Image from http://www.carleton.ca/~tpatters/teaching/intro/cambrian/cambrianex8.html. Source: http://www.emc.maricopa.edu/faculty/farabee/BIOBK/BioBookPaleo2.html Compare the life forms visible in the above graphic, which is an artist’s impression of the Ediacaran forms, with the two graphics below, artists’ impressions of flora and fauna after the Cambrian explosion, and you will see two key things: 1. There’s a far greater variety of forms, though whether this is because there really were more forms after the Cambrian explosion, or whether pre-Cambrian life-forms just did not fossilise as well as Cambrian forms, is open to discussion; 2. The Cambrian forms seem to be more physiologically complex. At the beginning of the Paleozoic era, multi-cellular animals underwent an ‘explosion’ in diversity known as the ‘Cambrian explosion’. This artist’s rendering, based on fossil evidence, of some of the bizarre life forms, now extinct, that emerged during this time. Source: http://www.astrobio.net/pressrelease/2431/attack-of-the-giant-insects In the image above, trilobites (1) live among many species that are not normally preserved. A typical Cambrian outcrop might produce only trilobites, brachiopods (2), mollusks (3), and crinoids (4). That is a tiny fraction of the full Cambrian biota, better represented by the roster of the Burgess Shale Cambrian Konservat-Lagerstatten. That community includes sponges Vauxia (5), Hazelia (6), and Eifellia (7); brachipods Nisusia (2); priapulid worms Ottoia (8); trilobites Olenoides (1); other arthropods such as Sidneyia (9), Leanchoilia (10), Marella (11), Canadaspis (12), Helmetia (13), Burgessia (14), Tegopelte (15), Naraoia (16), Waptia (17), Sanctacaris (18), and Odaraia (19); lobopods Hallucigenia (20) and Aysheaia (21); mollusks Scenella (3); echinoderms Echmatocrinus (4); and chordates Pikaia (22); among other oddities, including Haplophrentis (23), Opabinia (24), Dinomischus (25), Wiwaxia (26), Amiskwia (27), and Anomalocaris (28). ©2002 by S.M. Gon III (composition & linework) & John Whorrall (color rendering) Source: http://users.path.ox.ac.uk/~wjames/Evolution/evolution4_files/frame.htm Looking at this vast and wildly varying array of creatures, many of which would look utterly outlandish to us today, it is easy to believe that at this stage of evolution, the cards of animal design were constantly being shuffled and dealt over and over again, with few of the hands having whatever it took to stay in the game, rather than fold and become extinct: In many ways in fact, the Cambrian was the time when the most radical designs were tried out: five eyes; multi-jointed legs; gills along the perimeter of carapaces of assorted designs – you name it, evolution most likely tried it during the Cambrian. Consequently, it is hardly surprising to know that it was during this period that, after a series of abortive experiments in various evolutionary forms (punctuated by five major bouts of mass extinction), all the major lines of plant and animal types, whose descendants fill the world today, were established – leaving by the evolutionary wayside an imponderable assortment of other forms that became extinct along the way. It is the story of these “survivors” that I now wish to take up. Extinctions, note the Cambrian ones. Source: http://wpcontent.answers.com/wikipedia/commons/thumb/0/06/Extinction_intensity.svg/320pxExtinction_intensity.svg.png After the last of the five extinctions mentioned above, which together culled the Cambrian flora and fauna and pointed them in our present-day direction was over (about 500 million years ago), the surviving groups exploded in another evolutionary scramble that within 50 million years tripled the diversity of marine animal life. This marked the start of the Ordovician, and the animals that now emerged constituted the earliest members of a classification known as the Paleozoic fauna. Ordovician fauna & flora Source: http://www.columbia.edu/itc/eeeb/baker/N0316/Lecture%205/Images/ordovician.JPG These Ordovician animals - all of which were marine creatures - were of all sorts: some, like the Eumorphocysts, would have been strange and utterly unfamiliar to us, but there were also many that would have looked right at home in a dish of seafood stew! Of these latter, amongst the most common forms - at least as far as the fossil record goes - were the articulated brachiopods (which, superficially at least, looked much like our contemporary clams and oysters), and the nautiloid cephalopods (many of which are hardly distinguishable from latter-day squids). Pikaia (left) and Astraspis (right) Sources: http://www.karencarr.com/auto_image/auto_image_mid/Karen_Carr_Field_Museum_Pikaia_det ail.jpg http://www.ppne.co.uk/uploads/18449_icon.jpg Eventually, about 470 million years ago, Astraspis - a creature that may possibly be the great grand parent of all fishes evolved, deriving in its turn, as far as we can tell, from a Cambrian worm-like swimmer Pikaia, which is thus a likely candidate for the progenitor of all chordates (the phylum that encompasses all animals with a spinal cord), and hence all vertebrates, which are a sub-class of the former. In other words, great-great-great….grandpa/ma could well have looked like these guys. Sometimes, it can be quite instructive to meet the family! Astraspis was a peculiar looking customer by our standards: Of its 10 cm length, about two