The usage of ethanol
advertisement

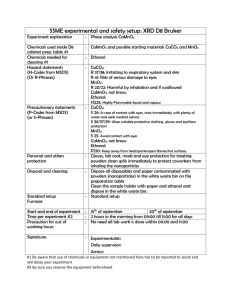
System Biology Project Biosynthesis of Ethanol Using E.coli System Group 11 生資所碩士班 924287 呂博凱 資工系博士班 928309 陳炯勳 資工系碩士班 924360 唐宗麟 Jan 20, 2005 The usage of ethanol Ethanol is a clear, colorless liquid with a characteristic, agreeable odor. In dilute aqueous solution, it has a somewhat sweet flavor, but in more concentrated solutions it has a burning taste. Ethanol, CH3CH2OH, is an alcohol, a group of chemical compounds whose molecules contain a hydroxyl group, –OH, bonded to a carbon atom. Ethanol melts at –114.1°C, boils at 78.5°C, and has a density of 0.789 g.mL-1 at 20°C. Its low freezing point has made it useful as the fluid in thermometers for temperatures below –40°C, the freezing point of mercury, and for other low-temperature purposes, such as for antifreeze in automobile radiators. Ethanol is used as an automotive fuel by itself and can be mixed with gasoline to form what has been called “gasohol”; the most common blends contain 10% ethanol and 85% ethanol mixed with gasoline. Over 1 billion gallons of ethanol are blended with gasoline every year in the United States. Because the ethanol molecule contains oxygen, it allows the engine to more completely combust the fuel, resulting in fewer emissions. Since ethanol is produced from plants that harness the power of the sun, ethanol is also considered a renewable fuel. Therefore, ethanol has many advantages as an automotive fuel. The metabolic pathways, the enzymes and the genes involved in the synthesis of ethanol During glycolysis, cells convert simple sugars, such as glucose, into pyruvic acid, with a net production of ATP and NADH. In the absence of a functioning electron transport system for oxidative phosphorylation, at least 95% of the pyruvic acid is consumed in short pathways which regenerate NAD+, an obligate requirement for continued glycolysis and ATP production. The waste products of these NAD+ regeneration systems are commonly referred to as fermentation products. Under conditions of oxygen insufficiency, microorganisms are particularly diverse in the array of fermentation products, which are produced by different genera. These products include organic acids, such as lactate, acetate, succinate, and butyrate, as well as neutral products, such as ethanol, butanol, acetone, and butanediol. Figure 1 represents the metabolic pathways involved in the biosynthesis of ethanol, generated by the KEGG website. The green background shows the enzymes that can be found in E.coli K-12 MG1655 strain. The red line shows the possible metabolic pathways involved in the biosynthesis of ethanol. The blue numbering shows the enzymes that may involve the biosynthesis of ethanol. There are nine enzymes in the glycolysis pathway: sugar phosphotransferas, glucose-6-phosphate isomerase, phosphofructokinase-2, fructose-bisphosphate aldolase, glyceraldehydes -3-phosphate dehydrogenase, acylphosphate phosphohydrolase, phosphoglycerate mutase, enolase, pyruvate kinase, respectively. They are encoded by ptsG, pgi, pfkB, dhnA, gapC_2, pgk, gpmA and pykF genes, respectively. There are two possible pathways in E.coli system that produce ethanol using pyruvate as the reactant. Figure 2 represents the two possible pathways. In the pathway 1, E coli is produced ethanol using pyruvate formate lyase (PFL) that converts pyruvate to acetyl-coA. But it is necessary to find some means that increase the flux of reaction 12, converting acetyl-coA to acetate. Acetate can convert to ethanol using aldehyde dehydrogenase and alcohol dehydrogenase class III. In the pathway 2, E coli is produced ethanol using pyruvate decarboxylase that converts pyruvate to acetaldehyde directly. Is E. Coli suitable for biosynthesis of ethanol?