TEACHER INSTRUCTIONS - California Lutheran University
advertisement

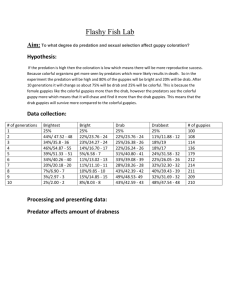
TEACHER INSTRUCTIONS- Guppies Galore! Jennifer Lockwood, Newbury Park High School (jlockwood@conejo.k12.ca.us) Nancy Monson, Sunset High School (Nancy_Monson@beavton.k12.or.us) SUBJECT: EVOLUTION Estimated Total Time: Three 50-minute periods or two 90-minute block periods California State Standards: Evolution The frequency of an allele in a gene pool of a population depends on many factors and may be stable or unstable over time. As a basis for understanding this concept: Students know variation within a species increases the likelihood that at least some members of a species will survive under changed environmental conditions. Evolution is the result of genetic changes that occur in constantly changing environments. As a basis for understanding this concept: Students know how natural selection determines the differential survival (and reproduction) of groups of organisms. Purpose/ Lesson Objectives: To examine competing evolutionary forces, natural vs. sexual selection, as well as revisit components of the scientific method using an excerpt from Jonathan Weiner’s The Beak of the Finch and John A. Endler’s 1980 guppy research as published in Evolution. An optional follow up activity will examine two different types of natural selection pressure on the evolution of human skin color. Key Vocabulary: Natural selection, sexual selection, heritable, headwater, camouflage, heterogeneous, phenotype, melanin, folic acid Assessment: Scientific method and conclusion question handouts, two graphs, Study questions TEACHER INSTRUCTIONS- Guppies Galore! Day 1 (traditional schedule) 1. Have students read text pg. 88-94 (up to stopping point #1) of “The Beak of the Finch.” 2. Pass out the handout titled “Identifying Components of the Scientific Method.” Students will complete this handout using Endler’s 10 pond greenhouse experiment at Princeton University. 3. Students should finish this for Homework. Day 2 4. Now that students have examined Endler’s experiment and how it was set up, pass out the Data Page which shows some of the data Endler collected in this experiment (data was pulled from Evolution, Vol. 34, Issue 1 (Jan. 1980) 76-91, Table 3: Means from the greenhouse experiment). Students will construct two bar graphs comparing 1) Relative length of spots for each color vs. predation intensity & 2) Relative height of spots for each color vs. predation intensity (using relative length and height since it removes guppy body size as a variable). 5. After student graphs are completed, have students analyze the graphs and list what conclusions they draw on the Handout from Day 1. Finish for Homework if necessary. Day 3 6. Pass out “The Beak of the Finch” reading #2, pg. 94-95 which describes the conclusions Endler drew when analyzing his data. Have students read and list Endler’s conclusions. After approximately 15 minutes, discuss with students- how similar was their list of conclusions to Endler’s conclusions? 7. Pass out the last segment, reading #3, of “The Beak of the Finch” (to pg. 96) and Handout 2: Conclusion Questions- Guppies Galore!. This segment discusses Endler’s field experiment testing this same hypothesis. 8. To illustrate two competing but different forces of natural selection, students can read the article titled: Skin Color Evolution in Naked Primates (US!) and answer the accompanying study questions. Students can begin the Follow-up activity in class after they turn in their Guppy Handouts 1 & 2 attached to their two graphs. They can finish the follow up reading and questions for Homework. READING #1: Excerpt from The Beak of the Finch, by Jonathan Weiner (pg. 88-96) This is a struggle of struggles, or war of wars, that Darwin could only imagine, a war in which the forces of sexual selection wrestle with the forces of natural selection, pushing and pulling a living form this way and that, down through the generations. John Endler, the author of Natural Selection in the Wild, has been observing this conflict for years, in one of the most elegant and precise demonstrations of evolution in real time. What the Grants are to Darwin’s finches, Endler is to guppies. His guppies are not the variety they sell in pet stores (he considers those trash fish). His guppies live in northeastern South America, in the small streams that zigzag down the mountains of Venezuela, Margarita Island, Trinidad, and Tobago, flashing through steep, undisturbed green forests and then the broad spreads of the old cacao and coffee plantations, on their way to the Caribbean Sea and the Atlantic. The male guppies wear black, red, blue, yellow, green, and iridescent spots in varying sizes, shapes, hues, and combinations. In fact their spots vary so much that they are like fingerprints: no two guppies are alike. These spots, like the beaks of Darwin’s finches, are heritable. Although the exact placement and arrangement of spots is unique, each guppy inherits its particular palette of colors, and also the general size and brightness of the ensemble, from its parents. The spots only show up on the males (they can be made to appear on the female guppies with testosterone treatments). Like minute variations in the beaks of finches, the spots on a guppy are the sorts of details that one might imagine are beneath the notice of natural selection. Nature may scrutinize the slightest variation, but there are some things even Darwin’s process cannot see. Design could not possibly govern a thing so small. In the 1970s, while Peter and Rosemary Grant were watching the finches of the Galapagos, Endler began watching the guppies of Venezuela’s Paria Peninsula, and Trinidad’s Northern Range. There the streams run down the mountains roughly parallel, as if in a series of vertical stripes. The streams are clear, swift, and clean, deeply shaded by tropical evergreens and punctuated by waterfalls. Their beds are lined with brilliant, manycolored gravel, much like the floors of the fish tanks in the pet stores. It is obvious to anyone who has ever tried to watch a school of these guppies against the parti-colored sands and pebbles of a streambed that the spots are excellent camouflage. In fact you could watch one of these clear streams for quite a while before you noticed the guppies at all, because they tend to swim close to the gravel while the sun is out. The fish need this camouflage because they have seven enemies: six species of fish and one freshwater prawn. All seven of these enemies hunt guppies from dawn till dusk. The most dangerous is Crenicichla alta, a cichlid fish, which eats about three guppies an hour; the least dangerous is Rivulus hartii, which eats one guppy in about five