thirds, corresponding to its head and body section, was encased in thick bony shields which were pierced by gill and eye openings. It had no fins apart from its tail, and although it could swim in a fashion, by beating its somewhat less armoured tail from side to side, its jawless mouth – which could suck in water and filter it, though could not chew – indicates that it was probably a benthic feeder, which trawled slowly along the sea beds rather than swimming at mid-water levels like most fish do today. Although Astraspis was not a particularly efficient swimmer, it got on very well in the Ordovician seas in which it evolved because, initially at least, it had no competition. However, that was a state of affairs which wasn’t going to last for too long. The basic Astraspis design – that of a potentially mobile swimmer which propelled itself with a side to side motion of its body – held myriad possibilities; possibilities which the forces of evolution were quick to exploit. After fairly diverse variations had been tried, the hard protective bony plates were finally shed, and the job of body support, as well as the function of providing a rigid framework to which muscles could be attached, and against which they could work, was taken over by an endoskeleton (a skeleton that is encased within the body, rather than one that encases the body within itself (an exoskeleton – like the ones prawns and crabs have)). This was a crucial inovation. It was what made the Ordovician (495 - 438 million years ago), the Silurian (438 - 408 million years ago), and to a much lesser extent, the Devonian (408 - 360 million years ago), the ages of the fishes. Turning for the moment to plant evolution, there too we find some remarkable innovations under way: Living in water has many advantages: There is no risk of dehydration; a lot of nutrients are carried in the water; and an aquatic organism doesn’t have to support too much of its own body-weight by itself – the buoyancy conferred by water relieves much of the load bearing requirements that life on land would impose. Be that as it may however, before the end of the Ordovician, marine plants were invading freshwater rivers, streams, ponds and lakes. Adapting to freshwater imposed new demands on plants: Although freshwater contains plenty of the essential nutrients that any plant needs – such as carbon dioxide, phosphates, and nitrates – because of osmosis, it is also more difficult to regulate cellular water levels: cells are salty, and thus, in freshwater, there is an osmotic pressure that has to be balanced if the cell is to remain stable. Once this problem was solved however, it was a simple step to start exploiting other moist environments – seasonal ponds, tidal marshes, spring seeps etc. From freshwater adaptation to dry land adaptation was only a very short hop: cell-walls needed to become a bit tougher so as to prevent drying out; they had to become strong enough to fully support the weight of the plant, and corrugated tubular arrays of longish cells appeared to do this; systems of channels and roots were developed so as to extract and carry nutrients and water from the soil into the plant; and a semi-permeable sheathing evolved, to keep moisture in but still allow the plant to breathe. When all of these innovations were in place, sometime towards the end of the Silurian, about 410 million years ago, plants, probably starting with Cooksonia, began to invade the dry land, and to move gradually further and further away from any bodies of water. Cooksonia fossil and an artist’s reconstruction thereof. Sources: http://eurypterids.net/CooksoniaPG.jpg http://upload.wikimedia.org/wikipedia/commons/e/ee/Cooksonia.png Hot on their tracks there followed the arthropods (millipede or scorpion like creatures). These too had to adapt to conditions on dry land, but here their pre-existing exoskeletons came in very handy. With a little modification, these hard shells were able to both prevent dehydration in the absence of a watery environment, and provide the physical support which the loss of aquatic buoyancy that accompanied life out of the water necessitated. Eurypterus remipes, ancestor of scorpions, and possibly the progenitor of all early terrestrial forms, though not quadrupeds, and thus not a potential member of your family! Source: Personal collection. Isobuthus rakovnicensis. Though depicted here in a terrestrial environment, isobuthus was most likely amphibious. Size ca. 10 cm Source: http://www.palaeos.com/Invertebrates/Arthropods/Scorpionida/Isobuthus.jpg Reconstruction of a fossil claw from the ancient sea scorpion Jaekelopterus rhenaniae (e), and comparative sizes of a human male with the sea scorpion (a), the trilobite Isotelus rex (b), the dragonfly Meganeura monyi (c), and the millipede Arthropleura armata (d). Source: http://opa.yale.edu/news/article.aspx?id=2010 Some of these early creatures were huge by today’s standards - Gigantoscorpio for example was a metre-long scorpion, while Arthropleura was a two metre-long arthropod. Of course there were also mite-sized animals, and all sorts of intermediate sizes. One thing though, none of these various land-creatures was a vertebrate (possessed of a back-bone), and none of them were quadrupeds (four-footed). However these were not lagging all that far behind. Next post, I shall look at the invasion of the land by vertebrates – our slightly less distant ancestors.