(LO, T et al. 1987; BS, MA et al. 2003) The shortcoming of using the pathway 1 to produce ethanol This fermentation pathway is unbalanced because one NADH, H+ is generated for each pyruvate made from sugars, and two NADH, H+ are required for converting pyruvate into ethanol. E. coli needs to balance it fermentation by also producing acetic and succinic acids. This is the major disadvantage associated with using E. coli cultures, a narrow and neutral pH growth range (ca. pH 6.0–8.0). According to the reason, we think the pathway 1 in the E. coli system isn’t suitable for biosynthesis of ethanol. We chose the pathway 2 to produce the ethanol in E. coli system. The pathway is expression of pyruvate decarboxylase that exists in Yeasts and Z. mobilis, in E. coli system. Pyruvate decarboxylase converts pyruvate to acetaldehyde, the reactant of producing ethanol. It consumes only one NADH, H+ for each ethanol produced. The shortcomings of using pathway 2 to produce ethanol and the methods to solve Early attempts to introduce a yeast-like pathway for converting pyruvate to ethanol were unsuccessful because the native alcohol dehydrogenase (ADH) activity of E. coli was not sufficient to achieve high ethanol yields. Ingram et al. isolated adh II from a genomic library of Z. mobilis using a novel indicator medium for ADH activity. E. coli transformed with a plasmid expressing pdc and adh II produced ethanol almost exclusively. The adhII and pdc genes were co-expressed under the control of the native lac promoter, and the construct was named the PET (production of ethanol). While plasmids carrying the adhII and pdc genes were not readily lost in the absence of antibiotics, these strains were still too genetically unstable for use in industrial fermentations. Ingrams research group sought to stabilize ethanol production by integrating the genes into the chromosome of strain ATCC11303. The PET operon was inserted into the pyruvate formate lyase (pfl) gene, in an attempt to eliminate an enzyme competing for pyruvate. However, the resulting construct yielded much lower amounts of ethanol than the plasmid bearing strains. This was attributed to reduced gene dosage. A strategy was developed for selecting mutants with increased adhII and pdc expression. The integrated DNA contained a chloramphenicol resistance marker. As a result, the final strain (KO11) was able to convert glucose and xylose to ethanol at yields 103 to 106% of theoretical. The extra ethanol was thought to arise from fermentation of carbohydrates present in the rich medium that were not accounted for in the sugar balance. The strain has not been widely used for SSF (simultaneous saccharification and fermentation), probably because the optimal culture pH (6.5) is not compatible with T. reesei cellulase complex, which has a pH optimum of 4.6. Propose experiments to determine the metabolic fluxes of the network associated with ethanol(F, CM et al. 1999; Torres and Voit. 2002) The implementation of the optimization method is diagramatically represented in Fig. 3. This may be divided into three main steps. Firstly, we need to design the mathematical model. Secondarily, we need to optimize the solution. Finally, we can assess the solutions. Design of the mathematical model Firstly, we need to define the relevant variables. Anaerobic fermentation pathway of the E.coli containing the PDC and ADHII enzymes is represented in Fig. 4. The metabolites and enzymes involved in the pathway is showed in the figure legends. In the GMA representation each rate law is written as a product of power law functions. For a system consisting of n dependent and m independent variables, these rate laws are written: The power law representation can be derived from first principles for arbitrary laws and is obtained by specifying a rate constant and writing a power term for each variable that directly influences the rate law in question. The exponent parameters in each power term are referred to as kinetic orders and defined as: Where the subscript 0 indicates evaluation at a given steady state. The rate constants are defined as: Accordingly the standard form of the GMA system of equations is: The model is described by the following set of differential equations: There are two means to estimate the kinetic orders and rate constants. Direct estimation from flux data 1. If the effect of Xj on flux Vi+ is to be determined, it would be ideal to keep all variable but Xj at their typical values, and to vary Xj about its normial value.The Flux Vi+ becomes a unvariate function of Xj, and this function is linear when plotted as the logarithm of Vi+ against the logarithm of Xj. 2.The Slope of the function is equivalent to the kinetic order gij(hij) and can estimate the rate constants. Estimation