Progress Report: September 2012
advertisement

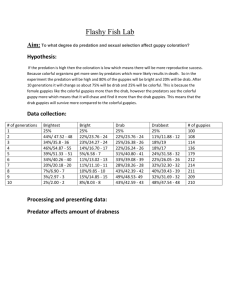
PROGRESS REPORT – 18 SEPTEMBER 2012 I. Focal Streams 1. Mark Recapture – Vital statistics: The core of our project is four replicate introduction experiments, each executed in a previously guppy-free headwater tributary of the Guanapo river. Each replicate was initiated with individually marked guppies derived from a high predation locality in the lower Guanapo River. We saved three scales from each individual to provide a source of DNA for genotyping and pedigree reconstruction. We exhaustively census each locality once per month, identify, measure and photograph all marked fish, plus photograph, measure and mark all new recruits (individuals that are >14mm standard length, which is just below the size at which the fish attain maturity). We initiated the first two introductions in March 2008 and the second two in March 2009. As of September 5, 2012, we completed our 55th consecutive monthly sampling. Data has been entered into our on-line data base for the first 54 censuses. We have completed collecting data on length and morphology from the photographs associated with our first 54 censuses, but have not yet linked the photographs to our on-line data base. At the end of the 52nd census (summarized in preparation for the August 2 submission of a NSF grant), we have marked the following number of individuals: Lower Lalaja: 6,183 (3,248 females) Upper Lalaja: 11,188 (5,781 females) Caigual: 2,874 (1476 females) Taylor: 5,058 (2,693 females) Total: 25,303 (13,198 females). Because we have multiple recaptures for most fish, the accumulated number of data entries (and associated photographs) were: Lower Lalaja: 33,577 Upper Lalaja: 60,137 Caigual: 12,476 Taylor: 14,330 Total: 120,520 We add approximately 8,000 new individuals to the data base per year and 3,500 new data points per monthly census. The figure on the right illustrates our estimated probably of recapture if alive. It shows that each census represents a very high quality sampling of each population. What we learn from each census: Each census provides a point estimate of population size, size structure, per capita recruitment rate, individual growth rate, population growth rate, and spatial distribution of guppies in the stream. All fish are collected in a spatially explicit fashion, kept separated by where they were collected throughout processing, then returned to the point of capture. These data can be combined with measurements of stream area and topography to provide information on population density, which can then be integrated with the associated work on the population biology of the killifish (Rivulus hartii) that is the only other fish species found in this portion of stream and with the ongoing sampling of the ecosystem. The underlying mark-recapture statistics reveal information on mortality rate, life span and, when combined with the pedigree, details on individual reproductive success, which in turn can lead to a diversity of new questions. Sample project derived from the monthly censuses: Here we present one example of the kind of questions that can be addressed with these census data, in addition to addressing the central goals of the proposal. The following work recently appeared as: Auer, S. K, A. Lopez-Sepulcre, T. Heatherly, T. J. Kohler, R. D. Bassar, S. A. Thomas and D. N. Reznick. 2012, J. Anim. Ecol. 81: 818-826. This project synthesized early results from the mark-recapture study with the periodic ecosystem assessments of resource availability. Abstract: 1. Current environmental conditions as well as those in the recent past and during the juvenile stage can have significant effects on adult performance and population dynamics, but their relative importance remains unexplored. 2. We examined the influence of food availability in the present, two months prior and during the juvenile stage on adult somatic growth rates in wild Trinidadian guppies (Poecilia reticulata). 3. We found that events occurring during early and later parts of an individual’s ontogeny both had important consequences for adult growth strategies, but the direction of their influence differed. Low food levels occurring in the present and 2 months prior in the adult stage both had negative effects on adult growth. In contrast, low food levels during the juvenile stage generally led to faster adult growth rates. However, these effects of juvenile food level diminished as the food level experienced two months prior increased. 4. These results suggest that the same conditions at different life stages can have different effects on the state of the organism and its subsequent short- and long-term growth strategies. They also suggest that, while juvenile conditions can have lasting effects on adult performance, the strength of their effects can be dampened by subsequent environmental heterogeneity in the adult stage. 5. A simultaneous consideration of events in both the adult and juvenile stage past may therefore improve predictions for individual- and population-level responses to environmental change. 