hours. Endler found guppies and at least a few of their enemies in almost every section of almost every stream, from the headwaters near the summit of each mountain to the plains and plantations below. Neither the guppies nor the guppy eaters can swim up a waterfall, and the population of each section of stream tends to stay put. (Sometimes a few fish get swept downstream, but none of them can get back up.) High up near the headwaters of each stream, the only enemy the guppies have is the comparatively mild-mannered Rivulus hartii. But moving downstream, section by section, the population of guppies lives and dies in the company of more and more of its enemies, until down near the base of each mountain, the stream is loaded with all seven of the guppy eaters. So a graph of risk and danger runs with the current. For the guppies, the higher in the stream, the lower the risk; and the lower in the stream, the higher the risk. In stream after stream the intensity of natural selection is graduated in the same way: gentle pressure among the guppies at the top, violent pressure among the guppies at the bottom. Endler saw that the streams would make a wonderful natural laboratory for the study of natural selection. He developed standardized methods of measuring guppy spots, as careful and ritualized as the Grants’ methods with Darwin’s finches. He learned to anesthetize and photograph each guppy he caught. (Like Darwin’s finches, the guppies have met very few human beings, so they are easy to catch.) From the photographs he recorded the color and position of each spot of each and every male guppy, dividing each guppy into dozens of sectors to make a standardized guppy map that is easy to read, to tally, and to enter into a computer. When Endler analyzed his surveys he discovered a pattern. The spots on each guppy look chaotic, but the spots of all of the populations of guppies in a stream, taken together, from the headwaters down to the base, have a kind of order. The spots on each population of guppies bear a simple relationship to the number of guppy eaters in their part of the stream. The more numerous the guppies’ enemies, the smaller and fainter the guppies’ spots. The fewer their enemies, the larger and brighter their spots. The lucky guppies in the headwaters wear sporty coats of many colors, and each color is represented by big clownish splotches. Many of their spots are blue. These blue spots are iridescent, like the Day-Glo patches that cyclists wear; they flash as the fish swim, and they can be seen a great distance through the clear water. Meanwhile the guppies downstream tend to wear conservative pin dots of black and red. The spots are almost vanishingly small. Most wear only a tiny amount of blue. Endler looked at his data from stream after stream. In every one of them, the size and number of spots ran steeply downhill. And Endler drew the same sort of conclusion that Lack did when he noticed the patterns of beaks in the Galapagos. Endler thought he could see the hand of natural selection at work among the guppies. The greater the pressure from their predators, the more camouflage they wear; the less the pressure, the slighter the camouflage. Of course, that interpretation did not explain why guppies are colorful at all. If they are in some danger everywhere, even in the headwaters, then why doesn’t natural selection favor the best camouflaged guppy everywhere? The answer is that a male guppy has more to do in life than merely survive. It also has to mate. To survive it has to hide among the colored gravel at the bottom of its stream and among the other guppies of its school. But to mate it has to stand out from the gravel and stand out from the school. It has to elude the eyes of the cichlid or the prawn while catching the eyes of the female guppy. The gaudier the male, the better his sex life. He is more popular among females, and he gets many chances to pass on his gaudy genes – as long as he lives. In a quiet spot near the headwaters of the stream his life is likely to be long and happy and he may father innumerable gaudy children. But in a spot near the base of the stream he may not father a single guppy before he vanishes down the gullet of a cichlid. The quieter the colors of a male, the less luck he has in courting females. On the other hand he is likely to have more time to try, because the less he stands out among his own kind, the less he stands out among his enemies. This is not just a problem for Trinidadian guppies. Wherever males court females, or females court males, whether the signals are a bright splash of color, as in guppies or redwinged blackbirds, or loud far-carrying songs, as in frogs and crickets, their broadcasts are always in danger of being interpreted by the enemy. Strong colors or loud calls can attract a mate from one side and a predator from the other. Every bullfrog calling in the night is in the dangerous spot of Romeo calling out beneath the balcony of the house of Capulet. A few species have found ways to finesse this problem. Among fish, some wrasses change color only very briefly, to flash a sexual signal in dangerous water – the equivalent of a sexy whisper, pssst! Looking at his guppy data, Endler read into them a struggle between two contending forces. Everywhere in the stream the gaudier fish produce gaudier young – pushing the next generation toward loud colors and self-advertisement. And everywhere in the stream the quieter fish produce quieter young, pushing the next generation toward modesty. In the relative safety of the headwater the gaudier guppies live long enough to win many females before they are eaten, so the population evolves in the direction of greater and greater gaudiness, and almost every male wears a coat of many colors. But in the dangerous waters at the base of the mountain the gaudy guppies live such a short time that they are outreproduced by the modest guppies. So the whole population evolves in the direction of greater and greater drabness. Males court females at distances of 2 to 4 centimeters, and from there the little spots are visible; but from farther away the males blend into the gravel. So the small-spotted guppies can blend into the background in the eyes of their predators, Endler says, “yet still be visible and stimulating to females.” When Endler first began studying the guppy streams, he was in the same position as David Lack after the Galapagos. Endler could see patterns that strongly suggested the forces of selection at work. He did not actually see selection shaping the patterns, but the closer he looked the more he was sure that the hand that shaped the patterns really was the hand of natural selection. Within the broad patterns he kept finding curious subsidiary