Biochemistry Ch 22 396-412 [4-20
advertisement

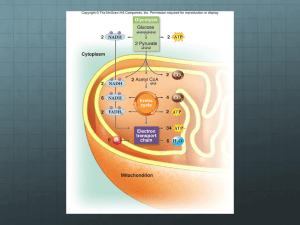
Biochemistry Ch 22 396-412 Glycolysis -principal pathways for generating ATP in cells and is present in all cell types and has ability to generate ATP with and without O2 -glucosepyruvate generates ATP via substrate-level phosphorylation and NADH -pyruvate can be oxidized to CO2 in TCA cycle and ATP made from electron transport -in anaerobic glycolysis, pyruvate and NADH are converted to lactate and ATP made without O2 -glucose can be found in diet, internal glycogen stores, and blood -glucose is major fuel for all tissues except intestinal mucosal cells, brain uses during fasting -glycolysis is an anabolic pathway providing precursors for synthesis of amino acids and 5P Reactions of Glycolysis – cleaves 1 mol glucose 2 mol pyrivate through two phases: 1. Preparative Phase – glucose phosphorylated 2x by ATP and cleaved into 2 triose phosphates 2. ATP-generating Phase – glyceraldehyde-3-phosphate (G3P) is oxidized by NAD+ and phosphorylated using inorganic phosphate, and ADP ATP a. The remaining phosphate is rearranged to form another ATP b. Since 2 mole of G3P (Trioses) are formed, 4 mol total ATP and 2 mol NADH formed, yielding a net 2 ATP, 2 NADH, 2 pyruvate from 1 glucose 1. Conversion of Glucose to Glucose-6-Phosphate- Glucose receives a phosphate from ATP to form G6P with the help of hexokinase or glucokinase in liver -commits the glucose to metabolism because G6P cannot travel back through membrane -G6P is a branch point in carb metabolism: glycolysis, pentose phosphate, glycogen synthesis -hexokinases phosphorylate glucose; various isozymes that have different kinetic properties -isozyme in pancreas (glucokinase) has higher Km than other hexokinases 2. Conversion of G6P to Triose Phosphates – G6P fructose-6-P by phosphoglucose isomerase, phosphorylated again, and cleaved into two 3-carbon fragments -F6P converted to fructose-1,6-bisP by phosphofructokinase-1, and is considered the first committed step of the pathway (requires ATP) -fructose-1,6-bisphosphate cleaved by aldolase into two triose phosphates -dihydroxyacetonephosphate (DHAP) is isomerized to glyceraldehyde-3-phosphate (a triose phosphate) -for every mole of glucose, 2 mol of G3P are produced to continue pathway 3. Oxidation and Substrate-level Phosphorylation – G3P oxidized and phosphorylated so that subsequent intermediates can donate phosphate to ADP ATP a. G3P-dehydrogenase – oxidizes aldehyde in G3P and transfers electrons to NAD+ NADH -forms high energy phosphate bond, which is the start of substrate-level phosph b. Phosphoglycerate kinase transfers phosphate from ADP to ATP and forms a 3phosphoglycerate product. c. 3-phosphoglycerate converted to 2-phosphoglycerate by phosphoglyceromutase which is then converted to phosphoenol pyruvate (PEP) by enolase and finally to pyruvate by pyruvate kinase and transfers phosphate to ADP ATP SUMMARY OF GLYCOLYSIS Glucose G6P F6P F1,6bisP G3P 1,3 BPG 3-PG 2-PG PEP Pyruvate Enzymes Hexokinase, phosphoglucose isomerase, PFK-1, aldolase, triose phosphate isomerase, glyceraldehyde 3-phosphate dehydrogenase, phosphoglycerate kinase, phosphogluceromutase, enolase, pyruvate kinase RED = ATP GENERATED BLUE = NADH GENERATED GLUCOSE + 2NAD + 2Pi + 2ADP 2 PYRUVATE + 2 NADH + 4H + 2 ATP + 2 H2O Oxidative Fates of Pyruvate and NADH – NADH must be reoxidized to NAD+ to continue reaction. NADH can be reoxidized aerobically and anaerobically -Aerobic NADH: shuttles NADH across mitochondrial membrane for electron transport (mit. Membrane impermeable to NADH) -Anaerobic