2. Laboratory life history assays: We continue to assay the life histories of the four experimental populations once per year. These assays are derived from approximately 40 juveniles collected from each focal stream in February. We begin with small juveniles because they have only a 10-20% chance of survival to recruitment, so removing them from the population has much less impact than removing adults. The fish are reared to a size of 12-15 mm in Trinidad, then transported to our lab in Fort Collins, Colorado (Cameron Ghalambor, co-PI). It then takes approximately 1 year to rear them to the second lab-born generation and quantify the life history. These same fish are used to assess the evolution of morphology and male coloration. We have completed these assays through the 1/0, 2/1 and 3/2 year cohorts (the first number refers to the number of years since the first pair of focal streams introductions was initated and the second number refers to years since the second pair of streams was initiated). We are currently collecting data from the 4/3 year sample. These assays have shown that males have evolved later ages and larger body sizes at maturity, as seen in earlier work on this system and as predicted by life history theory and that all did so within two years of the introduction. Project derived from the annual lab life history assays: Here I present one example of work that has built on the combined information that we are gathering from comparative studies, our focal stream experiments and the associated laboratory life history assays. The preliminary work on this project appeared as: Gordon, S. A. LopezSepulcre and D. N. Reznick. 2012. Predation-associated differences in sex-linkage of wild guppy coloration. Evolution 66: 912-918 (featured by Faculty of 1000) Abstract: Evolutionary theory predicts that the sex-linkage of sexually selected traits can influence the direction and rate of evolutionary change, and also itself be subject to selection. Theory abounds on how sex-specific selection, mate choice, or other phenomena should favor different types of sex-linked inheritance, yet evidence in nature remains limited. Here we use hormone assays in Trinidadian guppies to explore the extent to which linkage of male coloration differs among populations adapted to varying predation regimes. Results show there is consistently higher degree of X- and autosomal linkage in body coloration among populations adapted to low-predation environments. More strikingly, analyses of an introduced population of guppies from a high to a low predation environment suggest that this difference can change in 50 years or less. We are currently building on this work by performing an annual assay of the pattern of color expression in laboratory born females from the focal streams versus the high predation control site that they were derived from. These assays are giving us a high resolution record of how the linkage patterns of male colorations changes over time. This figure shows that there has been an apparent increase in the extent of x-linked cartenoid coloration within the first year of the introduction. Carotenoid HP Canopy Open Source Melanistic HP Canopy Source Open 3. Pedigree update: Our ability to generate an accurate pedigree for the replicate, evolving populations in the focal streams is essential to multiple goals associated with the original FIBR proposal. Pedigrees enable us to quantify individual reproductive success, estimated as the number of offspring produced by each individual, estimated as the number recruited into the adult population. These data are coupled with our assessment of individual phenotype, as derived from photographs and to in turn associate reproductive success with the evolution of key traits, including morphology, size and estimated age at maturity, and male coloration. The pedigrees also lend themselves to a diversity of other projects. One example is given below. State of the Pedigreee: More than 16,000 guppies from the four focal populations have been genotyped at 10 highly polymorphic microsatellite loci, Pedigree analyses are conducted using the program Colony [1-2], which uses maximum likelihood methods to reconstruct nested full- and half-sibship relationships and identify the most likely parents. Colony’s integrated method of sibship and parentage reconstruction, its sophisticated approach to dealing with genotyping error (two types of error specified individually for each locus), and other features, make it the most powerful software available for inferring parentage in complex systems. We obtained precise genotyping error rate estimates based on repeated genotyping of hundreds of individuals (>600 replicated observations per locus). Colony’s analytical power comes at a cost: analysis run times increase greatly with offspring number, per locus genotyping error, and polygamous mating (exemplified by guppies), and is also influenced by number of candidate parents and the number of loci employed. Consequently, Colony runs on monthly batches of guppy recruits can require up to a week (or