patterns. For instance in a few of the headwaters there are prawns. In these headwaters the guppies favor red spots. This red shift makes sense because although guppies and other fish see more or less the same colors that humans do, prawns and shrimp are red-blind – they cannot see the last band in our rainbows. So in those particular headwaters, male guppies with big red spots can show off to female guppies while hiding from the prawns. Back in the 1940s, Lack made his selectionist argument about Darwin’s finches without trying to measure it in the field to see if he was right. But Endler went the extra step: he decided to test the predictions of his theory by trying to detect these processes in action. He built ten ponds in a greenhouse at Princeton University. Four of the ponds were about as wide, deep, and long as the low-water territories of Crenicichla alta. The other six ponds were about the size of the headwater streams with the comparatively mild-mannered Rivulus hartii. Endler put black, white, green, blue, red, and yellow gravel in the bottom of his artificial ponds and pumped water through them to give them a current, like the streams in the wild. Meanwhile, Endler collected guppies from up and down a dozen streams in Trinidad and Venezuela. In some places he took guppies that lived with just one predator, in some places guppies that lived with two predators, and so on up to the maximum, seven. He wanted stocks of wild guppies that had evolved under the whole spectrum of guppy menace, that were coping in the wild with every level of danger. He bred each stock in a separate aquarium. When the artificial streams were ready for his guppies, Endler took five pairs at random from each stock and put them all together into two of his ponds to let them breed and mingle and get used to their new homes. Guppies can give birth at the age of five or six weeks, and a female guppy can spawn a lot of baby guppies, so it did not take long for the populations to double. After a month he took guppies from those two ponds and used them to stock two more ponds. A month after that, he had enough guppies to seed each of his ten ponds with two hundred fish per pond. What he had done, in effect, was to shuffle and reshuffle the deck. He now had a highly heterogeneous assortment of guppies. They had all kinds of spots, and their spots were completely random with respect to the gravel at the bottom of their homes. He let these guppies breed in their new streambeds for months. Then he added a few of their natural enemies to the streams, according to a careful plan. The evolutionary experiment had begun. According to his prediction, the guppies should now evolve rapidly. The guppies in each tank should begin to look more like guppies that live with that same set of predators in the wild, they should come to look more like the gravel in their particular stream, and those in the most dangerous tanks should come to mimic the gravel more closely than those in the safer tanks. After five months, Endler took his first census. He drained each stream, counting every male’s spots and noting their position, anesthetizing them, photographing them, as he had done in the wild, and then starting up the stream again. Nine months later he took a second census. By that time nine or ten generations had passed in the lives of his guppies. STOPPING POINT #1! (to be continued) “The Beak of the Finch” reading #2: Some of the guppies were safe, with no enemies. These guppies got gaudier between the foundation of the colony and the first census, and they got gaudier still by the time of the second census. The males evolved more and more spots, bigger and bigger spots, wilder and wilder palettes of spots. Meanwhile males in tanks with the dangerous cichlids evolved fewer and fewer, smaller and smaller spots. They were still visible to females, but they got less and less visible to cichlids, who strike from 20 to 40 centimeters away. These guppies mostly dropped the blue and the iridescent spots, their Day-Glo patches, just like guppies that live with cichlids in the wild. Endler measured these differences as meticulously as the Grants measure finch beaks. “Spot height, spot area, total area, and total spot area relative to body area also decreased significantly with increased predation intensity,” he reports. The fish themselves changed size, too. Full-grown guppies in the dangerous tanks were smaller, while mature guppies in the safe waters were larger – again, just as in the wild. Each tank had a different bottom: different mixes of gravel colors and different gravel sizes. In the pools with no predators the guppies did not change their spots to match the gravel – the opposite. Their spots evolved to be smaller than the big gravel and larger than the small gravel, making the males easier and easier to see, like chameleons in reverse. They carried more iridescent spots, and a wider palette of colors per fish, and generation after generation they looked less like their background, all of which is just what we would expect if they were competing for attention. Sexual selection was operating to make males as different from the gravel bottom as possible. If only one force or the other had been operating, just natural selection or just sexual selection, the guppies would not have evolved in this remarkable way. Without natural selection all of the fish would have gotten gaudier. Without sexual selection none of them would have gotten gaudier. But the safe ones did get much more colorful, adding, in particular, blue spots. It is probably not a coincidence that guppies’ retinas are exquisitely sensitive to blue. Almost all males carry some blue somewhere, even in the most dangerous waters – it may be a sine qua non of courtship. The fish had evolved in Endler’s greenhouse until they replicated the patterns that they display in nature, and they had done so in a very short time. Of course, Endler’s streams were artificial. He had not seen natural selection in the wild. A skeptic could still argue that Endler was wrong about his explanation for the pattern in the wild. So Endler figured out a way to run the same sort of evolutionary experiment in nature. STOPPING POINT #2! (to be continued) “The Beak of the Finch” reading #3: Early in his fieldwork he had found a Trinidadian stream that contained the guppy eater Rivulus hartii, but no guppies. About 2 kilometers away was a second stream that contained both guppy eaters and guppies. Endler took a random sample of about two hundred guppies from one of the high-danger zones in the second river. He measured each and every one, as usual, and then he transferred them to the safe place in the first river. He took a sample of their descendants more than a year