NADH: NADH reoxidized by lactate dehydrogenase reducing pyruvate to lactate -fate of pyruvate depends on route of NADH oxidation; if NADH is used aerobically, pyruvate can be used for TCA cycle by its oxidation to acetyl-CoA. -NADH reoxidized to NAD+ in cytosol by reaction that transfers electrons to DHAP in glycerol-3-P shuttle and oxaloacetate in malate-asparate shuttle. NAD formed in cytosol returns to glycolysis and glycerol-3-P or malate carries reducing equivalent across mitochondrial membrane (electrons, not NADH) -if NADH is used anaerobically, pyruvate is reduced to lactate Glycerol 3-Phosphate Shuttle – major shuttle in most tissues where NAD is regenerated by glycerol-3P dehydrogenase which transfers electrons from NADH to DHAP to form glycerol 3-P, whichdiffuses through outer mit. Membrane to the inner membrane where electrons are donated to a FAD-glycerophophsate dehydrogenase, donating electrons to coenzyme Q resulting in 1.5 ATP from OxPhos. NADH(cyt) + H + FAD (mit) NAD(cyt) + FADH2(mit) Malate-Aspartate Shuttle – cytosolic NAD is regenerated by malate dehydrogenase which transfers electrons from NADH to cytosolic oxaloacetate malate -malate is transferred across inner mit membrane and exchanges malate for alphaketoglutarate -in matrix, malate is oxidized back to oxaloacetate by mitochondrial malate dehydrogenase and NADH is generated (which can donate electrons to ETC) NADH (cytosol) + NAD (matrix) NAD (cytosol) + NADH (matrix) Anaerobic Glycolysis – NADH produced from glycolysis cannot be oxidized aerobically and thus is oxidized back to NAD+ by lactate dehydrogenase. GLUCOSE + 2ADP + 2Pi 2 LACTATE + 2 ATP + 2H2O + 2H -each mol of glucose generates 2 mol ATP, 2 mol NADH, and 2 pyruvate -in anaerobic metabolism, 1 glucose forms 2 moles of lactate and only 2 mol ATP per glucose -NADH nor pyruvate are used for further energy -oxidation of pyruvate in TCA generates 12.5mol ATP per mol pyruvate, 1.5 mol of ATP generated (Glycerol-3-phosphate shuttle) or 2.5 mol ATP from malateaspartate shuttle from aerobic oxidation of cytosolic NADH, therefore, TCA produces 30-32 mol ATP per glucose Acid Production in Anaerobic Glycolysis – results in acid production in the form of hydrogen ions. Glycolysis forms pyruvic acid lactic acid, which dissiates to form carboxylate anion, lactate, and H+, which can reduce pH Tissues Dependent on Anaerobic Glycolysis – RBCs, kidney medulla, eye, skeletal muscles, all rely on anaerobic glycolysis for a portion of ATP requirements, and they have a low ATP demand to begin with. -some lactic acid generated in skin is secreted as sweat, acting as antibacterial agent -tissues with mitrochondria show both aerobic and anaerobic glycolysis Fate of Lactate – lactate is taken up by other tissues (liver, heart, skeletal muscle) and oxidized back to pyruvate -in the liver, pyruvate synthesizes glucose (gluconeogenesis), cycling of lactate and glucose between liver and peripheral tissues is called the Cori Cycle -in other tissues, lactate is oxidized to pyruvate and then to CO2 in TCA cycle -LACTATE + NAD PYRUVATE + NADH + H -the heart uses lactate as fuel; similarly, skeletal muscles can use lactate during exercise -in the brain, glial cells and astrocytes produce lactate which is used by neurons -lactate dehydrogenase is tetramer composed of A-subunits, B subunits which combine in random combinations -M4 form (4 A subunits) is used in skeletal muscle, H4 in heart Other Functions of Glycolysis – provides precursors for biosynthetic pathways, such as for ribose-5-phosphate (for nucleotides), UDP-glucose, mannose, and sialic