more) on a multicore computer. However, pedigree reconstructions of the first three years (6-9 generations) of one of the focal populations (lower Lalaja) are illustrative of some of our progress to date. Two parent assignment rates averaged ~80% over 3345 guppies genotyped, and parentage assignments were robust, as measured by high levels of concordance obtained from replicate Colony runs using different random number seeds. However, although average assignment rates were reasonably high, two-parent assignment rates were negatively correlated with the log of the number of candidate parents, and as a consequence, declined to 60-70% in the third year. This decline in assignment rate was not attributable to a loss of genetic variation, since genetic diversity showed no decline over 3 years (year 1 (3) mean He = 0.766 (0.772); year 1 (3) mean # alleles/locus =12.0 (13.3). Simulations conducted in Cervus [3] assuming ~3000 candidate parents (typical of the focal populations in their current state) suggest that our existing panel of 10 loci should be sufficient to confidently assign parentage to 95% of offspring with greater than 80% confidence. If we increase our genotyping to 20 loci, then the simulations indicate that we can assign parents to 100% of offspring with at least 95% confidence. We believe the gap between simulations and our actual results stem from computational complications imposed by the large number of candidate parents. Going forward, we plan two remedies: we will make better use of our resampling data to refine our candidate parental pools, and we will double the number of loci genotyped. More genetic markers is expected to improve assignment rates, and by strengthening the genetic ‘signal’, may actually reduce analysis run times (Wang 2012). Importantly, the fact that no loss of genetic variation has been observed in the focal populations suggests that ample scope remains for further evolution of these populations. Project derived from pedigree data – “Beyond lifetime reproductive success. The posthumous reproductive dynamics of male Trinidadian guppies”. (manuscript submitted to Proc. Roy. Soc. London, Ser. B on 10 September 2012): Abstract: Evolution in fluctuating environments requires the long-term maintenance of genetic diversity in the population, yet selection erodes it. In semelparous populations, dormant germ banks (e.g. seeds) have been proposed as important in maintaining genetic diversity and preserving genotypes that are adaptive at different times. Such hidden storage of genetic diversity need not be exclusive to dormant banks. Effective population size and genetic diversity may be preserved in many iteroparous animals through sperm storage mechanisms in females. This allows males to reproduce posthumously and increase the effective sizes of seemingly female biased populations. Although long-term sperm storage has been demonstrated in many organisms, the understanding of its importance in the wild is very poor. We here show the prevalence of male posthumous reproduction in wild Trinidadian guppies, through the combination of mark-recapture and pedigree analyses of a multigenerational individual-based dataset. A significant proportion of the reproductive population consisted of dead males, who could conceive up to 10 months after death (the maximum allowed by the length of the dataset), and which is more than twice the estimated generation time. Demographic analysis shows that the fecundity of dead males has an important contribution to population growth and selection. Abundance (# individ. m-2) 4. Impact of guppies on the ecosystem: The 12000 ecosystems ecologists affiliated with the Total abundance 10000 project have continued with their bimonthly assessments of the ecosystem in the sections 8000 of the four focal streams where guppies were 6000 introduced and in the control reaches, upstream of the guppy introduction. Guppies 4000 are excluded from the control reaches by waterfalls. Thus far, they have documented Gatherers 8000 that the guppies significantly depress the LOL control abundance of invertebrates two years after LOL introduction 6000 UPL control their introduction (see Figure on right). This UPL introduction figure illustrates the results for just the first 4000 pair of streams that received the guppy 2000 introduction in 2008. “UPL has a thinned canopy while “LOL” had an intact forest 0 canopy. These results show that the overall 2008 2009 2010 abundance of invertebrates is higher under the thinned canopy, but that the impact of guppies in invertebrate abundance is similar in both streams. The second pair of streams generated a similar canopy effect after the first year. We have yet to complete data collection for 2011 which means that we only have data for the first year after the guppy introduction in the second pair of streams. As with the first pair, there was not an impact of guppies on invertebrate abundance after the first year. There is a tendency for primary productivity to increase in the guppy introduction sites relative to the controls, but this result (not illustrated