from the literature presentations Take the yeast system for example; the fluxes for the GMA take the following power-law form: Due to the inherent non-linearity of MM and GMA equations an assessment of the model quality and robustness is not as straightforward as in the linear case. Once the model has been established, those constraints necessary to guarantee cell viability must be also defined. These are of two main types: the steady-state constraints and the enzyme and metabolite concentration constraints. The former ensures that the optimized solution is at steady state while the latter constrains variables to remain within prescribed bounds. Additionally an appropriate objective function to be optimized (e.g. yield or flux) must be selected and formulated in mathematical terms. The optimization: a multi-start search algorithm We use a multi-start search algorithm, an improved hillclimbing. As all hillclimbing algorithms, the present one uses an iterative technique of improvement, meaning that during a single iteration a new activity is selected from the neighborhood of the current steady state. If the new activity set provides an improved value of the objective function and the metabolite concentrations agree with the imposed constraints, the new activity set becomes the current solution. Otherwise, some other neighbor is selected randomly and tested against the current solution. The search terminates if no further improvement is achieved. In the steady-state the model reduces to a set of non-linear equations, where the independent variables (Yj) are the enzyme activities and the dependent variables (Xi) are the metabolite concentrations. The optimization routine was represented in Fig. 5. Quality assessment of the solutions Since the optimization itself does not address the question of stability, the steady-state of the optimized solution must be checked with respect to its stability. With this purpose the differential equations are integrated numerically after a perturbation has been applied. The system is said to be stable (or metastable) if after a perturbation recovers its initial (steady) state. Unstable steady-state solutions are discarded. If this is the case the system is to be revisited after imposing more stringent constraints. Propose the ways to maximize yield The maximization of the ethanol production rate requires modulation of up to six enzymes. There are sugar transport system, VHK (hexokinase), VPFK (phosphofructokinase), VGADP (glyceraldehyde 3-phosphate dehydrogenase), VPK (pyruvate kinase), VATPase (ATPase), respectively. The increase in the rate of ethanol production, VPK/(VPK) basal, is more than 50 times, and the yield of the optimized solution is around 100% of theoretical. Figure 1. The metabolic pathways involved in the biosynthesis of ethanol. Green background: Enzymes can be found in E.coli K-12 MG1655 strain. Red line: The possible metabolic pathways involved in the biosynthesis of ethanol. Figure 2.The two possible pathways involved in the biosynthesis of ethanol. Figure 3. Flow diagram of the non-linear optimization Figure 4. Anaerobic fermentation pathway of the E.coli containing the PDC and ADHII enzymes. Metabolites: X1 (Glcin, intracellular glucose), X2 (G6P,glucose-6-phosphate), X3 (FDP, fructose diphosphate), X4 (PEP, phosphoenol pyruvate), X5 (ATP). Enzymes: Y1 (Vin, sugar transport system) Y2 (VHK, hexokinase), Y3 (VPFK,phosphofructokinase), Y4 (VGADP, glyceraldehyde 3-phosphate dehydrogenase), Y5 (VPK, pyruvate kinase), Y6 (Vcarb, glycogen synthetase), Y7 (VATPase, ATPase) Figure5. The optimization routine. Reference BS, D., C. MA, et al. (2003). "Bacteria engineered for fuel ethanol production: current status." Appl Microbiol Biotechnol. 63: 258-266. F, R.-A., R. CM, et al. (1999). "Non-linear optimization of biotechnological processes by stochastic algorithms: application to the maximization of the production rate of ethanol, glycerol and carbohydrates by Saccharomyces cerevisiae." Journal of Biotechnology 68: 15-28. LO, I., C. T, et al. (1987). "Genetic Engineering of Ethanol Production in Escherichia coli." APPLIED AND ENVIRONMENTAL MICROBIOLOGY 53(10): 2420-2425. Torres, N. V. and E. O. Voit. (2002). Pathway analysis and optimization in metabolic engineering.