later, after a passage of fifteen generations. The males in the safe stream were now much gaudier than their immediate ancestors, who were still living in the stream next door and coping with many enemies. The immigrant males wore bigger spots, and more of them, and each male sported a wider assortment of colors. Natural selection had acted just as predicted. Evolution had run as fast in the wild as in the greenhouse. Everywhere in those streams, daily and hourly, natural selection in the form of cichlids and prawns is not just metaphorically but literally scrutinizing the male guppies. The result of enemy predations on each generation keeps pushing the males to blend in with the stream bottom. At the same time, daily and hourly, sexual selection in the form of female guppies is scrutinizing those same males. The result of their choices is that generation after generation of males is pushed to stand out. Now it is clear why there is such virtually infinite range of variation in the way each individual male guppy is spotted. Many different random patterns of splotches will be equally good camouflage, because the streambed patterns are random too. It would not help the guppies to sport the same pattern as all the others, and in fact it would hurt them. If the guppy males all looked alike, their enemies could develop a search image – an inner template. They would search for that pattern, as we search for the face of a friend in a crowd. The rare misfit would have a great advantage. Meanwhile the females would go for the unusual males, too, and that would drive more and more diversity of patterns, So in this respect natural selection and sexual selection cease to oppose each other and push in the same direction: toward almost infinite diversity. We see here an example of what Darwin saw in the wide world. He understood that his simple process can lead to the most bewildering and chaotic-looking diversification and variety – but underneath, the driver is as simple and plain and commonsense as ever, “small consequences of one general law leading to the advancement of all organic beings, -- namely, multiply, vary, let the strongest live and the weakest die.” The guppy experiments suggested to Endler what Darwin’s finches were suggesting at about the same time to the Galapagos finch watchers: that natural selection can be swift and sure. The process is following along, all around us, much faster than Darwin ever dreamed. Endler’s study is leading him into deeper and deeper waters. He now suspects that the guppies’ spots, their mating habits, and their color vision are all evolving simultaneously, with change in any one of these factors driving change in all the others. To measure variations in the guppies’ retinas, Endler is collaborating with physiologists. These “hard science” types often remind him how “soft” the science of evolution is perceived to be by the outside world – even by biologists. “I was talking with someone in vision physiology the other day,” Endler says, “and he told me, ‘Wow, I had no idea that the subject was so rigorous. I had no idea that you actually did experiments.’ “We have a serious public-relations problem,” Endler says. “People don’t realize this is real science.” THE END! Data Table: Relative Spot Length and Spot Height in Male Guppies (Poecilia reticulate) Exposed to Different Predation Pressures (Endler, 1980) Trait Relative spot length* Relative spot height** Black Spot color Red Blue Predation pressure‡ C R K Iridescent 12.58 13.99 18.85 16.87 17.81 19.51 16.00 19.54 21.57 14.94 17.91 52.65 All spots*** 15.94 17.19 17.49 C R K 26.79 30.41 32.02 30.89 34.93 37.04 30.45 36.82 37.31 24.58 32.47 90.18 29.22 33.51 34.31 (fish in fine-grain gravel pond) ‡Predation pressure: C = intense predation R = weak predation K = no predation *Relative spot length = arc sin √spot length / body length (as measured in mm) **Relative spot height = arc sin √spot height / body height (as measured in mm) ***Mean spot size per fish for black, red, blue, yellow, gold, cream, green, and iridescent spots. Source: Endler, John; Natural Selection on Color Patterns in Poecilia reticulate; Evolution, Volume 34, Issue 1 (Jan., 1980), 76-91. Name _____________________________________ Period __________ Date _______ Identifying Components of the Scientific Method- Guppies Galore! Directions: After reading “The Beak of the Finch” reading #1 (pg. 88-94) go back through the reading and identify the components of the scientific method that John A. Endler used in his classic evolution experiment using guppies in northeastern South America. Fill in those components in the outline provided below. 1. What was the question Endler was trying to answer? (write this as a question): 2. What was Endler’s hypothesis? (write in “If…. and… then….” format): 3. What background information did Endler need to know about guppies and their environment (including predators) in order to design an experiment using guppies? List 5 pieces of specific information Endler (and now you!) learned about guppies while observing them in nature. 1. 2. 3. 4. 5. 4a. In any experiment, the set up is critical to testing the hypothesis. As a visual representation of the procedure, draw a sketch of Endler’s 10-pond experimental set up (in Princeton’s greenhouse). Be sure to include the following: label which ponds have high/low predator intensity; label which ponds represent headwater/downstream; the number of initial fish in each pond; what type of bottom surface the ponds contained; and how often the fish were counted. 4b. Describe the initial phenotypic mixture of the original guppy population for each pond in this experimental set up. 5. In order to find patterns and analyze their experimental results, the data must first be organized into data tables. Using a ruler, draw a data table Endler could have used to organize his results. Be sure to give your data table a title! (ex: Data Table 1: An Earthworm’s Heart Rate as it Relates to Soil Temperature) Hint: Think about what information can be measured quantitatively and also tests the hypothesis. **STOPPING POINT #1! (to be continued)** 6. The information in data tables is often displayed in a graph. You will be constructing two bar graphs using some of the actual data Endler collected from his 10 pond experiment. Be sure to draw the graphs big enough and be sure to complete the graph by labeling both the x and y-axes and by giving the graph a title! a) Relative length of spots for each color vs. predation intensity b) Relative height of spots for each color vs. predation intensity 7. After you have completed both graphs, analyze them. What conclusions can you draw? What did the experimental results tell Endler (and now you!)? List 4 of these below. 1. 2. 