here) falls short of statistical significance. Guppies have also significantly depressed the abundance of Rivulus in all four focal streams. These results are illustrated in the figures on the right. The upper panel illustrates the population density of Rivulus in the introduction site minus the Rivulus population density in control region, upstream of the guppy introduction site. Open symbols indicate streams with experimentally thinned canopies and closed symbols are the streams with intact canopies. “LOL” and “UPL” are the pair of streams for which guppies were introduced in 2008. Guppies were introduced to “CAI” and “TAY” in 2009. This figure illustrates Rivulus population densities for the 12 month period before guppies were introduced (-12 to 0 on the xaxis) then for either 48 (LOL and UPL) or 36 (CAI 36 (Cai and TAY) months after the guppies were introduced. Values less than zero represent those sampling periods when Rivulus population densities were higher in the control region than the introduction sites. These results show that guppies have had a larger impact on Rivulus abundance in streams with intact with intact forest canopies. The lower panel of this figure illustrates the increase in guppy population density over this same time period. Guppies depress Rivulus abundance by constricting the recruitment of juveniles. They do so either by preying on the newborn Rivulus or competing with the juveniles. The results for the invertebrates are contained in a paper, senior authored by Thomas Heatherly (PhD candidate, U. Nebraska), soon to be submitted. The first paper that reports on the results for Rivulus will be co-authored by Doug Fraser and Brad Lamphere and is accepted, pending revision for Ecology. Continued below with “Other ecosystem results” for the focal streams, No v Ja 200 n 7 M 200 ar 8 M 20 ay 08 2 Ju 00 l 8 Se 20 p 08 No 20 v 08 Ja 200 n 8 M 200 ar 9 M 20 ay 09 2 Ju 00 l2 9 Se 0 p 09 20 09 Total invert. abund. (#/m 2) 2 Chlorophyll- (mg/m ) Stage (cm) 100 5. Other ecosystem results – Seasonality and canopy effects: The two prominent 80 ecosystem effects that emerged from the study, other than the impact of guppies on 60 the ecosystem, are the effects of the rainy 40 versus dry season and canopy thinning. The gray bars block out the main dry season, 20 which runs from approximately February 3 until April. This interval represents a time when flooding events are less likely and in Pool 2 turn when the abundance of algae and Riffle invertebrates increases. The first panel in 1 this figure represents stream depth over time. The middle panel depicts the standing crop 0 of algae in pools versus riffles. The third panel represents the total abundance of -1 invertebrates. Each large rainfall event scours the 10000 pools and riffles of both algae and 8000 invertebrates. The benthic communities recover to full abundance over an interval of 6000 approximately 30 days. The difference 4000 between the thinned versus intact canopies 2000 is that primary productivity, and hence the standing crop of algae and abundance of 0 invertebrates, are higher under the thinned canopies. Guppy populations respond to seasonality by increasing reproductive output during the dry season and virtually ceasing reprocution in the wet season. The figure on the right depicts population growth rate over time from March 2008 through Sept. 2009. Population growth is concentrated during the dry season, when resources are abundant. Growth is also more rapid under the thinned canopy (red line) again because resources are more abundant. The figure below depicts the per capita recruitment of new individuals. Peaks that correspond to the main dry season (February through April) and the lesser dry season in September and October. Recruitment increases during each dry interval then declines during rainy periods, presumably tracking food availability. This same pair of figures illustrates the effects if thinned versus intact canopies. The red line again represents the population size and recruitment in the stream with a thinned canopy while the black line represents the data for the stream that has the intact canopy. The data are just for the first pair of streams and represents the time period from March 2008 through August 2009. The rate of recruitment was equal in both sites for the first dry season, presumably because resources were not limiting in either stream. The rate of recruitment was higher for the treatment with the thinned canopy in the remaining two dry seasons, presumably reflecting a significantly higher level of per capita food availability under the thinned canopy. Literature Cited 1. 2. 3. Jones, O.R. and J.L. Wang, COLONY: a program for parentage and sibship inference from multilocus genotype data. Molecular Ecology Resources, 2010. 10(3): p. 551-555. Wang, J.L., Computationally efficient sibship and parentage assignment from multilocus marker data. Genetics, 2012. 166: p. in press. Kalinowski, S.T., M.L. Taper and T.C. Marshall, Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Molecular Ecology, 2007. 16(5): p. 1099-1106.