acid -serin is synthesized from 3-phosphoglycerate, and alanine from pyruvate -triacylglycerols, glycerol-3-P converted from DHAP -liver is site of biosynthetic pathways in the body -bisphosphoglycerate shunt is a side reaction of glycolytic pathway in which 1,3 BPG is converted to 2,3BPG to serve as an allosteric inhibitor of O2 binding to heme -2,3 BPG reenters glycolysisas 3-phosphoglycerate Regulation of Glycolysis by the need for ATP – pathway is designed to regulate and maintain ATP homeostasis in cells -PFK-1 and PDH links glycolysis and TCA cycle are major regulatory sites that respond to feedback indicators of ATP utilization -supply of G6P is tissue dependent and is regulated by glycogenolysis and rate of phosphorylation by hexokinase enzymes Relationships among ATP, ADP, AMP Concentrations – AMP levels provide a better indicator of rate of ATP utilization than ATP concentrations itself 2 ADP AMP + ATP -AMP activates glycolysis, glycogenolysis, fatty acid oxidation to maintain ATP homeostasis Regulation of Hexokinases – hexokinases are normally inhibited by glucose-6-phosphate, and if G6P does not enter glycolysis or other pathways, it accumulates and inhibits hexokinase -in liver, Km of glucokinase is high, so glycolysis can continue even if energy is high Regulation of Phosphofructokinase-1 – PFK-1 is a rate-limiting step of glycolysis that regulates rate of glucose-6-P entry into glycolysis -PFK1 is an allosteric enzyme with 6 binding sites, two for substrates and 4 for regulatory sites -AMP and F-2,6-bisP are ACTIVATORS -ATP, citrate are INHIBITORS -PFK comes as isozymes called M (muscle), L (liver), and C (other tissues) -AMP increases affinity of PFK-1 for fructose-6-P (shifts kinetic curve to the left) -F-2,6-bisP opposes ATP inhibition and is not an intermediate in glycolysis by is synthesized by an enzyme that phosphorylates F6P at 2-position (PFK-2) -PFK2 is regulated through change in ratio of activity of two domains (phosphatase and kinase) -PFK2 can also be regulated through phosphorylation, which can increase kinase activity and increase fructose 2,6-bisP levels to contribute to glycolysis activation Allosteric Inhibition of PFK-1 at Citrate Site – function of citrate allosteric site on PFK1 is to integrate glycolysis with other pathways Regulation of Pyruvate Kinase – pyruvate kinase exists are R (RBC), L (Liver), M1/2 (muscle) isozymes have various allosteric sites -liver pyruvate kinase can be inhibited through phosphorylation by cAMP kinase and activated by other effectors (F-1,6-bisP) Pyruvate Dehydrogenase Regulation and Glycolysis – PDH is regulated principally by rate of ATP utilization through rapid phosphorylation to an inactive form -glycolysis and TCA are regulated together in a cell in adequate oxygen. In low O2, NADH/NAD ratio inhibits PDH, but AMP activates glycolysis -portion of pyruvate is then reduced to lactate to allow glycolysis Lactic Acidemia – in lactic acidosis, lactic acid accumulates in blood to levels that significantly affects pH -lactic acidosis results from greatly increased NADH/NAD ratio in tissues -increased NADH concentrations prevens pyruvate oxidation in TCA cycle and directs pyruvate to lactate -to compensate for decreased ATP from OxPhos, PFK-1 is activated and glycolysis activated -drinking alcohol increases NADH levels and can lead to lactic acidosis -hypoxia increases lactate production -Impaired PDH activated from inherited deficiency of E1 (subunit in complex), or from severe thiamine deficiency, increases blood lactate levels -Pyruvate carboxylase deficiency can lead to lactic acidosis