3. 4. 8. Read a second excerpt from “The Beak of the Finch” (pg. 94-95 up to stopping point #2). Now that you have read the conclusions Endler himself drew from his data, what were those conclusions? (Don’t worry if they’re different from what you wrote!) 1. 2. 3. 4. **STOPPING POINT #2! (to be continued)** Name _____________________________________ Period __________ Date _______ Conclusion Questions- Guppies Galore! Directions: Read the third (and last!) segment of “The Beak of the Finch” (to pg. 96) and answer the questions below. 1a. Endler also tested his hypothesis in nature. He took about 200 guppies from a highdanger zone in a river and transferred them to a safe place in another river. What is the phenotype of these high danger zone guppies before they are released into the safe river? 1b. What is the advantage to this appearance while living in a high danger zone? 1c. What is the advantage of spots (in general)? 1d. Do the above advantages (from 1b and 1c) work together in the same direction or against each other? 2. After 15 generations in a safe section of a river, all of the males in the safe river were much gaudier than their immediate ancestors taken from the high-danger zone. What caused this shift in phenotype? 3a. If all of the guppies had the same exact type of spot pattern (not necessarily colors), would this increase or decrease the guppies chances of survival? 3b. Why? 4. How is Endler’s work an example of “real science?” Skin Color Evolution in Naked Primates (US!)- TEACHER VERSION (complete info.) ~overview~ Human skin color can range from almost black to pinkish white in people. In general, people with ancestors from sunny regions have darker skin than people with ancestors from regions with less sunlight. (However, this is complicated by the fact that there are people whose ancestors come from both sunny and less-sunny regions; and these people may have skin colors across the spectrum). On average, women have slightly lighter skin than men. Skin color is determined by the amount and type of the pigment melanin in the skin. Melanin comes in two types: phaeomelanin (red to yellow) and eumelanin (dark brown to black). Both amount and type are determined by four to six genes which operate under incomplete dominance. One copy of each of those genes is inherited from the father and one from the mother. Each gene comes in several alleles, resulting in a great variety of different skin colors. Dark skin protects against skin cancer, mutations in skin cells induced by ultraviolet light. Light-skinned persons have about a tenfold greater risk of dying from skin cancer under equal sun conditions. Furthermore, dark skin prevents UV-A radiation from destroying the essential B12 vitamin folate (folic acid). Folate is needed for the synthesis of DNA in dividing cells and too low levels of folate in pregnant women are associated with birth defects. While dark skin protects vitamin B, it can lead to a vitamin D deficiency. The advantage of light skin is that it lets more sunlight through, which leads to increased production of vitamin D3, necessary for calcium absorption and bone growth. The lighter skin of women results from the higher calcium needs of women during pregnancy and lactation. The evolution of the different skin colors is thought to have occurred as follows: the haired ancestor of humans, like modern great apes, had light skin under their hair. Once the hair was lost, they evolved dark skin, needed to prevent low folate levels since they lived in sun-rich Africa. (The skin cancer connection is probably of secondary importance, since skin cancer usually kills only after the reproductive age and therefore doesn't exert much evolutionary pressure.) When humans migrated to sun-poorer regions in the north, low vitamin D3 levels became a problem and light skin color evolved. Dark-skinned people who live in sun-poor regions often lack vitamin D3, one reason for the fortification of milk with vitamin D in some countries. The Inuit are a special case: even though they live in an extremely sun-poor environment, they have retained their relatively dark skin. This can be explained by the fact that their traditional animal-based diet provides plenty of vitamin D. Albinism is a condition characterized by the absence of melanin, resulting in white skin and hair; it is caused by a genetic mutation. Research on skin color variability The color of human skin varies from dark brown to pale pink. In attempting to discover the mechanisms that have generated such a wide variation in human skin color, Nina Jablonski and George Chaplin (2000) discovered that there is a high correlation between the coloration of the human skin of indigenous peoples and the average annual ultraviolet (UV) radiation available for skin exposure where the indigenous peoples live. Accordingly, Jablonski and Chaplin plotted the whiteness (W) of skin coloration of indigenous peoples who have stayed in the same geographical area for the last 500 years versus the annual UV available for skin exposure (AUV) for over 200 indigenous persons and found that whiteness (W) of skin coloration is related to the annual UV available for skin exposure (AUV). According to Jablonski and Chaplin’s study in 2000 (p. 67, formula coefficients have been rounded to one-figure accuracy) where the whiteness (W) of skin coloration is measured as the percentage of light reflected from the upper inner arm at which location on humans there should be minimal tanning of human skin due to personal exposure to the sun, a lighter skinned human would reflect more light and would have a higher W number. Judging from the above linear fit to the empirical data, the theoretical maximum whitest human skin would reflect only 70 per cent of incident light for a hypothetical indigenous human-like population that lived where there was zero annual UV available for skin exposure (AUV = 0 in the above formula on pg. 67). Jablonski and Chaplin evaluated average annual UV available for skin exposure (AUV) from satellite measurements that took into consideration the measured daily variation in the thickness of the ozone layer that blocked UV hitting the earth, measured daily variation in opacity of cloud cover, and daily change in angle at which the sunlight containing UV radiation strikes the earth and passes through different thicknesses of earth's atmosphere at different latitudes for each of the different human indigenous peoples' home areas from 1979 to 1992. Jablonski and Chaplin proposed an explanation for the observed variation of untanned human skin with annual UV exposure. By Jablonski and Chaplin's explanation, there are two competing forces affecting human skin color: 1. the melanin that produces the darker tones of human skin serves as a light filter to protect against too much UV light getting under the human skin where too much UV causes sunburn and disrupts the synthesis of precursors (vitamin B12 folate/ folic acid) necessary to make human DNA; versus 2. humans need at least a minimum threshold of UV light to get deep under human skin to produce vitamin D3, which is essential for building and maintaining the bones of the human skeleton. Jablonski and Chaplin note that when human indigenous peoples have migrated, they have carried with them a sufficient human gene pool so that within a thousand years, the skin of their descendants living today has turned dark or turned white to adapt to fit the formula given above--with the notable exception of dark-skinned peoples moving north, such as to populate the seacoast of Greenland, to live where they have a year-round supply of food, such as fish, rich in vitamin D, so that there was no necessity for their skin to turn white to let enough UV under their skin to synthesize the vitamin D that humans need for healthy bones. In considering the color of human skin in the long span of human evolution, Jablonski and Chaplin note that there is no empirical evidence to suggest that the human ancestors six million years ago had a skin color different from the skin color of today's chimpanzees— namely pale-skinned under black hair. But as humans evolved to lose their body hair a parallel evolution permitted human populations to turn their base skin color dark or white over a period of less than a thousand years to adjust to the competing demands of 1) increasing eumelanin to protect from UV that was too intense and 2) reducing eumelanin so that enough UV would penetrate to synthesize enough vitamin D. By this explanation, in the time that humans lived only in Africa, humans had dark skin to the extent that they lived for extended periods of time where the sunlight is intense. As some humans migrated north, over time they developed white skin, though they retained within the gene pool the capability to develop black skin when they migrated to areas with intense sunlight again, such as across the Bering Strait and south to the Equator. Dissenting views Ashley Robins (1991) has argued that the white skin of indigenous peoples living north of 40° north latitude need not arise necessarily from the natural selection pressures on those humans to synthesize enough vitamin D. Robins argues that people with dark skin can get enough vitamin D if they spend enough time in the spring and summer sun, and he hypothesizes that the fatty tissues of humans would store enough vitamin D to maintain human health during the northern-most winters. Origins of black skin in humans Scientists have correlated the wide variations in human skin color with the mutations in one gene, the MC1R gene (Harding et al 2000:1351). The "MC1R" label for the gene stands for melanocortin 1 receptor, where "melano" refers to black, "melanocortin" refers to the hormone stimulant produced by the pituitary gland that orders cells to produce the melanin that makes skin cells black, the "1" in the MC1R gene name specifies the first family of melanocortin genes, and "receptor" indicates that the protein from the gene serves as a signal relay from outside the cell to inside the cell --to the place in the cell where the black melanin is synthesized. Accordingly, the MC1R gene specifies the amino acid sequence in the receptor protein that relays through the cell membrane the hormone signal from the pituitary gland to produce the melanin that makes human skin black. Many variations in the amino acid sequence of this receptor protein result in whiter or darker skin. The human MC1R gene consists of a string of 954 nucleotides, where each nucleotide is one of the four bases Adenosine (A), Guanine (G), Thymine (T), or Cytosine (C). But 261 of the nucleotides in the MC1R gene can change with no effect on the amino acid sequence in the receptor protein produced from the gene. For example, the nucleotide triplets GGT, GGC, GGA, and GGG are all synonymous and all produce the amino acid Glycine, (See a Codon Table) so a mutation in the third position in the triplet GGT is a "silent mutation" and has no effect on the amino acid produced from the triplet. Harding et al (2000:1355) analyzed the amino acid sequences in the receptor proteins from 106 individuals from Africa and 524 individuals from outside Africa to find why the color of all the Africans' skin was black. Harding found that there were zero differences among the Africans for the amino acid sequences in their receptor proteins, so the skin of each individual from Africa was black. In contrast, among the non-African individuals, there were 18 different amino acid sites in which the receptor proteins differed, and each amino acid that differed from the African receptor protein resulted in skin lighter than the skin of the African individuals. Nonetheless, the variations in the 261 silent sites in the MC1R were similar between the Africans and non-Africans, so the basic mutation rates among the Africans and non-Africans were the same. Why were there zero differences and no divergences in the amino acid sequences of the receptor protein among the Africans while there were 18 differences among the populations in Ireland, England, and Sweden? Harding (2000:1359-1360) concluded that the intense sun in Africa created an evolutionary constraint that reduced severely the survival of progeny with any difference in the 693 sites of the MC1R gene that resulted in even one small change in the amino acid sequence of the receptor protein--because any variation from the African receptor protein produced significantly whiter skin that gave less protection from the intense African sun. In contrast, in Sweden, for example, the sun was so weak that no mutation in the receptor protein reduced the survival probability of progeny. Indeed, for the individuals from Ireland, England, and Sweden, the mutation variations among the 693 gene sites that caused changes in amino acid sequence was the same as the mutation variations in the 261 gene sites at which silent mutations still produced the same amino acid sequence. Thus, Harding concluded that the intense sun in Africa selectively killed off the progeny of individuals who had a mutation in the MC1R gene that made the skin whiter. However, the mutation rate toward whiter skin in the progeny of those African individuals who had moved North to areas with weaker sun was comparable to the MC1R mutation rate of the white folks whose ancient ancestors grew up in Sweden. Hence, Harding concluded that the whiteness of human skin was a direct result of random mutations in the MC1R gene that were non-lethal at the latitudes of Ireland, England, and Sweden. Even the mutations that produce red hair with little ability to tan were non-lethal in the northern latitudes. Rogers, Iltis, and Wooding (2004) examined Harding's data on the variation of MC1R nucleotide sequences for people of different ancestry on the earth to determine the most probable progression of the skin color of human ancestors over the last 5 million years. Comparing the MC1R nucleotide sequences for humans and chimpanzees in various regions of the earth, Rogers concluded that the common ancestors of all humans on earth had white skin color under dark hair--similar to the skin and hair color pattern of today's chimpanzees. That is 5 million years ago, the human ancestors' dark hair protected their white skin from the intense African sun so that there was no evolutionary constraint that killed off the progeny of those who had mutations in the MC1R nucleotide sequences that made their skin white. However, over 1.2 million years ago, judging from the numbers and spread of variations among human and chimpanzee MC1R nucleotide sequences, the human ancestors in Africa began to lose their hair and they came under increasing evolutionary pressures that killed off the progeny of individuals that retained the inherited whiteness of their skin. By 1.2 million years ago, all people having descendants today had exactly the receptor protein of today's Africans; their skin was black, and the intense sun killed off the progeny with any whiter skin that resulted from mutational variation in the receptor protein (Rogers 2004:107). However, the progeny of those humans who migrated North away from the intense African sun were not under the evolutionary constraint that keeps human skin black generation after generation in Africa. Tracking back the statistical patterns in variations in DNA among all known people sampled who are alive on the earth today, Rogers concluded the following: 1) from 1.2 million years ago for a million years, the ancestors of all people alive today were as black as today's Africans, 2) for that period of a million years, human ancestors lived naked without clothing, and 3) the descendants of any people who migrate North from Africa will mutate to become white over time because the evolutionary constraint that keeps Africans' skin black generation after generation decreases generally the further North a people migrates (Rogers 2004). References Harding, Rosalind M., Eugene Healy, Amanda J. Ray, Nichola S. Ellis, Niamh Flanagan, Carol Todd, Craig Dixon, Antti Sajantila, Ian J. Jackson, Mark A. Birch-Machin, and Jonathan L. Rees. 2000. "Evidence for variable selective pressures at MC1R." American Journal of Human Genetics 66: 1351-1361. Jablonski, Nina G., and George Chaplin. 2000. "The evolution of human skin coloration." Journal of Human Evolution 39: 57-106. Jablonski, Nina G., and George Chaplin. 2002. "Skin deep." Scientific American 287 (4) (October): 74-82. Rees, J.L., and N. Flanagan. 1999. "Pigmentation, melanocortins, and red hair." Q. J. Med." 92: 125-131. Robins, A.H. 1991. Biological Perspectives on Human Pigmentation. Cambridge: Cambridge University Press. Rogers, Alan R., David Iltis, and Stephen Wooding. 2004. "Genetic variation at the MC1R locus and the time since loss of human body hair." Current Anthropology 45 (1): 105-108. External links Nicholas Wade. 2003. " Why Humans and Their Fur Parted Ways." New York Times (Science Times) (August 19). Summary of clues to the saga in which humans evolved to lose their hair and had to adjust, including turning from white skin to black skin, together with an estimation of the time at which humans invented clothing. The source of this article is Wikipedia, the free encyclopedia. The text of this article is licensed under the GFDL. www.absoluteastronomy.com/encyclopedia/h/hu/human_skin_color.htm Skin Color Evolution in Naked Primates (US!) ~overview~ Human skin color can range from almost black to pinkish white in people. In general, people with ancestors from sunny regions have darker skin than people with ancestors from regions with less sunlight. (However, this is complicated by the fact that there are people whose ancestors come from both sunny and less-sunny regions; and these people may have skin colors across the spectrum). On average, women have slightly lighter skin than men. Skin color is determined by the amount and type of the pigment melanin in the skin. Melanin comes in two types: phaeomelanin (red to yellow) and eumelanin (dark brown to black). Both amount and type are determined by four to six genes which operate under incomplete dominance. One copy of each of those genes is inherited from the father and one from the mother. Each gene comes in several alleles, resulting in a great variety of different skin colors. Dark skin protects against skin cancer, mutations in skin cells induced by ultraviolet light. Light-skinned persons have about a tenfold greater risk of dying from skin cancer under equal sun conditions. Furthermore, dark skin prevents UV-A radiation from destroying the essential B12 vitamin folate (folic acid). Folate is needed for the synthesis of DNA in dividing cells and too low levels of folate in pregnant women are associated with birth defects. While dark skin protects vitamin B, it can lead to a vitamin D deficiency. The advantage of light skin is that it lets more sunlight through, which leads to increased production of vitamin D3, necessary for calcium absorption and bone growth. The lighter skin of women results from the higher calcium needs of women during pregnancy and lactation. The evolution of the different skin colors is thought to have occurred as follows: the haired ancestor of humans, like modern great apes, had light skin under their hair. Once the hair was lost, they evolved dark skin, needed to prevent low folate levels since they lived in sun-rich Africa. (The skin cancer connection is probably of secondary importance, since skin cancer usually kills only after the reproductive age and therefore doesn't exert much evolutionary pressure.) When humans migrated to sun-poorer regions in the north, low vitamin D3 levels became a problem and light skin color evolved. Dark-skinned people who live in sun-poor regions often lack vitamin D3, one reason for the fortification of milk with vitamin D in some countries. The Inuit are a special case: even though they live in an extremely sun-poor environment, they have retained their relatively dark skin. This can be explained by the fact that their traditional animal-based diet provides plenty of vitamin D. Albinism is a condition characterized by the absence of melanin, resulting in white skin and hair; it is caused by a genetic mutation. Research on skin color variability The color of human skin varies from dark brown to pale pink. In attempting to discover the mechanisms that have generated such a wide variation in human skin color, Nina Jablonski and George Chaplin (2000) discovered that there is a high correlation between the coloration of the human skin of indigenous peoples and the average annual ultraviolet (UV) radiation available for skin exposure where the indigenous peoples live. Accordingly, Jablonski and Chaplin plotted the whiteness (W) of skin coloration of indigenous peoples who have stayed in the same geographical area for the last 500 years versus the annual UV available for skin exposure (AUV) for over 200 indigenous persons and found that whiteness (W) of skin coloration is related to the annual UV available for skin exposure. According to Jablonski and Chaplin’s study in 2000, where the whiteness (W) of skin coloration is measured as the percentage of light reflected from the upper inner arm at which location on humans there should be minimal tanning of human skin due to personal exposure to the sun, a lighter skinned human would reflect more light and would have a higher W number. Jablonski and Chaplin evaluated average annual UV available for skin exposure (AUV) from satellite measurements that took into consideration the measured daily variation in the thickness of the ozone layer that blocked UV hitting the earth, measured daily variation in opacity of cloud cover, and daily change in angle at which the sunlight containing UV radiation strikes the earth and passes through different thicknesses of earth's atmosphere at different latitudes for each of the different human indigenous peoples' home areas from 1979 to 1992. Jablonski and Chaplin proposed an explanation for the observed variation of untanned human skin with annual UV exposure. By Jablonski and Chaplin's explanation, there are two competing forces affecting human skin color: 1. the melanin that produces the darker tones of human skin serves as a light filter to protect against too much UV light getting under the human skin where too much UV causes sunburn and disrupts the synthesis of precursors (vitamin B12 folate/ folic acid) necessary to make human DNA; versus 2. humans need at least a minimum threshold of UV light to get deep under human skin to produce vitamin D3, which is essential for building and maintaining the bones of the human skeleton. Jablonski and Chaplin note that when human indigenous peoples have migrated, they have carried with them a sufficient human gene pool so that within a thousand years, the skin of their descendants living today has turned dark or turned white to adapt to fit the formula given above--with the notable exception of dark-skinned peoples moving north, such as to populate the seacoast of Greenland, to live where they have a year-round supply of food, such as fish, rich in vitamin D, so that there was no necessity for their skin to turn white to let enough UV under their skin to synthesize the vitamin D that humans need for healthy bones. In considering the color of human skin in the long span of human evolution, Jablonski and Chaplin note that there is no empirical evidence to suggest that the human ancestors six million years ago had a skin color different from the skin color of today's chimpanzees—namely pale-skinned under black hair. But as humans evolved to lose their body hair a parallel evolution permitted human populations to turn their base skin color dark or white over a period of less than a thousand years to adjust to the competing demands of 1) increasing eumelanin to protect from UV that was too intense and 2) reducing eumelanin so that enough UV would penetrate to synthesize enough vitamin D. By this explanation, in the time that humans lived only in Africa, humans had dark skin to the extent that they lived for extended periods of time where the sunlight is intense. As some humans migrated north, over time they developed white skin, though they retained within the gene pool the capability to develop black skin when they migrated to areas with intense sunlight again, such as across the Bering Strait and south to the Equator. References *Harding, Rosalind M., Eugene Healy, Amanda J. Ray, Nichola S. Ellis, Niamh Flanagan, Carol Todd, Craig Dixon, Antti Sajantila, Ian J. Jackson, Mark A. Birch-Machin, and Jonathan L. Rees. 2000. "Evidence for variable selective pressures at MC1R." American Journal of Human Genetics 66: 13511361. *Jablonski, Nina G., and George Chaplin. 2000. "The evolution of human skin coloration." Journal of Human Evolution 39: 57-106. *Jablonski, Nina G., and George Chaplin. 2002. "Skin deep." Scientific American 287 (4) (October): 74-82. *Rees, J.L., and N. Flanagan. 1999. "Pigmentation, melanocortins, and red hair." Q. J. Med." 92: 125131. *Robins, A.H. 1991. Biological Perspectives on Human Pigmentation. Cambridge: Cambridge University Press. *Rogers, Alan R., David Iltis, and Stephen Wooding. 2004. "Genetic variation at the MC1R locus and the time since loss of human body hair." Current Anthropology 45 (1): 105-108. The source of this article is Wikipedia, the free encyclopedia. The text of this article is licensed under the GFDL. www.absoluteastronomy.com/encyclopedia/h/hu/human_skin_color.htm Name ______________________________________ Period _______ Date _________ Study Questions- Skin Color Evolution in Naked Primates (US!) 1. List the two types of melanin and the shades of color associated with each type. 1) 2) 2a. What is incomplete dominance? 2b. Does incomplete dominance increase genetic variety or decrease genetic variety? Why? 3. Complete the following chart illustrating the health advantages and disadvantages of having generally darker or lighter skin. Skin Color Advantages Disadvantages Darker Lighter 4a. Why is vitamin B12 (folic acid) necessary for human survival? 4b. Why is vitamin D3 necessary for human survival? 5. How are the Inuit able to survive with dark skin in an extremely sun-poor environment? 6a. What corresponding two sets of data did Jablonski and Chaplin collect in their study of human skin color? (Fill in the table below.) Explain the method Jablonski and Chaplin used to collect these data. Two Sets of Collected Data How did Jablonski and Chaplin Collect Data? 6b. 1. 2. 7. Summarize the two competing natural selection pressures acting upon skin color. 8a. A lack of vitamin D is associated with a skeletal disease called rickets. Immigrants who have moved from equatorial regions to extreme northern or southern latitudes have experienced an increase in the occurrence of rickets. Explain why. 8b. Women who currently live in equatorial regions can also develop rickets. What is a cultural practice that may cause this increased risk?